metabolismo de los glúcidos
DESCRIPTION
Ciclo de Krebs, Glucólisis, Gluconeogénesis, Glucógenolisis, Glucogenogenésis, Vía de las Pentosas (Fosfato),TRANSCRIPT

PDF generated using the open source mwlib toolkit. See http://code.pediapress.com/ for more information.PDF generated at: Sun, 03 Jun 2012 06:21:59 UTC
Metabolismo GlucidosERA 2

ContentsArticles
Ciclo de Krebs 1Glucólisis 5Gluconeogénesis 14Glucogenolisis 18Glucogénesis 18Ruta de la pentosa fosfato 19
ReferencesArticle Sources and Contributors 24Image Sources, Licenses and Contributors 25
Article LicensesLicencia 26

Ciclo de Krebs 1
Ciclo de Krebs
Esquema didáctico del ciclo del ácido cítrico.
El ciclo de Krebs (también llamadociclo del ácido cítrico o ciclo de losácidos tricarboxílicos) es una rutametabólica, es decir, una sucesión dereacciones químicas, que forma partede la respiración celular en todas lascélulas aeróbicas. En células eucariotasse realiza en la mitocondria. En lasprocariotas, el ciclo de Krebs se realizaen el citoplasma, específicamente en elcitosol.
En organismos aeróbicos, el ciclo deKrebs es parte de la vía catabólica querealiza la oxidación de glúcidos, ácidosgrasos y aminoácidos hasta producirCO2, liberando energía en formautilizable (poder reductor y GTP).
El metabolismo oxidativo de glúcidos, grasas y proteínas frecuentemente se divide en tres etapas, de las cuales, elciclo de Krebs supone la segunda. En la primera etapa, los carbonos de estas macromoléculas dan lugar a moléculasde acetil-CoA de dos carbonos, e incluye las vías catabólicas de aminoácidos (p. ej. desaminación oxidativa), la betaoxidación de ácidos grasos y la glucólisis. La tercera etapa es la fosforilación oxidativa, en la cual el poder reductor(NADH y FADH2) generado se emplea para la síntesis de ATP según la teoría del acomplamiento quimiosmótico.
El ciclo de Krebs también proporciona precursores para muchas biomoléculas, como ciertos aminoácidos. Por ello seconsidera una vía anfibólica, es decir, catabólica y anabólica al mismo tiempo.El Ciclo de Krebs fue descubierto el por el alemán Hans Adolf Krebs, quien obtuvo el Premio Nobel.
Ciclo de Krebs 2
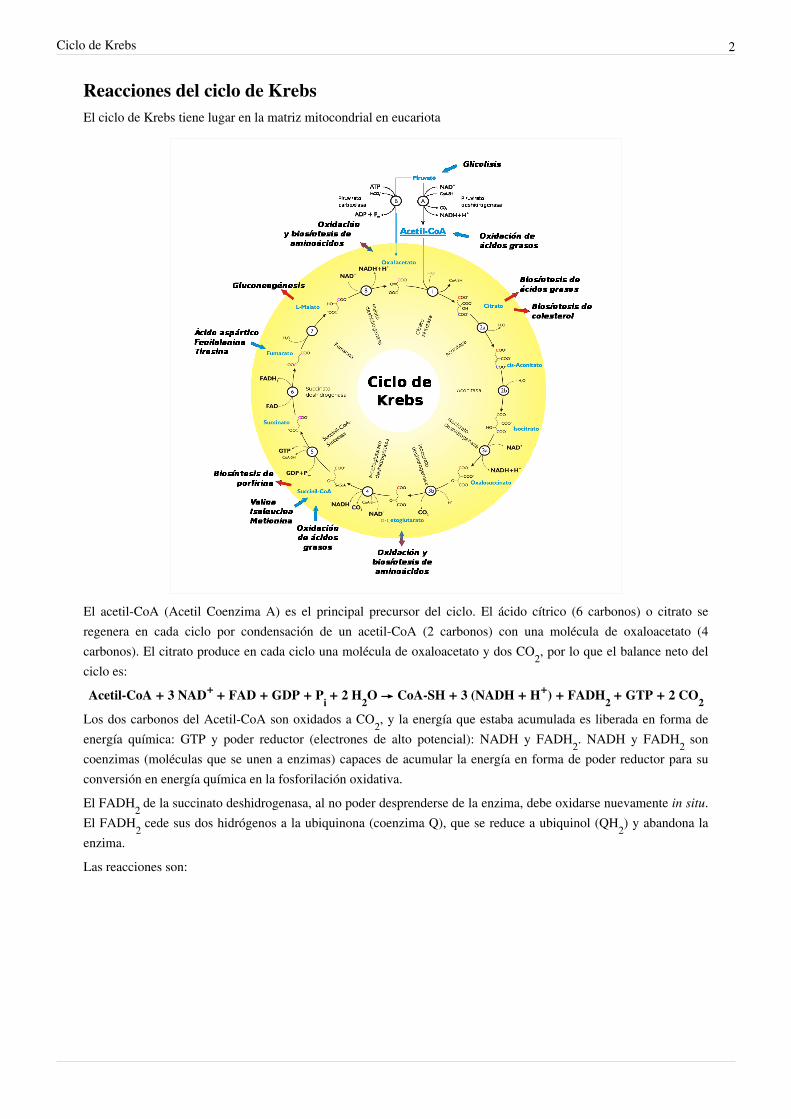
Reacciones del ciclo de KrebsEl ciclo de Krebs tiene lugar en la matriz mitocondrial en eucariota
El acetil-CoA (Acetil Coenzima A) es el principal precursor del ciclo. El ácido cítrico (6 carbonos) o citrato seregenera en cada ciclo por condensación de un acetil-CoA (2 carbonos) con una molécula de oxaloacetato (4carbonos). El citrato produce en cada ciclo una molécula de oxaloacetato y dos CO2, por lo que el balance neto delciclo es:Acetil-CoA + 3 NAD+ + FAD + GDP + P
i + 2 H
2O → CoA-SH + 3 (NADH + H+) + FADH
2 + GTP + 2 CO
2
Los dos carbonos del Acetil-CoA son oxidados a CO2, y la energía que estaba acumulada es liberada en forma deenergía química: GTP y poder reductor (electrones de alto potencial): NADH y FADH2. NADH y FADH2 soncoenzimas (moléculas que se unen a enzimas) capaces de acumular la energía en forma de poder reductor para suconversión en energía química en la fosforilación oxidativa.El FADH2 de la succinato deshidrogenasa, al no poder desprenderse de la enzima, debe oxidarse nuevamente in situ.El FADH2 cede sus dos hidrógenos a la ubiquinona (coenzima Q), que se reduce a ubiquinol (QH2) y abandona laenzima.Las reacciones son:
Ciclo de Krebs 3
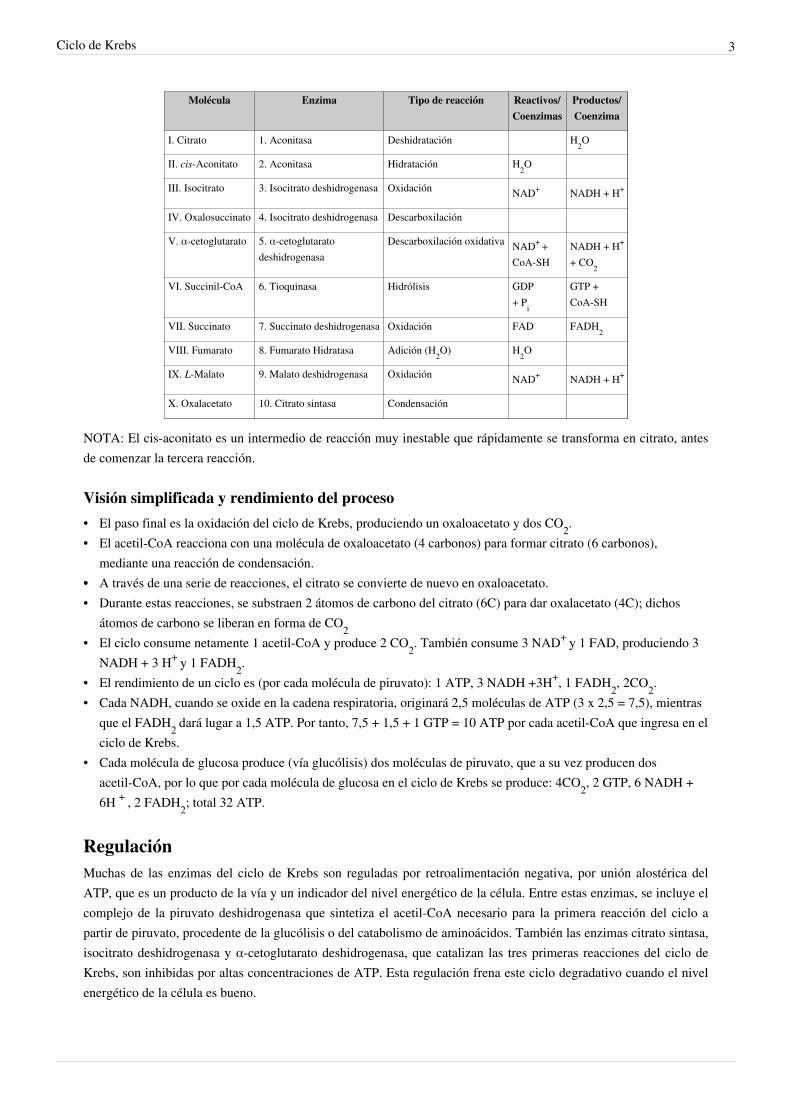
Molécula Enzima Tipo de reacción Reactivos/Coenzimas
Productos/Coenzima
I. Citrato 1. Aconitasa Deshidratación H2O
II. cis-Aconitato 2. Aconitasa Hidratación H2O
III. Isocitrato 3. Isocitrato deshidrogenasa Oxidación NAD+ NADH + H+
IV. Oxalosuccinato 4. Isocitrato deshidrogenasa Descarboxilación
V. α-cetoglutarato 5. α-cetoglutaratodeshidrogenasa
Descarboxilación oxidativa NAD+ +CoA-SH
NADH + H+
+ CO2
VI. Succinil-CoA 6. Tioquinasa Hidrólisis GDP+ Pi
GTP +CoA-SH
VII. Succinato 7. Succinato deshidrogenasa Oxidación FAD FADH2
VIII. Fumarato 8. Fumarato Hidratasa Adición (H2O) H2O
IX. L-Malato 9. Malato deshidrogenasa Oxidación NAD+ NADH + H+
X. Oxalacetato 10. Citrato sintasa Condensación
NOTA: El cis-aconitato es un intermedio de reacción muy inestable que rápidamente se transforma en citrato, antesde comenzar la tercera reacción.
Visión simplificada y rendimiento del proceso• El paso final es la oxidación del ciclo de Krebs, produciendo un oxaloacetato y dos CO2.• El acetil-CoA reacciona con una molécula de oxaloacetato (4 carbonos) para formar citrato (6 carbonos),
mediante una reacción de condensación.•• A través de una serie de reacciones, el citrato se convierte de nuevo en oxaloacetato.• Durante estas reacciones, se substraen 2 átomos de carbono del citrato (6C) para dar oxalacetato (4C); dichos
átomos de carbono se liberan en forma de CO2• El ciclo consume netamente 1 acetil-CoA y produce 2 CO2. También consume 3 NAD+ y 1 FAD, produciendo 3
NADH + 3 H+ y 1 FADH2.• El rendimiento de un ciclo es (por cada molécula de piruvato): 1 ATP, 3 NADH +3H+, 1 FADH2, 2CO2.• Cada NADH, cuando se oxide en la cadena respiratoria, originará 2,5 moléculas de ATP (3 x 2,5 = 7,5), mientras
que el FADH2 dará lugar a 1,5 ATP. Por tanto, 7,5 + 1,5 + 1 GTP = 10 ATP por cada acetil-CoA que ingresa en elciclo de Krebs.
• Cada molécula de glucosa produce (vía glucólisis) dos moléculas de piruvato, que a su vez producen dosacetil-CoA, por lo que por cada molécula de glucosa en el ciclo de Krebs se produce: 4CO2, 2 GTP, 6 NADH +6H + , 2 FADH2; total 32 ATP.
RegulaciónMuchas de las enzimas del ciclo de Krebs son reguladas por retroalimentación negativa, por unión alostérica delATP, que es un producto de la vía y un indicador del nivel energético de la célula. Entre estas enzimas, se incluye elcomplejo de la piruvato deshidrogenasa que sintetiza el acetil-CoA necesario para la primera reacción del ciclo apartir de piruvato, procedente de la glucólisis o del catabolismo de aminoácidos. También las enzimas citrato sintasa,isocitrato deshidrogenasa y α-cetoglutarato deshidrogenasa, que catalizan las tres primeras reacciones del ciclo deKrebs, son inhibidas por altas concentraciones de ATP. Esta regulación frena este ciclo degradativo cuando el nivelenergético de la célula es bueno.

Ciclo de Krebs 4
Algunas enzimas son también reguladas negativamente cuando el nivel de poder reductor de la célula es elevado. Elmecanismo que se realiza es una inhibición competitiva por producto (por NADH) de las enzimas que empleanNAD+ como sustrato. Así se regulan, entre otros, los complejos piruvato deshidrogenasa y citrato sintasa.
Principales vías que convergen en el ciclo de KrebsEl Ciclo de Krebs es una via metabólica central en la que convergen otras, tanto anabólicas como catabólicas.Igresan al ciclo por diferentes metabolitos:•• Acetil-CoA:
•• Glucolisis•• Oxidacion de ácidos grasos
•• Malato:•• Gluconeogénesis
•• Oxalacetato:•• Oxidación y biosíntesis de aminoácidos
•• Fumarato:•• Degradación de ácido aspártico, fenilalanina y tirosina
•• Succinil-CoA• Biosíntesis de porfirina•• Degradación de valina isoleucina y metionina•• Oxidación de ácidos grasos
•• Alfa-cetoglutarato•• Oxidación y biosíntesis de aminoácidos
•• Citrato•• Biosíntesis de ácidos grasos y colesterol
•• NADH y FADH•• Fosforilación oxidativa y cadena de transporte electónico
Véase también•• Descarboxilación oxidativa•• Ácido cítrico•• Glucólisis•• Fosforilación oxidativa• Ciclo de Krebs invertido (reductor)
Enlaces externos• Animación sobre el ciclo de Krebs [1] (castellano)• CiclodeKrebs.com [2] Información sobre el ciclo de Krebs• [3] Ciclo de krebs en el deporte• An animation of the citric acid cycle [4] (inglés)• A more detailed tutorial animation [5] (inglés)• A citric-acid cycle self quiz flash applet [6] (inglés)• The chemical logic behind the citric acid cycle [7] (inglés)

Ciclo de Krebs 5
References[1] http:/ / portales. educared. net/ wikiEducared/ index. php?title=Ciclo_de_Krebs[2] http:/ / www. ciclodekrebs. com[3] http:/ / www. todonatacion. com/ ciclo-de-krebs/[4] http:/ / www. science. smith. edu/ departments/ Biology/ Bio231/ krebs. html[5] http:/ / www. johnkyrk. com/ krebs. html[6] http:/ / www. pitt. edu/ AFShome/ j/ b/ jbrodsky/ public/ html/ 1820/ tca. htm[7] http:/ / www2. ufp. pt/ ~pedros/ bq/ tca. htm
Glucólisis
Reacción global de la glucólisis[1]
+
Glucosa + 2NAD+ + 2ADP + 2 2Piruvato + 2NADH + 2ATP + 2H+ + 2H2O
La glucólisis o glicolisis (del griego glycos, azúcar y lysis, ruptura), es la vía metabólica encargada de oxidar laglucosa con la finalidad de obtener energía para la célula. Consiste en 10 reacciones enzimáticas consecutivas queconvierten a la glucosa en dos moléculas de piruvato, el cual es capaz de seguir otras vías metabólicas y así continuarentregando energía al organismo.[1]
El tipo de glucólisis más común y más conocida es la vía de Embden-Meyerhof, explicada inicialmente por GustavEmbden y Otto Meyerhof. El término puede incluir vías alternativas, como la vía de Entner-Doudoroff. No obstante,glucólisis se usa con frecuencia como sinónimo de la vía de Embden-Meyerhof. Es la vía inicial del catabolismo(degradación) de carbohidratos.
DescubrimientoLos primeros estudios informales de los procesos glucolíticos fueron iniciados en 1860, cuando Louis Pasteurdescubrió que los microorganismos son los responsables de la fermentación,[2] y en 1897 cuando Eduard Buchnerencontró que cierto extracto celular puede causar fermentación. La siguiente gran contribución fue de Arthur Hardeny William Young en 1905, quienes determinaron que para que la fermentación tenga lugar son necesarias unafracción celular de masa molecular elevada y termosensible (enzimas) y una fracción citoplasmática de baja masamolecular y termorresistente (ATP, ADP, NAD+ y otras coenzimas). Los detalles de la vía en sí se determinaron en1940, con un gran avance de Otto Meyerhoff y algunos años después por Luis Leloir. Las mayores dificultades endeterminar lo intrincado de la vía fueron la corta vida y las bajas concentraciones de los intermediarios en las rápidasreacciones glicolíticas.En eucariotas y procariotas, la glucólisis ocurre en el citosol de la célula. En células vegetales, algunas de lasreacciones glucolíticas se encuentran también en el ciclo de Calvin, que ocurre dentro de los cloroplastos. La ampliaconservación de esta vía incluye los organismos filogenéticamente más antiguos, y por esto se considera una de lasvías metabólicas más antiguas.[3]

Glucólisis 6
Visión general
Esquema completo de la glucólisis
Durante la glucólisis se obtiene un rendimiento neto de dos moléculasde ATP y dos moléculas de NADH; el ATP puede ser usado comofuente de energía para realizar trabajo metabólico, mientras que elNADH puede tener diferentes destinos. Puede usarse como fuente depoder reductor en reacciones anabólicas; si hay oxígeno, puedeoxidarse en la cadena respiratoria, obteniéndose tres ATPs; si no hayoxígeno, se usa para reducir el piruvato a lactato (fermentación láctica),o a CO2 y etanol (fermentación alcohólica), sin obtención adicional deenergía.
La glucólisis es la forma más rápida de conseguir energía para una célula y, en el metabolismo de carbohidratos,generalmente es la primera vía a la cual se recurre. Se encuentra estructurada en 10 reacciones enzimáticas quepermiten la transformación de una molécula de glucosa a dos moléculas de piruvato mediante un proceso catabólico.La glucólisis es una de las vías más estudiadas, y generalmente se encuentra dividida en dos fases: la primera, degasto de energía y la segunda fase, de obtención de energía.La primera fase consiste en transformar una molécula de glucosa en dos moléculas de gliceraldehído (una moléculade baja energía) mediante el uso de 2 ATP. Esto permite duplicar los resultados de la segunda fase de obtenciónenergética.En la segunda fase, el gliceraldehído se transforma en un compuesto de alta energía, cuya hidrólisis genera unamolécula de ATP, y como se generaron 2 moléculas de gliceraldehído, se obtienen en realidad dos moléculas deATP. Esta obtención de energía se logra mediante el acoplamiento de una reacción fuertemente exergónica despuésde una levemente endergónica. Este acoplamiento ocurre una vez más en esta fase, generando dos moléculas depiruvato. De esta manera, en la segunda fase se obtienen 4 moléculas de ATP.
Reacciones posteriores
Véanse también: Fermentación y Ciclo de KrebsLuego de que una molécula de glucosa se transforme en 2 moléculas de piruvato, las condiciones del medio en quese encuentre determinarán la vía metabólica a seguir.En organismos aeróbicos, el piruvato seguirá oxidándose por la enzima piruvato deshidrogenasa y el ciclo de Krebs,creando intermediarios como NADH y FADH2. Estos intermediarios no pueden cruzar la membrana mitocondrial, ypor lo tanto, utilizan sistemas de intercambio con otros compuestos llamados lanzaderas (en inglés, shuttles). Losmás conocidos son la lanzadera malato-aspartato y la lanzadera glicerol-3-fosfato. Los intermediarios logran entregarsus equivalentes[4] al interior de la membrana mitocondrial, y que luego pasarán por la cadena de transporte deelectrones, que los usará para sintetizar ATP.De esta manera, se puede obtener hasta 30 moles de ATP a partir de 1 mol de glucosa como ganancia neta.Sin embargo, cuando las células no posean mitocondrias (ej: eritrocito) o cuando requieran de grandes cantidades deATP (ej.: el músculo al ejercitarse), el piruvato sufre fermentación que permite obtener 2 moles de ATP por cadamol de glucosa, por lo que esta vía es poco eficiente respecto a la fase aeróbica de la glucólisis.El tipo de fermentación varía respecto al tipo de organismos: en levaduras, se produce fermentación alcohólica,produciendo etanol y CO2 como productos finales, mientras que en músculo, eritrocitos y algunos microorganismosse produce fermentación láctica, que da como resultado ácido láctico o lactato.

Glucólisis 7
FuncionesLas funciones de la glucólisis son:• La generación de moléculas de alta energía (ATP y NADH) como fuente de energía celular en procesos de
respiración aeróbica (presencia de oxígeno) y fermentación (ausencia de oxígeno).• La generación de piruvato que pasará al ciclo de Krebs, como parte de la respiración aeróbica.•• La producción de intermediarios de 6 y 3 carbonos que pueden ser utilizados en otros procesos celulares.
Etapas de la glucólisisLa glucólisis se divide en dos partes principales y diez reacciones enzimáticas, que se describen a continuación.
Fase de gasto de energía (ATP)Esta primera fase de la glucólisis consiste en transformar una molécula de glucosa en dos moléculas degliceraldehído.
1er paso: Hexoquinasa
Véase también: Hexoquinasa
La primera reacción de la glucólisis es la fosforilación de la glucosa, para activarla(aumentar su energía) y así poder utilizarla en otros procesos cuando sea necesario. Estaactivación ocurre por la transferencia de un grupo fosfato del ATP, una reacción catalizadapor la enzima hexoquinasa,[5] la cual puede fosforilar (añadir un grupo fosfato) a moléculassimilares a la glucosa, como la fructosa y manosa. Las ventajas de fosforilar la glucosa son2: La primera es hacer de la glucosa un metabolito más reactivo, mencionado anteriormente,y la segunda ventaja es que la glucosa-6-fosfato no puede cruzar la membrana celular -adiferencia de la glucosa-ya que en la célula no existe un transportador de G6P. De estaforma se evita la pérdida de sustrato energético para la célula. Técnicamente hablando, lahexoquinasa sólo fosforila las D-hexosas, y utiliza de sustrato MgATP2+, ya que este catiónpermite que el último fosfato del ATP (fosfato gamma, γ-P o Pγ) sea un blanco más fácilpara el ataque nucleofílico que realiza el grupo hidroxilo (OH) del sexto carbono de laglucosa, lo que es posible debido al Mg2+ que apantalla las cargas de los otros dosfosfatos.[1]
Glucosa + ATP Glucosa-6-fosfato + ADP
[6]
[7] Esta reacción posee un ΔG negativo, y por tanto se trata de una reacción en la que sepierde energía en forma de calor. En numerosas bacterias esta reacción esta acoplada a laúltima reacción de la glucólisis (de fosfoenolpiruvato a piruvato) para poder aprovechar laenergía sobrante de la reacción: el fosfato del fosfoenolpiruvato se transfiere de una a otraproteína de un sistema de transporte fosfotransferasa, y en última instancia, el fosfato pasaráa una molécula de glucosa que es tomada del exterior de la célula y liberada en forma deG6P en el interior celular. Se trata por tanto de acoplar la primera y la última reacción deesta vía y usar el excedente de energía para realizar un tipo de transporte a través demembrana denominado translocación de grupo.
2° paso: Glucosa-6-P isomerasa
Véase también: Fosfohexosa isomerasa

Glucólisis 8
Éste es un paso importante, puesto que aquí se define la geometría molecular que afectarálos dos pasos críticos en la glucólisis: El próximo paso, que agregará un grupo fosfato alproducto de esta reacción, y el paso 4, cuando se creen dos moléculas de gliceraldehido quefinalmente serán las precursoras del piruvato.[1] En esta reacción, la glucosa-6-fosfato seisomeriza a fructosa-6-fosfato, mediante la enzima glucosa-6-fosfato isomerasa. Laisomerización ocurre en una reacción de 4 pasos, que implica la apertura del anillo y untraspaso de protones a través de un intermediario cis-enediol[8] Puesto que la energía librede esta reacción es igual a +1,7 kJ/mol la reacción es no espontánea y se debe acoplar.
Glucosa-6-fosfato Fructosa-6-fosfato
[6]
3er paso: Fosfofructoquinasa
Véase también: Fosfofructoquinasa-1
Fosforilación de la fructosa 6-fosfato en el carbono 1, con gasto de un ATP, a través de laenzima fosfofructoquinasa-1 (PFK1). También este fosfato tendrá una baja energía dehidrólisis. Por el mismo motivo que en la primera reacción, el proceso es irreversible. Elnuevo producto se denominará fructosa-1,6-bifosfato. La irreversibilidad es importante, yaque la hace ser el punto de control de la glucólisis. Como hay otros sustratos aparte de laglucosa que entran en la glucólisis, el punto de control no está colocado en la primerareacción, sino en ésta. La fosfofructoquinasa tiene centros alostéricos, sensibles a lasconcentraciones de intermediarios como citrato y ácidos grasos. Liberando una enzimallamada fosfructocinasa-2 que fosforila en el carbono 2 y regula la reacción. Fructosa-6-fosfato + ATP
Fructosa-1,6-bifosfato + ADP[6]
4° paso: Aldolasa
Véase también: Aldolasa
La enzima aldolasa (fructosa-1,6-bifosfato aldolasa), mediante una condensación aldólicareversible, rompe la fructosa-1,6-bifosfato en dos moléculas de tres carbonos (triosas):dihidroxiacetona fosfato y gliceraldehído-3-fosfato. Existen dos tipos de aldolasa, quedifieren tanto en el tipo de organismos donde se expresan, como en los intermediarios dereacción. Esta reacción tiene una energía libre (ΔG) entre 20 a 25 kJ/mol, por lo tanto encondiciones estándar no ocurre de manera espontánea. Sin embargo, en condicionesintracelulares la energía libre es pequeña debido a la baja concentración de los sustratos, loque permite que esta reacción sea reversible.[1]
Fructosa-1,6-bifosfato Dihidroxiacetona-fosfato + Gliceraldehído-3-fosfato
[6]
5° paso: Triosa fosfato isomerasa

Glucólisis 9
Puesto que sólo el gliceraldehído-3-fosfato puede seguir los pasos restantes de la glucólisis,la otra molécula generada por la reacción anterior (dihidroxiacetona-fosfato) es isomerizada(convertida) en gliceraldehído-3-fosfato. Esta reacción posee una energía libre encondiciones estándar positiva, lo cual implicaría un proceso no favorecido, sin embargo aligual que para la reacción 4, considerando las concentraciones intracelulares reales delreactivo y el producto, se encuentra que la energía libre total es negativa, por lo que ladirección favorecida es hacia la formación de G3P.Éste es el último paso de la "fase de gasto de energía". Sólo se ha consumido ATP en elprimer paso (hexoquinasa) y el tercer paso (fosfofructoquinasa-1). Cabe recordar que el 4topaso (aldolasa) genera una molécula de gliceraldehído-3-fosfato, mientras que el 5to pasogenera una segunda molécula de éste. De aquí en adelante, las reacciones a seguir ocurrirándos veces, debido a las 2 moléculas de gliceraldehído generadas de esta fase. Hasta estareacción hay intervención de energía (ATP).
Dihidroxiacetona-fosfato Gliceraldehído-3-fosfato
[6]
Fase de beneficio energético (ATP, NADH)Hasta el momento solo se ha consumido energía (ATP), sin embargo, en la segunda etapa, el gliceraldehído esconvertido a una molécula de mucha energía, donde finalmente se obtendrá el beneficio final de 4 moléculas deATP.
6° paso: Gliceraldehído-3-fosfato deshidrogenasa
Esta reacción consiste en oxidar elgliceraldehído-3-fosfato utilizando NAD+ paraañadir un ion fosfato a la molécula, la cual esrealizada por la enzima gliceraldehído-3-fosfatodeshidrogenasa o bien, GAP deshidrogenasa en 5pasos, y de ésta manera aumentar la energía delcompuesto.
Técnicamente, el grupo aldehído se oxida a ungrupo acil-fosfato, que es un derivado de uncarboxilo fosfatado. Este compuesto posee unaenergía de hidrólisis sumamente alta (cercana alos 50 kJ/mol) por lo que se da inicio al procesode reacciones que permitirán recuperar el ATPmás adelante.
Mientras el grupo aldehído se oxida, el NAD+ sereduce, lo que hace de esta reacción una reacciónredox. El NAD+ se reduce por la incorporación dealgún [H+] dando como resultado una moléculade NADH de carga neutra.
NAD+ NADH+ Pi + H+
Gliceraldehído-3-fosfatodeshidrogenasa
Gliceraldehído-3-fosfato+ Pi + NAD+
1,3-Bisfosfoglicerato+ NADH + H+
[6]
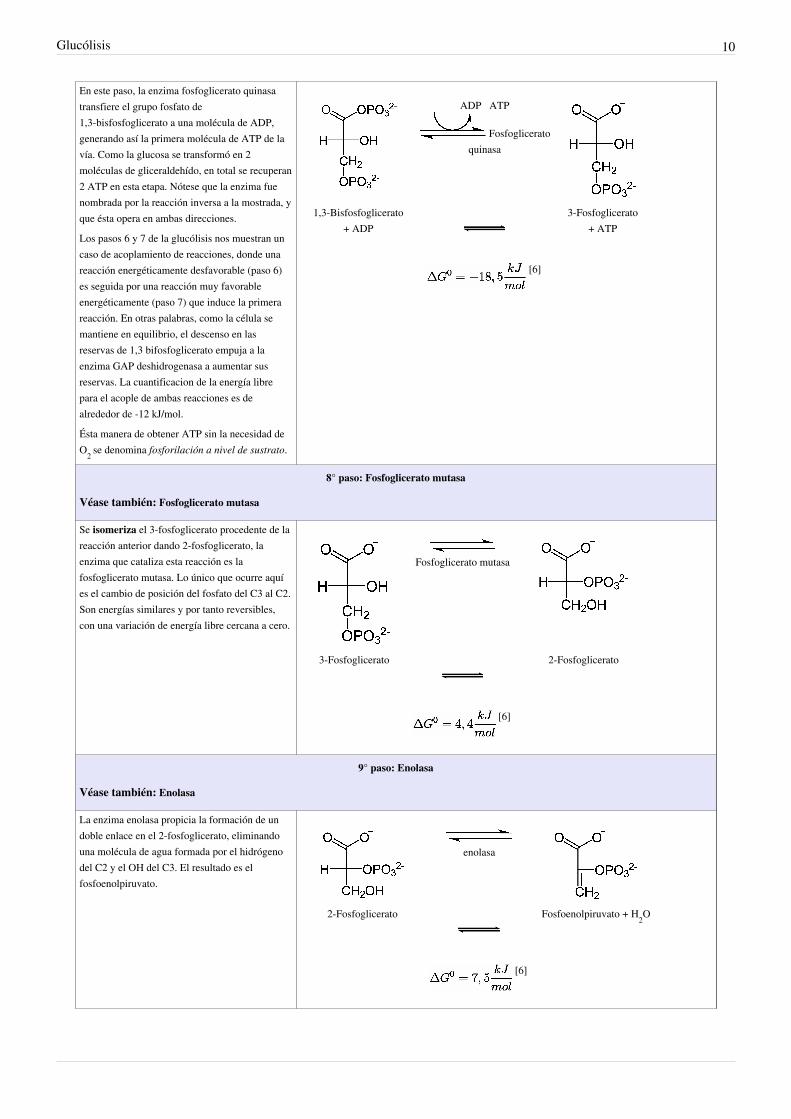
7° paso: Fosfoglicerato quinasa
Véase también: Fosfoglicerato quinasa
Glucólisis 10
En este paso, la enzima fosfoglicerato quinasatransfiere el grupo fosfato de1,3-bisfosfoglicerato a una molécula de ADP,generando así la primera molécula de ATP de lavía. Como la glucosa se transformó en 2moléculas de gliceraldehído, en total se recuperan2 ATP en esta etapa. Nótese que la enzima fuenombrada por la reacción inversa a la mostrada, yque ésta opera en ambas direcciones.
Los pasos 6 y 7 de la glucólisis nos muestran uncaso de acoplamiento de reacciones, donde unareacción energéticamente desfavorable (paso 6)es seguida por una reacción muy favorableenergéticamente (paso 7) que induce la primerareacción. En otras palabras, como la célula semantiene en equilibrio, el descenso en lasreservas de 1,3 bifosfoglicerato empuja a laenzima GAP deshidrogenasa a aumentar susreservas. La cuantificacion de la energía librepara el acople de ambas reacciones es dealrededor de -12 kJ/mol.
Ésta manera de obtener ATP sin la necesidad deO2 se denomina fosforilación a nivel de sustrato.
ADP ATP
Fosfogliceratoquinasa
1,3-Bisfosfoglicerato+ ADP
3-Fosfoglicerato+ ATP
[6]
8° paso: Fosfoglicerato mutasa
Véase también: Fosfoglicerato mutasa
Se isomeriza el 3-fosfoglicerato procedente de lareacción anterior dando 2-fosfoglicerato, laenzima que cataliza esta reacción es lafosfoglicerato mutasa. Lo único que ocurre aquíes el cambio de posición del fosfato del C3 al C2.Son energías similares y por tanto reversibles,con una variación de energía libre cercana a cero.
Fosfoglicerato mutasa
3-Fosfoglicerato 2-Fosfoglicerato
[6]
9° paso: Enolasa
Véase también: Enolasa
La enzima enolasa propicia la formación de undoble enlace en el 2-fosfoglicerato, eliminandouna molécula de agua formada por el hidrógenodel C2 y el OH del C3. El resultado es elfosfoenolpiruvato.
enolasa
2-Fosfoglicerato Fosfoenolpiruvato + H2O
[6]

Glucólisis 11
10° paso: Piruvato quinasa
Véase también: Piruvato quinasa
Desfosforilación del fosfoenolpiruvato,obteniéndose piruvato y ATP. Reacciónirreversible mediada por la piruvato quinasa.
ADP ATP
piruvato quinasa
Fosfoenolpiruvato Piruvato
[6]
El enzima piruvato quinasa es dependiente de magnesio y potasio. La energía libre es de -31,4 kJ/mol, por lo tanto lareacción es favorable e irreversible.El rendimiento total de la glucólisis de una sola glucosa (6C) es de 2 ATP y no 4 (dos por cadagliceraldehído-3-fosfato (3C)), ya que se consumen 2 ATP en la primera fase, y 2 NADH (que dejarán los electronesNc en la cadena de transporte de electrones para formar 3 ATP por cada electrón). Con la molécula de piruvato,mediante un paso de oxidación intermedio llamado descarboxilación oxidativa, mediante el cual el piruvato pasa alinterior de la mitocondria, perdiendo CO2 y un electrón que oxida el NAD+, que pasa a ser NADH más H+ yganando un CoA-SH (coenzima A), formándose en acetil-CoA gracias a la enzima piruvato deshidrogenasa, sepuede entrar al ciclo de Krebs (que, junto con la cadena de transporte de electrones, se denomina respiración).
Regulación
El efecto PasteurEl efecto Pasteur es la visualización del poder que posee el O2 en la fermentación mediada por levadura, que fuedescubierto por Luis Pasteur al observar la relación entre la tasa de fermentación y la existencia de aire. El determinóque éstas tenían una relación inversa, y además observó que en condiciones aeróbicas, las células de levaduraaumentaban y la fermentación disminuía.De esta manera, el efecto Pasteur fue una de las primeras observaciones que alguien realizó al proceso de laglucólisis de manera indirecta, pero observando que el metabolismo primario de glucosa se podía realizar conpresencia o ausencia de oxigeno, y que en este último ocurre la fermentación alcohólica.

Glucólisis 12
Regulación del sustratoVéase también: Transportador de glucosaLa membrana plasmatica de las celulas es impermeable a la glucosa. Para llevarla dentro de ella utilizatrasportadores especiales llamados GLUT, de los cuales hay diferentes tipos y algunos especializados para cadacélula.
Regulación de la actividad enzimática
Regulación glucolisis
La glucólisis se regula enzimáticamente en los tres puntos irreversiblesde esta ruta, esto es, en la primera reacción (G → G-6P), por medio dela hexoquinasa; en la tercera reacción (F-6P → F-1,6-BP) por mediode la PFK1 y en el último paso (PEP → Piruvato) por la piruvatoquinasa.
• La hexoquinasa es un punto de regulación poco importante, ya quese inhibe cuando hay mucho G-6P en músculo. Es un punto pocoimportante ya que el G-6P se utiliza para otras vías.
• La fosfofructoquinasa-1 es la enzima principal de la regulación dela glucólisis, actúa como una llave de agua, si está activa catalizamuchas reacciones y se obtiene más Fructosa 1,6 bisfosfato, lo quepermitirá a las enzimas siguientes transformar mucho piruvato. Siestá inhibida, se obtienen bajas concentraciones de producto y porlo tanto se obtiene poco piruvato. Esta enzima es controlada porregulación alostérica mediante: Por un lado se activa gracias a niveles energéticos elevados de ADP y AMP,inhibiendose en abundancia de ATP y citrato, y por otro se activa en presencia de un regulador generado por laPFK2 que es la Fructosa-2,6-Bisfosfato (F-2,6-BP), que no es un metabolito ni de la glucolisis ni de lagluconeogénesis, sino un regulador de ambas vías que refleja el nivel de glucagón en sangre.
La lógica de la inhibición y activación son las siguientes:•• ATP: inhibe esta enzima pues si hay una alta concentración de ATP entonces la célula no necesita generar más.•• Citrato: Si la concentración de citrato es alta el Ciclo de Krebs va más despacio de lo que el sustrato
(acetil-CoA) llega para degradarse, y la concentración de glucosa será más alta. En el Ciclo de Krebs seproduce mucho NADH y FADH2, para que funcionen se han de reoxidar en la cadena de transporteelectrónico creando gradiente de protones, si el gradiente no se gasta los coenzimas no se reoxidan y el Ciclode Krebs se para.
•• AMP, ADP: la alta concentración de estas moléculas implica que hay una carencia de ATP, por lo que esnecesario realizar glucólisis, para generar piruvato y energía.
• La piruvatoquinasa se regula distintamente según el tejido en el que trabaje, pero en hígado se inhibe enpresencia de ATP y Acetil Coenzima-A (Acetil-CoA), y se activa gracias de nuevo ante la F-2,6-BP y laconcentración de fosfoenolpiruvato.

Glucólisis 13
Regulación hormonalAl aumentar la glucosa en la sangre, después de una comida, las células beta del páncreas estimulan la producción deinsulina, y ésta a su vez aumenta la actividad de la glucocinasa en los hepatocitos.Las concentraciones altas de glucagon y las bajas de insulina disminuyen la concentración intracelular de fructosa2,6 bisfosfato. Esto trae por consecuencia la disminución de la glicólisis y el aumento de la gluconeogenésis.
Producción de glucosaLa gluconeogénesis es la ruta anabólica por la que tiene lugar la síntesis de nueva glucosa a partir de precursores noglucosídicos (lactato, piruvato, glicerol y algunos aminoácidos). Se lleva a cabo principalmente en el hígado, y enmenor medida en la corteza renal. Es estímulada por la hormona glucagón, secretada por las células α (alfa) de losislotes de Langerhans del páncreas y es inhibida por su contrarreguladora, la hormona insulina, secretada por lascélulas β (beta) de los islotes de Langerhans del páncreas, que estímula la ruta catabólica llamada glucogenólisis paradegradar el glucógeno almacenado y transformarlo en glucosa y así aumentar la glucemia (azúcar en sangre).Desde el punto de vista enzimático, producir glucosiliosas desde lacticosinidas cuesta más de lo que produjo sudegradación fosfórica. La ecuación extrafundamental es:
2 piruvato + 4 ATP + 2 GTP + 9 NADH + 7 H + 3 H2O → Glucosa + 4 ADP + 2 GDP + 6 P + 2 NAD+
Glucólisis en plantasLas plantas tienen la capacidad de realizar la fotosintesis, y entre los subproductos de este proceso está la glucosa.Ésta es usada por las plantas, entre muchas cosas, como fuente de energia en el proceso de respiración, el cual adiferencia de la fotosintesis es ejecutado independientemente de la luz. Al respirar las plantas absorben oxigeno delaire y expulsan dioxido de carbono y vapor de agua. El intercambio de sustancias lo realizan las estomas; aberturasque actuan como compuertas en las plantas que además tienen la caracteristica de cerrarse ante un descenso excesivodel vapor atmosferico.[9]
Referencias[1] David Nelson & Michael Cox (2004). «Glycolysis, Gluconeogenesis and the Pentose Phosphate Pathway». Lehningher's Principles of
Biochemistry. W.H.Freeman. 0716743396.[2] Papers de Pasteur (http:/ / biotech. law. lsu. edu/ cphl/ history/ articles/ pasteur. htm)[3] Romano AH & Conway T. Evolution of carbohydrate metabolic pathways. Res Microbiol. 147(6-7):448-55 (1996) PMID 9084754[4] No se usan los intermediarios generados, sino que por medio de las lanzaderas se vuelven a crear dentro de la mitocondria. Por esto se les
llama sus equivalentes. Para una visión química, visitar equivalentes[5] Meyerhof, O. Ueber die enzymatische Milch-säurebildung im Muskelextrakt; die Milch-säurebildung aus den gärfähigen Hexosen. Biochem
Z. 183:176 (1927)[6] Valores tomados de Lehningher's Principles of Biochemistry (ISBN 0-7167-4339-6) y del Volumen 3 de Biochemistry por J. Stenesh (ISBN
0-306-45733-4)[7] Colowick, S. y Kalckar H.. The role of myokinase in trans-phosphorylations; the enzymatic phosphorylation of hexoses by adenyl
pyrophosphate. J. Biol. Chem. 148: 117 (1943).[8] Irwin A. Rose (2006). «Mechanism of the Aldose-Ketose Isomerase Reactions». Advances in Enzymology - and Related Areas of Molecular
Biology, Volume 43. Wiley Interscience. ISBN 0471591788. - doi 10.1002/9780470122884.ch6 (http:/ / dx. doi. org/ 10. 1002/9780470122884. ch6)
[9] « Funciones de las plantas (http:/ / www. botanical-online. com/ funcionesplantas. htm)» (en español). Consultado el 30 de agosto de 2011.

Glucólisis 14
Véase también•• Metabolismo•• Ciclo de Krebs•• Fermentación•• Fosforilación oxidativa•• Vía metabólica
Enlaces externos• soko.com.ar: Explicación más extensa (http:/ / soko. com. ar/ Biologia/ Glucolisis. htm)• www2.ufp.pt: A lógica química da Glicólise (Portugués) (http:/ / www2. ufp. pt/ ~pedros/ bq/ glicolise. htm)• www.pdb.org: The Glycolytic Enzymes, información en Protein Data Bank (http:/ / www. pdb. org/ pdb/ static.
do?p=education_discussion/ molecule_of_the_month/ pdb50_1. html) (en inglés)• Explicación del proceso de glucólisis (Youtube) (http:/ / www. youtube. com/ watch?v=56tu7sKFh0w&
list=PLE21A11C7E7D1D26D)
• Wikimedia Commons alberga contenido multimedia sobre Glucólisis. Commons
Gluconeogénesis
Nombres en azul indican los sustratos de la vía, flechas en rojo lasreacciones únicas de esta vía, flechas cortadas indican reacciones dela glucolisis, que van en contra de esta vía, flechas en negrita indican
la dirección de la gluconeogénesis.
La gluconeogénesis es una ruta metabólica anabólicaque permite la biosíntesis de glucosa a partir deprecursores no glucídicos. Incluye la utilización devarios aminoácidos, lactato, piruvato, glicerol ycualquiera de los intermediarios del ciclo de los ácidostricarboxílicos (o ciclo de Krebs) como fuentes decarbono para la vía metabólica. Todos losaminoácidos, excepto la leucina y la lisina, puedensuministrar carbono para la síntesis de glucosa. LosÁcidos grasos de cadena par no proporcionan carbonospara la síntesis de glucosa, pues el resultado de suβ-oxidación (Acetil-CoA) no es un sustratogluconeogénico; mientras que los ácidos grasos decadena impar proporcionarán un esqueleto deCarbonos que derivarán en Acetil-CoA y Succinil-CoA(que sí es un sustrato gluconeogénico por ser unintermediario del ciclo de Krebs).
Algunos tejidos, como el cerebro, los eritrocitos, elriñón, la córnea del ojo y el músculo, cuando elindividuo realiza actividad extenuante, requieren de unaporte continuo de glucosa, obteniéndola a partir delglucógeno proveniente del hígado, el cual solo puedesatisfacer estas necesidades durante 10 a 18 horascomo máximo, lo que tarda en agotarse el glucógeno

Gluconeogénesis 15
almacenado en el hígado. Posteriormente comienza la formación de glucosa a partir de sustratos diferentes alglucógeno.La gluconeogénesis tiene lugar casi exclusivamente en el hígado (10% en los riñones). Es un proceso clave puespermite a los organismos superiores obtener glucosa en estados metabólicos como el ayuno.
Reacciones de la gluconeogénesisLas enzimas que participan en la vía glucolítica participan también en la gluconeogénesis; ambas rutas se diferencianpor tres reacciones irreversibles que utilizan enzimas específicas de este proceso y que condicionan los dos rodeosmetabólicos de esta vía.Estas reacciones son:1. De glucosa a glucosa-6P.2. De fructosa-6P a fructosa-1,6-bisfosfato.3. De fosfoenolpiruvato a piruvato.
Conversión del piruvato en fosfoenolpiruvatoEl oxaloacetato es intermediario en la producción del fosfoenolpiruvato en la gluconeogénesis. La conversión depiruvato a fosfoenolpiruvato en la gluconeogénesis se lleva a cabo en dos pasos. El primero de ellos es la reacción depiruvato y dióxido de carbono para dar oxaloacetato. Este paso requiere energía, la cual queda disponible porhidrólisis de ATP.La enzima que cataliza esta reacción es la piruvato carboxilasa, una enzima alostérica que se encuentra en lamitocondria. El acetil-CoA es un efector alostérico que activa la piruvato carboxilasa. Cuando hay más acetil-CoAdel necesario para mantener el ciclo del ácido cítrico, el piruvato se dirige a la gluconeogénesis. El ion magnesio y labiotina son necesarios para una catálisis eficaz.La biotina, enlazada covalentemente con la enzima, reacciona con el CO2, que se une de manera covalente. Despuésel CO2 se incorpora al piruvato, formando así oxaloacetato.La conversión de oxaloacetato a fosfoenolpiruvato la cataliza la enzima fosfoenolpiruvato carboxiquinasa, que seencuentra en la mitocondria y en el citosol. Esta reacción también incluye la hidrólisis de un nucleósido-trifosfato, eneste caso el GTP en vez del ATP.
Conversión de la fructosa-1,6-bisfosfato en fructosa-6-fosfatoLa reacción de la fosfofructoquinasa 1 de la glucólisis es esencialmente irreversible pero sólo debido a que estáimpulsada por la transferencia de fosfato del ATP. La reacción que tiene lugar en la gluconeogénesis para evitar estepaso consiste en una simple reacción hidrolítica, catalizada por la fructosa-1,6-bisfosfatasa.La enzima con múltiples subunidades requiere la presencia de Mg2+ para su actividad y constituye uno de losprincipales lugares de control que regulan la ruta global de la gluconeogénesis. La fructosa-6-fosfato formada en estareacción experimenta posteriormente la isomerización a glucosa-6-fosfato por la acción de la fosfoglucoisomerasa.

Gluconeogénesis 16
Conversión de la glucosa-6-fosfato en glucosaLa glucosa-6-fosfato no puede convertirse en glucosa por la acción inversa de la hexoquinasa o la glucoquinasa; latrasferencia de fosfato desde el ATP hace a la reacción virtualmente irreversible. Otra enzima específica de lagluconeogénesis, la glucosa-6-fosfatasa, que también requiere Mg2+, es la que entra en acción en su lugar. Estareacción de derivación se produce también mediante una simple hidrólisis.La glucosa-6-fosfatasa se encuentra fundamentalmente en el retículo endoplásmico del hígado con su lugar activosobre el lado citosólico. La importancia de su localización en el hígado es que una función característica del hígadoes sintetizar glucosa para exportarla a los tejidos a través de la circulación sanguínea.
RegulaciónLa regulación de la gluconeogénesis es crucial para muchas funciones fisiológicas, pero sobre todo para elfuncionamiento adecuado del tejido nervioso. El flujo a través de la ruta debe aumentar o disminuir, en función dellactato producido por los músculos, de la glucosa procedente de la alimentación, o de otros precursoresgluconeogénicos.La gluconeogénesis está controlada en gran parte por la alimentación. Los animales que ingieren abundantes hidratosde carbono presentan tasas bajas de gluconeogénesis, mientras que los animales en ayunas o los que ingieren pocoshidratos de carbono presentan un flujo elevado a través de esta ruta.Dado que la gluconeogénesis sintetiza glucosa y la glucólisis la cataboliza, es evidente que la gluconeogénesis y laglucólisis deben controlarse de manera recíproca. En otras palabras, las condiciones intracelulares que activan unaruta tienden a inhibir la otra.
Regulación por los niveles de energíaLa fructosa 1,6-bisfosfatasa es inhibida por concentraciones altas de AMP, asociadas con un estado energéticamentepobre. Es decir, la elevada concentración de AMP y reducida de ATP estimulan la gluconeogénesis.
Regulación por fructosa 2,6-bisfosfatoLa fructosa 1,6-bisfosfatasa es inhibida por la fructosa 2,6-bisfosfato, un modulador alostérico cuya concentraciónviene determinada por la concentración circulante en sangre de glucagón; la fructuosa 1,6-bisfosfatasa está presentetanto en el hígado como en los riñones.
Regulación de la fosforilaciónEste proceso es dependiente de la concentración de ATP; al disminuir la concentración de ATP, la fosforilacióntambién se observa disminuida y viceversa. En el hígado, este proceso aumenta al aumentar la síntesis deglucocinasa, proceso que es promovido por la insulina. La membrana de los hepatocitos es muy permeable a laglucosa, en el músculo y el tejido adiposo la insulina actúa sobre la membrana para hacerla permeable a ella.
Regulación alostéricaLa inanición aumenta el acetil-CoA y éste estimula la piruvato carboxilasa y por lo tanto la gluconeogénesis, almismo tiempo que inhibe la Piruvato Deshidrogenasa; la elevación de alanina y glutamina estimulan lagluconeogénesis. El cortisol aumenta la disponibilidad de sustrato y la fructosa 2,6-bisfosfato inhibe a la fructosa1,6-bisfosfatasa.

Gluconeogénesis 17
Balance energéticoHemos resaltado que las rutas catabólicas generan energía, mientras que las anabólicas comportan un costeenergético. En el caso de la gluconeogénesis podemos calcular este coste; la síntesis de glucosa es costosa para lacélula en un sentido energético. Si partimos desde piruvato se consumen seis grupos fosfato de energía elevada 4ATP (debido a las reacciones de la piruvato carboxilasa y a la de fosfoglicerato quinasa) y 2 GTP (consecuencia dela descarboxilación del oxalacetato), así como 2 de NADH, que es el equivalente energético de otros 5 ATP (ya quela oxidación mitocondrial de 1 NADH genera 2,5 ATP).En cambio, si la glucólisis pudiera actuar en sentido inverso, el gasto de energía sería mucho menor: 2 NADH y 2ATP
Reacción Global
2 Ácido pirúvico + 4 ATP + 2 GTP + 2 NADH + 6 H2O + 2H+ -----------> Glucosa + 4ADP + 2GDP +6Pi + 2NAD+
Importancia biomédicaLa gluconeogénesis cubre las necesidades corporales de glucosa cuando no está disponible en cantidades suficientesen la alimentación. Se requiere un suministro constante de glucosa como fuente de energía para el sistema nervioso ylos eritrocitos. Además, la glucosa es el único combustible que suministra energía al músculo esquelético encondiciones de anaerobiosis. La glucosa es precursora del azúcar de la leche (lactosa) en la glándula mamaria y secapta activamente por el feto. Por otro lado, los mecanismos gluconeogénicos se utilizan para depurar los productosdel metabolismo de otros tejidos desde la sangre; por ejemplo, lactato, producido por el músculo y los eritrocitos, yglicerol, que se forma continuamente por el tejido adiposo.
Referencias• Álvarez Rodríguez Bertha Adriana y colb. Bioquímica: Metabolismo de carbohidratos. Academia de Bioquímica.
Pags: 64-70• Benyon S., Roach J. O`Neale. Lo esencial en metabolismo y nutrición. Editorial Harcourt Brace. Madrid, España.
Pags: 89-91• Campbell Mary K., Farrell Shaw O. Bioquímica, 4a Edición. Editorial Thomson International. México D.F., 2004.
Pags. 497-501• Murray Robert K., Mayes Peter A., Granner Daryl K., Rodwell Victor W. Bioquímica de Harper, 14ª edición.
Editorial Manual Moderno, México D.F., 2001, pags 233 – 244• Mathews, Van Holde, Adhern. Bioquímica, 3ª edición. Editorial Pearson Addison Wesley. Madrid, España, 2002,
Pag. 628-639

Glucogenolisis 18
GlucogenolisisLa glucogenólisis es un proceso catabólico llevado a cabo en el citosol que consiste en la remoción de un monómerode glucosa de un glucógeno mediante fosforólisis para producir glucosa 1 fosfato, que después se convertirá englucosa 6 fosfato, la glucólisis. Es antagónica de la glucogénesis. Estimulada por el glucagón en el hígado, epinefrina(adrenalina) en el músculo e inhibida por la insulina.Es un proceso que requiere un grupo específico de enzimas citosolíticas: la glucógeno fosforilasa que segmentasecuencialmente los enlaces glucosídicos, la fosfoglucomutasa que convierte la G1P en G6P la cual puedehidrolizarse a glucosa (en hígado) o seguir la vía glucolítica (hígado y músculo) y por último la Glucosil Transferasaα(1→4) y la amilo-1,6-glucosidasa, que se encarga de hidrolizar las ramificaciones. Su deficiencia produce laEnfermedad de Cori y la Enfermedad de Pompe
Véase también•• Glucólisis
GlucogénesisLa glucogénesis es la ruta anabólica por la que tiene lugar la síntesis de glucógeno (también llamado glicógeno) apartir de un precursor más simple, la glucosa-6-fosfato. Se lleva a cabo principalmente en el hígado, y en menormedida en el músculo, es activado por insulina en respuesta a los altos niveles de glucosa, que pueden ser (porejemplo) posteriores a la ingesta de alimentos con carbohidratos.Se forma por la incorporación repetida de unidades de glucosa, la que llega en forma de UDP-Glucosa a un partidorde glucógeno preexistente que consiste en la proteína glucogenina, formada por 2 cadenas, que al autoglicosilarsepuede unir cada una de sus cadenas a un octámero de glucosas. Para que la glucosa-6-fosfato pueda unirse a la UDPrequiere de la participación de dos enzimas, la primera, fosfoglucomutasa, modifica la posición del fosfato aglucosa-1-fosfato.La glucosa-1-fosfato es el precursor para la síntesis de glucógeno pero también es el producto de su degradación. Lasíntesis de glucógeno requiere de aporte energético. El dador de glucosa para la sínteis de glucógeno es laUDP-glucosa donde el residuo glucosilo está activado para su transferencia, por su combinación con un compuestode alta energía como el UTP.
Pasos• La Glucosa se convierte en glucosa-6-fosfato mediante una reacción irreversible catalizada por la glucoquinasa o
hexoquinasa dependiendo del tejido en cuestión.glucosa + ATP → glucosa-6-P + ADP
• Glucosa-6-fosfato se convierte en glucosa-1-fosfato por la acción de la Fosfoglucomutasa, mediante la formaciónobligada de un compuesto intermediario, glucosa-1,6-bisfosfatasa.
glucosa-6-P ←→ glucosa-1-P
• Glucosa-1-fosfato se convierte en UDP-glucosa por la acción de la UDP-glucosa pirofosforilasa (llamada tambiénuridil transferasa).
glucosa-1-P + UTP → UDP-glucosa + PPi
• Las moléculas de glucosa son acopladas en cadena por la glucógeno sintasa, este paso debe realizarse sobre unprimer preexistente de glucogeno que contiene una pequeña proteína llamada glucogenina.

Glucogénesis 19
• Las ramificaciones son producidas por la enzima ramificadora del glucógeno, la cual transfiere un fragmento de 6a 8 unidades del extremo no reductor y lo une a una glucosa por un enlace α-1,6. Esto posibilita que ambascadenas puedan continuar alargándose mediante uniones α-1,4 de glucosas hasta poder producir nuevasramificaciones.
Enlaces externos• The chemical logic behind glycogen synthesis [1] (inglés)
Véase también•• Glucógeno•• Glucogenólisis
References[1] http:/ / www2. ufp. pt/ ~pedros/ bq/ glycogen. htm
Ruta de la pentosa fosfatoLa ruta de la pentosa fosfato, también conocida como lanzadera de fosfatos de pentosas, es una ruta metabólicaestrechamente relacionada con la EMP durante la cual se utiliza la glucosa para generar ribosa, que es necesaria parala biosíntesis de nucleótidos y ácidos nucleicos. Además, también se obtiene poder reductor en forma de NADP+ quese utilizará como coenzima de enzimas propias del metabolismo anabólico.De esta manera, este proceso metabólico, el cual es regulado por insulina, tiene una doble función, ya que la glucosase usa para formar NADPH, mientras que también se puede transformar en otros componentes del metabolismo,especialmente pentosas, utilizadas para la síntesis de nucleótidos y de ácidos nucleicos. Así, se forma un puente entrerutas anabólicas y catabólicas de la glucosa.[1]
La ruta de la pentosa fosfato tiene lugar en el citosol, y puede dividirse en dos fases:• Fase oxidativa: se genera NADPH.• Fase no oxidativa: se sintetizan pentosas-fosfato y otros monosacáridos-fosfato.
Fase oxidativaDurante fase oxidativa, a partir de glucosa-6-fosfato obtenida mediante la fosforilación de la glucosa libre, se obtieneNADPH y finalmente se forma la pentosa ribulosa-5-fosfato, motivo por el cual este proceso metabólico sedenomina “la ruta de la pentosa fosfato”.La primera reacción es la oxidación de la glucosa-6-fosfato, llevada a cabo por la enzima glucosa-6-fosfatodeshidrogenasa. En este primer paso se deshidrogena el grupo C1 para dar un grupo carboxilo, el cual, junto al C5,forma una lactona, es decir, un éster intramolecular. Es aquí donde se liberan dos hidrógenos de los cuales setransfiere un protón (H+) y dos electrones (e-) (hidridión) al NADP+ que actua como aceptor de electronesreduciéndose hasta formar la primera molécula de NADPH; el protón sobrante queda libre en el medio.Acto seguido, se produce la hidrólisis de la lactona gracias a la actuación de la lactonasa, con lo que se obtiene elácido libre 6-fosfogluconato. Seguidamente, éste último se transforma en ribulosa-5-fosfato por acción de la6-fosfogluconato deshidrogenasa. Aquí se obtiene la segunda molécula de NADPH, además de la liberación de unamolécula de CO2 debido a la descarboxilación oxidativa del ácido libre.

Ruta de la pentosa fosfato 20
Finalmente, la enzima pentosa-5-fosfato isomerasa, mediante un intermediario endiol, isomeriza la ribulosa-5-fosfatoy la convierte en ribosa-5-fosfato, gracias a la transformación del grupo cetosa en aldosa. Esta última reacciónprepara un componente central de la síntesis de nucleótidos para la biosíntesis de RNA, DNA y cofactores denucleótidos. Al mismo tiempo, lleva a cabo la transición hacia la fase no oxidativa de la ruta metabólica de lapentosa fosfato.De este modo se acaba obteniendo dos moléculas de NADPH que, además de su uso en la biosíntesis reductiva,también es responsable del mantenimiento de un medio reductor en la célula. Esto puede verse si hay un déficit deglucosa-6-fosfato deshidrogenasa, producido por un defecto en un gen que se encuentra en el cromosoma X,pudiendo afectar con mayor proporción a los varones.Los glóbulos rojos de la sangre necesitan grandes cantidades de NADPH para la reducción de la hemoglobinaoxidada y para poder regenerar el glutatión reducido, un antioxidante que presenta importantes funciones como laeliminación de peróxidos y la reducción de ferrihemoglobina (Fe3+). Estas necesidades se ven cubiertas gracias a laruta de la pentosa fosfato con el intermediario de reducción NADPH. Sin embargo, si existe este defecto genético,debido a la ingesta de algún determinado medicamento, como el antimalárico primaquina, o algunos vegetales, comopor ejemplo las habas, los eritrocitos se distribuyen en un lugar de debilidad, pudiendo desenvolver en una graveanemia hemolítica. Esta mutación genética podría aumentar la producción de peróxidos y con ello también habría laoxidación de los lípidos de membrana, junto a la aceleración de la degradación de los eritrocitos. De este modo, sepuede observar como la ruta de la pentosa fosfato es la única vía metabólica por la cual estas células pueden producirNADPH.A pesar de todo, los afectados por este problema congénito se ven altamente favorecidos en un aspecto. Estos suelenvivir en zonas tropicales, ya que son mejores protectores contra infecciones de malaria. Esto puede verse explicadopor la necesidad inmediata de los plasmodios hacia un medio reducido para su metabolismo, ya que los parásitosresisten mucho menos el estrés oxidativo respecto a sus células huésped.
Fase oxidativa de la ruta de la pentosa fosfato.
Todas las reacciones de esta primera parte del proceso metabólico se ven resumidas en la siguiente tabla:
Reactivos Productos Enzima Descripción
Glucosa-6-fosfato +NADP+
→ 6-Fosfoglucanato;-Lactona+ NADPH
Glucosa-6-fosfatodeshidrogenasa
Deshidrogenación. El grupo hidroxilo localizado en el C1de la glucosa-6-fosfato es convertido en un grupocarbonilo, generando una lactona y una molécula deNADPH durante el proceso.
6-Fosfoglucanato;-Lactona+ H2O
→ 6-Fosfoglucanato + H+ 6-Fosfoglucolactonasa Hidrólisis
6-Fosfoglucanato + NADP+ → Ribulosa-5-fosfato +NADPH + CO2
6-Fosfoglucanatodeshidrogenasa
Descarboxilación. El NADP+ es el aceptor de electrones,generando otra molécula de NADPH, un CO2 iRibulosa-5-fosfato.
La reacción general de esta primera fase es:Glucosa-6-fosfat + 2 NADP+ + H2O → Ribulosa-5-fosfat + 2 NADPH + 2 H+ + CO2

Ruta de la pentosa fosfato 21
Así, se puede ver como el NADPH es usado en la síntesis de ácidos grasos y colesterol, reacciones de hidroxilaciónde neurotransmisores, detoxificación de peróxidos de hidrógeno, así como en el mantenimiento del glutatión en suforma reducida.
Fase no oxidativaLa fase no oxidativa de la ruta de la pentosa fosfato se inicia en caso que la célula necesite más NADPH queribosa-5-fosfato. En este segundo proceso se encuentran una compleja secuencia de reacciones que permiten cambiarlos azúcares C3, C4, C5, C6 y C7 de las pentosas para poder formar finalmente gliceraldehído-3-fosfato yfructosa-6-fosfato, los cuales podrán seguir directamente con la glucólisis.Esta fase conlleva toda una serie de reacciones reversibles, el sentido de las cuales depende de la disponibilidad delsustrato. Asimismo, la isomerización de ribulosa-5-fosfato a ribosa-5-fosfato es también reversible. Esto nos permitepoder eliminar el excedente de ribosa-5-fosfato para acabar transformándolo en productos intermediarios de laglucólisis.La primera reacción llevada a cabo es la epimerización, regulada mediante la enzima pentosa-5-fosfato epimerasa,que convertirá la ribulosa-5-fosfato, producto de la fase oxidativa, en xilulosa-5-fosfato, generando así el sustratonecesario para la siguiente reacción controlada por la transcetolasa, la cual actúa junto a la coenzima pirofosfato detiamina (TPP). Ésta convertirá la xilulosa-5-fosfato en ribosa-5-fosfato y, mediante la transferencia de una unidad deC2 de la cetosa a la aldosa, se producirá gliceraldehído-3-fosfato y sedoheptulosa-7-fosfato.Sucedido esto, la transaldolasa, con la ayuda de un resto lisina en su centro activo, transfiere una unidad C3 de lasedoheptulosa-7-fosfato a gliceraldehído-3-fosfato, con lo que se formarán la tetrosa eritrosa-4-fosfato, además deuno de los primeros productos finales: la hexosa fructosa-6-fosfato, la cual se dirigirá hacia la glucólisis.Acto seguido, la enzima transcetolasa vuelve a transferir una unidad C2, desde la xilulosa-5-fosfato aeritrosa-4-fosfato, consiguiendo así formar otra molécula de fructosa-6-fosfato y un gliceraldehído-3-fosfato, ambosintermediarios de la glucólisis. De esta manera, se cierra la fase no oxidativa de esta ruta metabólica.[2]
Esta fase de la ruta conectará los procesos metabólicos que generan NADPH con los que originan NADH/ATP. Porotra parte, el gliceraldehído-3-fosfato y la fructosa-6-fosfato pueden intervenir, en vez de en el glucólisis, en lagluconeogénesis para formar una nueva síntesis de glucosa.

Ruta de la pentosa fosfato 22
Fase no oxidativa de la ruta de la pentosa fosfato.
Todas las reacciones de esta segunda parte del proceso metabólico se ven resumidas en la siguiente tabla:
Reactivos Productos Enzima
Ribulosa-5-fosfato → Ribosa-5-fosfato Ribulosa-5-fosfato Isomerasa
Ribulosa-5-fosfato → Xilulosa-5-fosfato Ribulosa-5-fosfato 3-Epimerasa
Xilulosa-5-fosfato + Ribosa-5-fosfato → Gliceraldehído-3-fosfato + Sedoheptulosa-7-fosfato Transcetolasa
Sedoheptulosa-7-fosfato + Gliceraldehído-3-fosfato → Eritrosa-4-fosfato + Fructosa-6-fosfato Transaldolasa
Xilulosa-5-fosfato + Eritrosa-4-fosfato → Gliceraldehído-3-fosfato + Fructosa-6-fosfato Transcetolasa
La célula y la ruta de la pentosa fosfatoLa ruta de la pentosa fosfato tiene una gran flexibilidad, de hecho, es un módulo ideal del metabolismo, que seadapta continuamente a las cantidades requeridas de ATP, NADPH, ribosa-5-fosfato, piruvato o acetil-CoA, segúnlas necesidades de la célula.Esta ruta se ve regularizada mediante la enzima glucosa-6-fosfato deshidrogenasa. El regulador más importante es laoferta de NADP+, el cual actúa como activador alostérico, mientras que el NADPH disminuye la actividad de laenzima como inhibidor competitivo. En condiciones fisiológicas el NADPH se encuentra en mayor proporción(70:1) respecto NADP+, si hubiese una utilización de equivalentes de reducción conduciría rápidamente a laestimulación de la deshidrogenasa debido al aumento de la cantidad de NADP+.Consecuentemente, esta ruta metabólica transcurre fuertemente en el tejido adiposo, donde hay una gran oferta deglucosa y una alta necesidad de NADPH, requerido para la biosíntesis de ácidos grasos. Por el contrario, en el tejidomuscular, se encuentra una baja necesidad de NADPH, por lo que se realiza la inversión de la ruta.

Ruta de la pentosa fosfato 23
En el caso del tejido adiposo, se dará lugar a NADPH para las células del tejido, pero, la formación deribosa-5-fosfato no dará suficiente síntesis de nucleótidos, hecho que provocará la conversión de las pentosas engliceraldehído-3-fosfato y fructosa-6-fosfato. Por lo general, estas biomoléculas se incorporarán en la glucólisis,con la ayuda de la enzima piruvato deshidrogenasa, para formar, finalmente, acetil-CoA necesario para la síntesis deácidos grasos. Así pues, en la glucólisis simultáneamente se forman equivalentes de reducción (NADPH, NADH) ytambién de energía (ATP). Este proceso se detiene cuando ya hay suficiente y, además, se han cubierto lasnecesidades de ATP. En este momento, los productos finales de la fase no oxidativa de esta ruta metabólica podránincorporarse en la gluconeogénesis, para formar nuevamente glucosa-6-fosfato y cerrar el ciclo.Por último, hay otro tipo de células, las proliferantes que también se aprovechan de la gran flexibilidad de esteproceso metabólico. Éstas necesitan una gran cantidad de ribosa-5-fosfato para poder sintetizar ácidos nucleicos y,así, replicarse con facilidad y rapidez. De este modo, la ruta puede invertirse, gracias a la reversibilidad de susreacciones y, a partir de una molécula de gliceraldehído-3-fosfato y dos de fructosa-6-fosfato, obtendremos comoproducto tres moléculas de ribosa-5-fosfato, sin formarse ningún NADPH.[3]
Véase también• Deficiencia de glucosa-6-fosfato deshidrogenasa• NADPH
Referencias[1] Kruger NJ, von Schaewen A (June 2003). « The oxidative pentose phosphate pathway: structure and organisation (http:/ / linkinghub.
elsevier. com/ retrieve/ pii/ S1369526603000396)». Curr. Opin. Plant Biol. 6 (3): pp. 236–46. PMID 12753973 (http:/ / www. ncbi. nlm. nih.gov/ pubmed/ 12753973). .
[2] The pentose phosphate pathway (http:/ / www. whfreeman. com/ stryer4/ con_index. htm?22) Biochemistry, Stryer, L. 4th edition[3] http:/ / bloodjournal. hematologylibrary. org/ cgi/ content/ full/ 92/ 7/ 2527 Early Phagocytosis of Glucose-6-Phosphate Dehydrogenase
(G6PD)-Deficient Erythrocytes Parasitized by Plasmodium falciparum May Explain Malaria Protection in G6PD Deficiency]
• Müller Stern, W. (2008)“Bioquímica: Fundamentos para Medicina i Ciencias de la Vida” Ed. Reverté, páginas522-527.

Article Sources and Contributors 24
Article Sources and ContributorsCiclo de Krebs Source: http://es.wikipedia.org/w/index.php?oldid=56634806 Contributors: AC99C5EC.ipt.aol.com, Acratta, Anassesduses, Apcpca, Appuertag, AstroNomo, Banfield, Beto29,BlackBeast, Camilo, Canyq, Carmin, Cecirockea, Cinabrium, CommonsDelinker, Dalton2, Diegusjaimes, Diucón, Dodo, El Moska, Eli22, Eloy, Emiduronte, Emijrp, Emilio Juanatey, F.A.A,Floripaint, Fmariluis, Gonn, HE12, HUB, Ilario, Isha, J. A. Gélvez, Jkbw, Jlosada, Jmcalderon, Jorge 2701, Jorge c2010, Jorgechp, Joseaperez, Juanjolalala, Jurock, KnightRider, Laura Fiorucci,Leonpolanco, Lycaon83, Matdrodes, Mel 23, Mirabilary, Miss Manzana, Montgomery, Moriel, Mutabaruca, Netito777, Nicop, Nosce, Noventamilcientoveinticinco, OboeCrack, Opinador, Oscar., PaulZhiiToO, Pedvi, Petruss, Pólux, Quesete, Rickynoram, Roberto Fiadone, Robpanta, Sahaquiel9102, Sauron, Savh, Shahriyar alavi, Snakeyes, Srbanana, Superzerocool, Svisorweb,Tamorlan, Tomatejc, Urband, VanKleinen, Varano, Vitamine, Will vm, Xuankar, Xvazquez, Yeza, conversion script, 274 anonymous edits
Glucólisis Source: http://es.wikipedia.org/w/index.php?oldid=56658919 Contributors: 3coma14, 4lex, Ahambhavami, Airunp, Alanesalan, Anarcosocialista1, AngelHerraez, Antur, Aozomek,Armenterosa, Baiji, BlackBeast, Boku wa kage, Cookie, Danielrengelm, Dark-lunacy, Delphidius, Derki, Devorador, Dianai, Diegusjaimes, Dreitmen, El Ágora, Eldar, Fiquei, Floripaint, Gregor0492, HUB, Haitike, HernanMC, Humberto, Ialad, Interwiki, Isha, Jean Victor Bernier, Jkbw, Jorge c2010, Joseaperez, Karmillina, Kyorai, L'irie, Leonpolanco, Luckas Blade, Mafores,Maizaisabela, Mar del Sur, MarcoAurelio, Matdrodes, Mel 23, Morquenstein, Netito777, Nioger, Opinador, Ortisa, Pabloes, Poco a poco, Pólux, Queninosta, Retama, Rosarino, RoyFocker,Ruzha, Savh, Schmtterling, Scuellar, Sgpsaros, Skadia, Snakeyes, SuperBraulio13, Superzerocool, Taichi, Tano4595, Tirithel, Tomatejc, Trafek, Triku, UA31, Ugly, VanKleinen, Verdecito,Vitamine, Xtabay, Xuankar, Xvazquez, Yeza, Youssefsan, Zaka, 322 anonymous edits
Gluconeogénesis Source: http://es.wikipedia.org/w/index.php?oldid=56269184 Contributors: Acratta, Ahambhavami, AngelHerraez, AntonioEMM, Ariel ahmed12, BiobulletM, CASF,Centroamericano, Cookie, Diegusjaimes, Fcrespo, GermanX, Halcón, Jlosada, Jmcalderon, MadriCR, Murphy era un optimista, Netito777, Peter-seg, Pitxulin, Pitxulin1, Queninosta, SUL,Schummy, Shahriyar alavi, Tirithel, Trafek, Vic Fede, Xvazquez, 70 anonymous edits
Glucogenolisis Source: http://es.wikipedia.org/w/index.php?oldid=53249282 Contributors: Ahambhavami, Alemany88, Jlosada, Mordekar, Wikiléptico, Xvazquez, 9 anonymous edits
Glucogénesis Source: http://es.wikipedia.org/w/index.php?oldid=56384773 Contributors: Acid burn25, Alemany88, Aparecio12, Camilo, Diegusjaimes, Digigalos, Dodo, El Moska, Elimedina,Gabymoreno, Gerardo Rascón Valverde, Jlosada, Matdrodes, OregMR, OshoX2008, Pilaf, Rjgalindo, Santiperez, Technopat, Wipijnm, Xadjax, Xuankar, Xvazquez, 55 anonymous edits
Ruta de la pentosa fosfato Source: http://es.wikipedia.org/w/index.php?oldid=56505890 Contributors: Ahambhavami, Airunp, BQmUB2009132, Boja, Boku wa kage, Diegusjaimes, Gusgus,Jorge c2010, Matdrodes, Morquenstein, Shahriyar alavi, Susana Márquez, Xvazquez, 20 anonymous edits

Image Sources, Licenses and Contributors 25
Image Sources, Licenses and ContributorsArchivo:Citric acid cycle with aconitate 2-es.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:Citric_acid_cycle_with_aconitate_2-es.svg License: GNU Free DocumentationLicense Contributors: Narayanese, WikiUserPedia, YassineMrabet, TotoBagginsArchivo:Ciclo de Krebs-es.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:Ciclo_de_Krebs-es.svg License: Public Domain Contributors: derivative work: r@ge (talk)Ciclo_de_Krebs.svg: User Minutemen on de.wikipediaArchivo:Alpha-D-Glucopyranose.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:Alpha-D-Glucopyranose.svg License: Public Domain Contributors: NEUROtikerArchivo:Pyruvat.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:Pyruvat.svg License: Public Domain Contributors: NEUROtikerFile:Glucólisis.png Source: http://es.wikipedia.org/w/index.php?title=Archivo:Glucólisis.png License: Creative Commons Attribution 3.0 Contributors: YassineMrabet (usuario WikipediaArchivo:Reaction-Glucose-Glucose-6P oc.png Source: http://es.wikipedia.org/w/index.php?title=Archivo:Reaction-Glucose-Glucose-6P_oc.png License: Copyrighted free use Contributors:SolonArchivo:Biochem reaction arrow foward NNNN horiz med.png Source: http://es.wikipedia.org/w/index.php?title=Archivo:Biochem_reaction_arrow_foward_NNNN_horiz_med.png License: GNU Free Documentation License Contributors: Richard Wheeler (Zephyris)Archivo:Reaction-Glucose-6P-Fructose-6P oc.png Source: http://es.wikipedia.org/w/index.php?title=Archivo:Reaction-Glucose-6P-Fructose-6P_oc.png License: Copyrighted free use Contributors: User:SolonArchivo:Biochem reaction arrow reversible NNNN horiz med.png Source: http://es.wikipedia.org/w/index.php?title=Archivo:Biochem_reaction_arrow_reversible_NNNN_horiz_med.png License: GNU Free Documentation License Contributors: Richard Wheeler (Zephyris)Archivo:Reaction-Fructose-6P-F16BP oc.png Source: http://es.wikipedia.org/w/index.php?title=Archivo:Reaction-Fructose-6P-F16BP_oc.png License: Copyrighted free use Contributors:User:SolonArchivo:Reaction-F16BP-DOAP-GA3P unlabel.png Source: http://es.wikipedia.org/w/index.php?title=Archivo:Reaction-F16BP-DOAP-GA3P_unlabel.png License: Copyrighted free use Contributors: User:SolonArchivo:Reaction DHAP to GAP.png Source: http://es.wikipedia.org/w/index.php?title=Archivo:Reaction_DHAP_to_GAP.png License: Copyrighted free use Contributors: SolonArchivo:D-Glycerinaldehyd-3-phosphat2.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:D-Glycerinaldehyd-3-phosphat2.svg License: Public Domain Contributors:NEUROtikerArchivo:GG-Pfeil 1-4.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:GG-Pfeil_1-4.svg License: Public Domain Contributors: GG-Pfeil_1-3.svg: NEUROtiker derivative work:Matt (talk)Archivo:D-1,3-Bisphosphoglycerat2.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:D-1,3-Bisphosphoglycerat2.svg License: Public Domain Contributors: NEUROtikerArchivo:D-3-Phosphoglycerat2.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:D-3-Phosphoglycerat2.svg License: Public Domain Contributors: NEUROtikerArchivo:GG-Pfeil_1.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:GG-Pfeil_1.svg License: Public Domain Contributors: NEUROtikerArchivo:D-2-Phosphoglycerat2.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:D-2-Phosphoglycerat2.svg License: Public Domain Contributors: NEUROtikerArchivo:Phosphoenolpyruvat Fischer2.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:Phosphoenolpyruvat_Fischer2.svg License: Public Domain Contributors: NEUROtikerArchivo:R-Pfeil rechts 1-3.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:R-Pfeil_rechts_1-3.svg License: Public Domain Contributors: NEUROtikerArchivo:Pyruvat Fischer.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:Pyruvat_Fischer.svg License: Public Domain Contributors: NEUROtikerFile:Regulación glucolisis.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:Regulación_glucolisis.svg License: Creative Commons Attribution-Sharealike 3.0 Contributors:User:Gregor 0492Archivo:Commons-logo.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:Commons-logo.svg License: logo Contributors: SVG version was created by User:Grunt and cleanedup by 3247, based on the earlier PNG version, created by Reidab.Imagen:Glucogenesis-es.jpg Source: http://es.wikipedia.org/w/index.php?title=Archivo:Glucogenesis-es.jpg License: Creative Commons Attribution-Sharealike 3.0 Contributors: BiobulletMArchivo:Oxidative phase of pentose phosphate pathway esp.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:Oxidative_phase_of_pentose_phosphate_pathway_esp.svg License: Creative Commons Attribution-ShareAlike 3.0 Unported Contributors: XvazquezArchivo:Non-oxidative phase of pentose phosphate pathway.svg Source: http://es.wikipedia.org/w/index.php?title=Archivo:Non-oxidative_phase_of_pentose_phosphate_pathway.svg License: Creative Commons Attribution-ShareAlike 3.0 Unported Contributors: Andrzej Małkowski

Licencia 26
LicenciaCreative Commons Attribution-Share Alike 3.0 Unported//creativecommons.org/licenses/by-sa/3.0/