amphibian community response to variation in habitat structure and competitor density
TRANSCRIPT

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofitpublishers, academic institutions, research libraries, and research funders in the common goal of maximizing access tocritical research.
Amphibian Community Response to Variation in HabitatStructure and Competitor DensityAuthor(s): Jennifer L. Purrenhage and Michelle D. BooneSource: Herpetologica, 65(1):14-30. 2009.Published By: The Herpetologists' LeagueDOI: http://dx.doi.org/10.1655/08-017R1.1URL: http://www.bioone.org/doi/full/10.1655/08-017R1.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated contentindicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercialuse. Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder.

AMPHIBIAN COMMUNITY RESPONSE TO VARIATION IN HABITATSTRUCTURE AND COMPETITOR DENSITY
JENNIFER L. PURRENHAGE1,2
AND MICHELLE D. BOONE1
1Department of Zoology, Miami University, Oxford, OH 45056, USA
ABSTRACT: Habitat destruction and alteration is a major cause of population declines; thus, habitatrestoration is essential for preventing species extinctions. Habitat restoration, however, may be hindered byan inadequate understanding of species’ habitat requirements. Habitat structure may play a major role ininfluencing the biotic interactions among species. Pond-breeding amphibians are one group that hasexperienced loss and alteration of critical habitats. Vegetation structure in wetland environments may beimportant to amphibian larvae if its presence mediates biotic interactions, such as competition, which canaffect growth and survival to metamorphosis. Despite the potential importance of habitat structure andcompetition, few studies have examined these factors simultaneously. In a full-factorial pond mesocosmexperiment, we tested for the effects of variation in habitat structure and density of a larger tadpolecompetitor (Rana pipiens, northern leopard frog) on development and survival of three species of larvalamphibians: Bufo americanus (American toad), Hyla versicolor (gray treefrog), and Ambystoma maculatum(spotted salamander). Survival of Bufo was greater in mesocosms with live or artificial cattails than in thoselacking vegetation. Although responses varied among species, our data suggest that time to metamorphosis,survival, and species evenness can be influenced by both aquatic habitat structure and competition, whereassize at metamorphosis is affected primarily by competition. Our study is the first to demonstrate thatvegetation structure alone can affect the expression of metamorphic traits for some anuran species.
Key words: Ambystoma maculatum; Bufo americanus; Habitat complexity; Hyla versicolor; Rana pipiens
AMPHIBIANS are currently experiencingworldwide population declines due, in part,to habitat loss and alteration (Alford andRichards, 1999; Collins and Storfer, 2003;Dodd and Smith, 2003; Kiesecker et al., 2001;Semlitsch, 2003; Stuart et al., 2004). Wetlandhabitats have been especially vulnerable toloss and alteration through human-mediatedchanges such as physical disturbance, alter-ation of hydroperiod, and biological invasions(Mitsch and Gosselink, 2007; Whigham, 1999;Zedler and Kercher, 2004). Such habitatmodifications can result in altered wetlandplant communities, which can in turn affectthe chemical, biological, and structural qualityof the aquatic environment. For instance,secondary compounds leached from invasiveplants (e.g., purple loosestrife, Lythrum sali-caria) have been shown to slow larvalamphibian development (Maerz et al., 2005).Biological attributes of the environment, suchas algal community composition, may also varyamong different plant communities and canaffect size of metamorphs and length of thelarval period in some amphibians that dependon periphyton as a primary food resource
(Brown et al., 2006; Skelly et al., 2002).Finally, habitat structure varies among andwithin wetland sites (Colburn, 2004), anddifferences in habitat structure may alterbehavior and the outcome of biotic interac-tions. Each of these changes can affectamphibian population dynamics and trans-form local community structure, potentiallyinfluencing species diversity at the landscapelevel (Gray et al., 2004; Knutson et al., 1999;Schiesari, 2006; Semlitsch, 2003; Skelly et al.,1999; Van Buskirk, 2005; Wellborn et al.,1996).
Although loss of suitable wetland habitatpresents a general threat to amphibians(Dodd and Smith, 2003), many species areknown to readily use newly constructedwetlands (Lehtinen and Galatowitsch, 2001;Pechmann et al., 2001; Petranka et al., 2003),which suggests that habitat restoration orconstruction of new wetlands could help offsetpast or present effects of habitat destruction.However, effective restoration, construction,and protection of wetland habitats require anunderstanding of which habitat features moststrongly influence wetland fauna. Pond hydro-period is known to be a strong predictor of theaquatic community that assembles at a site2 CORRESPONDENCE: e-mail, [email protected]
Herpetologica, 65(1), 2009, 14–30
E 2009 by The Herpetologists’ League, Inc.
14

(Wellborn et al., 1996), and changes in pondcanopy cover through time have been corre-lated with changes in amphibian communitystructure (Skelly et al., 1999). However, theeffects of other physical attributes of ponds,including vegetation structure, are unclearand could play an important role in determin-ing community dynamics. Habitat structurehas emerged as a key determinant of habitatsuitability for many taxa, including fishes(Diehl and Eklov, 1995; Eklov, 1997; Gre-nouillet et al., 2002; Persson and Eklov, 1995;Werner et al., 1983), invertebrates (Langel-lotto and Denno, 2004), reptiles (e.g., Pounds,1988), mammals (e.g., Wilder and Meikle,2005), and birds (e.g., Whittingham andEvans, 2004). The effects of habitat structureon amphibians have been largely overlooked(McDiarmid and Altig, 1999), but the limitedavailable data suggest that habitat variationwithin ponds can have important consequenc-es for growth and survival of larval amphibians(Alford, 1986; Babbitt and Jordan, 1996;Babbitt and Tanner, 1997, 1998; Baber andBabbitt, 2004; Smith and Doupnik, 2005;Sredl and Collins, 1992; Tarr and Babbitt,2002; Warkentin, 1992).
Variability in the distribution and complex-ity of habitat structure can result in differingavailability of microhabitat and food resourc-es, which may alter competitive and trophicinteractions (Bell et al., 1991). For instance,past studies have shown that larger ranids canaffect metamorphic traits of smaller hetero-specific amphibians (Boone et al., 2004a;Kupferberg, 1997; Lawler et al., 1999); largerranids can also affect larval traits of smallerheterospecifics, consequently altering interac-tions between the smaller species (Peacor andWerner, 1997). The presence of habitatstructure may mediate these competitive andtrait-mediated indirect interactions via habitatpartitioning and food supplementation. Alter-ation of biotic interactions may be especiallyimportant for species with complex life cyclesthat are restricted to specific habitats duringdistinct life stages, such as larvae of pond-breeding amphibians that must completemetamorphosis prior to pond drying (Wilbur,1980). Information about the consequences ofvariation in aquatic habitat structure onamphibians is valuable for managers interest-
ed in taking data-driven approaches to habitatconservation and restoration. In the absenceof comprehensive and accurate data, we riskprotecting and restoring habitat that is insuf-ficient to support many pond-dependentspecies.
We conducted an outdoor, pond mesocosmexperiment to examine the effects of habitatstructure and density of a larger tadpole (Ranapipiens) in larval amphibian communitiescomposed of two anurans (Bufo americanus,Hyla versicolor) and one salamander (Ambys-toma maculatum). We tested the followinghypotheses in our experimental communities:(1) the presence of aquatic habitat structurepositively affects amphibian metamorphosisby increasing survival to and size at metamor-phosis and decreasing length of the larvalperiod; (2) the presence of physical structurealone, rather than the increased living plantmatter, positively affects amphibian metamor-phosis; (3) increasing Rana density negativelyaffects amphibian metamorphosis by decreas-ing survival to and size at metamorphosis andincreasing length of the larval period; and (4)the presence of habitat structure mediates theeffect of increased Rana density on growthand survival to metamorphosis.
MATERIALS AND METHODS
Study System
The amphibian species used in our meso-cosm experiment occur as larvae in temporaryand permanent ponds, their breeding seasonsoverlap, and in some areas they breed in thesame wetlands (Babbitt et al., 2003; Trenhamet al., 2003; J. Purrenhage, personal observa-tion). Therefore, these communities have thespatial and temporal potential to occur innatural ponds, and have been used here torepresent an assemblage of species employinga range of larval period lengths and trophicpositions. Larval densities vary widely innature, and the densities used in our exper-iment fall well within the observed range (14–4238 per 1000 l; e.g., Morin, 1983; Petranka,1989); densities at the study site are notknown. Initial stocking densities for the threefocal species (45 Bufo, 45 Hyla, and 10Ambystoma per pond mesocosm) reflect therelative availability of each species’ eggs at the
March 2009] HERPETOLOGICA 15

collection sites, and thus, approximate theirnatural larval densities.
Field Collection and Rearing
We collected eggs from Bufo americanus(American toad) and Hyla versicolor (graytreefrog) from a fishless, temporary pond atRush Run Wildlife Area (Preble County,Ohio, U.S.A.) on 17 May 2005, and Ambys-toma maculatum (spotted salamander) eggsfrom a fishless, temporary pond at IndianCreek Preserve MetroPark (Butler County,Ohio, U.S.A.) on 25 April 2005. Anuran eggswere maintained in the lab at 22–24 C untilhatching. Upon hatching, tadpoles werereared in plastic bins filled with dechlorinated(aged 48 h) tap water. Water was changeddaily and tadpoles were given food (Tetra-MinH Tropical Flakes) ad libitum. Bufo andHyla tadpoles were added to the experimentat Gosner stage 25 (Gosner, 1960). Salaman-der eggs were maintained in a cattle tank(containing water, leaf litter, and plankton)outdoors at the Miami University EcologyResearch Center (ERC), Oxford, Ohio,U.S.A., until hatching. Larvae were raisedfrom multiple egg masses (7 Bufo egg strings,15 Ambystoma egg masses, and Hyla eggs werecollected from multiple sites within a pondwhere many males were calling) and mixedprior to addition to pond mesocosms toincorporate genetic diversity from the respec-tive species’ populations. Using dipnets, wecollected Rana pipiens (northern leopard frog)larvae for Rana density treatments from RushRun Wildlife Area on 1 June 2005. Rana larvaehad recently developed hind limbs when theywere added to the experimental ponds (Gosnerstage 33–36, Gosner, 1960). Northern leopardfrogs are explosive breeders that often precedebreeding of American toads or gray treefrogs.Northern leopard frog larvae grow fast andreach a large size by the time the other anuranshatch, thereby representing a natural compet-itor of toads and treefrogs; we therefore refer attimes to northern leopard frog (Rana) larvae ascompetitors. Although Rana larvae do notcompete directly with the zooplanktivoroussalamander larvae in our experiment, theycould indirectly affect salamanders via the foodweb (i.e., changes in algal abundance can affectabundance of zooplankton food resources).
Experimental Design
A mesocosm experiment was conductedoutdoors at the Miami University ERC fromMay through July 2005. To test the effects ofhabitat structure and density of a largerheterospecific anuran (Rana pipiens), we useda full-factorial design and manipulated threelevels of habitat structure (none, live cattailstructure, and artificial cattail structure) andfour levels of Rana density (0, 6, 12, or 24Rana larvae) in cattle-tank pond mesocosms(hereafter referred to as mesocosms). Weincluded four replicates of each treatment,and treatments were randomly assigned to agrid of 48 mesocosms (1.85-m diameter, 1480-l capacity), each containing 1000 l of water,1 kg of leaf litter from a mixed deciduousforest (primarily Acer), and inoculations ofalgae and zooplankton from natural ponds.Mesocosms were covered with fiberglassmesh lids to inhibit colonization of predatorsand escape of metamorphs.
Fourteen days prior to the addition ofamphibian larvae, one of three habitat-struc-ture treatments was randomly assigned toeach mesocosm: controls (no structure) con-tained five 20-cm diameter plastic pots withsoil and pea gravel; live-cattail treatmentscontained five pots with soil, pea gravel, andthree individual Typha laxmannii (gracefulcattail) plants per pot (total of 15 plants permesocosm); and artificial-cattail treatmentscontained five pots with soil, pea gravel, andthree individual artificial plants per pot (totalof 15 plants per mesocosm). We used cattails(Typha) in our habitat structure treatmentsbecause Typha species are common in themajority of wetlands in our area, and becausethey provide a relatively simple, uniformstructure similar to that of other commonwetland species (e.g., Phragmites, Iris). Tomimic common cattails, each artificial cattailwas constructed of one 61-cm long, 2-cmdiameter PVC stem and two green, artificialleaf blades attached with two transparentplastic cable ties. Graceful cattails wereexpected to have a similar structure tocommon cattails (and our artificial cattails),but the stems were more slender and theplants were more structurally complex thancommon cattails. For all treatments, pots werearranged in a tight cluster in one quadrant of
16 HERPETOLOGICA [Vol. 65, No. 1

the mesocosm; the location of the structurequadrant was randomized such that the fourreplicates of each treatment representedstructure treatments in each of the fourcardinal directions (i.e., N, S, E, W structurequadrants).
As we mentioned previously, we manipu-lated Rana pipiens density as an experimentaltreatment in our pond mesocosms because ofthe expectation that this species may exertstrong competitive effects on anuran hetero-specifics due to its large size; in nature, theremay be priority effects because this species isgenerally the earliest breeder in the commu-nity (Alford and Wilbur, 1985). The treatmentlevels of Rana density (0, 6, 12, or 24 Ranalarvae per 1000 l) were based on expectedfield densities with high early larval mortalityand are comparable to the densities used inprevious studies of large ranid competitors(Boone et al., 2004a; Kupferberg, 1997;Lawler et al., 1999). Rana larvae wereintroduced to mesocosms in the specifiedtreatment densities on 2 June. On 3 June,each mesocosm was stocked with 100 focallarvae: 45 Hyla, 45 Bufo, and 10 Ambystoma(3 June represented day zero for all measure-ments of time to metamorphosis). Mesocosmswere checked daily for metamorphs, andanurans with at least one front limb emerged(Gosner stage 42, Gosner, 1960) or salaman-ders with resorbed gills were collected andtransported to the laboratory at Miami Uni-versity for processing. Anurans were held inthe lab until tail resorption (#3 days), atwhich point we recorded time to metamor-phosis and size at metamorphosis. All meta-morphs were released at the egg-collectionsites within 48 h of metamorphosis.
We measured response for all species assurvival to metamorphosis, mass at metamor-phosis (g), and time to metamorphosis. Timeto metamorphosis represents the number ofdays from the start of the experiment until fullresorption of the tail for anurans (Gosnerstage 46, Gosner, 1960) or the gills forsalamanders. We calculated proportion sur-vived for each species in each treatment as thenumber of individuals that successfully meta-morphosed by the end of the experiment (29July) divided by stocking density for thatspecies. We calculated mesocosm means for
mass and time to metamorphosis, as meso-cosms were the experimental units in allanalyses.
We scraped uniform samples of periphyton(3 cm below the water surface 31.7 cm widthof scraped area) from both the mesocosmedge and the vegetation structure, and col-lected zooplankton samples, from each meso-cosm three times during the experiment todetermine if habitat structure and Ranadensity treatments affected availability of foodresources for anuran and salamander larvae,respectively. Measurements of chlorophyll aconcentration (mg/l), determined via fluorom-etry of periphyton samples, were used as aproxy for relative abundance of algal resourc-es. Periphyton samples were collected fromthe mesocosms on 9 June, 22 June, and 13July. Zooplankton samples were collectedfrom the mesocosms on 8 June, 29 June, and28 July. We also measured pH, watertemperature (C), and dissolved oxygen (DO)of each mesocosm weekly to determine anytreatment effects on these water characteris-tics. Finally, we monitored whether thebehavior of amphibian larvae differed amongtreatments by recording the number andlocation of amphibian larvae visible in eachmesocosm on four separate occasions (6, 10,11, and 14 June) before larvae began tometamorphose. Bufo and Hyla larvae weregrouped in these counts because the twospecies could not be reliably distinguishedduring all observations. Number of larvae wasthen divided by the initial stocking density tocalculate the proportion of larvae visible andactive above the leaf litter in each mesocosm(only larvae that were either hidden in the leaflitter or resting on the mesocosm bottom werenot counted as active); the proportion activewas used as the response variable in statisticalanalyses.
Data Analyses
Using multivariate analyses of covariance(MANCOVA) and univariate analyses ofcovariance (ANCOVA), we tested for mainand interaction effects of habitat structure andRana density on mass and time to metamor-phosis of the focal amphibians (Bufo, Hyla,Ambystoma) and the Rana larvae used incompetitor-density treatments. We used sur-
March 2009] HERPETOLOGICA 17

vival to metamorphosis as the covariate inthese analyses so that treatment differenceswere not confounded by differential survivalamong treatments and because it explained asignificant amount of the variation in the data.We used ANOVA to test for treatment effectson survival to metamorphosis of each species.Differences in periphyton abundance, zoo-plankton abundance, and water characteristics(pH, temperature, DO) among treatmentswere analyzed separately using repeatedmeasures ANOVA. We tested for treatmenteffects on the proportion of larvae that wereactive and visible during each of the fourobservations using repeated measures AN-OVA. To identify potential relationships insurvival of focal amphibian species, we rancorrelation analysis on percent survival for allpairs of species; we were particularly interest-ed in the potential role of Ambystoma larvaeas predators of anuran larvae. The correlationswere all positive and were not significant(0.0166 , r , 0.2596, P . 0.07), indicatingthat Ambystoma was unlikely to be acting as apredator of anurans in this experiment. Weevaluated the community-level response (spe-cies evenness) to experimental treatments bycalculating Simpson’s Diversity Index for eachmesocosm based on final survival of the focalamphibian species. Simpson’s Diversity Index(D) equals 1/S (proportion of species i)2. Wetested for treatment effects on values of Dusing ANOVA. For all analyses, data weretransformed as necessary to meet the assump-tion of normality. Data expressed as propor-tions (i.e., survival, activity) were angularlytransformed; mass and time data were log-transformed. Statistical analyses were per-formed using SAS version 9.1.
RESULTS
Community Response
Species evenness in our experimental pondcommunities was significantly lower in meso-cosms with a high density of Rana pipienstadpoles (F3,36 5 5.68, P , 0.01; Fig. 1A) andin mesocosms lacking habitat structure (F2,36
5 6.44, P , 0.01; Fig. 1B). In mesocosmswith decreased evenness, the skew was due toan increase in the ratio of Hyla versicolorrelative to Bufo americanus. There was
approximately a 10% change in the relativeabundances of Hyla and Bufo between Ranacontrols (Hyla, 0.574 6 0.026; Bufo, 0.348 60.026) and high Rana-density treatments(Hyla, 0.679 6 0.019; Bufo, 0.242 6 0.021).There was a similar shift in relative abun-dances of Hyla and Bufo between mesocosmswith either live vegetation (Hyla, 0.580 60.026; Bufo, 0.353 6 0.026) or artificialvegetation (Hyla, 0.568 6 0.022; Bufo, 0.3526 0.022) and mesocosms lacking habitatstructure (Hyla, 0.664 6 0.018; Bufo, 0.2576 0.019).
Effects of Habitat Structure and CompetitorDensity on Bufo
Habitat structure significantly influencedthe multivariate response (i.e., mass and timeto metamorphosis) of Bufo (Wilks’ lambda 50.6797, F4,68 5 3.62, P , 0.01), which wasmainly attributable to effects on time tometamorphosis (Fig. 2A). Bufo in artificialstructure treatments had a longer larval period(time to metamorphosis) than individuals inlive structure and control (no structure)treatments (F2,35 5 7.60, P , 0.01; Fig. 2A),and individuals in artificial structure treat-ments tended to be smaller at metamorphosisrelative to those in live cattail and controltreatments (F2,35 5 1.53, P 5 0.23; Fig. 2A).Density of Rana competitors significantlyinfluenced the multivariate response of Bufo(Wilks’ lambda 5 0.6347, F6,68 5 2.89, P 50.01). Univariate analyses revealed that Bufoin lower competitor-density treatments weregenerally larger at metamorphosis than thosein higher competitor-density treatments (F3,35
5 4.42, P , 0.01), regardless of time tometamorphosis (F3,35 5 0.81, P 5 0.50)(Fig. 3A).
Habitat structure had a significant effect onsurvival to metamorphosis (F2,36 5 7.63, P ,0.01); Bufo survivorship was approximately20% higher in mesocosms with artificial andlive cattail structure than in controls (Fig. 4).Increased competitor density had a negativeeffect on survival to metamorphosis (F3,36 57.70, P , 0.01; Fig. 4). Moreover, there was asignificant interaction effect of habitat struc-ture and competitor density on survival tometamorphosis for Bufo (F6,36 5 2.35, P 50.05; Fig. 4). Bufo exhibited increased survival
18 HERPETOLOGICA [Vol. 65, No. 1

when either live or artificial cattail structurewas present, particularly at low Rana density,except in mesocosms with live cattails and noRana competitors (Fig. 4).
Effects of Habitat Structure and CompetitorDensity on Hyla
There was no significant multivariate re-sponse (Wilks’ lambda 5 0.9158, F4,68 5 0.76,P 5 0.55) and no significant univariate effectsof habitat structure treatments on any of themeasured response variables for Hyla (time,
F2,35 5 1.31, P 5 0.28; mass, F2,35 5 0.72, P 50.49; survival, F2,36 5 0.03, P 5 0.97)(Fig. 2B). However, density of Rana compet-itors significantly influenced the multivariateresponse (Wilks’ lambda 5 0.4345, F6,68 55.86, P , 0.001). Univariate analyses revealedthat as competitor density increased, time tometamorphosis increased (F3,35 5 11.70, P ,0.001) and mass at metamorphosis decreased(F3,35 5 7.23, P , 0.001) (Fig. 3B). There wasnot a significant effect of competitor densityon Hyla survival (F3,36 5 2.58, P 5 0.07; LS
FIG. 1.—Simpson’s Diversity Index (D) for experimental larval amphibian communities at each level of Rana pipiensdensity (1A) and at each level of habitat structure (1B). Simpson’s Diversity Index incorporates final survival of the threefocal amphibians: Ambystoma maculatum, Bufo americanus, and Hyla versicolor. Based on initial stocking densities, themaximum possible value of D is 2.41, and is represented by the thick, horizontal line. Different letters indicatesignificant differences in D between treatments based on Scheffe’s post hoc test.
March 2009] HERPETOLOGICA 19
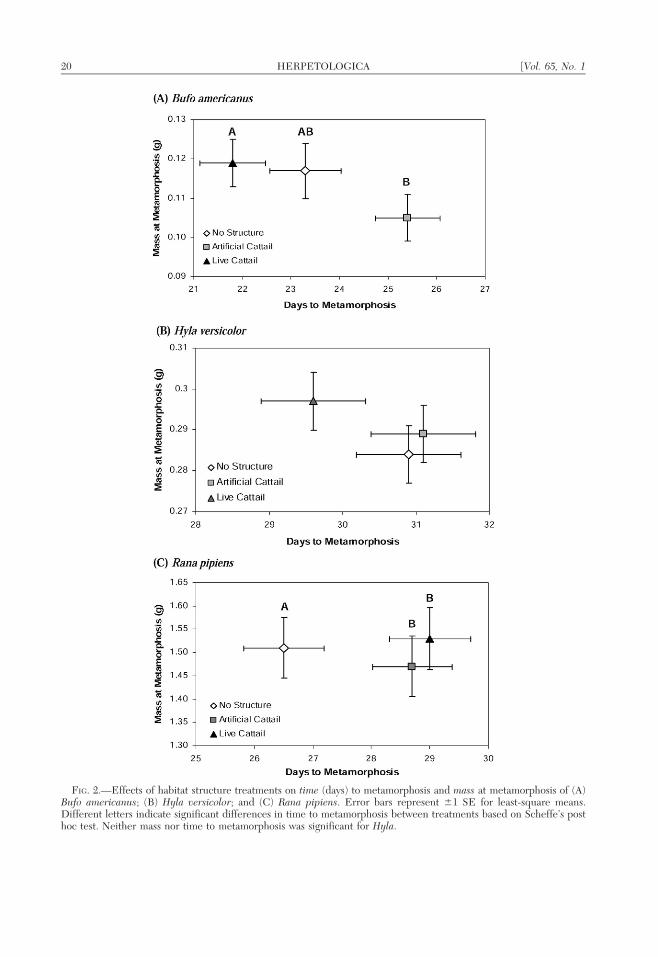
FIG. 2.—Effects of habitat structure treatments on time (days) to metamorphosis and mass at metamorphosis of (A)Bufo americanus; (B) Hyla versicolor; and (C) Rana pipiens. Error bars represent 61 SE for least-square means.Different letters indicate significant differences in time to metamorphosis between treatments based on Scheffe’s posthoc test. Neither mass nor time to metamorphosis was significant for Hyla.
20 HERPETOLOGICA [Vol. 65, No. 1
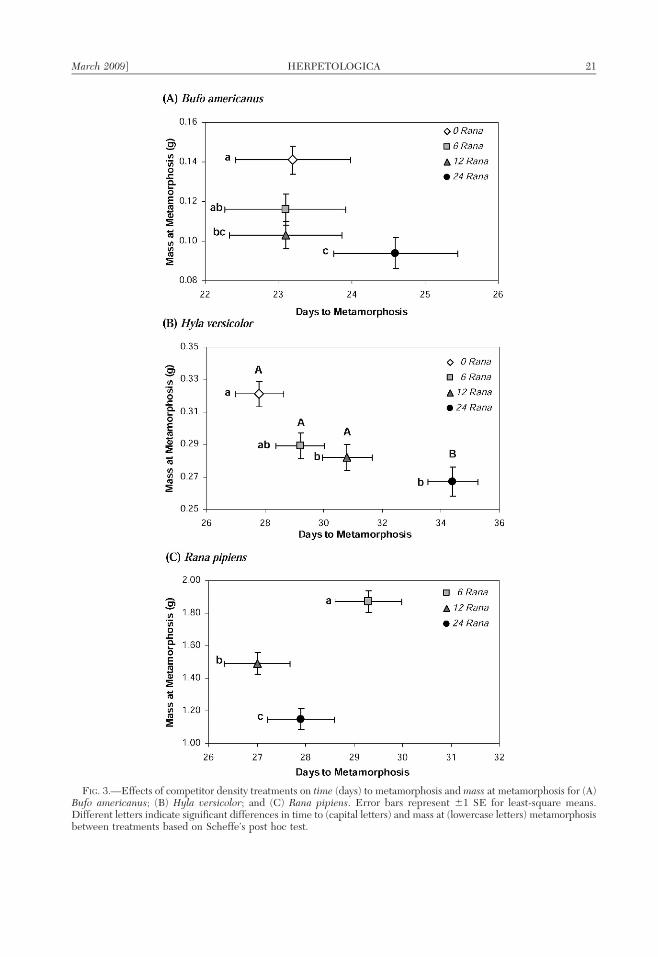
FIG. 3.—Effects of competitor density treatments on time (days) to metamorphosis and mass at metamorphosis for (A)Bufo americanus; (B) Hyla versicolor; and (C) Rana pipiens. Error bars represent 61 SE for least-square means.Different letters indicate significant differences in time to (capital letters) and mass at (lowercase letters) metamorphosisbetween treatments based on Scheffe’s post hoc test.
March 2009] HERPETOLOGICA 21

mean percent survival 6 SE: 0 Rana, 0.846 60.03; 6 Rana, 0.778 6 0.03; 12 Rana, 0.767 60.03; 24 Rana, 0.854 6 0.03). There were nosignificant treatment interaction effects on anyof the response variables measured for Hyla(Wilks’ lambda 5 0.6695, F12,68 5 1.26, P 50.26; time, F6,35 5 0.34, P 5 0.91; mass, F6,35
5 1.73, P 5 0.14; survival, F6,36 5 0.67, P 50.67).
Effects of Habitat Structure and Rana Densityon Ambystoma
Habitat structure (Wilks’ lambda 5 0.9914,F4,68 5 0.07, P 5 0.99) and Rana density(Wilks’ lambda 5 0.9235, F6,68 5 0.46, P 50.84) did not influence the multivariateresponse of Ambystoma. Univariate analysessimilarly showed no significant effects ofhabitat structure (mass: F2,35 5 0.12, P 50.89; time: F2,35 5 0.03, P 5 0.97; survival:F2,35 5 1.32, P 5 0.28) or Rana density (mass:F3,35 5 0.81, P 5 0.50; time: F3,35 5 0.09, P 50.96; survival: F3,35 5 1.06, P 5 0.38) on anyresponse variables measured.
Effects of Habitat Structure and PopulationDensity on Rana
Habitat structure did not significantlyinfluence the multivariate response of Rana(Wilks’ lambda 5 0.7520, F4,50 5 1.91, P 50.12). However, univariate analyses indicated
a significant effect of habitat structure on timeto metamorphosis in Rana (Fig. 2C). Rana inboth live and artificial structure treatmentshad a longer larval period than individuals inthe control treatment (F2,26 5 4.03, P 5 0.03;Fig. 2C), but there was no difference in massat metamorphosis (F2,26 5 0.17, P 5 0.84;Fig. 2C) or survival (F2,27 5 0.86, P 5 0.44)among treatments.
Intraspecific density significantly influ-enced the multivariate response of Rana(Wilks’ lambda 5 0.2490, F4,50 5 12.55, P ,0.001). Univariate analyses revealed that asintraspecific density increased, Rana exhibiteda dramatic decrease in mass at metamorphosis(F2,26 5 29.9, P , 0.001), and there was atrend suggesting that time to metamorphosisincreased (F2,26 5 2.77, P 5 0.08) in the low-density treatment (Fig. 3C). There was not asignificant effect of intraspecific density onRana survival (F2,27 5 0.64, P 5 0.53). Therewere no significant effects of treatmentinteractions on any of the response variablesmeasured for Rana.
Behavioral Observations
During behavioral observations, the pro-portion of combined Bufo and Hyla larvae thatwere visible and active (above leaf-littersubstrate) varied among sampling dates(F3,108 5 70.0, P , 0.001). Where significantly
FIG. 4.—Significant interaction of habitat structure and competitor density treatments for percent survival of Bufoamericanus. Error bars represent 61 SE for least-square means.
22 HERPETOLOGICA [Vol. 65, No. 1

different, the proportion of active Bufo andHyla larvae was greater in low (0 or 6 Rana/mesocosm) relative to high (12 or 24 Rana/mesocosm) competitor-density treatments.These differences were significant during thefirst (F3,36 5 3.73, P 5 0.02; LS means 6 SE:0 Rana, 47.7 6 2.48; 6 Rana, 43.1 6 2.48; 12Rana, 36.0 6 2.48; 24 Rana, 37.6 6 2.48individuals/mesocosm) and last (F3,36 5 3.49,P 5 0.03; LS means 6 SE: 0 Rana, 22.7 61.73; 6 Rana, 26.2 6 1.73; 12 Rana, 22.7 61.73; 24 Rana, 17.6 6 1.73 individuals/mesocosm) of the four behavioral observa-tions. The proportion of Rana visible andactive also varied across sampling dates (F3,81
5 7.64, P , 0.001); however, there were nosignificant treatment effects on activity ofRana larvae.
Periphyton and Zooplankton Resources
Repeated-measures ANOVA revealed thatthe abundance of periphyton from the meso-cosm edge varied greatly through time (F2,72
5 34.77, P , 0.001). Periphyton abundancedecreased initially during the peak growthperiod for anuran larvae (between the firsttwo sampling dates, on days 6 and 19 of theexperiment) and then increased dramaticallyonce most of the anuran larvae had metamor-phosed (between the second and final sam-pling dates, days 19 and 40), except in the highRana density (24 Rana/mesocosm) treatmentwhere algal abundance remained suppressed(Fig. 5A). There were significant time 3 Ranadensity (Fig. 5A; F6,72 5 3.91, P , 0.01) andtime 3 habitat structure (F4,72 5 2.58, P 50.04) interaction effects on periphyton abun-dance on the mesocosm edge. Neither inter-action revealed a clear pattern.
Repeated-measures ANOVA also revealedthat the abundance of periphyton growing onthe vegetation differed between the live andartificial cattail structure treatments (F1,24 57.28, P 5 0.01; Fig. 5B). Although we wereunable to quantify the total additional periph-yton available in vegetation structure treat-ments, the chlorophyll data indicate that thepresence of structure augmented total periph-yton; this effect was approximately 0.89 mg/l,2.34 mg/l, and 12.78 mg/l greater per unit areain artificial than in live cattail treatments ateach of the three sampling dates (days 12, 26,
and 56), respectively (Fig. 5B). Overall, abun-dance of periphyton on vegetation structureincreased over time (F2,48 5 14.68, P , 0.001;Fig. 5B), and there was a trend of decreasedabundance of periphyton on vegetation struc-ture with increased Rana density (F3,24 52.47, P 5 0.09).
Repeated measures ANOVA revealed asignificant time effect on total zooplanktonabundance (F2,72 5 44.48, P , 0.001), whichrepresented a decrease in abundancethroughout the experiment. The decrease inzooplankton abundance was observed acrossvegetation structure and Rana density treat-ments.
Water Characteristics
The pH of the water in pond mesocosmsgenerally declined over time (F7,252 5 92.33, P, 0.001), but the difference between meso-cosms was never more than 0.5 units. Simi-larly, pH was generally lower in live cattailstructure treatments (F2,36 5 6.36, P , 0.01),but the difference between treatments wasnever more than 0.1 units. Water temperaturefluctuated among sampling dates, rangingfrom 26.8 to 30.8 C (F7,252 5 4187.2, P ,0.001). Repeated-measures ANOVA revealeda significant interaction between Rana densityand habitat structure on water temperature(F6,36 5 2.38, P 5 0.05). The interaction effectseems to reflect the pattern that, in theabsence of Rana (0 Rana treatments), tem-perature was greater in habitat structurecontrols (mesocosms lacking vegetation) thanin live or artificial structure treatments;whereas, in the presence of Rana (6, 12, and24 Rana treatments), temperature did notappear to differ with varying habitat structure.However, temperature differences amongtreatments were generally small (,0.5 C).Dissolved oxygen (DO) concentration fluctu-ated through time (F7,252 5 20.67, P , 0.001).There was also a significant interaction ofRana density and time on DO (F21,252 5 3.50,P , 0.001), which seems to reflect generallyhigher DO in Rana controls (0 Rana/meso-cosm) relative to moderate and high (12 or 24Rana/mesocosm) Rana-density treatments.Actual significant differences in DO concen-tration between Rana density treatmentsranged from a 0.30-mg/l to a 2.08-mg/l
March 2009] HERPETOLOGICA 23

difference. We observed significant differenc-es in DO between habitat-structure treat-ments on several mid-season sampling dates:30 June (day 27; F2,36 5 3.20, P 5 0.05); 6 July(day 33; F2,36 5 4.09, P 5 0.03); and 13 July(day 40; F2,36 5 5.93, P , 0.01). On all threedates, DO was lower in mesocosms with livecattails than in mesocosms with no vegetationstructure or artificial cattails; however, the
greatest difference in average DO betweenstructure treatments was 0.5 mg/l.
DISCUSSION
Our study is the first to demonstrate thatthe presence of vegetation structure alone canaffect the expression of metamorphic traits forsome anuran species. The presence of artifi-
FIG. 5.—Relative abundance of periphyton (represented by chlorophyll a concentration) in samples taken from (A)the pond edge in ponds with different Rana-competitor densities and (B) the vegetation structure in ponds with artificialand live cattail structure. Error bars represent 61 SE for least-square means.
24 HERPETOLOGICA [Vol. 65, No. 1

cial or live structure increased Bufo survivor-ship in mesocosms where the density of Ranacompetitors was low. That this effect was notobserved for other amphibian species may bea reflection of the different life-history strat-egies employed by the species in our exper-imental community. Werner (1986) suggestedthat amphibian life histories differ based ontwo main factors: the opportunity for growthand the risk of mortality in the aquatic andterrestrial phases. In general, bufonids haveshorter larval periods and metamorphosesmaller relative to their adult size than hylidsand ranids; therefore, bufonids may be moresensitive than species with longer larvalperiods to changes in food resources. Theaddition of vegetation structure appears to beassociated with increased periphyton foodresources, and toads may experience suchbenefits of structure when the competitiveenvironment is more favorable (i.e., at lowcompetitor density). It is interesting that weobserved a significant effect of vegetationstructure on survival in Bufo americanus, thespecies that spends the least amount of time inthe pond environment. However, the shortlarval period of Bufo also reflects the stronginfluence that the aquatic environment canhave on this species in a short timeframe. Aclosely related species, Bufo woodhousii, hasbeen shown to respond to environmentalchanges even when other species in thecommunity showed little to no response(e.g., insecticide exposure: Boone et al.,2004b). Similarly, we found in this study thatBufo americanus responded to variation inhabitat structure despite brief exposure dur-ing their relatively short larval stage.
In many studies, the advantages of habitatstructure are attributed to a mediation ofpredator-prey dynamics. Prey detection byforaging predators (e.g., many fish predators)is often compromised in structurally complexenvironments, which confers clear benefits tosurvivorship and condition of prey (Babbittand Tanner, 1997, 1998; Crowder and Coo-per, 1982). In this experiment, however, weobserved a positive effect of habitat structureon larval survival and community evenness inthe absence of predators, which suggests thatthe advantages of habitat structure are notlimited to the availability of refugia for
predator evasion. Other possible explanationsof the benefits of habitat structure includeincreased foraging behavior due to the per-ception of safety in the presence of vegetationstructure and increased food resources pro-vided by periphyton growing on vegetation.Although we cannot empirically evaluatedifferences in the perception of predation riskamong habitat structure treatments based onour dataset, others have documented thisbehavior in a range of terrestrial, freshwater,and marine systems (Horat and Semlitsch,1994; Lima, 1990; Nicieza, 2002; Sih, 1986;Werner and Hall, 1988; Wirsing et al., 2007).We did not directly observe differences inbehavior among habitat-structure treatments;however, increased foraging due to perceptionof safety cannot be ruled out as a possiblemechanism for the observed differences inBufo survivorship. Alternatively, benefits toBufo larvae could be explained by an allevi-ation of exploitative competition with Hylaand Rana due to supplemental food resourcesprovided by periphyton growing on vegetationstructure. For instance, the community shifttoward Hyla dominance in mesocosms lackingvegetation structure could be due to Hyla’ssuperior exploitation of shared resources instructurally simple habitats. Chlorophyll anal-ysis of periphyton samples collected from thevegetation in our mesocosms confirmed addi-tional periphyton growth in mesocosms withboth live and artificial cattails, althoughperiphyton was more abundant on artificialcattails. Thus, the availability of supplementalperiphyton growing on vegetation may con-tribute to increased survival in some cases. Athird possible explanation is that the presenceof habitat structure may reduce interferencecompetition between species. Behavioral ob-servations of Bufo americanus and Hylaversicolor larvae did not reveal changes inactivity or avoidance behavior when they wereobserved in the presence of Rana pipienslarvae, which suggests that interference com-petition is not important between thesespecies and further supports the formerhypotheses about the mechanisms of habitatstructure effects.
Our results also demonstrate that habitatstructure can affect the length of the larvalperiod for some anurans. Larvae of pond-
March 2009] HERPETOLOGICA 25

breeding amphibians have been shown tomake trade-offs between the length of thelarval period and body size at the time ofmetamorphosis in response to changes inenvironmental attributes (e.g., Boone et al.,2004b; Crump, 1989; Laurila and Kujasalo,1999). Different species employ different lifehistory strategies; Bufo generally represent thestrategy of metamorphosing earlier and small-er relative to their adult size, Rana andAmbystoma remain in the aquatic environ-ment much longer and metamorphose at alarger size relative to their adult size, and Hylatend to fall somewhere in the middle of thiscontinuum. Shortening the larval periodreduces exposure to aquatic predators andminimizes the risk of desiccation, both ofwhich increase the likelihood of surviving tometamorphosis. However, larvae that leavethe pond environment early often do so at asmaller size, and size at metamorphosis isconsidered an indicator of future fitness formany species (e.g., Semlitsch et al., 1988).Thus, factors that influence plasticity in thesetraits can have potentially serious effects onpopulations.
We had expected to see larger metamorphsize and a shorter larval period in mesocosmswith habitat structure because of the supple-mental periphyton resource available on thevegetation. In contrast, we observed anincrease in the length of the larval period ofBufo in mesocosms with artificial cattails andof Rana in mesocosms with artificial or livecattails. For both species, the larval periodsbetween mesocosms with and without habitatstructure differed by approximately threedays. It is interesting that variation in habitatstructure can significantly influence the tim-ing of metamorphosis, but it remains unclearwhy this difference occurred and whetherthere are biologically significant consequencesof a three-day difference in larval period. If,for instance, the presence of habitat structureinduced a trade-off between time spent in thelarval stage and size at metamorphosis, wewould have expected that metamorphs emerg-ing later would be larger in size; however, ourdata do not support such a trade-off.
The observed difference in Bufo responseto artificial versus live cattails could beattributed to either biological or structural
differences between the two vegetation treat-ments. A biological explanation could be thatartificial and live cattails differ in theircapacities to support growth of microorgan-isms and algae that benefit amphibian larvae.Live cattails appeared to be associated with abiofilm at the water surface and larvae wereobserved feeding in these areas (J. Purrenh-age, personal observations). If live cattails didsupport microorganisms or algae that en-hanced the food resources for Bufo, this mayhelp to explain the apparent size and devel-opmental advantage conferred to larvae inmesocosms with live cattail structure. Alter-natively, the relevant difference between liveand artificial cattails could have been struc-tural. The live cattails in our mesocosms grewthroughout the experiment and provided amore structurally complex habitat than did theartificial cattails, which more closely approx-imated the structure of most Typha speciescommonly found in Midwestern ponds. Con-sequently, Bufo may have responded to levelsof structural complexity rather than to somebiological factor associated with live, but notartificial, cattails. However, we cannot defin-itively say whether one or both of thesemechanisms drove the observed effect.
As predicted, and consistent with previousfindings (Werner, 1986; Wilbur, 1977; Wilburand Collins, 1973), the anurans in ourexperiment exhibited smaller sizes at meta-morphosis in high-competition environments(higher Rana-density treatments), althoughtrade-offs between size at metamorphosisand length of the larval period differed amongspecies. On average, Bufo metamorphosed ata smaller size with increasing Rana density,but showed no significant alteration in thelength of the larval period. This pattern isconsistent with a strategy that minimizes thetime spent in the aquatic, larval stage,regardless of the resultant size at metamor-phosis (Werner, 1986), and provides furtherevidence for metamorphic synchrony in Bufo(Breden and Kelly, 1982; DeVito, 2003), evenin the absence of predation or changing waterlevels. In contrast, Hyla exhibited both agraded decrease in size at metamorphosis anda graded increase in the length of the larvalperiod with increasing Rana density. Thispattern suggests that increased competition
26 HERPETOLOGICA [Vol. 65, No. 1

with Rana inhibits growth of Hyla larvae, suchthat they must delay metamorphosis until theyreach a minimum size required by this speciesfor metamorphosis. Competition among larvalamphibians may occur via both interference andexploitation. However, as has been seen withother ranids (Boone et al., 2008; Kupferberg,1997), the observed suppression of periphytonabundance in mesocosms with high Ranadensity suggests that the competitive effect ofthe larger Rana pipiens larvae on smallerheterospecifics may be exerted mainly viaexploitation of the shared algal food resource.As previously mentioned, behavioral observa-tions suggest that interference by Rana is notthe mechanism of competition between thesespecies, which further supports the alternativehypothesis of exploitative competition.
The nature of the response by Rana tohabitat structure and larval density differedfrom the responses of Bufo and Hyla.Whereas Bufo metamorphosed earlier inmesocosms with live cattails than in thosewith artificial cattails, Rana larvae metamor-phosed earlier in mesocosms lacking vegeta-tion than in those with either artificial or livecattails. This difference suggests that thebiological or structural aspect of live cattailstructure that benefited Bufo does not confera similar advantage to Rana. If Bufo and Ranaare competing for the same food resource,then we might expect to see this differentialresponse since a condition that favors onespecies should have the opposite effect on itscompetitor. Rana also differed in their re-sponse to population density. Whereas Hylaexhibited both a smaller size at metamorpho-sis and a longer larval period with increaseddensity of Rana competitors, Rana exhibited adifferent pattern in these traits. Rana diddemonstrate a strong density-dependent re-sponse of greatly reduced size at metamor-phosis in moderate- and high-density treat-ments, but the density of Rana competitorsdid not significantly influence the larvalperiods of either Rana or Bufo. However,unlike Bufo, Rana did exhibit a trend of alonger larval period in low-density treatments,which may indicate a trade-off between size atmetamorphosis and time spent in the larvalstage. In accordance with the general life-history strategy of ranids, Rana larvae in our
experiment may have weighed the cost of along larval period against the benefit of a largesize at metamorphosis (Werner, 1986; Wilburand Collins, 1973). Alternatively, the differentresponse by Rana could be a consequence ofthe environmental conditions they were ex-posed to during the early larval stage in thenatural pond from which they were collected.However, even if exposure to unknown factorsin the natural environment did influence Ranaresponse, they nevertheless showed responseto experimental treatments after shorter-termexposure than the other species, relative totheir own developmental period.
In summary, variation in habitat structureand density of Rana larvae (competitordensity) significantly influenced length of thelarval period, mass at metamorphosis, andsurvival to metamorphosis of larval anurans, aswell as species evenness of the experimentalcommunities. Our hypothesis that habitatstructure increases survival to and mass atmetamorphosis and decreases length of thelarval period was partially supported for Bufo,which exhibited increased survival in thepresence of vegetation, but not for the otherspecies in our experimental community. Ourhypothesis that density of Rana pipiens, aheterospecific competitor, decreases survivalto and mass at metamorphosis and increaseslength of the larval period was supported forBufo and Hyla. Finally, our hypothesis thathabitat structure mediates the negative effectof competitor density was partially supportedfor Bufo, which exhibited increased survival atlow densities of the Rana competitor andshowed a similar trend in high Rana-densitytreatments. Habitat structure has been shownto affect the outcome of biotic interactions fora range of taxa, but has been largely over-looked in studies of larval amphibians (Bell etal., 1991; McDiarmid and Altig, 1999). Ourstudy is the first to show that for larvalamphibians in structurally complex habitatsthere may be delayed time to metamorphosisand increased probability of survival, even inthe absence of predation. Moreover, thepresence of habitat structure increased spe-cies evenness and may increase the likelihoodfor diverse communities over time.
Understanding the role of habitat structurefor pond-breeding amphibians is critical for
March 2009] HERPETOLOGICA 27

successful restoration and management prac-tices. We have demonstrated that aquaticvegetation has the potential to affect survivaland length of the larval period for someanuran larvae, but that species may differ intheir habitat needs and that habitat structuremay be important for mediating differentbiotic interactions (e.g., predation) and struc-turing communities (e.g., species evenness).While it may be premature to label vegetationstructure, with hydroperiod and predatorcommunity composition (Wellborn et al.,1996), as an important factor affecting thestructure and dynamics of larval amphibiancommunities, aquatic vegetation has beenshown to be important for courtship andoviposition in pond-breeding amphibians(Egan and Paton, 2004; Howard, 1978). Thus,we should continue to explore the role ofvegetation structure for amphibians in multi-ple life stages (e.g., metamorphs, breedingadults). Moreover, our finding that habitatstructure can enhance survival to metamor-phosis of anurans even in the absence ofpredators, justifies further examination of theeffects of habitat structure on amphibians inthe aquatic stage.
Acknowledgments.—This research was made possible bythe help of many people in the field and lab, especially G.Roman, H. Puglis, S. Provenzano, S. Wilder, J. Stoeckel, S.Reed, and the staff of the Miami University EcologyResearch Center, especially R. Kolb. We thank the BooneLab and the Ecology and Evolution Graduate Group atMiami University for suggestions and improvements toearlier versions of this manuscript. This research was fundedby Miami University Department of Zoology, an REU grantfrom the National Science Foundation (DBI-0353915), andan IUCN/SSC Amphibian Specialist Group Seed Grantawarded to J. Purrenhage. The experiments described herecomply with current U.S. laws and were approved underanimal care protocol #652 by the Institutional Animal Careand Use Committee at Miami University.
LITERATURE CITED
ALFORD, R. A. 1986. Habitat use and positional behavior ofanuran larvae in a Northern Florida temporary pond.Copeia 1986:408–423.
ALFORD, R. A., AND H. M. WILBUR. 1985. Priority effects inexperimental pond communities: competition betweenBufo and Rana. Ecology 66:1097–1105.
ALFORD, R. A., AND S. J. RICHARDS. 1999. Globalamphibian declines: a problem in applied ecology.Annual Review of Ecology and Systematics 30:133–165.
BABBITT, K. J., AND F. JORDAN. 1996. Predation on Bufoterrestris tadpoles: effects of cover and predatoridentity. Copeia 1996:485–488.
BABBITT, K. J., AND G. W. TANNER. 1997. Effects of coverand predator identity on predation of Hyla squirellatadpoles. Journal of Herpetology 31:128–130.
BABBITT, K. J., AND G. W. TANNER. 1998. Effects of coverand predator size on survival and development of Ranautricularia tadpoles. Oecologia 114:258–262.
BABBITT, K. J., M. J. BABER, AND T. L. TARR. 2003. Patternsof larval amphibian distribution along a wetlandhydroperiod gradient. Canadian Journal of Zoology81:1539–1552.
BABER, M. J., AND K. J. BABBITT. 2004. Influence of habitatcomplexity on predator-prey interactions between thefish (Gambusia holbrooki) and tadpoles of Hylasquirella and Gastrophryne carolinensis. Copeia2004:173–177.
BELL, S. S., E. D. MCCOY, AND H. R. MUSHINSKY (Eds.).1991. Habitat Structure: The Physical Arrangement ofObjects in Space. Chapman and Hall, London,England.
BOONE, M. D., E. E. LITTLE, AND R. D. SEMLITSCH. 2004a.Overwintered bullfrog tadpoles negatively affect sala-manders and anurans in native amphibian communities.Copeia 2004:683–690.
BOONE, M. D., R. D. SEMLITSCH, J. F. FAIRCHILD, AND B. B.ROTHERMEL. 2004b. Effects of an insecticide onamphibians in large-scale experimental ponds. Ecolog-ical Applications 14:685–691.
BOONE, M. D., R. D. SEMLITSCH, AND C. MOSBY. 2008.Suitability of golf course ponds for amphibian meta-morphosis when bullfrogs are removed. ConservationBiology 22:172–179.
BREDEN, F., AND C. H. KELLY. 1982. The effect ofconspecific interactions on metamorphosis in Bufoamericanus. Ecology 63:1682–1689.
BROWN, C. J., B. BLOSSEY, J. C. MAERZ, AND S. J. JOULE.2006. Invasive plant and experimental venue affecttadpole performance. Biological Invasions 8:327–338.
COLBURN, E. A. 2004. Vernal Pools: Natural History andConservation. The MacDonald & Woodward Publish-ing Company, Blacksburg, Virginia, U.S.A.
COLLINS, J. P., AND A. STORFER. 2003. Global amphibiandeclines: sorting the hypotheses. Diversity and Distri-butions 9:89–98.
CROWDER, L. B., AND W. E. COOPER. 1982. Habitatstructural complexity and the interaction betweenbluegills and their prey. Ecology 63:1802–1813.
CRUMP, M. L. 1989. Effect of habitat drying ondevelopmental time and size at metamorphosis in Hylapseudopuma. Copeia 1989:794–797.
DEVITO, J. 2003. Metamorphic synchrony and aggregationas antipredator responses in American toads. Oikos103:75–80.
DIEHL, S., AND P. EKLOV. 1995. Effects of piscivore-mediated habitat use on resources, diet, and growth ofperch. Ecology 76:1712–1726.
DODD, C. K., JR., AND L. L. SMITH. 2003. Habitatdestruction and alteration: historical trends and futureprospects for Amphibians. Pp. 94–112. In R. D.Semlitsch (Ed.), Amphibian Conservation. SmithsonianBooks, Washington, D.C., U.S.A.
EGAN, R. S., AND P. W. C. PATON. 2004. Within-pondparameters affecting oviposition by wood frogs andspotted salamanders. Wetlands 24:1–13.
28 HERPETOLOGICA [Vol. 65, No. 1

EKLOV, P. 1997. Effects of habitat complexity and preyabundance on the spatial and temporal distributions ofperch (Perca fluviatilis) and pike (Esox lucius).Canadian Journal of Fisheries and Aquatic Sciences54:1520–1531.
GOSNER, N. 1960. A simplified table for staging anuranembryos and larvae with notes on identification.Herpetologica 16:83–190.
GRAY, M. J., L. M. SMITH, AND R. I. LEYVA. 2004. Influenceof agricultural landscape structure on a Southern HighPlains, USA, amphibian assemblage. Landscape Ecol-ogy 19:719–729.
GRENOUILLET, G., D. PONT, AND K. L. SEIP. 2002.Abundance and species richness as a function of foodresources and vegetation structure: juvenile fish assem-blages in rivers. Ecography 25:641–650.
HORAT, P., AND R. D. SEMLITSCH. 1994. Effects ofpredation risk and hunger on the behaviour of twospecies of tadpoles. Behavioral Ecology and Sociobiol-ogy 34:393–401.
HOWARD, R. D. 1978. The influence of male-defendedoviposition sites on early embryo mortality in bullfrogs.Ecology 59:789–798.
KIESECKER, J. M., A. R. BLAUSTEIN, AND L. K. BELDEN.2001. Complex causes of amphibian population de-clines. Nature 410:681–684.
KNUTSON, M. G., J. R. SAUER, D. A. OLSEN, M. J. MOSSMAN,L. M. HEMESATH, AND M. J. LANNOO. 1999. Effects oflandscape composition and wetland fragmentation onfrog and toad abundance and species richness in Iowaand Wisconsin, USA. Conservation Biology 13:1437–1446.
KUPFERBERG, S. J. 1997. Bullfrog (Rana catesbeiana)invasion of a California river: the role of larvalcompetition. Ecology 78:1736–1751.
LANGELLOTTO, G. A., AND R. F. DENNO. 2004. Responsesof invertebrate natural enemies to complex-structuredhabitats: a meta-analytical synthesis. Oecologia 139:1–10.
LAURILA, A., AND J. KUJASALO. 1999. Habitat duration,predation risk and phenotypic plasticity in common frog(Rana temporaria) tadpoles. Journal of Animal Ecology68:1123–1132.
LAWLER, S. P., D. DRITZ, T. STRANGE, AND M. HOLYOAK.1999. Effects of introduced mosquitofish and bullfrogson the threatened California red-legged frog. Conser-vation Biology 13:613–622.
LEHTINEN, R. M., AND S. M. GALATOWITSCH. 2001.Colonization of restored wetlands by amphibians inMinnesota. American Midland Naturalist 145:388–396.
LIMA, S. L. 1990. Protective cover and the use of space:different strategies in finches. Oikos 58:151–158.
MAERZ, J. C., C. J. BROWN, C. T. CHAPIN, AND B. BLOSSEY.2005. Can secondary compounds of an invasive plantaffect larval amphibians? Functional Ecology19:970–975.
MCDIARMID, R. W., AND R. ALTIG (Eds.). 1999. Tadpoles:The Biology of Anuran Larvae. The University ofChicago Press, Chicago, Illinois, U.S.A.
MITSCH, W. J., AND J. G. GOSSELINK. 2007. Wetlands, 4thed. John Wiley and Sons, Hoboken, New Jersey, U.S.A.
MORIN, P. J. 1983. Predation, competition, and thecomposition of larval anuran guilds. Ecological Mono-graphs 53:119–138.
NICIEZA, A. G. 2002. Interacting effects of predation riskand food availability on larval anuran behaviour anddevelopment. Oecologia 123:497–505.
PEACOR, S. D., AND E. E. WERNER. 1997. Trait-mediatedindirect interactions in a simple aquatic food web.Ecology 78:1146–1156.
PECHMANN, J. H. K., R. A. ESTES, D. E. SCOTT, AND W. J.GIBBONS. 2001. Amphibian colonization and use ofponds created for trial mitigation of wetland loss.Wetlands 21:93–111.
PERSSON, L., AND P. EKLOV. 1995. Prey refuges affectinginteractions between piscivorous perch and juvenileperch and roach. Ecology 76:70–81.
PETRANKA, J. W. 1989. Density-dependent growth andsurvival of larval Ambystoma: evidence from whole-pond manipulations. Ecology 70:1752–1767.
PETRANKA, J. W., C. A. KENNEDY, AND S. S. MURRAY. 2003.Response of amphibians to restoration of a SouthernAppalachian wetland: a long-term analysis of commu-nity dynamics. Wetlands 23:1030–1042.
POUNDS, A. J. 1988. Ecomorphology, locomotion, andmicrohabitat structure: patterns in a tropical mainlandAnolis community. Ecological Monographs 58:299–320.
SCHIESARI, L. 2006. Pond canopy cover: a resourcegradient for anuran larvae. Freshwater Biology51:412–423.
SEMLITSCH, R. D. 2003. Conservation of pond-breedingamphibians. Pp. 8–23. In R. D. Semlitsch (Ed.),Amphibian Conservation. Smithsonian Books, Wash-ington, D.C., U.S.A.
SEMLITSCH, R. D., D. E. SCOTT, AND J. H. K. PECHMANN.1988. Time and size at metamorphosis related to adultfitness in Ambystoma talpoideum. Ecology 69:184–192.
SIH, A. 1986. Antipredator responses and the perceptionof danger by mosquito larvae. Ecology 67:434–441.
SKELLY, D. K., L. K. FREIDENBURG, AND J. M. KIESECKER.2002. Forest canopy and the performance of larvalamphibians. Ecology 83:983–992.
SKELLY, D. K., E. E. WERNER, AND S. A. CORTWRIGHT.1999. Long-term distributional dynamics of a Michiganamphibian assemblage. Ecology 80:2326–2337.
SMITH, G. R., AND B. L. DOUPNIK. 2005. Habitat use andactivity level of large American bullfrog tadpoles:choices and repeatability. Amphibia-Reptilia 26:549–552.
SREDL, M. J., AND J. P. COLLINS. 1992. The interaction ofpredation, competition, and habitat complexity instructuring an amphibian community. Copeia1992:607–614.
STUART, S. N., J. S. CHANSON, N. A. COX, B. E. YOUNG,A. S. L. RODRIQUES, D. L. FISCHMAN, AND R. W.WALLER. 2004. Status and trends of Amphibian declinesand extinctions worldwide. Science 306:1783–1786.
TARR, T. L., AND K. J. BABBITT. 2002. Effects of habitatcomplexity and predator identity on predation of Ranaclamitans larvae. Amphibia-Reptilia 23:13–20.
TRENHAM, P. C., W. D. KOENIG, M. J. MOSSMAN, S. L.STARK, AND L. A. JAGGER. 2003. Regional dynamics ofwetland-breeding frogs and toads: turnover and syn-chrony. Ecological Applications 13:1522–1532.
VAN BUSKIRK, J. 2005. Local and landscape influence onamphibian occurrence and abundance. Ecology86:1936–1947.
March 2009] HERPETOLOGICA 29

WARKENTIN, K. M. 1992. Microhabitat use and feedingrate variation in green frog tadpoles (Rana clamitans).Copeia 1992:731–740.
WELLBORN, G. A., D. K. SKELLY, AND E. E. WERNER. 1996.Mechanisms creating community structure across afreshwater habitat gradient. Annual Review of Ecologyand Systematics 27:337–363.
WERNER, E. E. 1986. Amphibian metamorphosis: growthrate, predation risk, and the optimal size at transfor-mation. The American Naturalist 128:319–341.
WERNER, E. E., AND D. J. HALL. 1988. Ontogenetic habitatshifts in bluegill: the foraging rate – predation risktrade-off. Ecology 69:1352–1366.
WERNER, E. E., J. F. GILLIAM, D. J. HALL, AND G. G.MITTELBACH. 1983. An experimental test of the effectsof predation risk on habitat use in fish. Ecology64:1540–1548.
WHIGHAM, D. F. 1999. Ecological issues related to wetlandpreservation, restoration, creation, and assessment. TheScience of the Total Environment 240:31–40.
WHITTINGHAM, M. J., AND K. L. EVANS. 2004. The effects ofhabitat structure on predation risk of birds in agricul-tural landscapes. Ibis 146:210–220.
WILBUR, H. M. 1977. Density-dependent aspects ofgrowth and metamorphosis in Bufo americanus. Ecol-ogy 58:196–200.
WILBUR, H. M. 1980. Complex life cycles. Annual Reviewof Ecology and Systematics 11:67–93.
WILBUR, H. M., AND J. P. COLLINS. 1973. Ecologicalaspects of amphibian metamorphosis. Science182:1305–1314.
WILDER, S. M., AND D. B. MEIKLE. 2005. Reproduction,foraging, and the negative density-area relationship of ageneralist rodent. Oecologia 144:391–398.
WIRSING, A. J., M. R. HEITHAUS, AND L. M. DILL. 2007.Fear factor: do dugongs (Dugong dugon) trade food forsafety from tiger sharks (Galeocerdo cuvier)? Behavior-al Ecology 153:1031–1040.
ZEDLER, J. B., AND S. KERCHER. 2004. Causes andconsequences of invasive plants in wetlands: opportu-nities, opportunists, and outcomes. Critical Reviews inPlant Sciences 23:431–452.
.Accepted: 13 February 2009
.Associate Editor: Kimberly Babbitt
30 HERPETOLOGICA [Vol. 65, No. 1