adrenergic influence of uterine ;musc1e …nopr.niscair.res.in/bitstream/123456789/19026/1/ijeb...
TRANSCRIPT

Indian Journal of Experimental Biology Vol. 37, April 1999, pp. 350-354
Adrenergic influence of uterine ;musc1e contractions stimulated by a glycoside from the root of Dalbergia saxatilis
C N Uchendu & B FLeek·
Department of Veterinary Physiology & Pharmacology, University of Nigeria, Nsukka, Nigeria
*Department of Veterinary Physiology & Biochemistry, University College Dublin, Ballsbridge, Dublin 4
Received 22 January 1998; revised 7 January 1999
The mechanism of uterine muscle contraction stimulated by a triterpenoid glycoside (dalsaxin) isolated from the root of D. saxarilis was investigated by in vitro methods in the rat. Dalsaxin caused a dose-related increase in uterine muscle contraction . The contraction was single and transient and was abolished by moderate doses of isoprenaline (1.80 nmol-O.40 ~mol) and salbutamol (0.13-25 ~mol). Adrenaline (9.10 nmol) also caused a reversible decrease (92.6%; P<O.OI) in myometrial contraction stimulated by this glycoside (0.24 mglmJ). Uterine muscle responses to dalSaxin (0.24 mglmJ) were enhanced by the p-adrenergic receptor antagonist, propranolol , in a dose related manner. Atipamezole (1 .50 nglmJ) but not prazos if) (7.72 nmol-15 .60 nmol) substantially reduced (80%; P<O.OI) myometrial contractions induced by this uterine spasmogen. The results suggest that dalsaxin enhances uterine muscle contraction by stimulating post junctional ~adrenergic receptors. presumably by inhibiting plasma membrane adenylate cyclase system and its associated increase in intracellular cAMP content.
Aqueous root extract of Dalbergia saxatilis, subfamily Leguminosae, is widely used in native medicine to accelerate birth and to expel the placenta in human subjects in South-eastem Nigeria. During the course of screening the root extracts of this medicinal plant for biological activity, a colourless, rhomboid . crystalline glycoside, melting at 211 °-213°C (uncorrected) was isolated as the active uterotonic principle. The isolation and partial chemical characterization of this active glycoside, now known by the trivial name dalsaxin , have been described elsewhere! . Influence of catecholaminergic receptors on dalsaxin modulated uterine force is reported in this communication.
Materials and Methods Animals-Non-pregnant Wistar rats of breeding age,
weighing about 200-250g were used for the experiments. The rats were supplied by the Faculty of Agriculture, University College Dublin, and reared under specific-pathogen-free (SPF) conditions. Standard rat chow and tap water were provided ad libitum . Each female rate received 0.1 mg/kg body weight of estradiol benzoate (Sigma, USA) in paraffin subcutaneously 24 hr prior to the experiments.
Tissue preparation arul isometric contraction studies-Uterine horns from the rats were trimmed free of fat and connective tissue. Uterine strip measuring
about 12 rom long was cut out and attached at one end to a specimen holder and at the other to a myograph attached to a Lectromed Multitrace 2 isometric force displacement transducer/amplifier. The tissue was suspended vertically in a 30 m1 conventional organ bath containing Krebs solution (37°C) and perfused continuously with O2 (95%), CO2 (5%) gas mixture. The physiological solution had (romol): KCI, 4 .7; NaC!., 118; CaCh, 2.5; KH2P04, 1.2; NaHC03, 2.5; MgS04, 1.2; and glucose, II . Recordings of isometric myometrial contractions were displayed on an LC 475 Macintosh computer screen connected in parallel to a MacLab/2e instrument and to the transducer.
All strips used were put under a small amount of resting tension (0.21 g) and experiments were started within 45 min following the development of stable, rhythmic, spontaneous contractions.
Drugs-Following drugs were used: salbutamol (Allen & Hanburys, England), isoprenaline (Pharmax, England), propranolol (Antigen Pharmaceuticals, Ireland), adrenaline (Sigma, USA), Prazosin (Invicta Pharmaceuticals, England), atipamezole (SmithKlime Beecham, Ireland), and acetylcholine (Sigma, USA) .
A minimum of 3 min was allowed for tissue responses in each experiment following which the preparation. was washed 2-3 times with Krebs solution in preparation for the subsequent experiment. Except

UCHENDU & LEEK: ADRENOCEPTORS & MYOME1RIAL CONlRAcnON BY DALBERGIA GLYCOSIDE 351
otherwise indicated, concentrations of test substances given in the text are all final nutrient bath concentrations.
Analysis of data-Differences between the means of the control and the experimental groups were compared using paired and unpaired Students's t-test as appropriate.
Results Effects of selective and non-selective {3-adrenoceptor
agonists on dalsaxin activity-Application of salbutamol (0.13-0.25 Jlmol) and isoprenaline (1.80 nmol-0.40 Jlmol) as selective and non-selective ~-adrenergic receptor agonists respectively, followed simultaneously by dalsaxin (0.11-0.24 mglml) resulted in complete abolition of dalsaxin mediated increases in uterine force (Fig. I) .
Adrenaline (9.10 nmol) also caused a sustainable
Dalsaxin
inhibition of acetylcholine (1.82-3.64 Jlmol; 100% inhibition, P<O.OI) and dalsaxin (0.24 mglm1; 92.6% inhibition; P<O.Ol) induced myometrial contraction (Fig. 2). However, the ~-adrenergic receptor blocker, propranolol ('I Jlmol) was able to reverse the inhibition caused by adrenaline. In addition, the frequency of contraction induced by dalsaxin (0.24 mglml) was potentiated by propranolol in a dose-related manner (Fig. 3), provided a fixed dose of the spasmogen was preceded by increasing but non-cumulative doses of propranolol (0.3-3.0 Jlmol).
Effects of ar and aradrenoceptor antagonists on dalsaxin mediated uterine force-To verify whether uterine muscle responses to dalsaxin were mediated via cell membrane a-receptor activation, varying doses of prazosin (7.72-15.60 nmol) and atipamezole (1.39-2.7 nglml) which are specific al- and aradrenoceptor
"."'WI""'"
Isopr
5min r--l
Salb.
5min r--r
Fig. I-Inhibitory effects of isoprenaline (1.80 nmol) and salbutarpol (0.13 ~mol ) on in vitro uterine muscle contraction stimulated by dalsaxin (0.1 I mg!ml). Note the absence of spontaneous spikes following the introduction of dalsaxin (arrow) 60s after the p-adrenoceptor agonists respectively. Dalsaxin was applied for the period indicated on the horizontal bars (n = 5).
Adre.(9.l0 om) ----------____________ _
6
:§
jWjAcetylchOline (lUll; 1.82 3.64
o --
, + .... (a)
5 min
(b)
Dalsaxin (0.24 mglmJ)
Propr. ljUl1 -(c) (d)
Fig. 2-Effect of adrenaline (9.10 nmol) on acetylcholine and dalsaxin-induced myometrial contraction. Acetylcholine (a), dalsaxin (b,d) and propranolol (c) were applied at the points indicated by the arrows respectively. (n = 5).
352 fNOlAN J EXP BIOL, APRlL 1999
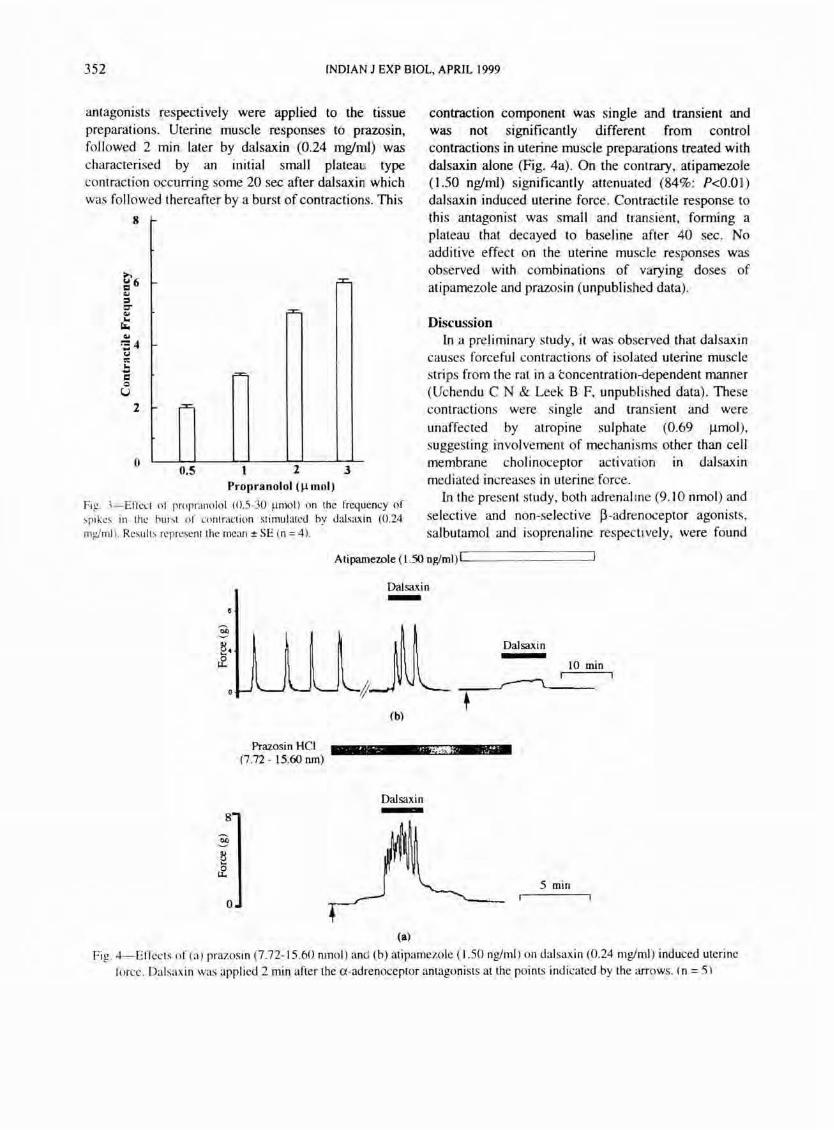
antagonists respectively were applied to the tissue preparations. Uterine muscle responses to prazosin, followed 2 min later by dalsaxin (0.24 mglml) was characterised by an initial small plateaU! type contraction occurring some 20 sec after dalsaxint which was followed thereafter by a burst of contractions. This
8
e-6 = .., :I 0-.., ... r.. .., ~4 .... 0:: ...
I -c 0 U
2 r=,
o 0.5 2 3
Propranolol (~mol)
Fig. 3-EITect of propranolol (0.5-30 ~mol ) on the frequency of ~pikes in the hurst of contrac tion stimu lated by dalsaxin (0.24 Illg/Illl ). Re~ ult s represent the mean ± SE (n = 4).
contraction component was single and transient and was not significantly different from control contractions in uterine muscle preparations treated with dalsaxin alone (Fig. 4a). On the contrary, atipamezole (1.50 nglml) significantly attenuated (84%: P<O.O I) dalsaxin induced uterine force . Contractile response to this antagonist was small and transient, forming a plateau that decayed to baseline after 40 sec. No additive effect on the uterine muscle responses was observed with combinations of varying doses of atipamezole and prazosin (unpublished data).
Discussion In a preliminary study, it was observed that dalsaxin
causes forceful contractions of isolated uterine muscle strips from the rat in a concentration-dependent manner (Uchendu C N & Leek B F, unpubl ished data) . These contractions were single and transient and were unaffected by atropine sulphate (0.69 Ilmol), suggesting involvement of mechanisms other than cell membrane cholinoceptor activation In dalsaxin mediated increases in uterine force.
In the present study, both adrenal ine (9.10 nmol) and selective and non-selective ~-adrenoceptor agonists, salbutamol and isoprenaline respectively, were found
Atipamewle (1.50 ng/ml)!L----- -----'
8
3 8 &
0
Prazosin HCl (7.72 - 15.60 run)
Dalsaxin -
(b)
. , ....... ,~~ .. :../~'
Dalsaxin
(a)
Dalsaxin
10 min ·
5 min
Fig. 4-Effects o f (a) prazosin (7.72-15 .60 nmol ) and (b) atipamezo!e (1.50 ng/ml) o n dalsaxin (0.24 mg/mI) induced uterine
force. Dalsaxin was applied 2 min after the a-adrenoceptor antagonists at the po ints indicated by the arrows. (n = 5)

UCHENDU & LEEK: ADRENOCEPTORS & MYOMETRIAL CONTRACDON BY DALBERGIA GLYCOSIDE 353
to block uterine muscle contractions induced by moderate doses of this active glycoside isolated from the root of DaLbergia saxatiLis. This finding is consistent with the observations of other investigators using rat uterine muscle preparations2.3. Like in other smooth muscle cells4
.5, ~-adrenergic relaxation of uterine muscle is thought to involve primarily the activation of membrane bound adenylate cyclase system with a concomitant increase in cytosolic cAMP concentration6
, leading to phosphorylation of myosin light chain kinase (MLCK) via a cAMP-dependent protein kinase enzyme7
. Phosphorylation of MLCK is concurrent with failure of myosin-actin interaction and hence uterine force. The usual increase in calcium (Ca2+) extrusion across the plasma membrane that accompanies elevated cytosolic cAMP concentration is not excluded in this relaxation mechanism.
Although smooth muscle relaxation has also been accomplished without elevation of intracellular cAMp8
, adrenergic inhibition of dalsaxin modulated uterine force may be related to the elevation of uterine cAMP content since propranolol (I J..lmol) was able to reverse the effect of adrenaline on dalsaxin (0.24 mglm) mediated myometrial contraction (Fig. 2); and to potentiate, in a dose related manner, dalsaxin modulated force (Fig. 3), provided increasing but noncumulative doses of the ~-adrenoceptor antagonist were preceded by a fixed dose of the spasmogen. Although a direct measurement of uterine cytosolic cAMP level was not undertaken in the present study, there is evidence that propranolol neither elicits uterine muscle contraction nor affects cAMP but does significantly reduce the inhibitory effect of isoprenaline on myometrial contraction9 and its associated increase in cAMP content lO
• In that study, these two effects of isoprenaline were found to be completely abolished when a higher concentration of propranolol (7 J..lmollkg) was used. These views have been further strengthened by the observations that ~adrenergic agonist-induced changes in muscle contraction can be reproduced by treatment with exogenous cAMp JO or by direct application of cAMP intracellularly by micro injection J J.
Under the conditions of the present experiments, it was intriguing that prazosin, a predominantly uJ-adrenoceptor antagonist, was unable to inhibit uterine muscle responses to dalsaxin (Fig. 4a) . This would suggest that mechanisms other than hydrolysis of phosphatidyl inositol-4,5-bisphosphate and generation of inositol-IA,S-trisphosphate (P,), that is , a
phosphoinositide-phospholipase mechanisml2.13 are involved in uterine force .
C transducing dalsaxin modulated
In sharp contrast, contractions stimulated by dalsaxin (0.24 mglmJ) were significantly (P<O.Ol) attenuated by 1.50 nglmJ atipamezole, a selective ~adrenoceptor antagonist (Fig. 4b), suggesting that dalsaxin stimulates uradrenoceptors to cause contraction of the uterus. Although uterine muscle cells contain both UI and uradrenoceptor subtypesl4, contraction is known to be mediated solely by ul-adrenoceptor activation l5 . The reason for the discrepancy between the present results and that of other investigators is unknown. While some workers have adduced an indirect physiological role for postjunctional uradrenoceptor.s, precisely to augment stimulus arising from ul-adrenoceptor activation l
6-18,
the results of the present study suggest a direct involvement of uradrenoceptors in the contractile process. As its main mechanism of action it is proposed that dalsaxin activates surface membrane u 2-adrenoceptors, resulting in the inhibition of the following three cellular components: membrane-bound adenylate cyclase system, intracellular cAMP, and intracellular Ca2+ binding to the sarcoplasmic reticulum. As a result, there is an influx of Ca2+ from the extracellular millieu, with subsequent increase in cytosol ie-free Ca2+ concentration. This increase in intracellular Ca2+ is detected by intracellular highaffinity Ca2+-binding protein (calmodulin) which then converts the calcium signal to specific cellular responses.
Further, it is considered that in a system where two receptor subtypes are present on the cell plasma membrane, the expected overall effect of a combination of two antagonists is to elicit a greater inhibition of the responses to the agonist (that is, a synergistic response is expected). However, since the inhibitory effect of prazosin (7.72-15.60 nmol) and atipamezole (1.39-2.70 nglmJ) put together was not significantly different from the effect of atipamezole alone, it could be inferred that dalsaxin contractile activity was mediated predominantly through postjunctional uradrenoceptor activation. Templeton et aL.19 had postulated that tissue stimulants are required to enhance or ' uncover' a hitherto queiscent postjunctional u 2-adrenoceptor expression. In their experiment with rat tail artery, both noradrenaline and phenylephrine were found to evoke dose-dependent

354 INDIAN J EXP BIOL, APRIL 1999
pressor responses on the ti ssue whereas UK-I 4,304, an
u 2-adrenoceptor agonist, was inactive. However, following the induction of tone with arginine vasopressin (AVP), responses to UK, 14-304 and
noradrenaline in the presence of 0 .05 Jlmol prazosin were significantly potentiated, showing leftward displacement of the dose-response curve. These results were taken to indicate that responses potentiated by A VP-induced tone actually arose from stimulation of
receptors other than ul-adrenoceptor (prazos in resistant, rauwolscine sensitive) . Similar findings were reported in rabbit isolated distal saphenous . artery using the physiological stimulants, angiotensin II and UK-14,30420 and in canine saphenous artery and portal vein, using the Ca2+-channel facilitator Bay K 8644 and
I d · F . I 2122 prostag an In 2a respectl ve y . .
Taken together, these results are considered
complimentary to the results of the present study for a
functional post-junctional u2-adrenoceptor (prazosin
res istant , atipamezole sensti ve) in the rat uterine
muscle cell , but in the present context, with a dual
physiological role for dalsax in both as the 'activator'
and the fac ilitator' of myometrial u radrenoceptor
function . The fact that uradrenoceptors were found in
high concentration in human term uterus, and were
functionally linked to inhibiti on of adenyly l cyclase
system via a Gi protein 2.1 further lend support to this
concept.
Acknowledgement Financial support for thi s project from the World
Bank in collaboration with the National Universities Commission in Nigeria is gratefully acknowledged.
References I Uchendu C N, Isolation, characterization and physiological
effects of active glycaside f rom the root of Dalbergia saxatilis. Ph .D Thesis, Universily of Nigeria, Nsukka, 1996.
2 Kroegger E A & Marshal J M, Am J Physiol, 226 ( 1974) , 1298.
3 Johansson S R M & Andersson R G A, Acta Pharmacol. Toxicol, 42 ( 1978), 347.
4 Jones A W, Bylund D B & Forte L R , Am. 1. Physiol, 246 (1984), H306.
5 Jones A W, Smith J M, Bylund D B & Jones S B, Vascular neuroeffector mechanisms (Elsevier, Amsterdam), 1985 .
6 Conti M A & Adelstein R S, Fed Proc, 39 (1980), 1569 . 7 Fortier M, Chase D, Korenman S G & Krall J F, Am J
Physiol, 245 ( 1983) , C84. 8 Do Khac L, Mokhtari A, Renner M & Harbon S, Mol
Pharmacal, 41 ( 1992),509 . 9 Johansson S R M & Andersson R G G, Acta Pharmacol
Toxicol, 47 ( 1980), 5. 10 Honeyman T , Merriam, P & Fay F S, Mol Pharmacol, 14
( 1978),86. II Fay F S & Taylor S R, Biophys J, 21 ( 1'978), 184a. I 12 Anwer K. Hovin gton J A & Sanborn B M, Endocrinology,
124 ( 1l) ~9 ) 2<)<)5 . 13 Breuiller-Fouche M, Douall a- Bell F, Makaa K, Geny B &
Ferre F, J Pharmacol Exp Tiler, 258 ( 199 1), 82. 14 Roberts J M, Insel P A, Goldlien R D & Goldli en A, Nature,
270 ( 1977), 624. 15 Hoffman B B, Lavni T N, Lefkowi tz R J & Fuffolo R R, 1
Pharmacal Exp Ther, 219 ( 198 1 ), 290. 16 Shepperson N B, Brl Pharmacal, 8 1 ( i 984), 15p. 17 Xiao X H, Medgett I C & Rand M J li n, Exp Pharmacal
Physiol, 14 (1987), 90 I. 18 Xiao X H & Rand M J , / Q'h /111 Cong Pharmacol, ( 1987), .
abstract 0 I 02. 19 Templeton A, Macmillan J, McGrath J C, Storey N D &
Wilson V G, Br 1 Pharmacol, 97 (1989), 563. 20 Dun W R, McGrath J C & Wilson V G. Br J Pharmacal, 96
( 1989), 259. 2 1 Su lpi zio A & Hieble J P, Eur J Pharmacal, 135 ( 1987), 107. 22 Furuta T , Naunym Schmie Arch Pharmacal, 337 ( 1988),525. 23 Breuiller M. Rouot B, Litime M N, Leroy M J & Ferre F, 1
Clin Endocrinol Metab, 70 ( 1990) , 1299 .