survivorship, development, and dna damage in echinoderm embryos and larvae exposed to ultraviolet...
TRANSCRIPT

www.elsevier.com/locate/jembe
Journal of Experimental Marine Biology and Ecology
292 (2003) 75–91
Survivorship, development, and DNA damage
in echinoderm embryos and larvae exposed to
ultraviolet radiation (290–400 nm)
Michael P. Lesser*, Thomas M. Barry
Department of Zoology and Center for Marine Biology, University of New Hampshire, Durham, NH 03824, USA
Received 11 September 2002; received in revised form 12 March 2003; accepted 14 March 2003
Abstract
Laboratory experiments utilizing ecologically relevant irradiances of ultraviolet radiation (UVR)
known to occur in shallow Gulf of Maine waters were conducted on the planktonic embryos and
larvae of two common benthic echinoids; the green sea urchin Strongylocentrotus droebachiensis
and the sand dollar Echinarachnius parma. Significant decreases in survivorship were observed in
freshly fertilized embryos of both species with greater mortality in E. parma that was associated with
the absence of UVR-absorbing compounds, the mycosporine-like amino acids. Experiments on
blastula, gastrula, and prism larval stages of S. droebachiensis also showed significant decreases in
survivorship, delays in development, and abnormal embryos and larvae associated with exposure to
UVR. Additionally, all developmental stages of S. droebachiensis experimentally exposed to UVR
resulted in significant increases in DNA damage, measured as cyclobutane pyrimidine dimer
photoproducts. The observed delays in early cleavage and subsequent developmental stages for S.
droebachiensis are correlated with DNA damage. It is postulated that cell cycle arrest at critical
checkpoints after DNA damage, mediated by a suite of cell cycle genes, is a component of the
observed UVR induced developmental delays.
D 2003 Elsevier Science B.V. All rights reserved.
Keywords: DNA damage; Echinoids; Embryos; Larvae; Ultraviolet radiation
1. Introduction
The effects of exposure to ultraviolet radiation (UVR: 290–400 nm) have become a
major concern in the marine environment. In particular, the decrease of stratospheric
0022-0981/03/$ - see front matter D 2003 Elsevier Science B.V. All rights reserved.
doi:10.1016/S0022-0981(03)00141-2
* Corresponding author. Tel.: +1-603-862-3442; fax: +1-603-862-3784.
E-mail address: [email protected] (M.P. Lesser).

M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–9176
ozone from anthropogenic inputs of chlorinated fluorocarbons has resulted in an increase
in the amount of biologically damaging ultraviolet-B radiation (UV-B: 290–320 nm)
reaching the sea surface. Attention has largely been focused on the Antarctic, and more
recently the Arctic, where the autocatalytic destruction of stratospheric ozone (‘‘the ozone
hole’’) leads to enhanced fluxes of UV-B (Smith and Baker, 1989; Madronich et al.,
1998) and a decrease in primary productivity (Smith et al., 1992). Recent global
assessments of the depletion of stratospheric ozone have shown increases of unweighted
UV-B radiation of up to 130% in the Antarctic, 22% in the Arctic, and 4–7% at temperate
latitudes (Madronich et al., 1998). The temperate latitudes include coastal areas that
contain many productive estuarine, intertidal, and shallow subtidal habitats such as those
found in the Gulf of Maine. Current predictions suggest that higher irradiances of UV-B
radiation will affect the marine environment for at least the next 25 years, if not longer
(Madronich et al., 1998). Of particular concern at any latitude is that ozone depletion
results in an increase in damaging UV-B wavelengths without a proportional increase in
longer UV-A and blue wavelengths involved in photoreactivation and photorepair (Smith,
1989; Halliwell and Gutteridge, 1999). In the Gulf of Maine, UVR can penetrate to
depths as deep as 7–12 m depending on the optical properties of the water column
(Banaszak et al., 1998; Adams and Shick, 2001; Lesser et al., 2001). The transmission of
UVR in the Gulf of Maine varies with changes in the stability and optical properties of
the water column due to seasonal variation in solar irradiation, phytoplankton blooms, or
increases in dissolved organic matter (Kirk, 1994). These changes in the attenuation of
UVR could have significant biological effects on the planktonic life-history phases of
invertebrates, fish, and macrophytes. Ultraviolet radiation is known to have a detrimental
effect on phytoplankton, bacteria, macrophytes, and zooxanthellate corals (Jokiel and
York, 1984; Cullen et al., 1992; Gleason and Wellington, 1993; Herndl et al., 1993;
Lesser et al., 1996; Lesser and Lewis, 1996; Shick et al., 1996; Franklin and Forester,
1997), and the harmful effects of UVR may involve damage to DNA and proteins,
oxidation of membrane lipids, and photooxidation of chlorophyll or damage to photo-
system II (Halliwell and Gutteridge, 1999). Structural damage to DNA can be caused
directly by UV-B and indirectly by UV-A (320–400 nm) wavelengths. The cyclobutane
pyrimidine dimer (CPD) is the predominant photoproduct of direct UV-B exposures,
while UV-A wavelengths cause damage indirectly to DNA through the photodynamic
production of hydroxyl radicals causing strand breaks and DNA cross-links (Peak and
Peak, 1990). The amount of photodamage to DNA is dependent on the equilibrium
between damage, which is dependent on the absorbed dose of UVR, rates of repair, and
rates of cell division.
Additionally, there are significant synergistic effects of solar UVR with oxidative stress
in marine organisms. Reactive oxygen species (ROS) are formed by the univalent
reduction of molecular O2, yielding superoxide radicals (O2�), and further reduction of
superoxide yields hydrogen peroxide (H2O2) and hydroxyl radicals (HO�) via the iron-
catalyzed Haber–Weiss reaction (Fridovich, 1986). These reactions are known to occur
normally in many metabolic processes in the cytosol, mitochondria, and chloroplasts
(Asada and Takahashi, 1987; Halliwell and Gutteridge, 1999) but concentrations of these
radicals increase under stressful conditions (Fridovich, 1986; Asada and Takahashi, 1987).
Defenses against ROS include various non-enzymatic antioxidants and quenchers (e.g.,

M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–91 77
ascorbate, tocopherols, carotenoids), and enzymes such as superoxide dismutase which
removes O2� but in so doing produces H2O2, catalase which then removes H2O2, and
ascorbate and glutathione peroxidases which remove H2O2 and other peroxides (Asada
and Takahashi, 1987; Halliwell and Gutteridge, 1999).
Reactive oxygen species also have multiple adverse effects on marine organisms as
described for UVR, including damage to DNA/RNA, inhibition of photosynthesis, and
bleaching of zooxanthellate corals (Dykens et al., 1992; Shick et al., 1995, 1996; Lesser,
1996, 1997; Lesser et al., 2001). Additionally, oxidative stress is known to play a role in
apoptosis or programmed cell death through several cell cycle genes such as p53. The
principal function of p53 is to promote the repair or deletion of cells exposed to agents that
cause DNA damage, like hypoxia, UVR, ROS, or mutagens (Graeber et al., 1996; Renzing
et al., 1996; Griffiths et al., 1997).
Defenses against solar UVR include blocking potentially harmful wavelengths by using
UV-absorbing compounds. Many of these compounds have been identified as mycospor-
ine-like amino acids (MAAs), and are found in a wide variety of marine organisms
(Karentz et al., 1991; Shick and Dunlap, 2002). A protective role of these compounds has
been inferred from their UVR absorbing properties and their decrease in concentration
with increasing depth or after experimental shielding from UVR (Shick and Dunlap,
2002). Eggs, embryos, and larvae from temperate sea urchins contain MAAs (Banaszak et
al., 1998; Carroll and Shick, 1996; Adams and Shick, 1996, 2001), as do sea star and
holothuroid eggs and larvae (Lesser unpublished).
Several adverse effects of UVR have already been documented for marine organisms,
including species of crustacean zooplankton that are normally found in near-surface waters
where high fluxes of UVR are present (Karansas et al., 1979; Damkaer and Dey, 1983;
Dey et al., 1988). Recent data have shown a significant increase in DNA damage,
measured as an increase in CPD formation, in ice fish and krill collected from shallow
waters in Antarctica during the occurrence of the ozone hole despite active DNA repair
capabilities (Malloy et al., 1997). DNA damage and up-regulation of p53 has also been
reported in laboratory experiments on the embryos of cod fish (Gadus morhua) in
response to UVR irradiances similar to exposures where developing embryos would be
found in the Gulf of Maine (Lesser et al., 2001). Near-surface waters also contain the
gametes and larvae of many species of benthic invertebrates. Larvae of the sand dollar
Dendraster excentricus exposed to UVR resulted in negative phototaxis and significant
mortality (Pennington and Emlet, 1986). Gleason and Wellington (1995) described
significant depth-dependent mortality of planula larvae from the coral, Agaricia agaricites
by UV-B radiation that was associated with differences in the MAA concentrations that
larvae obtained from their parent colonies. Urchin larvae also obtain their MAAs through
the eggs from their parents and the concentration of these compounds can be manipulated
by pre-conditioning adults with algal sources of these compounds (Adams and Shick,
1996; Carroll and Shick, 1996). Exposure of gametes and larvae to high doses of UVR
could potentially affect fertilization success, the timing of cleavage, and development time
for embryos that survive as observed for the temperate sea urchin, Strongylocentrotus
droebachiensis (Adams and Shick, 1996, 2001). Many of these effects on gametes and
larvae might be ameliorated if sufficient MAAs can be translocated from adults to gametes
(Adams and Shick, 1996).

M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–9178
The present study is the first of a series from laboratory and field experiments on the
effects of UVR on survival, development, and the biochemical and molecular response to
UVR in echinoderm larvae from the Gulf of Maine. Laboratory experiments examining the
effects of UVR on survivorship, development, and DNA damage on the embryos and
larvae from the green sea urchin S. droebachiensis and the sand dollar, Echinarachnius
parma are presented here.
2. Materials and methods
2.1. Experimental animals
Adult green sea urchins, S. droebachiensis (Muller), and sand dollars, E. parma
(Lamarck), were collected from around the Isles of Shoals, Gulf of Maine (42j59.29VN,70j37.01VW) at a depth of 10 m in June of 1999 for sand dollars and March of 2000 for sea
urchins. All animals were brought back to the University of New Hampshire Coastal
Marine Laboratory where they were maintained at 4–5 jC for sea urchins and 10 jC for
sand dollars. Animals were not feed and all were used within 2 weeks of collection.
Reproductively mature animals were brought into the laboratory and induced to spawn by
intracoelomic injection of 0.5 M KCl at 4 jC. Eggs from at least five females were
combined and washed three times with 0.22-Am filtered seawater and collected using a
100-Am Nitex filter after each wash. Sperm were left ‘‘dry’’ until used and the sperm of at
least three males combined and diluted in 0.22 Am filtered seawater at f 1:10,000 to
fertilize eggs at a density of 500 individuals ml� 1 in 2 l sterile plastic containers (Corning).
After 5 min, freshly fertilized embryos (FFE) were washed to remove excess sperm and
maintained at a density of 20–30 individuals ml� 1 until used for experiments. Both S.
droebachiensis and E. parma FFE were used within 15 min of fertilization in experimental
exposures to UVR, and samples taken at this time showed that fertilization success was
over 95% (three subsamples of 5 ml and counting 100 embryos in each sample). For
experiments on other stages of development in S. droebachiensis, embryos were allowed to
develop to blastula, gastrula, and prism stages and then exposed to UVR, as described
below. Embryos were aerated and gently stirred using magnetic stir bars. Larvae were fed
with phytoplankton (Isochrysis galbana) grown in nutrient replete f/2 medium as the prism
stage was reached. No larvae were fed during the experiments and all experiments were
conducted at 10 jC for E. parma and 4–5 jC for S. droebachiensis. A temperature-
controlled water bath was used to maintain (F 0.5 jC) these seawater temperatures.
2.2. Embryo and larval experiments
Freshly fertilized embryos, blastula, gastrula, and prism stage larvae were exposed to
artificial visible and UVR using four UV-340 lamps (Q-Panel) and four F40 Sun lamps
(General Electric) suspended f 20 cm from the top of the filters (see below) to provide a
downwelling mixed field (visible and UVR) exposure. Glass beakers (400 ml) containing
f 5 individuals ml� 1 were used for testing the effects of UVR on embryos and larvae.
Three treatments were used to partition the effects of UV-B from UV-A and visible

M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–91 79
radiation with three replicates per treatment. Schott WG and GG long pass filters (6� 6W)with nominal cutoffs (50% T) at 280, 320, and 400 nm were used to cover the beakers
containing 400 ml of embryos or larvae. Treatments were designated as UVO (GG 400
filter), UVA (WG 320 filter), and UVT (WG 280 filter). All experiments on embryos and
larvae tested were conducted for 3 days on a 12:12-h light/dark cycle. Using this design,
successive replicate experiments were carried out using embryos and larvae at different
developmental stages for S. droebachiensis, while only FFE were analyzed experimentally
for E. parma. During the 3-day experiments, samples were taken at T0, 30, 60, 180 min; 6,
12, 24, 48, and 72 h after fertilization for S. droebachiensis and E. parma to assess percent
survivorship under each treatment. Subsamples (10 ml) were fixed in 3% glutaraldehyde
and filtered (0.22 Am) seawater and counts of normally developed stages of development
were recorded as survivors using light microscopy. Percent survivorship for all treatments
was determined by comparing the counts of embryos or larvae for each sampling period
against the T0 counts. In addition, the developmental stage at the end of the experiment in
each treatment, and whether embryos and larvae were developing normally was recorded.
Digital light micrographs of S. droebachiensis were taken for each developmental stage
and for some stages during exposure to UVR. At the end of 3 days, all remaining embryos
or larvae were collected for additional analyses, as described below.
A duplicate set of experiments was run to determine the effects of UVR on delays in
cleavage for the FFE of S. droebachiensis and E. parma as described above with the
following exceptions. Preliminary experiments indicated that S. droebachiensis and E.
parma take from 2.5–3.0 h and 30–60 min, respectively, to begin their first mitotic
division, as indicated by the start of a cleavage furrow, at the experimental temperatures
described above. To assess the effects of UVR on delays in cleavage, samples of FFE were
taken every 15 min beginning at 105 min after fertilization for S. droebachiensis and every
10 min for E. parma immediately after fertilization until all FFE had cleaved and fixed to
determine the percent cleaved as a function of time as described by Adams and Shick
(1996). From 50 to 100 embryos were counted for each sample and the time at which FFE
for each species reached 50% cleavage was determined by a nonlinear regression fit of the
data using a logistic (exponential) equation.
2.3. Measurements of ultraviolet radiation
For laboratory experiments, UVR (UV-B and UV-A) and photosynthetically active
radiation (PAR: 400–700 nm) were measured using a wavelength and radiometrically
calibrated (NIST traceable standards) CCD spectrometer and fiber optics (Ocean Optics).
Three scans were taken and the mean reported was in units of W m� 2 nm� 1. Integrated
values of unweighted UVR (W m� 2) were calculated for each treatment and biologically
weighted irradiances (W m� 2) were obtained by multiplying the unweighted irradiance
with the DNA weighting function of Setlow (1974).
2.4. UV absorbing compounds (mycosporine-like amino acids)
Samples of embryos and larvae were analyzed for MAAs using the methods described
in Shick et al. (1995). Individual MAAs were separated with reverse-phase isocratic high

M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–9180
performance liquid chromatography (HPLC) on a Brownlee RP-8 column (Spheri-5: 4.6
mm ID� 25 cm) protected with an RP-8 guard column (Spheri-5: 4.6 mm ID� 3 cm)
using an aqueous mobile phase of 40–55% methanol (v/v) and 0.1% acetic acid at a flow
rate of 0.6 ml min� 1. Detection of peaks is by UVabsorbance at 313 and 340 nm. Identity
of peaks is accomplished using co-chromatography with known secondary standards
prepared from primary standards provided by Dr. Walt Dunlap via Dr. Deneb Karentz.
Quantification of individual peaks is accomplished using response factors calculated from
known amounts of standard. Soluble protein composition was assessed using Coomassie
brilliant blue, and bovine serum albumin as a standard (Bio-Rad Laboratories) to
normalize data.
2.5. Detection of DNA photoproducts using an enzyme-linked immunoabsorbent assay
Cyclobutane pyrimidine dimer (CPD) formation was measured using the procedures
and monoclonal antibody (TDM-2) of Mori et al. (1991). Embryo or larval genomic DNA
was isolated using commercially available kits (Easy-DNA, Invitrogen) and quality and
concentration determined spectrophotometrically using 260:280 nm ratios. Subsequently,
50 ng of DNA from each sample was used in an enzyme-linked immunoabsorbent assay
(ELISA) technique with TDM-2 as the primary antibody. An affinity purified goat anti-
mouse IgG secondary antibody conjugated with horseradish peroxidase was used and the
final color development read in flat-bottomed 96-well microtiter plates using a plate
reader (Bio-Rad) at 490 nm as described by Mori et al. (1991). As in Mori et al. (1991),
we present the absorbance results from the ELISA. Although differences between
treatments can be explored within an experiment, differences in DNA damage between
developmental stage experiments are confounded by differences in total DNA content,
cell shading in different stages, difference in rates of cell division between stages, as well
as differences in maternal versus zygotic transcript expression that are likely to affect rates
of photorepair.
2.6. Statistical analysis
Survivorship, cleavage delay, and CPD formation were statistically analyzed using a
one-way ANOVA at a significance level of 5%. No unequal variances were detected using
the Fmax test, and individual treatment differences were assessed using the Student–
Newman–Keuls (SNK) multiple comparison test. Where appropriate, ratios and percen-
tages were arcsine or log transformed for analysis and back transformed for presentation.
3. Results
Table 1 shows the UVR irradiances for each treatment along with the total dose for the
experiments on both the embryos and larvae of S. droebachiensis and E. parma. Year to
year differences were less than 3% and for all treatments, the irradiance of PAR was f 70
Amol quanta m� 2 s� 1. Additionally, the biologically effective irradiance (Table 1) for
each treatment was calculated using the DNA damage weighting function of Setlow
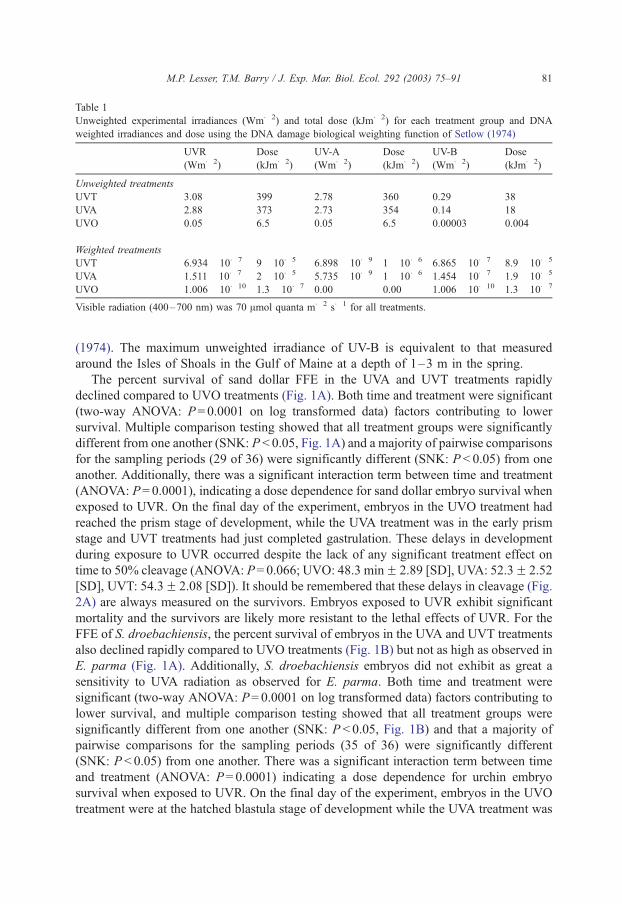
Table 1
Unweighted experimental irradiances (Wm� 2) and total dose (kJm� 2) for each treatment group and DNA
weighted irradiances and dose using the DNA damage biological weighting function of Setlow (1974)
UVR
(Wm� 2)
Dose
(kJm� 2)
UV-A
(Wm� 2)
Dose
(kJm� 2)
UV-B
(Wm� 2)
Dose
(kJm� 2)
Unweighted treatments
UVT 3.08 399 2.78 360 0.29 38
UVA 2.88 373 2.73 354 0.14 18
UVO 0.05 6.5 0.05 6.5 0.00003 0.004
Weighted treatments
UVT 6.934� 10� 7 9� 10� 5 6.898� 10� 9 1�10� 6 6.865� 10� 7 8.9� 10� 5
UVA 1.511�10� 7 2� 10� 5 5.735� 10� 9 1�10� 6 1.454� 10� 7 1.9� 10� 5
UVO 1.006� 10� 10 1.3� 10� 7 0.00 0.00 1.006� 10� 10 1.3� 10� 7
Visible radiation (400–700 nm) was 70 Amol quanta m� 2 s� 1 for all treatments.
M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–91 81
(1974). The maximum unweighted irradiance of UV-B is equivalent to that measured
around the Isles of Shoals in the Gulf of Maine at a depth of 1–3 m in the spring.
The percent survival of sand dollar FFE in the UVA and UVT treatments rapidly
declined compared to UVO treatments (Fig. 1A). Both time and treatment were significant
(two-way ANOVA: P= 0.0001 on log transformed data) factors contributing to lower
survival. Multiple comparison testing showed that all treatment groups were significantly
different from one another (SNK: P < 0.05, Fig. 1A) and a majority of pairwise comparisons
for the sampling periods (29 of 36) were significantly different (SNK: P < 0.05) from one
another. Additionally, there was a significant interaction term between time and treatment
(ANOVA: P= 0.0001), indicating a dose dependence for sand dollar embryo survival when
exposed to UVR. On the final day of the experiment, embryos in the UVO treatment had
reached the prism stage of development, while the UVA treatment was in the early prism
stage and UVT treatments had just completed gastrulation. These delays in development
during exposure to UVR occurred despite the lack of any significant treatment effect on
time to 50% cleavage (ANOVA: P= 0.066; UVO: 48.3 minF 2.89 [SD], UVA: 52.3F 2.52
[SD], UVT: 54.3F 2.08 [SD]). It should be remembered that these delays in cleavage (Fig.
2A) are always measured on the survivors. Embryos exposed to UVR exhibit significant
mortality and the survivors are likely more resistant to the lethal effects of UVR. For the
FFE of S. droebachiensis, the percent survival of embryos in the UVA and UVT treatments
also declined rapidly compared to UVO treatments (Fig. 1B) but not as high as observed in
E. parma (Fig. 1A). Additionally, S. droebachiensis embryos did not exhibit as great a
sensitivity to UVA radiation as observed for E. parma. Both time and treatment were
significant (two-way ANOVA: P= 0.0001 on log transformed data) factors contributing to
lower survival, and multiple comparison testing showed that all treatment groups were
significantly different from one another (SNK: P < 0.05, Fig. 1B) and that a majority of
pairwise comparisons for the sampling periods (35 of 36) were significantly different
(SNK: P < 0.05) from one another. There was a significant interaction term between time
and treatment (ANOVA: P= 0.0001) indicating a dose dependence for urchin embryo
survival when exposed to UVR. On the final day of the experiment, embryos in the UVO
treatment were at the hatched blastula stage of development while the UVA treatment was

Fig. 1. (A) Mean survival (F S.D.) of sand dollar (E. parma) freshly fertilized embryos exposed to UVR in the
three treatment groups. (B) Mean survival (F S.D.) of sea urchin (S. droebachiensis) freshly fertilized embryos
exposed to UVR in the three treatment groups. Each curve represents the percent survivors after exposure to the
UVR irradiances in Table 1 over the 3-day duration of the experiment.
M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–9182
in the blastula stage and UVT treatments were in the early blastula stage. These delays in
development during exposure to UVR, unlike the delays observed in E. parma, were
accompanied by significant treatment effects on time to 50% cleavage (ANOVA:
P < 0.0001; UVO: 193.7 minF 3.22 [SD], UVA: 202.3F 2.52 [SD], UVT: 261.0F 3.61
[SD]).

Fig. 2. Developmental stages of the green sea urchin (S. droebachiensis) (A) first cleavage (f 175 Am) (B)
blastulae (f 200 Am), (C) gastrula (f 300 Am), (D) prism stage larvae (325–350 Am), (E) four arm pluteus larva
(f 550 Am), and (F) packed blastula (f 125 Am).
M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–91 83

M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–9184
Only trace amounts of the MAA mycosporine-glycine were detected in the FFE of
embryos from E. parma which were too low and variable to be quantified accurately. In
the FFE of S. droebachiensis, large amounts of shinorine and porphyra-334 were observed
and trace amounts of mycosporine-glycine and palythine. Only shinorine and porphyra-
334 were quantified and analyzed. Neither shinorine (UVO; 82.2 nmol mg
protein� 1F1.05 [SD], UVA: 75.8F 6.61 [SD], UVT 86.7F 7.68 [SD]) nor porphyra-
334 (UVO: 562.6 nmol mg protein� 1F 56.80 [SD], UVA: 535.1F 21.32 [SD], UVT:
671.5F 121.69 [SD]) concentrations were significantly different (ANOVA: P>0.05) in
time zero samples of FFE.
Three additional survivorship experiments were conducted on the blastula, gastrula, and
prism stages of the green sea urchin S. droebachiensis. For the experiment on blastula (Fig.
2B), both time and treatment were significant (two-way ANOVA: P= 0.0001 on log-
transformed data) factors contributing to lower survival and multiple comparison testing
showed that all treatment groups were significantly different from one another (SNK:
P < 0.05, Fig. 3A) and that all pairwise comparisons for the sampling periods (36 of 36)
were significantly different (SNK: P < 0.05) from one another. There was a significant
interaction term between time and treatment (ANOVA: P= 0.0001) indicating a dose
dependence for blastula survival when exposed to UVR. Overall mortality in each
treatment group was less than that observed in the FFE experiment. On the final day of
the experiment, blastula in the UVO and UVA treatments was at the early gastrula stage
(Fig. 2C) of development while the UVT treatment were in the hatched blastula stage.
Survivorship results for the experiment on gastrula (Fig. 2C) were similar to those
observed on blastula. Both time and treatment were significant (two-way ANOVA:
P= 0.0001 on log transformed data) factors contributing to lower survival, and multiple
comparison testing showed that all treatment groups were significantly different from one
another (SNK: P < 0.05, Fig. 3B) and that all pairwise comparisons for the sampling
periods (36 of 36) were significantly different (SNK: P < 0.05) from one another. There
was a significant interaction term between time and treatment (ANOVA: P= 0.0001),
indicating a dose dependence for gastrula survival when exposed to UVR. Again, overall
mortality in each treatment group was less than that observed in the FFE experiment and
similar to the blastula experiment. On the final day of the experiment, gastrula in the UVO
and UVA treatments were at the late gastrula and early prism stage of development while
the UVT treatment was in the late gastrula stage. Lastly, the survivorship results for the
experiment on prism stage larvae (Fig. 2D) were again similar to those observed on
blastula and gastrula. Both time and treatment were significant (two-way ANOVA;
P= 0.0001 on log transformed data), and multiple comparison testing showed that all
treatment groups were significantly different from one another (SNK: P < 0.05, Fig. 3C)
and that all pairwise comparisons for the sampling periods (36 of 36) were significantly
different (SNK: P < 0.05) from one another. There was a significant interaction term
between time and treatment (ANOVA: P= 0.0001) indicating a dose dependence for prism
larvae survival when exposed to UVR. Overall mortality in each treatment group was less
than that observed in the FFE experiment and similar to experiments on other devel-
opmental stages. On the final day of the experiment, prism larvae in the UVO and UVA
treatments were at the four-arm pluteus stage (Fig. 2E) of development while the UVT
treatment was in the late prism, early four-arm plutei stage.
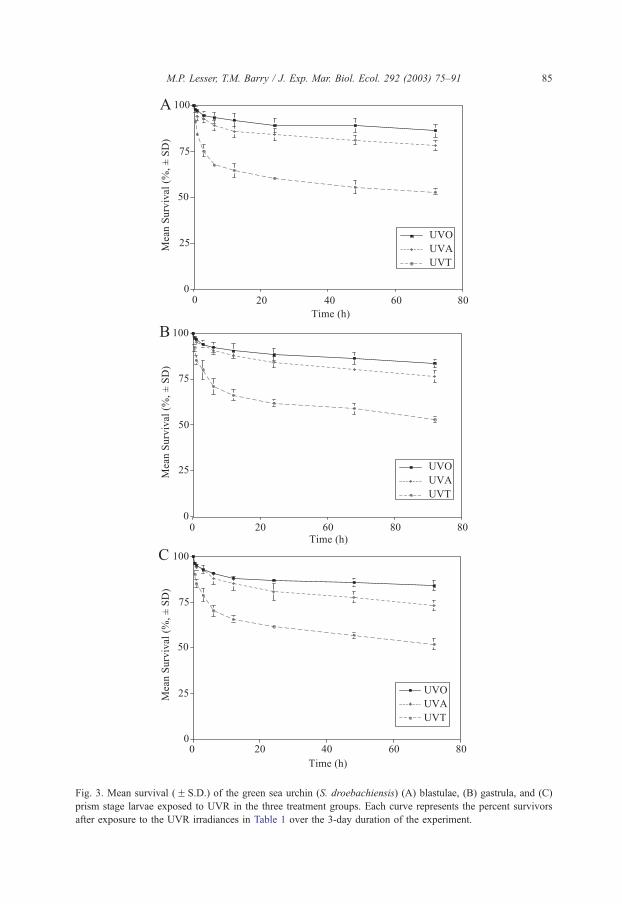
Fig. 3. Mean survival (F S.D.) of the green sea urchin (S. droebachiensis) (A) blastulae, (B) gastrula, and (C)
prism stage larvae exposed to UVR in the three treatment groups. Each curve represents the percent survivors
after exposure to the UVR irradiances in Table 1 over the 3-day duration of the experiment.
M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–91 85

Fig. 4. DNA damage (meanF S.D.), measured as cyclobutane pyrimidine dimmers (CPDs) of green sea urchin
(S. droebachiensis) embryos and larvae exposed to UVR in the three treatment groups. Treatment groups with
common superscripts are not significantly different from one another using multiple comparisons testing (SNK)
at a significance level of 0.05%.
M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–9186
At the end of each experiment, CPD photoproducts were measured, as described above.
All experiments exhibited significant (ANOVA: P < 0.05 on log-transformed values)
treatment effects on CPD formation (Fig. 4). For FFE, blastula, and gastrula, multiple
comparison testing of CPD formation revealed that all treatment groups were significantly
different from one another (SNK: P < 0.05, Fig. 4) and that UVT treatments in all cases
exhibited higher concentrations of CPDs followed by UVA and UVO treatments. For the
prism stage experiments, the UVO and UVA treatments were indistinguishable from each
other (SNK: P>0.05) but significantly different (SNK: P < 0.05) than the UVT treatment
(Fig. 4) that exhibited the highest concentration of CPDs.
4. Discussion
The sea urchin, S. droebachiensis, and the sand dollar, E. parma, are common
echinoderms found in the Gulf of Maine. These benthic echinoderms inhabit hard and
soft-bottom environments from shallow to deep-water in the GOM. The depth distribution of
these species in the shallow subtidal is relevant to the studies reported here. UVR penetration
to depths of 7–12 m has been reported in the GOM. Both of the target species have
significant populations at these depths where their principal food sources, macrophytes and
phytoplankton, are available. It is not unreasonable to hypothesize that the buoyant,
swimming, or vertically advected planktonic phases of these benthic echinoderms are
exposed to UVR during the spring and early summer in the GOM, the amount of which will

M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–91 87
be greatly affected by their depth of occurrence, and the stability and optical properties of the
water column. In this regard, an important result of the experiments described above is the
dose dependency of the decrease in survivorship with time. This has important consequences
for embryos and larvae in the plankton whose residence time within the upper portion of the
water column (< 10 m) is controlled by physical processes. An important component of
assessing the effects of a longer residence time in the upper portion of the water column is
whether reciprocity holds, that is, does a short-term exposure to high irradiances to UVR
have the same effect as a longer-term, lower irradiance exposure where the total dose is
equivalent.
Previous work on echinoid embryos and larvae has shown significant effects of UVR
on cell division, development, morphology, and DNA damage (Rustad, 1960; Eima et al.,
1984; Akimoto and Shiroya, 1987a,b). However, all of these studies utilized artificial light
sources containing germicidal UV-C (254 nm) radiation that is not ecologically relevant, as
UVR below 290 nm does not reach sea level. Recent studies by Adams and Shick (1996,
2001) have confirmed the occurrence of some of these effects on urchin embryos when
exposed to environmentally relevant wavelengths of UV-A and UV-B. Additionally, the
work reported here also confirms the adverse effects of environmentally relevant UVR on
echinoid survivorship, development, morphology, and DNA damage. Freshly fertilized
embryos of S. droebachiensis and E. parma, which have markedly different concentrations
of MAAs, exhibit clear species-specific differences in survivorship. Surprisingly, there
were no significant effects of UVR on cleavage delay in E. parma. One interpretation of
this result is that cleavage delay was measured on the low number of highly resistant
survivors in the experiment. Action spectra for the effect of UVR on cleavage delay in a
conspecific of the green sea urchin, Strongylocentrotus purpuratus, showed that UVR of
302 nm caused greater delays in cell division than the UVR of either 313 nm or 366 nm
(Geise, 1939). These results are similar to the results reported here for UVR effects on
cleavage delay in S. droebachiensis. On a daily basis, the unweighted and weighted
irradiances of UV-B used in this study were greater than that reported in Adams and Shick
(2001) but the proportional amounts of visible and UV-A, which are important in the
photorepair of DNA damage (Smith, 1989), are higher. The exposures described here also
simulate irradiances of UVR measured in the shallow (1–3 m) waters of the Gulf of Maine
in the spring which these embryos and larvae are likely to be exposed to.
For the subsequent developmental stages, blastula, gastrula, and prism, exposure to
UVR also resulted in significant decreases in survivorship but not as high as observed for
FFE and most of which can be explained by exposure to UV-B versus UV-A radiation. The
two dominant MAAs in all of the experiments were shinorine and porphyra-334 that
absorb maximally at 334 nm. Even though MAAs have broad absorption spectra
extending into the UV-B portion of the spectrum, and very high molar extinction
coefficients, the effects of UV-B in these experiments are significant for all developmental
stages. Previous work has shown that MAAs are invariant during development up to the
pluteus stage (Adams and Shick, 1996) or when exposed to UVR (Adams and Shick,
2001), and can have has high as an 86% efficiency of self-shading at 334 nm with high
MAA concentrations, as reported above. This suggests that the embryos and larvae in the
experiments reported here, up to the prism stage, have relatively stable MAA concen-
trations and should provide some protection from UVR if present.

M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–9188
During the experiments reported above only normally developing embryos and larvae
were counted as survivors because of the assumption that abnormal embryos and larvae
would not complete their developmental program. Beyond any delay in early cleavage,
there were also delays in the remainder of the developmental program when exposed to
UVR. Qualitative observations of the abnormal embryos showed similar types of abnormal
embryos as described by Adams and Shick (2001) including ‘‘packed blastula’’ (Fig. 2F),
exogastrula, and abnormal larvae. In contrast, the percentage of abnormal embryos or
larvae never exceeded 20% of the survivors at the end of the experiment, while Adams and
Shick (2001) report as high as 50% abnormal development in embryos with low
concentrations of MAAs and 10–15% abnormal development in embryos with high
concentrations of MAAs.
The DNA damage, measured as CPD photoproducts, observed in these experiments is
the direct result of exposure to UVR. DNA damage was not linearly related to survivorship
in any developmental stage. That is, in early developmental stages, it appears that small
amounts of DNA damage had greater effects on survivorship compared to the larger
amounts of DNA damage observed in later developmental stages. One reason for this
could be that differences in repair capacity exist as a result of differing amounts of
maternally derived transcripts of important repair enzymes such as photolyase compared to
the available zygotic transcripts in later developmental stages. Additionally, later stages of
development may be more tolerant of accumulating DNA damage than early stages (Epel
et al., 1999). Also, DNA damage may have also been caused indirectly by exposure to
UVR through the photodynamic production of ROS to a greater degree in early embryos
that resulted in decreased survivorship (Imlay and Linn, 1988). Either type of DNA
damage can lead to apoptosis if repair is not possible or delays in cell division while repair
is taking place (Paulovich et al., 1997). p53 can initiate cell cycle delays through either the
G1/S or G2/M checkpoints (Geyer et al., 2000; Innocente et al., 1999). Cleavage delays in
the FFE of S. droebachiensis and E. parma are the result of exposure to UVR. The
evidence presented here supports DNA damage as a precursor to those delays in cell
division and suggests the involvement of cell cycle genes in the observed delays. Delays in
development in the experiments on other developmental stages could be the result of the
time required to repair UVR-induced DNA damage, or the result of direct damage to
essential genes in the developmental program that are not repaired (Epel et al., 1999).
These factors, and our interpretation of experimental results, require us to put the effects of
UVR on DNA damage and subsequent delays in cell division in the context of the
equilibrium between damage and repair. Many cells of a developing embryo may require
the repair of all DNA damage or the repair of just enough DNA damage to proceed with
the developmental program (Epel et al., 1999). For fast developing embryos, such as sea
urchins, many cells may incur DNA damage that cannot be repaired but instead leads to
apoptosis and the removal of those cells. Sea urchin embryos may be able to tolerate the
loss of these cells and still complete their developmental program (Epel et al., 1999).
If DNA damage is the principal insult to developing embryos exposed to ecologically
relevant UVR, we should develop biological weighting functions to better compare our
experimental exposures to UVR, and to determine whether the irradiances of UVR in very
dynamic temperate waters will have similar effects on developing embryos in situ.
Weighting functions will also tell us something about the efficacy of MAAs in preventing

M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–91 89
DNA damage. Lastly, an examination of the genes involved in repair of DNA damage
(e.g., photolyase) and the differential expression of cell cycle genes will provide critical
insights into the effects of UVR on the cell cycle in developing echinoids.
Acknowledgements
Technical assistance from Mary Maravic is gratefully acknowledged, and a special
thanks to Dr. Toshio Mori for generously supplying the monoclonal antibodies to CPD.
This project was supported by the National Science Foundation (Biological Oceanography
Program, OCE-9818918). [SS]
References
Adams, N., Shick, J.M., 1996. Mycosporine-like amino acids provide protection against ultraviolet radiation in
eggs of the green sea urchin Strongylocentrotus droebachiensis. Photochem. Photobiol. 64, 149–158.
Adams, N., Shick, J.M., 2001. Mycosporine-like amino acids prevent UV-B induced abnormalities during early
development of the green sea urchin Strongylocentrotus droebachiensis. Mar. Biol. 138, 267–280.
Akimoto, A., Shiroya, T., 1987a. Photoreversability of UV-induced thymine dimmers and abnormal morpho-
genesis in sea-urchin embryos. Photochem. Photobiol. 45, 403–406.
Akimoto, A., Shiroya, T., 1987b. Photoreversal of abnormal morphogenesis in sea-urchin embryos caused by
UV-radiation. Photochem. Photobiol. 45, 407–412.
Asada, K., Takahashi, M., 1987. Production and scavenging of active oxygen in photosynthesis. In: Kyle, D.J.,
Osmond, C.B., Arntzen, C.J. (Eds.), Photoinhibition. Elsevier, Amsterdam, pp. 228–287.
Banaszak, A.T., Lesser, M.P., Kuffner, I.B., Ondrusek, M., 1998. Relationship between ultraviolet (UV) radiation
and mycosporine-like amino acids (MAAS) in marine organisms. Bull. Mar. Sci. 63, 617–628.
Carroll, A.K., Shick, J.M., 1996. Dietary accumulation of UV-absorbing mycosporine-like amino acids (MAAs)
by the green sea urchin (Strongylocentrotus droebachiensis). Mar. Biol. 124, 561–569.
Cullen, J.J., Neale, P.J., Lesser, M.P., 1992. Biological weighting function for the inhibition of phytoplankton
photosynthesis by ultraviolet radiation. Science 258, 646–651.
Damkaer, D.M., Dey, D.B., 1983. UV damage and photoreactivation potentials of larval shrimp, Pandalus
platyceros, and adult euphausiids Thysanoessa raschii. Oecologia 60, 169–175.
Dey, D.B., Damakaer, D.M., Heron, G.A., 1988. Dose/dose rate responses of seasonally abundant copepod of
Puget Sound. Oecologia 76, 321–329.
Dykens, J., Shick, J.M., Benoit, C., Buettner, G.R., Winston, G.W., 1992. Oxygen radical production in the sea
anemone Anthopleura elegantissima and its endosymbiotic algae. J. Exp. Biol. 168, 219–241.
Eima, Y., Ikenaga, M., Shiroya, T., 1984. Action spectrum for photoreactivation of ultraviolet-induced morpho-
logical abnormality in sea urchin eggs. Photochem. Photobiol. 40, 461–464.
Epel, D., Hemela, K., Shick, M., Patton, C., 1999. Development in the floating world: defenses of eggs and
embryos against damage from UV radiation. Am. Zool. 39, 271–278.
Franklin, L.A., Forester, R.M., 1997. The changing irradiance environment: consequences for marine macrophyte
physiology, productivity and ecology. Eur. J. Phycol. 32, 207–232.
Fridovich, I., 1986. Biological effects of the superoxide radical. Arch. Biochem. Biophys. 247, 1–11.
Geise, A.C., 1939. The effects of ultra-violet radiations of various wave-lengths upon cleavage of sea urchins.
Biol. Bull. 75, 238–247.
Geyer, R.K., Nagasawa, H., Little, J.B., Maki, C.G., 2000. Role and regulation of p53 during ultraviolet
radiation-induced G1 cell cycle arrest. Cell Growth Differ. 11, 149–156.
Gleason, D.F., Wellington, G.M., 1993. Ultraviolet radiation and coral bleaching. Nature 365, 836–838.
Gleason, D.F., Wellington, G.M., 1995. Variation in UV-B sensitivity of planula larvae of the coral Agaricia
agaricites along a depth gradient. Mar. Biol. 123, 693–703.

M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–9190
Graeber, A.G., Osmanian, C., Jack, T., Housman, D.E., Koch, C.J., Lowe, S.W., Graccia, A.J., 1996. Hypoxia-
mediated selection of cells with diminished apoptotic potential in solid tumors. Nature 379, 88–91.
Griffiths, S.D., Clarke, A.R., Healy, L.E., Ross, G., Ford, A.M., Wyllie, A.H., Greaves, M., 1997. Absence of p53
permits propagation of mutant cells following genotoxic damage. Oncogene 14, 523–531.
Halliwell, B., Gutteridge, J.M.C., 1999. Free Radicals in Biology and Medicine. Oxford Science Publications,
London. 936 pp.
Herndl, G.J., Muller-Niklas, G., Frick, J., 1993. Major role of ultraviolet-B in controlling bacterioplankton
growth in the surface layer of the ocean. Nature 361, 717–719.
Imlay, J.A., Linn, S., 1988. DNA damage and oxygen radical toxicity. Science 240, 1302–1309.
Innocente, S.A., Abrahamson, J.L.A., Cogswell, J.P., Lee, J.M., 1999. p53 regulates a G2 checkpoint through
cyclin B1. Proc. Natl. Acad. Sci. U. S. A. 96, 2147–2152.
Jokiel, P.L., York Jr., R.H., 1984. Importance of ultraviolet radiation in photoinhibition of microalgal growth.
Limnol. Oceanogr. 29, 192–199.
Karansas, J.J., van Dyke, H., Worrest, R.C., 1979. Mid-ultraviolet (UV-B) sensitivity of Acartia clausii Gies-
brecht (Copepoda). Limnol. Ocenogr. 24, 1104–1116.
Karentz, D., McEuen, F.S., Land, M.C., Dunlap, W.C., 1991. Survey of mycosporine-like amino acid compounds
in Antarctic marine organisms: potential protection from ultraviolet exposure. Mar. Biol. 108, 157–166.
Kirk, J.T.O., 1994. Optics of UV-B radiation in natural waters. Arch. Hydrobiol. Ergeb. Limnol. 43, 1–16.
Lesser, M.P., 1996. Exposure of symbiotic dinoflagellates to elevated temperatures and ultraviolet radiation
causes oxidative stress and inhibits photosynthesis. Limnol. Oceanogr. 41, 271–283.
Lesser, M.P., 1997. Oxidative stress causes coral bleaching during exposure to elevated temperatures. Coral Reefs
16, 187–192.
Lesser, M.P., Lewis, S., 1996. Action spectra for the inhibition of photosynthesis by ultraviolet radiation in the
hermatypic coral, Pocillopora damicornis. Mar. Ecol. Prog. Ser. 134, 171–177.
Lesser, M.P., Neale, P.J., Cullen, J.J., 1996. Acclimation of Antarctic phytoplankton to ultraviolet radiation: UV
absorbing compounds and carbon fixation. Mol. Mar. Biol. Biotechnol. 5, 314–325.
Lesser, M.P., Farrell, J.H., Walker, C.W., 2001. Oxidative stress and p53 expression in the larvae of Atlantic cod
(Gadus morhua) exposed to ultraviolet (290–400 nm) radiation. J. Exp. Biol. 204, 157–164.
Madronich, S., McKenzie, R.L., Bjorn, L.O., Caldwell, M.M., 1998. Changes in biologically active ultraviolet
radiation reaching the Earth’s surface. Photochem. Photobiol., B. 46, 5–19.
Malloy, K.D., Holman, M.A., Mitchell, D., Detrich III, H.W., 1997. Solar UV-B-induced DNA damage and
photoenzymatic DNA repair in Antarctic zooplankton. Proc. Natl. Acad. Sci. U. S. A. 94, 1258–1263.
Mori, T., Nakane, M., Hattori, T., Matsunaga, T., Ihara, M., Nikaido, O., 1991. Simultaneous establishment of
monoclonal antibodies specific for either cyclobutane pyrimidine dimers or (6–4) photoproduct from the
same mouse immunized with ultraviolet radiated DNA. Photochem. Photobiol. 54, 225–232.
Paulovich, A.G., Toczyski, D.P., Hartwell, L.H., 1997. When checkpoints fail. Cell 88, 315–321.
Peak, M.J., Peak, J.G., 1990. Hydroxyl radical quenching agents protect against DNA breakage caused by both
365-nm and gamma radiation. Photochem. Photobiol. 51, 649–652.
Pennington, J.T., Emlet, R.B., 1986. Ontogenetic and diel vertical migration of a planktonic echinoid larva,
Dendraster excentricus: occurrence, cause, and probable consequences. J. Exp. Mar. Biol. Ecol. 104, 69–95.
Renzing, J., Hansen, S., Lane, D.P., 1996. Oxidative stress is involved in the UV activation of p53. J. Cell Sci.
109, 1105–1112.
Rustad, R.C., 1960. Changes in the sensitivity to ultraviolet-induced mitotic delay during the cell division cycle
of the sea urchin egg. Exp. Cell Res. 21, 596–602.
Setlow, R.B., 1974. The wavelengths in sunlight effective in producing skin cancer: a theoretical analysis. Proc.
Natl. Acad. Sci. U. S. A. 71, 3363–3366.
Shick, J.M., Dunlap, W.C., 2002. Mycosporine-like amino acids and related gadusols: biosynthesis, accumula-
tion, and UV-protective functions in aquatic organisms. Annu. Rev. Physiol. 64, 223–262.
Shick, J.M., Lesser, M.P., Dunlap, W.C., Stochaj, W.R., Chalker, B.E., Wu Won, J., 1995. Depth-dependent
responses to solar ultraviolet radiation and oxidative stress in the zooxanthellate coral Acropora microphthal-
ma. Mar. Biol. 122, 41–51.
Shick, J.M., Lesser, M.P., Jokiel, P.L., 1996. Effects of ultraviolet radiation on corals and other coral reef
organisms. Global Change Biol. 2, 527–545.

M.P. Lesser, T.M. Barry / J. Exp. Mar. Biol. Ecol. 292 (2003) 75–91 91
Smith, R.C., 1989. Ozone, middle ultraviolet radiation, and the aquatic environment. Photochem. Photobiol. 50,
459–468.
Smith, R.C., Baker, K.S., 1989. Stratospheric ozone, middle ultraviolet radiation and phytoplankton productivity.
Oceanogr. Mag. 2, 4–10.
Smith, R.C., Prezelin, B.B., Baker, K.S., Bidigare, R.R., Boucher, N.P., Coley, T., Karentz, D., MacIntyre, S.,
Matlick, H.A., Menzies, D., Ondrusek, M., Wan, Z., Waters, K.J., 1992. Ozone depletion: ultraviolet radiation
and phytoplankton biology in Antarctic waters. Science 255, 952–959.