1.einleitung · 17-alpha-progesteron zu progesteron, dann zu androgen und schließlich zu Östrogen...
TRANSCRIPT

1
1.Einleitung
Der menschliche Embryo stellt das Fortpflanzungsprodukt zweier histoinkompatibler
Individuen dar. Daher handelt es sich bei jeder Schwangerschaft um einen einzigartigen
Lebensabschnitt, in dem das Immunsystem der Mutter den zur Hälfte mit väterlichen
Fremdantigenen beladenen Embryo akzeptiert. Da hierbei das mütterliche Immunsystem in
direktem Kontakt mit den für sie fremden, paternalen Zellen steht, wurde die
Schwangerschaft lange Zeit mit einer „Allotransplantation“ verglichen [1].
Das maternale Immunsystem muss während der Schwangerschaft vielfältige Aufgaben
übernehmen: zum einen den Schutz der Mutter vor Infektionen. Zusätzlich ist auch das Kind
vor eindringenden Erregern zu schützen. Zum andern darf der Fetus vom maternalen
Immunsystem nicht als „fremd“ erkannt werden, damit eine Abstoßungsreaktion verhindert
und der Erhalt der Schwangerschaft gewährleistet werden kann. Diese Fähigkeit ist als
essentielle Voraussetzung für das Überleben des Fetus und somit für eine erfolgreiche
Schwangerschaft anzusehen. Deshalb findet sich während der Schwangerschaft eine
einzigartige, veränderte maternale Immunitätslage, woran insbesondere das adaptive
Immunsystem beteiligt ist [2].
Es stellt sich nun die Frage nach den regulatorischen Mechanismen, welche im Rahmen des
Schwangerschaftsgeschehens eingreifen und diese Veränderung der Immunitätslage der
Mutter bewirken.
Im frühen Stadium der Schwangerschaft kommt es zu multiplen morphologischen,
biochemischen und immunologischen Veränderungen im uterinen Milieu der Mutter, welche
die Implantation der Blastozyste in das Endometrium ermöglichen [3]. Dies erfordert eine
subtile Kommunikation und Interaktion zwischen Mutter und Embryo. Betrachtet man
zunächst den Hormonhaushalt der Mutter, so ist zu beobachten dass sich das Endokrinium
umstellt um die Gegebenheiten für den Embryo zu optimieren. Wichtig sind hierbei vor allem
die Hormone Humanes Choriongonadotropin, Östrogen und Progesteron, die im Folgenden
näher beschrieben werden:
1.1 Humanes Choriongonadotropin:
Das Humane Choriongonadotropin (beta-HCG) stellt ein in der Plazenta gebildetes
Gonadotropin dar, ein Hormon das denen des Hypophysenvorderlappen ähnelt. Beta-HCG ist
ein Glykoprotein mit einem Molekülgewicht von 38 kDa, bei dem eine ausgeprägte
Mikroheterogenität vorliegt. Es bindet an den selben Rezeptor wie das Luteinisierende

2
Hormon (LH), weist allerdings eine erheblich höhere Affinität zu diesem Rezeptor auf und
wird wegen seiner LH-ähnlichen Wirkung auch als luteotropes Hormon bezeichnet. Es
zeichnet sich durch eine lange Halbwertszeit aus, die bei 96 Stunden liegt [4].
Beta-HCG wird von der menschlichen Blastozyste bereits während der Implantationsphase in
geringen Mengen gebildet [6, 57]. Nach Abschluss der Implantation wird das vom
Synzytiotrophoblasten gebildete beta-HCG in den maternalen Kreislauf abgegeben und kann
im Blut der Mutter ca. 6 Tage post ovulationem nachgewiesen werden. Maximale
Konzentrationen von beta-HCG liegen nach ungefähr 10 Schwangerschaftswochen vor, mit
Werten über 100000 IU/L. Danach fallen sie wieder ab auf einen Plateauwert von 10000-
20000 IU/L, der in der 16. SSW erreicht wird und bis zum Geburtstermin auf diesem
konstanten Niveau bestehen bleibt [5].
Die Hauptaufgabe des beta-HCG besteht darin den Fortbestand der Schwangerschaft zu
gewährleisten, was durch die Stimulation des Corpus luteum geschieht. Durch die Bindung an
den LH-Rezeptor induziert beta-HCG im Corpus luteum die Bildung der
Schwangerschaftshormone Progesteron und Estradiol. Dies wiederum bewirkt die Zunahme
der endometrialen Sekretion und des uterinen Wachstums um die Überlebensbedingungen für
den Embryo zu sichern.
Auch für den Fetus spielt Humanes Choriongonadotropin eine wichtige Rolle, da es die
fetalen Gonaden stimuliert. Beim männlichen Feten werden die Leydig-Zellen zur
Testosteronproduktion anregt, die Ovarien der weiblichen Feten werden zur Follikelreifung
bis hin zum Tertiärfollikel stimuliert [6].
1.2 Östrogen:
Östrogene sind Steroidhormone, die aus 18 Kohlenstoffatomen aufgebaut sind und sich durch
ihre Hydroxylgruppen unterscheiden. Beim Menschen unterscheidet man Östron, Östriol,
sowie die biologisch aktivste Substanz, das Östradiol. Sie entstehen aus Cholesterol, das via
17-alpha-Progesteron zu Progesteron, dann zu Androgen und schließlich zu Östrogen
umgewandelt wird. Der Abbau der Östrogene erfolgt in der Leber, die Ausscheidung über die
Nieren [7]. Bei der Frau werden die Östrogene vor allem in den Theka-interna-Zellen und den
Granulosazellen der Follikel, und im Stratum granulosum der Nebennierenrinde gebildet.
Während der Schwangerschaft wird Östrogen unter dem Einfluss von beta-HCG im 1.
Trimenon vom Corpus luteum, später von der Plazenta produziert. In der Menopause ist es
vor allem das Fettgewebe, das den Hauptproduktionsort für Östrogen darstellt.

3
Während der Follikelphase steigt die Östrogenkonzentration bis auf 350 pg/ml an.
Postovulatorisch kommt es zum kurzfristigen Absinken der Konzentration, um etwa eine
Woche post ovulationem in der Lutealphase auf Werte bis zu 200 pg/ml anzusteigen. Gegen
Zykusende kommt es wieder zum Abzusinken der Östrogenkonzentration. Im Verlauf der
Schwangerschaft kommt es zu einem konstanten Ansteigen der Östrogenkonzentration, wobei
die Werte zum Zeitpunkt der Geburt mit bis zu 25 ng/ml etwa 1000 mal höher liegen als zum
Beginn der Schwangerschaft [5].
Östrogene haben vielfältige Wirkungen auf den weiblichen Körper, die über
Östrogenrezeptoren auf den Zielzellen vermittelt werden: Östrogene verleihen der Stimme
einen weichen Klang, Fördern das Wachstum und die Proliferation der Brustdrüse, dienen der
Entwicklung des subkutanen Fettgewebes und seiner Verteilung. Am Uterus bewirken sie den
zyklusabhängigen Aufbau der neuen Schleimhaut und fördern das Dickenwachstum des
Myometriums. An der Zervix öffnet sich präovulatorisch der Muttermund, die Menge und
Spinnbarkeit des Zervixsekretes nimmt zu und die Viskosität des Zervixschleims nimmt ab. In
der Vagina lassen die Östrogene das Epithel proliferieren und der Karyopyknoseindex steigt
an. In den Knochen werden die Osteoblasten aktiviert. Die sexualunspezifischen Wirkungen
lassen sich als allgemein protein-anabole zusammenfassen. Die Wasserretention unter
Östrogeneinfluss führt zu einer Ablagerung in Haut und Schleimhäuten, und durch die
Stimulation der Prolaktinsekretion kommt es zu einer gesteigerten Synthese der
Hormontransportproteine in der Leber [4].
1.3 Progesteron:
Progesteron ist der wichtigste Vertreter der Gestagene, welches Steroidhormone mit 21 C-
Atomen sind. Die Bildung von Progesteron erfolgt im Gelbkörper in den Granulosa- und
Theka-Luteinzellen, in der Plazenta und in geringer Menge auch in der Nebennierenrinde [4].
Die Progesteronkonzentration ist zyklusabhängig: in der Follikelphase findet sich ein
niedriger Progesteronspiegel mit einem Normwert <1ng/ml, der mit Beginn des
präovulatorischen LH- Anstiegs zunimmt. In der anschliessenden Lutealphase kommt es zu
einem konstant hohen Niveau der Progesteronkozentration >12 ng/ml. Mit Rückbildung des
Corpus-luteum fällt auch der Progesteronspiegel wieder ab, und der Zyklus beginnt von
neuem. Progesteron wird zu Pregnandiol abgebaut und als solches über die Nieren
ausgeschieden. Die Aufgaben von Progesteron sind die sekretorische Transformation des
Endometriums und die Glykogeneinlagerung in die Deziduazellen. An der Zervix führt es zu
einem postovulatorischen Verschluss des Muttermundes, zur verminderten Sekretion und

4
Spinnbarkeit des Zervixschleims. Die Alveoli der Brustdrüse werden angeregt zur
Proliferations- und Sekretionsbereitschaft. Auf das Myometrium wirkt Progesteron
tonusmindernd. Die Körpertemperatur wird durch Progesteron um 0,5 °C erhöht.
Zusammenfassend dienen die Wirkungen des Progesterons der Vorbereitung, sowie dem
Schutz und Erhalt der Schwangerschaft, wobei die Wirkungen rezeptorvermittelt sind. In der
Schwangerschaft liegen die Normwerte von Progesteron im 1. Trimenon bei 10-50 ng/ml, im
2.Trimenon bei 20-130 ng/ml und bei 130-260 ng/ml im 3. Schwangerschafts-Trimenon [7].
1.4 Dexamethason:
Zusätzlich wurde in der vorliegenden Arbeit auch der Einfluss von Dexamethason untersucht,
welches als Glucocorticoid einen Vertreter der Steroidhormone darstellt.
Neben den physiologischen Wirkungen, die sich vor allem auf den Kohlenhydrat-, Fett- und
Proteinstoffwechsel beziehen, sollen hier ganz besonders die Wirkungen betrachtet werden,
die sich erst in höheren Dosen finden. Die katabole Wirkung auf das Blut- und Lymphsystem,
bei der es zu einer Lymphopenie im peripheren Blut kommt, welche auf eine
Umkompartimentierung der Lymphozyten zurückzuführen ist und vor allem die T-Zellen
betrifft. Die Zahl der Eosinophilen und Basophilen wird ebenfalls vermindert, jedoch nimmt
die Leukozytengesamtzahl durch Anstieg der Neutrophilen zu. Ausserdem kommt es unter
dem Einfluss von Dexamethason zu einer Thrombo- und Erythrozytose.
Die antiphlogistischen, immunsuppressiven und antiallergischen Wirkungen sind unspezifisch
und symptomatisch, d.h. sie sind nicht gegen eine bestimmte Noxe gerichtet, sondern es
werden die allgemeinen Reaktionen des Organismus gegen ein schädigendes Agens
unterdrückt [4]. Immunsuppressiv wirksam ist Dexamethason durch den Einfluss auf die
dendritischen Zellen. Es steigert die Produktion der antiinflammatorischen Cytokine und
unterdrückt die Produktion der cytotoxischen Cytokine, wie z.B. Interleukin-12, die eine Th-1
Immunantwort hervorrufen würden [8]. Zusätzlich inhibieren Glucocorticoide auch die
Reifung der dendritischen Zellen [9].
Glucocorticoide reduzieren die gesteigerte Gefäßpermeabilität und vermindern den Austritt
von Makrophagen und Leukozyten in das Gewebe, wodurch eine Ansammlung von
Entzündungszellen verhindert wird. Sie hemmen verschiedene Phospholipasen und die
Cyclooxygenase COX 2 und reduzieren so das entzündliche Geschehen.
Das Immunsystem beeinflussen die Glucocorticode durch eine Verminderung der T-
Lymphozytenzahlen. Sie hemmen die Phagozytosefähigkeit der Makrophagen und hindern
diese daran proinflammatorische Cytokine wie z.B. IL-1 oder TNF-alpha zu produzieren.

5
Die Synthese der Glucocorticoide verläuft wie folgt: ausgehend vom Cholesterin entstehen
über Pregnenolon und Progesteron die Glucocorticoide. Sie werden in einem zirkadianen
Rhythmus ins Blut abgegeben mit einer maximalen Ausschüttung in den Morgenstunden.
Glucocorticoide diffundieren in die Zielzellen um dort an spezifische, zytoplasmatische
Rezeptoren zu binden. Dexamethason stellt im Bezug zu Cortisol das Glucocorticoid mit der
stärksten antiphlogistischen Wirkung dar und besitzt keinerlei mineralocorticoide Wirkung
[4].
Auch der Embryo ist in der Lage Hormone zu bilden um mit der Mutter in Kontakt zu treten
[10]. Die hormonale Interaktion zwischen Mutter und Kind ist von mütterlicher Seite aus
durch die Trophoblastenzellen der Plazenta gewährleistet, da diese in der Lage sind Hormone
zu produzieren, wie z.B. das beta-HCG in den ersten 12-14 Wochen der Schwangerschaft. Als
wichtige Vertreter dieser Hormone ist neben beta-HCG das GnRH zu nennen. Aber auch
Prostaglandin E2 und Plättchen- aktivierender Faktor sind neben vielen anderen Substanzen
wie Cytokinen, Interferon-gamma oder dem „early pregnancy-factor“ wichtige, von
kindlicher Seite ausgehende Kommunikationsmittel [3].
Diese Art der Signalübertragung zwischen Mutter und Kind ist ebenso faszinierend wie
effizient und Gegenstand aktueller Forschungen. In der vorliegenden Arbeit sind vor allem die
Hormone Progesteron, Östrogen, humanes Choriogonadotropin, sowie Dexamethason näher
betrachtet worden, da das Hauptaugenmerk im Verlauf der Schwangerschaft auf diesen
Hormonen liegt. Bei einer normal verlaufenden Schwangerschaft treten jedoch nicht nur
Hormonveränderungen auf, sondern auch das vorherrschende Cytokinprofil der Mutter ist
verschoben. So dominieren die Th2 Cytokine bei einer erfolgreichen Schwangerschaft,
während ein Th1 Cytokinprofil mit einer normalverlaufenden Schwangerschaft nicht zu
vereinbaren ist [11].
Unter Cytokinen versteht man humorale Stoffe, die der unspezifischen Immunität zugerechnet
werden und die als Botenstoffe besonders der Kommunikation zwischen dem spezifischen
und unspezifischen Immunsystem dienen [12]. Es ist eine heterogene Stoffgruppe mit
gemeinsamen Strukturprinzipien: so handelt es sich meist um kleine Proteine bzw.
Monomere, die glykolysiert sind und zu denen man Interferone, Interleukine und
Wachstumsfaktoren rechnet. Die Cytokine wirken nach dem pleiotropen Wirkprinzip, das
bedeutet dass je nach Zielzelle ein und dasselbe Cytokin unterschiedliche Wirkung
hervorrufen kann. Ausserdem können die Wirkungen in ein und der selben Zelle
unterschiedlich sein. Die Redundanz der Cytokine wird als das Phänomen bezeichnet,

6
welches zu gleichen Wirkungen in Zielzellen durch unterschiedliche Cytokine führt. Auf
diese Art und Weise entsteht ein sehr komplexes Signalübermittlungssystem zwischen den
einzelnen Zellen des Immunsystems, dessen Klassifizierung zu Verwirrung geführt hat [13].
Die Einteilung erfolgt heute in Th1- und Th2 Cytokine, wobei das Th1-Profil während
immunologischer Aktivität beispielsweise bei Infekten vorrangig ist um eine cytotoxische
Immunantwort einzuleiten und den Körper auf diese Weise vor Erregern zu schützen.
Zur Gruppe der Th1 Cytokine gehören Interleukin (IL)-6, IL-18, IL-12, IFN-gamma und
TNF-alpha, während typische Vertreter der Th2-Klasse die Interleukine IL-4, IL-5 und IL-10
sind. Als Vertreter der Chemokine bezeichnet man IL-8 und MDC [14]. Im Folgenden soll
auf die einzelnen Cytokine näher eingegangen werden:
1.5 Interleukin-6:
Interleukin-6 (IL-6), ein Vertreter der proinflammatorischen Th1-Cytokine, wurde
ursprünglich definiert als B-Zell-stimulierender Faktor 2. Die Aufgabe der
proinflmmatorischen, also der entzündungsauslösenden Cytokine, ist es, durch Aktivierung
der körpereigenen Abwehr den Körper vor Erregern zu schützen. IL-6 wird hauptsächlich von
Monozyten, B-Zellen, T-Zellen, Endothelzellen, Fibroblasten und Keratinozyten produziert,
aber auch Tumorzellen, wie z.B. das Blasen- oder auch das Zervixkarzinom sind in der Lage,
IL-6 zu bilden. IL-6 ist ein essentieller Faktor für die finale B-Zell-Reifung zu antikörper-
produzierenden Plasmazellen und ist somit grundlegend für die gezielte körpereigene Abwehr
innerhalb des erworbenen Immunsystems. Es wirkt außerdem als Kofaktor bei der
Aktivierung von T-Zellen mit, und fördert zusammen mit IL-2 die Differenzierug der T-
Zellen in cytotoxische T-Zellen [15]. Diese spielen wiederum eine wichtige Rolle in der
Körperabwehr. Weiterhin ist IL-6 ein wichtiger Regulator der Akute-Phase-Reaktion, einem
Instrument der unspezifischen Abwehr, und stimuliert in diesem Zusammenhang die Synthese
und Freisetzung der Akute-Phase-Proteine wie z.B.: CRP, Fibrinogen oder Haptoglobin [15].
Auch auf die Hämatopoese hat das IL-6 vielfältige Wirkungen und kann somit auf die
Produktion der für das Immunsystem wichtigen Effektorzellen direkt einwirken. Es stimuliert
die hämatopoetischen Vorläuferzellen, steigert die Thrombozytopoese und fördert die
Ausreifung der Megakaryopoese. Im Tiermodel beschleunigt IL-6 die Erholung des
Knochenmarks nach cytotoxischer Behandlung und nach Knochenmarktransplantation [13].
Man kann somit das IL-6 als Stimulans für Zellwachstum und Zellreifung ansehen, was
jedoch nicht nur Vorteile mit sich bringt, denn auch bei pathologischen Prozessen ist das IL-6
von großer Wichtigkeit. Es kann als Wachstumsfaktor für verschiedene Tumorerkrankungen
gelten. In diesem Zusammenhang ist IL-6 mit Tumoren, wie dem maligen Myelom oder dem

7
Endometriumkarzinom assoziiert, deren unkontrolliertes Wachstum unter dem Einfluss dieses
Cytokins steht [16].
Besonders hervorzuheben ist auch die antiinflmmatorische Potenz des IL-6, wodurch die
Komplexität der Interaktionen des humanen Immunsystems verdeutlicht werden kann. Durch
die Fähigkeit des IL-6 die Spiegel der inflammatorischen Cytokine TNF-a und IL-1 zu senken
wird per se eine antiinflammatorische Reaktion hervorgerufen und somit eine weitere
Kontrolle der Immunantwort möglich [15].
1.6 Interleukin -18:
Interleukin-18 (IL-18) ist ein Monomer, welches aus 157 Aminosäuren aufgebaut ist. Es wird
von zahlreichen Zellen, z.B. aktivierten Makrophagen, Kupferzellen, oder auch
von dendritischen Zellen produziert und reguliert die Immunantworten sowohl im
angeborenen, als auch im erworbenen Immunsystem. Erkannt wird das IL-18 vom IL-18
Rezeptor, durch den es seine Wirkungen in den Zellen vermitteln kann. Dieser Rezeptor ist
weit verbreitet und befindet sich auf der Oberfläche der verschiedenen Zellen der beiden
Untereinheiten des humanen Immunsystems. Aus diesem Grund kann das IL-18 als
Bindeglied zwischen den beiden Schenkeln des Immunsystems angesehen werden [4]. IL-18
moduliert die Aktivität von Th1- und Th2-Zellen, B-Zellen, Natürlichen-Killer-Zellen sowie
dendritischen Zellen und ist in der Lage, in Abhängigkeit vom umgebenden Cytokinprofil,
sowohl Typ1-, als auch Typ2-Immunantworten hervorzurufen. Dies ist zum einen wichtig in
der Abwehr von Infekten, bei denen durch Beeinflussung der Zellen in ihrer
Cytokinproduktion das Interleukin-18 cytotoxische Immunantworten hervorrufen kann [17].
Zum anderen ist es während der Schwangerschaft wichtig durch das Cytokinprofil eine
protektive Immunantwort zu erhalten, was durch die Präsenz von IL-18 in der
Amnionflüssigkeit gewährleistet zu sein scheint. Mit zunehmendem Alter der physiologischen
Schwangerschaft steigt IL-18 in seiner Konzentration an, jedoch können zu hohe IL-18
Spiegel im Schwangerschaftsverlauf zu Aborten führen [18].
Unter dem Einfluss des IL-18 kommt es zu einer Expressionssteigerung von TNF-alpha, IL-1
beta und IFN-gamma bei den T-Zellen und NK-Zellen, wodurch die cytotoxische
Immunantwort gefördert und die tolerogene Immunantwort unterdrückt wird. Weiterhin teilt
IL-18 die biologische Aktivität mit IL-12, um eine Th1-Antwort hervorzurufen [19]. Von
ganz besonderer Bedeutung ist der Einfluss des IL-18 auf die dendritischen Zellen. Diese
werden unter dem Einfluss von IL-18 befähigt T-Zellen zu aktivieren und haben somit eine
herausragende Rolle im immunologischen Geschehen [31].

8
1.7 Interleukin-12:
Dieses ist ein cytotoxisches Interleukin (IL-12), welches eine Th1-Immunantwort hervorruft.
Es handelt sich um ein Heterodimer aus einer IL-12p70 und einer IL-12p40 Untereinheit und
wird auch als NK-Zell stimulierender Faktor bezeichnet. Produziert wird IL-12 vor allem von
B-Zellen, Makrophagen und dendritischen Zellen. Seine Funktionen sind die Aktivierung von
natürlichen Killerzellen und es induziert die Differenzierung der T-Zellen zu Th1-Zellen.
Diese produzieren dann Interferon-gamma (IFN-gamma), wodurch eine cytotoxische
Immunantwort zustande kommt. IFN-gamma selbst wirkt wiederum als verstärkendes Agens
auf die Ausschüttung von IL-12 [20]. IL-12 entsteht bei Infektionen in der frühen Phase der
Immunreaktion und kann durch IL-10, sowie durch Fibroblasten in seiner Funktion blockiert
werden [21]. Induziert wird die Bildung von IL-12p70 durch die Interaktion von DC 40+
Zellen mit dem CD 40- Ligand auf den T-Zellen nach deren Aktivierung. Jedoch sind es vor
allem die unreifen dendritischen Zellen die von IL-12p70 aktiviert werden, auf reife DC hat
dieses Interleukin keinen Einfluss [22]. Mittels IL-12 ist es den DC möglich naive CD 40+ B-
Zellen und auch Gedächtnis-B-Zellen zu aktivieren und in Antikörper-produzierende
Plasmazellen umzuwandeln.
Im Rahmen einer Infektion produzieren unreife dendritische Zellen das IL-12 um auf diese
Weise chemotaktische Anreize für die Immunzellen zu liefern, um z.B. Makrophagen, T-
Zellen, oder NK-Zellen zum Ort des immunologischen Geschehens zu locken. Reife DC
hingegen benutzen IL-12 um die Differenzierung der T-Zellen in die Wege zu leiten [23]. So
könnte der Th2-Weg, der nicht cytotoxische Weg der Immunantwort als „Fehler“ des
Immunsystems betrachtet werden, der gewählt wird wenn kein IL-12 ausgeschüttet wird.
IL-12 könnte inhibitorische Effekte auf die Ausbildung von Th2-Zellen haben, aber es
aktiviert die Th1-Zellen und regt diese zur Bildung von IFN-gamma an. Auf die IL-4
Produktion hat das Interleukin-12 keine Effekte [24]. Interleukin-12 bildet ein wichtiges
Bindeglied zwischen der angeborenen und der adaptiven Immunantwort da es im Rahmen
einer Entzündung folgende Reaktionen hervorruft: in der frühen Phase wird es von NK-Zellen
und T-Zellen produziert und dann induziert es deren IFN-gamma Produktion. Außerdem ist
das IL-12 für den Wechsel zu Th1 verantwortlich und führt zur optimalen IFN-gamma
Produktion und Verteilung durch die Th1-Zellen als Antwort auf das angreifende Antigen.
1.8 Tumornekrosefaktor-alpha:

9
Tumornekrose-Faktor-alpha (TNF-alpha) zählt zu den klassischen Vertretern der Th1-Gruppe
der Cytokine und führt somit ebenfalls zu einer cytotoxischen Immunantwort. Es handelt sich
um ein nicht-glykolysiertes Molekül mit Disulfidbrücken und ist aus 157 Aminosäuren
aufgebaut. Die Bildung dieses Interleukins erfolgt vor allem in Neutrophilen, aktivierten
Lymphozyten, Makrophagen, NK-Zellen und dendritischen Zellen auf inflammatorische
Stimuli hin. Die Rezeptoren für TNF-alpha werden auf der Oberfläche einer Vielzahl von
Zellen exprimiert, wobei 2 Arten von TNF-alpha Rezeptoren existieren. Der 55 kDa Rezeptor
ist hierbei verantwortlich für die lytische Aktivität, der 75 kDa Rezeptor für die proliferativen
und regulatorischen Signale in Interaktion mit den T-Zellen [25].
Die biologischen Funktionen des Tumornekrosefaktors sind vielfältig: er stimuliert die
cytotoxische Aktivität von Neutrophilen und fördert deren Adhärenz and das Endothel. TNF-
alpha wirkt als endogenes Pyrogen, da es durch die Freisetzung von Prostaglandin E2 und
Interleukin-1 im Körper Fieber auslösen kann. Durch das Zusammenspiel mit IL-1, dessen
Ausschüttung in den Makrophagen und Monozyten durch TNF-alpha gefördert werden,
kommt es außerdem zu einem Aktivierungssignal für Makrophagen, die dadurch in ihrer
Aktivität noch gesteigert werden [26]. TNF-alpha ist ein wichtiges Interleukin für die
regelrechte Entwicklung des lymphatischen Gewebes. Es ist an der Regulation der B-Zellen
beteiligt, da es die B-Zellen zur Proliferation anregt und deren Differenzierung fördert.
Im hämatopoetischen System wirkt es inhibitorisch auf Progenitorzellen, stimuliert aber
gleichzeitig die Freisetzung von Wachstumsfaktoren. Es stimuliert die T-Zellen und induziert
die Expression von MHC-Molekülen und von Interleukinrezeptoren [27].
TNF-alpha steigert die Migration der DC und erhöht somit die Anzahl der Zellen in den
Lymphknoten. Es führt zur vermehrten Bildung von Chemotaktischen Interleukinen wie IL-8
und führt zu einer bis zu 100- fachen Steigerung der Antigenprästentation durch die DCs.
TNF-alpha stellt selbst einen wichtigen Reifungsfaktor für die dendritischen Zellen dar und ist
somit essentiell für den Lebenszyklus dieser Zellen und das Ausführen der Immunantwort des
Menschen [28].
1.9 Interleukin-10:
Das Interleukin-10 (IL-10), oder auch der Cytokinsythese Inhibitor Faktor, ist ein Homodimer
aus 160 Aminosäuren mit einem Molekulargewicht von 18 kDa und zählt zu der Klasse der
Th2-Cytokine. Es wird von Makrophagen, DC und Th2-Zellen produziert und bewirkt durch
die Blockade von IL-12 im Makrophagen, dass diese Zellen die Th1-Zellen nicht aktiveren
können und es somit nicht zu einer cytotoxischen Immunantwort kommt. IL-10 gilt als

10
wirksamster Inhibitor der Makrophagenfunktion und Gegenspieler des cytotoxischen IL-12
[29]. Es hemmt weiterhin die Synthese der cytotoxischen Interleukine gamma-Interferon und
TNF-alpha, die von den Th1-Zellen produziert werden. Auf Proteinebene hemmt Interleukin-
10 die Monokine IL-4, IL-6 und IL-8 und greift auf diese Weise regulatorisch in das
immunologische Geschehen ein, im Sinne einer toleranten Immunantwort. Hinzu kommt eine
Proliferationshemmung der T-Zellen, was der Cytotoxizität entgegensteht.
Auf die Th2-Zellen übt das IL-10 keinen Einfluss aus [30]. Autoregulativ ist IL-10 wirksam
indem es im Sinne einer negativen Rückkopplung die eigene Produktion reduziert, wodurch
eine Limitierung der toleranten Immunantwort gegeben ist und das Immunsystem wieder auf
neue Antigene reagieren kann. IL-10 hat keinen Einfluss auf die MHC-II –Expression der
dendritischen Zellen, oder deren Fähigkeit naive T-Zellen zu stimulieren. Es verhindert aber
die verstärkte Produktion von kostimulatorischen Molekülen in den DC. Und es inhibiert die
Induktion von proinflammatorischen Cytokinen [31].
IL-10 blockiert die Cytokinsynthese und wirkt somit als Suppressor der Effektorfunktionen
von Makrophagen, T-Zellen und NK-Zellen. Auf diese Weise reguliert es die Differenzierung
von B-Zellen, Mastzellen und Thymozyten. Für die B-Zellen wirkt IL-10 als Kostimulans für
deren Proliferation und es führt als Differenzierungsfaktor dazu, dass die B-Zellen Antikörper
produzieren. Während der Schwangerschaft ist IL-10 wichtig für deren Erhalt, da es die Th1-
Zellen davon abhält cytotoxische Cytokine zu produzieren, welche zu einem Abort führen
würden [32]. IL-10 ist ein multifunktionales Cytokin mit unterschiedlichen Wirkungen auf die
meisten hämatopetischen Zelltypen. Die Hauptfunktion zeigt sich in einer Unterdrückung der
inflammatorischen Reaktion durch herabregulieren der cytotoxischen Immunantwort.
1.10 Interleukin-8:
Interleukin-8 (IL-8) gehört zur Klasse der Chemokine, worunter man Cytokine versteht, die in
den ersten Phasen entzündlicher Reaktionen im Körper freigesetzt werden, um eine gerichtete
Chemotaxis zu induzieren. D.h. sie sind dafür zuständig die für die Entzündung wichtigen
Zellen an den Ort des Geschehens zu locken. IL-8 ist ein von Monozyten abstammendes
Peptid, das chemotaktisch auf neutrophile Granulozyten wirkt und diese Zellen aktiviert. Die
Hauptproduzenten des IL-8 sind die dendritischen Zellen, daneben wird es aber auch von
Monozyten, Makrophagen, Fibroblasten und anderen Zellen produziert. Seine Synthese wird
durch TNF-alpha und IL-1 induziert [33].
IL-8 ist ein nicht-glykolysiertes Protein, das aus 72 Aminosäuren besteht und eine Molmasse
von 8 kDa hat. Es besitzt außerdem 4 Cysteinreste, die für seine biologische Aktivität wichtig

11
sind und zur Einteilung in die CXC-Chemokine geführt haben. Die Funktionen von IL-8 sind
neben der Aktivierung von Granulozyten, die Bildung von Sauerstoffradikalen, Verstärkung
der Chemotaxis und Expression von Adhäsions-molekülen. Dies sind Reaktionen die vor
allem zu Beginn der entzündlichen Reaktion in Körper wichtig sind um eine gezielte Abwehr
zu ermöglichen [31].
1.11 Macrophage-derived chemokine:
Macrophage-derived chemokine (MDC) zählt zur Subklasse der CC Chemokine und wird von
Makrophagen und vor allem von dendritischen Zellen produziert. Sein genetischer Pool ist
bereits vollständig codiert und liegt auf Chromosom 16. Es wirkt dosisabhängig als potentes
Chemotaxin für DC und aktivierte NK-Zellen mit einem Wirkungsmaximum bei 1 ng/l [34].
Vor allem unreife dendritische Zellen werden von MDC schnell an den Ort der Entzündung
gelockt und sind dann sehr effizient in der Antigenaufnahme. Auf den Reifungsvorgang der
DC hat das MDC jedoch keinen Einfluss. Im Thymus hat MDC die Aufgabe die dendritischen
Zellen entweder anzulocken oder zurückzuhalten, um die T-Zell-Stimulation zu verstärken.
Chemotaktisch wirkt MCD ebenfalls auf T2-Zellen, die CD4+, bzw. CD8+ sind. MDC ruft
eine Typ II Immunantwort hervor, wobei entzündliche Reize zu einem erhöhten MDC-
Spiegel führen [35].
Während der Schwangerschaft ist ein Shift von Th1 in Richtung Th2 zu beobachten. Dieser
soll den Embryo vor Angriffen des mütterlichen Immunsystems schützen, da die Th1
Cytokine die Plazenta direkt, bzw. indirekt mittels Aktivieren von cytotoxischen Zellen,
schädigen können [36]. Im Zusammenhang mit der Cytokinproduktion und
Cytokinsignalverarbeitung von herausragender Bedeutung erscheinen die dendritischen Zellen
(DC), welche eine zentrale Rolle im menschlichen Immunsystem einnehmen. Da diese Zellen
in der Lage sind Cytokine zu produzieren und auf Cytokinsignale hin zu reagieren, können sie
direkt auf das immunologische Geschehen einwirken und die Immunantwort modulieren.
Diese Eigenschaft der DC, die Fähigkeit Cytokine zu produzieren, war der zentrale Punkt der
vorliegenden Arbeit, in der die Cytokinproduktion von unreifen dendritischen Zellen nach
Einwirken von schwangerschaftsassoziierten Hormonen gemessen wurde.
1.12 dendritische Zellen
Erstmals entdeckt 1973 von Steinman und Cohn in der Milz von Mäusen, wurden diese Zellen
nach ihrem charakteristischen Aussehen mit den vielen Ausläufern, den sogenannten

12
Dendriten, benannt [37]. Wenig später wurde erkannt dass diese Zellen dem gleichen
Zellsystem angehörten wie die schon vor Hundert Jahren von Langerhans entdeckten
Langerhans-Zellen in der Epidermis, welche dort eine wichtige Funktion im Sinne der lokalen
Erregerabwehr haben. Bald wurden die dendritischen Zellen auch in anderen Organen, sowohl
in lymphatischen, als auch in nicht-lymphatischen, nachgewiesen, und die Expression einer
Vielzahl von HLA-Antigenen legte nahe dass es sich bei den DC um spezialisierte,
antigenpräsentierende Zellen handelt, die eine zentrale Stellung im menschlichen
Immunsystem innehaben.
Diese hochspezialisierten Abkömmlinge von Leukozyten kommen in fast allen Geweben des
Körpers, und so auch im Uterus vor. Sie bilden ein Netz von Zellen aus, das Fremdmaterial
erkennen kann und für dessen Beseitigung sorgen soll. Auf diese Weise erfüllen die
dendritischen Zellen eine Art „Wächterfunktion“, da sie den Körper vor eindringenden
Erregern schützen und über die Integrität der körpereigenen Zellen wachen [38, 39, 40]. Sie
können Fremdmaterial, in der Regel Proteine, mit Hilfe der Phagozytose aufnehmen und
intrazellulär prozessieren, d.h. verarbeiten. Die so zerkleinerten Moleküle werden an HLA-
Moleküle gebunden und an die Zelloberfläche transportiert damit T-Lymphozyten das fremde
Material erkennen können. Während dieser Vorgänge liegen die DC noch in unreifer Form
vor. Wichtig ist weiterhin die Fähigkeit mit dem internalisierten Antigen zum Lymphknoten
zu wandern, zum Ort des immunologischen Geschehens mit Sitz der T-Lymphozyten. Auf
diesem Weg durchlaufen die DCs einen Reifungsprozess. Dieser wird durch Signale der
Antigene und Signale der aktivierten T-Zellen mittels CD 40-Ligand getriggert [41].
Im Lymphknoten findet nun die Aktivierung der naiven T-Zellen statt. Die dendritischen
Zellen zünden die Immunantwort und wirken als natürliches Adjuvans bei der T-Zell
Aktivierung. Sie gelten als stärkste natürliche T-Zell Stimulatoren, die in ihrer Wirksamkeit
alle anderen Antigen-Präsentierenden-Zellen um ein Vielfaches an Effizienz und Potenz
übertreffen [42]. Aufgrund dieser Erkenntnisse setzt man hohe Erwartungen in die
dendritischen Zellen, um diese im Sinn von Immunmodulatoren einsetzen zu können.
Denkbar wären beispielsweise der Einsatz bei der Therapie von Autoimmunerkrankungen, bei
Tumoren oder in der Transplantaionsmedizin.
Um die Kontrolle bei einer Immunantwort nicht zu verlieren und das Immunsystem nach
Beseitigung der Antigene wieder herunterzufahren, haben die dendritischen Zellen eine
Apoptosefunktion, d. h. sie können sich selbst ausschalten und so die Immunantwort des
Körpers beenden [43].

13
Funktionsprinzip von APC:
Antigenaufnahmeund VerarbeitungWanderung
undReifung
Im Lymhknoten:Aussuchen derpassenden T-Zellen
Aktivierung derpassenden T-Zellen
T-Zellen wandern insGewebe, bekämpfendie fremden Antigene
Unreife Wächterzellen
Abbildung1: Schematischer Lebenszyklus von dendritischen Zellen.
Unreife Wächterzellen nehmen fremde Antigene (orange Punkte) auf und werden aktiviert.
Während sie die Antigene verarbeiten, wandern sie über die Lymphgefäße in die
Lymphknoten. Hier „screenen“ sie die ruhenden T-Zellen, bis sie diejenigen finden, welche
das fremde Antigen erkennen (rote Zellen). Diese T-Zellen werden darauf hin zur Teilung
angeregt und aktiviert (große rote Zellen). Die im Lymphknoten aktivierten T-Zellen wandern
dann über das Blutgefäßsystem in das Gewebe mit den fremden Antigenen (z.B. infiziertes
Gewebe) ein, um die eingedrungenen Fremdstoffe und Organismen zu bekämpfen.
Seit es gelungen ist die dendritischen Zellen aus Monozyten der peripheren Blutes zu isolieren
und anzuzüchten, wurde noch mehr über die Wirkungsweise dieser einzigartigen
Zellpopulation erforscht. So konnte man viel über die Cytokinproduktion erfahren, die je nach
ihrer Art, zu einer Aktivierung der cytotoxischen T-Zellen führt oder eben keine solche
Antwort hervorruft. Von besonderer Wichtigkeit ist das umgebende Cytokinprofil wie bereits
erwähnt während der Schwangerschaft, da eine vorherrschende Th1- Immunität mit einer
erhöhten Abortrate assoziiert ist [44, 11].
Die Schwangerschaft hier als Model für Onkologie und Autoimmunität heranzuziehen
erscheint logisch: führen doch die Th2-Cytokine, die zum Großteil von den dendritischen
Zellen produziert werden, zum Erhalt der teils „fremden Zellmasse“ Embryo, d.h. es findet
trotz der paternalen Signale auf der Zelloberfläche keine cytotoxische Reaktion statt, sondern

14
das Immunsystem ist tolerant gegen den „Fremdling“. Bleibt die Frage offen was die
maternalen Immunzellen dazu veranlasst, diesem bestimmten, weil bei jeder
normalverlaufenden Schwangerschaft gleichen, Muster zu folgen.
Da jedoch nicht nur das Immunsystem, sondern auch der Hormonhaushalt der Mutter
während der Schwangerschaft charakteristische Veränderungen erfährt, drängte sich im
Rahmen der Grundlagenforschung die Frage auf inwieweit die maternale Hormonlage mit
dem Immunsystem interagiert und welcher Zusammenhang zwischen der Hormon- und der
Immunlage der Mutter während einer Schwangerschaft besteht. Es stellte sich die Frage ob
durch die hormonellen Veränderungen im Schwangerschaftsverlauf auch die Veränderungen
im Immunsystem der Mutter beeinflusst werden.
Und so war es das Ziel dieser Studie zu untersuchen, inwieweit die im Körper überall
verbreiteten unreifen dendritischen Zellen unter dem Einfluss von Hormonen während der
Schwangerschaft stehen. Ob der Schlüssel zur einziartigen Immunitätslage während der
Schwangerschaft in dem Zusammenspiel von DC und Schwangerschaftshormonen liegt. Und
die Frage zu beantworten ob die hormonelle Lage der Mutter Einwirkungen auf die
Cytokinproduktion der dendritischen Zellen hat, um eventuell mit Hilfe von
schwangerschaftsassoziierten Hormonen das Immunsystem modulieren zu können.

15
2. Material und Methoden
2.1 Material
Alle Prozentangaben zu Lösungen verstehen sich für Feststoffe als Gewichtsprozent, für
Flüssigkeiten als Volumenprozent.
2.1.1 Geräte
Zellkulturausstattung:
Sterilwerkbank SterilGARD® Class II TypA/B3
Brutschrank Incubator NAPCO 5420-1
Lichtmikroskop Leica DM IRB
Zählkammer Brand Neubauer-Zählkammer Tiefe 0,1mm Fläche 0,0025mm
Zentrifugen :
Hettich EBA 12
Hettich Universal 16R
Waagen:
Kern 510-33
Scaltec SAC 51
Ohaus LS 5000
Sonstige Geräte:
Biophotometer, Eppendorf
ELISA Reader MRX ,Dynex Technologies
Heizblock Techne Dri-Block® DB2A
Magnetrührer Variomag® Electronicrührer, Monotherm
PH-Meter Greisinger Electronic GPRT 1400A
Vortex MS1 Minishaker IKA®
Software:
CellQuest, Becton Dickinson

16
Excel, Microsoft
WinMDI, Joseph Trotter, The Scripps Research Institute, La Jolla, CA
Winword, Microsoft
Statistica StatSoft
GraphpadPrism Prism
2.1.2 Chemikalien und Verbrauchsmaterial
geordnet nach Herstellern
Verbrauchsmaterial:
American National Can™ Parafilm “M”
Brand Reaktionsgefäße, Zentrifugenröhrchen 15ml und 50ml,
Braun Original Perfusorspritze OPS 50ml
Costar Zellkulturplatten: Cell culture Cluster 24 well
Eppendorf Combitips, Pipettenspitzen, Plastikküvetten
Greiner Pipettenspitzen, Reaktionsgefäße, Zentrifugenröhrchen,
Kryoröhrchen, Petrischalen Cellstar®
Hartmann Pehazell® Zellstoff
Nunc ELISA-Platten Nunc-Immuno™ Platte 96-well
TPP Petrischalen für Zellkultur 100*20mm
96-well- Zellkulturplatten für MLR

17
Chemikalien und Medienzusätze:
Biochrom PBS Dulbecco, Alsever Lösung, Zellkulturmedium RPMI1640,
Gentamycinsulfat, Penicillin/Streptomycin
Boehringer Neuraminidase
CAI Biochem Prostaglandin E2
Centeon Beriglobin
DAKO Ziegenserum (normal)
Endogen Merrettichperoxidase konjugiertes Streptavidin
Merck Zitronensäure ( Monohydrat)
Pharmingen Substrat Reagentien A und B (für ELISA)
ELISA-Sets: OptEIA™Set Human IL-8, IL-10, IL-12p70, IL-
13, TNF-alpha
PAN Fetales Kälberserum (FCS)
Sandoz GM-CSF (Leukomax400®)
Serono beta-HCG Pregenesin
Sigma Histopaque–1077, EDTA, Tween20, 2-Mercaptoethanol,
Dexamethason (9a-Fluoro-16a-methylprednisolon), Ethanol,
Trypanblau-Lösung 0,4%, Rinder Serumalbumin, DMSO, 17β-
Estradiol (4-Pregnene-3,20-dione), KHCO3, NH4CL,
Dexamethason
Strathmann Zellkulturzusätze IL-4, IL-6, IL-1β, TNF-alpha
Antikörper:
BD Pharmingen Fluorescein isothiocyanat (FITC)-konjugierter anti-IgG1
Antikörper
FITC-konjugierter anti-human monoklonaler Anti-CD40
Antikörper
FITC-konjugierter anti-human monoklonaler Anti-HLA-DR
Antikörper
Coulter Immunotech Phycoerythrin (PE)-konjugierter antihumaner monoklonaler
Anti-CD83 Antikörper
Cymbus Biotechnology PE-konjugierter anti-IgG1 Antikörper

18
PE- konjugierter anti-human monoklonaler anti-HLA-DR
Antikörper
R&D Systems Beschichtungs- und Dedektions- Antikörper für MDC und IL-18
ELISA
2.1.3 Puffer und Lösungen
Soweit nicht anders angegeben wurde destilliertes Wasser als Lösungsmittel verwendet.
Medienzusätze (Stocklösungen):
Pen/Strep 10000 E/ml Penicillin + 10000µl/ml Streptomycin
Gentamycin 10mg/ml Gentamycinsulfat
GM-CSF 100 U/µl gelöst in RPMI 1640 mit Gentamycin (1:200) mit
0,1% B SA
IL-4 500 U/µl gelöst in PBS mit 0,1% BSA
IL-1β 1000 U/µl in RPMI 1640 mit Gentamycin (1:200) mit 10% FCS
IL-6 1000 U/µl in RPMI 1640 mit Gentamycin (1:200) mit 10% FCS
TNF-alpha 1000 U/µl in RPMI 1640 mit Gentamycin (1:200) mit 10% FCS
PGE2 PBS
Lösungen zur Zellisolierung:
PBS Aus Pulverkonzentrat angesetzt (9,55g gelöst in 1l Wasser)
Alsever Lösung unverdünnt eingesetzt
PBS mit Pen / Strep PBS mit Penicillin/Streptomycin (1:200)
PBS / Citrat 1 Volumenteil 0,106M Na-Citrat-Lösung mit 9 Teilen PBS
PBS / EDTA PBS mit 2mM EDTA
PBS mit Neuraminidase 0,01 U/ml
Lysepuffer für Schafserys 8M NH4Cl
Hormone (Stocklösungen):
Östrogen 5mg/ml gelöst in Ethanol (MW 272,4 g/mol)
Progesteron 5mg/ml gelöst in Ethanol (MW 314,5 g/mol)

19
beta-HCG 5000IE/ml gelöst in physiologischer NaCl-Lösung
Dexamethason 50mM gelöst in DMSO
ELISA:
Beschichtungs- Puffer für IL-6, IL-18 PBS, pH 7,4
Beschichtungs-Puffer für MDC PBS, pH 7,4
Beschichtungs-Puffer für IL-8 8,4g NaHCO3 + 3,56g Na2CO3 auf 1l H2O, pH
9,5
Beschichtungs-Puffer für IL-12p70 11,8g Na2HPO4 + NaH2PO4 auf 1l H2O, pH 9,0
Verdünnungsmittel für OptEIAsets PBS mit 10% FCS
Verdünnungsmittel für MDC PBS mit 1% BSA
Verdünnungsmittel für IL-18 20mM TBS (20mM Tris-Base, 150mM NaCl) mit
0,1% BSA und 0,05% Tween 20, pH 7,3
Verdünnungsmittel für IL-6 PBS mit 4% BSA, pH 7,2 – 7,4
Wasch-Puffer für IL-6 50mM Tris, 0,2% Tween-20, pH 7,9 – 8,1
Wasch-Puffer für IL-18, MDC PBS mit 0,05% Tween-20, pH 7,4
Wasch-Puffer für OptEIAsets PBS mit 0,05% Tween-20, pH 7,4
Substrat-Lösung Tetramethylbenzidine und Hydrogen Peroxid
gemischt im Verhältnis 1:1
Stop-Lösung 2M H2SO4
2.1.4 Zellen, Zellkulturmedien und Medienzusätze
Zellen:
dendritische Zellen isoliert aus Buffycoats von Blutspendern des
Blutspendedienstes des BRK, Institut Wiesentheid und
der Abteilung für Transfusionsmedizin des
Universitätsklinikums Würzburg.
Die Buffycoats wurden ca. 24h nach der Spende
verarbeitet.
T- Zellen isoliert aus Buffycoats (s.o.), in FCS/DMSO
eingefroren und in flüssigem Stickstoff gelagert.

20
Schafserythrozyten isoliert aus Schafblut von Schafen aus den Tierställen des
Universitätsklinikums Würzburg. Das Schafblut wurde
ca. 2h nach Blutabnahme weiterverarbeitet.
Medien:
Differenzierungsmedium RPMI-1640 mit 10% FCS und Gentamycin (1:50µl/ml)
mit GM-CSF (1000 U/ml) und IL-4 (800 U/ml)

21
2.2 Methoden
2.2.1 Reinigung der Schaferythrozyten
Schaferythrozyten mit speziellen Oberflächeneigenschaften erlauben eine Rosettierung von T-
Zellen, die damit eine Isolation von T- Zellen aus einer Zell-Suspension ermöglicht.
Ausgegangen wird von einer Mischung aus 30ml frisch gewonnenem Schafblut, das zur
Gerinnungshemmung direkt bei der Blutabnahme mit 20ml Alseverlösung versetzt wird. Das
Schafblut wird in PBS gewaschen, bei 1100xg 10 min ohne Bremse zentrifugiert und durch
sorgfältiges Abnehmen des Überstandes vom Buffycoat, d.h. allen leukozytären Bestandteilen
des Blutes befreit. Anschließend erfolgt die Inkubation der Schafserythrozyten mit 0,005 U
Neuraminidase pro ml eingesetzten Schafblutes für 30 min bei 37°C. Nach drei weiteren
Waschschritten mit PBS mit Penicillin und Streptomycin wird das Pellet in 0,4 ml
RPMI1640/Genta/10% FCS pro ml eingesetztem Schafblut aufgenommen und bei 4°C im
Kühlschrank gelagert. Die gereinigten Schafserythrozyten können bis circa eine Woche nach
der Isolation verwendet werden. Eine Ausgangsmenge von 50ml Schafblut mit Alseverlösung
reicht für die Isolation von vier humanen Buffycoats.
2.2.2. Isolation dendritischer Zellen und Hormonansätze
Die Isolation dendritischer Zellen erfolgte weitgehend nach dem Standard- Protokoll von
Romani et al., 1996 [58].
2.2.2.1 Isolation von peripheral blood mononuclear cells (PBMC)
Die Isolation der PBMC erfolgte ausgehend von Buffycoats der Blutspendedienste, die
jeweils dem leukozytären Anteil von 500ml Vollblut entsprachen. Die Buffycoats wurden
steril geöffnet und nach Verdünnen mit PBS/Citrat auf Histopaque-Gradienten aufgetragen.
Pro Buffycoat wurden drei 50ml Röhrchen mit je 15ml vorgelegtem Histopaque mit dem
Leukozytengemisch beladen. Nach 30min Zentrifugation bei 400xg ohne Bremse bildeten
sich drei Phasen. In der obersten Phase befand sich das Plasma, welches abgenommen wurde
und nach Abzentrifugieren der Thrombozyten bei 3000g zur weiteren Verwendung kühl
gelagert werden konnte. In der mittleren weißlichen Phase befanden sich die PBMCs die
sorgfältig abgenommen und anschließend in PBS/Citrat gewaschen und 10 min bei 300xg
zentrifugiert wurden. Um die T-Zellen aus dem PBMC- Zell-Gemisch zu entfernen wurde das
PBMC- Pellet in 5ml der vorbehandelten Schafserythrozyten aufgenommen und bei 4°C eine
Stunde lang inkubiert. Die T-Zellen, die mit den Schafserythrozyten Rosetten-ähnliche

22
Vernetzungen bildeten, konnten nun über eine weitere Zentrifugation über einen Histopaque-
Gradienten abgetrennt werden: Die oberste Phase wurde verworfen, die mittlere weißliche
Phase die nunmehr Monozyten, Granulozyten und B-Zellen enthielt wurde vorsichtig
abgenommen und nach einem Waschschritt mit PBS/EDTA weiterverarbeitet. Die unterste
Phase enthielt das Erythrozyten- Pellet mit den T-Zellen, die zur T-Zell-Isolation verwendet
werden konnten. Um etwaige Schafblutspuren aus der Monozytenfraktion zu entfernen
wurden die Zellen kurz in 8M NH4Cl gespült, was zur Lyse der Erythrozyten führte, und
anschließend in PBS/EDTA gewaschen und gezählt.
Nach Bestimmung der Zellzahl wurde der Adhärenzschritt zur Entfernung nicht plastik-
adhärenter Zellen vorbereitet. Je 30 Millionen Zellen wurden in 10ml RPMI/Genta mit 2%
autologem Plasma in einer Petrischale 60min bei 37°C und 5% CO2 inkubiert. Nach Ablauf
der Inkubationszeit wurden die nicht-adhärenten Zellen abgenommen und die
Zellkulturschalen zweimal kräftig mit PBS/EDTA gewaschen. Die verbleibende adhärente
Zellfraktion wurde nun 7 Tage lang in 8ml Differenzierungsmedium pro Schale bei 37°C und
5% CO2 kultiviert. Medienwechsel erfolgten am Tag 3 und 5 der Kultur wobei jeweils nur die
Hälfte des Mediums durch frisches Medium mit IL-4 und GM-CSF ersetzt wurde.
2.2.2.2 Zugabe der Hormone
Für den Beginn der Stimulationsversuche wurden die DC gezählt und zu je 5x105 Zellen pro
well in 24well Platten umgesetzt. In jedem well wurden 500µl altes Medium mit 500µl
frischem Medium aufgefüllt. Von der jeweiligen Hormonverdünnung wurden je 20 µl
zugegeben, so dass die gewünschte Endkonzentration im jeweiligen Ansatz vorlag. Die
Hormonstimulationsansätze wurden weiter bis zum 10. Tag kultiviert, dann wurden die Zellen
abzentrifugiert (10min bei 250xg), die Überstände aliquotiert, in flüssigem Stickstoff
schockgefroren und bis zur Untersuchung im ELISA bei -80°C gelagert.
Progesteron und 17β-Östradiol wurden jeweils in den Konzentrationen 0,1ng/ml, 10ng/ml und
1000ng/ml getestet. Darüber hinaus wurden Versuche mit Kombinationen aus 0,1ng/ml,
10ng/ml und 1000ng/ml Progesteron mit 10ng/ml 17β-Östradiol durchgeführt.
Beta-HCG wurde in Konzentrationen von 10mU/ml, 50mU/ml und 250mU/ml getestet.
Zusätzlich wurden Kombinationen aus 0,1ng/ml, 10ng/ml und 1000ng/ml Progesteron mit
50mU/ml untersucht.
Dexamethason wurde in Verdünnungen von 10-10mol/l, 10-8mol/l und 10-6mol/l getestet, die
Kontrolle mit DMSO durchgeführt.

23
Da Progesteron und 17β-Östradiol in Ethanol gelöst wurden, musste der Einfluss des
Alkohols auf die Zellen untersucht werden. Hierfür wurden alle Experimente zusätzlich zu
den Hormonansätzen auch mit Lösemittelkontrollen durchgeführt, deren
Ethanolkonzentration den in den Hormonansätzen vorliegenden Ethanolanteilen entsprach.
2.2.3 Isolation der T-Zellen
Für die MLR mussten die T-Zellen isoliert werden. Die benötigten Zellen wurden aus den mit
den Schaferythrozyten rosettierten T-Zellen gewonnen, die bei der Monozytenisolierung
zurückblieben. Die Schafserythrozyten wurden hierfür mit NH4Cl-haltigem Lysepuffer
zerstört und die Zellen sofort in PBS gewaschen um die T-Zellen nicht zu schädigen. Nach
einem zweiten Waschschritt wurden die Zellen gezählt und in Aliquots zu je 20 mio. Zellen in
FCS/DMSO eingefroren.
2.2.4 Cytokin-ELISAs
Als erster Schritt der Untersuchung der Hormonwirkungen auf DCs wurde die
Cytokinproduktion der hormonstimulierten Zellen untersucht.
Die Cytokinmessungen erfolgten in Zellkulturüberständen, die am Tag 10 der Kultur
entnommen, aliquotiert und bei –80°C eingefroren wurden. Insgesamt wurden Überstände
von 20 gesunden Blutspendern gemessen, von denen 10 männlich und 10 weiblich waren.
Gemessen wurden die Cytokinspiegel von Interleukin-12(p70), da dies die funktionelle Form
des Cytokins darstellt, Interleukin-18, Interleukin-10, Tumornekrosefaktor-alpha, und
Interleukin-6, sowie die Chemokine Interleukin-8 und macrophage derived chemokine
(MDC). Verwendet wurden OptEIA ELISA-Sets von Pharmingen, die Antikörper für MDC
und IL-18 ELISA wurden von R&D Systems bezogen. Die ELISAs wurden nach den
Herstellerangaben etabliert und optimiert.
Vorgegangen wurde nach einem einheitlichen Schema:
Das Beschichten der Platten erfolgte am Abend vor dem Versuchstag: Für eine 96well- Platte
wurden 10 ml Beschichtungspuffer mit der entsprechenden Antikörpermenge gemischt und je
100µl der Lösung pro well aufgetragen. Die Beschichtungs-Antikörper der OptEIA Sets
wurden in einer Verdünnung von 1:250 zugesetzt, die Platten mit Folie zugeklebt und über
Nacht bei 4°C inkubiert. Für den IL-18 und MDC ELISA wurden die Antikörper in der
Konzentration 2µg/ml aufgetragen und die zugeklebten Platten bei Raumtemperatur stehen

24
gelassen. Nach Beendigung der Inkubation wurde die Beschichtungslösung entfernt und die
Platten dreimal mit Waschpuffer gewaschen und auf Zellstoff trockengeklopft. Anschließend
erfolgte das 60 minütige Blocken der Platte mit 200µl Blocking-Puffer pro well. Nach drei
Waschschritten wurden je 100µl der Standards und Proben aufgetragen.
Die Ermittlung der optimalen Standardreihe erfolgte in den Optimierungsversuchen und lag
im Bereich zwischen 2000pg/ml und 0pg/ml. Entsprechend mussten die Proben je nach
erwartetem Gehalt des jeweiligen Cytokins verdünnt werden. Nach 2h Inkubation und drei
anschließenden Waschschritten wurde der Detektions-Antikörper zugegeben: Im Fall der
OptEIA- Sets wurden 100µl Antikörper plus Streptavidin- horseradish peroxidase (HRP)-
Konjugat, beide jeweils in einer 1:250 Verdünnung, zugegeben und für eine Stunde inkubiert.
Bei den IL-18 und MDC- ELISAs erfolgte die Zugabe von Antikörper und HRP- Konjugat
getrennt: Zunächst wurde der Detektions- Antikörper aufgetragen, für die MDC- ELISAs in
einer Konzentration von 200 ng/ml, für die IL-18- Bestimmung in 250 ng/ml wobei hier die
Antikörperlösung 1 Stunde zuvor mit 2% hitze-inaktiviertem Ziegenserum versetzt werden
musste. Nach 2h Inkubation bei Raumtemperatur wurde die Platte dreimal gewaschen und
anschließend 20 Minuten lang mit 100µl Streptavidin konjugierter Meerrettich- peroxidase
pro well versetzt. Die optimalen Verdünnungen für die Streptavidinlösung wurden in
Vorversuchen ermittelt und lagen bei 1: 12000 für den MDC und 1:8000 für den IL-18-
ELISA.
Um das Messergebnis sichtbar zu machen, wurde nun nach drei (bzw. sieben bei den OptEIA-
Sets) weiteren Waschschritten je 100µl Substrat zugegeben. Da das Substrat lichtempfindlich
ist, musste die nun erfolgende enzymatische Reaktion im Dunkeln stattfinden. Nach der
Unterbrechung der Reaktion, die nach 20 Minuten mit je 50µl Stoplösung durchgeführt
wurde, konnte bei 450nm die optische Dichte und damit die Cytokinmenge photometrisch
bestimmt werden.
Die erhaltenen Daten wurden für valide erachtet, wenn die photometrisch gemessenen Werte
mindestens 2 Standardabweichungen über dem Nullwert der Standardkurve lagen.
2.2.5 statistische Auswertung
Die gemesenen Daten wurden mit dem Shapiro- Wilkins-Test auf eine Normalverteilung
überprüft. Da die Messergebnisse keiner Normalverteilung entsprachen, musste die weitere
statistische Auswertung der Daten mit einem nicht- parametrischen Test für gepaarte Proben
erfolgen. Hierfür wurde der „two-tailed Wilkoxon matched pair log rank Test“ gewählt.

25
Werte mit p< 0.05 wurden als statistisch signifikant betrachtet und mit * gekennzeichnet,
Werte mit p< 0.01 wurden mit ** als hochsignifikant gekennzeichnet. Alle statistischen
Auswertungen wurden mit dem Statistikprogramm Statistica Version 5.1 durchgeführt.

26
3. Auswertung und Ergebnis
Die jeweiligen Cytokinkonzentrationen in den Überständen der Zellkulturen von unreifen
dendritischen Zellen (uDC), die von den uDC nach Zugabe der unterschiedlichen
Hormonkonzentrationen gebildet wurden, wurden mittels ELISA Technik bestimmt. Es
wurden hier folgende, von den DC typischerweise produzierte Cytokine analysiert: IL-6, IL-
10, IL-12p70, IL-18, die Chemokine IL-8 und MDC, sowie der Tumornekrosefaktor TNF-
alpha.
Aus der Gruppe der Th2-Cytokine wurde exemplarisch folgendes Mitglied analysiert:
3.1 Interleukin-10:
Dieses Interleukin der anitinflammatorischen Subklasse Th-2, wirkt auf das Immunsystem
indem es die Fähigkeit der DC hemmt, die cytotoxische CD4+-T-Zell-Antwort auszulösen.
Min-Max25%-75%Median
IL-10 unreife DC
Ver
ände
rung
des
Zyt
okin
spie
gels
(in%
)
0
20
40
60
80
100
120
140
160
180
200
220
240
260
E2
0,1
E2
10
E2
1000
P 0
,1
P 1
0
P 1
000
BH
CG
10
BH
CG
50
BH
CG
250
P+E
0,1
P+E
10
P+E
100
0
P+H
0,1
P+H
10
P+H
100
0
DE
XA
10-1
0
DE
XA
10-8
DE
XA
10-6
**
*
*
*
Abbildung 2: Box- und Whisker Plots der ELISA-Ergebnisse der IL-10 Messung
Der Messbereich liegt bei IL-10 im Intervall von 16,24 bis 125,4 pg/ml und ist somit gut zu
verwerten, da es mit der Sensitivität des IL-10-ELISAs übereinstimmt. Diese liegt im Bereich
von 7,8 pg/ml bis 1000 pg/ml.

27
Unter dem Einfluss von Dexamethason kommt es sowohl in der höchsten, als auch in der
mittleren Konzentration zu einem signifikanten Anstieg des IL-10 Spiegels. Ebenso erzielen
die Hormone Östrogen und Progesteron verschiedene Wirkungen: so ist bei Östrogen eine
spiegelsenkende Tendenz zu erkennen, die sich in der höchsten Konzentration als statistisch
signifikant erweist. Progesteron steigert den IL-10 Spiegel der unreifen DC, vor allem in der
mittleren Konzentration. Die Kombination der beiden Hormone hat keinen statistisch
signifikanten Einfluss auf den IL-10 Spiegel. Bei beta-HCG scheint jedoch eine senkende
Tendenz erkennbar zu sein, die mit zunehmender Hormonkonzentration ansteigt. Der
Kombination aus beta-HCG und Progesteron gegenüber bleibt der von den unreifen DC
produzierte IL-10 Spiegel in allen drei Konzentrationen konstant.
Aus der Gruppe der Chemokine wurden folgende Vertreter analysiert:
3.2 Interleukin-8:
Das Interleukin-8 zählt zur Klasse der Chemokine und ist durch die gerichtete Chemotaxis
besonders wichtig für eine gerichtete cytotoxische Immunantwort.
Seine Konzentration wurde im Bereich von 11,6 pg/ml bis 301,1 pg/ml gemessen. Dieses
Ergebnis liegt vollständig im Rahmen des angewendeten ELISAs, der einen Messbereich von
3,1 pg/ml bis 200 pg/ml besitzt. Somit sind die ermittelten Daten gut verwertbar.

28
Min-Max25%-75%Median
IL-8 unreife DCÄ
nder
ung
des
Zyto
kins
pieg
els
(in%
)
0
20
40
60
80
100
120
140
160
180
200
220
E2
0,1
E2
10
E2
1000
P 0
,1
P 1
0
P 1
000
P+E
2 0,
1
P+E
2 10
P+E
2 10
00
P+H
0,1
P+H
10
P+H
100
0
HC
G 1
0
HC
G 5
0
HC
G 2
50
DE
X 1
0-10
DE
X 1
0-8
DE
X 1
0-6
** **
Abbildung 3: Box- und Whisker Plots der ELISA-Ergebnisse der IL-8 Messung
Dexamethason senkt in der mittleren und höchsten Konzentration in hochsignifikanter Art
und Weise, den Cytokinspiegel von IL-8. Alle anderen Hormonzugaben lassen leider keine
eindeutig signifikanten Einflüsse auf die IL-8 Produktion der unreifen dendritischen Zellen
erkennen.
Jedoch sind einige Tendenzen durchaus ersichtlich, auch wenn diese nicht eindeutig mit
statistischer Signifikanz belegbar sind: Spiegelsteigerung bei der Kombination von
Progesteron und Östrogen; die grösste Senkung des IL-8 Spiegel findet bei jedem Hormon in
der jeweils mittleren Konzentration statt. Ebenso wäre eine Senkung des Il-8 Spiegels bei
beta- HCG in allen Konzentrationen denkbar.
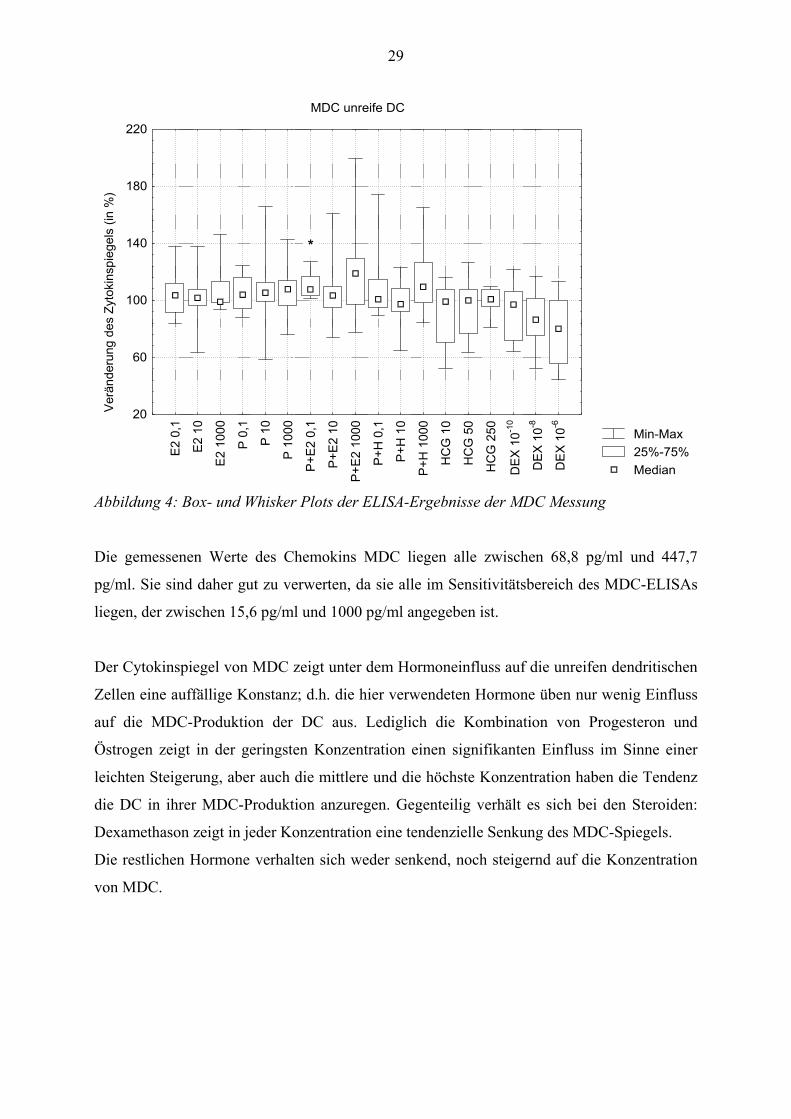
3.3 Macrophage-derived chemokine:
MDC gehört zu der Klasse der Chemokine und wird durch die schwangerschaftsassoziierten
Hormone wie folgt in seiner Ausschüttung beeinflusst:
29
Min-Max25%-75%Median
MDC unreife DCV
erän
deru
ng d
es Z
ytok
insp
iege
ls (i
n %
)
20
60
100
140
180
220
E2
0,1
E2
10
E2
1000
P 0
,1
P 1
0
P 1
000
P+E
2 0,
1
P+E
2 10
P+E
2 10
00
P+H
0,1
P+H
10
P+H
100
0
HC
G 1
0
HC
G 5
0
HC
G 2
50
DE
X 1
0-10
DE
X 1
0-8
DE
X 1
0-6
*
Abbildung 4: Box- und Whisker Plots der ELISA-Ergebnisse der MDC Messung
Die gemessenen Werte des Chemokins MDC liegen alle zwischen 68,8 pg/ml und 447,7
pg/ml. Sie sind daher gut zu verwerten, da sie alle im Sensitivitätsbereich des MDC-ELISAs
liegen, der zwischen 15,6 pg/ml und 1000 pg/ml angegeben ist.
Der Cytokinspiegel von MDC zeigt unter dem Hormoneinfluss auf die unreifen dendritischen
Zellen eine auffällige Konstanz; d.h. die hier verwendeten Hormone üben nur wenig Einfluss
auf die MDC-Produktion der DC aus. Lediglich die Kombination von Progesteron und
Östrogen zeigt in der geringsten Konzentration einen signifikanten Einfluss im Sinne einer
leichten Steigerung, aber auch die mittlere und die höchste Konzentration haben die Tendenz
die DC in ihrer MDC-Produktion anzuregen. Gegenteilig verhält es sich bei den Steroiden:
Dexamethason zeigt in jeder Konzentration eine tendenzielle Senkung des MDC-Spiegels.
Die restlichen Hormone verhalten sich weder senkend, noch steigernd auf die Konzentration
von MDC.

30
Aus der Gruppe der cytotoxischen Th1-Cytokine wurden folgende Vertreter analysiert:
3.4 Interleukin-18:
Interleuin-18 zählt zu den cytotoxischen Interleukinen der Klasse Th1 und ruft als
sogenannter Interferon-gamma induzierender Faktor, die inflammatorische Immunantwort
hervor.
Min-Max25%-75%Median
IL-18 unreife DC
Ver
ände
rung
en d
es Z
ytok
insp
iege
ls (i
n%)
0
20
40
60
80
100
120
140
160
180
200
220
E2
0,1
E2
10
E2
1000
P 0
,1
P 1
0
P 1
000
P+E
2 0,
1
P+E
2 10
P+E
2 10
00
P+H
0,1
P+H
10
P+H
100
0
HC
G 1
0
HC
G 5
0
HC
G 2
50
DE
X 1
0-10
DE
X 1
0-8
DE
X 1
0-6
**
*
Abbildung 4: Box- und Whisker Plots der ELISA-Ergebnisse der IL-18 Messung
Die Konzentrationen des Cytokins IL-18 lagen zwischen 15,94 pg/ml und 816,08 pg/ml. Sie
entsprechen somit dem Sensitivitätsbereich des IL-18 ELISAs, der zwischen 31,2 pg/ml und
4000 pg/ml liegt, und sind vollständig auswertbar.
Unter dem Einfluss der Hormonkonzentrationen ändert sich der von den dendritischen Zellen
sezernierte Cytokinspiegel von IL-18 wie folgt:
Progesteron steigert in der mittleren Konzentration den IL-18 Spiegel, senkt diesen jedoch in
Kombination mit Östrogen. Ebenfalls im Sinn einer Herabregulierung wirkt Dexamethason,
welches in der mittleren Konzentration den IL-18 Spiegel senkt. Bei beta-HCG zeigen sich
zwar keine statistisch signifikanten Ergebnisse, jedoch könnte man von einer Tendenz im
Sinne einer Steigerung der IL-18 Produktion von DC sprechen, da der Spiegel in allen drei
Konzentrationen ansteigt. Östrogen, sowie die Kombination aus Progesteron und beta-HCG,

31
zeigen ein ausgeglichenes Bild, so dass davon auszugehen ist, dass diese beiden Hormone ohne
Einfluss auf die IL-18-Produktion von dendritischen Zellen bleiben.
3.5 Interleukin-6:
Folgender Einfluss der schwangerschaftsassoziierten Hormone konnte auf die IL-6
Produktion von dendritischen Zellen herausgefunden werden:
Min-Max25%-75%Median
IL-6 unreife DC
Ver
ände
rung
en d
es Z
ytok
insp
iege
ls (i
n%)
0
20
40
60
80
100
120
140
160
180
200
220
E2
0,1
E2
10
E2
1000
P 0
,1
P 1
0
P 1
000
P+E
2 0,
1
P+E
2 10
P+E
2 10
00
P+H
0,1
P+H
10
P+H
100
0
HC
G 1
0
HC
G 5
0
HC
G 2
50
DE
X 1
0-10
DE
X 1
0-8
DE
X 1
0-6
*
*
*
*
Abbildung 5: Box- und Whisker Plots der ELISA-Ergebnisse der IL-6 Messung
Mit ermittelten Messwerten von 15,7 pg/ml bis 153,5 pg/ml liegt der IL-6 Spiegel im
Sensitivitätsbereich des IL-6 ELISAs. Dieser liegt zwischen 15 pg/ml und 1000 pg/ml. Die
Messergebnisse sind somit auswertbar. Progesteron steigert in der geringsten Konzentration
den IL-6-Spiegel von unreifen dendritischen Zellen. Die gleiche Tendenz lässt sich ebenfalls
für Östrogen feststellen, auch wenn der statistische Beweis nicht gegeben ist. Senkend auf die
IL-6 Produktion wirken sich in den höchsten Konzentrationen Dexamethason und beta-HCG
aus. Östrogen, die Kombination Östrogen mit Progesteron und beta-HCG folgen dem gleichen
Schema was die Beeinflussung des Cytokinspiegels im Zusammenhang mit der
Hormonkonzentration angeht: die niedrigsten Konzentrationen steigern die IL-6 Produktion
der unreifen DC am meisten.
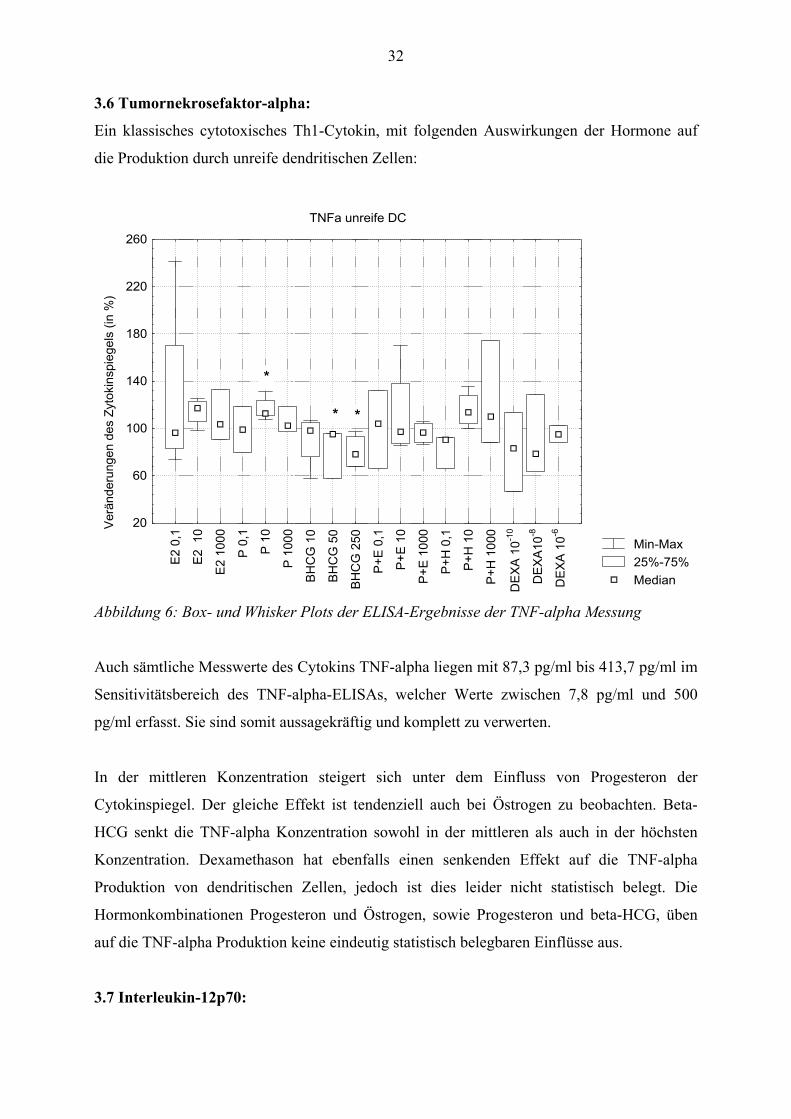
32
3.6 Tumornekrosefaktor-alpha:
Ein klassisches cytotoxisches Th1-Cytokin, mit folgenden Auswirkungen der Hormone auf
die Produktion durch unreife dendritischen Zellen:
Min-Max25%-75%Median
TNFa unreife DC
Ver
ände
rung
en d
es Z
ytok
insp
iege
ls (i
n %
)
20
60
100
140
180
220
260
E2
0,1
E2
10
E2
1000
P 0
,1
P 1
0
P 1
000
BH
CG
10
BH
CG
50
BH
CG
250
P+E
0,1
P+E
10
P+E
100
0
P+H
0,1
P+H
10
P+H
100
0
DE
XA
10-1
0
DE
XA
10-8
DE
XA
10-6
*
* *
Abbildung 6: Box- und Whisker Plots der ELISA-Ergebnisse der TNF-alpha Messung
Auch sämtliche Messwerte des Cytokins TNF-alpha liegen mit 87,3 pg/ml bis 413,7 pg/ml im
Sensitivitätsbereich des TNF-alpha-ELISAs, welcher Werte zwischen 7,8 pg/ml und 500
pg/ml erfasst. Sie sind somit aussagekräftig und komplett zu verwerten.
In der mittleren Konzentration steigert sich unter dem Einfluss von Progesteron der
Cytokinspiegel. Der gleiche Effekt ist tendenziell auch bei Östrogen zu beobachten. Beta-
HCG senkt die TNF-alpha Konzentration sowohl in der mittleren als auch in der höchsten
Konzentration. Dexamethason hat ebenfalls einen senkenden Effekt auf die TNF-alpha
Produktion von dendritischen Zellen, jedoch ist dies leider nicht statistisch belegt. Die
Hormonkombinationen Progesteron und Östrogen, sowie Progesteron und beta-HCG, üben
auf die TNF-alpha Produktion keine eindeutig statistisch belegbaren Einflüsse aus.
3.7 Interleukin-12p70:

33
Interleukin-12p70 zählt zu den proinflammatorischen Th1-Interleukinen. Seine Produktion
durch unreife dendritische Zellen, die unter dem Einfluss von schwangerschaftsassoziierten
Hormonen stehen, lässt sich leider nicht graphisch darstellen, da die ermittelten Messwerte
zwischen 2,8 pg/ml und 8,4 pg/ ml lagen. Diese Werte liegen ausserhalb des
Sensitivitätsbereiches für den IL-12-ELISA, der von 7,8 pg/ml bis 1000 pg /ml festgelegt ist.
Aufgrund dessen können die ermittelten Messergebnisse leider nicht ausgewertet werden .
3.8 Gesamtauswertung:
Insgesamt gesehen bleiben die von den dendritischen Zellen produzierten Cytokinspiegel
unter dem Einfluss von unterschiedlichen schwangerschaftsassoziierten Hormonen in
verschiedenen Konzentrationen und in unterschiedlichen Kombinationen, bemerkenswert
stabil.
Es lassen sich auch nur wenig statistisch signifikanten Ergebnisse darstellen, so dass davon
ausgegangen werden muss, dass es nicht die während der Schwangerschaft vorherrschende
Hormonlage ist, die Einfluss auf die dendritischen Zellen nimmt. Das heißt es werden die DC
nicht durch die Hormone angeregt um das Immunsystem des Menschen in der Art und Weise
zu beeinflussen, dass die Toleranz des „allogenen Transplantats“ Embryo gewährleistet ist.

34
4. Diskussion
Die Schwangerschaft stellt im Leben des Menschen einen immunologischen Balanceakt dar.
Das mütterliche Immunsystem muss tolerant sein gegenüber väterlichen Fremdantigenen auf
der Zelloberfläche des Embryos, und trotzdem muss es kompetent in der Erregerabwehr
bleiben.
Da die Plazenta während der Schwangerschaft keine unüberwindbare Barriere für kindliche
Zellen in den mütterlichen Organismus darstellt, kommt es zum Übertritt von Zellen in den
Kreislauf der Mutter. Das Ausbleiben von Angriffen gegen diese Zellen macht die
Veränderung im Immunsystem während der Zeit der Schwangerschaft deutlich und noch
immer sind diese Veränderungen, denen das menschliche Immunsystem während einer
Schwangerschaft unterliegt, wenig verstanden. Doch wie hängen die Schwangerschaft und
das Immunsystem zusammen, und wer ermöglicht diese einzigartige immunologische
Ausnahmesituation?
Die dendritischen Zellen gelten als zentrale Zellen im menschlichen Immunsystem. Sie
vermitteln zwischen dem angeborenen und adaptiven System und haben somit entscheidenden
Anteil an der Immunantwort, sei sie cytotoxisch oder toleranzauslösend. In der vorliegenden
Arbeit wurde nach dem Zusammenhang zwischen Immunsystem und Schwangerschaft
gesucht, um einen Erklärungsansatz zu finden, warum der Embryo vom maternalen
Immunsystem in der normal verlaufenden Schwangerschaft akzeptiert wird.
Bei jeder Schwangerschaft ändert sich der Hormonhaushalt der Mutter um einerseits den
Körper auf die Einnistung der befruchteten Eizelle vorzubereiten und optimale Bedingungen
für die Entwicklung des Kindes zu schaffen, und um andererseits auf diese Weise eine
Möglichkeit der Kommunikation zwischen Mutter und Kind zu ermöglichen. Es scheint daher
logisch, das Augenmerk darauf zu richten, welche Wirkung die maternale Hormonlage
während einer Schwangerschaft auf das Immunsystem ausübt.
In der vorliegenden Arbeit wurde der Einfluss von schwangerschaftsassoziierten Hormonen
auf die Cytokinproduktion unreifer dendritischer Zellen untersucht, weil das Cytokinprofil,
welches während der Schwangerschaft vorherrscht, entscheidend sein kann für den Erhalt,
bzw. den Verlust des Embryos [11].
Die unreifen dendritischen Zellen wurden hierbei aus menschlichen Blutspendeüberresten
angezüchtet und nach Anzucht und Kultur mit verschiedenen schwangerschaftsassoziierten
Hormonen in unterschiedlichen Konzentrationen versetzt. Um die immunologischen
Vorgänge im Rahmen einer Schwangerschaft zu untersuchen, wurden die in der

35
Schwangerschaft wichtigen Hormone Östrogen, Progesteron, beta- HCG und das
Corticosteroid Dexamethason eingesetzt. Dann wurden mittels ELISA-Technik die von den
unreifen dendritischen Zellen produzierten Cytokinspiegel gemessen, welche unter dem
Einfluss der Hormone entstanden sind. Dies erlaubt eine Aussage darüber, ob die unreifen
dendritischen Zellen, durch die maternale Hormonlage während der Schwangerschaft
beeinflusst werden ein bestimmtes Cytokinprofil hervorzubringen und dadurch
schwangerschaftsprotektiv, bzw. abortiv wirksam sein können.
4.1 Östrogene:
Östrogene wurden in drei unterschiedlichen Konzentrationen zu den unreifen dendritischen
Zellen gegeben: 0,1 ng/ml als physiologische Konzentration wie es im Zyklus der
Nichtschwangeren im Körper im Serum vorliegt; 10 ng/ml als Konzentration, die die
Schwangerschaft simulieren sollte und während der Schwangerschaft so vor allem im zweiten
Trimenon im Serum vorkommt; und mit 1000 ng/ml als supraphysiologische Konzentration,
wie sie im Körper normalerweise nicht vorkommt und Situationen maximaler
Östrogenausschüttung nachahmen sollte, wie es beispielsweise bei hormonproduzierenden
Tumoren oder bei einer Hormonbehandlung der Fall sein kann.
Die Cytokinproduktion von unreifen dendritischen Zellen wird durch Östrogen kaum
beeinflusst. Vor allem sind wenige statistisch signifikante Effekte zu verifizieren. Einzig auf
das Th2-Interleukin IL-10 zeigt Östrogen in der supraphysiologischen Konzentration von
1000 mg/ml eine signifikante Senkung. Da in den beiden anderen Konzentrationen jedoch
kein Effekt zu sehen ist, ist davon auszugehen, dass Östrogen die IL-10 Produktion in
physiologischen Situationen nicht beeinflusst. Es führt also nicht zu einem
schwangerschaftsprotektiven Cytokinprofil.
Betrachtet man nun die Th1-Cytokine TNF-alpha, IL-6, und Il-18, stellt man fest, dass auch
die Th1-Cytokin-Produktion der unreifen dendritischen Zellen nicht unter dem Einfluss von
Östrogen steht, da hier keine Effekte im Sinne einer Senkung oder Steigerung nachgewiesen
werden konnten. Ebenso verhält es sich mit dem Einfluss von Östrogen auf die Chemokine
MDC und Interleukin-8. Auch diese bleiben unbeeinflusst von Östrogen.
So kann man zusammenfassend sagen, dass das schwangerschaftsassoziierte Hormon
Östrogen keinen Einfluss, weder einen steigernden, noch einen hemmenden, auf die
Cytokinproduktion von humanen dendritischen Zellen hat. Es lässt sich somit keiner eindeutig
schwangerschaftsprotektiven Funktion zuordnen, was den Erhalt des Embryos aufgrund
immunologischer Akzeptanz von paternalen Fremdantigenen angeht.

36
Früheren Studien zufolgen befinden sich auf den unreifen dendritischen Zellen viele
Rezeptoren für Östrogene, durch welche unterschiedliche Effekte in den DC erzielt werden
können [45]. So inhibiert Östrogen die Antigenpräsentation der unreifen dendritischen Zellen.
Außerdem senkt Östrogen die Fähigkeit der DC naive T-Zellen zu induzieren. Auf die
Cytokine hat Östrogen folgende Wirkungen: es zeigten sich keine Effekte auf das Th2-
Cytokin IL-10, was auch dem Ergebnis der vorliegenden Arbeit entspricht. Man kann
demzufolge davon ausgehen dass die bei einer normalen Schwangerschaft erhöht gefundenen
Th2-Cytokinspiegel nicht als Folge des erhöhten Östrogenspiegels der Mutter auf dendritische
Zellen anzusehen sind.
Die Spiegel der Th1-Cytokine TNF-alpha und IL-12 fanden Liu et al. unter dem Einfluss von
Östrogenen gesenkt. Dies steht im Gegensatz zur vorliegenden Arbeit, da sich hier durch
Östrogeneinfluss kein statistisch signifikanter Effekt auf die Th1-Cytokine nachweisen lies.
Die Studienlage ist jedoch kontrovers. So konnte in anderen Studien belegt werden dass
Östrogene die Antigenpräsentation der dendritischen Zellen, und deren Migrationsfähigkeit
supprimieren und es unter dem Einfluss von Östrogen zu einer Th2-Immunantwort kommt,
Östrogene also antiinflammatorische Wirksamkeit besitzen [46]. Dies steht wiederum im
Gegensatz zu anderen Studien, in denen unter dem Einfluss von Östrogen eine eindeutig
cytotoxische Th1-Immunantwort bewiesen wurde [47].

37
4.2 Progesteron:
Progesteron wurde ebenfalls in drei unterschiedlichen Konzentrationen zu den unreifen
dendritischen Zellen gegeben: 0,1 ng/ml, die physiologische Konzentration, die sich im
Zyklusgeschehen in der follikulären Phase widerspiegelt; 10 ng/ml, was einer
Schwangerschaft entsprechen sollte, und 1000 ng/ml, welches die supraphysiologische
Konzentration darstellt, wie sie eventuell bei hormonproduzierenden Tumoren auftreten
könnte.
Von allen schwangerschaftsassoziierten Hormonen zeigte Progesteron die deutlichsten
Effekte auf die Cytokinprduktion unreifer DC, so dass dieses Hormon eine Schlüsselstellung
einzunehmen scheint. Folgende Einflüsse auf die Cytokinproduktion der unreifen
dendritischen Zellen konnten für Progesteron gezeigt werden:
In der mittleren Konzentration, d.h. mit dem physiologischen Progesteronspiegel der
Schwangerschaft, beeinflusst Progesteron alle gemessenen Cytokinspiegel der DC in
statistisch signifikanter Art und Weise. Es führt zu einer Steigerung sowohl der Konzentration
der Th1-, als auch der Th2-Cytokine. Das bedeutet dass Progesteron die unreifen
dendritischen Zellen zu einer gesteigerten Cytokinproduktion anregen kann. Allein die
Chemokine bleiben in ihrer Produktion unbeeinflusst. Es ist erwiesen dass Progesteron mit
Glucocorticoidrezeptoren kreuzreagiert und deshalb ähnliche antiinflammatorische
Wirkungen hervorruft [48]. Daher ist es kontrovers zu betrachten das Progesteron die
Produktion der inflammatorischen Cytokine in gleicher Weise steigert wie die Produktion der
anti-inflammatorischen Th2- Cytokine. Dass Progesteron eine Th2-Immunantwort fördert und
somit auch auf immunologischem Weg schwangerschaftsprotektiv wirksam ist, ist durch
Studien belegt [49]. Gleichzeitig wurde in diesen Studien auch herausgefunden dass
Progesteron durch direkte Interaktion mit den T-Zellen die cytotoxische Th1-Immunantwort
unterdrückt.
Die jeweils niedrigsten und die höchsten Progesteronkonzentrationen hatten keinen
signifikanten Einfluss auf die Cytokinproduktion. Dies könnte dadurch erklärt werden, dass
eine Einflussnahme von Progesteron auf die dendritischen Zellen am deutlichsten während
der Schwangerschaft stattfindet, jedoch in den anderen Phasen, während des normalen Zyklus
oder im supraphysiologischen Bereich, die dendritischen Zellen nicht vom Progesteron
beeinflusst werden.

38
4.3 Beta-Humanes-Chorion-Gonadotropin:
Auch beta-HCG wurde in drei verschiedenen Konzentrationen gemessen: die niedrigste
Konzentration mit 10 IE/ml, zu Beginn einer Schwangerschaft im Serum zu finden, die
mittlere Konzentration mit 50 IE/ml, die etwa der 7. Schwangerschaftswoche entspricht und
die höchste Konzentration mit 250 IE/ml, welche sich während der maximalen beta-HCG-
Ausschüttung um die 10. Schwangerschaftswoche im Serum findet.
Folgende Einflüsse von dem beta-HCG auf die Cytokinproduktion von unreifen dendritischen
Zellen wurde ermittelt: Auf die Chemokinspiegel von MDC und IL-8 hat beta- HCG keinen
Einfluss, das heisst dass deren Produktion durch dendritische Zellen nicht unter Einfluss
dieses Hormons steht. In aktuellen Studien konnte jedoch gezeigt werden dass beta-HCG
durch Aktivierung von Monozyten, also den Vorläuferzellen der unreifen DC, zu einem
Ansteigen des Chemokins IL-8 führt, dies scheint sich aber im Fall der unreifen dendritischen
Zellen in unserem Experiment nicht zu bestätigen [50].
In den mittleren und hohen Dosen senkt beta-HCG in signifikanter Art und Weise den
Cytokinspiegel der Th1-Cytokine IL-6 und TNF-alpha. Das Cytokin IL-18 bleibt jedoch
unbeeinflusst. Dies bedeutet, dass die Produktion bestimmter cytotoxischer Cytokine unter
dem Einfluss von beta-HCG gemindert wird, was sich protektiv auf das
Schwangerschaftsgeschehen auswirken kann. Dies steht im Gegensatz zu früheren Studien,
denen zufolge beta-HCG die cytotoxischen Cytokine wie IL-6 und TNF- alpha in ihrer
Produktion steigert [51].
Zusammenfassend kann festgestellt werden, dass die als potentiell schwangerschaftsprotektiv
anzusehenden Th2-Cytokine durch beta-HCG zwar nicht statistisch signifikant beeinflusst
werden, jedoch ist eine Tendenz zur Spiegelsenkung bei ansteigender Konzentration des
Hormons zu beobachten. Dies ließe sich insofern interpretieren, als dass beta-HCG die
gesamte Cytokinproduktion der dendritischen Zellen herabreguliert.
4.4 Dexamethason:
Als Coticosteroid mit der grössten Potenz, wurde Dexamethason ebenfalls in drei
verschiedenen Konzentrationen zu den unreifen dendritischen Zellen gegeben:
10-10 mol/l als niedrigste Konzentration mit der höchsten Verdünnung, 10–8 mol/l als mittlere
Konzentration und 10–6 mol/l als höchste Konzentration mit der geringsten Verdünnung.

39
Corticosteroide beeinflussen das Immunsystem auf verschiedenen Ebenen, so wird die
Cytokinproduktion von unterschiedlichen Zellen, z.B. den T-Zellen, Makrophagen,
Monozyteten oder dendritischen Zellen verändert.
Studienergebnissen zufolge supprimiert Dexamethason die Produktion der Th2-Cytokine [52].
Dies ist ein Effekt, der in der vorliegenden Arbeit ganz gegensätzlich gefunden wurde. Genau
wie bei Ramirez F. et al, 1996, fand sich in der vorliegenden Arbeit, dass Corticosteroide zu
einer Th2-Immunantwort führen [53]. Es zeigte sich dass Dexamethason in der mittleren und
der höchsten Konzentration den IL-10 Spiegel steigert. Das heisst unter dem Einfluss von
Steroiden kommt es zur Ausschüttung schwangerschaftsprotektiver Cytokine durch die
dendritischen Zellen. Im Gegensatz dazu werden die potentiell schwangerschaftsschädigenden
Th1-Cytokine in ihrer Produktion durch unreife dendritische Zellen, unter dem Einfluss von
Dexamethason gehemmt.
Statistisch signifikante Effekte zeigen sich besonders deutlich bei IL-18 in der mittleren
Konzentration, sowie bei IL-6 in der mittleren und höchsten Konzentration. Bei TNF-alpha
war ebenfalls eine Tendenz zur Spiegelsenkung zu erkennen, so dass man davon ausgehen
kann dass Dexamethsaon die Produktion der cytotoxischen Cytokine während der
Schwangerschaft hemmt. Gleiches konnte schon durch verschiedenen Studien belegt werden,
so fanden zum Beispiel Viera et al., 1998, das Glucocorticoide die Produktion cytotoxischer
Cytokine durch humane dendritische Zellen hemmen, und Piemonti und Kollegen wussten
schon 1999, dass die dendritischen Zellen in ihrer Differenzierung und Reifung durch
Glucocorticoide beeinflusst werden [8, 54].
Das Ergebnis der vorliegenden Arbeit entspricht auch den Studien von de Jong et al., 1999,
die herausfanden dass unreife dendritische Zellen zu einer Hemmung der Th1-
Cytokinprodukion führen und im Gegensatz dazu die Th-2 Cytokine vermehrt von den
unreifen DC sezerniert werden [55]. Auch die Reifung der DC wird von Corticosteroiden
blockiert. Ohne Einfluss bleiben die Steroide laut de Jong, auf die Antigenaufnahme und die
Expression kostimulatorischer Moleküle wie CD 40-Ligand oder MHC-II-Produktion. Dies
steht im Gegensatz zu Pan J. et al., 2001, welcher Beweise erbrachte, dass Dexamethason die
Expression kostimulatorischer Moleküle inhibiert, und auf diese Weise die Aktivierung von
T-Zellen durch dendritische Zellen verhindert wird [56].
Werden die unreifen dendritischen Zellen mit Dexamethason inkubiert, kommt es zu einer
Herabregulation der kostimulatorischen Merkmale wie den MHC II-Rezeptoren und dem B7-
Ligand. Ausserdem sind die unreifen DC nach einer Dexamethasoninkubation refraktär in
ihrer Reifung. Insgesamt konnten wir zeigen dass die unreifen dendritischen Zellen unter dem

40
Einfluss von Glucocorticoiden in der Produktion der cytotoxischen Th1-Cytokine unterdrückt
werden. Die schwangerschaftsprotektiven, antiinflammatorischen Th2-Cytokine werden in
ihrer Produktion gesteigert.
4.5 Progesteron und Östrogen:
Da Hormone in vivo nie einzeln auftreten, sondern immer in Kombination auf den Körper
einwirken, wurden verschiedene Kombinationen von Hormonen ausgetestet und deren
Einfluss auf die Cytokinproduktion der unreifen dendritischen Zellen untersucht, um die in
vitro Bedingungen den Bedingungen im Körper anzupassen. Ziel war es hierbei
herauszufinden, ob es zu einer eventuellen Summation der Einzeleffekte der getesteten
Hormone kommen kann und ob sich diese Effekte durch Kombination sinnvoll nutzen lassen.
So wurden zuerst die beiden weiblichen Sexualhormone kombiniert, die den Zyklus der Frau
im Wesentlichen mitbestimmen und die während einer Schwangerschaft dominieren. Wieder
wurden drei unterschiedliche Konzentrationen ausgetestet:
Die geringste Konzentration mit Progesteron 1000 mg/ml und Östrogen 0,1mg/ml, was
annähernd der Follikelphase des weiblichen Zyklus entspricht, die mittlere Konzentration mit
Progesteron 1000 mg/ml und Östrogen 10 mg/ml, die sich während der lutealen Zyklusphase
findet, und die höchste, die supraphysiologische Konzentration mit Progesteron 1000 mg/ml
und Östrogen 1000 mg/ml.
Die Kombination der beiden Hormone lies in der mittleren Konzentration eine Tendenz zur
Steigerung des IL-10-Spiegels erkennen, das heisst das Zusammenspiel von Progesteron und
Östrogen ruft ein schwangerschaftsprotektives Cytokinmuster durch dendritische Zellen
hervor. Ebenfalls positiv auf den Verlauf der Schwangerschaft würde sich auswirken dass IL-
18, ein Th1-Cytokin, in der mittleren Konzentration gesenkt wird. Auf die anderen
gemessenen Th1-Cytokine zeigten sich keine eindeutigen Effekte. Und auch die niedrigste
oder die höchste Konzentration der Hormonkombination brachte keine statistisch
signifikanten Ergebnisse. Bei den Chemokinen war auffällig dass es unter dem Einfluss von
Progesteron und Östrogen zu einer signifikanten Spiegelsteigerung von MDC in der
niedrigsten Konzentration kommt. Da MDC zu einer Th2-Immunantwort führt, könnte man
dies als positiv für die Schwangerschaft werten.
Insgesamt ist festzustellen, dass sich direkte summatorische Effekte der Einzelfunktionen der
beiden Hormone Progesteron und Östrogen nicht nachweisen lassen.

41
4.6 Progesteron und beta-HCG:
Auch diese Hormonkombination wurde in drei unterschiedlichen Konzentrationen zu den
unreifen dendritischen Zellen gegeben, um deren Verhalten während unterschiedlicher
hormoneller Stimulation zu untersuchen. Wie in den Versuchsansätzen vorher wurde versucht
mit den beiden unteren Konzentrationen physiologische Situationen im Körper zu imitieren,
mit der höchsten Konzentration jedoch eine Situation zu erreichen, wie sie im Körper
normalerweise nicht vorkommt, um Aussagen über die DC-Funktion unter maximaler
Hormonstimulation zu treffen. Die niedrigste Konzentration lag bei Progesteron 1000 mg/ml
und beta-HCG 10 IE/ml; die mittlere bei Progesteron 1000 mg/ml und beta-HCG 50 IE/ml,
und die höchste, supraphysiologische Konzentration lag bei Progesteron 1000 mg/ml und
beta-HCG 250 IE/ml.
Die Kombination dieser beiden Hormone lässt leider nur wenige Aussagen über die
Cytokinproduktion der unreifen dendritischen Zellen zu. Es stehen in keiner Konzentration
statistisch signifikante Ergebnisse zur Verfügung. Die Effekte der aus den Einzelmessungen
gewonnenen Erkenntnisse konnten auch hier nicht summiert werden. Man muss in diesem
Fall annehmen dass die Cytokinproduktion der unreifen DC nicht durch die
Hormonkombination von Progesteron mit beta-HCG beeinflusst wird und man keine
Aussagen bezüglich der Wirkung auf die Schwangerschaft treffen kann.
Es lassen sich insgesamt nur wenige statistisch signifikante Einflüsse speziell der
schwangerschaftsassoziierten Hormone auf die Cytokinproduktion der unreifen dendritischen
Zellen feststellen. Man kann somit festhalten dass es nicht die mütterlichen Hormone sind, die
im Verlauf der Schwangerschaft dazu führen, dass ein bestimmtes, förderliches Cytokinprofil,
nämlich das der Th2-Cytokine, vorliegt. Sondern dass es von bisher noch unbekannten
Mechanismen abhängt, dass diese Klasse der Cytokine von den DC produziert werden und die
Schwangerschaft ungestört verlaufen kann.
Alles in allem handelte es sich natürlich um einen Versuch mit hohem experimentellen
Aufwand mit einer Probandenzahl von n=20. Obwohl dies in einer Versuchsanordnung auch
für die statistische Aussagekraft eine ausreichende Testanzahl darstellt, wären natürlich
Versuche mit mehr Patientenproben wünschenswert. So könnte man konkretere Aussagen
über die in dieser Arbeit sichtbaren Tendenzen treffen. Auch könnte man verschiedenen

42
unvermeidbaren Ereignissen besser begegnen, wie zum Beispiel der hohen biologischen
Variabilität aufgrund der Verwendung biologischen Materials.
Die in der Arbeit verwendeten Zellen stammten von gesunden Blutspendern, wobei 10
männliche und 10 weibliche Proben weiterverarbeitet wurden. Hierbei blieben auch viele
Aspekte unklar, die im Zusammenhang mit der Hormonlage hätten wichtig sein können.
Beispielsweise wurden keine Angaben über das Alter der Blutspender gemacht, ob sich
Frauen in der Menopause unter den Spendern befanden. Auch könnte z.B. die Einnahme
oraler Kontrazeptiva, oder die Zyklusphase der gebärfähigen Frau, über die man keine
Aussage treffen kann, die Zellen beeinflussen.
Letztlich bleibt es weiterhin unklar was die genauen Ursachen für die Spiegelerhöhung der
Th2-Cytokine während einer normalverlaufenden Schwangerschaft sind. Man kann jedoch
davon ausgehen dass es nicht alleine die schwangerschaftsoziierten Hormone der Mutter sind,
die zu dem Th2-gewichteten Cytokinprofil führen, bzw. dass die dendritischen Zellen ihr
Verhaltensmuster im Hinblick auf die hormonelle Beeinflussung nur in begrenztem Ausmaß
ändern.

43
5. Zusammenfassung
Die Schwangerschaft ist aus immunologischer Sicht ein Phänomen: trotz der fremden,
väterlichen Antigene, die der Embryo auf seinen Zellen exponiert, wird er von Immunsystem
der Mutter akzeptiert, und im Normalfall nicht abgestossen. Die Frage nach der Ursache
dieses Phänomens beschäftigt die Wissenschaft seit Jahren.
Die Schwangerschaft geht jedoch nicht nur mit einer Veränderung des Immunsystems einher,
sondern es müssen sich viele Systeme im Organismus der Mutter umstellen, damit das Leben
und Überleben des Kindes gesichert werden kann. Ein wichtiges Beispiel stellt der
Hormonhaushalt der Mutter dar, der sich bei jeder normalverlaufenden und somit
erfolgreichen Schwangerschaft stets in gleicher Weise ändert. So stellt sich die Frage ob das
mütterliche Immunsystem und die veränderte Hormonlage zusammenhängen, und wenn ja,
auf welche Art und Weise?
Hierzu ist es wichtig das Immunsystem näher zu betrachten. Schnell wird man auf die
aussergewöhnlichen dendritischen Zellen kommen, die ubiquitär im menschlichen Körper
verbreitet sind. Diese Zellen, zurecht als herausragende Immunzellen des Menschen
bezeichnet, dirigieren durch die von ihnen produzierten Cytokine die Immunantwort. Sie sind
fähig durch Interaktion mit den T-Lymphozyten die Reaktion des Körpers in eine
cytotoxische, bzw. tolerogene Richtung zu lenken.
Man hat herausgefunden, dass bei einer Schwangerschaft das umgebende Cytokinprofil von
immenser Bedeutung für deren Verlauf ist. So kann es bei vorliegen von Th1-Cytokinen zu
einem Abortgeschehen kommen, bei überwiegen der Th2-Cytokine im mütterlichen Milieu,
ist von einer normalverlaufenden Schwangerschaft auszugehen. Ausgehend von diesen
Kenntnissen wurde in der vorliegenden Arbeit untersucht, wie die maternale Hormonlage die
Cytokinproduktion der unreifen dendritischen Zellen beeinflusst. Hierzu wurden aus den
Buffycoats von 20 menschlichen Blutspenden gemäß dem Standartprotokoll von Romani et
al., die PBMC, die peripheren Monocyten im Blut isoliert. In der Zellkultur konnten aus
diesen Zellen dendritischen Zellen angezüchtet werden.
Die DC wurden dann mit den schwangerschaftsassoziierten Hormonen Östrogen, Progesteron,
beta-HCG und Dexamethason versetzt, wobei man durch die Gabe unterschiedlicher
Konzentrationen, Situationen im Schwangerschaftsverlauf simulieren wollte. Dann wurden
die dendritischen Zellen zwei weitere Tage zusammen mit den Hormonen kultiviert. Mittels
ELISA-Technik wurde abschließend die Cytokinproduktion der dendritischen Zellen im
Kulturüberstand bestimmt, um eine Aussage über das Cytokinprofil dieser wichtigen

44
menschlichen Immunzellen unter dem Einfluss der schwangerschaftsasoziierten Hormone
treffen zu können. Die Daten wurden mit einem nicht-parametrischen Test für gepaarte
Proben statistisch ausgewertet.
Als Ergebnis dieser Arbeit liegt vor, dass es keinen eindeutig statistisch belegbaren
Zusammenhang zwischen der Cytokinproduktion der dendritischen Zellen und den
schwangerschaftsassoziierten Hormonen gibt; d. h. man kann den maternalen Hormonen
keine direkte immunologisch-protektive Funktion hinsichtlich einer Beeinflussung
dendritischer Zellen zuordnen.
Das heißt aber auch dass das Geheimnis der Toleranz des Embryos weiterhin bestehen bleibt
und auch in Zukunft die Forschung beschäftigen wird.

45
6. Literaturverzeichnis
[1] Medawar P.B.: Some immunological and endocrinological problems raised by
evolution of viviparity in vertebrates. Society for Exp Biol, Evolution Symposia 11.
Academic Press, New York, 1953, 320-338.
[2] Herz U., Renz H.: Fetomaternale Immunität - Neue Konzepte zur Plastizität des
Immunsystems während der Schwangerschaft. Gynäkologe, 2001, 34: 494-502.
[3] Duc-Goiran P., Mignot T.M., et al.: Embryo-maternal interactions at the implantation
site: a delicate equilibrium. Europ. J. Obstetr. Gynecol. Reprod. Biol., 1999, 83: 85-
100.
[4] Forth, Henschler, Rummel: allgemeine und spezielle Pharmakologie und Toxikologie,
8.Auflage, 2001.
[5] Stauber M., Weyerstahl T.: Gynäkologie und Geburtshilfe, 2001 Georg-Thieme
Verlag, S. 90, 460 f., 87, 461, 509.
[6] Keck C., Neulen J., Behre H., Breckwoldt M.: Endokrinologie, Reproduktionsmedizin,
Andrologie, 2.Auflage, 2002 Georg-Thieme Verlag, S. 31-41.
[7] Pfleiderer A., Beckwoldt M., Martius G.: Gynäkologie und Geburtshilfe, 4. Auflage
2001, Georg-Thieme Verlag, S. 49 f., 56 f., 269.
[8] Vieira P., Kalinski P., Wierenga E., Kapsenberg M., de Jong E.: Glucocorticoids
inhibit bioactive IL-12p70 production by in vitro-generated human dendritic cells
without affecting their t- cell stimulatory potential. J. Immunol., 1998, 161: 5245-
5251.
[9] Kim K., Choe Y., Choe I., Lim J.: Inhibition of glucocorticoid-mediated, caspase-
independent dendritic cell death by CD 40 activation.
J. Leuk. Biol., 2001, 69(3): 426-34.
[10] Gerhard I., Runnebaum B.: Endokrinologie in der Schwangerschaft
[11] Raghupathy R.: Th1-type immunity is incompatile with successful pregnancy
Immunol. Today, 1997, 10: 478-482.
[12] Renz-Polster H., Braun J.: Kap. 4; Immunologie. Basislehrbuch Innere Medizin, 2000,
2.Auflage, Urban&Fischer Verlag
[13] Niederle N., Bergmann L., Ganser A.: Zytokine. Gustav-Fischer Verlag, 1996.
[14] Sharkey, A.: Cytokines and embryo/endometrial interactions. Reprod. Med. Rev.,
1995, 4: 87-100.
[15] Kishimoto, T.: The biology of interleukin-6. Blood, 74: 1, 1998.

46
[16] Vincent T., Mechti N.: IL-6 regulates CD44 cell surface expression on human
myeloma cells. Leukemia, 2004, 18(5): 967-75.
[17] McInnes I.B., Gracie J., Leung B., Wei X., Liew F.: Interleukin-18: a pleiotropic
participant in chronic inflammation. Trends Immunol. Today, 2001, 21(7): 312-5.
[18] Kruse N., Greif M., Moriabadi F., Marx L., Toyka K., Rieckmann P.: Variations in
cytokine mRNA expression during normal human pregnancy. Clin. Exp. Immunol.,
2000, 119:317-322.
[19] Stoll S., Jonuleit H., Schmitt E., Müller G., Yamauchi H., Kurimoto M., Knopp J., Enk
A.: Production of functional IL-18 different subtypes of murine and human dendritic
cells: DC-derived IL-18 enhances IL-12-dependent Th1 development. Eur. J.
Immunol., 1998, 28: 3231-3239.
[20] Skok J., Poudrier J., Gray D.: Dendritic cell-derived IL-12 promotes B cell induction
of Th2 differentiation: A feedback regulation of Th-1 development.
J. Immunol. 1999; 196:4284-4291.
[21] Berthier R., Rizitelli A., Martinon-Ego C, Laharie A., CollinV., Chesne S., Marche P.:
Fibroblasts inhibit the production of IL-12p70 by murine dendritic cells. Immunology,
2003, 108 (3): 391.
[22] Heystek H., den Drijver B., Kapsenberg M., von Lier R., de Jong E.: Type I IFNs
modulate IL-12p70 Production by human dendritic cells depending on the maturation
status of the cells and counteract IFN-gamma-mediated signalling. Clin. Immunol.,
2003, 107 (3): 170-7.
[23] Moser M., Murphy K.: Dendritic cell regulation of Th1-Th2-development. Nature
Immunol., 2000, 1(3): 199-205.
[24] Watford W., Moriguchi M., Morinobu A., O`Shea J.: The biology of Il-12:
coordinating innate and adaptive immune responses. Cytokine Growthfactor Rev.,
2003; 14 (5): 361-368.
[25] Beutler B.: The role of tumor necrosis factor in health and disease. J. Rheumatol.
1999; 26: 16-2.
[26] Rieser C., Bock G., Klocker H., Bartsch G., Thurnher M.: Prostaglandin E2 and tumor
necrosis factor alpha cooperate to activate human dendritic cells: Synergistic
activation of interleukin-12 production. J. Exp. Med. 1997; 186:1603-08.
[27] Balkwill F.: Tumor necrosis factor or tumor promoting factor? Cytokine &Growth
Factor Rev., 2002; 13:135-141.

47
[28] Sallusto F., Lanzavecchia A.: Efficient presentation of soluble antigen by cultured
human dendritic cells is maintained by granulocyte/macrophage colony-stimulating
factor plus interleukin-4 and downregulated by tumor necrosis factor alpha. J. Exp.
Med., 1994, 179: 1109-1118.
[29] Mosmann T.: Properties and functions of interleukin-10. Adv. Immunol., 1994, 56: 1-
26.
[30] Steinbrink K., Wölfl M., Jonuleit H., Knop J., Enk A.: Induction of tolerance by IL-10
treated Dendritic cells. J. Immunol., 1997, 159: 4772-4780.
[31] Jonuleit H., Knop J., Enk A.: Cytokines and their effects on maturation, differentiation
and migration of dendritic cells. Arch. Dermatol. Res., 1996, 289: 1-8.
[32] Robertson S., Seamark R., Guilbert L., Wegman T.: The role of cytokines in gestation.
Crit. Rev. Immunol., 1994, 14: 239-292.
[33] Janeway C., Travers P., Walpert M., Shlamichik M.: Immunologie. 5. Auflage, 2002,
Spektrumverlag.
[34] Godiska R., Chantry D., Raport C., Sozzani S., Allavena P., Leviten D., Mantovani A.,
Gray P.: Human Macrophage-derived Chemokine (MDC), a Novel Chemoattractant
for Monocytes, Monocyte-derived Dendritic Cells, and Natural Killer Cells. J. Exp.
Med., 1997, 185 (9): 1595-1604.
[35] Mantovani A., Gray P., Damme J., Sozzani S.: Macorphage-derived Chemokine (
MDC). J. Leuk. Biol., 2000, 68: 400-404.
[36] Hill J.A.: Cellular and Molecular Biology of the Maternal-Fetal Relationship.
INSERM/ John Libbey Eurotext, 1991, 269-27.
[37] Steinman R., Cohn Z.: Identification of a novel cell type in peripheral lymphoid
organs of mice. J. Exp. Med., 1973, 137: 1142-62.
[38] Hart D.: Dendritic cells: Unique Leukocyte Populations Which Control the Primary
Immune Response. Blood, 1997, 90(9): 3245-3287.
[39] Everson M., McDuffie D., Lemark D., Koopman W., McGhee J., Beagley K.:
Dendritic cells from different tissues induce production of different T cell cytokine
profiles. J. Leuk. Biol., 1996, 59: 494-498.
[40] Kämpgen E.: dendritische Zellen – „Zündfunken“ der Immunantwort. Blick,
Universität Würzburg, 1997 (1): 67-70.
[41] Satthaporn S., Eremin O.: Dendritic cells: biological functions. J.R. Coll. Surg. Edinb.,
2001, 46: 9-20.

48
[42] Banchereau J., Steinman R.: Dendritic cells and the control of immunity. Nature,
1998, 392: 245-252.
[43] Steinman R.: The dendritic cell system and its role in immunogenicity. Annu. Rec.
Immunol., 1991, 9: 271-96.
[44] Wegmann T., Lin H., Guilbert L., Mosmann T.: Bidirectional cytokine interactions in
the maternal-fetal relationship: is successful pregnancy aTH2 phenomenon? Immunol.
Today, 1993, 14: 353-356.
[45] Liu H., Buenafe A., Matejuk A., Ito A., Zamora A., Dwyer J., Vandenbark A., Offner
H.: Estrogen inhibition of EAE involves effects on dendritic cell function. J. Neurosci.
Res., 2002, 70: 238-248.
[46] Beagly K., Gockel C.: Regulation of innate and adaptive immunity by the female sex
hormones oestradiol and progesterone. FEMS Immunol. Med. Microbiol, 2003, 38:
13-22.
[47] Maret A.: Estradiol enhances primary antigen-specific CD 4 T cell responses and Th1
development in vivo. Essential role of estrogen receptor alpha expression in
hematopoietic cells. Eur. J. Immunol., 2003, 33: 512-521.
[48] Kontula K., Andersson L., Huttunen M., Pelkonen R.: Binding of progestins to the
glucocorticoid receptor. Correlation to their glucocorticiod-like effects on in vitro
functions of human mononuclear leukocytes. Biochem. Pharmacol., 1983, 32(9):
1511-18.
[49] Miyaura H., Iwata M: Direct and indirect inhibition of Th1 development by
progesterone and glucocorticoids. J. Immunol., 2002,168:1086-1094
[50] Kosaka K., Fujiwara H., Tatsumi K., Yoshioka S., Sato Y., Egawa H., Higuchi T.,
Nakayama T., Ueda M., Maeda M., Fuji S.: Human chorionic gonadotropin (HCG)
activates monocytes to produce Interleukin-8 via a different pathway from luteinizing
hormone/HCG receptor system. J. Clin. Endocrinol. & Metabol., 2002, 87(11): 5199-
5208.
[51] Schäfer A., Pauli G., Friedmann W., Dudenhausen J.: Human chorionic gonadotropin
(HCG) and placental lactogen (hPL) inhibit interleukin-2 (IL-2) and increase
interleukin-1 (IL-1), -6 and tumor necrosis factor (TNF) expression in monocyte cell
cultures. J. Perinat. Med., 1992, 20: 233-240.
[52] Fushimi T., Okayama H., Seki T., Shimura S., Shirato K.: Dexamethasone suppressed
gene expression and production of IL-10 by human peripheral blood mononuclear
cells and monocytes. Int. Arch. Allergy. Immunol., 1997, 112: 13-18.

49
[53] Ramirez F., Fowell D., Puklavec M., Simmonds S., Mason D: Glucocorticoids
promote a Th2 cytokine response by CD4+ cells in vitro. J. Immunol., 1996, 156:
2406-2412.
[54] Piemonti L., Monti P., Allavena P., Sironi M., Soldini L., Leone B., Socci C., Di Carlo
V.: Glucocorticoids affect humen dendritic cell differentiation and maturation. J.
Immunol., 1999, 162: 6473-6481.
[55] De Jong E., Viera P., Kalinski P., Kapsenberg M.: Corticosteroids inhibit the
production of inflammatory mediators in immature monocyte-derived DC and induce
the development of tolerogenic DC3. J. Leuk. Biol., 1999, 66: 201-204.
[56] Pan J., Ju D., Wang Q., Zhang M., Xia D., Zhang L., Yu H., Cao X.: Dexamethasone
inhibits the antigen presentation of dendritic cells in MHC class pathway. Immunol.
Lett., 2001, 76 (3): 153-61.
[57] Miller-Lindholm A., LaBenz C., Ramey J., Bedows E., Ruddon R.: Human Chorionic
Gonadotropin-beta Gene Expression in first trimester Placenta.
Endocrinology, 1997, 138: 5459-5465.
[58] Romani N., Reider D., Heuer M., Ebner S., Kämpgen E., Bock G., Klockner H.,
Bartsch G.: Generation of mature dendritic cells from human blood. An improved
method with special regard to clinic applicability.
J. Immunol. Met., 1996, 196: 137-151.