developmental modification and elimination of the larval stage in the ascidian, corella willmeriana
TRANSCRIPT

DEVELOPMENTAL MODIFICATION AND ELIMINA- TION O F THE LARVAL STAGE TN THE ASCIDIAN,
CORELLA WILLAIERIANA
C. M. CHILI) Hull Zoological Laboratory, the University of Chicago
SIXTY-FOUR FIGURES
AUTHOR'S ABSTRACT
The eggs of Corolla develop in tho atrial chamber of the parent a t a pH below that of nornial sea-water (pH 7.4 2). When removed to normal sea-water in early stages nxd under certain other experimental conditions, larval development is more or less inhibited, the tail being most inhibited, the dorsal region somewhat less. The free larval stage niay be eliminated and later development and metamorphosis may proceed normally to a n qdvanced stage in the chorion and give rise to normal ascidians. ?'he regions most inhiblted are, in general, those which possess the highest reducing power, a s indicated by KMn04. Experi- ments made in the attempt to control development all agree in indicating that the early stages a re adjusted to a certain GO, roncentraticn approximately that of the atrial cham. ber and presumably near that of the body. Solutions of the same p H may or may not inhibit development according lo their COn content.
The tail, the region of highest reducing power in the embryo during its development, is most inhibited; the dorsal region the next most rapidly reducing region, is next in degree of inhibition. 811 differences in reducing power disappear when, or soon after, the animals a r e killed by other agents before treatment with KMn04.
During the summers of 1918-1920, spent at the Puget Sound Biological Station, some time was devoted to study of the developmeiit of the solitary ascidian, Corella m-illmeriana, which is abundant on piles, on the lower surfaces of log floats, etc., at Friday Harbor. The animals breed during the summer and developmental material is readily obtained, but the course of development of eggs removed from the atrial chamber and developing in sea-water differs widely from that in nature. Development in the atrial cavity under natural conditions gives rise to a very active, long-tailed free-swim- ming larva which rapidly metamorphoses inlo a young ascidian. In eggs developing outside the atrial cavity in sea- water only a small proportion or none gives rise to swimming larvae. Under these conditions, development of the larval tail is more or less inhibited and in the more extreme cases other regions map also be affected. The inhibited forms
467
JOTRNAL OF XIOEP110L00Y AN1> PHYSIOLO(rY, V O L . 44 N O 3

468 C. M. C H I L D
sometimes hatch, but often the entire development up to a late stage of metamorphosis takes place within the chorion. Since tlie occurrence of both larval and more or less direct clevelopment in eggs of the same species and even in those of tlie same individual seemed in itself a matter of some interest and since the various degrees of inhibition of larval develop- ment appeared to represent differential inhibitions (Child, '24, pp. 80-84, 104-108), it was thought desirable to attempt to control the course of development by analyzing the condi- tions which determine normal development as a basis for further experimental work. This attempt was finally suc- cessful in the summer of 1920, but lack of time prevented further study of developmental modification, and publication has been delayed in the hope that opportunity for continuing the work might arise. It has not been possible, however, to return to Friday Harbor, and since the work did reach certain definite coiic1usions, it is being placed on record as a case of differential inhibition and also as an interesting case of a particular relation to environment.
In this connection I wish oiice more to express my apprecia- tion of the kiridiiess of the Director of the Puget Sound Biological Station, Ur. T. C. Frye, in placing the facilities of tlie station a t my disposal for this and other work.
M A T E R I A L AXD METIIODS
C'orella willmeriana is found below the level of all except extreme spring tides attached to piles and also on the lower surfaces of floating logs, always in dim light. The animals a rc attached with tlie longitudinal axis horizontal or with the distal elid obliquely or even vertically downward and with the atrial side of the body uppermost when the position is horizontal or oblique (fig. 1). Under natural conditions, a varying number of eggs, ranging from a very few to fifty or more, are extruded into the atrial chamber and fertilized about suiirise, apparently in reaction to light, for in the laboratory extrusion often occurs shortly after the light is

M O D I F I C A T I O N O F ASCIDIAN DEVELOPMEST 469
turned on in the evening, but has not been observed at other times. The follicle cells surrounding the eggs give them buoyancy, so that they accumulate in the highest part of the atrial chamber where they are kept in more or less continuous movement by the current when the siphons are open, but do not pass out until hatched as swimming larvae unless a violent contraction of the parent body occurs, in which case they may be ejected with the water. After hatching, they swim or are carried out and undergo metamorphosis. The animals are so transparent that eggs in the atrial chamber are readily seen with the naked eye, often before the animals are de- tached from the natural substratum. The eggs are collected
Fig. 1 The mature animal attached t o a vertical surface. The position of the embryos in the atrial chamber is indicated by the black dots.
either by cutting the atrial wall and allowing its contents to flow out or by placing the animals with siphons uppermost so that when the atrial siphon opens after the animal is left undisturbed f o r a time, the eggs pass out and rise to the surface.
The atrial chamber of an individual may contain two or even three different developmental stages which represent the fertilizations of one, two, or even three successive morn- ings, but in perhaps half the animals collected oiily one stage was found. By collecting the developmental stages from 8 to 9 A.M., it was usually possible to obtain from a number of animals sufficient material in stages preceding cleavage o r in early cleavage for several experimental lots of 100 to

4i0 C. M. CHILI)
200 eggs each. The method of recording results of experi- ments 011 developmental modification which was filially adopted consisted in the distinction of three form groups, normal larvae, forms with tails elongated but distorted, and forms with tails inhibited at an early stage and in determin- iiig the number of individuals in each group in a count of a sample, usually one hundred individuals taken at random. In some of thc larger lots more than one hundred were coiintecl, and in a few cases the lot consisted of less than one hundred eggs.
'IThen the eggs are ejected or removed from the parent body, the follicle cells keep them a t the surface, but after removal of the follicle cells, which is readily accomplished without injury to the eggs by gentle shaking or squirting from a dropper, they sink rapidly. This makes it possible readily to compare development at the surface with that at the bottom. Eggs removed about 8 A.M. from newly collected animals or from animals kept in the laboratory overnight range from undivided eggs to 2- to 4-cell stages. Gastrula- tion takes place in the laboratory a t a temperature of 18" to 30°C. shortly after noon, and by evening the tail is elongat- ing rapidly. In the laboratory hatching of the fully developed larvae occurs twenty-four to twenty-eight hours after fertili- zation. In nature development takes place a t a temperature of 11" t o 13°C. and is of course less rapid. 1\Ietamorphosis into the young ascidian is complete, except for the resorption of the stolons, in the laboratory in about four clays. Under ortliiiarv laboratory conditions, the death rate is zero, even though development may be considerably modified, as will a p p a r below. Jfetamorphosis proceeds normally, except as regards attachment, in finger-bowls, even without change of water .
111 the attempt to discover conditions outside the atrial chamher which would give normal larval development and to determine how development is altered under different cori- dit ions, various experimental modifications of environment have been used, starting from what could be determined con-

MODIFICATION O F ASCIDIAK DEVELOPMEKT 471
cerning conditions in the atrial chamber. Tests of water from the atrial chamber showed a pH ranging from 7.0 to 7.4 and occasionally to 7.6 in different individuals, with an aver- age of 7.3 to 7.4. With this fact in view., the eggs have been subjected to hydrogen-ion concentrations ranging from below 6.6 to somewhat above 8.6 and beginning with various stages from one to two cells to larvae about to hatch or hatched. I n these experiments eggs floating with follicle cells present and eggs at the bottom without follicle cells have been used and experimental temperatures have ranged from 11" to 23°C. The desired p1-l was usually obtained by adding HC1 or NaOH in various amounts to sea-water. I n the case of HC1 the effects of different concentrations brought to the same pH by shaking with air were also tested. H,SO,, CO,, and water from the atrial chambers of ascidians were also used in some experiments. Development of numerous lots of eggs has also been observed under the following conditions : at the surface with follicle cells in open bowls and in bowls covered with a glass plate with exclusion of a i r ; after removal of the follicle cells at the bottom of various depths of water from 2 to 15 em. with frequent agitation and without agitation; with con- tinuous agitation and aeration by bubbling air through the water and at the same time maintaining the pH nearly con- stant by frequent renewal of fluid of a certain pH; at various temperatures from 11" to 23°C.; in light of different in- tensities and in darkness. I n cases in which it was desired to keep the pH as constant as possible without disturbance, o r at least to keep it from inci-easing., the developmental stages were kept in full finger-bowls covered with a glass plate so as to exclude air, or in 500-cc. Erlenmeyer flasks almost full and corked, o r in some experiments in full corked tubes holding 40 cc. The concentrations of agents used are given in the text as indicated concentrations in sea-water, that is as the concentrations indicated by the volume or weight of the agent added to a given volume of sea-water. I n the case of I-IC1, the solutions a re made up from concentrated chemically pure HC1 as 10 m.

4'72 C. M. CHILD
Thus far little time has been spent in the direct investiga- tion of the gradients of developmental stages, and the only data presented here are those on differential reduction of KhfnC), (Child, '19, '24, p. 87). The modifications of develop- ment, however, constitute indirect evidence concerning the gradients, since the most susceptible regions are most in- hibited and the evidence from development agrees with that from differential reduction of liAInO,.
All figures are from drawings of individuals made at the time of observation, but follicle cells and test cells are not shown, and though eggs and embryos differ somewhat in size, the chorion is represented as a circle of the same diame- ter in all figures except when deformed by the embryo. Since the important points f o r present purposes concern primarily the external form and proportions, position of the sensory vesicle, and development of tail arid stoloiis, tlic: internal structures are omitted in most of the figures of the modified forms in stages preceding metamorphosis. The notochord cells of the tail are clearly visible in the living larvae, but are not drawn; the remains of the notochord and other caudal structures after resorption are indicated by dotted outlines in the posterior body region. In figures of later stages show- ing the tunic this is indicated by the thin membrane forming its outer surface, e.g., in figure 10.
NORMAL DEVELOPMENT AND METAMORPHOSIS
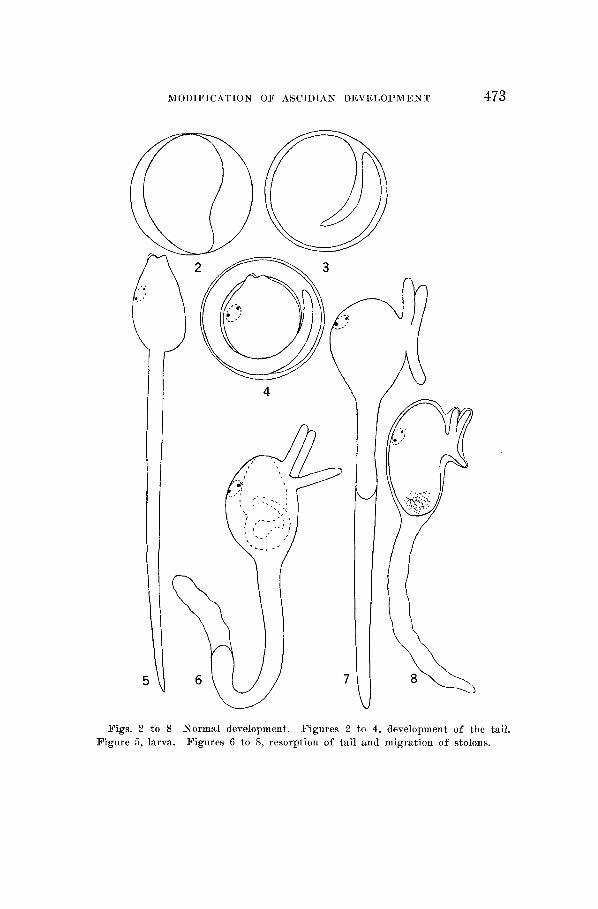
Soon after gastrulation, the tail region begins to elongate (fig. 2 ) ) and since the dorsal region apparently grows more rapidly than the ventral, becomes bent ventrally (fig. 3), and as it elongates is coiled around the body (fig. 4). Before hatching, the sensory vesicle with its two sensory pigment spots becomes clearly visible, the eye spot appearing first, and the three so-called adhesive papillae which are really stolon buds have arisen at the anterior end (fig. 4). Figure 5 shows the larva soon after hatching. It is a very active swimmer and the caudal movements are exceedingly rapid. Figures 6, 7, and 8 show7 stages in the resorption of the tail
M O D I F I C A T I O N O F ASCIDIAN D E V E L O P M E N T 473
5 7 \
Figs. 2 to 8 Figure 5, larva.
Normal development. Figures 6 to 8, resorption of tail and migration of stolons.
Figures 2 to 4, development of the tail.

474 C. M. C H I L D
which iisuallj- takes place withiii a few hours after hatching. The tunic of the tail region, which is usually little more than a dclicate membrane at this stage (figs. 6 to 8) map persist for a time after resorption of the tail, but is finally lost. Occasionally tlie separation of this membrane from the body surface by a jelly-like secretion beneath it takes place over h t l i larval body arid tail before resorption, hut usually it occurs during or after resorption of the tail (figs. 8 to 10; see also figs. 45, 46, 51, 52, 60, GI, 64).
A s resorption of the tail proceeds, the papillae of attacli- ment elongate into stolons, hut among the thousands of in- dividuals observed not one has been found attached, and in many cases the tunic is seen covering the stolons as wcll as tlie body (figs. 43, 46, 61, 64). Under laboratory condi- tions, these stolons do not function as organs of attachment. It seems probahle that they do fuiiction under natural condi- tioils, otherwise it is difficult to understand liow the animals become attached in such dcfinite manner, for after the tunic lias developed over tlie whole body a s in figure 10 they are certainly not able to attach. Apparently, thev do not react to the glass surface, and unfortunately they were mot tested with other surfaces. The outgrowth of the papillae into stolons may he a result of their failure to become attached. During metamorphosis they apparently migrate from the anterior end posteriorly along the ventral side. This change in position secms to be due to growth of the anterodorsal and dorsal, and reduction of the ventral and posteroventral regions of the body wall. The degenerating notochord cells are yellowish in the living animal and are indicated in figure 8 hy the dotted outlines in the posterior region. Figure 9 represents the young ascidian four days in tlie laboratory after fertilization. The stolons are now almost posterior and the incurrent opening lias developed. Development be- yond this stage is slow and does riot proceed far in the labora- tory, apparently because of lack of food. Figure 10 shows the most aclvaiiced stage attained in the laboratory. Degen- eration of the iiotochord cells has progressed until onlj- a

MO1)IPICATION O F ASCIDIAN DISVETAOPMENT 475
small mass remains on the posterior border of the stomach, and the stolons have now fused to a rounded mass at the posterior end, and this is undergoing resorption. The tunic has attained considerable thickness, but is still transparent.
DIFFERENTIAL REDUCTION O F KMnO, BY 1)EVELOPMENTAL STAGES
The differentials in reduction of KXnO, are so definite and uniform for particular stages and agree so well with the facts of normal and modified development that there seems to be no doubt of their significance as indicating differentials in physiological condition. These differentials are indicated
10
Pigs. 9 and 10 Normal development; the young ascidian.
in figures 11 to 23. In these figures the shaded portions indicate merely the regions which first become deeply stained by Mn0, or other oxides within one-half to two hours in KAlnO, m/5000 or m/4000. The point should be emphasized, however, that other parts of the body wall are not uncolored, but only less dceply colored. The permanganate apparently penetrates rapidly and all exposed parts become yellow within a few moments, but the regions indicated in the figures become deep brown or blackish much earlier than others.
In the two- and four-cell stages, the earliest stages tested with permanganate, the agent in concentration of m/5000

476 C. 31. CHILD
penetrates the chorion rapidly and stains the superficial cyto- plasm, but the deeper brown or blackish color appears first in the apical hemisphere. In this hemisphere a slight de- crease in depth of color from the apical region toward the equator is sometimes visible in optical section, but is not sufficiently distinct to be regarded as certainly significant. At the equator, however, a very marked decrease in reduc- tion takes place, so that the basal half of each cell is much less deeply colored than the apical half (fig. 11). At the third cleavage, division occurs in this region and separates approximately the two regions of different reduction rate into different cells, but in the two- and four-cell stages they are parts of the same cell in each half o r quadrant and there can be no doubt that the very marked difference in reduction iiidic2ates a difference in physiological condition in the two regions. This difference is in part, of course, a difference in the amount of yolk and in general cells containing less yolk show greater physiological activity. This apicobasal dif- ferential persists through the earlier cleavage stages es- sentially unchanged.
As gastrulation approaches, however, the region of most rapid reductioii in the ectoderm is anterodorsal (fig. 12) and at certain stages of development and of reduction appears to extend along the lateral margins of the blastopore (fig. 13). The reduction diff ereiitial a t the beginning of tail develop- ment is indicated in figure 14. At this stage the anterodorsal region of rapid reduction is still present, and a slight indica- tion of a second posterior region appears. As elongation of the tail proceeds, its tip becomes a region of very rapid reduction, showing deep color even before the aiiterodorsal region and a very distinct reduction gradient decreasing from tip to base of the tail appears at least in the ectoderm. Figure 15 shows an early stage of reduction in which the caudal gradient is already distinct, while the anterodorsal differential has not yet appeared, though it does appear after a few minutes’ further exposure (fig. 16). At this stage, and often earlier, a few isolated cells scattered over the body

MODIFICBTION O F ASCIDIAN DEVELOPMENT 477
surface, the tunic cells, become colored first of all as indicated in figure 15. In later stages they become more numerous (fig. 17) and are visible in the earlier stages of reduction, even
Figs. 11 to 18 11, four-eel1 stage. development, t h e tunic cells indicated in figures 15, 16, and 1 7 .
Differential reduction of KMnO, in prelarval stages. Figure Figures 15 to 18, stages of tail Figures 1 2 t o 14, gastrula.
in fully developed larvae, but are not visible after the body becomes deeply colored. Since they do not become more deeply colored than other cells, their early coloration is prob- ably due largely to the fact that they are more or less isolated on the surface of the body and are reached first by the agent.

478 C. ill. C H I L D
Figure 17 shows a somewhat later stage in side view, and figure 18 the same in dorsal view. In figure 18 the antero- dorsal region of rapid reduction is shown extending along the middorsal line. This mediolateral differential appears in tlie earlier stages of reduction, but is later obscured by the deep coloring of all parts. Figures 12 to 18 are drawn from individuals removed from the chorion, but repeated tests havc shown that the reduction gradients are the same within the chorion and that the permanganate penetrates the cliorion almost at once.
I n tlie full-grown larva (fig. 19) the caudal reduction gradient is still conspicuous, but reduction is relatively less rapid in the tail as compared with tlie anterodorsal region than in earlier stages. The stolon buds now appear as regions of more rapid reduction than adjoining parts, but otherwise the anterodorsal differential persists. Before resorption of the tail begins, the caudal gradient may become very slight or may disappear completely and the tail may show merely irregular areas of more rapid reduction (fig. 20). This cliange may occur within an hour or two. On the other hand, the tail undergoing resorption sometimes still shows traces of the original gradient and the body differential persists in all cases (fig. 21). Figures 22 and 23 show the regions of most rapid reduction in earlier and later metamorphosis. These two figures iridicate that the anterodorsal region of rapid reduction is extending around the anterior end of the body iis tlicl stolons change their position. Each stolon also shows an apicolnasal reduction gradient.
Observation of reduction of KlllnO, in large numbers of individuals shows a high degree of uniformity. Rarely an individual larva has been seen in which the anterodorsal dif - ferential was very slight or not visible, but these are so few that they are probably to be regarded as dying or in poor condition, rather than as exceptional individuals. Whether the differentials in reduction exist after reduction has pro- ceeded to completion, as they do in other forms investigated, was not determined, f o r even after dehydration and clearing

MODIFICATIOX O F ASCIDIAK DEVELOPMENT 479
the animals were still opaque. Sections should show whether the differential persists, but mere not made.
J
19 10
22 I I
Figs. 19 to 23 Differential reduction of KMnO, in larral and postlarval stages. Figure 19, swimming larva. Figure 20, disnppeciraiice of caudal gradient before resorption. Figure 21, a stage of resorption showing some caudal gradient. Figures 22 and 2 3 , stages of metamorphosis.
That the reduction differentials are characteristic features of the living but not of the dead organism is shown by the differences in reduction in individuals placed in permanganate alive and those which have been killed by means of other

480 C . M. C H I L D
agents before exposure to the permanganate. A few experi- ments of this sort were made with early and with fully devel- oped larvae and with early stages of metamorphosis. Of animals in 50 per cent alcohol for one minute and then re- turned to sea-water, washed, and brought into KiMn04, 50 per cent showed no differential redaction, but became colored rapidly and uniformly, though much less deeply than living animals. The other 50 per cent showed the normal differ- entials more or less distinctly, in most cases only as traces. After ten minutes in 50 per cent alcohol, 80 per cent showed uniform reduction, 20 per cent slight traces of the differ- entials. After one hour in 70 per cent alcohol, fifteen minutes and one-half hour in 95 per cent alcohol, followed by gradual return to water, 100 per cent showed uniform reduction with no trace of the differentials.
a f t e r one-half hour in saturated HgC1, and several wash- ings in sea-water, about 50 per cent showed uniform reduc- tion, the remainder, uniform reduction in the body wall, but slight traces of a differential in the notochord and pharynx. After two hours in HgCl,, 80 per cent showed uniform reduc- tion, the remainder slight traces of a differential in tlie pharynx. Individuals in HgSC1, for one-half hour, then washed and run up to 70 per cent alcohol, and finally back to sea- water after two to three hours in alcohol, showed uniform reduction in all cases. All animals that show uniform reduc- tion become less deeply colored than the living animals.
After twenty-four hours in KCN m/500,75 per cent showed uniform reduction, the remainder slight traces of the original differentials. Killing with boiling water obliterated all traces of the differentials. Evidently, the obliteration of the reduc- tion differentials takes place more rapidly with some agents than with others, but in all cases observed it occurs very soon, leaving no traces of them. These facts show that the reduc- tion differentials a re in some way associated with differentials characteristic of the living condition, and the fact that the regions of most rapid reduction are regions of greatest developmental activity indicates that here as elsewhere tlie

MODIFICSTION O F BSCIDIAN DEVELOPMENT 48 1
reduction differential is an indicator of a differential in such activity .
THE D E V E L O P M E N T A L MODIFICATIONS
Although the developmental modifications constitute a graded series representing various degrees of differential inhibition, they are separated for convenience of description into several groups.
The less id i ib i ted foi-ms
The most conspicuous and characteristic feature of the modifications observed is a greater or less degree of inhibi- tion in development of the larval tail. The lesser degrees of inhibition consist in failure of the tail to attain full length and apparently a lack of turgor in the cells, so that, instead of being regularly coiled before hatching, the tail is irregu- larly bent and kinked and in most cases its general course of growth is dorsally instead of ventrally around the body as in normal development. Figures 24 to 31 show examples of such inhibition at various stages before hatching and may be com- pared with figure 4, which represents normal development. I n some of these cases tail development may begin normally with ventral bending of the caudal outgrowth, but as elonga- tion continues the direction may change and the tail becomes variously and irregularly bent. Such forms result from a relatively slight degree of inhibition or from inhibition begin- ning late in development. With a somewhat greater degree of inhibition acting from early development, the tail grows dorsally from the beginning (fig. 26) and becomes variously bent and kinked on the dorsal surface (figs. 27 to 31). As these figures show, the development of the dorsal region of the body may also be inhibited, so that the tail arises more dor- sally and the sensory vesicle appears nearer the base of the tail than in normal development (compare figs. 26 to 31 with fig. 4). I n the more extreme cases, such as figure 31, the dorsal surface is considerably shortened in the anteroposte- rior direction and flattened instead of rouiided ; consequently,

482 C. M. CHILD
the stolon buds, as well as the tail, appear to arise farther dorsally than normally. In other words, the dorsal region is more inhibited than the ventral.
‘I’lic inhibited tails do not straighten entirely after hatch- i n g as the normal tails do, but retain their bent and distorted
30 31 Figs. 24 to 31 The less extreme dcgrees of inhibition in unliatolieil auim;tls.
Figures 24 and 2,5, sliglitlj- inhibitcd, tail elongated, bent ventrally a s in normal derelopment, Imt irot regularly coiled. Figurcs 26 to 31, soniewlrat inole inliibitecl, tail rlongntrtl, bmt dorsally, and distorted in various awys. The tail arises further dors:rlly ant1 the dist:iiice hetween its base and the sensory resicle is less tlian in nornr:rl :~ninials, and in figure 31 the wl10le dorsal region is Ant :mil shortened.

MODIFICATION O F ASCIDIAN D E V E L O P M E N T 483
forms to a greater or less degree and remain shorter and smaller than normal tails. Examples of such inhibited forms in the stages which approach normal larvae most closely are shown in figures 32 to 37. All possible gradations in the degree of inhibition and distortion of the tail appear and its motor activity shows a gradation more o r less parallel to the degree of morphological inhibition. The tail of the
37 36 Figs. 32 to 37 The less extreme tlegrers of inhihition in llatched animals.
Figure 33, tail reduced, Figures 34 to 37, tail bent dorsally, distance hetween tail
Figure 32, tail slightly bent, motor activity decreased. hut bent ventrally. and sensory vesicle decreased.
normal animal is extremely active, swimming movemcrits being so rapid as to be barely visible. As the degree of in- hibition of tail development increases, motor activity de- creases. In forms with slightly inhibited tails, such as figure 32, swimming movements powerful enough to propel the larva occur, but are weaker than the normal. With somewhat greater degree of inhibition (fig. 3 3 ) , the movements are still weaker and less in amplitude and accomplish little or no loco-
484 C . M. CHILI)
motion. 111 tlie forms with dorsally bent tails (figs. 3-2 t o 37) tlic motor activity consists of slight vibratory movements or is completely absent and tlic larvae lie motionless on the 1 ) o t t om.
40
41 44
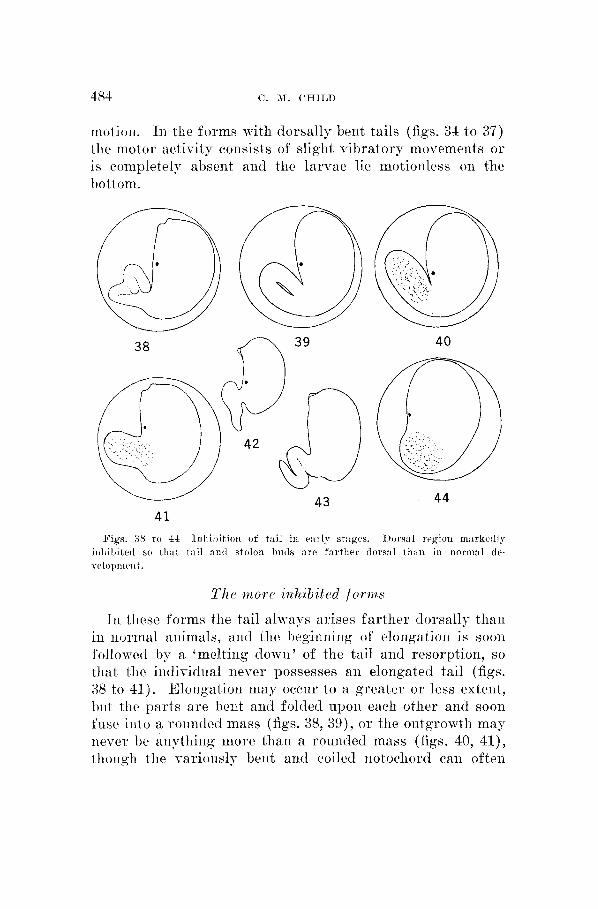
Figs. 38 t o 4.1- 1111~il~ition of tail in early st:iges. I>o~s:il region mnvkcdly i111iil)itetl so tli:rt t:til ant1 stolon l ~ u d s are farther i1ors:il tli:ni in normal tlc- velo~”ll‘~llt.
l’h P w i o re i?thib i t r cl f ( I r 111 s
In these forms tlie tail always arises farther dorsally than in iiormal animals, and the heginning of elorigation is sooii followed hy a ‘melting down’ of the tail and resorption, so that tlie iiitlividual never possesses an eloiigated tail (figs. 38 to 41). Eloilgation may occur to a greater or less extent, h i t the parts are beiit a i d folded upoii each other aiid soon fuse iiito a rouiitled mass (figs. 38, 39), o r the outgrowth may iiever bc aiiytliiiig more than a rounded mass (figs. 40, 41), though the varionslv bent arid coiled notochord can often

MODIFICATION O F A S C I D I A N 1~E\’EIAlPMEKT 485
he distinguished within this mass for a time. I n some cases, however, the iiotochord does not elongate, hut remains merely a mass of cells. Such forms usually remain within the chorioii until the development of tlie yoixng ascidiari is ad- vanced, but they may hatch, particularly in alkaline sea- water (figs. 42, 43).
I n these forms development of the whole dorsal region is inhibited in the anteroposterior axis to such an extent that the tail arises dorsally and the sensory vesicle is close to it, and even the stolon buds, in cases sufficiently advanced to show them, are more or less dorsal in position (figs. 38, 41 to 43). Caudal elongation may he inhibited from the earliest stages to such a degree that the tail never becomes anything more than a rounded mass in the posterior region of the embryo (fig. 44). When kept under tlie inhibiting conditions, such forms do not, as a rule, develop much further than thc stages figured and it is not certain that they ever complete metamorphosis.
Stololi modificatio?is i j a later stages The stolons which develop during metamorphosis show con-
siderable variation in normal animals as regards length at- tained, hut, with rare esceptioiis, their number and arrange- ment, two dorsal, one ventral, in their original position-later, two anterior, one posterior-are constant (figs. 5 to 9) . With the lesser degrees of differential inhibition the stolons are often longer or larger in diameter than under natural condi- tions (figs. 45, 46). TTith greater degrees of inhibition, the three stolons a re frequently replaced by a single large out- growth which involves the whole stolon-forming region. Figures 47 to 53 show cases of this modification. I n figures 47 to 51 the stolon outgrowth is still anterior, and various stages of tail resorption appear. In figure 52 the stoloiiic outgrowth is aiiteroventral and in figure 53 it is ventral.

486 C. M. C H I L D
Forws with bilaterally situated stwse orgaiis
I11 normal animals the sensory vesicle containing the two pigmented sense organs lies somewhat to the right of the
48
5 1
49 50
52 53
Figs. 4.5 to .?3 Stolon fornis whicli often occur in inliil,itetl i ~ ~ t l i r i t l n n l s .
MODIFICATION O F SSCII)IAP\' DEVELOPMEKT 487
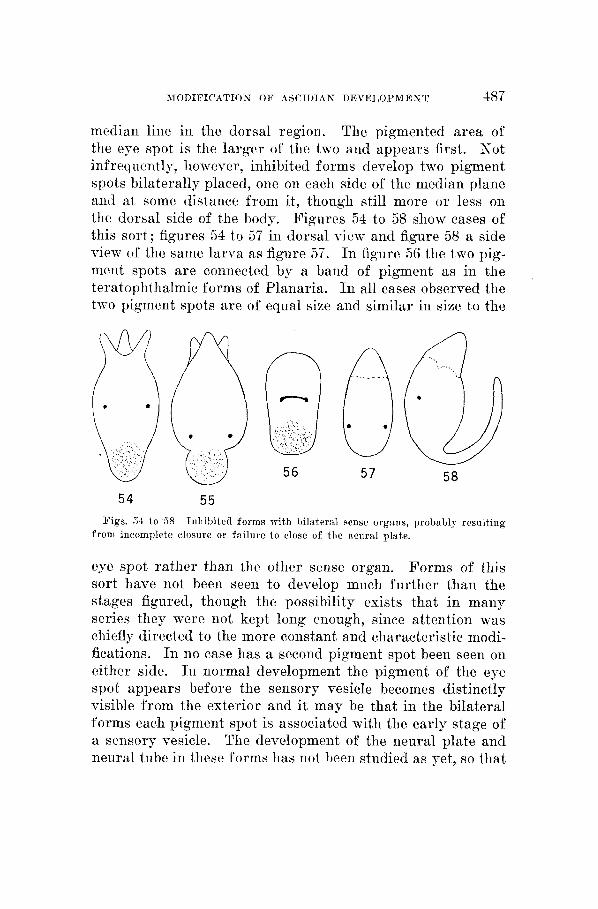
median line in the dorsal region. The pigmented area of the eye spot is the larger of the two and appears first. Not infrequently, however, inhibited forms develop two pigment spots bilaterally placed, one on each side of the median plane and at some distance from it, though still more or less on the dorsal side of the body. Figures 54 to 58 show cases of this sort; figures 34 t o 57 in dorsal view and figure 58 a side view of the same larva as figure 57. In figure 56 the two pig- ment spots are connected by a band of pigment as in the teratoplithalmic forms of Planaria. I n all cases observed the two pigment spots are of equal size and similar in size to the
56 57 58
54 55 Figs. 34 to 38 Inhibited forms with l~ilaternl sense organs, probably resulting
from incomplete closure or failure t o close of the neural plate.
eye spot rather than the other sense organ. Forms of this sort have not been seen to develop much further than the stages figured, though the possibility exists that in many series they were not kept long enough, since attention was chiefly directed to the more constant and characteristic modi- fications. In no case has a second pigment spot been seen on either side. In normal development the pigment of the eye spot appears before the seiisory vesicle becomes distinctly visible from the exterior and it may be that in the bilateral forms each pigment spot is associated with the early stage of a sensory vesicle. The development of the neural plate and neural tube in these forms has not been studied as yet, so that

488 C. M. CHILD
it is not possible a t present to do more tliaii call attention to their owiirrence. The bilateral position of the pigment spots, which are nntloubtedly early stages of sense organs a i d probably of eye spots, suggests, liowever, that closure of the neural plate has been delayed o r inhibited a i d that, iimler these conditions, two separate sensory primordia, bilaterally situated, develop. But wlietlier or not this sug- gestion is correct, it is a fact of some interest that under inhibiting conditions bilatei*al symmetry, at least as regards tlie p s i tion of these primordia, appears iiisteatl of tlie nor- mal asymmetry.
METAMORPIIOSIH OF THE: INHIRTTEI) FORhIS
Jletamorpliosis may take place in tlie inhibited forms a s iii tlie normal animals after hatching, or it may take place in tlie chorioii, and liatcliirig ma>- he tlclaycd until thc stage of the yoiuig ascidiaii. The course of metamorphosis is in all C N S ~ S essentially tlie same, thougli the dcgrcc of development of tlie tail preceding it may diflcr widcly. Figures 59 ant1 60 show cases in which tlerclopmeiit almost to the stage of Iiatc~liiiig took place uiider iiatural conditions, the emhryos I)ciiig a t the stage of figure 4 wlicw subjected to the inhihitiiig coiitlitions. ‘l’hese conditions prcveiited hatchiiig in most C;ISC’S a r id resorption of the tail began at or iw within thc c~lioi*ioii. The two fignres show stages attained after txeiity- two hoiirs ’ exposlire to tlie iiihibiting conditions, HCl m/1000, 1)II 7.1 t o 7.2. In figure 59 tlic tail is partly, in figure 60 w2iolly, resorhecl, arid in hot11 tlic caiidal tunic is indicated. Figure 61 shows a stage of metamorphosis after thirty-four hours in sea-water heginning with the gastrula stage. The l a ~ ~ a l tail was distorted aiicl with slight motor activity, similai. to figures 28 to 30, the larvae hatched in most cases, hut IICVCI’ swam and began metamorphosis at once. Figiirc 62 is a c ~ i s e of metamorphosis withiii the choriori in an animal in sea-water from early cleavage. CYauclal development was similar to tliat in figure 31. In the case of figure 63, exposure to stla-water lwgan with a11 advanccd cleavage stage, and

the tail developed to a somewhat greater extent tliaii in the case of figure 62 before resorption began, but was distorted aiicl folded on itself. Figure 64 sliows another case slightly iiiliibitctl from adraiicecl cleavage (HC1 m/2000, pH 7.3 to '7.6). 111 this case the stolons have developed to considerable length a i d tlie tunic surrounds both body and stolons iiisicle the chorion.
59 60 61
62 63 64
I n general, conditions wl~icli inhibit tail development to such an extent that tlie tail is distorted and more or less stunted and s h o w little or no motor activity have little effect on metamorphosis. With greater degrees of inhibition, metamorphosis may be retarded o r development may be stopped in some prelarval stage as soon as the inhibiting factor attains its full effect. Eveii under such conditions,

490 C. 11.1. C H I L D
however, metamorphosis may go on slowly for a time in many individuals, but may not be complete, apparently because the animals do not succeed in acclimating themselves to the con- ditions.
Dc~elopment of tlie tail, its increasing resistance to the coiled position, and probably also its motor activity appear to he factors in determining hatching, but increasing pressure resulting from growth in size of tail and body, or possibly f rom acciimiilation of fluid in the chorion, may also be con- cerned. In general, the inhibited forms are somewhat smaller than the normal animals, but hatching may occur even though tlie tail is riot fully developed and the larva does not fill tlie space within the chorion. Under these conditions, how- c~vc’r, hatching is much less frequent than in normal develop- meiit. If liatcliing does not occur at the larval stage, it usually does not occur until metamorphosis is advanced.
EXPEHlJIENTAL CONTliOL O F DEVELOPMENT
The data presented under this head are to be regarded as preliminary and as constituting merely a basis f o r further experimentation. So much time was spent in the attempt to discover and control experimentally conditions under which development would proceed as it does in the atrial chamber, that little was available for other forms of experiment. The first observations on eggs removed from the atrial chamber and developing in sea-water showed that differential inhibi- tion occiirred in sea-water, but it was believed a t first that this was due to some accidental condition, and only after repeated trials were the results accepted as convincing. De- velopment in sea-water from early stages gave only a small percentage of iiormal swimming larvae, o r often none a t all, and showed all degrees of differential inhibition figured above (figs. 24 to 58) except tlie extreme types observed in sea-water plus NaOH (figs. 41 to 43). As already noted, these differ- entially inhibited larvae proved to be highly resistant to laboratory conditions in later stages. Even when left in finger-bowls for several days without change of water, the

M O D I F I C A T I O N O F ASCIDIAN DEVELOPMENT 491
death rate was zero or almost zero, and all except the most inhibited forms gave rise to apparently normal young ascidians. Obviously, this inhibiting action of normal sea- water on larval development might he due to any one o r all of several factors, and in order to control development it is necessary to determine under what conditions the course of development that is characteristic in nature will occur. Most of the experimental work with Corella was concerned with this problem. The p H in the atrial cavity was deter- mined a t once and experiments with different pH were among the first performed, but both in connection with the pH ex- periments and independently of them the effects on develop- ment of control of other conditions were determined. By way of elimination, the experiments which led to results of little or no significance to the problem are briefly mentioned first.
Development in nature takes place a t a temperature of 11" to 13"C., but in the lahoratory the temperature was higher, the usual range being from 16" to 20"C., but occa- sionally on warm days going to 23". Developmental stages removed from the atrial chamber were allowed to develop a t different temperatures within these limits. Some lots were never exposed to laboratory temperatures a t any time, but developed in closed containers, both with and without air above the water, suspended from the logs on which the ascid- iaiis were found. Other lots were kept a t a temperature of 13" t o 15°C. by means of flowing water surrounding the con- tainers. Still others were exposed to the laboratory fluctua- tions. Repeated experiments along these lines showed, if anything, that development in sea-water a t the low tempera- tures a t which development took place in nature was slightly more inhibited than at laboratory temperatures up t o 20" to 21°C. I n some cases there was apparently a slightly in- creased inhibition a t 23", but so slight that i t could not be regarded as significant.
It was noted that the current of water passing through the atrial chamber kept the embryos in more or less con-

tiiiiims, though slight movement, arid it seemed possible that a tlPrelopmc~iit so rapid as tliis is, particularly at tlie higher laboratory temperatures, might require a coiitiiiiions change of water in order to supply adequate oxygen or to remove the W2. It was fourid that eggs remaining undistnrl~ed a t the bottom of open bowls in sea-water several centimeters deep after removal of tlie follicle cells showecl no greater, anid often less, inhihition than those floating uiidisturbed on tlic open surfacae. With frequent agitation and renewal of sea-water, Iiowerci., the eggs witliout follicle cells were as much inhibited as, or even more than, those floating imdis- t n r l d oii the open surface. L4gitation aiid frequent change of water did not decrease, lint seemed rather to inci-case the iiihibitiiig action on floating eggs. Floating eggs shomed much less inliihition when the howls were covered by glass plates without including any air than when they were left open to the air (table I). Continuous aeration and agitation by bubbling air through the water not only did not decrease the iiiliibiting action, but seemed sometimes to iiicrease it. In short, no evidence could be obtained that low oxygen or high CO, was iii any degree respoiisible for tlie inhibiting action of sea-water. Eggs developing in sea-water in different light intensities axid in darkness at natural and a t laboratory temperatures shoir.etl 110 appreciable differences.
Rq)eated colorimetric estimations of pH of the fluid from the atrial chamber shomctl that in freshly collected animals it xvas usnally 7.3 to 7.4, but might raiige above or below this. If the siphons had h e n long closed or if faeces were present, the pH might he a s low as 7.0 or occasionallp acid beyond the i*ange of plieiiol red. In animals which had been out of watcr for some time at lorn tide, the atrial fluid was often strongly acid hecause the siphons had been closed for some timc. The decreascl in pH of the water passing through the atrial chamber is ordinarily due to the CO, resulting from respiration and is readily removed by shaking with air, but the faeces coiitaiii some otlier acid.

M O D I F I C A T I O N O F ASCIDIAN D E V E L O P M E N T 493
I n the belief that the pH might be the important factor in determining the developmelit of swimming larvae in the atrial cliamher, data were obtained on the cffect on development of sea-water hrouglit to a certain pH by addition of acid, in most cases HC1, followed by shaking with air so fa r as that was necessary beyond the shaking required for mixture, to hring the solution to the required pH. &iny lots of eggs were treated with different pH in closed arid in open containers a t laboratory temperatures arid in the natural habitat of the animals. I n open containers the pH, of course, did not remain Coiistant, but in finger-bowls filled to the top and covered with glass plates so as to exclude air and in full closed flasks the change in pH within twenty-four hours cle- perided to some extent on the pH at the start, but was rarely more than 0.2. In a few experiments the p H was kept even more constant by testing arid bringing to the desired pEI every three to four hours.
The results of these experiments on the influence of pH appeared at first to be inconsistent aiitl puzzling. Occasion- ally, a n experimental lot would give a high percentage of normal larvae, mdiile another lot a t the same p H aiid ap- parently under the same conditions would show no normal forms at all. Finally, however, it became clear from repeated experiments with various concentrations of HC1 in sea-water ased at different pH or brought to a certain pH by shaking, that the p H was not the only or the most important factor. Sea-water-HC1 mixtures made with different concentrations of HCl and brought to the same pH might give very differ- ent results as regards developmelit. For example, a high percentage of normal larvae was obtained in sea-water hrouglit to a pH of 7.4 to 7.6 lo27 shaking an HC1-sea-water mixture of m/4000 or m/5000 indicated concentration until the required pH was attained, while an HC1-sea-water of m/1000 to m/2000 indicated concentration brought to the same p H by shaking inhibited development as much as or more than normal sea-water. According to these experiments, normal developmelit does not depend on pH alone, but never-

494 C. M. CHILD
theless apparently does depend on the amount of acid added to the sea-water, f o r with concentrations of HC1 m/3000 and abovc development is inhibited a t the same pH which with a lowi-er co-riceiitration of HC1 gives a high percentage of normal development. This being the case, two possibilities apparently exist : either the anion of the acid inhibits above a certain concentration or else change in the sea-water re- sulting from the addition of the acid is the effective factor. Since the amount of acid added is very small in all cases, inhibiting action of the anion appears to be excluded. The c1iic.f change in the sea-water brought about by the addition of acid is an increase in the CO, content. Up to a certain limit, the CO, set free increases with the amount of acid added ; consequently, the amount which must be removed by shaking to bring the mixture to a certain higher pH also increases. When this p H is below that of normal sea-water low concentrations of HC1 which require little or no shaking beyond that necessary for mixing to bring them to it will have a higher CO, content than normal sea-water and as mom HCl is added and more CO, must be removed to give the tlcsired pH the CO, content at this p H will decrease and above a certain HC1 concentration will be less than that of normal sea-water. For example, HCl m/4000 requires little shaking, HC1 m/2000 much shaking, to bring it to pH 7.4; that is, little of the CO, is removed from the lower, much from the higher concentration. Continued experiment along the lines suggested by these results and conclusions showed very clearly that the earlier developmental stages are ad- justed to a CO, content higher than that of normal sea-water arid approximately that of the atrial chamber under the usual conditions. It was not possible to determine the CO, tensions of the various mixtures used, but the various experiments apparently exclude other possibilities and afford a basis for the control of development. Some of the more significant data, chiefly from the summer of 1920, the last summer spent a t the Puget Xound Station, are given in tables 1 to 4. Tables 1 to 3 concern larval development and table 4 concerns

MODIFICATION O F ASCIDIAN DEVELOPMENT 495
metamorphosis. The column lieads in tables 1 and 2 are the same : the first seven columns give the characteristic features of each experiment and require no explanation. The last four columns give the percentages of the different forms and also the percentage of individuals which have hatched a t the end of the experiment. For purposes of tabulation, three groups of forms have been distinguished: first, ‘normal larvae,’ including forms like figures 4 and 5 with fully de- veloped tail regularly coiled before hatching and straight after hatching ; secondly, forms with ‘tails elongated but dis- torted,’ like figures 24 to 37; thirdly, forms with ‘tails in- hibited in early stages,’ like figures 38 to 44. The two groups of modified forms correspond essentially to the less inhibited and the more inhibited forms as distinguished above (pp. 381-483), except that the second group includes also the bilat- eral forms like figures 54 to 58. The grouping f o r the tables is based on the degree of inhibition of the tail, since that is the most conspicuous feature of the differential inhibition. There is, of course, no sharp line between the two groups of inhibited forms, bat, in general, individuals in which the tail outgrowth does not attain a length of more than one-half the body length are counted in the second group. Since the death rate is zero in the tabulated experiments, the three groups in the tables total 100 per cent. The last column, ‘hatched,’ affords in some experiments a further indication of the effects of the various conditions.
Table 1 gives a few data on development in sea-water under various conditions as regards exposure to air. In series IA the early stages were put into a large flat container in shallow water and were kept in almost continuous agitation during the first sixteen hours. The p1-I of the sea-water at the be- ginning of this experiment was slightly higher than usual, but never went below 8.0. Here, in spite of good aeration, two counts gave only 3 to 4 per cent normal and only one individual hatched. In series IB the eggs retained their follicle cells and floated undisturbed on the surface. Under these conditions, surface tension brings most of them to-

getlier and the? lie in contact in il single layer on the surface. T1m.e (winits gave 27, 29.6, arid 27 per cent normal larvae, hiit few were hateheti. In series I C arid I1 the eggs retained follicle cells aiicl floated oit the surface of a full finger-bowl iii contact with a glass plate wliicli covered the bowl so that air w a s excluded. Under these conditions, as on the free sitri'ace, they mass together in one or more groups aiicl form a laper. It will be rioted that in these two lots the percentage
2 g ? icz a
8.1
8.0
8.0
8.0
TABLE 1
I1c.i c Top?i ie i i t 1 1 1 sea-water tinder di f ferent coi idzt ions
r , c Z k
Y m" q;
s g 5 ;s 2
8.0 100 100
8.0 100 5 4
100
7.S 26 100
i.i 30
A t Imttoiii in 10 to 13 nim. watcr, frequeiit agitstic
Floating uiitlist~irlierl 011
opeir snrface
Floating u~rdistiirlrt.tl nu closctl surface
Floatiiig uiitlistur~ietl 011
closctl surface
of normal larvae is miicli higher, 64 aiid 70 in TC and 83.3 in 11, wliich consists, however, of a small number of' eggs. Similar experiments have been repeatedly performed and, in general, eggs which are massed together arid undistui.bet1 give a higher percentage of normals than those mhicli arc separated and agitated in open vessels and the percentage of normals is still liiglier w-hen the surface on which they float is closed. All these experiments suggest that for normal larval clevelopment more C'O, must be present tliaii in iiormal sea-water. Wlieit llie eggs clump together in masses, partic-

iLIODIFICATION O F ASCIDIAN DEVELOPiMENT 4%
C
2 5 z I
6.6- 6.9 7.35
a + 8.(i+
r -- i . i J
7 . 3 3 T.8 8.6
7 . 3 3
8.5 i . 3
~~
- 0- ,.-d
i.G
ularly 011 a closed surface arid are undisturbed, their own CO, accumulates about them to some extent a id the pH in their immediate vicinity is often 0.2 to 0.3 lower than else- where in the bowl. I n table 1 I C and I1 show a decrease in pH of 0.2 in one and 0.3 in the other, but the pH about the aggregations of embryos must have been still lower.
TABLE 2
Uevelopnzent zt i various coitceatrntions of I lC? and S a O E aird 2 7 1 sen-water
g 5 5 ; S ? 4 -
100 100 100 100 100 100
100 100 100
~
100
100 100
~
100
100
I A I R IC: I D I13 IF
IIA I IB I I C
1 to 16 cells 23 IICl i11/1150, closed 1 to 16 crlls 23 lICl111/2000, closed 1 to 16 cclls 23 HCI m/5000, closcc! 1 to 16 cells 23 Sea-water, c l o w l 1 to 16 cells 23 NaOH m,/3000, closed 1 to 16 cc~lls 23 'XaOH m/2000, czloseil
I
16 cells 1 2 1 IICI m/40m, closec1 16 cr,!ls 2 1 Sea-water, c~loscrl 16 cells 2 1 NaOH 111/4000, closeii
J 4
z e z < ?
c
z2 I
~
6.6-- i . 0
8.0 8.6f 8.6+
r - i . 3
I11 11-18 1 V B 8
VA
T'B
7.4 8.0 8.6+
2 t o 8 cells 24 HC1 ni/.i000, closed to 36 cells' 22 1HCIm/4000, closcd to 16 cells 22 NaOH ii1/4000, close11
2 t o 8 cclls 25 40 cc. sea-water fmni ascidiaii stock, close(
2 to 8 cells 25 40 cc. sea-watei, vlosec
i.6 7 . 3 8.5
7 .4
8.0
Table 2 gives comparative data on development in various concentrations of IIC1 and NaOH aiitl in sea-water in closed bowls or tubes without air. I n series I the percentage of iiormal larvae and the percentage of hatched individuals in- creases with decreasing coriceritration of HC1 to m/5000 in which 9 1 per cent normal larvae develop arid 26 per cent are hatched a t the elid of the experiment. All the normal

498 C. M. C H I L D
larvae a i d most of the others hatch a little later. I n sea- mxtcr and in the two concentrations of NaOH there are no normal larvae and the degree of inhibition increases with iiicreasiiig alkalinity until in NaOH m/2000 as in HC1 m/1150 all tails are inhibited in early stages.
In series IT 9’7 per cent normal larvae develop in HC1 m/4000, 8 per cent in sea-water, and none in NaOH m/4000. In series 111 100 per cent are normal in HC1 m/5000 and in series IV 90 per cent a re normal j i i HC1 m/4000. I n series V the water in which a stock of ascidiaiis had been standing overnight a t a p H of 7.4 is compared with normal sea-water, 40-cc. tubes closed without iiiclusion of air being used in both cases. Here the water from the ascidian stock gives 94 per cent of normal larvae with 50 per cent hatched, while in normal sea-water 100 per cent have distorted tails and none have hatched.
It must be pointed out that different lots of eggs collected on different days often differ widely as regards the per- centages of normal and inhibited forms in sea-water or under o t h c ~ c-wnditions. Fo r example, taking the different closed sea-water lots in tables 1 and 2, I C and I1 of table 1 give 64 to 83 per cent of normal larvae, I D of table 2 gives none and also gives 48 per cent with tails inhibited in early stages, I IR gives 8 per cent normal and 92 per cent of distorted tails, and VB, 100 per cent of distorted tails. Difference in snsczeptibility of different lots of eggs to sea-water uii- douhtedly accounts for at least a considerable par t of these clifferences. Eggs from different iridividuals map be adjusted t o slightly different CO, concentrations, but incidental factors not fully coiitrolled cannot be exclnded. For example, the degree to ~vliich the floating eggs clump together may deter- mine marked differences in the CO, in their immediate vi- cinity. Unfortunately, the importance of this point was not recognized until too late f o r most of the experiments. More- over, in some lots a par t of the eggs may lose their follicle cells and sink to the bottom, where they lie scattered. Such eggs were usnally included in counts with eggs from the

M O D I F l C A T l O N O F ASCIDIAN DEVELOPMENT 499
surface, but in table 1, IB, IC, and T I only floating eggs were counted. This probably accounts for the higher normal frequency in these lots than in the sea-water lots of table 2. Differences of temperature may also account for some of the differences in normal frequency. Some series developed at 16" to 18"C., but in some cases the temperature during the day was 23"('. In the different series of table 2, however, the changes are always in the same direction from acid to sea-water and to NaOH. I n all cases (TC, IIA, 111, IVA) HC1 m/5000 or m/4000 indicated concentration at pH 7.3 to 7.6 at the beginning of the experiment, gives 90 to 100 per cent of normal larvae. The highest percentage of normals under any other conditions in table 2 is 22 per cent in IB, HCl m,/2000, pH 6.9. In series IVB 12 per cent of normal larvae appear in NaOH m/4000 at pH 8.5. This case is un- accounted for , but may have resulted from clumping together of the eggs with a cbnsiderable decrease of pH in the middle region of the aggregation. The average normal percentage for all lots in HC1 m/5000 and m,/4000 and ascidian stock water in table 2 is 94.4 and that for all lots under other con- ditions is 4.7-a difference of 89.7 per cent. This difference is so great and the different series are so consistent that there can be no doubt of its significance.
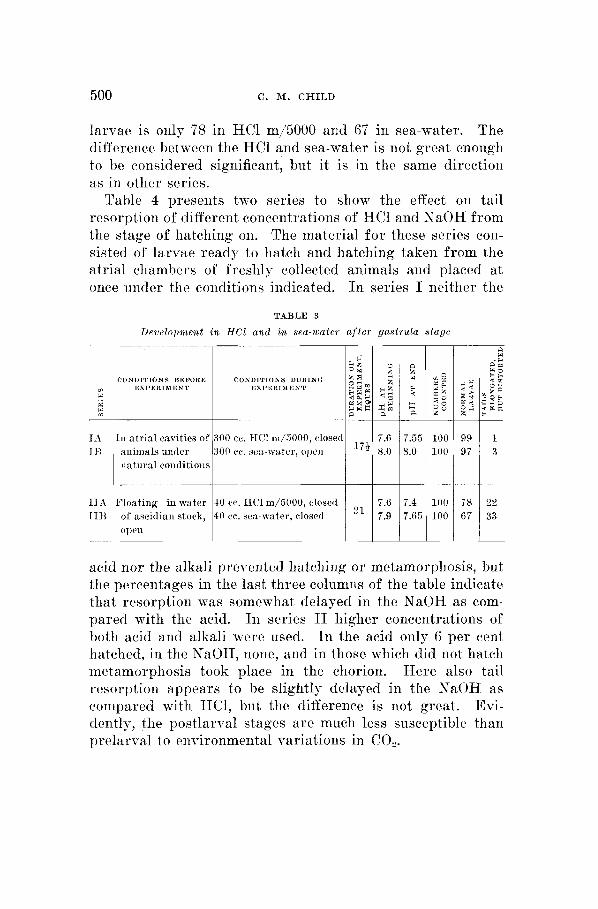
Table 3 shows that normal sea-watcr and HC3 m/5000 have little effect in altering the course of development when the period of exposure begins only at the gastrula stage. 111
series I development up to the gastrula took place in the atrial chambers under natural conditions ; that is, the ani- mals were collected in the early afternoon. In this series both the IICl lot, IA at pH 7.6 to 7.55 and the open sea-water lot at pH 8.0 give almost 100 per cent of normal larvae. The eggs of series I T were extruded in the early morning into the water of the stock aquarium at pH 7.2 to 7.0 from the CO, produced by the animals overnight and floated on the siirface of this water up to the gastrula stage. The two lots of series I1 were apparently slightly inhibited by the environment of the early stages, for the percentage of normal
500 C. M. C H I L D
larvae is only 78 in HC1 m/5000 and 67 in sea-water. The difference between the HC1 and sea-water is not great enough to lie considered significant, but it is in the same direction as in other series.
Table 4 presents two series to show the effect on tail resorption of different concentrations of HCl and NaOH from the stage of hatching on. The material for these series con- sisted of larvae ready to hatch and hatching taken from the atrial chambers of freshly collected animals and placed a t once under the conditions indicated. I n series I neither the
TABLE 3
Drvelopmeiit in HC1 and in sea-water a f t e r gastrula stage
CONDITIONS BEFORE I CONDITIONS DURTNCI EXPERIMENT EHI’EltIMIIENT EXPERIMENT hXI’h1tIMIILNT
In atrial cavities of 300 cc. IICl m/5000, closed 7.6 174 8.0 animals under
natural coiiditioiis 300 c e . seii-wat~r, open
TIA Floating in water 40 cc. EICl 1n/5000, closcd 1113 of nscitlian stock, 40 re. seil-water, closed
o p e 1 1
99 1 97 3
__
T U 22 67 33
acid nor the alkali prevented hatching or metamorphosis, but the percentages in the last three columiis of the table indicate that resorption was somewhat delayed in the NaOH as com- pared with thc acid. I n series I1 higher concentrations of both acid and alkali were used. I n the acid oiily 6 per cent hatched, in the NaOH, none, and in those which did not hatch metamorphosis took place in the chorion. Here also tail resorption appears to be slightly delayed in the NaOH as compared with HC1, but the difference is not great. Evi- dently, the postlarval stages a re much less susceptible than prelarval to environmental variations in CO,.

M O D I F I C A T I O N O F ASCIDIAN DEVELOPMENT 501
HCl m/1600, closed
XaOH ni/3U00,
Under certain conditions, larval development is inhibited in nature. 111 animals exposed to sunlight while out of water at extreme low tide the same sorts of inhibited forms have been found as under experimental conditions. As already noted, the atrial fluid may become strongly acid in such ani- mals, because the siphons remain closed. This and perhaps the high temperature are undoubtedly the factors which de- termine the inhibition in such cases.
6.8
24 8.6+
TABLE 4
Development in I I C l and A-aOH f rom stage j u s t preceding hatching
CONDITIONS BEFORE
EXPEEIMENT
In atrial cavities of animals under iiatural conditions
In atrial cavities of animals under iiatural coiiditioiie
HCl m/3500, closed
NaOH m/4000,
l 2 8.4
ope11 I I
14 short, stump only
29 less 71 than half
94 than half resorbed
thaii half resorbed
10 less 83
Experiments toward the end of the summer in which the desired pH was obtained by bubbling CO, through the water gave no results, because both experimental and control lots failed to develop beyond early stages. These experiments were made only after it became evident that CO, was the important factor. In obtaining material f o r them the locali- ties from which the animals had been previously collected were exhausted and no new localities were found. Moreover, the failure of the last lots of eggs to develop suggested that the end of the breeding season was at hand. As regards

502 C. M. CHILI)
coiitrol of tlevelopmeiit by CO,, tlicn, the final proof' is lacak- iiig, but tlie evideiice seems to he conclusive without it.
1)IRC"SSIOS
T I I ~ J i izhibitrd lama1 f o m i s a s cases of d i f r r e u t i a l idi i l j i t ion It is a question of some interest \vhether the inhibition of
larval development in Corella falls within tlie general cate- gory of differential inhibition ; that is, whether the inhibition results from the fact that the more active regions iire more susceptible to the inhibiting conditions than less active re- gions, or whether there is some specific effect. Thus f a r no large number of agents has h e n used iii experimental con- trol, but the same sorts of inhibited forms have beeii obserred iii the higher concentrations of acids, in normal sea-water and in NaOII-sea-water, the only differences being in degree of iiiliibition arid frequency of the different forms, according to concentration of agent, stage of development, etc. Tlius far, tlicn, there is no evidence of specificity of effect of c~xteriial factors. Moreover, the data on inhibition are in a gelicral way in accord with those on reduction of I<Mn04; that is, the regions which are most inhibited are those ~vliich the pel-maiigaiiate method indicates as the most active and Ihercfore as in general most susceptible to the more extreme actions of external agents. The tail is indicated by reduc- tion of I<JLnO, as the most active region of the embryo dur- iiig its outgrowth, and therefore most susceptible, and the iiiliihitcd forms show that the development of tlie tail is first of all and most strongly inhibited. The tail may be markedly iiiliibitecl, while other parts of the body are iiot appreciably affected (figs. 24, 25, 33), but commoiily tlie iiiliibited tail is more dorsal in position than normal and is bent o r coiled (lorsally instead of ventrally and the distance between the sensory vesicle and the base of the tail is less than normal (figs. 26 to 30, 34, 35). In the more extreme modifications the tlorsal region is flat instead of rounded in tlie anteroposterior clirthction and the papillae of attachment or stolen buds ap- 1)e;tr farther (lorsally than in normal animals; that is, the

XODIFICATION O F ASCIDIAN DEVELOPMENT 503
whole dorsal region is shortened as compared with the ventral (figs. 31, 38, 41 to 43). The more dorsal origin, the dorsal instead of ventral curvature of the tail, the altered position of the sensory vesicle, and the shortening of the whole dorsal region are obviously the result of a relatively greater degree of inhibition of development on the dorsal than on the ventral side. The different degrees of inhibition may all appear in the same lot of eggs under the same conditions, but in the higher concentrations of acid and in NaOH the frequency of the more inhibited forms is higher than in the lower acid concentrations and in normal sea-water.
Figures 26 to 30, 34, 35, show or suggest-most of them being too early to show the papillae-that the distance be- tween the base of the tail and the sensory vesicle may de- crease markedly, while that between sensory vesicle and the papillae remains unaltered and decreases only with more extreme inhibition (figs. 31, 36, 38, 41, 43). On the other hand, the region of high reducing power, as indicated by KMnO,, extends over the whole length of the dorsal region and is apparently most marked anterior to the sensory vesicle in the later stages. These two facts appear to be in conflict. The apparent discrepancy seems, however, to be due merely to the fact that the anterodorsal region is well advanced in very early stages, which are apparently less susceptible to the inhibiting conditions. Development to the gastrula stage is but little, if at all, retarded in the stronger acid mixtures or in sea-water, but by the time the posterodorsal elongation begins the susceptibility is in general higher and the postero- dorsal region is therefore more inhibited than those parts of the dorsal region which develop earlier. The NaOH mix- tures inhibit more strongly than others used and in these the effect on the anterodorsal region is clearly distinguishable (figs. 41 to 43). Undoubtedly, the fact that the tail develops relatively late when the whole embryo has a higher metab- olism and is more susceptible is a factor in its very high susceptibility to inhibiting conditions; that is, in order to grow as rapidly as it does at this relatively active stage, it

504 C. &I. CHILD
must attain a very high level of activity. The reduction method, its extremely rapid development, arid its very great muscular activity when not too much inhibited, all indicate that it is the most active and most susceptible region of the embryo.
The dorsal bending of the tail in most of the inhibited forms (figs. 26 to 31, 34 to 43) indicates a difference in susceptibility between dorsal and ventral regions of the tail, but 110 corresponding difTerence in rate of reduction of KMnO, on dorsal and ventral sides of the tail itself has been ob- served. Like the other lethal methods, tlie reduction method is of value chiefly in indicating the more marked differences, and as death takes place the slight differences are largely or wholly obliterated. lloreover, the differences in rate of re- duction ~vhich appear late are more or less obscured hy the general coloration of the body. The shortening of the whole dorsal region and the bending of tlie tail dorsally under the inhibiting conditions both indicate a higher susceptibility and a greater degree of inhibition. The reduction method indi- cates most of this region as a region of high reducing power, hut does not show any definite differences between dorsal and ventral regions of the tail. Modification of development tlirougli diff ereiitial susceptibility unquestionably brings to light quantitative differences in physiological condition which a re not shown by the lethal methods. In the present case the evidence from reduction is less complete than that from modification of development, but as f a r as it goes is in agree- ment with it. Noreover, the evidence from modification of clevclopment in Corella is in line with that in other chordates.
As regards their general localization in the body, tlie most active and most inliihited regions of the embryo, the dorsal region and the tail, correspond to the regions of highest susceptibility to lethal agents in other chordates on which observations have been made, viz., the lamprey (Hyman, '26), teleost embryos (Hyman, '21), the frog (Bellamy, '19, '22), the chick (IIyman, '27; Hinrichs, '27). These data are con- firmed by those on modification of development in the

MODIPICATION O F ASCIDIAN DEVELOPMENT 505
chordates obtained by various investigators with various agents. I n general, no evidence of specificity in these modi- fications has been found, the same modifications appearing with different conditions. Much of this literature has been discussed by Hyman in the papers referred to above. All the evidence at hand then indicates that the inhibited larval forms of Corella are typical cases of differeiitial inhibition; that is, inhibition corresponding to the physiological activity and therefore to the susceptibility of the different parts.
Aiateroposterior d e v e l o p m e n t in the c h o r d a t e s and the g r a d i e n t s
In Corella, as in other chordates, two embryonic regions are indicated by the methods employed as regions of high metabolic activity and high susceptibility, but attention must be called to certain differences. In the other chordates ex- amined the second ‘high’ region represents primarily the dorsal lip of the blastopore and the tail bud is a later develop- ment from it. I n Corella, however, according to the data at present available, the region corresponding to the dorsal blastopore lip appears to be a part of the anterior ‘high’ region. It may be, of course, that further work with im- proved technique will show a primary anterior and a second- ary blastoporal active region here, but in the light of the observations as they stand, a different interpretation is sug- gested. This is that the relation of the posterior growing region to the blastopore which we find in fishes and amphibia is not a necessary nor a primary relation, but rather a result of the spreading out of the embryo over the yolk. I n a holoblastic form like Corella with gastrular invagination in- stead of prolonged overgrowth, the appearance of the blasto- pore adjoining the primary active region does not, I think, involve us in any difficulties. If physiological isolation is a factor in the appearance of the secondary high region, the position at the posterior end of the embryo follows as a matter of course. I n connection with these data it seems pertinent to call attention briefly to certain points of Kings-

506 C. M. CII ILD
bur>T's recent discussions of tlie gradient conception as ap- plied t o vertebrate development (Kingsbury, '24, '25). The anterotlorsal active region of Corella appears to be continu- ous with the active apical hemisphere of early stages, or with some part of it, but the nervous system apparently does not arise from its most anterior part. According to Conklin ('05), the neural plate in Cynthia consists of cells which represent regions both above and below the plane of tlie third cleavage on the dorsal side, and the anterior end of the neural plate extends only about one-third of the distance from the equator to tlie animal pole. If the course of development in ('orella is similar, as in all prohability it is, the anterior end of the nervous system certainly does not represent the animal pole. Iii general, the region of the apical pole ap- pears to be the most active region in early developmental stages, though in the case of Corella no marked differences hare been observed within the apical hemisphere. I t is not iieccwary to assume, however, that the most active region remains exactly at the animal pole as development progresses. It may shift its position to other cells or cell groups, but it seems to he true that the anterior region of the nervous system arises from that region of the embryo which is most active at the time of origin of the nervous system and that this region is usually a part of, or continuous with, the primary apical active region. As regards the vertebrates, opinions still differ to some extent concerning the relation of the anterior end of the nervous system to the animal pole, and it is entirely possible that the distance of this region from the animal pole may differ widely in different forms. Moreover, it is conceivable that the active region from which the anterior nervous system develops may originate, not as a part of the primary apical region, but distinct from it. The point of chief importance for the gradient theory is that the most active region at a certain stage or a part of it develops as the anterior end. The question whether this region represents the animal pole is merely a question of fact in each particular case. In many invertebrates it cer-

M O D I F I C A T I O N O F ASCIDIAN DEVELOPMENT 507
tainly does, but in various others it does not. In the sea- urchin, f o r example, the anterior end of the larva represents one side of the egg and in certain hydroids the definitive apical end develops from the basal pole of the planula and this in turn from the vegetal region of the egg. I n Corella it appears that the active region of early stages extends o r shifts along the meridian which becomes dorsal.
As regards the two gradients of the annelid and the verte- brate, with which Kingsbury seems to find difficulty, it may be pointed out, first, that many lines of evidence demonstrate or indicate their existence and, secondly, that, as already noted (Child, '25), they differ in character, the anterior being a gradient in which differentiation begins at, and progresses from, the highest level; the posterior, a gradient in which growth is most rapid at, and decreases from, the highest level; but we do not know the physiological basis of these differences. It is important, however, to keep in mind the fact that differentiation always proceeds in the direction from anterior to posterior. The second gradient results merely from differences in growth activity and is temporary. It may for the time being dominate a large part of the body length, but as each segment in turn becomes older and less active it falls under the nervous control of the more anterior regions. This nervous dominance is fa r more effective than the primitive sort of dominance exercised by the embryonic re- gion of the posterior end. The temporary reflex apparatus of the amphibia, as described by Coghill, and its replacement by the definitive apparatus (Child, '21, pp. 252 to 255) ap- pear to be expressions of the relations of the two gradients. No one, I think, has discovered any other physiological basis for the character of the first apparatus and its later dis- appearance, but that the facts are anything more than sug- gestive o r indicative has not been maintained.
A gradient is merely the result of localization of an active region and dominance is the influence of the active region on other parts about it. Differences in degree of activity are the earliest distinguishable characteristics of physiological

508 C . M. CHILD
axes ; consecpeiitly, these axes represent gradients. The possihilily to which Kingsbury calls attention following Har- rison and Coiililin, that the gradients are only expressions of some more fundamental polarity and symmetry, need not bc denied, but the important point is that there is no evidence f o r such a polarity. All the evidence indicates that the gradients a re the primary factors. TVhe11 we alter them, polarity and symmetry a re altered; when we obliterate them, we can find no traces of the original polarity and symmetry, aiid when we determine new gradients in cells or cell masses capable of developing, new polarities and symmetries ap- pear. Also there is much evidence to indicate that dominance is primarily dynamic in character and none to indicate that it is not. The most active region of a gradient differs from others as an excited region differs from less excited or un- excited regions, and the external factors which determine gradirnts a re in general similar to those which bring about ciscitation, except that they act less intensely aiid for longer times. I t is not assumed that dominaiice consists in the transmission of ~vaves of actual excitation, but rather that the dominant region exercises a continuous effect similar, perhaps, to the iiervoiis maintenance of muscle tonus. This effect may be primarily electric in nature, since differences of potential apparently always exist between different levels of a gradient.
According to Kingsbury, the law of anteroposterior de- velopmeiit reveals only a time sequence. This, of course, is quite true from the descriptive standpoint, and the mor- phologist may be content with this revelation. To the physiologist, however, the so-called law represents the gross rcsult of a complex of processes about which he would like to learn something. The ‘law’ concerns him only as a starting- point for investigation. If there is a time sequence, it must have some basis. The gradient conception of axiate develop- ment, the outgrowth of many different lines of evidence, is an attempt to provide a consistent and effective physiological basis for the observed time sequence as well as for other

MODIFICATION O F ASCIDIAX DEVELOPMEKT 509
features of development. I n short, the morphology of de- velopment reveals only a time sequence, but the physiology of development reveals, among other things, gradients which seem to account for the time sequence. The case of Corella seems to add a little more evidence pointing in the same direction as that already at hand.
Metamwphos i s awl elimination of the larlcal stage
Metamorphosis begins normally with the extremely rapid regression and resorption of the tail, and as this takes place the papillae begin to develop into more or less elongated stolons (figs. 6 to 8). The sequence of events gives the im- pression of a relation between tail resorption and develop- ment of the stolons, as if the tail inhibited stolon development. The change in position by apparent migration through almost 180" (figs. 5 to 10) is very evidently the result of differential growth and reduction of regions of the body wall. The anterodorsal region extends anteriorly and ventrally and the ventral region decreases in extent. The differential reduction of KhlnO, in the later stages indicates that the anterodorsal region of high reducing power extends around the anterior end to the ventral side as the stolons migrate. The sen- sory vesicle becomes somewhat more anterior, but its change in position is slight compared with that of the stolons (figs. 8 to 10). A possible interpretation of this apparent migra- tion is lack of adequate nutrition for all par ts in these ad- vanced stages. Under such conditions, in other forms very generally the more active regions maintain themselves or even grow a t the expense of the less active. Here the more active anterodorsal region may be extending at the expense of the less active ventral region.
Attachment has not been observed in the laboratory in any case, and as the tunic develops i t surrounds the stolons as well as other parts, in some cases apparently covering the siphon openings (figs. 45, 46, 64). Probably in nature the larvae react by attachment to certain kinds of surface, and the glass of the containers does not initiate the reaction. Experiments to settle this point were not performed.

510 C. M. CHILD
The inhibited forms show all degrees of reduction and elimination of the larval stage and approach to direct de- velopment, and may hatch only at an advanced stage of meta- morphosis. In other words, direct development in this form may result from differential inhibition of larval development, and the less extremely inhibited forms are capable of normal metamorphosis. After resorption of the tail, development apparently progresses more s l o ~ l y as compared with the extreme rapidity of development of prelarval stages and the later stages are apparently much less susceptible. Under conditions which inhibit larval development markedly, meta- morphosis may proceed in the usual course at about the usnal rate, but under the more extreme inhibiting conditions metamorphosis may be retarded o r even stopped. Further experiments on the control and modification of metamorphosis would be of intcrest.
Failure of the inhibited forms l o hatch may conceivably be due to small size, to the absence of tail movement, or to some unknown condition in the inhibited forms. When hatcli- irig occiirs in advanced metamorphosis, it appears to be due to increase in size of the animal.
The coizrlitions determiizing decelopmmit of swiinming lnrcac The evidence concerning the influence on development of
different conditions indicates that CO, is an important factor ixi determining the course of development. Apparently, the optimum f o r the development of swimming larvae is a CO, concentration approximately that of the water passing through the atrial chamber. Development in various acid- sea-water mixtures brought to the same higher pH by shaking shows that, at least within certain limits, the CO, rather than the pH must be the significant factor, f o r the mixtures made with the larger amounts of HCl, and which therefore require more shaking and lose more of their CO, in being raised to the desired pH, inhibit larval development, while those made with lower concentrations of HCl do not inhibit at the same pH. On the other hand, at a pH about neutrality

MODIFICATION O F ASCIDIAN D E V E L O P M E N T 511
or lower, development is markedly inhibited, probably be- cause the amount of CO, liberated by the acid here becomes sufficient to inhibit. Increase of the pH above that of normal sea-water by addition of NaOH also increases the inhibiting action, presumably by decreasing the free CO,. The much higher percentage of swimming larvae from eggs floating on a closed surface as compared with those on an open surface points in the same direction. On the closed surface the CO, produced by the developing embryos accumulates about them and a slight decrease in pH usually occurs throughout the container, while on the open surface the CO, passes off. These observations were made before the work of Smith and Clowes ('24 a, b) on the influence of CO,, as compared with the hydrogen-ion concentration of sea-water, on cell division, and other processes, but they are apparently in agreement with this work and the later wvork of Hyman ('25) and R. S. Lillie ( '26), though it was not possible to undertake quanti- tative determination of the CO, in the various mixtures. This part of the work on Corella is merely preliminary, being concerned primarily with the problem of obtaining control of development as a basis for further investigation. All the data obtained, however, indicate that CO, is the important factor in determining the development of swimming larvae in the atrial chamber and differential inhibition of larval development in normal sea-water. Undoubtedly, the effective- ness of CO, here as elsewhere results from its high penetrat- ing power, as compared with other acids (Jacobs, '12, '20 a, b, '22; Smith and Clowes, '24). The adjustment of the eggs of Corella to a CO, concentration approximately that of the atrial chamber and higher than that of sea-water is a fact of some interest. For these eggs normal sea-water does not constitute the optimum environment, as far as larval de- velopment is concerned, though it does not prevent the de- velopment of young ascidians.

512 C. M. CHILD
SUMXARY
1. The eggs of the ascidian, Corella willmeriana, develop in nature in the atrial cavity to swimming larvae, which make their \+ray to the exterior and metamorphose.
2. The differential reduction of K&fn04 in different stages of development indicates the existence in the early stages of high reducing power in the apical hemisphere. As develop- ment proceeds to the gastrula stage, this rapidly reducing region becomes anterior, anterodorsal, and dorsal.
3. As posterior elongation and tail development begin, a second region of high reducing power appears in the caudal region and during the period of tail development this becomes the most rapidly reducing region of the body, but loses this characteristic as resorption of the tail begins.
4. During the period of its outgrowth each stolon bud shows a gradient in reduction, decreasing from the tip basip- etally.
5. In later metamorphosis the anterodorsal reducing region extends around the anterior end as the stolons migrate poste- riorly.
G. All regional differences in reducing power disappear when, o r soon after, the animals are killed by other agents before they are brought into Ei&YInO4.
7. When the early developmental stages are removed from the atrial cavity to normal sea-water, larval development is differentially inhibited, the tail being most strongly inhibited, the dorsal body region next. The tail may fail to develop heyond early stages, the larval period may be eliminated, and the individual may undergo metamorphosis to an ad- vanced stage in the chorion.
8. The same modifications of development as in normal sea- water occur in sea-water to which certain amounts of HC1 o r NaOH have been added, but their occurrence is not dependent on the pH of the mixtures.
9. That the differential inhibition of development results from differential susceptibility of different regions, which in turn is dependent on their phpsiological activity, is indicatctl

M O D I F I C A T I O N O F ASCIDIAN D E V E L O P M E N T 513
by the fact that the regions most inhibited are those of highest reducing power and theref ore presumably most active.
10. The pH of the atrial chamber is lower than that of sea-water and the experiments performed in the attempt to obtain development of swimming larvae outside the atrial chamber indicate that a certain CO, concentration higher than that of normal sea-water and approximately that of the atrial chamber is necessary f o r the development of normal swim- ming larvae. Whether a sea-water to which acid has been added inhibits development or does not, does not depend upon the pH, but upon the amount of CO, set free in the water by the acid or removed by shaking. At a given pH, e.g., 7.4 to 7.6, a sea-water-acid mixture from which much of the CO, has been removed by shaking inhibits, while another from which little CO, has been removed gives a high per- centage of swimming larvae. The addition of small amounts of NaOH to sea-water brings about more extreme inhibition than normal sea-water, presumably because of the decrease in CO,.
11. The early developmental stages of Corella are ap- parently adjusted to a CO, concentration approximately that of the atrial chamber and probably near that of the body of the animal. Because of its high penetrability, CO, is the most effective factor in determining the intracellular hydrogen-ion concentration. The larval and postlarval stages become acclimated to sea-water.
LITERATURE CITED
EELLAMY, A. W. 1919 Differential susceptibility as a basis for modification I. Biol. Bull., vol. 37.
1923 Differential susceptibility as a basis, etc. 11. Am. Jour.
CHILD, C. M. Demonstration of the axial gradients by means of potas- Biol. Bull., vol. 36.
~~ 1921 The origin and development of the nervous system. Chicago. 1924 Physiological foundations of behavior. New York. 1923 The physiological significance of the cephalocaudal differ-
ential in vertebrate development. The organization and cell lineage of the ascidian egg.
Jour. Acad. Nat. Sciences Phila., second series, vol. 13, par t 1.
and control of early development in the frog.
Anat., vol. 30. 1919
sium permanganate.
Anat. Rec., vol. 31. CONKLIK, 3:. G. 190.5

5 14 C. M. C H I L D
IIINRIC'HS, &I. A. Rfodification of development on tlie basis of differential susceptibility to radiation. IV. Jour . Exp. Zool., vol. 47.
HI'XAP*, I,. 11. 1921 The rnetalmlic gr:idients of rertpbrate embryos. I. Biol. Bull., vol. 40.
On tlie action of certain substances on oxygen consumption. PI. Biol. Bull., vol. 49.
.___ 1926 The metabolic gradients of vertehrnte embryos. 11. Jour . Morph. and Pliysiol., vol. 42.
____ 1927 The metabolic gradients of Tertehrate emhryos. 111. Biol.
JACOBS, h1. 11. 1912 Studies on the pli? siological cliaracteristies of species. I. Jour. EXIJ. Zool., vol. 12.
____ 1920 il To what extent arc tlie physiological effects of carbon dioside dnc t o liydrogen ions.
____ 1920 b The production of intracellular acidity by neutr:il or :ilkaline solutions containing ear lmi dioxide. Amer. Jour. Physiol., vol. 53.
____ 1922 The effects of carbon dioxide on the consistency of proto- plasm. Biol. Hull., vol. 42.
The significance of the so-called 1:tw of cephalocaudal tlifferential growth.
~- 1926 On the so-called lax of anteroposterior development. Anat. Rec., vol. 33.
LILLIE, R. S. 1926 The nctiration of st:irfisli eggs by acids. Jour . Gen. Physiol., rol. 8.
SaIITH, I[. w., AND (>LOTT*ES, G. H. A. 1924 n The influence of carlmn dioxide on the velocity of ilivisioii of marine eggs. Amer. Jour. Physiol., rol. 68.
The influence of hydrogen ion concentration on unfertilized Arhncia, Asterias and Chaetopterus eggs.
1927
._ 192.5
Bull., vol. 5 2 .
Amer. Jour . Physiol., vol. 51.
BTRY, R. F. 1924 Anat. Rec., vol. 27.
__-- _- - 1924 b Biol. Bull., rol. 47.