mechanisms controlling soil carbon turnover and their potential application for enhancing carbon...
TRANSCRIPT

Mechanisms controlling soil carbon turnover and theirpotential application for enhancing carbon sequestration
Julie D. Jastrow & James E. Amonette &
Vanessa L. Bailey
Received: 27 July 2005 /Accepted: 19 May 2006 / Published online: 21 December 2006# Springer Science + Business Media B.V. 2006
Abstract In addition to increasing plant C inputs, strategies for enhancing soil Csequestration include reducing C turnover and increasing its residence time in soils. Twomajor mechanisms, (bio)chemical alteration and physicochemical protection, stabilize soilorganic C (SOC) and thereby control its turnover. With (bio)chemical alteration, SOC istransformed by biotic and abiotic processes to chemical forms that are more resistant todecomposition and, in some cases, more easily retained by sorption to soil solids. Withphysicochemical protection, biochemical attack of SOC is inhibited by organomineralinteractions at molecular to millimeter scales. Stabilization of otherwise decomposable SOCcan occur via sorption to mineral and organic soil surfaces, occlusion within aggregates,and deposition in pores or other locations inaccessible to decomposers and extracellularenzymes. Soil structure is a master integrating variable that both controls and indicates theSOC stabilization status of a soil. One potential option for reducing SOC turnover andenhancing sequestration, is to modify the soil physicochemical environment to favor theactivities of fungi. Specific practices that could accomplish this include manipulating thequality of plant C inputs, planting perennial species, minimizing tillage and otherdisturbances, maintaining a near-neutral soil pH and adequate amounts of exchangeablebase cations (particularly calcium), ensuring adequate drainage, and minimizing erosion. Insome soils, amendment with micro- and mesoporous sorbents that have a high specificsurface – such as fly ash or charcoal – can be beneficial.
Climatic Change (2007) 80:5–23DOI 10.1007/s10584-006-9178-3
All authors contributed equally to this article.
J. D. Jastrow (*)Biosciences Division, Argonne National Laboratory, Argonne, IL 60439, USAe-mail: [email protected]
J. E. AmonetteChemical Sciences Division, Pacific Northwest National Laboratory, Richland, WA 99354, USAe-mail: [email protected]
V. L. BaileyBiological Sciences Division, Pacific Northwest National Laboratory, Richland, WA 99354, USAe-mail: [email protected]

1 Introduction
Accumulation of soil organic C (SOC) requires a positive imbalance between inputs to andoutputs from soil organic matter stocks. Carbon accrual can be driven by an increase inphotosynthetically derived C inputs, a decrease in C losses, or both. Decomposition,leaching, runoff, and erosion can all contribute to losses from any given location, but thelatter three processes also have the potential to add to C inputs elsewhere. Carbon seques-tration occurs when a positive disequilibrium is sustained over some period of time, withthe system eventually achieving a new, higher steady-state.
Because storage of SOC in soils is dynamic (i.e., C derived from the atmosphere isultimately cycled back to the atmosphere), the residence time (τ) of C in the soil is a majordeterminant of the capacity of a soil to sequester C (Luo et al. 2003). An increase in τ cansequester SOC even without an increase in inputs. Conversely, if τ remains unchanged thenincreased inputs can only be sequestered for as long as the mechanisms controlling τ remainunsaturated (Six et al. 2002a).
Under environmental extremes of moisture, temperature, pH, or nutrient availability, τ islargely controlled by how these factors limit decomposer activity. Inputs may besequestered seemingly without constraint in a relatively undecomposed and uncomplexedstate (e.g., boreal peat deposits) but the accumulated C is vulnerable to release from storageif environmental conditions become more moderate (Freeman et al. 2001). Fire is anothersignificant factor affecting τ, particularly in grassland and savanna soils and peat deposits.Although the primary effect of fire is the rapid loss of aboveground C, the ultimate effectson SOC vary (Johnson and Curtis 2001). The generation of charcoal sequesters C directlyand enhances soil fertility (Dai et al. 2005; Glaser et al. 2002). Furthermore, charcoal mayplay a beneficial role in the stabilization of SOC (Amonette et al. 2003a).
Under more tempered environmental conditions, two major mechanisms – (bio)chemicalalteration and physicochemical protection – are responsible for stabilizing SOC and therebycontrolling τ. With (bio)chemical alteration, SOC is transformed by biotic and abioticprocesses to chemical forms that are more resistant to decomposition and, in some cases,more easily retained by sorption to soil solids. The alteration process is generally referred toas humification, and the altered products as humic materials. With physicochemicalprotection, biochemical attack of SOC is inhibited by organomineral interactions atmolecular to millimeter scales. Stabilization of otherwise decomposable SOC can occur viasorption to soil surfaces, complexation with soil minerals, occlusion within aggregates, anddeposition in pores inaccessible to decomposers and extracellular enzymes. Both humifiedand non-humified SOC can be protected, although the greatest impact on τ is likely to arisefrom protection of humified SOC. Because of the specific nature of the protective sites,either transport of SOC to these sites or reorientation of mineral components around SOC isrequired for stabilization. The relative importance and potential saturation of these twostabilization mechanisms vary depending on soil type, vegetation, management practices,and environmental conditions.
Although increasing C inputs to soil organic matter is an important component ofstrategies for enhancing SOC sequestration, we focus in this paper on increasing τ. Becauseturnover is primarily a biological process, we first consider the overall role of the soil bioticcommunity and then briefly review how (bio)chemical alteration and physicochemicalprotection mechanisms control the stabilization of SOC. Lastly, we discuss how anunderstanding of the integrated functioning of these mechanisms within the context of soilstructure suggest specific manipulations to increase the residence time of C in soils and,thereby, promote SOC sequestration.
6 Climatic Change (2007) 80:5–23

2 Soil biota and carbon stabilization
The microbial community in soils is diverse, abundant, and only partly understood (Gans etal. 2005); it is often described as a “black box” (Tiedje et al. 1999). Some researchersestimate that as much as 90–99% of the soil microbial community is unculturable bycurrent technologies (Hill et al. 2000). Therefore, we are limited in our ability toconfidently and absolutely ascribe soil processes to specific microorganisms. For thepurpose of discussing biogeochemical processes, it is easier to consider the community interms of the greater biological domains, (e.g., fungi, bacteria, actinomycetes) or in terms ofbiochemical functions and specific processes (e.g., enzyme activities).
In general, two of the dominant groups of soil microorganisms, fungi and bacteria, havesubstantially different biochemistries with consequent effects on SOC stabilization. A clearexample, but by no means the exclusive cause of different sequestration potentials, is thedifference in C utilization efficiency (CUE). Although some researchers have reported thatCUE may not differ between fungi and bacteria (Payne 1970), others have found bacterialCUE to be lower than fungal CUE (Adu and Oades 1978; Claus et al. 1999). Organismswith a lower CUE respire a greater proportion of metabolized C as CO2. Thus, inmetabolizing the same amount of C, bacteria with a lower CUE would contribute less topools of newly stabilized SOC than fungi. However, as with most relationships detected insoils, these observations are highly dependent upon the soils studied, and generalizationsmust be interpreted broadly.
Fungi and bacteria also differ in the nature of the extracellular enzymes they produce.Fungi produce large amounts of phenoloxidases, laccases, and peroxidases that help attacklignitic materials and promote condensation reactions. Bacteria, on the other hand, are morelikely to produce lipases and cellulases needed to attack nonlignitic materials. Themonomers derived from degradation of lignitic materials are perhaps the most commonconstituents of humic materials and, thus an abundance of fungal enzymes should tend tofavor humification.
A primary contribution of fungi to C stabilization derives from the very nature of theirmetabolism and biomass formation. Fungal cell walls are constructed of complex moleculessuch as melanin and chitin, in contrast to bacterial membranes (which are dominantlyphospholipids). After cell death, phospholipids are rapidly metabolized by other bacteria,and overall these bacterial products are more vulnerable to grazers (Frey et al. 2001).Melanin and chitin residues are more recalcitrant and tend to persist in soils (Guggenbergeret al. 1999; Holland and Coleman 1987). For example, when microbes were grown on leaflitter, the amount of C stored by fungi was 26 times greater than the C stored by bacteria(Suberkropp and Weyers 1996).
The fungi discussed so far are largely saprotrophic in nature, i.e., they acquire Cthrough degradation of complex organic compounds. A second subset of fungi,mycorrhizal fungi, also play a distinct and unique role in C sequestration. Although,mycorrhizal fungi are not directly involved in the decomposition of soil organic matter;they are obligate symbionts that coexist with most terrestrial plants. In this relationship, theplant gains nutrients gathered from the greater soil volume explored by mycorrhizalhyphae, and the fungus obtains photosynthate-C directly from the plant. Thus, the primarycontribution of mycorrhizal fungi to C sequestration is as an additional source of C inputsto soil organic matter. This occurs, indirectly, through the generally positive effect of themycorrhizal symbiosis on plant growth, particularly in low phosphorus soils. However,with C standing stocks of 54–900 kg C ha−1 and cell walls composed largely of chitin, theturnover of mycorrhizal fungi also can be a significant direct source of C inputs that are
Climatic Change (2007) 80:5–23 7

relatively resistant to decomposition (Zhu and Miller 2003). Another form of C inputsproduced by mycorrhizal fungi is a glycoprotein called glomalin that appears to besomewhat resistant to degradation, perhaps due to interactions with soil minerals (Steinbergand Rillig 2003). In addition, mycorrhizal fungi contribute indirectly to SOC stabilizationvia their role in the formation and stabilization of soil aggregates (Jastrow et al. 1998;Miller and Jastrow 1990; Tisdall 1996; Tisdall and Oades 1982). Mycorrhizal hyphaetogether with fine roots create a “sticky-string bag” that enmeshes and entangles soilparticles, helping to stabilize macroaggregates (Miller and Jastrow 2000; Oades and Waters1991). The deposition of glomalin may also contribute to aggregate stabilization (Rillig2004), although this hypothesis is still subject to scrutiny (Franzluebbers et al. 2000).Lastly, mycorrhizal fungi are hypothesized to further enhance C sequestration bytranslocating nutrients from the bulk soil to the host plant, thereby competing with free-living decomposer microorganisms that would otherwise mineralize SOC to CO2 (Rilligand Allen 1999).
Our consideration of the potential significance of fungi to C sequestration is not tominimize the role played by bacteria in these processes. On the contrary, bacteria have beenextensively studied, and most of what is currently known about SOC dynamics and storageis derived from studies centered on bacteria (e.g., Hu et al. 1999; Marilley et al. 1999; Pelzet al. 1998; Schimel et al. 1999). We highlight the role of fungi in this paper as an emergingarea of research and one that merits deeper study. Several recent reports have calledattention to the distinct roles played by fungi in the C cycle. For example, fungal activitywas correlated with soil C content (Bailey et al. 2002); shifts in mycorrhizal communitystructure (Treseder and Allen 2000) and increased grazing of fungi (Allen et al. 2005) werefound in systems exposed to elevated atmospheric CO2 concentrations; and increases infungal biomass were observed immediately following land abandonment and in associationwith undisturbed, high-C heathlands (van der Wal et al. 2006).
In addition to fungi and bacteria, soil invertebrates play important roles in thefragmentation, comminution, and decomposition of plant litter and – through grazing andcomplex food web interactions – exert significant controls on microbial communitystructure, biomass, and activities (Beare et al. 1995; Mikola et al. 2002). Further, significanttransformations of SOC are performed by microbes living within the guts or excrement ofsoil invertebrates (Hättenschwiler et al. 2005; Martin and Marinissen 1993; Seastedt 1984).
3 (Bio)chemical alteration
In most soils, C stabilization necessarily involves alteration of organic matter to chemicalforms that are more recalcitrant to microbial attack, more likely to adsorb to soil solids, orboth. The alteration typically occurs in two stages. First, plant-derived residues arefragmented into particulate organic matter and then decomposed into smaller molecules.Similarly, the C assimilated by decomposers and used for growth is also decomposed whenthese organisms turn over. The decomposition stage is followed by condensation andpolymerization reactions that create new larger molecules from the small moleculesreleased during decomposition. Both stages of alteration are largely biologically driven,although there is also significant evidence for abiotic condensation and polymerizationreactions at mineral surfaces. Consequently, the nature of the soil biotic community and themineral phases present in the soil are key factors in the alteration process.
8 Climatic Change (2007) 80:5–23

3.1 Decomposition
After physical fragmentation to particulate organic matter, the first stage of (bio)chemicalalteration involves the decomposition of C inputs (chiefly cellulose and lignin biopolymerspresent in plant litter) into smaller molecules. This process is mediated principally bymicroorganisms and their extracellular enzymes. Cellulose is readily degraded by bothfungi and bacteria. Lignin, however, is more recalcitrant, and its complete degradation isrestricted to a select group of fungi that produce the extracellular lignin peroxidases(Conesa et al. 2002; Schubert 1965; ten Have and Teunissen 2001). As yet, no bacteria havebeen identified that are able to initiate degradation of lignin, and current knowledgesuggests that they can only mineralize the partly transformed intermediates produced by thefungi. For a given cohort of C inputs, selective degradation by fungi and bacteria willdecompose the labile portions first and lead to a progressive increase in the averagerecalcitrance of the remaining SOC with time.
Decomposition of themore recalcitrant C inputs relies heavily on the activity of extracellularenzymes, which have a limited lifetime in soils (on the order of days; e.g., Amonette et al.2003a, b; Dick and Tabatabai 1992; Shen et al. 2002). These enzymes typically sorb to soilsolids, with varying effects on their ability to catalyze decomposition. Although the effect ofsorption is enzyme- and mineral-specific, in many instances the enzyme activity decreasesinitially as a result of sorption, but longevity and total activity over the life of the enzymeincrease (Boyd and Mortland 1990; Shen et al. 2002). Enzyme sorption to minerals, however,also limits access to potential substrates (particularly the large insoluble biopolymers) andrestricts decomposition to regions near the source of the enzyme (George et al. 2005).
Sorption of the enzyme to the substrate is the key process. Schimel and Weintraub(2003) suggested that a limited number of sorption sites exist on a substrate surface; hence,once these sites are saturated by enzymes, additional enzymes must diffuse farther from thesource and return less product to the microorganism that synthesized them. This negativefeedback loop (simulated by a reverse Michaelis–Menton relationship) regulates theproduction of extracellular enzymes by microorganisms. One model of extracellularenzyme production by bacteria suggests that the typical extracellular enzyme foragingdistance is on the order of 10–50 μm (Vetter et al. 1998). At longer distances, the benefit tothe organism does not exceed the cost of producing the enzyme.
The limited zone of extracellular enzymatic activity is expanded, particularly inundisturbed soils, by fungi whose hyphal growth habit allows them to form “bridges”between the soil and surface litters (Beare et al. 1992; Holland and Coleman 1987). Inrelatively undisturbed systems, these hyphal networks can become a large component of thesoil fabric (Haynes and Beare 1997). Because the fungal biomass is made up of complexpolymers such as chitin and melanin (Guggenberger et al. 1999; Holland and Coleman1987) that are relatively resistant to degradation, its senesced residues contribute to soilorganic matter (West et al. 1987). In contrast, bacteria exhibit limited mobility in soils andthus seem to be restricted to using substrates located within their immediate proximity(Wolfaardt et al. 1994). Moreover, bacterial biomass is comprised of labile energy-richmolecules such as phospholipids and amino acids (Guggenberger et al. 1999). Thussenesced bacterial residues do not accumulate or contribute significantly to SOC in theirunaltered state (West et al. 1987).
In addition, wetting and drying cycles may have an effect on microbial communitystructure and, consequently, decomposition rates. Schimel and co-workers have shownsignificant long-term decreases in respiration for a grassland soil and for birch leaf litter
Climatic Change (2007) 80:5–23 9

subjected to repeated wetting and drying cycles (Clein and Schimel 1994; Fierer and Schimel2002). The relationship is by no means robust, however. Increases in respiration were seenin a soil formed under perennial oak vegetation (Fierer and Schimel 2002), whereas longperiods (e.g., six months) between cycles showed no effect (Haider and Martin 1981).
3.2 Condensation/polymerization
In the second stage of (bio)chemical alteration, new compounds form from some of themolecules released during the decomposition stage. These compounds are condensates of avariety of different “monomers” and, as a consequence, do not have well-defined compo-sition or structure. Indeed, there is increasing evidence to suggest that the condensates mayconsist of dynamic clusters of chemically altered and unaltered compounds held togetherloosely by hydrophobic interactions and hydrogen bonding (Sutton and Sposito 2005). Ingeneral, the condensates have higher aromatic character and oxygen content than theaverage biomass present in soils (Hayes and Malcolm 2001). When compared to plant litter,the condensates have higher levels of carboxylic and fatty-acid C and lower levels of poly-saccharide C (Chefez et al. 2002; Zech et al. 1997). Their most important attribute, however,is that they are more recalcitrant to decomposition, either as a result of their intrinsic bio-chemical properties or enhanced sorption affinities. As shown in Fig. 1, consensus hasemerged among researchers that the chemically altered fraction of the condensates areformed by the reactions of amino compounds (acids and sugars) with quinones or reducingsugars to form melanin-type compounds (Flaig 1975; Haider et al. 1975; Hedges 1988;Kononova 1961; Maillard 1916; Martin and Haider 1971; Martin et al. 1975; Stevenson1994; Tan 2003; Waksman 1932). The amino compounds and reducing sugars are readilyavailable from the lysing of microorganisms, whereas the quinones are the result of oxidationof polyphenols derived from lignin and other plant materials as well as from microorganisms.
The condensation of reducing sugars with amino compounds proceeds spontaneously butslowly at typical soil temperatures, and is greatly enhanced by repeated wetting and dryingcycles. Many investigators, however, consider the quinone/amino-compound condensationreaction to be the dominant humification pathway in soils (Stevenson 1994). The rate-determining step for this reaction is believed to be the oxidation of the polyphenol to formthe quinone. The reaction is faster at high pH, and at the near-neutral pH of most soilsassistance from a catalyst is needed for the reaction to proceed at a measurable rate. Bio-logical catalysts include polyphenol oxidase, peroxidase, and laccase enzymes produced byfungi, which mediate the electron transfer from the polyphenol to molecular oxygen (Sjobladand Bollag 1981, Tate 1992). Other substances in soils either catalyze the reaction – e.g.,amorphous silica, charcoal, iron-sorbed smectite (Amonette et al. 2003a, b, 2004; Booth et al.2004) – or serve as oxidants – e.g., oxides and hydroxides of manganese and iron, smectites(Amonette et al. 2000; Naidja et al. 1998; Shindo and Huang 1984; Wang and Huang 2005).Recent evidence suggests the overall reaction rate increases synergistically when bothbiological and inorganic catalysts are present (Amonette et al. 2000, 2003a, b).
Ultimately, the availability of molecular oxygen determines whether the polyphenoloxidation reaction occurs, as this species also oxidizes the metal oxides and phyllosilicates.Indeed, insufficient oxygen is likely to stop the quinone/amino-compound condensationreaction from occurring in soils, just as it prevents the enzymatic decomposition of peatbogs (Freeman et al. 2001). Too much oxygen, however, and the monomers and newlyformed condensates will continue to be altered until their C is fully oxidized to CO2. In
10 Climatic Change (2007) 80:5–23

contrast to the oxidation step, the subsequent condensation reaction with the amine groupoccurs spontaneously, although many factors (primarily pH) can affect its rate.
The mineral phases in soils can have an important impact on (bio)chemical alteration,principally as catalysts of condensation and as oxidants. The dominant minerals in soils ona mass basis are typically quartz and the feldspars, but these minerals have very lowspecific surfaces (ca. 0.1 m2 g−1) and correspondingly low impact on chemical alterationsof soil organic matter. Minerals with high specific surfaces (>10 m2 g−1) and high chemicalimpact on alterations include the phyllosilicate clays, allophanes, and the oxides andhydroxides of manganese and iron. The phyllosilicate clays can be grouped according towhether they are swelling (e.g., smectites, vermiculites) or non-swelling (e.g., kaolinite,illite), with swelling clays offering internal surfaces as high as several hundred squaremeters per gram. Although some studies have shown correlations between clay content (orclay plus silt) and the amount of soil organic matter (e.g., Six et al. 2002a, b), the surfacereactivity and specific surface of soil minerals appear to be better predictors of SOC(Baldock and Skjemstad 2000).
4 Physicochemical protection
Protection of (bio)chemically altered soil organic matter from further microbial decompo-sition or oxidation by molecular oxygen and extracellular enzymes is essential tosignificantly lengthen the residence time of C in soils. For protection of new C to occur,some change in the arrangement between this C and other soil particles must occur.Rearrangement can occur in a number of ways but the end state generally involves chemicalor physical sorption of the new C to an existing surface coupled with some sort of physical
Fig. 1 Major pathways for hu-mification showing C inputs andthe types of condensation reac-tions that are likely to occur.Pathways are (a) sugar-amine,(b) polyphenol-quinone, (c) lig-nin-quinone, and (d) modified-lignin (after Stevenson 1994,p. 189).
Climatic Change (2007) 80:5–23 11

barrier to prevent further access by agents that would decompose or oxidize the C. If notalready at a surface, the new C may diffuse or advect to a surface where it can be adsorbed.Alternatively, the soil particles may change position to become reoriented in associationwith SOC as a result of advection, the mechanical actions of plant and fungal growth,bioturbation by earthworms and other soil fauna, or changes in hydration status.
4.1 Chemical protection
Relatively labile organic compounds (e.g., microbial byproducts) as well as humified C canbe protected by strong chemical associations with soil minerals. Sorption occurs via avariety of organomineral associations, such as polyvalent cation bridges, hydrogen-bonding, van der Waals forces, and interactions with hydrous oxides and aluminosilicates.Drying also appears to play an important role in increasing the stability of organomineralassociations by enhancing hydrogen bonds in addition to polymerizing and denaturing theorganics. Thus, many sorption and complexation interactions are possible depending onvarious factors, including the chemical characteristics of the soil organic matter, thereactivity and specific surface of soil minerals, base-cation status, presence of iron andaluminum oxides, pH, and redox conditions (for overviews see Baldock and Skjemstad2000; Blanco-Canqui and Lal 2004; Oades 1984; Sollins et al. 1996).
Clay mineralogy plays an important role in the chemical protection of SOC (Baldockand Skjemstad 2000; Dalal and Bridge 1996; Sollins et al. 1996). In soils dominated by 2:1clay minerals, such as smectite and illite, complexation is believed to occur via theformation of multivalent cation bridges between negatively charged organic groups andnegatively charged clay platelets. In contrast, for soils dominated by 1:1 clays (kaolinite)and iron- and aluminum-oxides, surface-charge densities are generally lower and offerfewer sites for complexation with organics.
Thus, the availability of multivalent cations – particularly calcium, iron, aluminum, andmanganese – is an important factor in the chemical protection of SOC. In addition to theirrole as bridging agents, high saturation of clays with multivalent cations helps to keeporganomineral complexes more flocculated and condensed, thereby reducing the efficiencyof attack by microbes and enzymes (Baldock and Skjemstad 2000). This mechanism maybe particularly important in soils with low-charge clays.
Variations in specific surface are important to chemical protection as they determinepotential sites for interactions (Baldock and Skjemstad 2000; Kaiser and Guggenberger2003; Sollins et al. 1996). Specific surface, which can range from ca. 0.1 to several hundredsquare meters per gram, varies with clay mineralogy and also decreases with increasingparticle size. Thus, sandy or silty soils with small amounts of high-specific-surface clayscan likely sorb more C than a similarly textured soil with low-specific-surface clays.
Sorption does not appear to occur uniformly across mineral surfaces, but rather has beenshown to exist in patches (Baldock and Skjemstad 2000; Kaiser and Guggenberger 2003).The most reactive sites may be located at the mouths of micropores between two domainsof a mineral, where multiple organomineral attachments can occur. Sorption at such sites ishypothesized to contribute to observed variations in turnover rates for mineral-associatedSOC (Kaiser and Guggenberger 2003). The first molecules sorbed at these sites are the beststabilized, but after these pore openings are filled, additional organic matter loading is lessstrongly sorbed.
12 Climatic Change (2007) 80:5–23

4.2 Physical protection
Substantially greater protection is obtained when decomposer access to chemicallyprotected substrates is physically impeded or when soil structural controls on gas exchangeand moisture conditions inhibit decomposer activity (Elliot and Coleman 1988; Elliot et al.1980; Six et al. 2002a; Sollins et al. 1996; Young and Ritz 2000). Physical protectionmechanisms are intimately tied to the processes of aggregate turnover and stabilization atmultiple scales. Aggregates are formed and degraded as a result of physical processesincluding wetting and drying, freezing and thawing, the growth of plant roots andassociated mycorrhizal fungi (including physical entanglement and localized drying due towater uptake), bioturbation (i.e., the activities of earthworms, termites, ants, and other soilfauna), and tillage or other physical disturbance (Allison 1968; Dexter 1988; Kay 1990;Oades 1993; Six et al. 2004). In many soils, the deposition and transformation of organicmatter plays a major role in aggregate stabilization, and strong feedbacks exist betweenaggregate turnover and SOC dynamics (Dalal and Bridge 1996; Feller and Beare 1997;Jastrow and Miller 1998; Oades 1993; Six et al. 2004, 2002b; Tisdall 1996).
In soils where organic matter is a major aggregate-binding agent, plant growth anddecomposition of organic inputs (especially roots) lead to the development of a hierarchicalaggregate structure (Fig. 2), where the mechanisms of aggregate formation and stabilizationand their relative importance change with spatial scale (Jastrow et al. 1998; Oades andWaters 1991; Tisdall and Oades 1982). Primary particles and extremely stable fine-silt-sizedaggregates (<20 μm diameter) are bound together into microaggregates (20–250 μmdiameter) by plant and microbial debris and by humic materials or polysaccharide polymersstrongly sorbed to clays, with the most persistent clay–organic associations strengthened bypolyvalent cation bridges. Microaggregates along with silt-sized aggregates and primaryparticles are bound into macroaggregates (>250 μm diameter) by labile organic materialsand by fine roots, fungal hyphae, bacteria, and algae.
By its very nature, aggregate hierarchy creates a parallel hierarchy of pores between andwithin aggregates of varying sizes that control gas exchange, water movement, and soilfood web structure and dynamics (Elliot and Coleman 1988; Young and Ritz 2000). Thus,the nature and effectiveness of various aggregate-binding agents depend on their physicaldimensions relative to the dimensions of the pores (i.e., planes of weakness) being bridged,and smaller aggregated units are generally more stable and less susceptible to disturbance(Dexter 1988; Jastrow et al. 1998; Kay 1990; Tisdall and Oades 1982).
Current evidence indicates that microaggregates are formed inside macroaggregates(Fig. 3), particularly as root-derived particulate organic matter is decomposed (Angers et al.1997; Gale et al. 2000; Golchin et al. 1994; Six et al. 2000). Similarly, silt-sized aggregatesmay be stabilized (Fig. 3) by the deposition of microbial byproducts and condensation ofhumates to form complexes with mineral particles and decomposing organic debris insidemicroaggregates (Golchin et al. 1994). Disturbance and other factors that increase the rate ofmacroaggregate turnover can, thereby, decrease the formation and stabilization of micro-aggregates (Six et al. 2000).
Several lines of evidence indicate that protection of SOC by microaggregates is greaterthan the protection afforded by macroaggregates. The amount of C mineralized fromcrushed or dispersed aggregates in laboratory incubation studies is often greater formicroaggregates than for macroaggregates (Bossuyt et al. 2002; Gregorich et al. 1989). Intracer studies, labeled C preferentially accumulated in microaggregates compared to otherlocations within the soil (Angers et al. 1997; Besnard et al. 1996; Denef et al. 2001; Gale et al.
Climatic Change (2007) 80:5–23 13

2000). Furthermore, for soils exhibiting aggregate hierarchy, estimates of the mean residencetime of C in macroaggregates are shorter than for C in microaggregates (Six and Jastrow2002).
Although the amount of long-term protection provided by macroaggregates appears tobe minimal, the rate of macroaggregate turnover plays an important role in the stabilizationof SOC. Some level of macroaggregate turnover may be necessary to incorporate andprotect fresh C inputs from rapid mineralization (Plante and McGill 2002). Yet, if macro-aggregate turnover is too rapid, microaggregate formation and stabilization can be inhibited,leading to a reduction in the amount of microaggregate-protected SOC – especially micro-aggregate-occluded particulate organic matter (Six et al. 2004).
In addition to the physical barriers created by the encrustation and occlusion of soilorganic matter with soil mineral components, the size distribution and tortuosity of pores(which is related to aggregate size distribution and hierarchy) also plays an important role inthe physical protection of SOC. Pore size, and in particular pore neck size, controls thedistribution, movement and activity of decomposers and soil food web dynamics (Elliot andColeman 1988; Elliot et al. 1980; Young and Ritz 2000). Labile C substrates filling poressmaller than 1 μm or located inside larger pores with necks less than 1 μm are notaccessible to most soil microorganisms. Similarly, the diffusion of extracellular enzymesinto micron-sized pores can be limited and is completely excluded from nano-scale pores(Zimmerman et al. 2004). Pore size distributions and, hence, enzyme accessibility at thisscale differ with clay mineralogy. For example, pore size in clay tactoids of Ca-saturated illite is∼10 nm compared to ∼100 nm in kaolinite (Dalal and Bridge 1996).
Decomposer activity is also limited by localized oxygen and water availability. Steepdeclines in oxygen concentrations have been measured within small distances fromaggregate surfaces, and interactions between water films and small pore necks can lead to
Fig. 2 Conceptual diagram of aggregate hierarchy illustrating microaggregates inside a macroaggregate(modified from Jastrow and Miller 1998, p. 209).
14 Climatic Change (2007) 80:5–23

anaerobic patches within largely aerated aggregates (Sexstone et al. 1985; Young and Ritz2000). Recent work by Strong et al. (2004) demonstrated initial rates of decompositionwere most rapid in soils with large volumes of intermediate-sized pores of about 15–60 μm.In particular, decomposition was enhanced near the gas–water interface, most likelybecause these locations optimize (1) organism motility; (2) the diffusion of nutrients, toxins,and enzymes; and (3) oxygen supply. In larger pores (60–300 μm) oxygen was abundant,but decomposition was slower – probably because motility, diffusion, and direct contactbetween residues and mineral surfaces were all reduced. However, the greatest protectionand slowest rates of decomposition were found in soils that had large volumes of pores withneck diameters <4 μm.
Thus, the interactions of the microbial community with the complexity of pore space andsurfaces that compose their physical habitat leads to protection of SOC via what has beentermed “partial refuge” (Ekschmitt et al. 2005). In essence, this means that not alldegradable SOC is within range of decomposing organisms all the time because of (1) thelimited zones of microbial habitats – estimated by Nannipieri et al. (2003) to be <5% of theoverall space in soils, (2) the limited foraging distances for extracellular enzymes (Schimeland Weintraub 2003; Vetter et al. 1998), and (3) the specificity of enzyme and substratechemistries. Hence, the enzyme needed to decompose a specific functional group may notbe available because the right habitat for the appropriate microorganism is too far from thesubstrate. Although the difference can be subtle at some scales, decomposition is preventedby the effect of physical structure on local biological activity rather than by physicalocclusion of the substrate. This situation, however temporary, slows the overall degradationof SOC and thereby increases its residence time.
Fig. 3 Conceptual model ofmicroaggregate formation andturnover (based on Gale et al.2000, Golchin et al. 1994, Sixet al. 2000). Illustrated stages are(a) fragmented and decomposingroots and litter become encrustedwith mineral particles formingmicroaggregates; (b) decomposi-tion continues but at a slowerrate due to physical protec-tion; microaggregate is stabi-lized; (c) slowed decompo-sition and intimate contactwith soil minerals in stablemicroaggregate enables organicmatter to be humified orchemically protected by asso-ciation with mineral fraction;(d) organic binding agents de-compose sufficiently for aggre-gate to be destabilized; mineralfraction enriched with neworganomineral associationsbecomes available for incorpo-ration into new microaggregates.
Climatic Change (2007) 80:5–23 15

5 Soil structure and the stabilization of soil organic carbon
A master integrating variable that both controls and indicates the C stabilization status of asoil is soil structure (i.e., the arrangement of solids and pores in the soil). Soil structureinfluences (1) infiltration, runoff, drainage, and thus aeration, moisture availability, anderosion; (2) affects the availability of substrates, nutrients and other solutes; (3) controlssolute transport; and (4) serves as the habitat (Fig. 4) for roots, rhizosphere organisms, andthe decomposer community (Christensen 2001; Elliot and Coleman 1988; Elliot et al. 1980;Young and Ritz 2000). Consequently, soil structure plays important roles in both plantgrowth and the regulation of decomposer activity and, therefore, the cycling andstabilization of SOC.
The spatial organization of soil structure is hierarchical, with structural complexityaffecting functional organization and process (Fig. 5). As structural organization increasesin both scale and complexity – from the primary structures of organomineral complexes tothe architecture of the soil profile – progressively more dynamic responses toenvironmental perturbations and management practices occur.
Primary structure is a product of long-term soil forming factors, especially parentmaterial, climate, and topography (Jenny 1941), and secondary structure is determined bythe binding of primary particles into soil aggregates of varying sizes. Secondary structure isdynamic over much shorter time scales than primary structure with aggregate formation andturnover affected by many factors – including climate, vegetation, decomposer activity, soilmineralogy, base-cation status, and management practices (see overviews by Jastrow and
Fig. 4 Conceptual drawing of a cross-section (1 cm2) of a highly structured and biologically active soilmicrosite illustrating how different types of pore space and the water distribution within pores influence thehabitat and feeding relationships among various soil organisms (from Elliot and Coleman 1988, p. 27).
16 Climatic Change (2007) 80:5–23
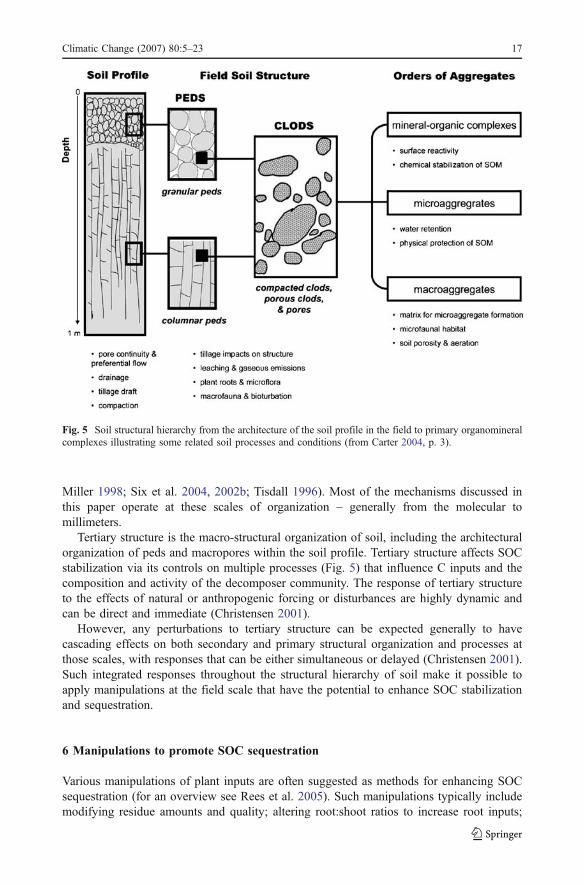
Miller 1998; Six et al. 2004, 2002b; Tisdall 1996). Most of the mechanisms discussed inthis paper operate at these scales of organization – generally from the molecular tomillimeters.
Tertiary structure is the macro-structural organization of soil, including the architecturalorganization of peds and macropores within the soil profile. Tertiary structure affects SOCstabilization via its controls on multiple processes (Fig. 5) that influence C inputs and thecomposition and activity of the decomposer community. The response of tertiary structureto the effects of natural or anthropogenic forcing or disturbances are highly dynamic andcan be direct and immediate (Christensen 2001).
However, any perturbations to tertiary structure can be expected generally to havecascading effects on both secondary and primary structural organization and processes atthose scales, with responses that can be either simultaneous or delayed (Christensen 2001).Such integrated responses throughout the structural hierarchy of soil make it possible toapply manipulations at the field scale that have the potential to enhance SOC stabilizationand sequestration.
6 Manipulations to promote SOC sequestration
Various manipulations of plant inputs are often suggested as methods for enhancing SOCsequestration (for an overview see Rees et al. 2005). Such manipulations typically includemodifying residue amounts and quality; altering root:shoot ratios to increase root inputs;
Fig. 5 Soil structural hierarchy from the architecture of the soil profile in the field to primary organomineralcomplexes illustrating some related soil processes and conditions (from Carter 2004, p. 3).
Climatic Change (2007) 80:5–23 17

modifying root depth distributions, morphology, architecture, and quality; altering mycor-rhizal associations; manipulating lifeform or species mixtures (e.g., grasses, legumes, trees;perennial vs annual; C3 vs C4 species) and the timing of inputs.
Here we focus on and summarize key factors controlling the accumulation of C in soilsvia an increase in τ and draw some conclusions about the types of manipulations that havethe potential to enhance this process. At the highest level, an increase in C residence timerequires increases in (1) the net rate of (bio)chemical alteration to recalcitrant forms, and(2) the degree of physicochemical protection of this newly altered C.
Seemingly obvious manipulations for enhanced C sequestration would include shiftingthe soil microbial community towards populations with increased CUE and humifyingcharacteristics, such as are found in certain fungi. This is not an easy task as it is difficult tointroduce and monitor microorganisms into soils with lasting effect. With currenttechnologies, better options are to (1) manipulate plant C inputs to enhance fungal growthand humification and (2) modify the soil physicochemical environment to directly enhanceC sequestration and indirectly favor the activities of microorganisms that enhance Csequestration.
As the rate-limiting step in (bio)chemical alteration seems to be the oxidation ofpolyphenolic monomers in most soils, manipulations focused on optimizing this stage inthe process are most likely to succeed. Specific goals include maximizing the availabilityand activity of phenol oxidase and related enzymes, while minimizing the availability andactivity of hydrolase enzymes, and ensuring adequate, but not excessive, levels of oxygen.The availability of phenol oxidase and related enzymes is determined in large part by theabundance of the fungi that synthesize them. The activity of these enzymes is optimal atnear-neutral pH and enhanced synergistically by their sorption to micro- and mesoporousminerals (oxidizing and non-oxidizing), including coal and charcoal. The availability ofhydrolase enzymes probably is more closely related to the abundance of soil bacteria,although clearly both fungi and bacteria require hydrolases for their survival. We know littleabout the effect of sorption on the activity of hydrolases, although it would be expected tofollow the same trend as the oxidase activity. The levels of oxygen in soil depend onmoisture content, depth, and pore structure. Avoidance of excessively wet or dry conditionsand minimizing disturbance of soil structure will promote the optimal oxygen levels for(bio)chemical alteration and at the same time enhance the growth of fungi. Specificpractices that could promote (bio)chemical alteration include minimizing tillage, maintain-ing soil pH near neutral, and ensuring adequate drainage. In some soils, this approach canbe further augmented by amendments with various high-specific-surface micro- andmesoporous sorbents (such as fly ash or charcoal) that stabilize enzymes and improvemoisture retention properties.
Manipulations to enhance the other primary (bio)chemical mechanism of humification(condensation of reducing sugars with amino compounds) involve maximizing the numberof wetting and drying cycles. In humid-zone agricultural soils, this might be achieved bycontrolling the flow from drainage tile systems. With irrigated agricultural soils, man-agement would involve timing irrigations to coincide with actual soil moisture conditionsthereby ensuring that the requisite degree of dryness is achieved, rather than irrigating on afixed schedule. The degree of drying required for maximum effect may vary from soil tosoil depending on the soil structure. Research to clarify the optimal conditions is needed.
With respect to enhancing physicochemical protection, the primary goals involvemaximizing the stability of soil microaggregates while ensuring the optimum rate of soilmacroaggregate turnover to allow incorporation and stabilization of new SOC. Somespecific practices include minimum tillage or planting perennials (particularly species with
18 Climatic Change (2007) 80:5–23

high-density fibrous root systems, such as grasses), maintenance of adequate amounts ofexchangeable base cations (particularly calcium), and, in some soils, amendments withmicro- and mesoporous solids to create a more favorable pore-size distribution. Practicesthat prevent erosion – such as cover crops, adequate drainage, protection from wind, andsustainable grazing practices – are also critical to maintenance of soil structure, and thereby,physicochemical protection of SOC.
Although we have focused strictly on increasing the residence time of C in soils, thepotential effects of any manipulation on the N status of the soil also should to be taken intoaccount, as the C and N cycles are very closely coupled (Schimel and Bennett 2004;Schimel and Weintraub 2003; Stevenson and Cole 1999). The availability or lack of oneelement drives the mineralization or immobilization of the other. For example, the releaseof N2O caused by over fertilization or maintenance of wet conditions can more than offsetany gains made from SOC stabilization.
In summary, the health of the microbial community is critical to the stabilization of SOC.Manipulations that enhance the fungal community and (bio)chemistry associated with thehumification process have the potential to increase the sequestration of C in soils. Chiefamong these is minimizing disturbance of the soil by tillage or erosion. Water managementis also important, with the goal being to cycle frequently between field moist and dryconditions and avoid long-term (i.e., greater than a few days) exposure to either extreme.The soil pH should be maintained near neutral, and adequate calcium should be present onthe cation exchange complex. Amendments with micro- and mesoporous materials such asfly ash and charcoal are recommended for soils with poor structure and low exchangecapacities. Given current knowledge, these recommendations appear to be promising meansfor reducing SOC turnover and increasing its residence time for the purpose of enhancingsoil C sequestration.
Acknowledgments We thank four anonymous reviewers for comments on an earlier version of themanuscript. This work was supported by the US Department of Energy, Office of Science, Office ofBiological and Environmental Research, Climate Change Research Division under contract W-31-109-Eng-38to Argonne National Laboratory and contract DE-AC05-76RL01830 to Pacific Northwest NationalLaboratory.
References
Adu JK, Oades JM (1978) Utilization of organic materials in soil aggregates by bacteria and fungi. Soil BiolBiochem 10:117–122
Allen MF, Klironomos JN, Treseder KK, Oechel WC (2005) Responses of soil biota to elevated CO2 in achaparral ecosystem. Ecol Appl 15:1701–1711
Allison FE (1968) Soil aggregation – some facts and fallacies as seen by a microbiologist. Soil Sci 106:136–143
Amonette JE, Capp JA, Lüttge A, Baer DR, Arvidson RS (2000) Geochemical mechanisms in terrestrialcarbon sequestration. In: Annual report 1999, environmental dynamics and simulation, PNNL-13206/UC-400. Pacific Northwest National Laboratory, Richland, Washington, pp 3-23 to 3-27
Amonette JE, Kim J, Russell CK (2004) Enhancement of soil carbon sequestration: a catalytic approach.Preprint paper – Am Chem Soc, Div Fuel Chem 49:366–367
Amonette JE, Kim J, Russell CK, Palumbo AV, Daniels WL (2003a) Enhancement of soil carbonsequestration by amendment with fly ash. In: Proceedings 2003 International, Ash UtilizationSymposium, Lexington, Kentucky, Paper #47
Amonette JE, Kim J, Russell CK, Palumbo AV, Daniels WL (2003b) Fly ash catalyzes carbon sequestration.In: Proceedings Second Annual Conference on Carbon Sequestration, Alexandria, Virginia, http://www.carbonsq.com/pdf/posters/TI3.pdf
Climatic Change (2007) 80:5–23 19

Angers DA, Recous S, Aita C (1997) Fate of carbon and nitrogen in water-stable aggregates duringdecomposition of 13C15N-labeled wheat straw in situ. Eur J Soil Sci 48:295–300
Bailey VL, Smith JL, Bolton H Jr (2002) Fungal-to-bacterial ratios in soils investigated for enhanced carbonsequestration. Soil Biol Biochem 34:1385–1389
Baldock JA, Skjemstad JO (2000) Role of the soil matrix and minerals in protecting natural organic materialsagainst biological attack. Org Geochem 31:697–710
Beare MH, Coleman DC, Crossley DA Jr, Hendrix PF, Odum EP (1995) A hierarchical approach toevaluating the significance of soil biodiversity to biogeochemical cycling. Plant Soil 170:5–22
Beare MH, Parmelee RW, Hendrix PF, Cheng W, Coleman DC, Crossley DA Jr (1992) Microbial andfaunal interactions and effects on litter nitrogen and decomposition in agroecosystems. Ecol Monogr62:569–591
Besnard E, Chenu C, Balesdent J, Puget P, Arrouays D (1996) Fate of particulate organic matter in soilaggregates during cultivation. Eur J Soil Sci 47:495–503
Blanco-Canqui H, Lal R (2004) Mechanisms of carbon sequestration in soil aggregates. Crit Rev Plant Sci23:481–504
Booth KJ, Patti AF, Scott JL, Wallis PJ (2004) Organic matter transformation catalyzed by clays: modelreactions for carbon sequestration in soils. In: SuperSoil 2004, 3rd Australian New Zealand SoilsConference, University of Sydney, Australia. http://www.regional.org.au/au/asssi/supersoil2004/s10/oral/1538_wallisp.htm
Bossuyt H, Six J, Hendrix PF (2002) Aggregate-protected carbon in no-tillage and conventional tillageagroecosystems using carbon-14 labeled plant residue. Soil Sci Soc Am J 66:1965–1973
Boyd SA, Mortland MM (1990) Enzyme interactions with clays and clay-organic matter complexes. In:Bollag J-M, Stotzky G (eds) Soil biochemistry, vol. 6. Marcel Dekker, New York, pp 1–28
Carter MR (2004) Researching structural complexity in agricultural soils. Soil Tillage Res 79:1–6Chefetz B, Tarchitzky J, Desmukh AP, Hatcher PG, Chen Y (2002) Structural characterization of soil organic
matter and humic acids in particle-size fractions of an agricultural soil. Soil Sci Soc Am J 66:129–141Christensen BT (2001) Physical fractionation of soil and structural and functional complexity in organic
matter turnover. Eur J Soil Sci 52:345–353Claus H, Gleixner G, Filip Z (1999) Formation of humic-like substances in mixed and pure cultures of
aquatic microorganisms. Acta Hydrochim Hydrobiol 27:200–207Clein JS, Schimel JP (1994) Reduction in microbial activity in birch litter due to drying and rewetting events.
Soil Biol Biochem 26:403–406Conesa A, Punt PJ, van den Hondel C (2002) Fungal peroxidases: molecular aspects and applications.
J Biotechnol 93:143–158Dai X, Boutton TW, Glaser B, Ansley RJ, Zech W (2005) Black carbon in a temperate mixed-grass savanna.
Soil Biol Biochem 37:1879–1881Dalal RC, Bridge BJ (1996) Aggregation and organic matter storage in sub-humid and semi-arid soils. In:
Carter MR, Stewart BA (eds) Structure and organic matter storage in agricultural soils. CRC, BocaRaton, Florida, pp 263–307
Denef K, Six J, Bossuyt H, Frey SD, Elliott ET, Merckx R, Paustian K (2001) Influence of dry–wet cycles onthe interrelationship between aggregate, particulate organic matter, and microbial community dynamics.Soil Biol Biochem 33:1599–1611
Dexter AR (1988) Advances in characterization of soil structure. Soil Tillage Res 11:199–238Dick WA, Tabatabai MA (1992) Significance and potential uses of soil enzymes. In: Metting FB Jr (ed) Soil
microbial ecology: applications in agricultural and environmental management. Marcel Dekker, NewYork, pp 95–127
Ekschmitt K, Liu M, Vetter S, Fox O, Wolters V (2005) Strategies used by soil biota to overcome soilorganic matter stability – why is dead organic matter left over in soil? Geoderma 128:167–176
Elliott ET, Coleman DC (1988) Let the soil work for us. Ecol Bull 39:23–32Elliott ET, Anderson RV, Coleman DC, Cole CV (1980) Habitable pore space and microbial trophic
interactions. Oikos 35:327–335Feller C, Beare MH (1997) Physical control of soil organic matter dynamics in the tropics. Geoderma 79:69–116Fierer N, Schimel JP (2002) Effects of drying–rewetting frequency on soil carbon and nitrogen
transformations. Soil Biol Biochem 34:777–787Flaig W (1975) An introductory review on humic substances: aspects of research on their genesis, their
physical and chemical properties, and their effect on organisms. In: Povoledo D, Golterman HL (eds)Humic substances: their structure and function in the biosphere. Centre for Agricultural Publishing andDocumentation, Wageningen, The Netherlands, pp 19–42
Franzluebbers AJ, Wright SF, Stuedemann JA (2000) Soil aggregation and glomalin under pastures in theSouthern Piedmont USA. Soil Sci Soc Am J 64:1018–1026
20 Climatic Change (2007) 80:5–23

Freeman C, Ostle N, Kang H (2001) An enzymic “latch” on a global carbon store – a shortage of oxygenlocks up carbon in peatlands by restraining a single enzyme. Nature 409:149
Frey SD, Gupta VVSR, Elliott ET, Paustian K (2001) Protozoan grazing affects estimates of carbonutilization efficiency of the soil microbial community. Soil Biol Biochem 33:1759–1768
Gale WJ, Cambardella CA, Bailey TB (2000) Root-derived carbon and the formation and stabilization ofaggregates. Soil Sci Soc Am J 64:201–207
Gans J, Wolinsky M, Dunbar J (2005) Computational improvements reveal great bacterial diversity and highmetal toxicity in soil. Science 309:1387–1390
George TS, Richardson AE, Simpson RJ (2005) Behaviour of plant-derived extracellular phytase uponaddition to soil. Soil Biol Biochem 37:977–988
Glaser B, Lehmann J, Zech W (2002) Ameliorating physical and chemical properties of highly weatheredsoils in the tropics with charcoal – a review. Biol Fertil Soils 35:219–230
Golchin A, Oades JM, Skjemstad JO, Clark P (1994) Soil structure and carbon cycling. Aust J Soil Res32:1043–1068
Gregorich EG, Kachanoski RG, Voroney RP (1989) Carbon mineralization in soil size fractions after variousamounts of aggregate disruption. J Soil Sci 40:649–659
Guggenberger G, Frey SD, Six J, Paustian K, Elliot ET (1999) Bacterial and fungal cell-wall residues inconventional and no-tillage agroecosystems. Soil Sci Soc Am J 63:1188–1198
Haider K, Martin JP (1981) Decomposition in soil of specifically 14C-labeled model and cornstalk ligninsand coniferyl alcohol over two years as influenced by drying, rewetting, and additions of an available Csubstrate. Soil Biol Biochem 13:447–450
Haider K, Martin JP, Filip Z, Fustec-Mathon E (1975) Contribution of soil microbes to the formation ofhumic compounds. In: Povoledo D, Golterman HL (eds) Humic substances: their structure and functionin the biosphere. Centre for Agricultural Publishing and Documentation, Wageningen, The Netherlands,pp 71–85
Hättenschwiler S, Tiunov AV, Scheu S (2005) Biodiversity and litter decomposition in terrestrial ecosystems.Annu Rev Ecol Evol Syst 36:191–218
Hayes MHB, Malcolm RL (2001) Considerations of compositions and aspects of the structures of humicsubstances. In: Clapp CE, Hayes MHB, Senesi N, Bloom PR, Jardine PM (eds) Humic substances andchemical contaminants. Soil Science Society of America, Madison, Wisconsin, pp 3–39
Haynes RJ, Beare MH (1997) Influence of six crop species on aggregate stability and some labile organicmatter fractions. Soil Biol Biochem 29:1647–1653
Hedges JI (1988) Polymerization of humic substances in natural environments. In: Frimmel FH, ChristmanRF (eds) Humic substances and their role in the environment. Wiley, New York, pp 45–57
Hill GT, Mitkowski NA, Aldrich-Wolfe L, Emele LR, Jurkonie DD, Ficke A, Maldonado-Ramirez S, LynchST, Nelson EB (2000) Methods for assessing the composition and diversity of soil microbialcommunities. Appl Soil Ecol 15:25–36
Holland EA, Coleman DC (1987) Litter placement effects on microbial and organic matter dynamics in anagroecosystem. Ecology 68:425–433
Hu SJ, van Bruggen AHC, Grünwald NJ (1999) Dynamics of bacterial populations in relation to carbonavailability in a residue-amended soil. Appl Soil Ecol 13:21–30
Jastrow JD, Miller RM (1998) Soil aggregate stabilization and carbon sequestration: feedbacks throughorganomineral associations. In: Lal R, Kimble JM, Follett RF, Stewart BA (eds) Soil processes and thecarbon cycle. CRC, Boca Raton, Florida, pp 207–223
Jastrow JD, Miller RM, Lussenhop J (1998) Contributions of interacting biological mechanisms to soilaggregate stabilization in restored prairie. Soil Biol Biochem 30:905–916
Jenny H (1941) Factors of soil formation. McGraw-Hill, New YorkJohnson DW, Curtis PS (2001) Effects of forest management on soil C and N storage: meta analysis. For
Ecol Manag 140:227–238Kaiser K, Guggenberger G (2003) Mineral surfaces and soil organic matter. Eur J Soil Sci 54:219–236Kay BD (1990) Rates of change of soil structure under different cropping systems. Adv Soil Sci 12:1–52Kononova MM (1961) Soil organic matter. Nowakowski TZ, Greenwood GA (transl.) Pergamon, OxfordLuo Y, White LW, Canadell JG, DeLucia EH, Ellsworth DS, Finzi A, Lichter J, Schlesinger WH (2003)
Sustainability of terrestrial carbon sequestration: a case study in Duke Forest with inversion approach.Glob Biogeochem Cycles 17(1):1021
Maillard LC (1916) Synthese de matieres humiques par action des acides amines sur les sucres reducteures.Ann Chem Phys 5:528–317
Marilley L, Hartwig UA, Aragno M (1999) Influence of an elevated atmospheric CO2 content on soil andrhizosphere bacterial communities beneath Lolium perenne and Trifolium repens under field conditions.Microb Ecol 38:39–49
Climatic Change (2007) 80:5–23 21

Martin JP, Haider K (1971) Microbial activity in relation to soil humus formation. Soil Sci 111:54–63Martin JP, Haider K, Bondietti E (1975) Properties of model humic acids synthesized by phenoloxidase and
autoxidation of phenols and other compounds formed by soil fungi. In: Povoledo D, Golterman HL (eds)Humic substances: their structure and function in the biosphere. Centre for Agricultural Publishing andDocumentation, Wageningen, The Netherlands, pp 171–186
Martin A, Marinissen JCY (1993) Biological and physico–chemical processes in excrements of soil animals.Geoderma 56:331–347
Mikola J, Bardgett RD, Hedlund K (2002) Biodiversity, ecosystem functioning and soil decomposer foodwebs. In: Loreau M, Naeem S, Inchausti P (eds) Biodiversity and ecosystem functioning–synthesis andperspectives. Oxford University Press, Oxford, pp 169–180
Miller RM, Jastrow JD (1990) Hierarchy of root and mycorrhizal fungal interactions with soil aggregation.Soil Biol Biochem 22:579–584
Miller RM, Jastrow JD (2000) Mycorrhizal fungi influence soil structure. In: Kapulnik Y, Douds DD Jr (eds)Arbuscular mycorrhizas: physiology and function. Kluwer, Dordrecht, The Netherlands, pp 3–18
Naidja A, Huang PM, Bollag J-M (1998) Comparison of reaction products from the transformation ofcatechol catalyzed by birnessite or tyrosinase. Soil Sci Soc Am J 62:188–195
Nannipieri P, Ascher J, Ceccherini MT, Landi L, Pietramellara G, Renella G (2003) Microbial diversity andsoil functions. Eur J Soil Sci 54:655–670
Oades JM (1984) Soil organic matter and structural stability: mechanisms and implications for management.Plant Soil 76:319–337
Oades JM (1993) The role of biology in the formation, stabilization and degradation of soil structure.Geoderma 56:377–400
Oades JM, Waters AG (1991) Aggregate hierarchy in soils. Aust J Soil Res 29:815–828Payne WJ (1970) Energy yields and growth of heterotrophs. Annu Rev Microbiol 24:17–52Pelz O, Cifuentes LA, Hammer BT, Kelley CA, Coffin RB (1998) Tracing the assimilation of organic
compounds using δ13C analysis of unique amino acids in the bacterial peptidoglycan cell wall. FEMSMicrobiol Ecol 25:229–240
Plante AF, McGill WB (2002) Soil aggregate dynamics and the retention of organic matter in laboratory-incubated soil with differing simulated tillage frequencies. Soil Tillage Res 66:79–92
Rees RM, Bingham IJ, Baddeley JA, Watson CA (2005) The role of plants and land management insequestering soil carbon in temperate arable and grassland ecosystems. Geoderma 128:130–154
Rillig MC (2004) Arbuscular mycorrhizae, glomalin, and soil aggregation. Can J Soil Sci 84:355–363Rillig MC, Allen MF (1999) What is the role of arbuscular mycorrhizal fungi in plant-to-ecosystem
responses to elevated atmospheric CO2? Mycorrhiza 9:1–8Schimel JP, Bennett J (2004) Nitrogen mineralization: challenges of a changing paradigm. Ecology 85:591–602Schimel JP, Weintraub MN (2003) The implications of exoenzyme activity on microbial carbon and nitrogen
limitation in soil: a theoretical model. Soil Biol Biochem 35:549–563Schimel JP, Gulledge JM, Clein-Curley JS, Lindstrom JE, Braddock JF (1999) Moisture effects on microbial
activity and community structure in decomposing birch litter in the Alaskan taiga. Soil Biol Biochem31:831–838
Schubert WJ (1965) Lignin biochemistry. Academic, New YorkSeastedt TR (1984) The role of arthropods in decomposition and mineralization processes. Annu Rev
Entomol 29:25–46Sexstone AJ, Revsbech NP, Parkin TB, Tiedje JM (1985) Direct measurement of oxygen profiles and
denitrification rates in soil aggregates. Soil Sci Soc Am J 49:645–651Shen S, Tu S, Taylor RM (2002) Interactions of enzymes with clays and applications in bioremediation. In:
Dixon JB, Schulze DG (eds) Soil mineralogy with environmental applications, SSSA Book Series 7. SoilScience Society of America, Madison, Wisconsin, pp 795–817
Shindo H, Huang PM (1984) Catalytic effects of manganese(IV), iron(III), aluminum, and silicon-oxides onthe formation of phenolic polymers. Soil Sci Soc Am J 48:927–934
Six J, Jastrow JD (2002) Organic matter turnover. In: Lal R (ed) Encyclopedia of soil science. MarcelDekker, New York, pp 936–942
Six J, Bossuyt H, Degryze S, Denef K (2004) A history of research on the link between (micro)aggregates,soil biota, and soil organic matter dynamics. Soil Tillage Res 79:7–31
Six J, Conant RT, Paul EA, Paustian K (2002a) Stabilization mechanisms of soil organic matter: implicationsfor C-saturation of soils. Plant Soil 241:155–176
Six J, Elliott ET, Paustian K (2000) Soil macroaggregate turnover and microaggregate formation: amechanism for C sequestration under no-tillage agriculture. Soil Biol Biochem 32:2099–2103
Six J, Feller C, Denef K, Ogle SM, de Moraes Sa JC, Albrecht A (2002b) Soil organic matter, biota andaggregation in temperate and tropical soils – effects of no-tillage. Agronomie 22:755–775
22 Climatic Change (2007) 80:5–23

Sjoblad RD, Bollag J-M (1981) Oxidative coupling of aromatic compounds by enzymes from soilmicroorganisms. In: Paul EA, Ladd JN (eds) Soil biochemistry, vol 5. Marcel Dekker, New York,pp 113–152
Sollins P, Homann P, Caldwell BA (1996) Stabilization and destabilization of soil organic matter:mechanisms and controls. Geoderma 74:65–105
Steinberg PD, Rillig MC (2003) Differential decomposition of arbuscular mycorrhizal fungal hyphae andglomalin. Soil Biol Biochem 35:191–194
Stevenson FJ (1994) Humus chemistry: genesis, composition, reactions, 2nd edn. Wiley, New YorkStevenson FJ, Cole MA (1999) Cycles of soil: carbon, nitrogen, phosphorus, sulfur, micronutrients. Wiley,
New YorkStrong DT, DeWever H, Merckx R, Recous S (2004) Spatial location of carbon decomposition in the soil
pore system. Eur J Soil Sci 55:739–750Suberkropp K, Weyers H (1996) Application of fungal and bacterial production methodologies to
decomposing leaves in streams. Appl Environ Microbiol 62:1610–1615Sutton R, Sposito G (2005) Molecular structure in soil humic substances: the new view. Environ Sci Technol
39:9009–9015Tan KH (2003) Humic matter in soil and the environment: principles and controversies. Marcel Dekker,
New YorkTate RL III (1992) Soil organic matter: biological and ecological effects. Krieger, Malabar, Floridaten Have R, Teunissen PJM (2001) Oxidative mechanisms involved in lignin degradation by white-rot fungi.
Chem Rev 101:3397–3413Tiedje JM, Asuming-Brempong S, Nüsslein K, Marsh TL, Flynn SJ (1999) Opening the black box of soil
microbial diversity. Appl Soil Ecol 13:109–122Tisdall JM (1996) Formation of soil aggregates and accumulation of soil organic matter. In: Carter MR,
Stewart BA (eds) Structure and organic matter storage in agricultural soils. CRC, Boca Raton, Florida,pp 57–96
Tisdall JM, Oades JM (1982) Organic matter and water stable aggregates in soils. J Soil Sci 33:141–163Treseder KK, Allen MF (2000) Mycorrhizal fungi have a potential role in soil carbon storage under elevated
CO2 and nitrogen deposition. New Phytol 147:189–200van der Wal A, van Veen JA, Smant W, Boschker HTS, Bloem J, Kardol P, van der Putten WH, de Boer W
(2006) Fungal biomass development in a chronosequence of land abandonment. Soil Biol Biochem38:51–60
Vetter YA, Deming JW, Jumars PA, Krieger-Brockett BB (1998) A predictive model of bacterial foraging bymeans of freely released extracellular enzymes. Microb Ecol 36:75–92
Waksman SA (1932) Humus. Williams & Wilkins, Baltimore, MarylandWang MC, Huang PM (2005) Cleavage of 14C-labeled glycine and its polycondensation with pyrogallol as
catalyzed by birnessite. Geoderma 124:415–426West AW, Sparling GP, Grant WD (1987) Relationships between mycelial and bacterial populations in
stored, air-dried and glucose-amended arable and grassland soils. Soil Biol Biochem 19:599–605Wolfaardt GM, Lawrence JR, Robarts RD, Caldwell DE (1994) The role of interactions, sessile growth, and
nutrient amendments on the degradative efficiency of a microbial consortium. Can J Microbiol 40:331–340Young IM, Ritz K (2000) Tillage, habitat space and function of soil microbes. Soil Tillage Res 53:201–213Zech W, Sensei N, Guggenberger G, Kaiser K, Lehmann J, Miano TM, Miltner A, Schroth G (1997) Factors
controlling humification and mineralization of soil organic matter in the tropics. Geoderma 79:117–161Zhu YG, Miller RM (2003) Carbon cycling by arbuscular mycorrhizal fungi in soil–plant systems. Trends
Plant Sci 8:407–409Zimmerman AR, Chorover J, Goyne KW, Brantley SL (2004) Protection of mesopore-adsorbed organic
matter from enzymatic degradation. Environ Sci Technol 38:4542–4548
Climatic Change (2007) 80:5–23 23