the caecilian cardio-vascular system and intra-amphibian relationships
TRANSCRIPT

J . Zool., Lond. (1970) 160, 199-229
The caecilian cardio-vascular system and intra-amphibian relationships
RONALD LAWSON
Department of Biology, University of Salford, Salford, 5, England
(Accepted 11 August 1969)
(With 8 figures in the text)
The cardio-vascular system of Hypogeophis is described and compared with that of other caecilians. The extant caecilians are divisible into two geographical groups in which the varying degrees of asymmetry of the arterial arches evolved in parallel. The nomenclature of the cephalic veins is discussed and the cranial circulation of caecilians is compared with that of some urodeles and anurans. The general similarities which exist between the vascular system of Mecilians and urodeles may be indicative of a close relationship. However, it is pointed out that more detailed information is required before intra-amphibian relation- ships can be established with any degree of certainty.
Contents
Introduction . . . . . . . . . . Materials and methods . . . . . . Heart . . . . . . . . . . Arterial system . . . . . . . . Venous system . . . . . . . . Vessels of the vertebral and body musculature Vessels of the brain and spinal cord . . Discussion . . . . . . . . . . summary . . . . . . . . . . References . . . . . . . . . .
. . . .
.. '.
. . . .
. . . .
. . . .
. . . .
. . . .
. . . .
. . . .
. . . .
..
..
..
..
..
..
..
.. ,. ..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
..
page .. 199 . . 200 .. 200 .. 202 . . 209 .. 214 .. 216 .. 218 .. 225 . . 226
Introduction
A large volume of literature on the monophyletic, diphyletic or even polyphyletic nature of the amphibia has been produced, but there is no general agreement on the problem. A good deal of the uncertainty is due to an inadequate fossil record but the position is aggravated by the lack of anatomical information on extant forms.
The paucity of data is particularly marked in relation to the caecilians and some of the papers which discuss relationships within the Amphibia completely ignore this group (e.g. Schmalhausen, 1959 and Holmgren, 1952). The present study is a continuation of a series providing information on one caecilian species (Lawson, 1963,1965; 1966). It also seeks to correlate, at least to some degree, these data with those available for other caecilians, urodeles and anurans. It is clear that many more detailed anatomical studies are required to effect any realistic appraisal of intra-amphibian relationships.
199

200 R. LAWSON
Materials and methods The major portion of the detail of the circulatory system was obtained by dissection of a large
number of specimens of Hypogeophis rostrutus rostratus (Parker, 1958). The material from the Seychelles had been preserved in formalin for some time and in general injections were not possible. However, in some cases it was possible to inject small areas which were void of blood using latex, Indian ink or Prussian blue in gelatin. In order to establish the minute course of the cranial blood vessels microscopical serial sections were made, and in addition similar sections of various parts of the body were produced to facilitate the description of the circulatory system of the vertebral and body musculature.
In the text the major vessels are given numbers, the large branches of these vessels are desig- nated by capital letters and the subdivisions of these by small letters. For example, the internal carotid artery is 2, the stapedial artery (a major branch of the internal carotid) is 2A and a small branch of the stapedial artery (muscular ramus) which supplies the m. levator mandibulae and the m. levator quadrati is therefore 2A(a).
The heart The heart of Hypogeophis rostratus lies ventrally at about one third of the body length
from the anterior end. It is completely enclosed in a tough semi-transparent pericardium which is attached to the apex of the ventricle by three bundles of connective tissue. These connections consist of a large central bundle and two smaller lateral ones. Such apical bundles (gubernaculum cordis) are a characteristic feature of the heart in fishes and are also known to occur in some urodeles and chelonians. The ventral surface of the ventricle, conus and the lower part of the truncus is covered by an extensive network of blood vessels which are often visible through the pericardium. These vessels also extend to the dorsal side of the ventricle where they again form an extensive network. This elaborate series of coronary vessels opens into the sinus venosus and is therefore a network of coronary veins which also communicates with an extensive system of spaces within the myocardium. It is evident that such spaces in turn are continuous with the cavity of the ventricle and that during the contraction of the ventricle blood is squeezed into the coronary spaces in the myocardium, then into the coronary veins on the surface of the heart and returned to the general circulation via the sinus venosus. The heart of Hypogeophis rostratus therefore possesses a complex coronary circulation quite unlike that described in any other group of amphibia. The details of the structure of the heart, and the coronary system have been described by the author (Lawson, 1966) and some attempt was made to describe the mechanism of passage of blood through the heart.
The truncus arteriosus gives rise to two common carotid arteries. The left common carotid has its origin a few millimetres closer to the heart than the right and where the two carotids lie together their cavities are separated by a common wall (Lawson, 1966).
A single systemico-pulmonary arch is present and this curves sharply over the apex of the right atrium. As it passes above the atria the arch gives rise to a pulmonary branch which in turn divides into a large right pulmonary artery (to the well developed right lung) and a smaller left pulmonary artery to the much reduced left lung. The single systemico- pulmonary arch is said to represent the left systemic and pulmonary arches (Marcus, 1935). Between the common carotid arteries and the pulmonary arch the systemico- pulmonary arch gives rise to a series of vessels. The first two of these extend cranially to
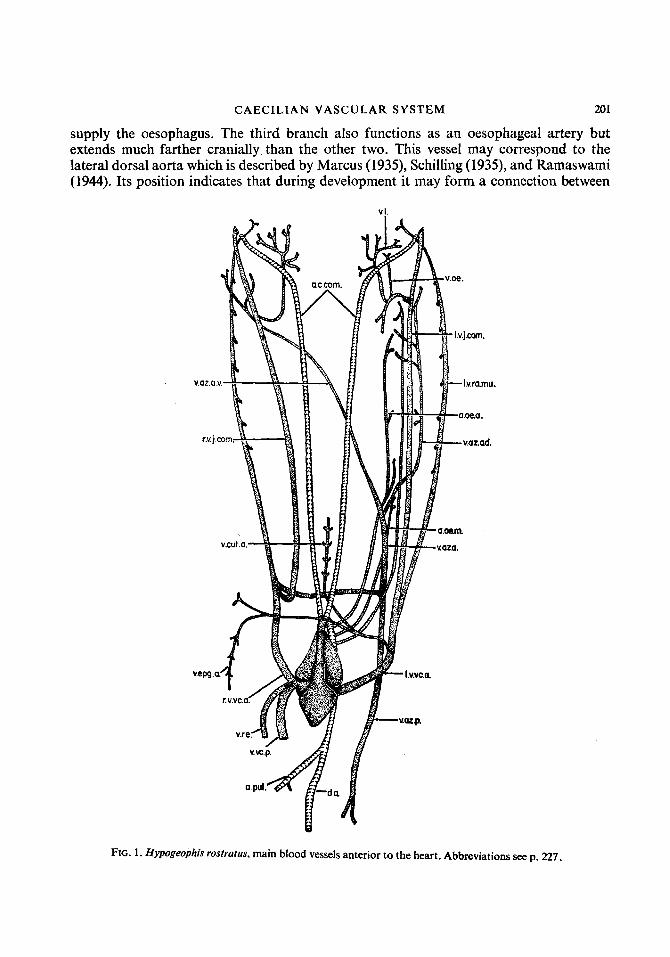
CAECILIAN VASCULAR SYSTEM 201
supply the oesophagus. The third branch also functions as an oesophageal artery but extends much farther cranially than the other two. This vessel may correspond to the lateral dorsal aorta which is described by Marcus (1939, Schilling (1935), and Ramaswami (1944). Its position indicates that during development it may form a connection between
v. I
FIG. 1 . Hypogeophis rostratus, main blood vessels anterior to the heart. Abbreviations seep. 227.

202 R. LAWSON
the carotid and systemic arches and hence represent a lateral dorsal aorta (ductus caroticus). In the adult, however, no such connection can be demonstrated and the vessel breaks up into a plexus on the anterior portion of the oesophagus. Generally two further branches of the systemico-pulmonary arch are found in this region and these are vertebral arteries (Fig. 1).
The arterial system 1 . COMMON CAROTID ARTERY
The common carotid arteries carry blood to the head and run together until they reach the pharyngeal region where they diverge and each artery then curves dorsolaterally. At this point each common carotid artery gives rise to two vessels which curve abruptly posteriorly (Figs 1 and 2). 1A. Vertebral artery
After leaving the common carotid this vessel curves posterodorsally and then runs medially below the ventral ramus of the first spinal nerve. At this point the vertebral artery then accompanies the ventral ramus of the first spinal nerve and gives rise to a number of small vessels which supply the muscles immediately surrounding the vertebral column of this region (Fig. 2). 1 B. Oesophageal artery
This artery stems from the common carotid with the vertebral artery and forms a plexus on the dorsal side of the anterior limits of the oesophagus.
After the origin of these vessels each common artery continues anteriorly until the level of the thyroid gland is reached. At this point the common carotid has two major divisions, the internal and external carotid (Fig. 2).
2. THE INTERNAL CAROTID A R ~ R Y runs dorsomedially below the posterior fibres of the m. levator mandibulae posterior (Fig. 2) and then divides into a stapedial artery and a palato- cerebral artery. 2A. The stapedial artery runs medial to the retroarticular process of the lower jaw and then accompanies the lateral head vein (vena capitis lateralis) through the cranio-quadrate passage to the region of the post-orbital sinus where it subdivides (Figs 2 and 3). 2A(a). The muscular rami are small branches of the stapedial artery which supply the levator mandibulae muscles and the m. levator quadrati (Fig. 4). 2A(b). The internal maxillary artery is accompanied by the maxillary division of the trigeminus nerve, and is also associated with the maxillary vein and maxillary sinus (see below). The internal maxillary artery supplies the retractor muscle of the tentacle (m. retractor tentaculi) and the skin surrounding the tentacular pore (Fig. 2). 2A(c). The mandibular artery accompanies the mandibular ramus of the trigeminus through the levator muscles of the mandible and enters the lower jaw via a large foramen immediately anterior to the articular facet of the mandible. On entering the lower jaw the mandibular artery divides into a labial artery and an alveolar artery. The labial artery leaves the lower jaw with the intermandibular ramus of the fifth nerve and supplies the intermandibularis muscle and the skin overlying it. The alveolar branch accompanies the

FIG
. 2. C
epha
lic v
ascu
lar s
yste
m o
f H
ypog
eoph
is ro
stra
tus.
Abb
revi
atio
ns s
ee p
. 227
.
h) 8

204 R. LAWSON
alveolar rami of the fifth and seventh cranial nerves. These nerves and the alveolar artery run in a groove in the medial border of the pseudodentary and supply the dental tissues and the lateral epithelium of the floor of the mouth (Fig. 2). 2B. Palato-cerebral artery
This vessel is the second major subdivision of the internal carotid artery and runs dorsally and medially to lie in a groove in the large 0s basale which forms the posterior part of the skull (see Lawson, 1963). The artery then runs anteriorly in this groove and then in the carotid canal within which it divides into the cerebral and palatine vessels (Fig. 2). The distribution of the cerebral artery will be described (see p. 216).
1.rn.i. P r.
ch
FIG. 3. T.S. Head of Hypogeophis rostratus in the region of the ophthalmic ganglion. Abbreviations see p. 227.
2B(a). Palatine artery The palatine artery leaves the carotid canal and runs along a ventrolateral shelf on the
0s basale, below the trigeminal and is accompanied by the vena palatina and a mesial

CAECILIAN VASCULAR SYSTEM 205
branch of the palatine nerve. At the level of the posterior limit of the internal nostril the palatine artery curves anteromesially around the rim of the choana and together with the palatine vein and the mesial ramus palantinus enters a canal in the vomer (Fig. 4). As it runs through the vomerine canal the palatine artery gives rise to the posterior nasal
FIG. 4. T.S. Head of Hypogeophis rostrutus in the region of the tentacle. Abbreviations see p. 227.
artery which supplies Jacobson’s organ and the nasal epithelium. The palatine artery leaves its canal in the vomer and then enters a similar canal in the naso-premaxilla. At the level of the posterior limit of the tooth row on the naso-premaxilla the palatine artery curves mesially (still enclosed within the naso-premaxilla) and in the midline unites with that from the other side to form a vessel which supplies the skin on the tip of the snout (Fig. 2). Throughout its course from the carotid canal to the tip of the snout each palatine artery gives rise to a number of blood vessels which supply the dental tissues and the mucous membrane covering the palate. In addition to the posterior nasal artery a branch of each palatine vessel supplies the anterior nasal epithelium.
3. THE EXTERNAL CAROTID ARTERY is the second major division of the common carotid and carries blood to some of the cranial muscles, skin and the tongue. It has several branches. 3A. The muscular ramus leaves the main trunk of the external carotid and it supplies the m. rectus cervicis and interhyoideus posterior muscle.

206 R. LAWSON
3B. The laryngeal artery curves caudally and mesially above the rectus cervicis muscle to supply the laryngeal muscles. 3C. The lingual arteries carry blood to the tongue and the floor of the mouth. The most anterior of the lingual arteries runs laterally through the rectus cervicis muscle and then runs anteriorly to supply the m. ceratohyoideus internus. It then courses dorsally through the fibres of the m. ceratohyoideus and eventually breaks up into a large number of small vessels which form a plexus among the fibres of the genioglossus and geniohyoideus muscles. The second lingual artery forms a plexus above the first ceratobranchial and the third lingual artery forms a similar plexus above the second ceratobranchial (Fig. 2).
The second lingual artery also supplies the m. subarcualis rectus I1 and 111 (Lawson, 1965). The fourth lingual artery forms an extensive plexus which starts above the third ceratobranchial and extends posteriorly to supply the floor of the buccal cavity and the pharynx. These arterial plexuses are connected by many small blood vessels which run above the ceratobranchials and there is, therefore, an almost continuous arterial network in the tongue, buccal cavity floor and the pharyngeal floor. 3D. The internal mandibular artery is the largest branch of the external carotid. It curves around the lateral border of the thyroid gland and then runs anteriorly with the lateral ramus of the first spinal nerve. The artery extends anteriorly to the level of the ceratohyal and then turns abruptly dorsally through the fibres of the m. genioglossus. It then curves medially to fuse with the corresponding artery from the opposite side. The resulting vessel soon divides into a number of small arteries supplying the genioglossus and genio- hyoideus muscles and also the tissue overlying the mental symphysis. As the main stem of the internal mandibular artery curves around the thyroid gland it produces two small branches. The first supplies the m. rectus cervicis and the more posterior fibres of the m. intermandibularis. The second branch is an extremely short vessel which runs directly to the thyroid gland (Fig. 2).
4. BRANCHES OF THE SYSTEMICO-PULMONARY ARCH
4A. The anterior oesophageal artery is a large vessel which runs anteriorly over the ventral surface of the oesophagus until it reaches the laryngeal region. During its course this vessel supplies the ventral wall of the oesophagus via a large series of small arteries which form an extensive oesophageal plexus. As indicated above the extreme anterior end of the oesophagus is supplied with a further oesophageal artery which is a branch of the common carotid. 4B. The middle oesophageal artery arises from the systemico-pulmonary arch immediately posterior to the anterior oesophageal artery and supplies the mid-portion of the oesophagus (Fig. 1). 4C. Theposterior oesophageal artery supplies the lower portion of the oesophagus and also the body musculature in this region as it subdivides to form a series of vertebral arteries (Fig. 1). 4D. The vertebral arteries arise from the systemico-pulmonary arch as it passes above the heart. There are usually two but there may be three or four (Fig. 1). These arteries run dorsally and medially where they break up and supply the vertebral and body musculature (p. 21 5).

CAECILIAN VASCULAR SYSTEM 207
5. THE PULMONARY ARTERY stems from the systemico-pulmonary arch and runs posteriorly and medially until it reaches the anterior limit of the lungs where it gives rise to a small artery to the diminutive left lung. The main branch of the pulmonary artery curves ventrally over the surface of the right lung and forms a rich pulmonary plexus (Fig. 1).
The single systemic arch is continued as the dorsal aorta which lies mid-dorsally adjacent to the renal vein (Fig. 5). As the aorta progresses posteriorly it has a number of branches which may be conveniently described in terms of their distribution.
6. VESSELS OF THE ALIMENTARY CANAL
6A. The anterior gastric artery leaves the dorsal aorta at the level of the posterior limit of the right lung and supplies the proximal portion of the stomach and the gastro-oeso- phageal junction. 6B. The anterior median gastric artery leaves the dorsal aorta just posterior to the anterior gastric artery and a posterior median gastric artery is a branch of the coeliaco-mesenteric complex. 6C. Coeliaco-mesenteric complex leaves the dorsal aorta at the level of the pancreas and turns anteriorly as a large trunk which soon subdivides into a number of smaller vessels. The first of these is the posterior median gastric artery (see above). 6C(a). The lieno-gastric artery divides into a posterior gastric artery, which supplies the pylorus, and a splenic artery. 6C(b). The duodeno-pancreatic artery runs across the face of the pancreas and then sub- divides into a pancreatic artery and a duodenal artery. 6C(c). The anterior mesenteric artery leaves the dorsal aorta as it reaches the posterior end of the pancreas and runs directly to the distal portion of the duodenum. 6C(d). Mesenferic arteries.
There are usually three mesenteric arteries which run ventrally and supply the gut (Fig. 5).
6C(e). The posterior mesenteric artery supplies the caudal portion of the intestine (Fig. 5) . 6D. The rectal artery arises from the dorsal aorta at the posterior limit of the kidney and supplies the rectum (Fig. 5).
7. VESSELS OF THE URINO-GENITAL SYSTEM
7A. Gonadial arteries.
which leave the dorsal aorta (Fig. 5). 7B. The arteries supplying the fat bodies are branches of the gonadial arteries (Fig, 5). 7C. The renal arteries (usually four or five) leave the dorsal aorta as it runs between the kidneys (Fig. 5). 7D. The oviducul arteries leave the dorsal aorta anteriorly and supply the oviducal funnel and the oviduct.
The gonads (in both male and female) are supplied by a series of vessels (usuaHy seven)
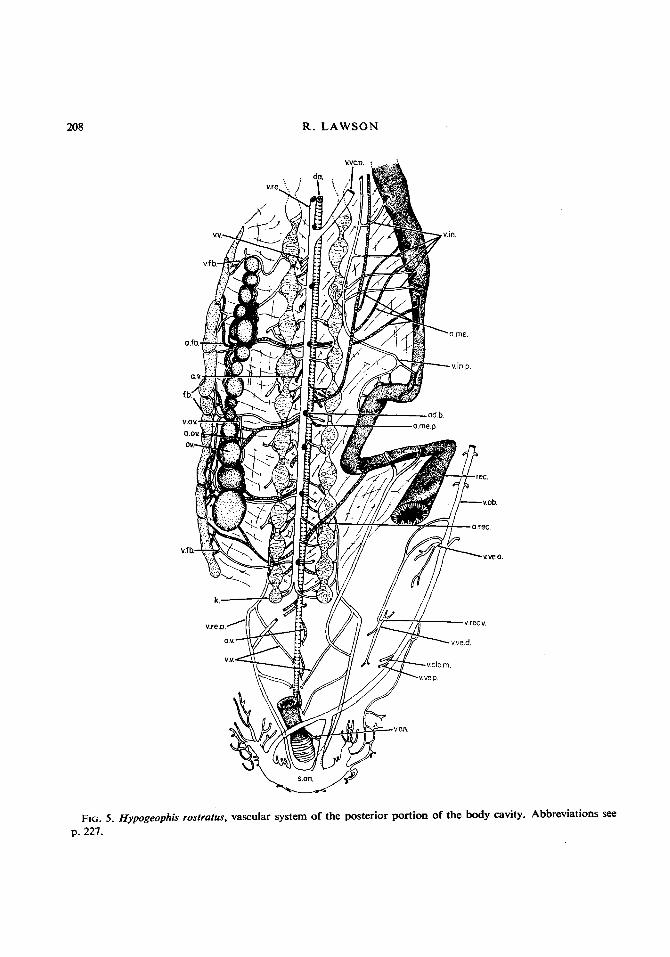
208 R. LAWSON
FIG. 5. p. 227.
Hypogeophis ' rostrutus, vascular system of the posterior portion of the body cavity. Abbreviations see

CAECILIAN VASCULAR SYSTEM 21 1
thus formed, which is then joined by the lateral jugular vein and the resulting vessel, is the internal jugular vein. The internal jugular runs caudally, accompanied by the ramus intestino-accessorius of the vagus; it then curves laterally through the fibres of the m. cephalo-dorso-subpharyngeus to unite with the external jugular.
10. THE EXTERNAL JUGULAR vEiN has complex origins and is formed by the fusion of a large number of subsidiary vessels (Figs 1 and 2). 1OA. The common facial vein carries blood from the superficial intermandibular region, the thymus, m. interhyoideus and the skin. It has its origin in distinct divisions (Fig. 2). 10A(a). The internal mandibular vein arises among the fibres of the m. genioglossus as an anastomosis with a corresponding vessel from the other side of the animal. It runs poster- iorly in the connective tissue on the ventral side of the lower jaw and receives numerous large factors from the intermandibular muscle and the venous plexus within the lower jaw. At the posterior end of the m. intermandibularis the internal mandibular vein opens into a large sinus (Fig. 2). In addition to the internal mandibular vein this sinus is formed from the external mandibular vein, deep mandibular vein and the lingual vein. The internal mandibular vein is then continued dorsally and joins the post-orbital sinus (Fig. 2). Posteriorly the large mandibular sinus narrows slightly and corresponds to the inferior facial vein. This expanded vessel collects blood from the ventral portion of the m. inter- hyoideus and the skin. It then joins the superior facial vein (Fig. 2). 10A(b). The superior facial vein collects blood from a series of small vessels which drain the posterior fibres of the m. cephalo-dorso-subpharyngeus and the vertebral musculature. The superior facial vein soon divides into two branches. The first branch turns ventrally and collects blood from the interhyoideus and interhyoideus posterior muscles before it joins the common facial vein. The second branch becomes closely applied to the skin, curves anteriorly and joins the inferior facial vein immediately behind the sinus of the internal mandibular vein. The vessel thus formed is the common facial vein which runs through the fibres of the m. interhyoideus and collects blood from the m. interhyoideus, m. interhyoideus posterior and m. cephalo-dorso-subpharyngeus. The common facial vein then continues caudally, collects blood from the thymus, runs below the ventral rami of the first two spinal nerves and is then joined by the ventral branch of the superior facial vein. The venous trunk thus formed extends caudally as the external jugular vein which is almost immediately joined by the internal jugular and the large vessel thus formed curves medially as the common jugular vein (Fig. 2).
1 1. THE COMMON JUGULAR VEIN, formed by the union of the internal and external jugular veins, runs caudally at the lateral edge of the oesophagus and is accompanied by the intestinal ramus of the vagus nerve. Anteriorly the common jugular receives two branches. 11A. The laryngeal vein collects blood from the laryngeal muscles, the fibres of the m. rectus cervicis, runs above the common carotid artery and then joins the common jugular (Fig. 1). 1lB. The oesophageal vein drains the anterior limit of the oesophagus and then joins the laryngeal vein as it enters the common jugular trunk.

212 R. LAWSON
The right and left common jugular veins run caudally to open into the sinus venosus. The left common jugular is joined by three vessels before it reaches the level of the sinus venosus. 11C. The muscu/ar r’amus originates at the level of the ventral occipital musculature among the posterior fibres of the m. interhyoideus posterior and the anterior fibres of the m. rectus abdominis. It then runs caudally below the m. transversus, is joined by a number of vessels draining the body musculature and joins the left common jugular at the level of the apex of the right atrium (Fig. 1). 11D. The anterior cutaneous vein arises in the ventral region of the pharynx and runs caudally between the common carotid arteries. This vein collects blood from the ventral portion of the skin anterior to the heart and receives a large number of smaller cutaneous veins (Fig. 1).
1 1 E. Anterior epigastric vein Superficial examination suggests that the anterior epigastric vein is the direct forward continuation of the abdominal vein (Fig. 1). However, careful inspection shows that blood from the skin in the region of the liver is collected by a series of veins and the anterior epigastric is the most anterior of the vessels of this group. It originates in the superficial tissues at the anterior level of the liver and collects blood from the skin and body muscles and then runs cranially. The anterior epigastric vein has a small branch which enters the anterior end of the liver and the remaining portion of the vessel curves anterolaterally to join either the anterior cutaneous vein or the left common jugular.
The right common jugular trunk is also joined by three subsidiary vessels before it enters the sinus venosus. 11F. The muscuZar ramus of this vessel has essentially the same distribution as that of the left common jugular. 1lG. The azygos vein joins the right common jugular at the same level as the muscular ramus and is formed by the junction of anterior and posterior azygos veins (Fig. 1). 1 lG(a). The anterior azygos vein collects the major portion of the blood from the vertebral region anterior to the heart and also drains the inner fibres of the anterior portion of the m. rectus abdominis. The muscular elements of this system originate among the fibres of the muscles in the pharyngeal region (Fig. 1) and the vessel thus formed runs caudally to join a vertebral ramus. The vertebral ramus of the anterior azygos system consists of a longitudinal vessel which runs caudally and collects blood from a number of vertebral veins. These drain the vertebral musculature and are also in communication with the spinal veins. 1 lG(b). The posterior azygos vein lacks a muscular ramus (similar to that of the anterior azygos) and consists only of the vertebral ramus which collects blood from the vertebral musculature posterior to the heart. 11H. The rend vein is an anterior continuation of the post-caval system (see below) and runs between the kidneys. In the region of its origin from the post-caval the renal vein receives blood from several vertebral veins (Fig. 5). As it extends anteriorly it receives many factors carrying blood from kidney and hence also from vertebral musculature. The renal vein opens into the base of the right common jugular.

CAECILIAN VASCULAR SYSTEM 21 3
12. THE POSTERIOR VENA CAVA (POST-CAVAL VEIN), arises at the caudal end of the kidneys by the fusion of a large vessel from each kidney and continues cranially between the kidneys, ventral to the dorsal aorta, until it reaches a point just behind the origin of the anterior mesenteric artery (Fig. 5). The posterior vena cava then curves ventrally to enter the liver and then runs forward along its ventral edge to leave its anterior end as a large hepatic vein, which opens into the sinus venosus principale. As the post-caval vein runs between the kidneys it receives blood from the kidney by a number of short renal venules. It also collects blood from several small vessels which drain the gonads and the fat bodies (Fig. 5). Anteriorly the renal vein (an extension of the post-caval) continues to receive blood from the kidney and also from the oviducal funnel in the female (Fig. 5) . The renal vein and the hind portion of the posterior vena cava therefore receive blood from the kidney, gonads and fat body. Posteriorly blood is carried to the kidney by the renal portal veins and more anteriorly by a number of vertebral veins draining the body musculature (see below).
13. ABDOMINAL VEIN
When the skin is removed from the ventral side of the anal region of Hypogeophis a huge sinus is reveaIed. In addition there is a large subcutaneous venous plexus on either side of the anus. The vessels of this anal plexus ramify among the cutaneous and muscular tissues and drain into the anal sinus. The anal sinus is also in direct communication with the spinal veins. The abdominal vein arises by the fusion of two veins, which originate at the anterior limit of the anal sinus, and then continues anteriorly in the mid-ventral line immediately below the skin. It collects blood from a large number of cutaneous veins (see below) and then opens into the posterior end of the liver (Fig. 5). 13A. The anal veins drain the anal sphincter muscle (Fig. 5). 13B. The posterior vesical vein collects blood from a rich vascular network on the surface of the posterior lobe of the bladder (Fig. 5). 13C. The median cloacal vein arises from the medial region of the cloaca as several fine vessels, which unite into a single vein, to lead cranially and join the abdominal vein immediately in front of the posterior vesical vein. The posterior abdominal vein also collects blood from the skin and anastomoses with the terminal vessels of the left side of the anal plexus (Fig. 5). 13D. The vesico-haemorrhoidul vein enters the abdominal vein at the level of the anterior end of the bladder and is formed by the union of the dorsal vesical and the ventral rectal veins (Fig. 5). 13D(a). The dorsal vesical vein is formed from several factors draining the dorsal wall of the bladder (Fig. 5). 13D(b). The ventral rectal vein lies ventral to the gut but dorsal to the bladder and collects blood from the ventral wall of the rectum before joining the dorsal vesical vein to form the vesico-haemorrhoidal vein (Fig. 5). 13E. The anterior vesical vein collects blood from the anterior lobe of the bladder and joins the abdominal vein immediately posterior to the vesico-haemorrhoidal vein (Fig. 5).
After collecting blood from the bladder and the cloaca the abdominal vein continues cranialjy, in the mid-ventral line, to open into the Iiver. However, between the anterior

214 R. LAWSON
end of the bladder and the liver the abdominal vein receives numerous segmental vessels carrying blood from the skin and it also communicates with the ventral longitudinal vessel draining the rectus abdominis muscle (see p. 215).
14. THE MEDIAN EPIGASTRIC VEINS can only be located with careful examination since superficially they seem to be a forward continuation of the abdominal vein. They do however, form a short series of three veins which collect blood from the skin and m. rectus abdominis and run directly to the liver.
The fourth member of this series is continued cranially as the anterior epigastric vein (see common jugular vein).
15. THE HEPATIC PORTAL SYSTEM is composed of an extensive series of vessels draining the gut. 15A. The posterior intestinal vein arises from several branches draining the posterior portion of the intestine and the intestino-rectal junction. These small vessels open into a longitudinal vessel (hepatic portal) which runs anteriorly suspended in the dorsal ligament (Fig. 5). 15B. The omphalo-mesenteric vein arises from a number of factors in the wall of the duo- denum and the anterior part of the intestine. It passes medial to the pancreas, to be joined by a pancreatic vein and opens immediately into the hepatic portal vein. 15C. The Zieno-gastric vein runs caudally through the gastro-splenic ligament, and collects blood from the stomach, spleen and the pancreas. 15D. The posterior gastric vein drains the pylorus and enters the hepatic portal vein just anterior to the lieno-gastric vessel.
After its junction with the posterior gastric vein the hepatic portal vein runs directly to the liver without receiving further tributaries.
16. THE ANTERIOR GASTRIC VEINS carry blood from the anterior portion of the stomach directly to the liver.
17. THE RENAL PORTAL VEINS originate from the anal sinus and run cranially above the cloaca and rectum. Each renal portal collects blood from the vertebral muscle via two large vertebral vessels. Before they reach the kidneys each renal portal vein is joined by a haemorrhoidal vessel from the posterior part of the intestine (Fig. 5).
18. THE PULMONARY VEIN arises from the dorso-medial aspect of the right lung and then runs anteriorly and then transversely across the proximal portion of the lung. At this level it is joined by a small vessel draining the rudimentary left lung and the vessel thus formed runs forward to enter the right atrium.
Vessels of the vertebral and body musculature The complex body musculature of Hypogeophis is supplied with blood by an extensive
system of vertebral arteries which leave the common carotids, oesophageal arteries and the dorsal aorta. These arteries arise separately and although there are many anastomoses

CAECILIAN VASCULAR SYSTEM 215
between successive vessels there is no large longitudinal vertebral artery. Each vertebral artery runs dorsally, penetrates the m. intertransversarius and continues until the level of the lower head of the rib is reached. At this point the vertebral artery divides into dorsal and ventral branches. The dorsal portion runs over the lower head of the rib and takes up a
FIG. 6 . Hypogeophis rosfratus, diagram of typical body segment showing the main venous drainage channels. Abbreviations see p. 227.
position very close to the lateral border of the segmental ganglion. The artery then con- tinues dorsally after giving rise to a small vessel which supplies the m. intertransversarius, until the dorso-lateral limit of the ganglion is reached. It then divides into a medial and a lateral branch. The medial division accompanies the medial branch of the dorsal ramus of the spinal nerve and supplies the m. dorsalis trunci and the skin. The lateral branch

216 R. LAWSON
accompanies the ramus dorsalis lateralis of the spinal nerve and supplies the m. cutaneous dorsalis and the skin.
The ventral division of the vertebral artery accompanies the ventral ramus of the spinal nerve, runs through the fibres of the m. intertransversarius and m. transversus and then curves dorsolaterally to reach the region of the centrum tendineum. At this point it divides into dorsal, medial and lateral branches. The dorsal branch accompanies the ramus cutaneous lateralis of the spinal nerve and supplies the m. obliquus abdominis and the skin. The medial vessel lies close to the ventral muscular ramus of the spinal nerve and supplies the superficial fibres of the m. rectus abdominis, and the skin. It also runs to the inner fibres of the m. rectus abdominis, the m. obliquus abdominis internus and the m. obliquus abdominis internus piofundus.
Blood is returned from the m. dorsalis trunci by an extensive series of segmental vessels which open into the circumspinal veins. Before piercing the intervertebral membrane these vessels are joined by small veins draining the interspinal muscles (Fig. 6). The cir- cumspinal veins communicate with the longitudinal spinal veins which carry blood from the vertebral canal and join the main venous channels such as the renal vein, posterior vena cava or the anterior caval system (see above).
Blood from the musculature of the remainder of the body wall largely drains into three longitudinal vessels (Fig. 6). No attempt has been made to homologize these vessels with the cutaneous veins described in urodeles (Francis, 1934) since in caecilians the cutaneous drainage system is much more complex. The longitudinal vessels are therefore simply termed the dorsal, median and ventral longitudinal veins. The dorsal longitudinal veins are situated at the apex of the m. rectus abdominis and collect blood from the m. cutaneous dorsalis (Fig. 6). This blood is carried in each dorsal longitudinal vessel and then enters a series of veins which run below the m. rectus abdominis until they reach the centrum tendineum. At this point they are joined by a vessel draining the m. transversus. The composite vessels thus formed curve ventrally and receive veins draining the m. obliquus abdominis internus and the m. obliquus internus profundus. In addition they receive a large tributary from the ventral longitudinal vein and open into the abdominal vein or the anterior cutaneous veins (see above).
The median longitudinal veins collect blood from the m. obliquus externus by a series of dorsoventral vessels. The median longitudinal vessels run superficially at the ventral limit of the m. obliquus abdominis externus and open into the ventral longitudinal veins by a series of segmental vessels (Fig. 6).
The ventral longitudinal vessels also collect blood from the m. rectus abdominis and the skin and then join the segmental branches of the median longitudinal veins as indica- ted below.
Blood from the skin is collected by an elaborate series of small veins which open into one of the longitudinal veins, directly into the abdominal vein or an epigastric vein. Anterior to the heart the cutaneous drainage is via the anterior caval system or the median anterior cutaneous vein (Fig. 1).
Vessels of the brain and spinal cord 19. THE CEREBRAL ARTERY leaves the main trunk of the palato-cerebral vessel within the carotid canal, enters the cranial cavity and divides into an anterior and a posterior ramus (Fig. 2).

CAECILIAN VASCULAR SYSTEM 217
19A. The posterior ramus (posterior cerebral artery) runs medially and then subdivides. The anterior subdivision of the posterior cerebral artery supplies the ventrolateral border of the mid brain. A more posterior branch forms a ramus pituitarius and a branch which unites with a similar artery from the opposite side to form the basilar artery (Fig. 2). 19B. The anterior ramus (anterior cerebral artery) supplies the brain, especially in the region of the telencephalon (Fig. 2).
20. THE SAGITTAL SINUS arises between the anterior ends of the olfactory lobes and runs posteriorly in the mid dorsal line to the nodus chorioideus which forms a triangular plexus and also collects blood from the dorsal and lateral regions of the fore brain by the lateral cerebral veins (Fig. 2).
2 1. OBLIQUE CRANIAL VEINS (VENAE CRANIALES OBLIQUAE) Blood is carried from the nodus chorioideus by a single median vessel which runs
directly posteriorly and opens into the chorioid plexus (Fig. 2). This consists of a large network in the region of the hemispheres and a posterior plexus which covers the roof of the mesencephalon and the fossa rhomboideus. The anterior and posterior divisions of the chorioid plexus are connected by a pair of dorsal longitudinal vessels (venae craniales obliquae) which in turn open on each side into a large post-otic cranial vein which drains into the post-orbital sinus (Fig. 2). Blood from the posterior chorioid plexus leaves the cranial cavity by two routes.
22. POSTERIOR CEREBRAL VEIN (VENA CRANIALIS POST OTICA) carries blood from the postero- medial border of the chorioid plexus and curves ventrally, issues from the skull, and joins the vena capitis lateralis (Figs 2 and 8).
23. THE ANTERIOR CIRCUMSPINAL VEIN collects blood from the posterior chorioid plexus, and lies between the anterior limit of the atlas and the edge of the foramen magnum. It communicates ventrally with the posterior cerebral veins and the spinal vein. Most of the blood from the anterior circumspinal vein is passed to the posterior cerebral vein and the spinal vein, but in addition blood is also passed to the second circumspinal vein by a median longitudinal vessel (Figs 2 and 6).
24. THE ANTERIOR CEREBRAL VEIN (VENA CRANIALIS PRE-OTICA) drains the anterior portion of the cranial cavity and opens into the post-orbital sinus (Fig. 2).
25. DRAINAGE OF POST ORBITAL SINUS Blood from the post-orbital sinus is carried by two veins, the vena capitis lateralis
(lateral head vein) and a vein which runs in a distinct groove in the floor of the cranial cavity and probably represents the embryonic vena capitis medialis (see below). This vessel collects blood from a short pituitary vein and opens into the posterior cerebral vein at the level of the roots of the IXth and Xth cranial nerves. After this junction the posterior cerebral vein divides into medial and lateral vessels (Fig. 2). The median vessel thus formed divides almost immediately into the right and left spinal veins which run in the vertebral canal, below the spinal cord, for the entire length of the animal.

21 8 R . LAWSON
26. CIRCUMSPINAL VEINS
The longitudinal spinal veins are connected intersegmentally by a series of circumspinal veins which collect blood from the body wall (see p. 215) and from the spinal cord itself (Figs 2 and 6).
CUTANEOUS LYMPHATIC SYSTEM
Injection difficulties made a complete study of the lymphatic system impossible, but some detail of the superficial lymphatics could be seen in sectioned material. The cutaneous lymphatic vessels form a rich reticulum with irregular meshes which open into a main longitudinal lymphatic trunk (truncus Iymphaticus longitudinalis Iateralis (Francis, 1934)) which runs along the dorsomedial edge of the m. rectus abdominis on either side of the animal. The longitudinal lymph vessel receives numerous lateral tributaries and is in communication with a series of intersegmental lymph hearts. The lymph hearts are con- tractile sacs which are composed of an endothelial layer within a muscular coat and they force fluid into the dorsal longitudinal veins. Any regress of blood is prevented by simple valves. The longitudinal lymphatic trunk also communicates with a subvertebral lymphatic system by a series of transverse intersegmental lymph vessels.
Discussion Extant caecilians may be divided into two groups on the basis of geographical distribu-
tion.
1. NEW WORLD GENERA Caecilia, Chthonerpeton, Gymnopis, Rhinatrema, Siphonops and Typhlonectes (Gorham,
1962).
2. OLD WORLD GENERA
Boulengerula, Gegeneophis, Geotrypetes, Herpele, Hypogeophis, Ichthyophis, Idiocranium Indotyphlus, Praslinia, Schistometopum, Scolecomorphus, and Uraeotyphlus (Gorham, 1962).
The list of caecilian genera presented by Gorham must now be extended to include those described by Taylor (1967) who has erected 13 new genera and re-established three others. The additional genera are :
NEW WORLD
Brasioltyphlus, Cryptopsophis, Copeotyphlinus, Dermophis, Epicrionops, Luetkeno- typhlus, Microcaecilia, Mimosiphonops, Nectocaecilia, Oscaecilia, Parvicaecilib, Pota- ~ o t y p h l ~ and Pseudosiphonops.
OLD WORLD Afrocaecilia, Caudacaecilia and Grandisonia. Unfortunately information on the vascular system is not available for all the genera
listed by Gorham (and for none of those erected by Taylor) but it is clear from the present study and from the literature that there is considerable variation in morphology of the major arteries. In Caecilia the long truncus arteriosus gives rise to a single common

CAECILIAN VASCULAR SYSTEM 219
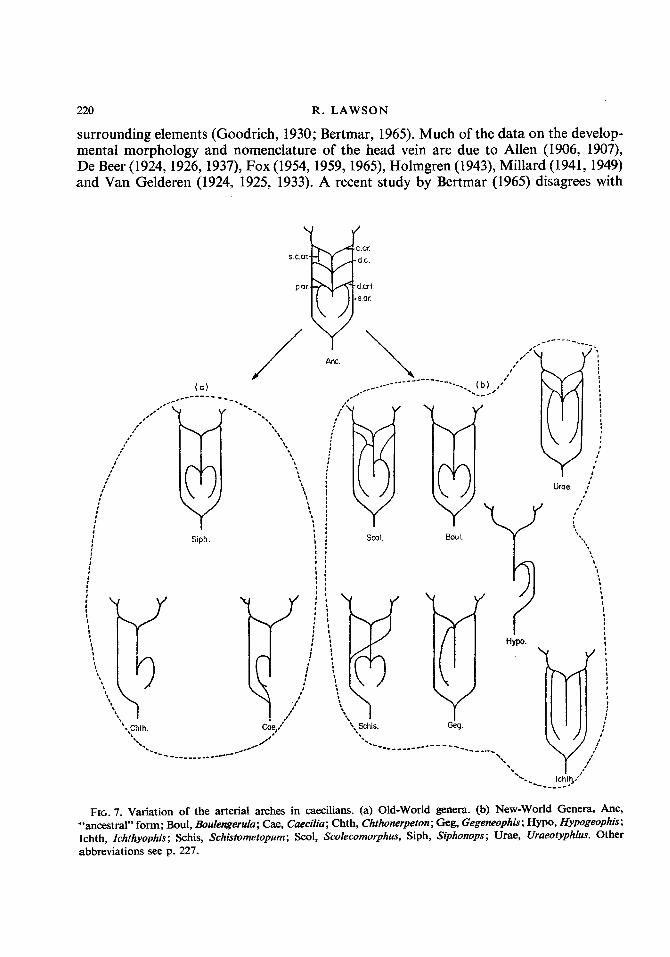
carotid vessel which divides anteriorly into left and right carotid arches. A single (right) systemic arch is present and this fuses with a posterior extension of the pulmonary artery (Ramaswami, 1944). Siphonops has a fairly symmetrical arrangement except that the right pulmonary artery is larger than the left (Ramaswami, 1944). Chthonerpeton lacks the left systemic and right pulmonary arches. In Schistometopum the two systemic arches meet in front of the heart as the left systemic is strongly reflected to the right (Fig. 7). The arterial arches are more or less symmetrically arranged in Scolecomorphus and Boulengerula. However, in the former the systemic and pulmonary arches are connected by a ductus arteriosus and the right pulmonary arch arises more anteriorly than the left (Ramaswami, 1944). In Gegeneophis the right systemic arch is absent (Ramaswami, 1944). Uraeotyphlus has a symmetrical arrangement and has a right and left ductus arteriosus. In Ichthyophis the common arterial trunk leading from the heart is very long and the pulmonary arteries arise far anteriorly. The arterial system of Hypogeophis differs considerably from that of the other genera in having a single systemico-pulmonary arch. Previous accounts of the system in Hypogeophis by Marcus (1935), Schilling (1935) and Ramaswami (1944) describe the presence of a lateral dorsal aorta (ductus caroticus) which runs from the carotid arch to the systemico-pulmonary arch. The author has been unable to find this vessel (see Lawson, 1966 for full discussion). The artery which corresponds most closely to the posi- tion of the lateral dorsal aorta is an oesophageal artery. Marcus (1935) has also suggested that the systemico-pulmonary arch is the left one. If this is the case then Hypogeophis differs from all other known genera since in other forms it is the right arch which is retained.
It seems likely that the least specialized arrangement of the arterial arches would be a symmetrical one which retained a right and left ductus caroticus. Variation of this basic pattern has occurred in both geographical groups of caecilians. It is generally agreed that Old World and New World caecilians each consist of a number of related genera, but that the members of one geographical group are not closely related to the members of the other. (See Parker, 1956, 1958; Parker & Dunn, 1964; Wake, 1967.) The structure of the vascular system suggests that this might well be the case and that within the two groups varying degrees of asymmetry have been evolved in parallel (Fig. 7).
The cephalic circulation in caecilians is elaborate and this is partly the result of the complex network in the vascular membranes of the buccal cavity. The main blood supply to the head of Hypogeophis is the internal carotid artery, a major branch of which forms a stapedial artery which is closely associated with the maxillary branch of the trigeminus nerve. A subdivision of the stapedial artery, the mandibular artery, supplies the lower jaw. The remainder of the internal carotid is the palato-cerebral artery which forms the cerebral artery to the brain and the palatine artery which is associated with the palatine vein and nerve and supplies the palatal tissues. Unfortunately there is no complete account of the cranial arteries in other genera and caecilians but De Villiers (1938) comments on the existence of a stapedial artery supplying the branches of the trigeminal nerve and an internal carotid (palato-cerebral) artery accompanying the ramus palatinus VII in Scole- comorphus. De Jager (1938, 1939) describes a similar pattern in Gymnopis mexicana (= Dermophis mexicanus of De Jager) and also comments on the presence of a mandibular branch of the internal carotid.
The morphology of the head veins in the tetrapods has been considered to be important in establishing affinities since they are said to retain a relatively constant relationship with
16
220 R. LAWSON
surrounding elements (Goodrich, 1930; Bertmar, 1965). Much of the data on the develop- mental morphology and nomenclature of the head vein are due to Allen (1906, 1907), De Beer (1924, 1926, 1937), Fox (1954, 1959, 1965), Holmgren (1943), Millard (1941, 1949) and Van Gelderen (1924, 1925, 1933). A recent study by Bertmar (1965) disagrees with
I , v Geg.
Hypo. 7, FIG. 7. Variation of the arterial arches in caecilians. (a) Old-world genera. (b) New-World Genera. Anc,
"ancestral" form; Boul, Boulengerula; Cae, Caecilia; Chth, Chthonerpeton; Geg, Gegeneophis; Hypo, Hypogeophis; Ichth, Zchthyophis; Schis, Schistometopum; Scol, Scolecomorphus, Siph, Siphonops; Urae, Uraeotyphlus. Other abbreviations see p. 227.

221
some of the findings of some of the previous authors. The main area of dissent concerns the contribution of the median head vein (vena capitis medialis) to the adult cephalic drainage andespecially the homologies of the median cerebral vein (vena cranialis medialis). In vertebrates the median head vein (vena capitis medialis) is usually the first vein to develop in the head during ontogeny and lies within the cranial cavity. Later it is super- seded to some degree by a lateral head vein (vena capitis lateralis) which is essentially extracranial. However, regardless of the details of the embryonic development, the head is eventually drained by an internal jugular system which is basically produced from four components.
CAECILIAN VASCULAR SYSTEM
(1) The remnants of the intracranial vena capitis medialis. (2) An extracranial vena capitis lateralis. (3) Interconnections between the two in the form of cerebral veins. (4) A component derived from the anterior end of the anterior cardinal vein. In Neoceratodus the vena capitis medialis persists as the anterior cerebral vein (vena
cerebralis anterior), infra-orbital vein, mandibular vein and pituitary vein (Bertmar, 1965). Bertmar also describes the presence of a median cerebral vein (vena cerebralis medialis) which communicates with the vena capitis lateralis. Posteriorly, a portion of the vena capitis medialis persists as the “vena capitis medialis proper” which drains the inner side of the rhombencephalon and the otic region. This vessel is connected to the posterior cerebral vein (vena cerebralis posterior) which is apparently considered by Bertmar to be part of the anterior cardinal. The development of the venous system of Neoceratodus has also been studied by Fox (1965) whose description largely agrees with that of Bertmar. Fox (1954) has also described the development of the head of Cryptobrunchus and has indicated that there are several major developmental differences between fishes and tetra- pods. The more important of these are:
(1) The infra-orbital vein in tetrapods is part of the vena capitis lateralis (rather than the vena capitis medialis as in fish).
(2) The vena capitis lateralis in tetrapods is short and fuses with the vena capitis medialis at the level of the otic process and the resulting vessel which runs lateral to the otic capsule is the “head vein”.
(3) The head vein in the tetrapods is joined by a posterior cerebral vein (or veins) which emerge from the skull with the IXth and Xth cranial nerves.
(4) The “vena capitis medialis proper” (sensu Bertmar) is absent in tetrapods. The “head vein” in Cryptobranchus (Fox, 1954) seems to be homologous with the
vessel described by De Beer (1926) and Bertmar (1965) as an anterior extension of the anterior cardinal which grows lateral to the cranium to establish a connection with the vena capitis medialis, and is termed the vena capitis lateralis. Francis (1934) also uses this term to describe the vessel.
Millard (1941, 1949) has produced a clear account of the development of the cranial vascular system in Xenopus luevis. In this form the internal jugular vein is also a compound structure which is derived from the vena capitis medialis, anterior cardinal vein and vena capitis lateralis. In a 5 mm embryo the major cranial vessel is the intra-cranial vena capitis medialis. This soon establishes contact with a vena capitis lateralis which is an anterior

222 R. LAWSON
continuation of the anterior cardinal. During ontogeny cross connections between the two form anterior, median and posterior cerebral veins. In the larval stages a pharyngeal vein is present which drains the thymus. At metamorphosis the pharyngeal vein is lost and the thymus is drained by the facial vein which empties into the vena capitis lateralis. The anterior cerebral vein (a derivative of the vena capitis medialis) no longer reaches the brain and drains the dorsal part of the orbit and persists as the ophthalmic vein. The posterior cerebral vein is lost. The adult cerebral return is therefore largely mediated via a median cerebral vein which issues from the cranium, through the foramen pro-oticum, to join the lateral head vein. The median cerebral vein has an anterior branch which drains the brain, a median branch from the pituitary and a posterior branch from the hind brain. Millard suggests that the median cerebral vein in the adult is not the homo- logue of the embryonic median cerebral vein (vena cerebralis medialis Vera) since it changes its position during ontogeny so that it emerges anteriorly and dorsally to the trigeminus. The anterior portion of the embryonic median head vein persists in Xenopus as the orbito-nasal vein. The posterior portion of the internal jugular (derived from the anterior cardinal) receives an occipital vein and a vertebral vein.
The cranial drainage of Leiopelmu hochstetteri is basically similar to that described for Xenopus luevis by Millard, and the main exit for blood is a large vena cerebralis medialis (Stephenson, 1951). Stephenson comments on the existence of a small posterior cerebral vein, in L. hochstetteri, and this issues from the jugular foramen to join the posterior portion of the lateral head vein. An occipital vein is also present. She points out that this primitive species is the only anuran known to possess a posterior cerebral vein. The presence of an anterior cerebral vein (vena cerebralis anterior) in the embryo of Leiopelmu urcheyi is also recorded by Stephenson, but this vessel disappears in the adult and its area of drainage is taken over by the median cerebral vein.
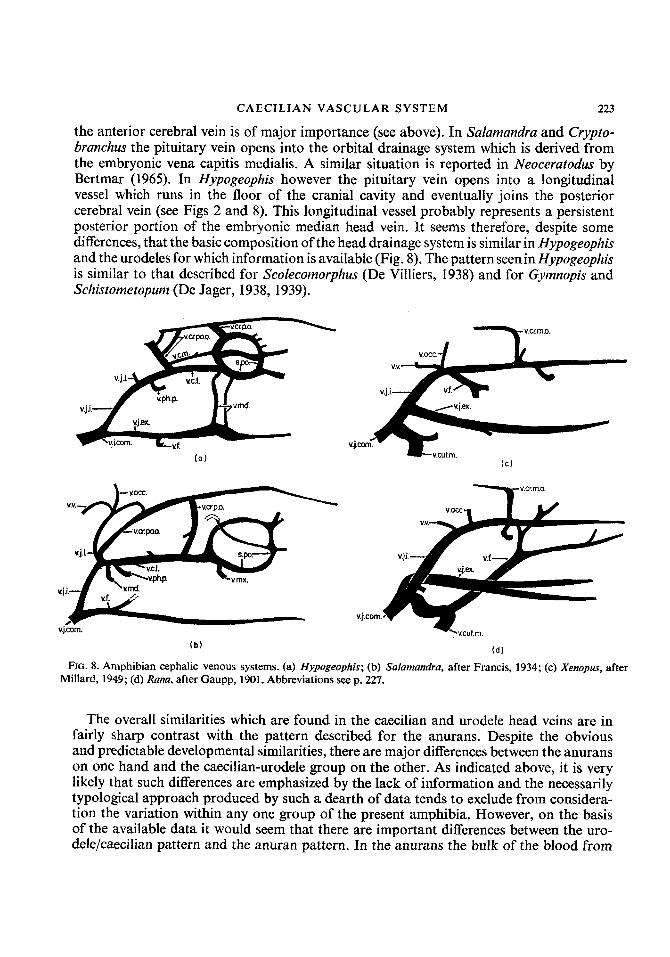
It is clear therefore that despite the obvious basic developmental similarities of the cephalic venous systems there are a number of ways in which the adult system has evolved. The morphology of the cephalic veins in Hypogeophis is largely similar to that described for Cryptobrunchus (Fox, 1954), Sulumundru (Francis, 1934) and Triturus (Rex, 1893). In all cases the vena capitis lateralis (head vein of Fox) is joined by a large factor from the palate (vena pharyngo-palatina) and a superficial lateral element, the lateral jugular vein. In Sulumundru the importance of the anterior cerebral vein is noted by Francis who describes it as the main exit for the blood from the cranium. A posterior cerebral vein is present in Sulumandra, Cryptobrunchus and Triturus and this issues from the cranium via the jugular foramen along with the IXth and Xth cranial nerves. A median cerebral vein is also present in Sulumundru and Triturus but apparently not in Cryptobrunchus. Posteriorly, the lateral jugular vein in Sulumundru receives a post-occipital cranial vein (vena cranialis post- occipitalis) which leaves the vertebral canal via a foramen in the first vertebra (Francis, 1934). In Hypogeophis the posterior cerebral vein drains directly into the vena capitis lateralis and it is much more extensive than in Sulumandra. Further, the posterior cerebral vein in Hypogeophis is in direct communication with the spinal veins and there is no vertebral vein (Fig. 2). In both caecilians and urodeles the posterior cerebral vein is retained in the adult (although the extent and importance of the vessel is variable) and in Cryptobrunchus Fox describes a pair of such vessels on either side of the head. In Triturus (Rex, 1893) the main avenue of cranial drainage seems to be the posterior cerebral vein and it is therefore rather closer to the caecilian pattern than that of Sulumundra in which
CAECILIAN VASCULAR SYSTEM 223
the anterior cerebral vein is of major importance (see above). In Sulumundra and Crypto- branchus the pituitary vein opens into the orbital drainage system which is derived from the embryonic vena capitis medialis. A similar situation is reported in Neocerutodus by Bertmar (1965). In Hypogeophis however the pituitary vein opens into a longitudinal vessel which runs in the floor of the cranial cavity and eventually joins the posterior cerebral vein (see Figs 2 and 8). This longitudinal vessel probably represents a persistent posterior portion of the embryonic median head vein. It seems therefore, despite some differences, that the basic composition of the head drainage system is similar in Hypogeophis and the urodeles for which information is available (Fig. 8). The pattern seenin Hypogeophis is similar to that described for Scolecomorphus (De Villiers, 1938) and for Gymnopis and Schistometopum (De Jager, 1938, 1939).
v.cr.p.0. v.upo.0.
v.j.1. uc.1.
.,ii L v.ph.p. V . M . - 1 Y ”.,.’.--- I
(0)
VOCC.
v.j.i.--/ v.f. . vjCom.
(b)
y v . c r r n . o .
FIG. 8. Amphibian cephalic venous systems. (a) Hypogeophis; (b) Sulumandru, after Francis, 1934; (c) Xenopus, after Millard, 1949; (d) Runu, after Gaupp, 1901. Abbreviations see p. 227.
The overall similarities which are found in the caecilian and urodele head veins are in fairly sharp contrast with the pattern described for the anurans. Despite the obvious and predictable developmental similarities, there are major differences between the anurans on one hand and the caecilian-urodele group on the other. As indicated above, it is very likely that such differences are emphasized by the lack of information and the necessarily typological approach produced by such a dearth of data tends to exclude from considera- tion the variation within any one group of the present amphibia. However, on the basis of the available data it would seem that there are important differences between the uro- dele/caecilian pattern and the anuran pattern. In the anurans the bulk of the blood from

224 R. LAWSON
the cranium leaves via the posterior ophthalmic vein which collects blood from the interior orbital, orbito-nasal and anterior orbital veins, and is derived from the embryonic median head vein. The lateral head vein in this group is therefore relatively small, but it is joined posteriorly by the post-occipital vein and a vertebral vein and therefore resembles the urodele rather than the caecilian. A major portion of the cephalic blood passes into the common facial vein and then into the vena cutana magna (Fig. 8). Considerable variation in the distribution of the facial vein occurs within the group and the facial vein may empty into the cutaneous vein (Ram esculenta, R. temporaria) cutaneous vein and the external jugular (Rana tigrina) or the external jugular only (Hyla aurea, Bufo melanostictus). Millard (1941, 1949) indicates that Xenopus resembles urodeles rather than other anurans, in that the anterior portion of the vena capitis lateralis is derived from the vena capitis medialis and lies within the trigeminus. In other anurans the anterior portion of the head vein develops outside the cranium and lateral to the trigeminus (Millard, 1949; Goodrich, 1930). The posterior portion of the head vein in Xenopus becomes incorporated into the wall of the otic region. Millard also emphasizes the fact that the median cerebral vein, which is the functional cerebral vessel in the adult, is not a true vena cerebralis medialis since it changes its position during development (see above). The facial vein in Xenopus is unlike that of most other anurans in that it opens directly into the internal jugular system at the point where the vena capitis lateralis joins the anterior cardinal. This situation is also found in Pipa pipa (Klinckowstrom, 1893), while in LRiopelma hochstetteri the facial vein and the vena palatina join thevenacapitis lateralis as it unites with the median cerebral vein (Stephenson, 1951).
The posterior venous system of caecilians is similar to that of other amphibians, but there is modification related to the body form, loss of limbs and limb girdles, the elongate, kidney and the large number of fat bodies. The abdominal vein in caecilians originates from the anal sinus via a pair of vessels which soon unite (Fig. 5). Chatterjee (1935) con- siders these to be homologous with the pelvic veins of other forms. The somatic venous drainage is effected in the caecilians by a series of epigastric and cutaneous veins and there is no large vena cutana magna. The posterior limit of the kidney is supplied by a renal portal vein, but anteriorly there is a series of renal arteries and a number of vessels from the body wall and gonads. The blood from the anterior end of the kidney is carried directly to the sinus venosus by an anterior continuation of the post-caval vein (Fig. 5 ) which I have termed the “renal vein”. Such a vein has also been reported in the genera described by Ramaswami (see above) and Hevpele (Chatterjee, 1935) is reported to have two renal veins. Although embryological data are not available it seems likely that the renal veins represent the persistent posterior cardinal(s) since they also drain the body wall and fat bodies (Fig. 5).
Although caecilians and urodeles rely, at least to some degree, on the skin for respiration they do not have a large cutaneous artery and a vena cutana magna. However, the cae- cilians and urodeles have extensive cutaneous circulation which is essentially segmental. It is likely that the segmental cutaneous circulation of caecialians and urodeles is related to the elongate form of the body in these groups and that the anuran system is associated with the extreme shortening of the body. A further similarity between caecilians and uro- deles, again related to body form, is the nature of the cutaneous lymphatic system and the large number of lymph hearts. This contrasts sharply with the large lymph spaces and small lymph sacs in the anurans.

CAECILIAN VASCULAR SYSTEM 225
The available data suggest that there are many similarities between the vascular systems of caecilians and urodeles. It is also clear that many of the features of these two groups differ quite markedly from the anurans. The assessment of the significance of such characters in the context of intra-amphibian relationships is difficult since it seems to the author that as information on a greater number of species becomes available the differences will tend to become less well marked. It may also be true that the similarities which exist between caecilians and the urodeles are due to the parallelism within the two groups. In view of the long history of the amphibia and the likelihood of the early divergence of the three extant groups it is entirely possible that parallel evolution would have occurred especially in two groups which have retained an elongate body form. However, in view of the similar nature of the vascular system, especially the cephalic portion, the author is inclined to think that the urodeles are more closely related to the caecilians than to the anurans, and that the anurans and the caecilians are widely separated. However, in view of the long time periods involved, the possibility of parallel evolution and the paucity of data the conclusion can only be tentative and does not obviate a single ancestor for the modern groups, nor indeed three separate ones.
Summary The heart in Hypogeophis has an extensive system of coronary veins which communicate
with a series of myocardial spaces. A single systemico-pulmonary arch is present and this gives rise to a series of oesophageal arteries and a pulmonary artery. More posteriorly the dorsal aorta has a series of branches to the viscera and a number of vertebral arteries. There are many anastomoses between the vertebral arteries but there is no large longi- tudinal vessel. The vertebral arteries supply the vertebral musculature and eventually the skin. Blood is returned from the skin by an extensive series of vessels which initially communicate with a system of longitudinal cutaneous veins. There is no large vena cutana magna. A segmental series of lymph hearts is present. A large venous sinus is found in the cloaca1 region and this communicates with the anterior abdominal vein. Paired carotid arteries supply the head and each internal carotid has a prominent palato- cerebral artery which divides into the anterior and posterior cerebral arteries. Each posterior cerebral artery has a ramus pituitarius and a branch which unites with a similar artery from the opposite side to form the basilar artery. The main venous return from the cranium is via an anterior cerebral vein (vena cranialis pro-otica) and a posterior cerebral vein (vena cranialis post-otica). The former drains into the postorbital sinus while the latter leaves the cranial cavity with the IXth and Xth cranial nerves via the jugular foramen. The posterior cerebral vein joins the lateral head vein (vena capitis lateralis), and it also communicates with an occipital vein (vena cranialis occipitalis). The occipital vein also drains into the anterior circumspinal vein. This vein is the most anterior of a segmental series of vessels which surround the spinal cord and which com- municate ventrally with the longitudinal spinal veins.
The general arrangement of the vascular system in Hypogeophis is similar to that of other genera of caecilians for which information is available. There is, however, a great deal of variation (in the disposition of the major arterial arches) within the group. It is evident that these variations have occurred in parallel in the two geographical groups of caecilians. i.e. the Old World forms and the New World forms.

226 R. LAWSON
A detailed examination of the cephalic venous system in the caecilians suggests that there are a number of similarities between them and the urodeles. Further, the caecilian/ urodele arrangement shows marked differences from that found in anurans. The anurans have no posterior cerebral vein whereas in other groups this is a very important vessel. The main cephalic drainage in the anurans is via a median cerebral vein which may open into the lateral head vein or into the facial vein. The facial vein and the external jugular system as a whole shows considerable variation within the anura and the facial vein may open into the internal jugular vein or in some cases into the vena cutana magna.
The structure of the cephalic system in caecilians and urodeles as well as the overall similarities in their vascular system suggests that either these similarities have been acquired by parallel evolution within the groups, or that they are in reality closely related to each other, but not to the anura.
I would like to thank Mr J. F. Guy Lionnet, Department of Agriculture, Seychelles who pro- vided most of the specimens of Hypogeophis. Additional material was kindly given by Lord Richard C. Percy, Department of Zoology, University of Newcastle on Tyne.
Mr M. J. Parr and Mr R. E. Merry, Department of Biology, University of Salford made many useful suggestions. My gratitude is also due to Dr D. B. Wake, Department of Anatomy, Uni- versity of Chicago; Professor M. Jollie, Department of Zoology, University of Northern Illinois; Dr H. Fox, Department of Zoology, University College of London and Dr H. J. de Jongh, University of Leiden; who were kind enough to read the manuscript prior to the publication.
Part of the research was carried out (in the Department of Anatomy, University of Chicago) during the tenure of a Fulbright Scholarship. I am especially indebted to Professor R. Singer, Chairman of Anatomy, for allowing me to work in his Department and to Dr D. B. Wake and Dr M. Wake for their kindness. My thanks are also due to Professor W. E. Kershaw and the Senate of the University of Salford and to Dr R. Koppleman, Master of College Biology, Uni- versity of Chicago.
R E F E R E N C E S
Allen, W. F. (1906). Distribution of the lymphatics in the head and in the dorsal, pectoral and ventral fins of
Allen, W. F. (1907). Distribution of the subcutaneous vessels in the head region of the ganoids, Polyodon and
Bertmar, G. (1965). On the development of the jugular and cerebral veins in fishes. Proc. 2001. SOC. Lond. 144:
Chatterjee, B. K. (1935). The anatomy of Uraeotyphlus menoni. Part I, Digestive, circulatory, respiratory and
De Beer, G. R. (1924). Studies on the vertebrate head. Part I. Q. JI microsc. Sci. 68: 287-341. De Beer, G. R. (1926). Studies on the vertebrate head. Part 11. Q. JI microsc. Sci. 70: 263-370. De Beer, G. R. (1937). The development of the vertebrate skufl. Oxford: The University Press. De Jager, E. F. (1938). A comparison of the cranial nerves and blood vessels of Dermophis mexicanus and Dermophis
De Jager, E. F. (1939). Contributions to the anatomy of the skull of Gymnophiona. Anat. Anz. 88: 193-222. De Villiers, C. G. S. (1938). A comparison of the cranial features of the East Africa Gymnophiones, Boulengerula
Fox, H. (1954). Development of the skull and its associated structures in Amphibia with special reference to the
Fox, H. (1959). A study of the development of the head and pharynx of the larval urodele Hynobius and its bearing
Fox, H. (1965). Early development of the head and pharynx of Neoceratodus with a consideration of its phylogeny.
Scorpaenichthys marmoratus. Proc. Wash. Acad. Sci. 8: 41-90.
Lepidosteus. Proc. Wash. Acad. Sci. 9: 79-125.
87-130.
urinogenital systems. Amt . Anz. 81: 393-414.
gregorii. Anat. Anz. 86: 321-347.
boulengeri and Scolecomorphus ulugurensis. Anat. Anz. 86: 1-26.
urodeles. Trans. zool. SOC. Lond. 28: 241-304.
on the evolution of the vertebrate head. Phil. Trans. R. SOC. (B) 242: 155-205.
J. Zool., Lond. 146: 470-554.

CAECILIAN VASCULAR SYSTEM 227
Francis, E. T. B. (1934). The anatomy ofthe salamander. Oxford: The University Press. Gaupp, E. (1 901). Anatomie des Frosches. Braunschweig. Gelderen, C. van (1924). Die morphologie der Sinus durae matris. Erster Teil (Ontogonie). 2. Anat. EntwGesch.
Gelderen, C. van (1925). Die morphologie der Sinus durae matris. Dritter Teil. Vergleichendes, Erganzendes,
Gelderen, C. van (1933). Gefassystem. In Handbuch der vergleichenden Anatomie der Wirbeltiere 6 : 685-774.
Goodrich, E. S. (1930). Studies on the structure and development of vertebrates. London: Macmillan. Gorham, S. W. (1962). Liste der rezenten Amphibien und Reptilien. Gymnophiona. Tierreich No. 78: 1-25. Holmgren, N. (1943). Studies of the head of fishes. Part 4. AcfaZooL, Sfockh. 24: 1-188. Holmgren, N. (1952). An embryological analysis of the mammalian carpus and its bearing upon the question of
Klinckowstrom, A. (1893). Beitrag zur Kenntnis des Parietalauges. 2001. Jb. 1: 249-280. Lawson, R. (1963). The anatomy of Hypogeophis rostratus (Amphibia, Apoda) Part 1, The skin and skeleton.
Lawson, R. (1965). The anatomy of Hypogeophis rosfrafus (Amphibia, Apoda) Part 11, The musculature. Proc.
Lawson, R. (1966). The anatomy of the heart of Hypogeophis rostratus (Amphibia, Apoda) and its possible mode of
Marcus, H. (1935). Zur stammensgeschichte des Herzens. Morph. Jb. 76: 92-103. Millard, N. (1941). Vascular anatomy of Xenopus laevis. Trans. R. SOC. S. Afr. 28: 387439. Millard, N. (1949). The development of the venous system of Xenopus laevis. Trans. R. SOC. S. Afr. 32: 55-99. Parker, H. W. (1956). Viviparous caecilians and amphibian phylogeny. Nature, Lo&. 178: 250. Parker, H. W. (1958). The caecilians of the Seychelles Islands with a description of a new subspecies. Copeia 1958:
Parker, H. W. & DUM, E. R. (1964). Dentitional metamorphosis in the Amphibia. Copeia 1964: 75-86. Parsons, T. S. & Williams, E. E. (1963). The relationships of the modem Amphibia. Q. Rev. Biol. 38: 26-53. Ramaswami, L. S. (1944). An account of the heart and associated structures in some genera of Apoda. Proc. zool.
Rex, H. (1893). Beitrag zur morphologie der Hirnvenen der Amphibien. Morph. Jb. 19: 295-311. Schilling, C. (1935). Das Herz von Hypogeophis und seine Entwicklung. Morph. Jb. 76: 52. 91. Schmalhausen, I. I. (1959). Concerning monophyletism and polyphyletism in relation to the problem of the origin
Stephenson, E. M. (1951). The anatomy of the head of the New Zealand frog Leiopelma. Trans. zool. SOC. Lo&.
Taylor, E. H. (1967). The caecilians ofthe world. Univ. Kansas Press. Wake. M. (1967). Gill structure in the caecilian genus Gymnopis. Bull. Sth Calif: Acad. Sci. 66: 109-115.
73: 541-605.
Phyletisches and Zusammenfassendes. 2. Anat. EntwGesch. 75: 525-596.
Bolk, L. et al. (ed.). Berlin & Vienna: Urban & Schwarzenberg.
the origin of the tetrapod limb. Acta Zool., Stockh. 33: 1-115.
Proc. Univ. Durham Phil. SOC. (A) 8: 254-273.
Univ. Newcastle Phil. SOC. (A) 1: 52-63.
action. J. Zool., Lmd. 149: 320-336.
71-76.
SOC. LO&. 114: 117-138.
of land vertebrates. Byull. mosk. Obshch. ZspjV. Prir. (Biol.) 64: 15-33.
27: 255-305.
Abbreviations used in figures
a.al. a.b. a.c.com. a.c.in. ad.b. a.fb. a.md. a.md.in. a.me. a.me.p. a.mx.in. a m . a.oe.a. a.oe.m.
alveolar artery basilar artery common carotid artery internal carotid artery adrenal body artery to fat body mandibular artery internal mandibular artery mesenteric artery posterior mesenteric artery internal maxillary artery oesophageal artery anterior oesophageal artery middle oesophageal artery
a.oe.p. a.ov. a.p. a.p.ce. a.pul. a .m. a.sc.d. a.sc.v. as t . a.v. ad.b.
ba. car.
posterior oesophageal artery ovarian artery palatine artery posterior cerebral artery pulmonary artery rectal artery dorsal spinal artery ventral spinal artery stapedial artery vertebral artery adrenal body
0s basale carotid arch

228
c.n.p. ch.
d.art. d.c. da.
f.b.
€LOP. gh. gt . h.
im.
k.
1.m.a. 1.m.i. 1.m.p. 1.9. 1.v. j.com. I.v.ra.mu. 1.v.vc.a. li.
m.cu.d. m.d.tr. m.is. m.it. m.0.a.e. m.0.a.i. m.0.a.p. m.r.a. m.tra. md. mx.
n.pmx. no.ch.
OP. ov.
p.ar. p.lin. p.p.m. p.p.t. pl.ch. Pr.
r.v.j.com. r.v.vc.a. rb.
cavum nasale principale ceratohyal
ductus arteriosus ductus caroticus dorsal aorta
fat body
ophthalmic ganglion m. geniohyoideus gut
lobus hemisphericus
m. intermandibularis
kidney
R. LAWSON
s.an. s.ar. s.c.ar. s.md. s.mx. s.p.1. s.po. ssag. sh. sph. sq.
m. levator mandibdae anterior m. levator mandibulae internus m. levator mandibdae posterior m. levator quadrati left common jugular vein muscular ramus of left anterior vena cava left anterior vena cava liver
m. cutaneous dorsalis m. dorsalis trunci m. interspinalis m. intertransversarius m. obliquus abdominis externus m. obliquus abdominis internus rn. obliquus abdominis internus profundus m. rectus abdominis m. transversus mandibular nerve maxillary nerve
naso-premaxilla nodus chorioideus
ramus ophthalmicus V ovary
pulmonary arch lingual plexus median palatine plexus transverse palatine plexus chorioid plexus parietal
right common jugular vein right anterior vena cava rib
t. t.v. thm.
v.ab. v.an. v.az.a. v.az.a.d. v.az.a.v. v.az.p. V.C.1. v.c.m. v.cdsp.
v.cl0.m. v.cr.m.0.
v.cr.p.0.
v.cr.po.0.
v.cut.a. v.cut.m. v.epg.a. v.f. v.fb. v.f.c. v.f.s. v.fr. v.h.p. v.ih.p. v.in. v.in.p. v.j.com. v.j.ex. v.j.i. v.j.1. v.1. v.1.d. v.1.m. V.I.V.
v.md. v.md.in.
anal sinus systemic arch systemico-carotid arch mandibular sinus maxillary sinus lateral palatine sinus post-orbital sinus sagittal sinus m. subhyoideus sphenethmoid squamosal
tentacle trunk vertebra thymus
abdominal vein anal vein anterior azygos vein dorsal branch of anterior azygos vein ventral branch of anterior azygos vein posterior azygos vein vena capitis lateralis vena capitis medialis vein draining m. cephalodorso-subpharyn- geus median cloaca1 vein vena cranialis met-otica (median cerebral vein) vena cranialis pre-otica (anterior cerebral vein) vena cranialis post-otica (posterior cerebral vein) anterior cutaneous vein vena cutana magna anterior epigastric vein facial vein vein draining fat body common facial vein superior facial vein frontal vein hepatic portal vein vein draining m. interhyoideus posterior intestinal vein posterior intestinal vein common jugular vein external jugular vein internal jugular vein lateral jugular vein laryngeal vein dorsal longitudinal vein median longitudinal vein ventral longitudinal vein mandibular vein internal mandibular vein
rec. rectum v.md.ex. external mandibular vein
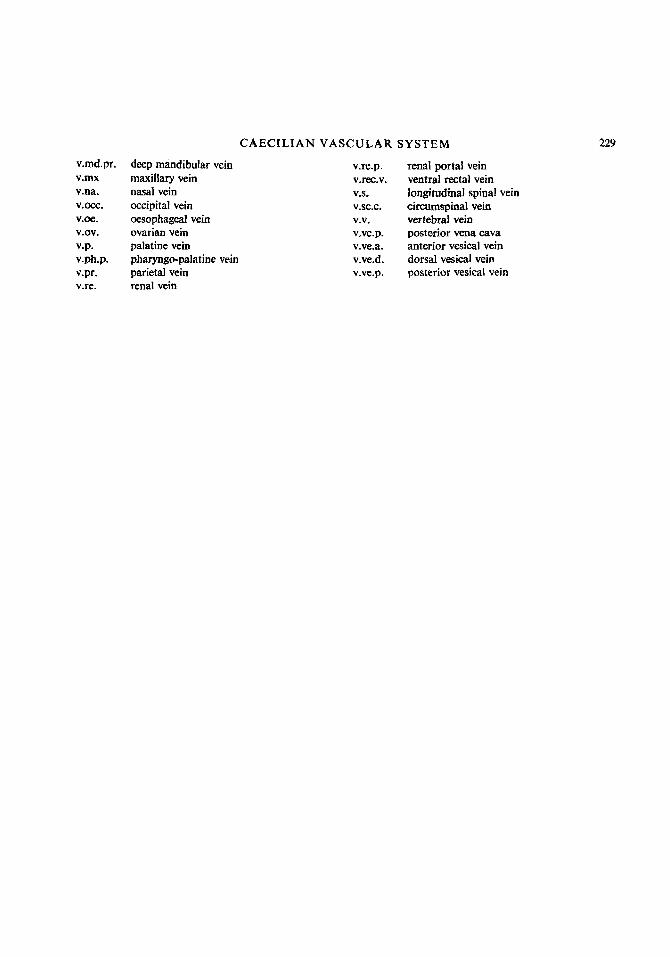
v.md.pr. v.mx v.na. V.0Cc. V.0e.
v.0v. v.p. v.ph.p. v.pr. v.re.
deep mandibular vein maxillary vein nasal vein occipital vein oesophageal vein ovarian vein palatine vein pharyngo-palatine vein parietal vein renal vein
CAECILIAN VASCULAR SYSTEM
v.re.p. v.rec.v. V.S. v.sc.c. V.V. v.vc.p. v.ve.a. v.ve.d. v.ve.p.
renal portal vein ventral rectal vein longitudinal spinal vein circumspinal vein vertebral vein posterior vena cava anterior vesical vein dorsal vesical vein Dosterior vesical vein
229
