project - matej znidaric
TRANSCRIPT

Role of NOPS proteins in the regulation of postsynaptic structure Matej Znidaric; ETH Zürich
October 2011 – June 2012
Mentors: dr. Steven Brown
Dennis Mircsof

Matej Znidaric Role of NOPS proteins in the regulation of postsynaptic structure ETH, UZH Zürich [email protected] Neuroscience
1
Summary
Synapses are very dynamic systems that play key role in neuronal functions like information processing and transmission. One of the major proteins in the inhibitory synapses is structural protein gephyrin, since it anchors GABA type-A and glycin receptors to the synaptic membrane and links them on the microtubules. We studied the effect of DBHS family of proteins (NONO, SFPQ and PSPC1) on the gephyrin clustering in-vitro in the rats’ primary hippocampal neurons, using biochemical assays and confocal microscopy. mCherry linked DBHS proteins as well as PSPC1 and SFPQ RNA recognition mutants were cloned and successfully expressed in HEK-293T and neuron cell lines. In addition, localization of cloned proteins in HEK cells was intranuclear and it did not change compared to endogenous protein. In neurons RRM mutants were found outside the nucleus and number of gephyrin clusters in the dendrites was reduced. Furthermore, there was a tendency toward larger clusters compared to non-mutated PSPC1 or SFPQ cloned proteins or negative control. With our study, phenotypic change in case of mCherry-PSPC1 RRM and mCherry-SFPQ RRM mutant, was shown but the mechanisms and underlying processes are yet to be described.
Introduction
Entire nervous system of the mammals is composed from basically 2 broad types of cells; glia cells and neurons. Neurons transmit and compute information by changing their trans-membrane potential. Glia on the other hand support and nourish neurons but their role in information processing is not yet understood (Purves 2012).
Neurons are dynamic, polarized cells in the nervous system. Their morphology is extremely diverse and directly reflects function in the neuronal network (Purves 2012). They connect to each other via specialised structures known as synapses. There is large complexity within the synapses with several hundred proteins involved in the process of information transduction and therefore they represent metabolically very demanding parts of the cell (Kirsch 1996; Purves 2012). Cell keeps in phase with metabolic demand with localized protein synthesis at polyribosomes, which stretch into the neurites. For their development, function and plasticity of the synapse, structural proteins that cluster membrane receptors play a crucial role (Prior and Schmitt 1992; Froehner 1993; Kirsch 1996).
In the inhibitory synapses, structural protein gephyrin anchors type-A receptor for the neurotransmitter GABA (γ-aminobutyric acid) and Glycin receptors (Gly-R), on the membrane (Froehner 1993; Kirsch 1996). 2 isoforms of gephyrin are known. Gephyrin protein has two major functional domains, E and G, linked by an C-domain. C-domain contains regulatory sites and binding sites for interacting proteins; it was shown that it binds to the microtubules (Prior and Schmitt 1992; Froehner 1993; Stanley 1993; Kirsch 1996; Wang, Bedford et al. 1999; Fritschy, Harvey et al. 2008) and several other synaptic proteins, namely: neuroligin and colibistin (Fritschy, Panzanelli et al. 2012). Post-translational modifications especially phosphorylation and acetylation of gephyrin were shown to be important for synaptic plasticity mechanisms (Wang, Bedford et al. 1999). Gephyrins structural significance for inhibitory synapses is reflected with in-vivo knock out study, where mice lacking the protein are not viable, thus making gephyrin an appropriate model protein to study effects on inhibitory synapses.

Matej Znidaric Role of NOPS proteins in the regulation of postsynaptic structure ETH, UZH Zürich [email protected] Neuroscience
2
DBHS family (Drosophila Behavior Human Splicing) of proteins consist of 3 members; NONO/nrb54, PSPC1/PSP1 and SFPQ/PSF. 2 RNA recognition motifs (RRM) and coiled-coil domain responsible for protein-protein interactions are common in all members of the family (Fox and Lamond 2010; Passon, Lee et al. 2012). In the nucleus they function as structural proteins of intra-nuclear bodies named paraspeckles (Fox, Lam et al. 2002; Bond and Fox 2009; Fox and Lamond 2010). They have function in RNA production and processing; transcriptional initiation, termination, alternative splicing and DNA repair (Shav-Tal and Zipori 2002). NONO was also shown to function as a transcriptional factor of clock genes (Brown, Ripperger et al. 2005) and together with SFPQ they bind and retain hyper-adenosine-inosine (A-to-I) edited RNA within the nucleus (Shav-Tal and Zipori 2002; Prasanth, Prasanth et al. 2005). It is assumed that phosphorylation status of the protein may play a role in its functions (Bruelle, Bedard et al. 2011; Sumanasekera, Kelemen et al. 2012). In addition, there was evidence that SFPQ and NONO have a role in cytoplasm of the dendrites in the RNA-transport granule (Kanai, Dohmae et al. 2004).
Aim of the project was to determine if DBHS proteins have an effect on gephyrin protein and consequently on synapse form and function. We tried to determine if the mutations in the RRMs motifs in the DBHS proteins cause a phenotypic change in the system. In order to acquire the data the constructs had to be cloned first. Confocal imaging was then used to visualize and gather the data.

Matej Znidaric Role of NOPS proteins in the regulation of postsynaptic structure ETH, UZH Zürich [email protected] Neuroscience
3
Results
Cloning of the constructs:
DBHS proteins were cloned in the vector containing mCherry protein. PSPC1, PSPC1 RRM mutant, SFPQ and SFPQ RRM mutant were amplified from vector containing myc tag using designed primers yielding in products PSPC1 and PSPC1 RRM (1582 bp) (figure 1a), SPFQ and SFPQ RRM (2110 bp) (Figure 1b). Test digestion of proteins in mCherry containing plasmid showed expected sizes of inserts; (Figure 2).
Figure 1: electrophoresis gel of PSPC1 / PSPC1-RRM (a) and SFPQ / SFPQ RRM (b) amplified with PCR from vector containing myc tag. Both amplified parts are appropriate size; PSPC1 / PSPC1 RRM (1582bp) and SFPQ / SFPQ RRM (2110bp)
Biochemical assay
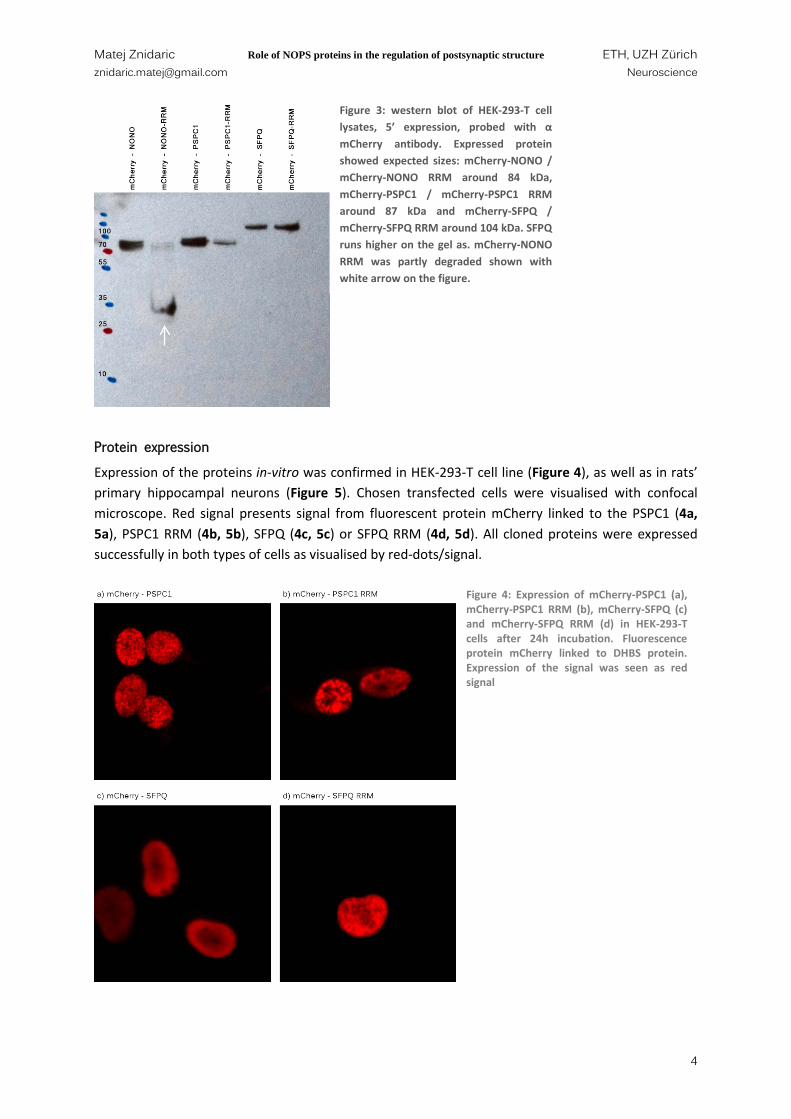
Wesern blot was performed to check the expression of cloned constructs in HEK-293-T cell line. All DBHS proteins and their RRM mutants (mCherry-NONO, mCherry-NONO RRM, mCherry-PSPC1, mCherry-PSPC1 RRM, mCherry-SFPQ and mCherry-SFPQ RRM respectively) were expressed as seen on Figure 3. Sizes of the proteins match expected sized of the protein summed with fluorescent protein mCherry; mCherry-NONO and mCherry-NONO RRM 84 kDa, mCherry-PSPC1 and mCherry PSPC1 RRM 87 kDa, mCherry-SFPQ and mCherry-SFPQ RRM 103 kDa but they run as 20 kDa higher as shown previously (Kuwahara, Ikei et al. 2006).
Figure 2: test digestion of PSPC1/PSPC1 RRM in mCherry vector, cut with Sac1, BamH1 (a) and SFPQ/SFPQ RRM in mCherry vector, cut with Kpn1, Sac1 (b). Fragments show expected sizes; PSPC1 / PSPC1 RRM (1582bp) and SFPQ / SFPQ RRM (2110bp)
a) b)
Matej Znidaric Role of NOPS proteins in the regulation of postsynaptic structure ETH, UZH Zürich [email protected] Neuroscience
4
Protein expression
Expression of the proteins in-vitro was confirmed in HEK-293-T cell line (Figure 4), as well as in rats’ primary hippocampal neurons (Figure 5). Chosen transfected cells were visualised with confocal microscope. Red signal presents signal from fluorescent protein mCherry linked to the PSPC1 (4a, 5a), PSPC1 RRM (4b, 5b), SFPQ (4c, 5c) or SFPQ RRM (4d, 5d). All cloned proteins were expressed successfully in both types of cells as visualised by red-dots/signal.
Figure 3: western blot of HEK-293-T cell lysates, 5’ expression, probed with α mCherry antibody. Expressed protein showed expected sizes: mCherry-NONO / mCherry-NONO RRM around 84 kDa, mCherry-PSPC1 / mCherry-PSPC1 RRM around 87 kDa and mCherry-SFPQ / mCherry-SFPQ RRM around 104 kDa. SFPQ runs higher on the gel as. mCherry-NONO RRM was partly degraded shown with white arrow on the figure.
Figure 4: Expression of mCherry-PSPC1 (a), mCherry-PSPC1 RRM (b), mCherry-SFPQ (c) and mCherry-SFPQ RRM (d) in HEK-293-T cells after 24h incubation. Fluorescence protein mCherry linked to DHBS protein. Expression of the signal was seen as red signal

Matej Znidaric Role of NOPS proteins in the regulation of postsynaptic structure ETH, UZH Zürich [email protected] Neuroscience
5
Figure 5: expression of mCherry-PSPC1 (a), mCherry-PSPC1 RRM (b), mCherry-SFPQ (c) and mCherry-SFPQ RRM (d) in primary rats’ hippocampal neurons. Fluorescence protein mCherry linked to DHBS protein. All cloned constructs were expressed successfully as visualised by red dots/signal. In case a) and c) images were taken on 2 channels; for mCherry and nuclear stain; DAPI.

Matej Znidaric Role of NOPS proteins in the regulation of postsynaptic structure ETH, UZH Zürich [email protected] Neuroscience
6
Colocalization results
HEK cells were transfected with mCherry-DBHS protein as well as with same myc-DBHS (stained with CY5 – blue signal) protein. Additionally, endogenous DBHS (PSPC1 in case of mCherry-PSPC1 and mCherry-PSPC1 RRM or SFPQ in case of mCherry-SFPQ and mCherry-SFPQ RRM) protein was visualised with Alexa 488 – green signal (Figure 6). Localization of endogenous protein overlaps with localization of myc linked protein and mCherry linked protein. Both PSPC1 and SFPQ proteins are localized intra-nucelarally in HEK cells. These results confirm that although overexpressed, both myc- bound as well as mCherry- bound DBHS protein retain their localization.
Figure 6: localization of mCherry-PSPC1 and myc-PSPC1 (a), mCherry-PSPC1 RRM and myc-PSPC1 RRM (b), mCherry-SFPQ and myc-SFPQ (c) and mCherry-SFPQ RRM and myc-SFPQ RRM (d) in primary rats’ hippocampal neurons. In a) and b) endogenous PSPC1 and in c) and d) endogenous SFPQ was stained (green). Fluorescence protein mCherry linked to DHBS protein (red) and myc-DHBS protein (blue). All transfected constructs were expressed successfully as visualised by different colours on the picture.

Matej Znidaric Role of NOPS proteins in the regulation of postsynaptic structure ETH, UZH Zürich [email protected] Neuroscience
7
Effect on gephryin clusters
Effect of the PSPC1 RRM and SFPQ RRM mutants on the gephyrin clustering was examined in-vitro in rats’ primary hippocampal neurons. Neurons were cotransfected with GFP labelled gephyrin and one of the DBHS proteins (PSPC1, PSPC1 RRM, SFPQ or SFPQ RRM) linked to mCherry fluorescence reporter (Figure 7). Control (Figure 7a) was cotransfected with GFP-gephyrin and PCR3.1, an empty plasmid vector. Both non-mutated DBHS proteins (PSPC1 and SFPQ) were localized in the nucleus, as seen by red signal on Figure 7b and 7d. Signal also overlaps with DAPI staining (figure not shown). PSPC1 RRM mutant as well as SFPQ RRM mutant were mainly localized in the soma but not in the nucleus (Figure 7c and 7e), red signal overlaps with green signal from GFP-gephyrin and results in yellow signal on the figures. Number of gephyrin clusters is reduced in neurons containing mutated proteins (PSPC1 RRM and SFPQ RRM) compared with negative control (neurons transfected with only with GFP-gephyrin; Figure 7a) or with non-mutated mCherry linked DBHS proteins (Figure 7b, 7d).
Figure 7: neurons were cotransfected with GFP labelled gephyrin (green signal) and DBHS protein tagged with red fluorescence protein mCherry (red signal). Negative control (a) was cotransfected with empty vector (PCR3.1). Gephyrin clusters seen as green dots localize primarily on the dendrites, with some larger clusters located in the soma (a, b, d). Number of gephyrin clusters in the neurons transfected with mCherry-PSPC1 (b) and mCherry-SFPQ (d) seem to be the same as in negative control on the other hand, number of clusters in neurons transfected with RRM mutants; mCherry-PSPC1 RRM (c) and mCherry-SFPQ RRM (e) seem to be lower. Also clusters seem to be larger. Non-mutated DBHS proteins were localized in the nucleus (b, d) while RRM mutatnts (c, e) were located in the soma but not in the nucleus where they overlapped with GFP signal and provided yellow signal on the pictures (c, e).

Matej Znidaric Role of NOPS proteins in the regulation of postsynaptic structure ETH, UZH Zürich [email protected] Neuroscience
8
Discussion
Expression of the cloned DBHS proteins (Figure 2) was successful in both HEK-293-T cell line (Figure 4) and primary hippocampus neurons (Figure 5). And as the size of the cloned proteins was confirmed on western blot (Figure 3), we can assume that DNA coding for proteins was correctly cloned, expressed and proteins were folded as predicted. Co-localization experiments performed in HEK cells (Figure 6) showed that the addition of mCherry domain or myc tag did not disturb localization of the protein, compared to endogenous DBHS proteins. In the cells is somehow hard to distinguish all 3 different channels and due to overexpression of inserted plasmids endogenous proteins might be under expressed. Due to highly overexpressed system, it also harder to spot parapseckle formations, which could be clearly seen where only cloned protein was imaged (Figure 4). However results indicate that both myc linked and mCherry linked DBHS proteins retain their localization, which is necessary information to proceed with further experiments. In the HEK cell lines the RRM mutants (mCherry-PSPC1 RRM; Figure 4b and mCherry-SFPQ RRM; Figure 4d) were localized in the nucleus as were non mutated proteins (Figure 4a, 4c). On the other hand, in the neurons RRM mutants were mainly located in the soma of the neuron but not in the nucleus (Figure 5b, 5d, 7c, 7e). It is hard to judge what caused the change in localization in neurons but isoform variants might be place to start, as it is known that many proteins had multiple slice variants that differ in different cell types.
Gephyrin forms clusters which could be seen as green dots on the image (Figure 7). It localizes mainly in the dendrites and in the soma as seen on the negative control (Figure 7a). Clusters in the soma usually appear larger than in the dendrites and it could be a result of the overexpression. Other explanation could be that it over-clusters and is therefore unusable in the synapse and targeted for the degradation in the soma. In co-transfected neurons with non-mutated form of the DBHS protein (mCherry-PSPC1; Figure 7b and mCherry-SFPQ; Figure 7d) changes in the gephyrin clustering are not very obvious. Especially in the case of the mCherry-PSPC1 the situation is more or less the same. In case of mCherry-SFPQ there is large build-up of gephyrin clusters in the soma, but clusters in the dendrites seem to be very similar compared to control. Also number of the clusters in the dendrites is comparable to control or PSPC1 overexpression. Build-up of gephyrin clusters in the soma could be again result of high overexpression rate or it could be an effect of the different process in the cell (neuronal activity, transportation, type of the neuron etc). Both PSPC1 and SFPQ are localized in the nucleus and no yellow signal could be seen (no overlapping with GFP-gephyrin). On the contrary, RRM mutants (mCherry-PSPC1 RRM; Figure 7c and mCherry-SFPQ RRM; Figure 7e), where both RNA recognition motifs have been mutated lost their nuclear localization and appear mainly in the soma, right next to the nucleus. Signal also heavily correlates with GFP-gephyrin and produces yellow signal as a result. Number of gephyrin clusters is in both cases reduced and there seem to be tendency towards bigger clusters (Figure 7c, 7e). It is hard to speculate what causes the phenotypic change in the gephyrin clustering in the neurons. Alternative splicing might play an important role in the phenotypic change, furthermore, SFPQ and paraspeckles were linked to alternative splicing. Another aspect could include function of DBHS proteins, mainly NONO and SFPQ in RNA retention in the nucleus. Since mutated DBHS proteins could not bind gephyrin RNA, it might be differently processed, or not processed as necessary. There are some evidences also in their applications in RNA trafficking, which could present another branch to study and link to phenotypic changes observed. It is also important not to forget that neurons represent very dynamic systems where synaptic activity

Matej Znidaric Role of NOPS proteins in the regulation of postsynaptic structure ETH, UZH Zürich [email protected] Neuroscience
9
plays a major role also in protein splicing and their coordination. Therefore it seems crucial to explore and study these different possible protein functions and link them to the phenotypic observations.

Matej Znidaric Role of NOPS proteins in the regulation of postsynaptic structure ETH, UZH Zürich [email protected] Neuroscience
10
Materials and methods
Cloning of the Constructs:
Two different sets of constructs were cloned. In first set, proteins (SFPQ, SFPQ RRM mutant, PSPC1 and PSPC1 RRM mutant) were in plasmid vector (ampicillin resistance) in front of the myc tag. Proteins without myc tag were amplified using designed primers with flanking restriction sites (Sac1 and BamH1 in case of PSPC1 and PSPC1 RRM, Sac1, Kpn1 in case of SFPQ, SFPQ RRM) and synthesised (Microsynth, SW). Gradient PCR was preformed (55°C-65°C 40 cycles, 2’ 72°C, added DMSO in case of SFPQ, Phusion polymerase provided by BioLabs was used), PCR products were purified using NucleoSpin II Gel Extraction Kit (Macherey-Magel) and cut (double digest with selected enzymes). Size was checked using standard gel electrophoresis and inserts of correct size were purified with NucleoSpin II Gel Extraction Kit (Macherey-Magel) and ligated into cut vector (kanamycin resistance). Vector contained fluorescent protein mCherry (707 bp) just in front of the restriction site. 10β strain of E.coli cells were transformed (using transfection solution; 100 mM CaCl2, 100 mM MgCl2 and 10’ heat shock) with ligation mixture and incubated on kanamycin plates (37°C, 16 h). Plasmids were purified from colonies using NucleoBond 100 kit (Macherey-Magel) according to manufacturer protocol and test restriction was performed.
Expression of plasmids
Expression of the plasmids was checked in HEK-293-T mammalian cell line. Cells were diluted 1:6 ration in 6-well plates 1 day before transfection and transfected with constructs using Polyethylenimine transfection protocol. Expression of fluorescent proteins was checked after 24h incubation (37°C, 5% CO2).
Primary rats’ hippocampal neurons were dissected from 18 day old embryos and cultivated for 11 days prior to transfection as previously described (Buerli, Pellegrino et al. 2007). Transfection was performed using magneto-transfection protocol.
Biochemical assay
Protein size was confirmed with standard western blot. HEK-293-T cells lysed in EBC buffer (50 mM Tris-HCl pH 8.0, 120 mM NaCl and 0.5% NP-40) containing complete mini-protease inhibitor (Roche Diagnostics) and phosphatase inhibitor cocktail 1 and 2 (Sigma–Aldrich). 2xSDS sample buffer were added and samples were boiled on 90°C for 3’. Samples run on 10% SDS gel and were transferred to the PVDF membranes using wet-transfer method and blocked (5% Roche blocking solution) and probed with primary antibodies (αmyc or αmCherry) over night at 4°C. Secondary antibodies coupled to the donkey horseradish peroxidase (HRP) were used to visualize the result. Membranes were than exposed up to 5’ and film was developed.
Confocal microscopy
Neurons were fixed and stained 7 days after transfection (neurons were fixed in 4% PFA solution 10’, and permeabilized in 0.1% Triton 6’, incubated for 1h in PBS and 10% NGS solution with primary

Matej Znidaric Role of NOPS proteins in the regulation of postsynaptic structure ETH, UZH Zürich [email protected] Neuroscience
11
antibodies, washed in PBS and probed with secondary antibodies and washed again). HEK-293-T cells were fixed and stained as neurons, 1 day post-transfection. Cover slips were mounted using DACO solution and left to dry out.
Primary antibodies used were polyclonal mouse anti PSPC1 (1:250 dilution), mouse anti SFPQ (1:250 dilution), mouse anti myc (1:1000 dilution), presynaptic markers vGlut1 (Synaptic Systems, 1:10000 dilution) and vGat (Synaptic Systems, 1:3000 dilution). Secondary antibodies were chosen according to primary and contained fluorescent signal CY3 (Jackson Immuno Research), CY5 (Jackson Immuno Research), or Alexa 488. Additionally the cells were stained with DAPI solution (c=1mg/ml, 1:5000 dilution).
Characteristics of fluorescent dyes used in experiments are shown in Table 1.
Table 1: excitation and emission wavelengths for fluorescent dyes Fluorescant dye Excitation (nm) Emission (nm) mCherry 587 610 eGFP 488 507 CY3 (512); 550 570; (615) CY5 (625); 650 670 Alexa488 499 519 DAPI 345 455
Fluorescent signals of different wavelengths were visualized by confocal microscopy (LSM 710; Zeiss AG, Jena, Germany) using a 63× or 40× Plan-Apochromat objective (N.A. 1.4). The pinhole was set to 1 Airy unit for each channel and separate colour channels were acquired sequentially. Images were taken by program ZEN 2009 (Carl Zeiss, Germany) version 5.5.0.305 and additionally cropped to the desired dimensions in Adobe Photoshop CS6. Minimal adjustments of contrast and brightness were applied to entire images.

Matej Znidaric Role of NOPS proteins in the regulation of postsynaptic structure ETH, UZH Zürich [email protected] Neuroscience
12
Bibliography
Bond, C. S. and A. H. Fox (2009). "Paraspeckles: nuclear bodies built on long noncoding RNA." J Cell Biol 186(5): 637-644. Brown, S., J. Ripperger, et al. (2005). "Period1-Associated Proteins Modulate the Negative Limb of the Mammalian Circadian Oscillator." Science 308. Bruelle, C., M. Bedard, et al. (2011). "The mitotic phosphorylation of p54(nrb) modulates its RNA binding activity." Biochem Cell Biol 89(4): 423-433. Buerli, T., C. Pellegrino, et al. (2007). "Efficient transfection of DNA or shRNA vectors into neurons using magnetofection." Nat. Protocols 2(12): 3090-3101. Fox, A. H., Y. W. Lam, et al. (2002). "Paraspeckles: A Novel Nuclear Domain." Curr Biol. Fox, A. H. and A. I. Lamond (2010). "Paraspeckles." Cold Spring Harb Perspect Biol 2(7): a000687. Fritschy, J. M., R. J. Harvey, et al. (2008). "Gephyrin: where do we stand, where do we go?" Trends Neurosci 31(5): 257-264. Fritschy, J. M., P. Panzanelli, et al. (2012). "Molecular and functional heterogeneity of GABAergic synapses." Cell Mol Life Sci. Froehner, S. C. (1993). "Regulation of ion channel distribution at synapses." Annual review of neuroscience 16: 347-368. Kanai, Y., N. Dohmae, et al. (2004). "Kinesin transports RNA: isolation and characterization of an RNA-transporting granule." Neuron 43(4): 513-525. Kirsch, J. (1996). "Synaptic Targeting of Ionotropic Neurotransmitter receptors." Molecular and Cellular Neuroscience 8(2-3): 93-98. Kuwahara, S., A. Ikei, et al. (2006). "PSPC1, NONO, and SFPQ are expressed in mouse Sertoli cells and may function as coregulators of androgen receptor-mediated transcription." Biol Reprod 75(3): 352-359. Passon, D. M., M. Lee, et al. (2012). "Structure of the heterodimer of human NONO and paraspeckle protein component 1 and analysis of its role in subnuclear body formation." Proc Natl Acad Sci U S A 109(13): 4846-4850. Prasanth, K. V., S. G. Prasanth, et al. (2005). "Regulating gene expression through RNA nuclear retention." Cell 123(2): 249-263. Prior, P. and B. Schmitt (1992). "Primary structure and alternative splice variants of gephyrin, a putative glycine receptor-tubulin linker protein." Neuron 8. Purves, D. (2012). Neuroscience, Fifth Edition. Shav-Tal, Y. and D. Zipori (2002). "PSF and p54nrb/NonO - multi-functional nuclear proteins." FEBS Letters. Stanley, C. F. (1993). "Anchoring glycine receptors." Nature. Sumanasekera, C., O. Kelemen, et al. (2012). "C6 pyridinium ceramide influences alternative pre-mRNA splicing by inhibiting protein phosphatase-1." Nucleic Acids Res 40(9): 4025-4039. Wang, H., F. K. Bedford, et al. (1999). "GABA(A)-receptor-associated protein links GABA(A) receptors and the cytoskeleton." Nature 397.