metamorphosis of the invasive ascidian cionasavignyi ... · pdf filethe larval settlement and...
TRANSCRIPT

Submitted 1 December 2015Accepted 5 February 2016Published 25 February 2016
Corresponding authorPatrick L. Cahill,[email protected]
Academic editorJoseph Pawlik
Additional Information andDeclarations can be found onpage 13
DOI 10.7717/peerj.1739
Copyright2016 Cahill et al.
Distributed underCreative Commons CC-BY 4.0
OPEN ACCESS
Metamorphosis of the invasive ascidianCiona savignyi: environmental variablesand chemical exposurePatrick L. Cahill, Javier Atalah, Andrew I. Selwood and Jeanne M. KuhajekCawthron Institute, Nelson, New Zealand
ABSTRACTIn this study, the effects of environmental variables on larval metamorphosis of thesolitary ascidian Ciona savignyi were investigated in a laboratory setting. The progres-sion of metamorphic changes were tracked under various temperature, photoperiod,substrate, larval density, and vessel size regimes. Metamorphosis was maximised at18 ◦C, 12:12 h subdued light:dark, smooth polystyrene substrate, and 10 larvae mL−1
in a twelve-well tissue culture plate. Eliminating the air-water interface by fillingculture vessels to capacity further increased the proportion of metamorphosed larvae;87 ± 5% of larvae completed metamorphosis within 5 days compared to 45 ± 5%in control wells. The effects of the reference antifouling compounds polygodial,portimine, oroidin, chlorothalonil, and tolylfluanid on C. savignyi were subsequentlydetermined, highlighting (1) the sensitivity of C. savignyi metamorphosis to chemicalexposure and (2) the potential to use C. savignyi larvae to screen for bioactivity in anoptimised laboratory setting. The compounds were bioactive in the low ng mL−1 tohigh µg mL−1 range. Polygodial was chosen for additional investigations, where it wasshown that mean reductions in the proportions of larvae reaching stage E were highlyrepeatable both within (repeatability= 14± 9%) and between (intermediate precision= 17 ± 3%) independent experiments. An environmental extract had no effect on thelarvae but exposing larvae to both the extract and polygodial reduced potency relativeto polygodial alone. This change in potency stresses the need for caution when workingwith complex samples, as is routinely implemented when isolating natural compoundsfrom their biological source. Overall, the outcomes of this study highlight the sensitivityof C. savignyi metamorphosis to environmental variations and chemical exposure.
Subjects Ecology, Marine BiologyKeywords Ciona savignyi, Metamorphosis, Antifouling, Bioactivity, Environmental variables
INTRODUCTIONThe Pacific transparent ascidian Ciona savignyi Herdman (Cionidae) is a Japanese nativewith an expanding invasive range that currently includes Argentina, British Columbia,California, NewZealand, Puget Sound, and Spain (Fofonoff et al., 2003; Lambert & Lambert,1998; Smith, Cahill & Fidler, 2010).Ciona savignyi is recognised as a problematic biofoulingorganism; it reproduces rapidly in invaded environments and can dominate man-madeand natural substrates (e.g., Cohen et al., 1998; Zvyagintsev, Sanamyan & Kashenko, 2007).This hermaphroditic species can spawn year-round in temperate regions (Nomaguchi etal., 1997; P Cahill, pers. obs.), with each individual releasing hundreds or thousands of eggsto be fertilised in the water column (Hendrickson et al., 2004; Jiang & Smith, 2005).
How to cite this article Cahill et al. (2016), Metamorphosis of the invasive ascidian Ciona savignyi: environmental variables and chemicalexposure. PeerJ 4:e1739; DOI 10.7717/peerj.1739

Any attempt to counteract the invasive tendencies of C. savignyi requires an in-depthunderstanding of this species life-history characteristics. A key step in the biofoulingprocess is the transition from free-swimming larva to sessile adult (Pawlik, 1992). Larvaemust contact a suitable surface upon which to settle, choose to attach, and then undergoa complex series of morphological changes to form established juveniles. Marine larvaetypically respond to a range of environmental and con-specific cues, with a high degree ofvariability in responses between species (Jackson et al., 2002; Rodriguez, Ojeda & Inestrosa,1993). Little information is available on the extrinsic factors that stimulate C. savignyi tometamorphose; identifying these cues will facilitate the development of targeted treatmenttechnologies and mitigation techniques.
In particular, the metamorphic process may be sensitive to exposure to chemicalcompounds. It has been shown that C. savignyi metamorphosis is inhibited by the naturalantifouling agent polygodial (Cahill & Kuhajek, 2014), with other natural (e.g., oridin orportimine; Selwood et al., 2013; Tsukamoto et al., 1996) and synthetic (e.g., chlorothaloniland tolyfluanid;Voulvoulis, Scrimshaw & Lester, 1999) compounds also likely to be effectiveagainst C. savignyimetamorphosis. Screening for effects on C. savignyimetamorphosis haspotential to identify targeted lead compounds, including both known compounds and novelnatural compounds. In the case of novel natural compounds, initial screening and isolationtypically involves working with complex biological extracts, with potential for interactionsbetween constituents of the extract (Colegate & Molyneux, 2007). Understanding howenvironmental factors influencemetamorphosis will improve our ability to reliably quantifyantimetamorphic effects.
In addition to the interest in C. savignyi as a marine invader, this species has beenincreasingly studied as a model organism for developmental biology (Corbo, Di Gregorio& Levine, 2001; Sasakura et al., 2012; Satoh, 2003). Ascidians occupy an intriguingevolutionary position as sister clade to the vertebrates (Lemaire, 2011; Satoh & Levine,2005; Schubert et al., 2006), meaning they a can afford insights into developmental biologyin general. Embryos and larvae of C. savignyi can be produced in large numbers in the lab,the latter undergoing a defined progression of metamorphic changes to form establishedjuveniles within 7 days (Cirino et al., 2002; Hendrickson et al., 2004; Kourakis, Newman-Smith & Smith, 2010). Many studies have examined the intrinsic determinants driving theprogression from egg, to free-swimming larvae, to sessile adult in C. savignyi (e.g., Imai,Satoh & Satou, 2002a; Imai, Satou & Satoh, 2002b; Kimura, Yoshida & Morisawa, 2003).Identifying extrinsic conditions that stimulate ascidian larvae to settle and metamorphosewill also provide a context for these molecular and biochemical investigations(Morse, 1990).
In this study, the effects of extrinsic environmental factors onC. savignyimetamorphosiswere examined in a laboratory setting. Variables investigated included temperature,photoperiod, substrate, larval density, and vessel size. The most susceptible stages ofthe larval settlement and metamorphosis processes were identified, providing a relevantexperimental end-point for investigating the effects of reference antifouling compoundson the larvae.
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 2/18

MATERIAL AND METHODSCulture and spawningAdult C. savignyi, collected from the underside of pontoons at Nelson Marina, Nelson,New Zealand, were housed in water lily baskets suspended in 10-L glass aquaria for up tothree weeks. Aquaria were supplied with 10 L seawater h−1 as part of a 1000-L recirculatingsystem held at 18 ± 1 ◦C, 34 ± 1 psu, and 300 ± 50 mV ORP; constant full-spectrumflorescent light prevented premature spawning. Daily, the flow of water to the aquaria wascut off for 3 h while C. savignyi were fed 250 mL of an 8−9 ×106 cells mL−1 Isochrysisgalbana Parke culture. Three gravid individuals with densely packed egg and sperm ductswere spawned according to Cirino et al. (2002). Ventral incisions were made to expose theegg and sperm ducts. The egg duct of each individual was pierced with a Pasteur pipetteand the eggs transferred to a glass Petri dish (90 mm dia., 68 mL vol.) filled with 20 mLof 0.3-µm filtered and UV-sterilized seawater (FSW). Sperm were then harvested andtransferred to a glass Petri dish (90 mm dia., 68 mL vol.) containing 50 mL FSW. Each dishof eggs received eight drops of sperm suspension from each of the two other individuals.After incubating at 18 ◦C for 1 h, fertilized eggs were strained through a 20-µm sieve,rinsed three times with 25 mL reconstituted seawater (RSW; 33 ± 0.5 psu; Red Sea Salt,Red Sea Aquatics, Cheddar, UK), transferred to a glass Petri dish along with 25 mL RSW,and held at 18 ◦C for 18 h to hatch. Hatched larvae were transferred to conical flasks anddiluted with RSW as necessary to yield desired larval densities.
Temperature, photoperiod, and substrateThe temperature, photoperiod, and substrate preferences of larvae were assessed in threeseparate experiments. Firstly, the effects of exposing larvae to 15, 18, 21, and 25 ◦C wereevaluated. For the photoperiod experiment, larvae were exposed to the following lightingregimes: 24:0, 12:12, 0:24 h intense (100± 10µmolm−2 s−1) or subdued (10± 5µmolm−2
s−1) light:dark. Substrate type (polystyrene, acrylic, or glass) and texture (smooth or rough)were assessed using unlined wells (polystyrene) or wells lined with acrylic discs (35-mmdiameter) or glass cover slips (35-mm diameter, Gerhard Menz GmbH, Saarbrücken, GE).Untreated unlined wells, acrylic discs, and glass cover slips comprised smooth treatments.Sandblasted acrylic and sanded (800-grit sandpaper) glass and polystyrene comprisedrough treatments; it should be noted that these surface treatments likely resulted indifferent feature sizes for each of the three substrate types and that the walls of the wellsremained untreated in all cases.
All three experiments (temperature, photoperiod, and substrate) were performed insix-well tissue culture plates (Corning R© Costar R©, Corning Inc., Corning, NY; 36 mm dia.;17.5 mL vol.) filled with 10-mL aliquots of 2.5 larvae mL−1 RSW. Default parameterswere 18 ◦C, 12:12 h subdued light:dark, and unlined smooth wells; three replicates wereperformed in all cases (n= 3). Culture waters were renewed with RSW every other dayand settled larvae were scored according to metamorphic stage (Fig. 1) after 1, 3, 5, and 7days. Unattached larvae or larvae adhered to the meniscus were not counted. A proxy fordaily metamorphic progress was calculated based on the number of larvae counted at each
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 3/18
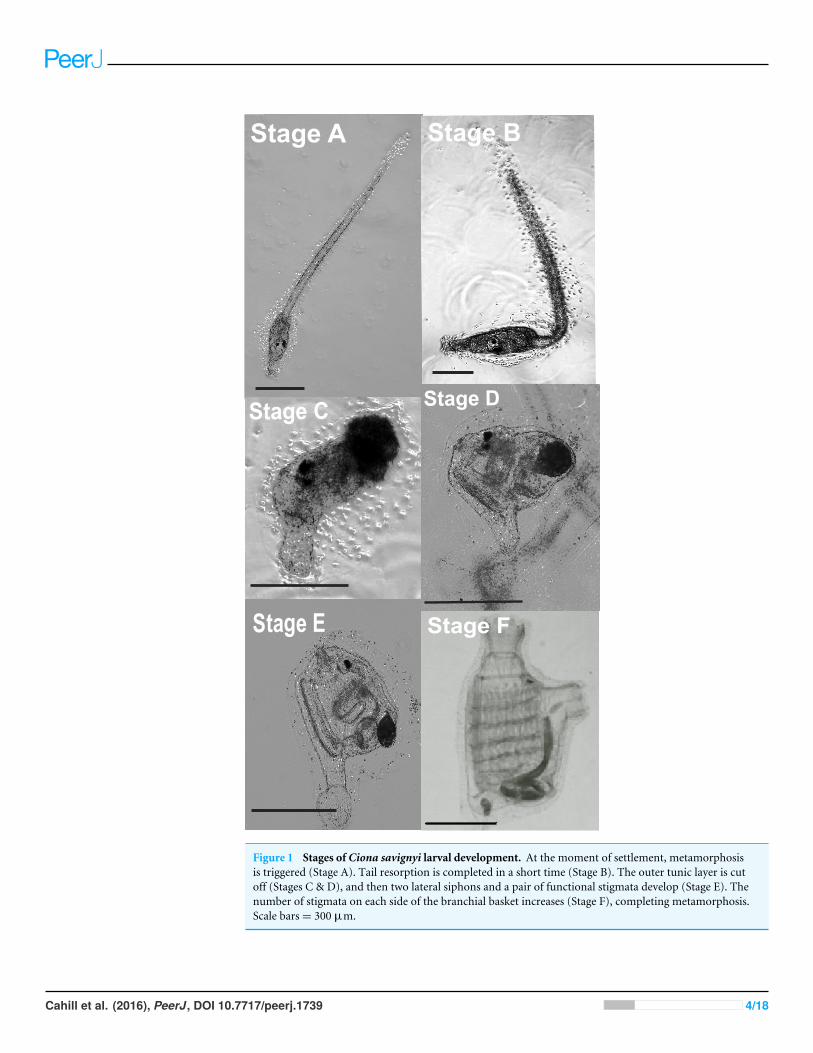
Figure 1 Stages of Ciona savignyi larval development. At the moment of settlement, metamorphosisis triggered (Stage A). Tail resorption is completed in a short time (Stage B). The outer tunic layer is cutoff (Stages C & D), and then two lateral siphons and a pair of functional stigmata develop (Stage E). Thenumber of stigmata on each side of the branchial basket increases (Stage F), completing metamorphosis.Scale bars= 300 µm.
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 4/18

developmental stage as follows:
Metamorphic progress= (Stage A larvae×1)+ (Stage B larvae×2)+ (Stage C larvae×3)
+(Stage D larvae×4)+ (Stage E larvae×5)+ (Stage F larvae×6).
Vessel size, larval density, and fill volumeVessel sizewas assessed in conjunctionwith larval density. Polystyrene Petri dishes (LabServ,Thermo Fisher Scientific, Melbourne, AU; 90 mm dia.; 68 mL vol.), six-well plates, andtwelve-well plates (Corning R© Costar R©; 23 mm dia.; 6.5 mL vol.) were filled with 25-, 10-,or 5-mL aliquots of larval suspension, respectively. Four larval densities (2.5, 5, 10, and 15larvae mL−1) were tested in each of the three vessel sizes, yielding twelve vessel size× larvaldensity treatments in total (n= 3). Vessels were held at 18 ◦C, 12:12h subdued light to darkand culture waters were renewed every other day. After 5 days, the number of larvae thathad completed stage E of metamorphosis were counted (Fig. 1).
Fill volume was assessed using twelve-well plates and a larval density of 10 larvaemL−1. Treatment wells were filled to capacity with 7.1 mL of larval suspension so that theseawater contacted the underside of the lid, eliminating the air-water interface. Controlswells were filled with 5 mL of larval suspension (n= 3). The temperature, photoperiod,and experimental end-point matched those used for the vessel size and larval densityexperiments but culture waters were not renewed for the duration of this experiment.
Carrier solvent and reference compoundsThe sensitivity of larvae to two common carrier solvents was assessed in twelve-well plateswith 7.1 mL aliquots of 10 larvae mL−1 RSW; held at 18 ◦C, 12:12 h subdued light to dark.Concentrations of ethanol and dimethyl sulfoxide (DMSO) evaluated were 0.05, 0.1, 0.25,0.5, 1, 2.5, 5, 10, 20, and 40 µL mL−1; controls contained no solvent (n= 3). Larvae thathad completed stage E of metamorphosis were counted after 5 days.
Five reference compounds, polygodial (ENZO Life Sciences, Farmingdale, NY, USA),portimine (Cawthron Natural Compounds, Nelson, NZ), oroidin (ENZO Life Sciences),chlorothalonil (Sigma-Aldrich, St Louis,MO,USA), and tolylfluanid (Sigma-Aldrich), werescreened using the protocol outlined for carrier solvent. With the exception of polygodial(1, 2.5, 5, 10, 15, 20, 40, 80 ng mL−1) and portimine (0.05, 0.1, 0.5, 1, 5, 10, 20 ng mL−1),concentrations tested against the larvae were 0.001, 0.005, 0.01, 1, 5, and 10 µg mL−1.Stock solutions prepared in 20% (v/v) ethanol (polygodial), RSW (portimine), or 20%(v/v) DMSO (oroidin, chlorothalonil, tolylfluanid) were added to wells to yield the desiredtest concentrations; control wells contained solvent only (n= 3).
Polygodial treatments (5, 10, and 50 ng mL−1; n= 3) were subsequently included ineight independent experiments performed over a 1-year period, allowing repeatability(i.e., intra-run variability) and intermediate precision (i.e., inter-run variability) to beestimated. In a separate experiment assessing the potential for interactions among complexmixtures of natural compounds, the potency of pure polygodial was compared to that ofpolygodial enriched with an environmental extract. The extract was produced by eluting10 L seawater that ∼400 g of green-lipped mussels, Perna canaliculus Gmelin, had been
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 5/18

cultured in for 1 day through a 20-cm3 column of DiaionHP20 resin (Mitsubishi Chemical,Tokyo, JP). The column was flushed with 50 mL ethanol, and the resulting extract driedand re-suspended in 1 mL 20% (v/v) ethanol. Polygodial stock solutions prepared in 1mL of 20% (v/v) ethanol or 1 mL of extract were added to wells to yield final polygodialconcentrations of 2.5, 5, 10, and 50 ng mL−1 (n= 3).
Data analysesTemperature, photoperiod, and substrate data were analyzed using one-way linearmixed-effects modelling (Bolker et al., 2009), with metamorphic progress (‘Temperature,photoperiod, and substrate’) as the response variable, treatment as a fixed factor, and timeas a continuous covariate. Replicate was included as a random effect to account for therepeated-measures experimental design. Reported p-values are based on the t distributionof the ratios between the estimates and their standard errors (Pinheiro & Bates, 2006).Principal response curves (PRC), a redundancy analysis for multivariate responses inrepeated-measures design (Van den Brink & Ter Braak, 1998; Van den Brink & Ter Braak,1999), were used with 999 permutations to identify the metamorphic stages that weredriving treatment effects (>0.5 signifies strong treatment effect). Mean absolute PRCcoefficients for the number of larvae in each metamorphic stage (A–F) at each time pointwere calculated by averaging the values from the temperature, photoperiod, and substratedatasets. The effects of larval density and vessel size were assessed using a two-way factorialANOVA with the number of Stage E larvae on day 5 as the response variable. The effectof fill-volume on the number of larvae reaching Stage E within 5 days was tested using aStudent’s t -test, and carrier solvent using one-way ANOVA followed byDunnett’s post-hoctest. Dose-response curves were plotted for the reference compounds using four parameterlogistic curve fitting, and the corresponding concentrations that reduced the number oflarvae reaching Stage E by 50% relative to blank controls (EC50) calculated (Kuo, Mitchell& Tuerke, 1993). Repeatability (relative bias) and intermediate precision for the replicatepolygodial experiments were calculated in accordance with USP <1033 >(2012). Analyseswere performed using R 2.13.1 (R Development Core Team, 2015) and SigmaPlot 11.0(Systat Software Inc, 2015).
RESULTSOverall, larvae reached stages A and B of metamorphosis within 1 day, while stage C took1.2 ± 0.06 days, stage D 3.1 ± 0.02 days, stage E 4.3 ± 0.03 days, and stage F 6.6 ± 0.04days.
Temperature, photoperiod, and substrateLarvae responded to variations in temperature, photoperiod, and substrate. Dailymetamorphic progress was comparable for larvae reared at 15, 18, or 21 ◦C but hinderedat 25 ◦C (t =−4.6, p< 0.001; Fig. 2A). Metamorphic progress was enhanced relativeto the other photoperiod regimes evaluated when exposed to 12:12 h subdued light:dark(t = 2.1,p= 0.04), and slowed by exposure to constant intense light (t = 3.0, p= 0.004;Fig. 2B). Metamorphic progress was reduced when the substrate was smooth acrylic
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 6/18

Figure 2 Effects of (A) temperature, (B) photoperiod, and (C) substrate on Ciona savignyi larvae.Metamorphic progress was calculated based on the sum of the number of individuals reaching each meta-morphic stage converted to numerals (A= 1, B= 2, C= 3, D= 4, E= 5, F= 6). Values are means (n =3)± 95% confidence interval.
(t =−3.5,p= 0.04; Fig. 2C) but there were no discernable differences between theother substrate types tested. The statistical differences observed for the temperature(PRC, F = 10.1,p= 0.005), photoperiod (PRC, F = 8.4, p= 0.005), and substrate (PRC,F = 1.9,p= 0.005) datasets were driven by the latter stages of metamorphosis. Mean
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 7/18
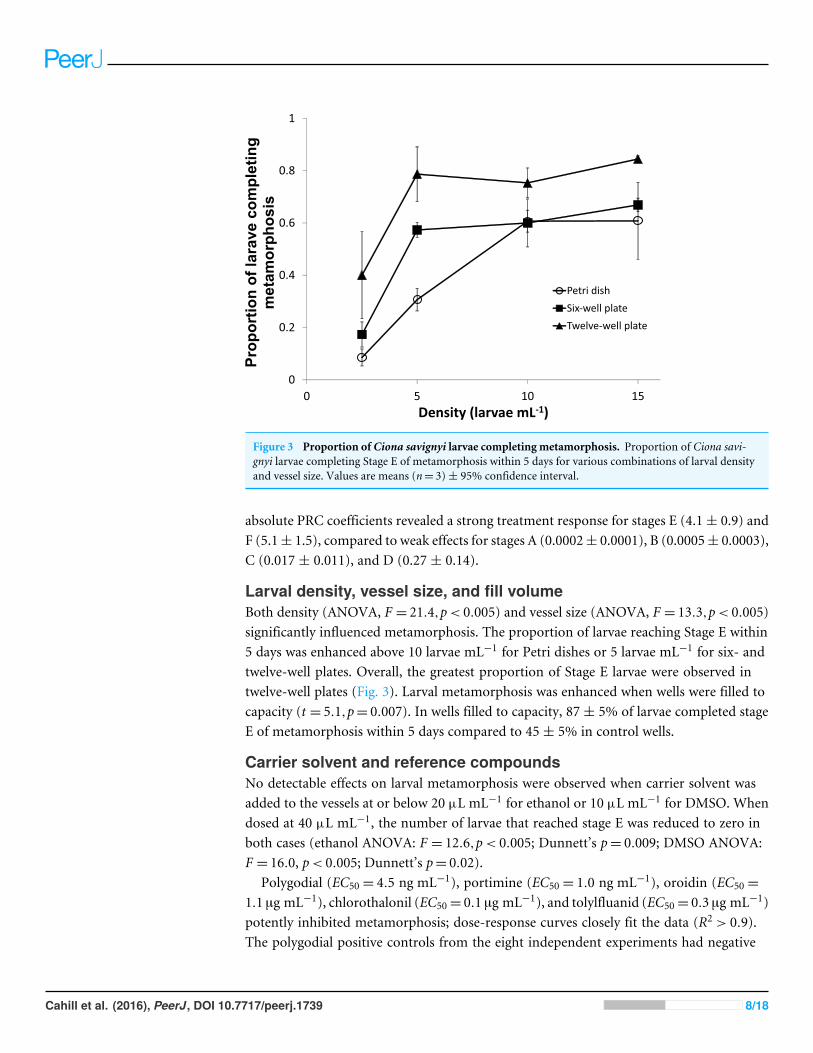
Figure 3 Proportion of Ciona savignyi larvae completing metamorphosis. Proportion of Ciona savi-gnyi larvae completing Stage E of metamorphosis within 5 days for various combinations of larval densityand vessel size. Values are means (n= 3)± 95% confidence interval.
absolute PRC coefficients revealed a strong treatment response for stages E (4.1± 0.9) andF (5.1± 1.5), compared to weak effects for stages A (0.0002± 0.0001), B (0.0005± 0.0003),C (0.017 ± 0.011), and D (0.27 ± 0.14).
Larval density, vessel size, and fill volumeBoth density (ANOVA, F = 21.4,p< 0.005) and vessel size (ANOVA, F = 13.3,p< 0.005)significantly influenced metamorphosis. The proportion of larvae reaching Stage E within5 days was enhanced above 10 larvae mL−1 for Petri dishes or 5 larvae mL−1 for six- andtwelve-well plates. Overall, the greatest proportion of Stage E larvae were observed intwelve-well plates (Fig. 3). Larval metamorphosis was enhanced when wells were filled tocapacity (t = 5.1,p= 0.007). In wells filled to capacity, 87 ± 5% of larvae completed stageE of metamorphosis within 5 days compared to 45 ± 5% in control wells.
Carrier solvent and reference compoundsNo detectable effects on larval metamorphosis were observed when carrier solvent wasadded to the vessels at or below 20 µL mL−1 for ethanol or 10 µL mL−1 for DMSO. Whendosed at 40 µL mL−1, the number of larvae that reached stage E was reduced to zero inboth cases (ethanol ANOVA: F = 12.6,p< 0.005; Dunnett’s p= 0.009; DMSO ANOVA:F = 16.0, p< 0.005; Dunnett’s p= 0.02).
Polygodial (EC50= 4.5 ng mL−1), portimine (EC50= 1.0 ng mL−1), oroidin (EC50=
1.1 µg mL−1), chlorothalonil (EC50= 0.1 µg mL−1), and tolylfluanid (EC50= 0.3 µg mL−1)potently inhibited metamorphosis; dose-response curves closely fit the data (R2 > 0.9).The polygodial positive controls from the eight independent experiments had negative
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 8/18

Figure 4 Relative bias for polygodial treatments. Relative bias from eight independent experiments forthe effects of for polygodial treatments on Ciona savignyimetamorphosis. Values are means (n = 8) rela-tive to a known dose-response relationship for polygodial± 95% confidence interval.
Table 1 Overall variability for polygodial treatments.Variance component estimates and overall vari-ability for the effects of polygodial treatments on Ciona savignyimetamorphosis.
5 ngmL−1 10 ngmL−1 Mean
Var (Run) −0.013 0.002 −0.005Var (Error) 0.033 0.030 0.031Overall 15% 20% 17%
dose-dependent effects on metamorphosis, with mean reductions in the proportions oflarvae reaching stage E varying by 14 ± 9% within experiments (i.e., repeatability) and17 ± 3% between experiments (i.e., intermediate precision; Fig. 4, Table 1). However,enrichment with an environmental extract decreased the potency of polygodial (Fig. 5).The extract alone had no detectable effect on metamorphosis but the observed shift inpotency for enriched vs. pure polygodial represents an approximately 80% increase in EC50
(7.8 vs. 4.3 ng mL−1).
DISCUSSIONThe larvae of C. savignyi were well suited to laboratory culture and, as has been reportedpreviously, they completedmetamorphosis to established juveniles inside seven days (Cirinoet al., 2002; Hendrickson et al., 2004). The proportion of larvae completing metamorphosisto stage E within five days exceeded 90% in some cases, but the performance of the larvaewas highly dependent on environmental factors. Temperature and photoperiod stronglyinfluenced metamorphic progress. Although no observable differences were found between
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 9/18
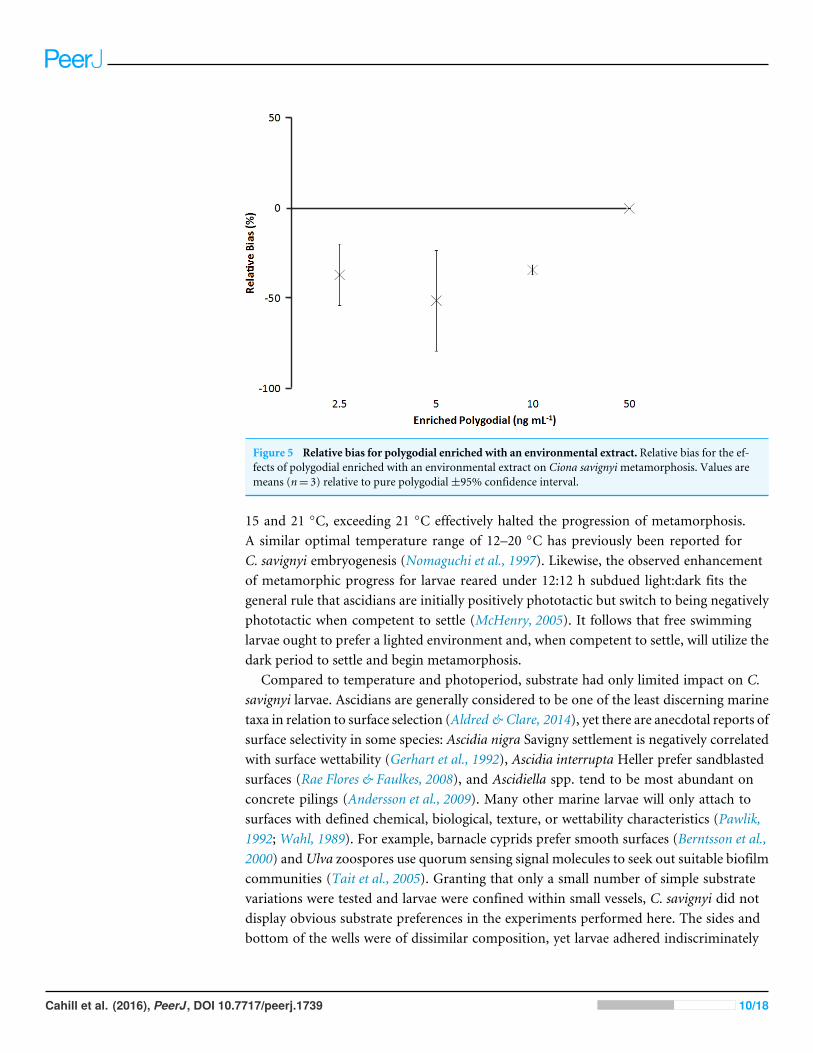
Figure 5 Relative bias for polygodial enriched with an environmental extract. Relative bias for the ef-fects of polygodial enriched with an environmental extract on Ciona savignyimetamorphosis. Values aremeans (n= 3) relative to pure polygodial±95% confidence interval.
15 and 21 ◦C, exceeding 21 ◦C effectively halted the progression of metamorphosis.A similar optimal temperature range of 12–20 ◦C has previously been reported forC. savignyi embryogenesis (Nomaguchi et al., 1997). Likewise, the observed enhancementof metamorphic progress for larvae reared under 12:12 h subdued light:dark fits thegeneral rule that ascidians are initially positively phototactic but switch to being negativelyphototactic when competent to settle (McHenry, 2005). It follows that free swimminglarvae ought to prefer a lighted environment and, when competent to settle, will utilize thedark period to settle and begin metamorphosis.
Compared to temperature and photoperiod, substrate had only limited impact on C.savignyi larvae. Ascidians are generally considered to be one of the least discerning marinetaxa in relation to surface selection (Aldred & Clare, 2014), yet there are anecdotal reports ofsurface selectivity in some species: Ascidia nigra Savigny settlement is negatively correlatedwith surface wettability (Gerhart et al., 1992), Ascidia interrupta Heller prefer sandblastedsurfaces (Rae Flores & Faulkes, 2008), and Ascidiella spp. tend to be most abundant onconcrete pilings (Andersson et al., 2009). Many other marine larvae will only attach tosurfaces with defined chemical, biological, texture, or wettability characteristics (Pawlik,1992; Wahl, 1989). For example, barnacle cyprids prefer smooth surfaces (Berntsson et al.,2000) andUlva zoospores use quorum sensing signal molecules to seek out suitable biofilmcommunities (Tait et al., 2005). Granting that only a small number of simple substratevariations were tested and larvae were confined within small vessels, C. savignyi did notdisplay obvious substrate preferences in the experiments performed here. The sides andbottom of the wells were of dissimilar composition, yet larvae adhered indiscriminately
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 10/18

to all surfaces. The exception was wells lined with smooth acrylic, where the overallprogression of metamorphosis was reduced by approximately 12% compared to the othersurfaces tested. This apparent insensitivity to surface characteristics may contribute toC. savignyi’s invasive character, whereby larvae will settle on a wide range of availablesurfaces.
When the temperature, photoperiod, and substrate datasets were combined, the earliestdevelopmental stage of metamorphosis to show strong treatment response was stage Ewhen the branchial basket, siphons, and stigmata develop. It has previously been notedthat tail-resorption is a particularly sensitive stage of ascidian metamorphosis (e.g., Bishop,Bates & Brandhorst, 2001; Eri et al., 1999; Green et al., 2002) but, in the case of C. savignyi,the tail resorption process (Stage B) was little affected by variations in temperature,photoperiod, or substrate. It is not clear why the later stages of metamorphosis were moresensitive to environmental variations but the lecithotrophic nature of C. savignyi maybe partly responsible. Energy reserves within the larvae may become depleted later inmetamorphosis (e.g., Jaeckle, 1994; Jaeckle & Manahan, 1989; Moran & Manahan, 2003),with energy deficient larvae becoming increasing susceptible to sub-optimal environmentalconditions.
When larval density and vessel size were subsequently evaluated, the proportion of larvaereaching Stage E of metamorphosis was enhanced above 5 or 10 larvae mL−1, with twelve-well plates having the greatest proportion of Stage E larvae overall. Density-dependentbehaviours are common among marine larvae (Hadfield & Paul, 2001); examples includethe gregariousness of larvae of the barnacle Balanus amphitrite Darwin (Head et al., 2003),the tubeworm Hydroides dianthus Verrill (Toonen & Pawlik, 1996), and the oyster Ostreaedulis L. (Bayne, 1969). Enhanced metamorphosis in smaller vessels could be a resultof conspecific settlement cues or reduced surface area to volume ratio. Under the laterscenario, larvae are more likely to contact, and thus adhere to, the internal surfaces of asmaller vessel.
Filling wells to capacity further increased the proportion of Stage E larvae observed after5 days. When wells were not filled to capacity, some C. savignyi larvae became trapped atthe air-water interface and where not considered to have successfully attached (P Cahill,pers. obs.). This phenomenon has been reported previously for other ascidians (e.g.,Fletcher & Forrest, 2011), and is probably due to the larvae swimming upwards against theforce of gravity for the first few hours after hatching (Hendrickson et al., 2004). Becauseculture waters contact the underside of the lid in wells that were filled to capacity, larvaeare provided with an inverted surface upon which to settle and are prevented fromthem becoming trapped at the air-water interface. In line with these findings, it has beenanecdotally observed that C. savignyi adults tend to be congregated on the underside ofman-made structures in the sea (P Cahill, pers. obs.).
Evaluating the effects of select reference compounds on C. savignyi metamorphosishighlighted the potential to use C. savignyi larvae to screen for bioactivity under anoptimised laboratory setting. The natural antifouling agent polygodial (Cahill & Kuhajek,2014),the algal biotoxin portimine (Selwood et al., 2013), and the synthetic antifoulingbiocides chlorothalonil and tolyfluanid (Voulvoulis, Scrimshaw & Lester, 1999) were
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 11/18

bioactive in the low ng mL−1 to high µg mL−1 ranges. Tolerance of C. savignyi larvae forthe carrier solvents ethanol (20 µL mL−1 max.) and DMSO (10 µL mL−1 max.) facilitatedscreening these compounds, which, with the exception of portimine, have only limitedwater solubility. The results reported here meet or exceed potency estimates previouslydetermined for these compounds against other organisms. For example, polygodial iseffective against fungal pathogens at approximately 1 µg mL−1 (Kubo & Himejima, 1991),chlorothalonil kills water fleas and fathead minnows at 0.03–0.2 µg mL−1 (Sherrard et al.,2002), and tolylfluanid controls seaweed zoospores at 0.03 µg mL−1 (Wendt et al., 2013).However, systems that test the bioactivity of compounds in solution must be carefullyinterpreted. In nature, marine larvae are typically only exposed to allelopathic metabolitesat surfaces, such as when they contact another benthic organisms (Pawlik, 1993). Perfusinglarval tissues with a compound, as was done here, provides a largely pharmacologicalrather than ecological context. This pharmacological data is often used as a first step toidentify promising bioactive compounds but should always be followed by additionalinvestigations, either laboratory or field based, where the compound of interest is boundto/released from a surface (Bressy et al., 2014).
When polygodial treatments were included in eight independent experiments performedover a one year period, mean reductions in the proportion of larvae reaching stage E variedby 14 ± 9% and 17 ± 3% within and between experiments, respectively. These valuesrepresent a relatively high degree of consistency for the effects of polygodial exposure,with comparable larval systems returning repeatability estimates in the order of 5–30%(e.g., Piazza et al., 2012; Ross & Bidwell, 2001; Stronkhorst et al., 2004). The larvae wereunaffected by a complex environmental extract produced from the culture waters ofgreen-lipped mussels but the same extract reduced the potency of polygodial to almosthalf. This result highlights the potential for changes in potency when working with complexchemical samples, as is common when attempting to isolate natural products (Colegate &Molyneux, 2007).
Overall, this study yielded insights into the environmental factors impactingmetamorphosis ofC. savignyi larvae. This increased understanding of the interplay betweenthe environment and the biology of C. savignyi is vital to understanding, and perhapscounteracting, the invasive tendencies of this organism. The findings also provide contextfor the growing body of research examining the biochemical and genetic determinants ofC. savignyimetamorphosis. Demonstrating that C. savignyi larvae are sensitive to a range ofnatural and synthetic bioactive agents highlighted one potential application for these larvae.Using C. savignyi larvae to assess the toxicity of compounds under optimised laboratoryconditions presents a potentially useful preliminary screening tool but the outcomes ofsuch experiments must be carefully interpreted and should be followed by ecologicallyrelevant investigations. Nevertheless, chemicals targeting C. savignyimetamorphosis couldpotentially be used to control this pest species.
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 12/18

ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis study was funded by the New Zealand Ministry of Business Innovation andEmployment (CAWX1315). The funders had no role in study design, data collectionand analysis, decision to publish, or preparation of the manuscript.
Grant DisclosuresThe following grant information was disclosed by the authors:New Zealand Ministry of Business Innovation and Employment: CAWX1315.
Competing InterestsThe authors declare there are no competing interests.
Author Contributions• Patrick L. Cahill conceived and designed the experiments, performed the experiments,analyzed the data, wrote the paper, prepared figures and/or tables.• Javier Atalah analyzed the data, reviewed drafts of the paper.• Andrew I. Selwood conceived and designed the experiments, contributed reagents/ma-terials/analysis tools, reviewed drafts of the paper.• Jeanne M. Kuhajek conceived and designed the experiments, reviewed drafts of thepaper.
Data AvailabilityThe following information was supplied regarding data availability:
The raw data has been supplied as Data S1.
Supplemental InformationSupplemental information for this article can be found online at http://dx.doi.org/10.7717/peerj.1739#supplemental-information.
REFERENCESAldred N, Clare AS. 2014.Mini-review: impact and dynamics of surface fouling by
solitary and compound ascidians. Biofouling 30:259–270DOI 10.1080/08927014.2013.866653.
AnderssonMH, BerggrenM,Wilhelmsson D, ÖhmanMC. 2009. Epibenthic coloniza-tion of concrete and steel pilings in a cold-temperate embayment: a field experiment.Helgoland Marine Research 63:249–260 DOI 10.1007/s10152-009-0156-9.
Bayne B. 1969. The gregarious behaviour of the larvae of Ostrea edulis L. at settlement.Journal of the Marine Biological Association of the United Kingdom 49:327–356DOI 10.1017/S0025315400035943.
Berntsson KM, Jonsson PR, Lejhall M, Gatenholm P. 2000. Analysis of behaviouralrejection of micro-textured surfaces and implications for recruitment by the barnacle
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 13/18

Balanus improvisus. Journal of Experimental Marine Biology and Ecology 251:59–83DOI 10.1016/S0022-0981(00)00210-0.
Bishop CD, BatesWR, Brandhorst BP. 2001. Regulation of metamorphosis in ascidiansinvolves NO/ cGMP signaling and HSP90. Journal of Experimental Zoology289:374–384 DOI 10.1002/jez.1019.
Bolker BM, Brooks ME, Clark CJ, Geange SW, Poulsen JR, Stevens MHH,White J-SS.2009. Generalized linear mixed models: a practical guide for ecology and evolution.Trends in Ecology & Evolution 24:127–135 DOI 10.1016/j.tree.2008.10.008.
Bressy C, Briand JF, Compère C, Réhel K. 2014. Efficacy testing of biocides and biocidalcoatings. In: Biofouling methods. Hoboken: Wiley Blackwell, 332–345.
Cahill P, Kuhajek J. 2014. Polygodial: a contact active antifouling biocide. Biofouling30:1035–1043 DOI 10.1080/08927014.2014.966305.
Cirino P, Toscano A, Caramiello D, Macina A, Miraglia V, Monte A. 2002. Labora-tory culture of the ascidian Ciona intestinalis (L.): a model system for moleculardevelopmental biology research. In:Marine models electronic records. Available athttp://www.mbl.edu/BiologicalBulletin/MMER/cirino/CirTit.htm.
Cohen A, Mills C, Berry H,WonhamM, Bingham B, Bookheim B, Carlton J, ChapmanJ, Cordell J, Harris L, Klinger T, Kohn A, Lambert C, Lambert G, Li K, Secord D,Toft J. 1998. A rapid assessment survey of non-indigenous species in the shallowwaters of Puget Sound. Report of the Puget Sound Expedition September 8–16, 1998.Prepared for Washington State Department of Natural Resources, Olympia, WA andUnited States Fish and Wildlife Service, Lacey, WA. United States Fish and WildlifeService, Washington, D.C. 41 pp. Available at http:// cdm16658.contentdm.oclc.org/cdm/ref/ collection/p267501ccp2/ id/712.
Colegate SM,Molyneux RJ. 2007. Bioactive natural products: detection, isolation, andstructural determination. Boca Raton: CRC Press.
Corbo JC, Di Gregorio A, Levine M. 2001. The ascidian as a model organism in develop-mental and evolutionary biology. Cell 106:535–538DOI 10.1016/S0092-8674(01)00481-0.
Eri R, Arnold JM, Hinman VF, Green KM, Jones MK, Degnan BM, LavinMF. 1999.Hemps, a novel EGF-like protein, plays a central role in ascidian metamorphosis.Development 126:5809–5818.
Fletcher LM, Forrest BM. 2011. Induced spawning and culture techniques for theinvasive ascidian Didemnum vexillum (Kott, 2002). Aquatic Invasions 6:457–464DOI 10.3391/ai.2011.6.4.11.
Fofonoff PW, Ruiz GM, Steves B, Carlton JT. 2003. Ciona savignyi. National ExoticMarine and Estuarine Species Information System. Available at http:// invasions.si.edu/nemesis/ browseDB/SpeciesSummary.jsp?TSN=-122 (accessed 18 July 2014).
Gerhart D, Rittschof D, Hooper I, Eisenman K, Meyer A, Baier R, Young C.1992. Rapid and inexpensive quantification of the combined polar compo-nents of surface wettability: application to biofouling. Biofouling 5:251–259DOI 10.1080/08927019209378246.
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 14/18

Green K, Russell B, Clark R, Jones M, GarsonM, Skilleter G, Degnan B. 2002. A spongeallelochemical induces ascidian settlement but inhibits metamorphosis.MarineBiology 140:355–363 DOI 10.1007/s002270100698.
Hadfield MG, Paul VJ. 2001.Natural chemical cues for settlement and metamorphosis ofmarine invertebrate larvae. Boca Raton: CRC Press, 431–461.
Head R, Overbeke K, Klijnstra J, Biersteker R, Thomason J. 2003. The effect of gregari-ousness in cyprid settlement assays. Biofouling 19:269–278DOI 10.1080/0892701031000101502.
Hendrickson C, Christiaen L, Deschet K, Jiang D, Joly J-S, Legendre L, Nakatani Y,Tresser J, SmithWC. 2004. Culture of adult ascidians and ascidian genetics.Methodsin Cell Biology 74:143–170 DOI 10.1016/S0091-679X(04)74007-8.
Imai KS, Satoh N, Satou Y. 2002a. Early embryonic expression of FGF4/6/9 gene andits role in the induction of mesenchyme and notochord in Ciona savignyi embryos.Development 129:1729–1738.
Imai KS, Satou Y, Satoh N. 2002b.Multiple functions of a Zic-like gene in the differen-tiation of notochord, central nervous system and muscle in Ciona savignyi embryos.Development 129:2723–2732.
Jackson D, Leys SP, Hinman VF,Woods R, LavinMF, Degnan BM. 2002. Ecologicalregulation of development: induction of marine invertebrate metamorphosis.International Journal of Developmental Biology 46:679–686.
JaeckleW. 1994. Rates of energy consumption and acquisition by lecithotrophiclarvae of Bugula neritina (Bryozoa: Cheilostomata).Marine Biology 119:517–523DOI 10.1007/BF00354313.
JaeckleWB, Manahan DT. 1989. Growth and energy imbalance during the developmentof a lecithotrophic molluscan larva (Haliotis rufescens). The Biological Bulletin177:237–246 DOI 10.2307/1541939.
Jiang D, SmithW. 2005. Self-and cross-fertilization in the solitary ascidian Cionasavignyi. The Biological Bulletin 209:107–112 DOI 10.2307/3593128.
Kimura Y, YoshidaM,MorisawaM. 2003. Interaction between noradrenaline oradrenaline and the β 1-adrenergic receptor in the nervous system triggers earlymetamorphosis of larvae in the ascidian, Ciona savignyi. Developmental Biology258:129–140 DOI 10.1016/S0012-1606(03)00118-0.
Kourakis M, Newman-Smith E, SmithW. 2010. Key steps in the morphogenesis of a cra-nial placode in an invertebrate chordate, the tunicate Ciona savignyi. DevelopmentalBiology 340:134–144 DOI 10.1016/j.ydbio.2010.01.016.
Kubo I, HimejimaM. 1991. Anethole, a synergist of polygodial against filamentousmicroorganisms. Journal of Agricultural and Food Chemistry 39:2290–2292DOI 10.1021/jf00012a040.
Kuo J, Mitchell D, Tuerke T. 1993. SigmaPlot: Scientific graph system. Transforms andcurve fitting reference. Corte Madera: Jandel Scientific.
Lambert CC, Lambert G. 1998. Non-indigenous ascidians in southern California harborsand marinas.Marine Biology 130:675–688 DOI 10.1007/s002270050289.
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 15/18

Lemaire P. 2011. Evolutionary crossroads in developmental biology: the tunicates.Development 138:2143–2152 DOI 10.1242/dev.048975.
McHenryMJ. 2005. The morphology, behavior, and biomechanics of swimming inascidian larvae. Canadian Journal of Zoology 83:62–74 DOI 10.1139/z04-157.
Moran AL, Manahan DT. 2003. Energy metabolism during larval development ofgreen and white abalone, Haliotis fulgens and H. sorenseni. The Biological Bulletin204:270–277 DOI 10.2307/1543598.
Morse DE. 1990. Recent progress in larval settlement and metamorphosis: closing thegaps between molecular biology and ecology. Bulletin of Marine Science 46:465–483.
Nomaguchi TA, Nishijima C, Minowa S, HashimotoM, Haraguchi C, Amemiya S,Fujisawa H. 1997. Embryonic thermosensitivity of the ascidian, Ciona savignyi.Zoological Science 14:511–515 DOI 10.2108/zsj.14.511.
Pawlik JR. 1992. Chemical ecology of the settlement of benthic marine invertebrates.Oceanography and Marine Biology: An Annual Review 30:273–335.
Pawlik JR. 1993.Marine invertebrate chemical defenses. Chemical Reviews 93:1911–1922DOI 10.1021/cr00021a012.
Piazza V, Ferioli A, Giacco E, Melchiorre N, Valenti A, Del Prete F, Biandolino F, Den-tone L, Frisenda P, Faimali M. 2012. A standardization of Amphibalanus (Balanus)amphitrite (Crustacea, Cirripedia) larval bioassay for ecotoxicological studies. Eco-toxicology and Environmental Safety 79:134–138 DOI 10.1016/j.ecoenv.2011.12.014.
Pinheiro J, Bates D. 2006.Mixed-effects models in S and S-PLUS NJ . New York: SpringerScience & Business Media.
RDevelopment Core Team. 2015. R: a language and environment for statistical comput-ing . Vienna: R Foundation for Statistical Computing. Available at http://www.R-project.org/ .
Rae Flores A, Faulkes Z. 2008. Texture preferences of ascidian tadpole larvae dur-ing settlement.Marine and Freshwater Behaviour and Physiology 41:155–159DOI 10.1080/10236240802360914.
Rodriguez SR, Ojeda FP, Inestrosa NC. 1993. Settlement of benthic marine inverte-brates.Marine Ecology Progress Series 97:193–207 DOI 10.3354/meps097193.
Ross K, Bidwell J. 2001. A 48-h larval development toxicity test using the marine poly-chaete Galeolaria caespitosa Lamarck (Fam. Serpulidae). Archives of EnvironmentalContamination and Toxicology 40:489–496 DOI 10.1007/s002440010201.
Sasakura Y, Mita K, Ogura Y, Horie T. 2012. Ascidians as excellent chordate mod-els for studying the development of the nervous system during embryogene-sis and metamorphosis. Development, Growth & Differentiation 54:420–437DOI 10.1111/j.1440-169X.2012.01343.x.
Satoh N. 2003. The ascidian tadpole larva: comparative molecular development andgenomics. Nature Reviews Genetics 4:285–295.
Satoh N, Levine M. 2005. Surfing with the tunicates into the post-genome era. Genes &Development 19:2407–2411 DOI 10.1101/gad.1365805.
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 16/18

Schubert M, Escriva H, Xavier-Neto J, Laudet V. 2006. Amphioxus and tunicatesas evolutionary model systems. Trends in Ecology & Evolution 21:269–277DOI 10.1016/j.tree.2006.01.009.
Selwood AI, Wilkins AL, Munday R, Shi F, Rhodes LL, Holland PT. 2013. Portimine:a bioactive metabolite from the benthic dinoflagellate Vulcanodinium rugosum.Tetrahedron Letters 54:4705–4707 DOI 10.1016/j.tetlet.2013.06.098.
Sherrard R, Murray-Gulde C, Rodgers Jr J, Shah Y. 2002. Comparative toxicity ofchlorothalonil and chlorpyrifos: Ceriodaphnia dubia and Pimephales promelas.Environmental Toxicology 17:503–512 DOI 10.1002/tox.10091.
Smith KF, Cahill PL, Fidler AE. 2010. First record of the solitary ascidian Ciona sav-ignyiHerdman, 1882 in the Southern Hemisphere. Aquatic Invasions 5:363–368DOI 10.3391/ai.2010.5.4.05.
Stronkhorst J, Ciarelli S, Schipper C, Postma J, DubbeldamM, VangheluweM, BrilsJ, Hooftman R. 2004. Inter-laboratory comparison of five marine bioassays forevaluating the toxicity of dredged material. Aquatic Ecosystem Health & Management7:147–159 DOI 10.1080/14634980490281579.
Systat Software Inc. 2015. SigmaPlot . San Jose: Systat Software Inc.Tait K, Joint I, DaykinM,Milton DL,Williams P, CamaraM. 2005. Disrup-
tion of quorum sensing in seawater abolishes attraction of zoospores of thegreen alga Ulva to bacterial biofilms. Environmental Microbiology 7:229–240DOI 10.1111/j.1462-2920.2004.00706.x.
Toonen RJ, Pawlik J. 1996. Settlement of the tube worm Hydroides dianthus (Poly-chaeta: Serpulidae): cues for gregarious settlement.Marine Biology 126:725–733DOI 10.1007/BF00351339.
Tsukamoto S, Kato H, Hirota H, Fusetani N. 1996.Mauritiamine, a new antifoulingoroidin dimer from the marine sponge Agelas mauritiana. Journal of NaturalProducts 59:501–503 DOI 10.1021/np960113p.
United States Pharmacopeial Convention. 2012. Biological assay validation. In:United States pharmocopeia. USP <1033>. Rockville: United States PharmacopeialConvention, p. 25.
Van den Brink PJ, Ter Braak CJ. 1998.Multivariate analysis of stress in experimentalecosystems by principal response curves and similarity analysis. Aquatic Ecology32:163–178 DOI 10.1023/A:1009944004756.
Van den Brink PJ, Ter Braak CJ. 1999. Principal response curves: Analysis of time-dependent multivariate responses of biological community to stress. EnvironmentalToxicology and Chemistry 18:138–148 DOI 10.1002/etc.5620180207.
Voulvoulis N, ScrimshawM, Lester J. 1999. Alternative antifouling biocides. AppliedOrganometallic Chemistry 13:135–143DOI 10.1002/(SICI)1099-0739(199903)13:3<135::AID-AOC831>3.0.CO;2-G.
WahlM. 1989.Marine epibiosis. I. Fouling and antifouling: some basic aspects.MarineEcology Progress Series 58:175–189 DOI 10.3354/meps058175.
Wendt I, Arrhenius Å, Backhaus T, Hilvarsson A, Holm K, Langford K, Tunovic T,Blanck H. 2013. Effects of five antifouling biocides on settlement and growth of
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 17/18

zoospores from the marine macroalga Ulva lactuca L. Bulletin of EnvironmentalContamination and Toxicology 91:426–432 DOI 10.1007/s00128-013-1057-9.
Zvyagintsev AY, Sanamyan K, Kashenko S. 2007. On the introduction of the ascidianCiona savignyiHerdman, 1882 into Peter the Great Bay, Sea of Japan. Russian Journalof Marine Biology 33:133–136 DOI 10.1134/S1063074007020083.
Cahill et al. (2016), PeerJ, DOI 10.7717/peerj.1739 18/18