supplementary information - media.nature.com · supplementary information . an orthogonalized...
TRANSCRIPT

Supplementary Information
An orthogonalized platform for genetic code expansion in both bacteria and eukaryotes
James S. Italia,1 Partha Sarathi Addy,1 Chester J.J. Wrobel,1 Lisa A. Crawford,1 Marc J. Lajoie,2 Yunan Zheng1 and Abhishek Chatterjee1*
1Department of Chemistry, Boston College, 2609 Beacon Street, Chestnut Hill, MA 02467, USA
2Department of Biochemistry, University of Washington, Seattle, WA 98195, USA.
*email: [email protected]
S1
Nature Chemical Biology: doi:10.1038/nchembio.2312

Supplementary Figures:
Supplementary Results
Supplementary Figure 1. (a) Comparing the number of UAAs genetically encoded using the E. coli based selection system relative to those using a yeast based selection system. Note: Nearly half of the UAAs genetically encoded using the yeast selection system use a single E. coli leucyl-tRNA synthetase variant that exhibits high polyspecificity towards structurally homologous substrates.1 (b) Number of UAAs genetically encoded in eukaryotes each year from 2003 to 2014 using the three established tRNA/aRS pairs: bacterial tyrosyl, bacterial leucyl, and the archaeal pyrrolysyl systems. The pyrrolysyl system has been exclusively used to genetically encode new UAAs in eukaryotes in the last six years. The numbers were compiled from the reviews by Liu et al.2and Dumas et al.1
S2
Nature Chemical Biology: doi:10.1038/nchembio.2312

Fluo
resc
ence
-aaRS +aaRS
Supplementary Figure 2. Fluorescence microscopy images showing the expression of EGFP-39-TAG reporter in HEK293T cells co-expressing tRNAEcTrp
CUA, in the presence and the absence of its cognate EcTrpRS. Robust reporter expression is only observed when the EcTrpRS is co-expressed, confirming the lack of cross-reactivity of tRNAEcTrp
CUA in these cells.
S3
Nature Chemical Biology: doi:10.1038/nchembio.2312

Supplementary Figure 3. Map of the plasmid used to complement the deletion of the endogenous E. coli Trp-tRNA/aaRS pair. pUltraG-ScW40CCA contains a glnS’ promoted wild-type yeast TrpRS, proK promoted engineered yeast Trp-tRNACCA,3 CloDF13 origin of replication, and a spectinomycin selectable marker.
S4
Nature Chemical Biology: doi:10.1038/nchembio.2312

Supplementary Figure 4. Colony PCR based verification of successful genomic alterations by λ-Red recombineering. The expected PCR product size for desired recombinant E. coli are shown using green arrows, while the same for the precursor E. coli strain is shown in red. Locations of primers to verify each recombination reaction are shown in the context of a map of the corresponding genomic locus, and the specific primer-combination used is indicated above each gel-image. (a) Replacing EcTrpRS with zeocin
S5
Nature Chemical Biology: doi:10.1038/nchembio.2312

resistance cassette: Lanes 1-6 are potential hits, lane 7 is EcNR1, and lane 8 is no E. coli control. (b) tRNAEcTrp
CCA (trpT) replaced with gentamycin resistance cassette: Successful hits will have a larger PCR product due to the introduction of the larger gentamycin cassette. Lanes 1-4 are potential hits, lane 5 is the precursor strain (EcNR1 pUltraG_ScW40CCA trpS::zeoR), and lane 6 is no E. coli control. (c) Deletion of endogenous galK: Successful hits will have a smaller PCR product. Lanes 1-8 are potential hits, lane 9 is the precursor strain, lane 10 is an E. coli strain where the galK was previously deleted using a similar strategy, and lane 11 is no E. coli control. (d) Deletion of the genomic λ-Red machinery using a galK cassette to generate ATMW1: Two different primer combinations are used. (A): Forward primer (dLambda-F) anneals 150bp upstream of the deletion site and the reverse primer anneals only to an internal site of galK (galK-iR). (B): Forward (dLambda-F) and reverse (dLambda-R) primers anneal 150bp upstream and downstream from the deletion site. 1-4 are successful ATMW1 hits, 5 is EcNR1 (precursor strain that has the λ-Red machinery), 6 is Top10 (negative control, does not have the λ-Red machinery), and 7 is PBS. The large size of the λ-Red machinery prevents the formation of a PCR product when using the primer combination (B): dLambda-F+ dLambda-R. The products from successful recombinants (1-4) are faint but clearly present.
S6
Nature Chemical Biology: doi:10.1038/nchembio.2312

Supplementary Figure 5. (a) ESI-MS analysis of sfGFP-151-TAG purified from ATMW1 co-expressing tRNAEcTrp
CUA alone. The observed mass (27562) is consistent with the incorporation glutamine, glutamate, or a lysine residue at the nonsense codon. Tryptic digestion of this protein followed by LC- MS/MS analysis of the resulting peptides confirms the incorporation of glutamine in response to TAG (shown in panel c). (b) ESI-MS analysis of sfGFP-151-TGA purified from ATMW1 co-expressing wild- type EcTrpRS and tRNAEcTrp
UCA. The mass is consistent with the incorporation of tryptophan in response to TGA.
S7
Nature Chemical Biology: doi:10.1038/nchembio.2312

CUG
Supplementary Figure 6. Predicted secondary structures (a) tRNAEcTrpCCA (trpT), and (b) tRNAEcGln
(glnV). Highlighted in glnV structure (with a red circle) is the U residue in the anticodon that has been shown to be a major identity element. While this U is absent in wild-type trpT, creating a TAG suppressor variant of this tRNA results in its introduction. (c) Homology alignment of these two tRNA sequences.
S8
Nature Chemical Biology: doi:10.1038/nchembio.2312

Supplementary Figure 7. While TGA termination is leaky in wild-type E. coli, it is not the case in ATMW1. Three chloramphenicol acetyl transferase (CAT) mutants, harboring TGA codons at three different positions, were expressed individually in ATMW1 or EcNR1 (the precursor strain for ATMW1), in the absence of any exogenous suppressor tRNAs, and the ability of the bacteria to survive increasing concentrations of chloramphenicol was tested. ATMW1 strains showed much lower tolerance to chloramphenicol, indicating significantly lower levels of TGA read-through.
S9
Nature Chemical Biology: doi:10.1038/nchembio.2312

Supplementary Figure 8. Evolution of EcTrpRS to create mutants capable of charging 5HTP (1; Figure 4a). (a) Structure of Geobacillus stearothermophilus TrpRS (PDB 1I6M) active site, showing the active site residues chosen for randomization to alter its substrate specificity. (b) Mutations associated with 5HTP-specific EcTrpRS variants.
S10
Nature Chemical Biology: doi:10.1038/nchembio.2312

Supplementary Figure 9. Deconvoluted whole-protein mass-spectrometry analysis of sfGFP reporters indicate the absence of TGG mischarging by tRNAEcTrp
UCA in ATM E. coli. Panels (a)-(d) represent sfGFP-151-TGA (harbors one TGG codon) co-expressed with EcTrpRS-h14/tRNAEcTrp
UCA in the presence of 1 mM 5HTP, 5MTP, 5PrW, or 5BrW, respectively. Panels (e) and (f) represent sfGFP-151-TGG (harbors two TGG codons) co-expressed with EcTrpRS-h14/tRNAEcTrp
UCA in the absence, or presence of 1 mM 5PrW. While the presence of low levels of oxidized protein species (+16, +32, +48) complicates the detection of potential TGG-mischarging product (expected mass for the single misincorporation product is indicated with a red arrow in each case) for 5HTP (a) or 5MTP (b), the absence of such misincorporation product can be confirmed for 5PrW (c and f), and 5BrW (d).
S11
Nature Chemical Biology: doi:10.1038/nchembio.2312

Supplementary Figure 10. Fluorescence microscopy images showing the expression of EGFP-39-TGA reporter in HEK293T cells co-expressing tRNAEcTrp
UCA, in the presence and the absence of its cognate EcTrpRS. Robust reporter expression is only observed when the EcTrpRS is co-expressed, confirming the lack of cross-reactivity of tRNAEcTrp
UCA in these cells.
S12
Nature Chemical Biology: doi:10.1038/nchembio.2312
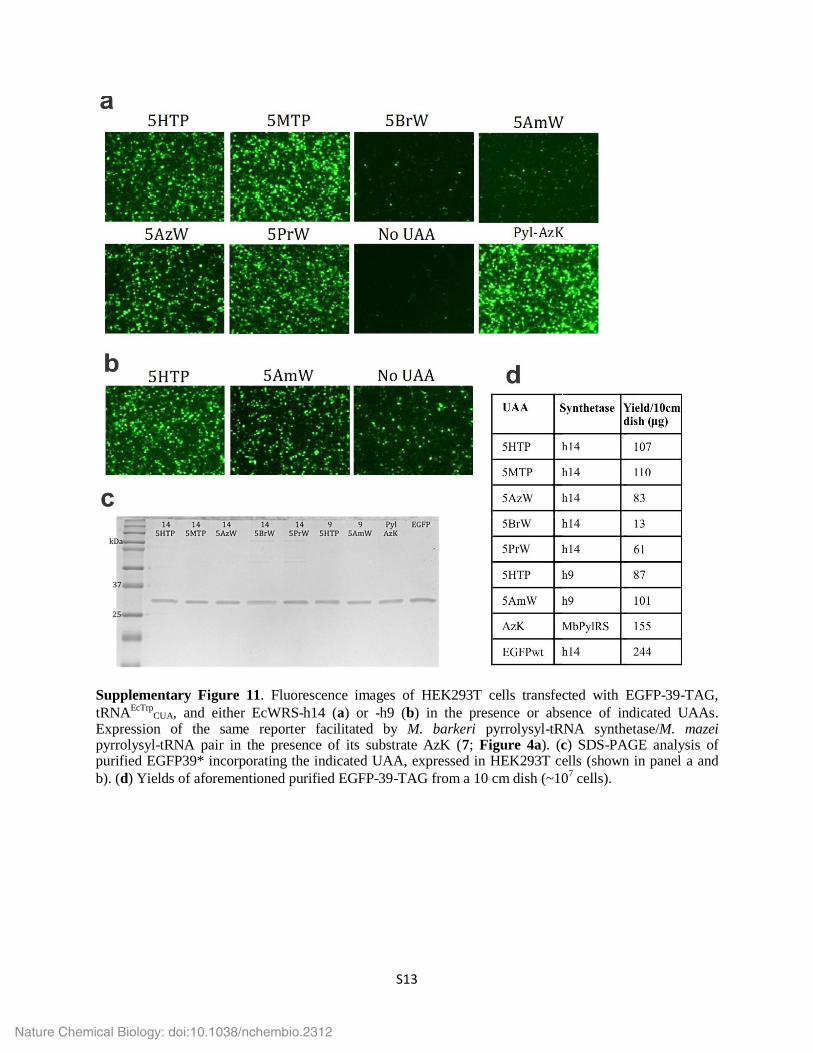
Supplementary Figure 11. Fluorescence images of HEK293T cells transfected with EGFP-39-TAG, tRNAEcTrp
CUA, and either EcWRS-h14 (a) or -h9 (b) in the presence or absence of indicated UAAs. Expression of the same reporter facilitated by M. barkeri pyrrolysyl-tRNA synthetase/M. mazei pyrrolysyl-tRNA pair in the presence of its substrate AzK (7; Figure 4a). (c) SDS-PAGE analysis of purified EGFP39* incorporating the indicated UAA, expressed in HEK293T cells (shown in panel a and b). (d) Yields of aforementioned purified EGFP-39-TAG from a 10 cm dish (~107 cells).
S13
Nature Chemical Biology: doi:10.1038/nchembio.2312

Supplementary Figure 12. Structures of DBCO-Cy5, and Alexa488-PCA.
S14
Nature Chemical Biology: doi:10.1038/nchembio.2312

Supplementary Figure 13. Expanded images of the SDS-PAGE analyses described in this report. The figure number in the manuscript corresponding to each expanded image is mentioned below each image.
S15
Nature Chemical Biology: doi:10.1038/nchembio.2312

Supplementary Tables:
Supplementary Table 1. ESI-MS analysis of reporter proteins isolated from ATMW1 or HEK293T cells. Reporter Amino
acid aaRS Expecte
d mass Observed
mass Note
sfGFP-151TAG Gln none 27620 27562 pBK/tac system in ATMW1 (Gln x-reactive)
sfGFP-151-TGA Trp EcWRSwt 27620 27621 pBK/tac system in ATMW1 sfGFP-151-TGA 5HTP h14 27636 27636 pBK/tac system in ATMW1 sfGFP-151-TGA 5MTP h14 27652 27650 pBK/tac system in ATMW1 sfGFP-151-TGA 5BrW h14 27700 27699 pBK/tac system in ATMW1 sfGFP-151-TGA 5AzW h14 27663 27660 pBK/tac system in ATMW1 sfGFP-151-TGA 5PrW h14 27676 27674 pBK/tac system in ATMW1 sfGFP-151-TGA 5HTP h9 27636 27637 pBK/tac system in ATMW1 sfGFP-151-TGA 5AmW h9 27636 27635 pBK/tac system in ATMW1 sfGFP-151-TGG Trp h14 27620 27620 pBK/tac system in ATMW1
EGFPwt Tyr None 29683 29683 pAcBac1 system in HEK293T EGFP-39-TAG BocK MbPyl 29748 29748 pAcBac1 system in HEK293T EGFP-39-TAG AzK MbPyl 29761 29761 pAcBac1 system in HEK293T EGFP-39-TAG Trp EcWRSwt 29707 29708 pAcBac1 system in HEK293T EGFP-39-TAG 5HTP h14 29723 29724 pAcBac1 system in HEK293T EGFP-39-TAG 5MTP h14 29736 29736 pAcBac1 system in HEK293T EGFP-39-TAG 5BrW h14 29786 29785 pAcBac1 system in HEK293T EGFP-39-TAG 5AzW h14 29748 29747 pAcBac1 system in HEK293T EGFP-39-TAG 5PrW h14 29761 29760 pAcBac1 system in HEK293T EGFP-39-TAG 5HTP h9 29723 29724 pAcBac1 system in HEK293T EGFP-39-TAG 5AmW h9 29722 29721 pAcBac1 system in HEK293T
sfGFP-151-TAG 5AzW h14 28368 28366 Labeled with DBCO-biotin EGFP-39-TAG 5AzW h14 30453 30453 Labeled with DBCO-biotin
S16
Nature Chemical Biology: doi:10.1038/nchembio.2312
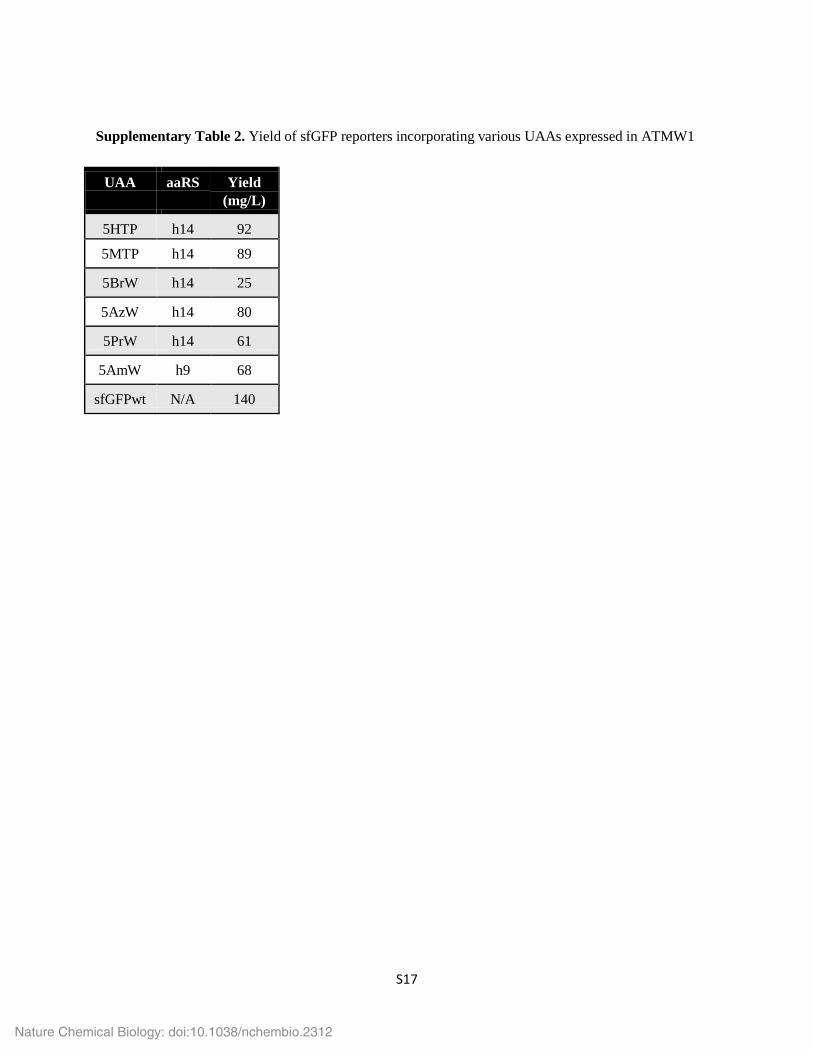
Supplementary Table 2. Yield of sfGFP reporters incorporating various UAAs expressed in ATMW1
UAA aaRS Yield (mg/L)
5HTP h14 92
5MTP h14 89
5BrW h14 25
5AzW h14 80
5PrW h14 61
5AmW h9 68
sfGFPwt N/A 140
S17
Nature Chemical Biology: doi:10.1038/nchembio.2312

Supplementary Table 3. Oligonucleotides TrpRS.Z.ab-F ATCAGTCTATAAATGACCTTCTGCCCGCATTAGGGCTTCCGCATAGCGAAAATCAGGAA
TCGAAAAAGGTGTTGACAATTAATCATCGGC TrpRS.Z.ab-R TGTAGGCCGGATAAGGCGTTCACGCCGCATCCGGCATGAACAAAGCGCAATTTGCCAG
CAATAGTGAAAGCTTGCAAATTAAAGCCTTCG TrpRS150F GTCGGCGACTCACGCAATGATATTCAGGCGGC TrpRS150R AGCGAGATGTGGAAACGGCGAGGCACTTCAC
Zeo-iR CTGGTCCTGGACCGCGCTGATGAAC TrpRS_sqiR ATCCTGGCGCACGGTGATCGCGTGTTG
trpTKO.Gent-F CAGTCGGTTAGAATACCTGCCTGTCACGCAGGGGGTCGCGGGTTCGAGTCCCGTCCGTT CCGCCACCCTAATTACGCACACCGTGGAAAC
trpTKO.Gent-R CGGTAGAAGGATTTACTTCGGAGAGGGTTATTTCAGATAAAAAAAATCCTTAGCTTTCG CTAAGGATGATTTCCCGGGAAGCCGATCTCG
trpT GsqF GGGGTCTCCCCATGCGAGAGTAGGGAAC trpT GsqR CCGTTGTCGATAGCACAACACTTTCACGGCC
galK.90 del CGCGCAGTCAGCGATATCCATTTTCGCGAATCCGGAGTGTAAGAACGCGCAGTCAGCG ATATCCATTTTCGCGAATCCGGAGTGTAAGAA
galK_KO_verf_F TGGCAGAGACCCAGCG galK_KO_verf_R ACCCCAGTCCATCAGCG dLambda.galK-F GCTATGAAATAGAAAAATGAATCCGTTGAAGCCTGCTTTTTTATACTAACTTGAGCGAA
ACGGGAAGCCTGTTGACAATTAATCATCGGC dlambda.galK dterm-R GCCGCGTTGATTTTCTCCTGCCAGCTCATAATGCTGCCGCGTTGTAATATTCAGCACTGT
CCTGCTCCTT dlambda.sqF GGTTTGATCAGAAGGACGTTGATCGGGCGG dlambda.sqR TTCAGATACTGGCGATCATCCGCCACCAG
dLambda.sqiR AGCCCATTGATAGTTTTCATGTGCGACAATGGGCG EcWRS_mut7_8-F GAATCCCATATGATGACTAAGCCCATCGTTNBTNSTGGCGCACAGCCCTCAGGTGAATT
G libEcWRS-NdeI-F TACGCTTTGAGGAATCCCATATGATGACTAAGCCCATCG
EcWRS1_mut-VPViR CAGATTAGTTTGATACAGCAGGATGTCCGCTGCCATC EcWRS1_mut144-6_F GATGGCAGCGGACATCCTGCTGTATCAAACTAATCTGnnknnknnkGGTGAAGACCAGAAA
CAGCACCTCGAACTGAGC EcWRS_NcoI_PstI_termR agcgtttgaaactgcagccatggtaccTTACGGCTTCGCCACAAAACCAATCGC
proK-F GTTAGCCTGCAGGTAATTCCGCTTCGCAACATGTGAG TrpH NcoI-R GGCCGCCATGGCAAATTCGACCCTG Trp40CCA-iR GCAACCAGGCGCTTTGGAGGCGCCAGCTCTACCCTTGAG Trp40CCA-iF AGCTGGCGCCTCCAAAGCGCCTGGTTGCAGGTTC
SmR-R CGCGCGCAGATCAGTTGGAAGAATTTGTCCACTACGTG pNP-SpeI-F ATAATGGACTAGTGCGCTTGTTTCGGC pNP-BAB-R CTCCTTAGATCTTCCTAGGTGGATCCACCATTCC
pEvol CmR SpeI-F AATAATACTAGTGTTGATACCGGGAAGCCCTGG pEvol_CmR PstI-R AATAATCTGCAGCGAATTTCTGCCATTCATCCGCTTATTATCAC
CmR-TGA-7 GCTAAGGAAGCTAAAATGGAGAAAAAAATCACTTGATATACCACCGTTGATATATCCC AATGGC
CmR-TGA-84 GCAATGAAAGACGGTGAGCTGGTGTGATGGGATAGTGTTCACCCTTGTTACACC CmR-TGAT-98 CCCTTGTTACACCGTTTTCCATGAGTGATCTGAAACGTTTTCATCGCTCTGGAG pRep-KpnI-tR-F AATAATaggtaccGTTCTGTTGCCCGTCTCACTGGTG
pRep-EcWtR-NdeI/AvrII-R. AATAATAcatatgCCTAGGTGGCAGGGGCGGAGAGACTC
EcW-TGA-MSDM GTTCAATTGGTAGAGCACCGGTCTTCAAAACCGGGTGTTGGGAGTTCGAG T7F1 CAGGTTCGCAGCGTCAGCCGGAATGGTACCG T7R3 GCGCCCGACAGCCTTCCAGTTCCTGTGAGAAATCAAGCCGGAAGCCGTAGCGTAC T7F3 GTACGCTACGGCTTCCGGCTTGATTTCTCACAGGAACTGGAAGGCTGTCGGGCGC
T7R4b CCATGACCATGATTACCGTGCACTGAAATACCATTAACATTGCTAAGAACG T7F4 CGTTCTTAGCAATGTTAATGGTATTUCAGTGCACGGTAATCATGGTCATGG T7R5 CGAAGGCGAAGCGGCATGCATAATGTGCCTGTCAAATGG
T7_mut-SbfI-F AATAATcctgcaggCTACTCAGGAGAGCGTTCACCGAC T7_mut-NotI/SbfI-R AATAATCCTGCAGGGCGGCCGCTACGGGAGGGCTTACCATCTGG
Barnase MSDM 3 TGA TTTAACTTTAAGAAGGAGATATACATATGGCATgaGTTATCAACACGTTTGACGGGGTTG Barnase MSDM 45 TGA GTGGCATCAAAAGGGAACCTTGCATGAGTCGCTCCGGGGAAAAGCATC
EcWtR-PstI-F TTAGCCTGCAGTGTGCTTCTCAAATGCCTGAGGCCAGTTTGCTC EcWtR-prok-oF GCGCCCCGCATTTAGGGGCGTAGTTCAATTGGTAGAGCACCGGTC EcWtR-proK-oR ACTACGCCCCTAAATGCGGGGCGCATCTTACTGCGC EcWtR-KpnI-R ATATATGGTACCAAAAAATGGCAGGGGCGGAGAGACTCG
GFPflip-NotI-F + TCGATCCCGCGAAATTAATACGACTCACTATAG sfGFP-T7+lam-PstI-R. ATATACTGCAGCGCCAAGCTAGCTTGGATTCTCACCAATAAAAAACGC
S18
Nature Chemical Biology: doi:10.1038/nchembio.2312

MjYtR del F TGGCAGGGGCGGAGAGACTCGAACTCC MjYtR del oR CGAGTCTCTCCGCCCCTGCCA AATTCGAAAAGCCTGCTCAACGAGCAGG
EcWtR TGA MSDM GTTCAATTGGTAGAGCACCGGTCTTCAAAACCGGGTGTTGGGAGTTCGAG sfGFP (pEvol) TGA151 CTCGAGTACAACTTTAACTCACACAATGTATGAATCACGGCAGACAAACAAAAGAATG
G EcWRS1.FA.NotI-F AATAATAgcggccgcATGACTAAGCCCATCGTTTTTGCTGGCGCAC
EcWRS-NotI-R AATAATAgcggccgcTTACGGCTTCGCCACAAAACCAATCGC pUltraII-tRsqR GGTGCCCTTAAACGCCTGGTTGC
EcTrpRS-NdeI-F AAtAAAcatatgATGACTAAGCCCATCGTTTTTAGTGGCGCAC EcTrpRS-PstI-R TTATTCTGCAGTTACGGCTTCGCCACAAAACCAATCGC TrpRS-R EcoRI ATTATTGAATTCTTACGGCTTCGCCACAA TrpRS-F-NheI AATAAATGCTAGCATGACTAAGCCCATC
U6-R tRNAtrp* AvrII AATTATTGCTAGCAAAAAATGGCAGGGGCG tRNAtrp* Nhe-R AATTATTGCTAGCAAAAAATGGCAGGGGCG
EcWRS_mamNheI-F
aataataGCTAGCgccaccATGACTAAGCCCATCGTT
EcWRS_mamEcoRI-R AATAATAgaattcTTACGGCTTCGCCACAAAACCAATCGC
Supplementary Note 1: Synthesis of 5PrW and associated characterization data. Supplementary Note 2: Sequences for plasmids and other gene cassettes.
References: 1 Dumas, A., Lercher, L., Spicer, C. D. & Davis, B. G. Designing logical codon reassignment–
Expanding the chemistry in biology. Chem. Sci. 6, 50-69 (2015). 2 Liu, C. C. & Schultz, P. G. Adding new chemistries to the genetic code. Annu. Rev. Biochem. 79,
413-444 (2010). 3 Ellefson, J. W. et al. Directed evolution of genetic parts and circuits by compartmentalized
partnered replication. Nat. Biotechnol. 32, 97-101 (2014).
S19
Nature Chemical Biology: doi:10.1038/nchembio.2312