kiss1 receptor is preferentially expressed in clinically non-functioning pituitary tumors
TRANSCRIPT

KISS1 receptor is preferentially expressed in clinicallynon-functioning pituitary tumors
Marianna Yaron • Ulrich Renner • Suzan Gilad •
Gunter K. Stalla • Naftali Stern • Yona Greenman
� Springer Science+Business Media New York 2014
Abstract
Objective KISS1 is a metastasis suppressor gene involved
in cancer biology. Given the high expression levels of KISS1
and KISS1R in the hypothalamus and the pituitary respec-
tively, we hypothesized that this system could possibly affect
tumor invasiveness and clinical behavior of pituitary tumors.
Methods Expression levels of KISS1 and KISS1R mRNA
were evaluated by RT-PCR. Clinical information pertaining
tumor characteristics was extracted from patients’ charts.
Results Tumors from 39 patients (21 females, mean age
47.5 years) were examined. KISS1R was expressed in 26
(67 %) of samples (94 % of NFPA, 42 % of GH-, 67 % of
ACTH-, and 25 % of PRL-secreting adenomas) and was
found more often in female patients (81 vs. 50 % males,
p \ 0.05); and in NFPA (94 vs. 45.5 % in secreting
tumors; p = 0.003). Patients expressing KISS1R were
older at presentation (50.5 ± 1.4 vs. 38.1 ± 1.3 years;
p = 0.008). In the multivariate analysis, factors signifi-
cantly associated with KISS1R expression included female
gender (OR 13.8, 95 % CI 1.22–155.9; p = 0.03) and
having a NFPA (OR 24.7, 95 % CI 1.50–406.4; p = 0.02).
Tumor size, invasiveness and age at presentation were not
independently associated with KISS1R expression.
Pituitary tumors and normal pituitary were negative for
KISS1 mRNA expression.
Conclusions The majority of human NFPA expressed
KISS1R with lower rates of expression in other types of
pituitary tumors. KISS1R expression did not impart a
clinical beneficial tumor phenotype, as it was not associ-
ated with tumor size or invasiveness. Additional studies are
required to elucidate the role of KISS1 receptor in pituitary
gland physiology and pathology.
Keywords KISS1 � GPR54 � KISS1R � Pituitary adenoma
Abbreviations
NFPA Non functional pituitary adenoma
KISS1R KISS1 receptor
GH Growth hormone
ACTH Adrenocorticotropic hormone
PRL Prolactin
Introduction
The KISS1 gene, localized to chromosome 1, was originally
isolated more than a decade ago from melanoma and breast
carcinoma cells that have lost their ability to metastasize
[1, 2]. Later, a family of peptides encoded by the KISS1 gene
and named kisspeptins were isolated from human placenta
and found to be the endogenous ligands for the than orphan G
protein-coupled receptor GPR54 [3–5], now named KISS1
receptor (KISS1R). KISS1R is highly expressed in placenta,
pituitary and in the brain, mostly within forebrain nuclei,
whereas KISS1 is predominantly expressed in the placenta,
hypothalamus and striatum with lower levels of expression
in testis, pancreas and liver [3–6].
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11102-014-0572-y) contains supplementarymaterial, which is available to authorized users.
M. Yaron � S. Gilad � N. Stern � Y. Greenman (&)
Institute of Endocrinology, Metabolism and Hypertension, Tel
Aviv-Sourasky Medical Center and Sackler Faculty of Medicine,
Tel Aviv University, 6 Weizmann Street, 64239 Tel Aviv, Israel
e-mail: [email protected]
U. Renner � G. K. Stalla
Neuroendocrinology Group, Max-Plank-Institute of Psychiatry,
Munich, Germany
123
Pituitary
DOI 10.1007/s11102-014-0572-y

There are strong data indicating that KISS1 is a metas-
tasis suppressor gene in melanoma [7], gastric [8], esoph-
ageal [9], bladder [10], papillary thyroid [11], and other
cancers [12–15], in that loss of KISS1 mRNA expression
was correlated with tumor progression, presence of
metastasis and a poor outcome. Nevertheless, these find-
ings are not universal as high KISS1 expression in hepa-
tocellular carcinoma is associated with a worse prognosis,
and the correlation of KISS1 expression with breast cancer
progression remains controversial [16].
In addition to its involvement in cancer biology, the
KISS1/KISS1R system plays a central role in the neuro-
endocrine regulation of reproduction. Loss of function
mutations of KISS1 were found to be associated with lack
of puberty onset and hypogonadotrophic hypogonadism in
humans and rodents, whereas activating mutations led to
precocious puberty [17–21]. There is compelling evidence
pointing to the hypothalamus, and specifically, the GnRH
neurons as the primary site of action of kisspeptins [22].
However, in addition to the GnRH mediated increase in LH
levels, there is mounting evidence that kisspeptins may
directly stimulate gonadotropin secretion at the pituitary
level [22]. KISS1 and KISS1R mRNA and protein have
been shown to be expressed in the rat pituitary [23, 24].
Furthermore, kisspeptin has been shown to elicit LH
secretion, albeit in a lower magnitude than the response to
GnRH, from pituitary cells in vitro from several species
[25], including non-human primates [26].
Given the high expression levels of KISS1 and KISS1R in
the hypothalamus and pituitary respectively, the involve-
ment of this system in gonadotropin secretion, and its effects
on cell growth, migration, motility and adhesiveness in other
organs, we hypothesize that KISS1 or its receptor may be
involved in pituitary tumorigenesis, and could possibly
affect the degree of tumor invasiveness and clinical behavior
of pituitary tumors. Although KISS1 and KISS1R expression
in pituitary tumors has been previously reported [27], its
relationship with tumor invasiveness has not been explored.
Therefore, in the present study, we set out to analyze the
pattern of mRNA expression of KISS1 and KISS1R in
pituitary adenomas, giving special emphasis to possible
associations with tumor clinical characteristics such as size,
invasiveness and hormonal secretion.
Materials and methods
Human normal pituitary gland and pituitary adenoma
tissue
A commercial panel of total RNA from various normal
pituitary tissues was obtained from CLONTECH (Human
Total Master Panel II and human pituitary gland poly-A
RNA; Palo Alto, Ca) pooled from 88 male/female Cauca-
sians. Pituitary tumor specimens were obtained during
transsphenoidal surgery at the Tel Aviv-Sourasky Medical
Center (n = 25). Following surgical excision, all tumors
were immediately snap-frozen in liquid nitrogen and stored
at -80 �C until analysis. Additional cDNA samples from
16 pituitary tumors were received from the Max-Plank
Institute of Psychiatry, Munich, Germany. This study was
approved by the institutional ethics committee of both
medical centers.
Tumor size and invasiveness were defined on the basis
of preoperative radiological investigations and operative
finding as follows: Grade I (microadenomas, \10 mm in
diameter), Grade II (enclosed macroadenomas with or
without suprasellar extension), Grade III (microadenomas
with local invasion of sphenoid and/or cavernous sinus)
and Grade IV tumors (macroadenomas with diffuse inva-
sion into the sella floor/cavernous sinus) [28, 29]. Tumors
of patients diagnosed with PRL-, ACTH- and GH- secret-
ing adenomas had positive staining for the respective over-
secreted hormones, as detected by routine immunohisto-
chemistry staining (IHC). Of the 17 clinically silent NPFA,
two stained positive for b-FSH, five were positive for both
b-LH and b-FSH, and five had negative staining for all
pituitary hormones. IHC data from five NPFA was missing.
Human placental tissue
Human placental tissues were obtained up to 30 min after
spontaneous placental delivery. The collected specimens
were transferred to Petri dishes and washed thoroughly with
physiological saline to remove any contamination with
maternal blood and amniotic fluid. Each tissue sample was
transferred to tubes containing 1.5 ml RNAlater solution
(RNA stabilization reagent, Qiagen GmbH, Germany) and
stored at -70 �C until tissue processing.
RNA isolation and reverse transcription (RT)
Total RNA was extracted from 30 to 200 mg frozen pitu-
itary tumor tissue homogenized in a Polytron 3000
homogenizer (Kinematica AG, Littau, Switzeland) using
Trizol reagent (Life Technologies, Inc., Gaithersburg,
MD), according to the manufacturer’s instructions. RNA
concentration was determined by absortiometry. One lg of
RNA was reverse transcribed using Advantage TM RT-for-
PCR kit (Clontech) and the final pellet was resuspended in
100 ll of DNA-free water. Five ll of this cDNA suspen-
sion together with specific primers in a final concentration
of 0.5 micromolar each, dNTPs 0.2 mM, MgCl2 1.5-
2.5 mM and AmpliTaq Gold polymerase (Perkin Elmer)
1 U were used in a hot start PCR reaction in a final volume
of 50 ll. PCR reactions were performed using the ABI
Pituitary
123
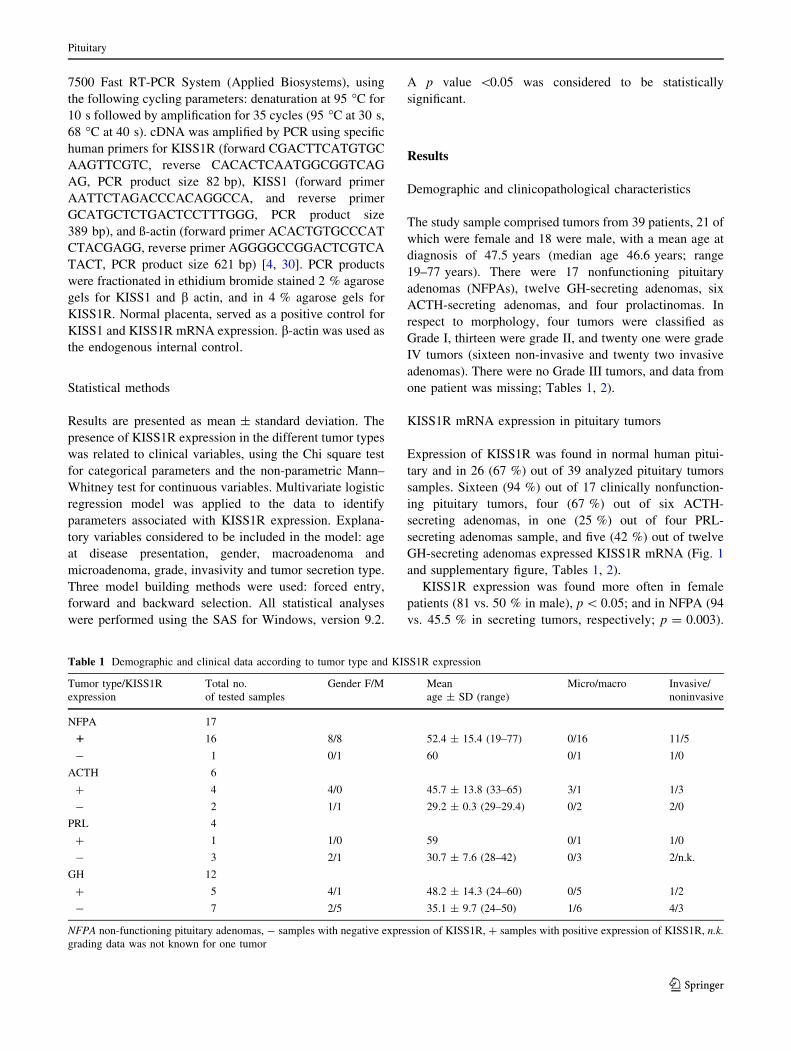
7500 Fast RT-PCR System (Applied Biosystems), using
the following cycling parameters: denaturation at 95 �C for
10 s followed by amplification for 35 cycles (95 �C at 30 s,
68 �C at 40 s). cDNA was amplified by PCR using specific
human primers for KISS1R (forward CGACTTCATGTGC
AAGTTCGTC, reverse CACACTCAATGGCGGTCAG
AG, PCR product size 82 bp), KISS1 (forward primer
AATTCTAGACCCACAGGCCA, and reverse primer
GCATGCTCTGACTCCTTTGGG, PCR product size
389 bp), and ß-actin (forward primer ACACTGTGCCCAT
CTACGAGG, reverse primer AGGGGCCGGACTCGTCA
TACT, PCR product size 621 bp) [4, 30]. PCR products
were fractionated in ethidium bromide stained 2 % agarose
gels for KISS1 and b actin, and in 4 % agarose gels for
KISS1R. Normal placenta, served as a positive control for
KISS1 and KISS1R mRNA expression. b-actin was used as
the endogenous internal control.
Statistical methods
Results are presented as mean ± standard deviation. The
presence of KISS1R expression in the different tumor types
was related to clinical variables, using the Chi square test
for categorical parameters and the non-parametric Mann–
Whitney test for continuous variables. Multivariate logistic
regression model was applied to the data to identify
parameters associated with KISS1R expression. Explana-
tory variables considered to be included in the model: age
at disease presentation, gender, macroadenoma and
microadenoma, grade, invasivity and tumor secretion type.
Three model building methods were used: forced entry,
forward and backward selection. All statistical analyses
were performed using the SAS for Windows, version 9.2.
A p value \0.05 was considered to be statistically
significant.
Results
Demographic and clinicopathological characteristics
The study sample comprised tumors from 39 patients, 21 of
which were female and 18 were male, with a mean age at
diagnosis of 47.5 years (median age 46.6 years; range
19–77 years). There were 17 nonfunctioning pituitary
adenomas (NFPAs), twelve GH-secreting adenomas, six
ACTH-secreting adenomas, and four prolactinomas. In
respect to morphology, four tumors were classified as
Grade I, thirteen were grade II, and twenty one were grade
IV tumors (sixteen non-invasive and twenty two invasive
adenomas). There were no Grade III tumors, and data from
one patient was missing; Tables 1, 2).
KISS1R mRNA expression in pituitary tumors
Expression of KISS1R was found in normal human pitui-
tary and in 26 (67 %) out of 39 analyzed pituitary tumors
samples. Sixteen (94 %) out of 17 clinically nonfunction-
ing pituitary tumors, four (67 %) out of six ACTH-
secreting adenomas, in one (25 %) out of four PRL-
secreting adenomas sample, and five (42 %) out of twelve
GH-secreting adenomas expressed KISS1R mRNA (Fig. 1
and supplementary figure, Tables 1, 2).
KISS1R expression was found more often in female
patients (81 vs. 50 % in male), p \ 0.05; and in NFPA (94
vs. 45.5 % in secreting tumors, respectively; p = 0.003).
Table 1 Demographic and clinical data according to tumor type and KISS1R expression
Tumor type/KISS1R
expression
Total no.
of tested samples
Gender F/M Mean
age ± SD (range)
Micro/macro Invasive/
noninvasive
NFPA 17
1 16 8/8 52.4 ± 15.4 (19–77) 0/16 11/5
- 1 0/1 60 0/1 1/0
ACTH 6
? 4 4/0 45.7 ± 13.8 (33–65) 3/1 1/3
- 2 1/1 29.2 ± 0.3 (29–29.4) 0/2 2/0
PRL 4
? 1 1/0 59 0/1 1/0
- 3 2/1 30.7 ± 7.6 (28–42) 0/3 2/n.k.
GH 12
? 5 4/1 48.2 ± 14.3 (24–60) 0/5 1/2
- 7 2/5 35.1 ± 9.7 (24–50) 1/6 4/3
NFPA non-functioning pituitary adenomas, - samples with negative expression of KISS1R, ? samples with positive expression of KISS1R, n.k.
grading data was not known for one tumor
Pituitary
123

Patients harboring tumors that expressed KISS1R mRNA
were older that those with negative expression (50.5 ± 1.4
vs. 38.1 ± 1.3 years at presentation, respectively;
p = 0.008; Table 1). There were no significant differences
in KISS1R expression according to invasiveness or tumor
grade (Table 2).
In the multivariate logistic regression model, factors
significantly associated with KISS1R expression included
female gender (OR 13.8, 95 % CI 1.22–155.9; p = 0.03)
and having a non-functioning pituitary adenoma (OR 24.7,
95 % CI 1.50–406.4; p = 0.02). Tumor size (microade-
noma or macroadenoma), invasivity and age at presentation
were not independently associated with KISS1R expres-
sion. The results of the multivariate analysis are summa-
rized in Table 3.
KISS1 mRNA was clearly expressed in normal human
placental tissue but was absent in normal pituitary tissue
and in 15 pituitary tumor samples analyzed (10 NFPA, two
GH secreting adenomas, two ACTH secreting tumors and
one prolactinomas), not shown.
Discussion
KISS1 neurons are central regulators of the hypothalamo-
pituitary gonadal axis through direct stimulation of
KISS1R expressing GnRH neurons [22]. The finding that
KISS1 and its receptors are expressed in rat pituitary cells,
and that kisspeptins may directly stimulate pituitary gon-
adotropin secretion in vitro, raised the hypothesis that there
may be another tier of regulation in addition to the well
established hypothalamic level [23, 24]. Kisspeptins have
also been shown to regulate cell growth and adhesion, and
loss of KISS1 expression in several human malignancies
has been linked to the development of a metastatic phe-
notype, and consequently a poor clinical outcome [7, 16].
Hence, the possibility that kisspeptins may also be involved
in the regulation of pituitary tumor development is attrac-
tive, as most pituitary tumors, even when large and locally
invasive do not metastasize.
Table 2 KISS1R expression in relation to clinicopathological tumor
characteristics
Variable No. of tested
samples
KISS1R expression (%)
N = 39
p
Expression ?
Number (%)
-
Number (%)
Adenoma type
NFPA 17 16 (94) 1 (6) 0.003*
ACTH 6 4 (67) 2 (33)
PRL 4 1 (25) 3 (75)
GH 12 5 (42) 7 (58)
Tumor size
Macro 35 23 (66) 12 (34) NS
Micro 4 3 (75) 1 (25) NS
Invasion
Invasive 22 14 (64) 8 (36) NS
Non-
invasive
16 12 (75) 4 (25) NS
Grade
I 4 3 (75) 1 (25) NS
II 13 10 (77) 3 (23) NS
III 0 0 0 NA
IV 21 13 (62) 8 (38) NS
Gender
Male 18 9 (50) 9 (50)
Female 21 17 (81) 4 (19) 0.008
NFPA non-functioning pituitary adenomas, - samples with negative
expression of KISS1R, ? samples with positive expression of
KISS1R
* p \ 0.003 NFPA versus other secreting tumors (ACTH-, PRL-,
GH-secreting tumors)
Fig. 1 PCR analysis of KISS1R mRNA expression in pituitary
adenomas. Lane 1 in all gels: ladder. a Lane 3 NFPA, lanes 6–7
prolactinoma, lane 8 ACTH-secreting adenoma, lane 9 GH-secreting
adenoma, lane 10 prolactinoma, lane 11 normal pituitary gland, lane
12 negative control; b lanes 2–6 NFPA, lane 8 NFPA, lanes 9–11
GH-secreting adenoma, lane 12 - negative control; c lanes 2–3
NFPA, lane 4 GH-secreting adenoma, lanes 5–6 NFPA, lane 7
prolactinoma, lane 8 NFPA, lane 10 NFPA, lane 11 placenta, lane 12
negative control; d lanes 2 and 4 NFPA, lanes 5–7 ACTH-secreting
adenoma, lane 8 NFPA, lanes 9–11 GH-secreting adenoma, lane 12
negative control; e lane 2 NFPA, lanes 3–6 GH-secreting adenoma,
lanes 7–8 ACTH-secreting adenoma, lane 9 negative control
Pituitary
123

In this study we found that KISS1R is expressed in
normal human pituitary, as previously reported by Muir
et al. [5]. Furthermore, the majority of NFPA also
expressed KISS1R, consistent with the gonadotroph cell
origin of these tumors, and with previous reports of
KISS1R co-localization with LH-b cells in rat pituitary
[23]. It should be noted, though, that in the latter study,
only 10 % of LH-b cells co-expressed KISS1R. Thus, it
could be postulated that NFPA gonadotrophs retained
KISS1R expression even after becoming adenomatous.
Nevertheless, the percentage of human gonadotrophs that
express KISS1R is unknown, and any pathophysiological
inference from another species could be inaccurate. Inter-
estingly, KISS1R expression was not restricted to NFPA,
but was also found, albeit at a lower rate, in GH, ACTH
and prolactin secreting tumors. Gutierrez-Pascual et al.
[24] have demonstrated that kisspeptin-10 induced a rise in
free cytosolic Ca2? concentration in rat somatotrophs.
Furthermore, kisspeptin-10 has been reported to stimulate
the secretion of both growth hormone and prolactin from
cultured bovine anterior pituitary cells [31]. Thus, although
this data indirectly implies functional KISS1R expression
in somatotrophs and lactotrophs from other species, it is
imperative that co-localization studies be performed in
normal human pituitary cells. Information thus obtained
could point to a possible mechanism by which KISS1R
could be involved in pituitary tumorigenesis, namely
through loss or de novo acquisition of its expression. The
lack of correlation between KISS1R expression and tumor
size or degree of invasiveness could infer that this gene is
not important for tumor progression. Nevertheless, our
analysis was not quantitative and we cannot rule out a more
subtle type of influence of KISS1R expression on tumor
behavior. That KISS1R expression was more prevalent in
female patients’ tumors suggests that sex steroids could
modulate its tumoral expression. Consistent with this line,
estrogen has been shown to modulate KISS1 and KISS1R
mRNA in rat pituitary [23].
In contrast to the information accumulated from rat
pituitary studies, we were unable to detected KISS1 mRNA
expression neither in the normal human pituitary nor in
pituitary tumor samples. Muir et al. [5], in their seminal
paper reporting the cloning of human KISS1R and the
identification of KISS1 as its ligand, also did not identify
KISS1 expression in the human pituitary. On the other hand,
Martınez-Fuentes et al. [27] reported KISS1 mRNA
expression both in normal human pituitary and roughly in
one third of pituitary tumors, irrespective of their secretory
profile. The discordance between our results may stem from
technical differences in tumor processing and primer sensi-
tivity. Similarly to our findings, Martınez-Fuentes et al. [27]
found KISS1R expression in all pituitary tumor types (with
the exception of two rare TSH-secreting tumors) irrespective
of size, albeit without the preponderant expression in NFPA
detected by us. Important limitations of our study are the
relatively small number of tumors examined, and the fact
that expression analysis was not quantitative.
In conclusion, we have demonstrated that the vast
majority of human NFPA express KISS1R, with lower
rates of expression in other types of pituitary tumors; and
that this expression was more often found in tumors
excised from female subjects and at a higher age of clinical
presentation. We could not prove the hypothesis that
KISS1R expression could impart a clinical beneficial tumor
phenotype, as it was not associated with tumor size or
invasiveness. Undoubtedly, additional studies are required
to elucidate the role of KISS1R in pituitary gland physi-
ology as well as in its pathological processes.
Conflict of interest The authors have no disclosures to declare and
they have no conflict of interest.
Ethical standard We declare that the experiments comply with the
current laws of our country.
References
1. Lee JH, Miele ME, Hicks DJ, Phillips KK, Trent JM, Weissman BE,
Welch DR (1996) KiSS-1, a novel human malignant melanoma
metastasis-suppressor gene. J Natl Cancer Inst 88:1731–1737
2. Lee JH, Welch DR (1997) Suppression of metastasis in human
breast carcinoma MDA-MB-435 cells after transfection with the
metastasis suppressor gene, KiSS-1. Cancer Res 57:2384–2385
3. Kotani M, Detheux M, Vandenbogaerde A, Communi D, Van-
derwinden JM, Le Poul E, Brezillon X, Tyldesley R, Suarez-
Table 3 Multivariate logistic analysis of the association between KISS1R expression and a set of demographic and clinical variables
Covariate Odds ratio (95 % CI) Estimate SE Wald beta p value
Macroadenoma versus microadenoma 0.678 (0.03–15.5) -0.194 0.7985 0.06 0.8
Grade 1.552 (0.01–223.8) 0.219 0.0301 0.03 0.9
Invasivity (invasive vs. non-invasive) 3.138 (0.01–713.5) 1.143 2.768 0.17 0.6
Secretory type (NFPA vs. others) 24.7 (1.50–406.4) 1.603 0.7143 5.04 0.02
Age at the presentation (\50 years vs. older) 1.076 (0.98–1.181) 0.073 0.0475 2.39 0.1
Gender (female vs. male) 13.79 (1.22–155.9) 1.312 0.6187 4.49 0.03
Pituitary
123

Huerta N, Vandeput F, Blanpain C, Schiffmann SN, Vassart G,
Parmentier M (2001) The metastasis suppressor gene KiSS-1
encodes kisspeptins, the natural ligands of the orphan G protein-
coupled receptor GPR54. J Biol Chem 276:34631–34636
4. Ohtaki T, Shintani Y, Honda S, Matsumoto H, Hori A, Kanehashi
K, Terao Y, Kumano S, Takatsu Y, Masuda Y, Ishibashi Y,
Watanabe T, Asada M, Yamada T, Suenaga M, Kitada C, Usuki
S, Kurokawa T, Onda H, Nishimura O, Fujino M (2001)
Metastasis suppressor gene KiSS-1 encodes peptide ligand of a
G-protein coupled receptor. Nature 411:613–617
5. Muir AL, Chamberlain L, Eshourbagy NA, Michalovich D,
Moore DJ, Calamari A, Szekeres PG, Sarau HM, Chambers JK,
Murdock P, Steplewski K, Shabon U, Miller JE, Middleton SE,
Darker JG, Larminie CGC, Wilson S, Bergsma DJ, Emson P,
Faull R, Philpott KL, Harrison DC (2001) AXOR12, a novel
human G protein-coupled receptor, activated by the peptide
KiSS-1. J Biol Chem 276:28969–28975
6. Lee DK, Nguyen T, O’Neill GP, Cheng R, Liu Y, Howard AD,
Coulombe N, Tan CP, Tang-Nguyen AT, George SR, O’Dowd
BF (1999) Discovery of a receptor related to the galanin recep-
tors. FEBS Lett 446:103–107
7. Shirasaki F, Takata M, Hatta N, Takehara K (2001) Loss of
expression of the metastasis suppressor gene KiSS1 during mel-
anoma progression and its association with LOH of chromosome
6q16.3-q23. Cancer Res 61:7422–7425
8. Dhar DK, Naora H, Kubota H, Maruyama R, Yoshimura H,
Tonomoto Y, Tachibana M, Ono T, Otani H, Nagasue N (2004)
Downregulation of KISS-1 expression is responsible for tumor
invasion and worse prognosis in gastric carcinoma. Int J Cancer
111:868–872
9. Ikeguchi M, Yamaguchi K, Kaibara N (2004) Clinical signifi-
cance of the loss of KiSS1 and orphan G-protein-coupled receptor
(hOT7T175) gene expression in esophageal squamous cell car-
cinoma. Clin Cancer Res 10:1379–1383
10. Sanches-Carbayo M, Capodieci P, Cordon-Cardo C (2003)
Tumor suppressor role of KiSS-1 in bladder cancer: loss of KiSS-
1 expression is associated with bladder cancer progression and
clinical outcome. Am J Pathol 162:609–617
11. Ringel MD, Hardy E, Bernet VJ, Burch HB, Schuppert F, Bur-
man KD, Saji M (2002) Metastin receptor is overexpressed in
papillary thyroid cancer and activates MAP kinase in thyroid
cancer cells. JCEM 87:2399
12. Nagai K, Doi R, Katagiri F, Ito T, Kida A, Koizumi M, Masui T,
Kawaguchi Y, Tomita K, Oishi S, Fujii N, Uemoto S (2009)
Prognostic value of metastin expression in human pancreatic
cancer. J Exp Clin Cancer Res 21:28–29
13. Hata K, Dhar DK, Watanabe Y, Nakai H, Hoshiai H (2007)
Expression of metastin and a G-protein-coupled receptor
(AXOR12) in epithelial ovarian cancer. Eur J Cancer
43:1452–1459
14. Ikeguchi M, Hirooka Y, Kaibara N (2003) Quantitative reverse
transcriptase polymerase chain reaction analysis for KiSS-1 and
orphan G-protein-coupled receptor (hOT7T175) gene expression
in hepatocellular carcinoma. J Cancer Res Clin Oncol
129:531–535
15. Martin TA, Watkins G, Jiang WG (2005) KiSS-1 expression in
human breast cancer. Clin Exp Metastasis 22:503–511
16. Makri A, Pissimissis N, Lembessis P, Polychronakos C, Koutsi-
lieris M (2008) The kisspeptin (KiSS-1)/GPR54 system in cancer
biology. Cancer Treat Rev 8:682–692
17. Seminara SB, Messager S, Chatzidaki EE, Thresher RR, Acierno
JS Jr, Shagoury JK, Bo-Abbas Y, Kuohung W, Schwinof KM,
Hendrick AG, Zahn D, Dixon J, Kaiser UB, Slaugenhaupt SA,
Gusella JF, O’Rahilly S, Carlton MB, Crowley WF Jr, Aparicio
SA, Colledge WH (2003) The GPR54 gene as a regulator of
puberty. N Engl J Med 349:1614–1627
18. Seminara SB (2005) Metastin and its G protein-coupled receptor,
GPR54: critical pathway modulating GnRH secretion. Front
Neuroendocrinol 26:131–138
19. de Roux N, Genin E, Carel JC, Matsuda F, Chaussain JL, Mil-
grom E (2003) Hypogonadotropic hypogonadism due to loss of
function of the KiSS1-derived peptide receptor GPR54. Proc Natl
Acad Sci USA 100:10972–10976
20. Teles MG, Bianco SD, Brito VN, Trarbach EB, Kuohung W, Xu
S, Seminara SB, Mendonca BB, Kaiser UB, Latronico AC (2008)
A GPR54-activating mutation in a patient with central precocious
puberty. N Engl J Med 358:709–715
21. Silveira LG, Noel SD, Silveira-Neto AP, Abreu AP, Brito VN,
Santos MG, Bianco SD, Kuohung W, Xu S, Gryngarten M,
Escobar ME, Arnhold IJ, Mendonca BB, Kaiser UB, Latronico
AC (2010) Mutations of the KISS1 gene in disorders of puberty.
JCEM 95:2276–2280
22. Pinilla L, Aguilar E, Dieguez C, Millar RP, Tena-Sempere M
(2012) Kisspeptins and reproduction: physiological roles and
regulatory mechanisms. Physiol Rev 92:1235–1316
23. Richard N, Galmiche G, Corvaisier S, Caraty A, Kottler ML
(2008) KiSS-1 and GPR54 genes are co-expressed in rat go-
nadotrophs and differentially regulated in vivo by oestradiol and
gonadotrophin-releasing hormone. J Neuroendocrinol 3:381–393
24. Gutierrez-Pascual E, Martınez-Fuentes AJ, Pinilla L, Tena-
Sempere M, Malagon MM, Castano JP (2007) Direct pituitary
effects of kisspeptin: activation of gonadotrophs and somato-
trophs and stimulation of luteinising hormone and growth hor-
mone secretion. J Neuroendocrinol 19:521–530
25. Navarro VM, Castellano JM, Fernandez-Fernandez R, Tovar S,
Roa J, Mayen A, Nogueiras R, Vazquez MJ, Barreiro ML, Magni
P, Aguilar E, Dieguez C, Pinilla L, Tena-Sempere M (2005)
Characterization of the potent luteinizing hormone-releasing
activity of KiSS-1 peptide, the natural ligand of GPR54. Endo-
crinology 146:156–163
26. Luque RM, Cordoba-Chacon J, Gahete MD, Navarro VM, Tena-
Sempere M, Kineman RD, Castano JP (2011) Kisspeptin regu-
lates gonadotroph and somatotroph function in nonhuman pri-
mate pituitary via common and distinct signaling mechanisms.
Endocrinology 152:957–966
27. Martınez-Fuentes AJ, Molina M, Vazquez-Martınez R, Gahete
MD, Jimenez-Reina L, Moreno-Fernandez J, Benito-Lopez P,
Quintero A, de la Riva A, Dieguez C, Soto A, Leal-Cerro A,
Resmini E, Webb SM, Zatelli MC, degli Uberti EC, Malagon
MM, Luque RM, Castano JP (2011) Expression of functional
KISS1 and KISS1R system is altered in human pituitary adeno-
mas: evidence for apoptotic action of kisspeptin-10. Eur J
Endocrinol 164:355–362
28. Wilson C (1990) Role of surgery in management of pituitary
tumors. Neurosurg Clin N Am 1:139–159
29. Asa SL (1998) Tumors of the pituitary gland. In: Rosai J (eds)
Atlas of tumor pathology, 3rd series Fascicle 22, Armed Forces
Institute of Pathology, Washington, pp 1–214
30. Yan C, Wang H, Boyd DD (2001) KiSS-1 represses 92-kDa type
IV collagenase expression by down-regulating NF-jB binding to
the promoter as a consequence of IjBa-induced block of p65/p50
nuclear translocation. J Biol Chem 276:1164–1172
31. Kadokawa H, Suzuki S, Hashizume T (2008) Kisspeptin-10
stimulates the secretion of growth hormone and prolactin directly
from cultured bovine anterior pituitary cells. Anim Reprod Sci
105:404–408
Pituitary
123