harbasins report: water management strategies for estuarine and
TRANSCRIPT

HARBASINS Report:
Water management strategies for estuarine and
transitional waters in the North Sea Region.
Prepared by: Research Institute for Nature and Forest
INBO.R.2008.3
HARBASINS is a project funded under the European Regional Development
Fund INTERREG IIIB North Sea Region Program – A European Community
Initiative concerning Trans National Co-operation on Spatial Development 2000-
2006.

HARBASINS – Conservation goals 2
HARBASINS: Harmonised River Basin Strategies North Sea
Estuarine Ecosystem Functioning and Health
Habitat needs to realise conservation goals for fish in estuaries:
case study of the tidal Schelde
Jan Breine, Joachim Maes, Maarten Stevens, Ilse Simoens, Mike Elliott, Krystal Hemingway
and Erika Van den Bergh
INBO.R.2008.3

HARBASINS – Conservation goals 3
Colofon
Jan Breine, Ilse Simoens
Instituut voor Natuur- en Bosononderzoek
Wetenschappelijke Instelling van de Vlaamse Gemeenschap
Duboislaan 14, 1560 Groenendaal
www.inbo.be
email: [email protected]
Joachim Maes
VITO, Vlaams Instituut voor Technologisch Onderzoek
Integrale Milieustudies
Boeretang 200
B-2400 Mol
Mike Elliott, Krystal Hemingway
Institute of Estuarine & Coastal Studies, University of Hull
Cottingham Road Hull
HU6 7RX
UK
Maarten Stevens, Erika Van den Bergh
Instituut voor Natuur- en Bosononderzoek
Kliniekstraat 25
1070 Brussel
wijze van citeren: Breine, J., Maes, J., Stevens, M., Simoens, I., Elliott, M., Hemingway, K. & E. Van
den Bergh, 2008. Habitat needs to realise conservation goals for fish in estuaries: case study of the
tidal Schelde. INBO.R.2008.3
Rapportnummer: INBO.R.2008.3
Depotnummer: D/2008/3241/009
Druk: Ministerie van de Vlaamse Gemeenschap, Departement L.I.N. A.A.D. Afd. Logistiek – Digitale
Drukkerij
Trefwoorden:Keywords: habitat needs, conservation goals, fish, estuaries, Schelde

HARBASINS – Conservation goals 4
Index
1. Introduction
2. Conservation goals
3. Guilds for estuarine fishes with relevance to the formulation of conservation goals 3.1. Introduction
3.2. Salinity preference guilds
3.3. Estuarine use guilds
3.3.1. Freshwater resident species
3.3.2. Estuarine resident species
3.3.3. Diadromous fish species
3.3.4. Marine adventitious species
3.3.5. Marine juvenile migrants
3.3.6. Marine seasonal migrants
3.4. Flow preference guilds
3.5. Reproductive special demands guild
3.6. Trophic guilds
3.7. Stratum guilds
3.8. Tolerance guilds
4. Conservation goals for fish in estuaries
4.1. Introduction
4.2. General quality needs within the estuary
4.3. Guild-specific goals and associated habitat needs
4.3.1. Introduction
4.3.2. Salinity preference guilds
4.3.3. Estuarine use guilds
4.3.3.1. Conservation goals and associated habitat needs for freshwater
resident species
4.3.3.2. Conservation goals and associated habitat needs for estuarine
resident species and marine juvenile migrants
4.3.3.3. Conservation goals and associated habitat needs for diadromous
fish species
4.3.3.4. Conservation goals and associated habitat needs for marine
adventitious species and marine seasonal migrants
4.3.4. Flow preference guilds
4.3.5. Reproductive special demands guild
4.3.6. Trophic guilds
4.3.7. Stratum guilds
4.3.8. Tolerance guilds
4.4. Habitat status
4.4.1. Introduction
4.4.2. Habitat loss in the Schelde
4.5. Mitigating measures
4.5.1. Introduction
4.5.2. Mitigating processes in the Schelde
5. Conclusion
Annex
Table A List of fish species found in the Zeeschelde and their guilds
Table B Essential fish habitats in relation to ecological guilds

HARBASINS – Conservation goals 5
1. Introduction
The protection of nature values and functioning of aquatic ecosystems is the subject of several legal
commitments and international agreements. European legislation includes the Water Framework
Directive (WFD) as well as the Wild Bird and Habitat Directives (BHD). In addition, different
competences derive from local, regional, national or multilateral and international initiatives, each
with its own objectives and targets. Clearly these commitments apply on different spatial scales
(Europe, North Sea area, country, river basin). Consequently, evaluations of present regulations that
aim to protect environments optimally adopt an integrated, hierarchical structure (Fig. 1). Under this
approach, particular commitments aim at sustainable and integrated management but may focus on a
different spatial level of the ecosystem and its functioning. Accordingly, objectives at each level aim
at proper functioning of the ecosystem in a way that the commitments are respected. Each level feeds
back to the higher level in order to avoid contradictions. Once the objectives are set, quantitative
indicators or measurement endpoints have to be defined to measure the actual status and to compare
the ecosystem state with references set by the conservation goals. Depending on the scale, indicators
are either based on integrated data or represent an explicit measure of the state of the ecosystem.
Therefore, any monitoring scheme should provide a wide range of information so that for each level of
assessment the necessary information can be deduced. Restoration measures will then aim at restoring
the processes that generate the required habitats and species populations. The potential for recovery
remains since most species and functional groups persist, albeit in greatly reduced numbers.
This proposal describes habitat needs for fish in estuaries, defined as parts of a river under tidal
influence (Fairbridge, 1980), to assure a good status of fish populations as defined by the Water
Framework Directive (EU Water Framework Directive, WFD, 2000). This good status is obtained
when the conservation goals are fulfilled. A good habitat quality, a proper ecosystem functioning and
a good carrying capacity will assure that conservation goals are achieved. Conservation goals for fish
described by Adriaensen et al. (2005) are refined. The tidal Schelde is taken as a case study but this
approach can be applied to all North Sea estuaries. As such, this report informs to the objectives of the
HARBASINS project. In particular, the HARBASINS project addresses the question as to how
different approaches by regions in Europe, with respect to legal implementation of European law and
site specific management experiences, result in improved estuarine functioning and health as reflected
by the structure of the estuarine fish community. In this context, estuarine fish are used as a top
component in the ecosystems and one which has a high public and political resonance, hence often
being (with wading birds) the end point for the management of the systems. An accompanying report
(Cutts et al., in prep.) will consider similar aspects for estuarine wading birds.

HARBASINS – Conservation goals 6
In this report, and in order to make the method widely applicable and thus not reliant on the particular
species inhabiting any single estuary, we firstly classify fishes into guilds with relevance for the
formulation of conservation goals. We describe general and guild-specific conservation goals for fish.
Based on literature, habitat requirements are defined and then this information is used to define habitat
needs allowing a general functioning of the estuarine ecosystem and provide some suggestions to
achieve these needs. In addition we describe habitat needs at a guild level. A fish-based tool to assess
the present status relative to the conservation goals will be developed and reported in another
document.
Fig. 1. Hierarchical integration of conservation objectives depending on the spatial scale
(adapted from Wolfstein & Van den Bergh, 2004)
2. Conservation goals
Van den Bergh et al. (2005) describe quality goals for key attributes in the Schelde estuary in a
hierarchical way, starting from ecosystem physical and chemical processes down to the level of
specific habitat types and species (groups). Conservation goals for fish are defined as: achieving the
good ecological status as required by the Water Framework Directive. This ecological status can be
assessed or quantified with the classification tools that were developed for this purpose (e.g. fish-based
index of biotic integrity (EBI, Breine et al., 2007a). Fish population attributes included are: total
number of species, percentage of smelt (Osmerus eperlanus) individuals, percentage marine migrating
juveniles, percentage omnivorous and piscivorous individuals. In other words: the different ecological
guilds or functional groups that are typically present in an estuary should be able to complete their
North sea region Environmental quality OSPAR, WFD, others
River basin Environmental quality WFD, others
Estuary
Habitat
Conditions for ecological processes WFD, Natura 2000, LTV
Environmental quality Type/species: conservation goals

HARBASINS – Conservation goals 7
lifecycles and establish sustainable populations. Hence we apply the guild approach to define the
habitat needs to meet these conservation goals.
3. Guilds for estuarine fishes with relevance for the formulation of
conservation goals
3.1. Introduction
Fish can be aggregated into functional groups (guilds) according to different characteristics or along
different gradients. Fish species are adapted to the different ecological gradients in rivers, transitional
waters and coastal areas. As such fish assemblages along a river course are structured according to the
slope, discharge, temperature and oxygen content of the corresponding river section (Léger, 1945).
The longitudinal distribution of fishes occurs according to their tolerances, in a manner that lets some
progressively displace others (Roule, 1927). Therefore, fish species diversity, trophic structure as well
as reference conditions change along a river gradient in a predictable way (RCC, River Continuum
Concept, Vanote et al., 1980) as is the case in estuaries (Maes, 2000). This concept corresponds with
the distinction of different zones in rivers using type species, wetted river width and slope (Huet,
1949). The RCC is a longitudinal concept and ignores interactions between river channel, riparian
zone and floodplain. Particularly in estuaries, the exchange of water along a transversal gradient is of
crucial importance providing temporary essential fish spawning and nursery habitats in periods of
flooding either by tides (Tide Pulse) or by peak discharges (Flood Pulse, Junk et al., 1989). In terms of
energy and nutrient exchange, this lateral interaction is locally probably more important than axial
processes. It follows that in particular estuaries, river landscapes under tidal influence, support a high
biological production in both the fresh and brackish water parts. We can use these theoretical concepts
to classify estuarine fish faunas according to their ecological guilds. Root (1967) defined a guild as a
group of species that exploit the same class of environmental resources in a similar way. Species
within a guild are defined by the similarities in the use of ecological niches. In this report we group
species along environmental variables into guilds. Different categories within one guild are not always
clearly separated i.e. depending on their life cycle, species can belong to different categories (Gerking,
1994). Table A in annex is a summary of a literature review classifying each species into guilds. Only
species occurring in the presence absence list for the tidal Schelde and its tributaries are considered
(Breine et al., 2007b). These are species that should be in the Schelde when it reaches its Maximal
Ecological Potential (MEP) or its Good Ecological Potential (GEP).
The guild structure of an ecosystem is often more stable in time than its species composition and
therefore guilds have been used to assess the biological integrity of estuaries (Borja et al., 2004;
Breine et al., 2007a; Coates et al., 2004; Deegan et al., 1997; Harrison et al., 2000; Harrison &
Whitfield, 2004; Hughes et al., 2002; Jager & Kranenbarg, 2004; Moy, 2004; Salas et al., 2004;

HARBASINS – Conservation goals 8
Whitfield & Elliott, 2002). We have defined the following guilds: salinity preference, estuarine use,
flow preference, reproduction, stratum use (adults), tolerance guilds and trophic guilds. These guilds
contain relevant ecological and functional information of the estuary. Guilds describe the main
features of the fishes’ biology and the way in which they use the estuary (Elliott et al., 2007).
Therefore conservation goals and the related habitat needs can be defined guild-specific.
3.2. Salinity preference guilds
In estuaries a clear salinity gradient is present and therefore the first guild in our classification defines
groups according to their salinity preference (following a longitudinal gradient). Indeed fish can
occupy different habitats during particular periods of their life history. Salinity is an important factor
influencing the dispersal of fish and other organisms (Elliott et al., 2007). For classification of the
estuarine ichthyofauna into salinity preference classes we combined information obtained from fish
surveys and from literature (Bulger et al., 1993; Elliott & Dewailly, 1995; Schiemer & Waidbacher,
1999; Pihl et al., 2002; Quak, 1994; van Emmerik, 2003). We distinguish different classes or salinity
zones in the Schelde (Table 1). Based on the salinity preference fish species can be attributed to these
classes (Table A in annex).
Table 1: Salinity classes in the Schelde
Salt guild Salinity range (Venice system, 1959)
Freshwater <0.5
Oligohaline 0.5-5
Mesohaline 5-18
Polyhaline 18-30
In the Schelde we defined five different zones based on the Venice system. We did not differentiate
between the freshwater zone with short and long retention time (Fig. 1).

HARBASINS – Conservation goals 9
Figure 1: Salinity zones and Omes segments (numbers, Hoffmann & Meire, 1979) in the Schelde
Information about the presence of each species, in a particular salinity zone, according to its age is
used to classify species into the salinity preference guild. E.g. a species can only be considered as a
freshwater species if it occurs as a juvenile and adult in freshwater while diadromous species should
occur in all zones according to their life stage. Age is taken into account because some species show
an ontogenetic shift of habitat e.g. adult cod (Gadus morhua) live in the sea but juveniles are found in
the mesohaline zone of the estuary. The habitat requirements for juveniles are different then for adults
and should be taken into account while defining conservation goals. The results are represented in the
table A in annex.
3.3. Estuarine use guilds
In this group we have guilds that correspond largely with the ‘estuarine use functional group’ defined
by Elliott et al. (2007). According to the use of habitat which is related to the life history strategy, we
distinguish between different guilds described below (Elliott & Dewailly, 1995; Mathieson et al.,
2000; Pihl et al., 2002; Thiel & Potter, 2001).

HARBASINS – Conservation goals 10
3.3.1. Freshwater resident species
The freshwater resident species occur in the freshwater part of the estuary during their complete life
cycle. They reproduce, grow up and feed in freshwater, but can also be found in the oligohaline zone.
However, there is a difference between fish assemblages in the mainstream and in the tributaries. The
freshwater fish fauna of lowland rivers under tidal influence is normally dominated by eurytopic
species. These are typical for the potamon, the river stretch with slow running water. This area
includes the bream and ruffe/flounder zone (Huet, 19491). From survey data in the Zeeschelde for a
period from 1995 to date, we observe that the freshwater part of the mainstream is dominated by roach
(Rutilus rutilus), pikeperch (Sander lucioperca), bream (Abramis brama) and white bream (Blicca
bjoerkna). Other species such as rudd (Scardinius erythrophthalmus), gibel carp (Carassius gibelio),
ruffe (Gymnocephalus cernuus), three-spined stickleback (Gasterosteus aculeatus), perch (Perca
fluviatilis) and other cyprinids frequent this river zone. In the tributaries typical rheophilic species
including dace (Leuciscus leuciscus) and chub (Leuciscus cephalus) may dominate further upstream
where the current velocity is higher. We differentiate between Rheophilic A and B. Rheophilic A
species are confined tot the main river channel or all life stages are dependant on running water.
Rheophilic B have stages in backwaters or tributaries that are well connected with the main stream and
prefer running water or part of their life stage depends on it. In the Schelde tributaries we also have a
dominance of eurytopic species that occur in all kind of stream conditions. In our survey data we have
a dominance of roach followed by three-spined stickleback. Further upstream rheophilic species were
collected such as bleak (Alburnus alburnus) and gudgeon (Gobio gobio). The freshwater migrants
defined by Elliott et al. (2007) are included in this group.
3.3.2. Estuarine resident species
These species complete their complete life cycle within the estuary (excluding the freshwater zone).
They are sensitive to the disappearance of specific habitat such as intertidal mudflats, creeks and
marshes, accumulation of toxic substances and often show specialised parental care (e.g.
Pomatoschistus microps and P. minutus).
1 Huet (1949) described for each river zone a typical fish assemblage. Stating that water temperature and stream
velocity are important factors influencing the distribution of fishes. Both factors are dependant on the slope. He concluded that in a given geographical area, rivers of similar width, depth and slope have identical properties and similar fish populations. He distinguished 5 river types depending on the width and for each of these different zones. In the trout zone salmonid fish occur (trout and salmon). In the grayling zone we have a mixed population dominated by salmonids. In a barbel zone we expect a mixed population dominated by cyprinoids and in the bream zone we find a cyprinid fish population with piscivores.

HARBASINS – Conservation goals 11
3.3.3. Diadromous fish species
Diadromous species form an ecological group in the migration guild. Diadromous migrant species
necessarily pass estuaries when migrating between the ocean and upstream fresh water areas. The
osmotic regulation is known to be adapted in the mesohaline zone. Some species are diadromous on a
facultative basis: i.e. they may complete their life cycle in the environment where they hatch or start
migrating. We differentiate between anadromous and catadromous species since conservation goals
will be different especially concerning their spawning habitat. Anadromous species spend the adult
life history stage at sea and subsequently migrate to fresh or brackish water to reproduce either
pelagically or on gravel or sand beds e.g. twaite shad (Alosa fallax) and mullet (Liza ramado).
Catadromous species live in freshwater and reproduce at sea e.g. eel (Anguilla anguilla).
3.3.4. Marine adventitious species
Marine adventitious species or marine stragglers (Elliott et al., 2007) coincidently occur in the
estuaries and only in small numbers e.g. brill (Scophthalmus rhombus). Estuaries are not considered
crucial habitats for these species. Although there are marine adventitious species that each high tide
visit the estuary to feed (Zander et al., 1999). Their presence in estuaries reflects what happens in the
coastal zone (Jager & Kranenbarg, 2004).
3.3.5. Marine juvenile migrants
These use estuaries as a nursery area within the first year of life. It is assumed that this estuarine
habitat use results in either increased survival or increased growth and hence, enhances recruitment
success. Some species in this group are intensively fished in coastal zones (e.g. herring Clupea
harengus, Sole Solea solea, plaice Pleuronectes platessa, Cod Gadus morhua and sea bass
Dicentrarchus labrax).
3.3.6. Marine seasonal migrants
Marine seasonal migrants use the estuary when conditions are favourable, usually during post juvenile
stages and on a seasonal basis. We can compare this group with the marine estuarine –opportunistic
group described by Elliott et al. (2007). They enter estuaries in substantial numbers. A typical marine
seasonal migrant in the Schelde is sprat (Sprattus sprattus).
3.4. Flow preference guilds
To define conservation goals for the fish species in the freshwater estuary we divide the fish fauna
according to their flow preference along a lateral gradient. This especially concerns habitat criteria for
freshwater species but also diadromous and some estuarine resident species. We use literature to

HARBASINS – Conservation goals 12
define species preferences (Aarts & Nienhuis, 2003; Elliott & Dewailly, 1995; Elliott & Hemingway,
2002; Huet, 1949; Kroes & Monden, 2005; van Emmerik, 2000; Quak, 1994; Schouten & Quak,
1994). In the Annex table A we have defined rheophilic species (obligate and partial) which need lotic
water, eurytopic species and limnophylic species which prefer rivers with a semi lentic conditions or
standing waters with macrophytes (stagnophilic). Species from the latter guild are not common in the
mainstream of estuaries.
3.5. Reproductive special demands guild
Spawning involves the presence of ripe adults and the production of eggs. It is a functional element of
the adult period in fish ontogeny. Therefore the functional status of an ecosystem should be reflected
in the composition of its reproducing fish assemblage (Copp, 1989). Although fishes will accept some
variation in spawning conditions, the known reproductive tolerances of various fishes or group of
fishes can be used to describe a functional change in an ecosystem’s development. If an ecosystem
changes it may no longer offer the conditions necessary for one species’ functional state and
processes, although the new conditions may be acceptable for a previously absent species. Several
attempts were made to develop a classification based on reproductive styles. The reproductive guild
concept is difficult especially due to insufficient information. Knowledge of preferred spawning
grounds and features of reproductive behaviour are essential to define conservation goals. We used
information from Aarts & Nienhuis (2003), Balon (1975, 1981), Costa et al. (2002), Elliott et al.
(2007) and van Emmerik (2003) to determine special spawning requirements. The special demands we
investigated are: special habitat demands (gravel, stone, plants and floodplains), parental care and nest
building. Gravel spawners depend on the presence of gravel to deposit their eggs (e.g. grayling,
Thymallus thymallus). Diadromous fish can be gravel bed spawners with benthic larvae e.g. river
lamprey (Lampetra fluviatilis) spawns on gravel or sand and the European sturgeon (Acipenser sturio)
females spawn above gravel and stones while the sea lamprey (Petromyzon marinus) need gravel and
build a nest high upstream. Stone spawners depend on the presence of rocks or stones to deposit their
eggs (e.g. Spirlin, Alburnoides bipunctatus). Sand spawners need the presence of a sand soil to deposit
their eggs (e.g. Burbot, Lota lota). Ostracophilic spawners deposit their eggs in living shells or houses
of other organisms (e.g. Bitterling, Rhodeus sericeus) while speleophilic spawners deposit eggs in
cavities (e.g. Bullhead, Cottus gobio). Soil spawners that deposit eggs on the bottom are not
specialised (e.g. White bream, Abramis bjoerkna) which applies also for diadromous species that are
open water spawners and are not included in the list. The results are presented in the Annex Table A.

HARBASINS – Conservation goals 13
3.6. Trophic guilds
Feeding guilds may be used to assess the structure and functioning of estuarine fish communities as a
stable trophic network is reflected by a healthy trophic composition (Breine et al., 2007). As Elliott et
al. (2007) mention, it is essential to understand the relations within the food webs so that the
conservation goals can ensure that the trophic network is optimal. The feeding guilds and feeding
preferences of dominant types are present in most estuaries (Costa & Elliott, 1991). Many estuarine
fish are either generalist feeders or opportunists. Therefore absolute separation is difficult and
combinations should be allowed (Elliott & Dewailly, 1995). To define the trophic guilds one is faced
with some problems:
1. The diet of many fish species varies with the availability of food.
2. Many species undergo an ontogenetic shift.
3. Some guilds overlap, e.g. piscivores are part of the vertivores and the planktivores are part of
the invertivores.
From literature we define the diet for juveniles and adult specimens (last step in the food chain) based
on the diet preference of the species. We use information from Batzer et al. (2000), Belpaire et al.
(2000), Breine et al. (2001), Breine et al. (2004), Breine et al. (2007a), Bruslé & Quignard (2001), De
Nie (1996), Elliott & Dewailly (1995), Elliott et al. (2002), Gerking (1994), Gerstmeier & Romig
(1998), Jager & Kranenbarg (2004), Maitland (2000), Mathieson et al. (2000), Muus et al. (1999),
OVB (1988) and van Emmerik (2003) to assign species to food classes. We have following groups
(Table A in annex):
Planktivores (P): feed mainly on zooplankton and phytoplankton
Benthivores (B): feed mainly on benthic invertebrates
Planktivores and benthivorous (PB): feed on both plankton and benthic invertebrates
Piscivores (F) feed mainly or only fish of the same or different species
Benthivorous and piscivorous (BF): feed on both fish and benthic invertebrates
Omnivores (O): feed on animals and plants
Benthivorous and herbivorous (BP): feed on benthic invertebrates and plants
Herbivores and planktivores (PPB): feed on plants, zooplankton and zoobenthos
Herbivores and phytoplankton (PP): feed on plants and phytoplankton
Vertivores and piscivores (VF): feed on fish and vertebrates
The feeding mode functional group presented by Elliott et al. (2007) takes some of these groups
together and considers also additional groups. However we think that the above groups explain
sufficiently the functioning of the estuary.

HARBASINS – Conservation goals 14
3.7. Stratum guilds
For the stratum guild or vertical distribution guild the use of the water column by adult species is taken
into account. We distinguish three groups. The benthic fishes or bottom dwellers live in or on the
bottom (e.g. flounder Platichthys flesus). Benthic species are sensitive to the degradation of benthic
habitats (dredging, sedimentation etc…). A second group consists of demersal fish that occur nearby
the bottom (e.g. bream). Pelagic fish occur in the open water sometimes close to the surface (e.g.
Twaite shad). Stratum guilds in the table A (annex) are defined according to Breine et al. (2004),
Breine et al. (2007a), Elliott & Hemingway (2002), Hughes & Oberdorff (1999), Mathieson et al.
(2000) and van Emmerik (2003).
3.8. Tolerance guilds
In Belgium the tolerance guilds have been used by Belpaire et al. (2000), Goffaux et al. (2001), Breine
et al. (2004) and Breine et al. (2007a). Within these guilds we differentiate between habitat sensitivity
and pollution intolerance. This includes information about sensitivity to canalisation, dredging and
fragmentation on the one hand and tolerance to oxygen deficiency and pollution sensitivity (organic
pollution) in general on the other hand. Oxygen demands are an important issue for conservation goals
(Maes et al., 2007a,b). In table A (annex) we compiled information from different authors: Belpaire et
al., 2000; Berrebi dit Thomas et al., 1998; Breine et al., 2001; Breine et al., 2007a; De Nie, 1996;
Grandmottet, 1983; Kruk, 2007; Mann, 1996; Muus et al., 1999; Nijsen & De Groot, 1987; OVB,
1997; OVB, 1988; Phillipart & Vranken, 1983a; Phillipart & Vranken, 1983b; Reitsma, 1992;
Riemersma, 2000; Turnpenny et al., 2004; Van Aelbroeck, 1910; Vandelannoote et al., 1998; Vanden
Auweele, 1995; van Emmerik, 2003; Verneaux, 1981 and Williot, 1991. We define each species as
tolerant or intolerant to pollution i.e. low oxygen and organic pollution. Habitat sensitivity is
expressed as fragmentation sensitivity and the need for undisturbed shelter since some species have
special shelter demands or strategies (van Emmerik, 2003).
4. Conservation goals for fish in estuaries
4.1. Introduction
Estuaries serve several ecological functions at different spatial and temporal scales. Conservation
goals and associated indicators are therefore appropriate only if they target a specific scale. Here, we
focus on four geographical scales. On a regional scale (i.e. North Sea), it is hypothesized, though yet
to be assessed in an analytical way, that the various North Sea estuaries contribute significantly to fish
recruitment to adult populations of especially marine fish but also of estuarine, diadromous and
freshwater fish species. Elliott et al. (1990), for example, indicate the way that estuaries support

HARBASINS – Conservation goals 15
nursery stages which become the adult populations. For the purpose of this exercise, fish recruitment
is defined here as the ingress of 0-group fish to the 1-group old, one year after hatching. Basin-wide,
the estuary (mesohaline and polyhaline) is per unit surface area the most productive part of the
watershed. The elevated estuarine productivity relates to the nursery function that estuaries perform
for juveniles of offshore spawned marine, estuarine and catadromous fishes and for upstream spawned
freshwater and anadromous fishes. Within the estuary different salinity zones can be identified.
Finally, within different salinity zones, species are adapted to particular habitats. Here we also
consider the essential estuarine habitats for fish. Essential habitats are habitats (waters and substrata)
that are necessary to spawn, breed, feed and grow to maturity and that act as pathways in diadromous
migration. In other words habitat demands depend on biological aspects. Table 2 illustrates the relation
between the biological aspect and habitat demand. Estuaries are subject to pronounced seasonal cycles
in species occupancy. Wintering waterfowl probably provide the best example but also fish
communities - as integral part of estuarine ecosystems - undergo seasonal changes in species
composition. We hypothesise that seasonality in estuarine fish community reflects the year-round
fulfilment of the nursery function.
Table 2: Biological aspect versus habitat demands, adapted from Milner (1984)
Biological aspect Habitat demand
Reproduction
Migration to spawning place Sufficient water depth, appropriate flow velocity
and absence of physical and chemical (water
quality) barriers
Spawning Appropriate substratum if not pelagic
Incubation Substratum stability, appropriate water
temperature and salinity, sufficient oxygen and
water movement
Feeding and growth
Availability of food Bank and aquatic vegetation, substratum allows
macro-fauna production, transport of organic
material
Optimal use of energy for movement and to seek
food
Shelter and protection (obstacles, tree roots)
diverse flow pattern, diverse substratum and
vegetation and an appropriate water temperature
Protection
Against physical displacement
Against predation
Against inter- and intraspecific competition
Habitat diversity: variable substratum (protection
and hiding places), flooded banks, tree roots,
branches, trees, debris, vegetation, pools,
turbidity etc…
The essential fish habitat concept derives from US legislation. In a European context it may be
interpreted as habitat directive area. Here, we identify essential fish habitats for the different

HARBASINS – Conservation goals 16
ecological guilds (Table B in annex). This list differs from the one proposed by Pihl et al. (2002) in
that we give more weight to tidal freshwater and freshwater fishes.
Conservation goals and restoration measures often target specific habitats or single species. However,
they have influence on larger spatial and temporal scales as well, so we argue that tools to evaluate the
effectiveness of restoration practices must comply with these scales. We will first set up general
functional needs for fishes and then, by considering the different functional groups, more guild-
specific goals and associated habitat needs.
4.2. General quality needs within the estuary
Fish need an essential level of estuarine water quality. This essential level will allow that the estuary
contains a healthy sustainable fish population. Van den Bergh et al. (2005) set quality goals to
evaluate the rehabilitation process for essential habitats within the Schelde estuary. From those we
select water temperature, turbidity, discharge flow rate, salinity and dissolved oxygen (DO) as key
ecological quality factors for the fish population (Maes et al., 2007a). The physical and chemical
conditions set up and reflect the main fundamental niches - for the substratum and the water column
(see figure in Borja & Elliott, 2007).
In addition different types of migration should be possible in one estuary.
Temperature directly or indirectly controls physiological rates, hatching success (Maitland & Hatton-
Ellis, 2003), juvenile growth rate (Aprahamian, 1988), year class strength (Aprahamian &
Aprahamian, 2001), juvenile seaward migration (Limburg, 1996), adult upstream movements
(Aprahamian, 1988). More generally, temperature also relates to natural mortality of teleost fishes
(Pauly, 1982) as well as to prey abundance. It is of note that with a temperature increase a double
challenge with respect to hypoxia is created: (1) the oxygen demand increases because of increased
metabolism at higher temperatures and (2) the solubility of oxygen is negatively correlated with the
water temperature. At present we are faced with a global warming resulting in a steady increase of the
water temperature. This is a worldwide effect and should be treated as such. On a basin-wide level we
should be aware that this temperature increase affects the DO. We should therefore implement
mitigating measures that increase the DO.
Turbidity decreases the light penetration which has a direct effect on the planktonic primary
production such as the development of diatoms. In addition photosynthesis will be reduced hence
lower DO concentrations. The high load of suspended matter can clog the gills although in some
estuaries, some species are adapted to it (Blaber & Blaber, 1980). It results also in sediment

HARBASINS – Conservation goals 17
accumulation which can have a negative effect on the benthic organisms although estuaries are
affected by natural erosion-deposition cycles and so the benthos is adapted to the elevated turbidity.
As many other estuaries the Schelde is turbid with poor light penetration. This is a natural
phenomenon limiting the production of estuarine phytoplankton and thus limiting the food availability
for zooplankton. As such much microplankton primary production may be the result of resuspended
microphytobenthos. Zooplankton is well adapted to high suspended sediment levels, together with
attached and free-living bacteria which form their food source. The high turbidity reduces visibility
and has an impact on visual predators since it reduces the most important visual characteristics of
prey: contrast with background, size, movement, shape, colour and unusual form (Wootton, 1992).
Dredging causes a resuspension of the bottom sediments and has an effect on the turbidity. Both
temperature and turbidity should be such that they do not hamper the life of fish occurring in
the estuary.
Another important factor influencing the habitat suitability for fish is the freshwater discharge. This
habitat suitability versus discharge for different stages of development of species is defined by depth,
velocity and substratum (Angermeier & Karr, 1983; Capra et al., 1995) as well as its influence on
salinity regime. In the downstream part of the estuary, where the cross-sectional area is large
compared to the cross-sectional area of the Schelde, the river discharge can be disregarded
(Horrevoets et al., 2004). Upstream, where the cross-sectional area approaches that of the river, the
fresh water discharge gains importance over the tidal flow and affects the tidal range (Horrevoets et
al., 2004). In the upstream area discharge should not create floods.
Salinity is influenced by water discharge and tidal intrusion and its importance has been described
above. The balance between freshwater (riverine) flows and the tide should assure the salinity
gradient. Any disrupter in salinity will have an impact on the distribution of fish. Salinity determines
the distance that a species is capable of penetrating the estuary (McLusky & Elliott, 2004). Its effects
on organisms is complex and is influenced by other variables e.g. temperature and discharge
(McLusky & Elliott, 2004).
The DO has an effect on the distribution of fish (Reincke et al., 1992) and too low values may result in
fish mortality (Alabaster et al., 1979; Erichsen Jones, 1964; Scholz, 1986; Turnpenny et al., 2004). Of
course certain fish can adapt to hypoxia or avoid it and move upwards or laterally but this reduces the
habitat availability. Fish larvae and juvenile fish are less successful at leaving regions with low DO
concentrations while adults mostly escape from those areas and form typical distributions along the
oxygen gradient. Young life stages of fish may be more susceptible to low DO effects (Turnpenny et

HARBASINS – Conservation goals 18
al., 2004). Adriaensen et al. (2005) propose a dissolved oxygen (DO) concentration of 5 mg l-1
as a
lower limit for essential water quality with respect to estuarine fish. Maes et al., (2007b) state this
threshold of 5 mg l-1
as the DO minimum for surface waters in Flanders. This limit is based on DO
criteria for US estuaries as outlined by USEPA and on empirical models describing the response of
estuarine fish to different oxygen concentrations (Maes et al., 2005, 2007a,b). Experiments to measure
the DO requirements of fish in the Thames estuary produced, however, limits that are significantly
lower than 5 mg l-1
(Turnpenny et al., 2004). Table 3 shows DO standards developed by the Thames
Tideway Strategy Group (TTSG) aimed specifically at the Thames Tideway but having more general
application in other British transitional waters (Turnpenny et al., 2006). However, fish populations in
the Tideway were likely to be sustainable (<10% mortality per annum) under this DO standards
regime. We propose to apply the same standards for the Schelde estuary. The objectives are directly
linked to these standards (Table 3).
Table 3: DO standards proposed by the TTSG (Turnpenny et al., 2006)
DO (mg l-1
) Return Period (years) Duration (# of 6 hour tides)
4 1 29
3 3 3
2 5 1
1.5 10 1
Note: the objectives apply to any continuous length of river ≥ 3 km.
Duration means that the DO must not fall below the limit for the
stated number of tides.
A tide is a single ebb or flood.
The bases for these standards are:
- The one week standard (4 mg l-1, 1 yrRP, >29 tides) was selected to ensure protection against chronic
effects such as depression of growth and avoidance of hypoxic areas.
- The 24 h standard (3 mg l-1, 3 yrRP, >3 tides) and the 6 h standard (2 mg l-1, 5 yrRP, > 1 tide) were
selected to provide protection to stocks.
- The lowest standard (1.5 mg l-1) was included to ensure protection from mass mortalities.
A migration possibility within the estuary and between the estuary and the ocean is an important
factor for the sustainability of migrating species. These species use the estuary as a migrating route
especially for spawning. According to Lucas and Barras (2001) besides horizontal and vertical
migrations we have other types towards essential habitats: feeding migrations, refuge-seeking
migrations, spawning migrations and post-displacement movements, recolonisation and exploratory

HARBASINS – Conservation goals 19
migrations. Physical and chemical (water quality) barriers can be present in different places along the
estuary and should therefore be alleviated, removed or made passable.
4.3. Guild-specific goals and associated habitat needs
4.3.1. Introduction
Sediment type, degree of vegetation cover, tidal elevation, slope of intertidal area and depth of subtidal
area determine the suitability of a habitat for a species. Water currents influence fish abundance and
distribution. Average current velocities determine the sediment characteristics and consequently the
resultant benthic communities. Dietary preferences of fish will in turn influence their occurrence on
specific sediments. The habitat complexity, productivity and the area of a single habitat type are major
contributors to determine the fish diversity of an area (Wootton, 1990). The width of the intertidal
zone will qualify its potential as a feeding ground or a nursery area. The depth of the subtidal area may
affect its suitability as a habitat for fish. Rocky shores and bottoms offer more niches than the uniform
habitat of soft sediments which is the case for the Schelde estuary. Estuaries are very dynamic systems
and geomorphological processes such as erosion and sedimentation affect the habitat value of areas for
fish. We take these criteria into consideration when defining habitat needs.
4.3.2. Salinity preference guilds
We defined five guilds of which three occur in the Zeeschelde: freshwater species, oligohaline and
mesohaline species. The conservation goals and associated habitat needs for these species are identical
to those defined for the estuary use guilds.
4.3.3. Estuary use guilds
4.3.3.1. Conservation goals and associated habitat needs for freshwater resident species
The presence of freshwater species is restricted to the freshwater, oligohaline and mesohaline parts of
the estuary. Conservation goals for this group specifically target the freshwater tidal part of the
estuary. For this group we define two conservation goals (CG):
CG1: Presence of a sustainable population of freshwater species (Annex Table A)
CG2: Presence of core populations of type specific species (for Schelde HD annex II and red list
species)
CG1 is the most essential goal since CG2 comprises species from lists that to our opinion should be
revised. In order to realise these goals particular conditions in the freshwater habitat should be present.
Tidal freshwater rivers and wetlands constitute rare habitats (Pihl et al., 2002). They are situated above
the saltwater boundary but still experience tides, which distinguish them from the more common

HARBASINS – Conservation goals 20
bream zone. The tidal amplitude creates a cross sectional gradient with a subtidal riverbed, intertidal
mudflats and higher located freshwater tidal marshes which are, in North Western Europe, typically
dominated by reed and willow species. Species compositions of freshwater tidal areas tend to be
influenced by landscape patterns just above the tidal limit and can thus be predicted using the zonation
concept. Through colonisation from upstream areas, tidal freshwater rivers may have fish faunas from
the adjacent barbel zone (as in the Thames) or from the bream zone (as in the Elbe or the Schelde).
The freshwater tidal area of the Thames is dominated by dace, a rheophilic A species. The freshwater
tidal area of the Schelde is dominated by representatives of eurytopic fishes such as roach, pikeperch
and bream, this as a result of the flat landscape and a river system that is categorized as bream zone.
Rheophilic B, species that need well connected backwaters, may lack as these habitats are absent and
replaced by intertidal marsh. According to Ward (1989) connectivity, succession and ecotones define
the functional and structural elements of the four-dimensional nature of running water. Increased
habitat and shoreline complexity is beneficially for most of the fish species (Shlosser, 1982; Lobb &
Orth, 1991; Grenouillet et al., 2000; Pollux et al., 2006, Kruk, 2007). Eurytopic fish benefits from a
good hydrological connection between the different components that together constitute the river
corridor (channel, marshes, floodplain) and from the presence of tributaries (Pollux et al., 2006).
Especially supratidal floodplains are considered essential habitats as they provide suitable spawning
and juvenile habitats (Junk et al., 1989, Ward et al., 1999, Scholten et al., 2003). In the Schelde the
rheophilic A species are all diadromous species except one (dace).
Given differences in life history characteristics and habitat use, habitat needs for rheophilic species
probably take into account ecological gradients along a longitudinal section. For eurytopic species, on
the other hand, the possibility of lateral exchanges between floodplains, marshes and the river channel
are crucial factors. As fish grow their habitat requirements change. Therefore, habitat preferences
often switch in the course of the development.
Limnophylic species may occur but estuarine habitats are not essential for them. Based on habitat
preferences, efforts should focus on rheophilic species as well as eurytopic fishes.
Using present knowledge of rheophilic A and eurytopic fishes, we suggest three habitat needs (HN):
HN1: On a basin-wide scale, the ecological connectivity along longitudinal and transversal river
gradients permits the development of a sustainable fresh water population of rheophilic A and
eurytopic fish species in the estuary. This includes an absence of physical and chemical (water quality
e.g. good DO) barriers.

HARBASINS – Conservation goals 21
HN2: On an ecosystem scale, the presence of floodplains along the tidal freshwater part of the estuary
assures the annual recruitment of freshwater eurytopic fishes. This goal relates to the hypothesis that
floodplains represent a critical factor in life history of eurytopic fishes.
HN3: The availability of sheltered diversified intertidal habitat surfaces and subtidal areas with a
diverse food supply in the freshwater estuary are essential as nursery and feeding grounds for
eurytopic and rheophilic species. Surface roughness is a measure of diversity for upstream parts of the
river (Chow, 1973) and the higher its value the more natural the river.
4.3.3.2. Conservation goals and associated habitat needs for estuarine resident species
and marine juvenile migrants
Marine juvenile migrants and estuarine resident species respectively use estuaries often during a
particular part or all of their life cycle. We consider two resources of importance: space and food and
the former can be subdivided into amount of area used and amount of time the area is used (extent
versus duration). The main difference between the guilds is the time spend in the estuary. Marine
species strictly spawn at sea while estuarine species may extend their spawning habitat to estuaries
sensu strictu. However, for the purpose of this exercise, marine juvenile migrants and estuarine fishes
are grouped together as they share many biological and ecological properties. Typical estuarine
species, such as common goby (Pomatoschistus microps), pipefishes (Syngnathidae) and eelpout
(Zoarcus vivparus) are therefore able to complete their life cycle both in the brackish water parts of
estuaries as well as outside estuaries in a fully marine environment. Similar conservation targets may
thus apply on both marine and estuarine fishes. For estuarine resident fishes, estuaries can be regarded
as essential habitats providing habitats for spawning, feeding and growing. It is situated in the salinity
range from the oligohaline to the marine. Marine juvenile fishes were defined as those species that use
estuaries as a nursery area as 0-group individuals. In particular, shallow areas in the marine and
brackish part of estuaries that are either turbid or vegetated may qualify as fish nurseries. The same
habitats as listed for estuarine resident species apply as essential habitats for marine juvenile migrants
and need preferential protection (Table B annex) another reason why these two groups are combined.
These habitats are essential and probably contribute significantly to fish recruitment or the survival of
recruits which have migrated into the estuary. The relationship between nursery size and fish
recruitment is postulated by Rijnsdorp et al. (1992). Under this hypothesis, we accept that increasing
total estuarine fish nursery habitat has a positive effect of the recruitment of marine juveniles and can
act as a mitigating measure. Estuaries are characterised by seasonal patterns in species composition as
a result of successive “migration waves” of estuarine resident and marine juvenile migrant individuals.
These migrations relate to species specific life history strategies and are highly influenced by

HARBASINS – Conservation goals 22
processes occurring in the sea. Complete seasonal niche partitioning of the particular estuarine
ecosystems suggests optimal functioning of the fish nurseries.
Based on the present knowledge of estuarine resident species and marine juvenile migrants, we
suggest following conservation goals:
CG4: On a regional and basin-wide scale marine juvenile migrants (0-group individuals) should be
present in accordance to the season as well as a sustainable population estuarine species.
CG5: Preserving the seasonal dynamics of the estuarine fish communities and marine juvenile
migrants in the estuary is a prior goal.
CG6: On habitat level different life stages of estuarine species should be present according to the
habitat type and young marine individuals are present according to the season.
In order to achieve these goals we propose following habitat needs:
HN4: On regional and basin-wide scales we aim at an estuarine nursery size that is sufficiently large
(temporal and spatial) such that it contributes significantly to the recruitment of young marine and
estuarine fish populations or at least have several small nursery areas within a geographic area which
are connected. Size is only one criterion, the estuary should have an appropriate water depth, its shape
should be convenient for the larvae, connectivity should allow the larvae to move to adult habitats and
the physical-chemical conditions (DO and salinity) should not be restrictive. Further research is
needed to quantify the nursery function of estuaries. An alternative is provided by the estimation of
carrying capacity of estuaries for fish i.e. the number of individuals an estuary can support per unit of
surface area. Elliott et al. (2007) indicate the methods of defining carrying capacity but as yet these
have not been tested for estuarine fishes. In order to examine the nursery function of the estuary for
juveniles of a particular species we should assess whether its contribution per unit area to the
production of individuals that recruit to adult populations is greater, on average, than production from
other habitats in which juveniles occur (Beck et al., 2001). According to these authors we should
assess different biotic and abiotic factors including landscape factors.
HN5: Seasonal migrations can only occur if conditions in the estuary are favourable for the fish i.e.
temperature and DO conditions should be optimal and food should be available. Physical and chemical
(water quality) barriers should be absent.
HN6: The presence of shallow, low dynamic (as opposed to high tidal energetic areas) habitats
(sheltered mud flats, salt marches; reed beds,…) that provide protection and a high and continuous
supply of food should be insured. We should therefore define the characteristics of mud flats and salt

HARBASINS – Conservation goals 23
marches with a good quality and define their dimensions allowing the development of a dynamic and
diversified habitat. The mudflats should have an optimal sediment and tidal position for the
maintenance of suitable prey biomass (Elliott et al., 1998 and McLusky & Elliott, 2004).
4.3.3.3. Conservation goals and associated habitat needs for diadromous fish species
The (sub)tidal transition zone between rivers and oceans is a vital habitat for diadromous fish linking
spawning grounds with adult habitat. Some anadromous fish species, for instance twaite shad (Alosa
fallax), use the freshwater intertidals as spawning habitat. Shallow areas or vegetated habitats
throughout the estuary serve as essential nurseries for 0-group anadromous and catadromous fish (see
intertidal creeks and marches Table B).
Based on the present knowledge of diadromous species, we suggest:
CG7: On a regional scale, endangered anadromous populations of in particular Atlantic salmon,
Atlantic sturgeon, houting (Coregonus oxyrhynchus) and allis shad (Alosa alosa) should have self
sustainable populations. At present eel is also a species that can be considered as being at risk (Robinet
& Feunteun, 2002; ICES, 2006). For the Convention on International Trade in Endangered Species of
Wild Flora and Fauna (CITES) a work document has been released in which it is proposed to
reconsider the status of eel in Europe (CITES, 2006).
CG8: Basin-wide, self sustaining populations of geographically-relevant diadromous species (E.g.
twaite shad, river lamprey, smelt, thinlip mullet and flounder for the Schelde) should frequent the
estuary.
CG9: Within the estuary diadromous individuals from the > 0-group should be present.
We suggest following habitat needs:
HN7: On a regional scale fishing activities should be reduced and controlled.
HN8: Within a basin-wide level and within the estuary a good water quality (DO concentration) and
ecological connectivity permit unconstrained movements of diadromous fish between spawning and
nursery grounds and the adult habitats.
HN9: To assure the nursery function of the estuary the necessary amount of space and the proper
morphology to provide shelter and foot for > 0-group diadromous individuals should be present. This
part should also have a good water quality. We need to quantify the concept of a fish nursery (Beck et
al., 2001).

HARBASINS – Conservation goals 24
HN10: The presence of upstream situated spawning habitats and sufficient intertidal habitat are
required for the success of the rehabilitation programs of diadromous populations. The upstream
habitat should have clear and oxygenated water. No dredging activities should occur but mud should
be absent so that spawning can occur on gravel or sand.
4.3.3.4. Conservation goals and associated habitat needs for marine adventitious and
marine seasonal migrant species
We will not further consider conservation goals for these two ecological guilds since estuaries are not
considered as essential habitats for these ecological groups although it is of note that if the above
Conservation Goals are achieved then these groups will also be supported in the estuaries. Marine
seasonal migrants facultative exploit estuaries if conditions are favourable. Sprat is a good example of
such a life history strategy. Their presence in estuaries, however, reflects a good hydrological
connection between estuarine habitats and the ocean. Ecological connectivity requires suitable water
quality and quantity between habitats. Connectivity issues include poor water quality for transient
fishes, temporal (un)availability of tidal creeks due to marsh rising as a result of channel deepening or
strong tidal currents preventing upstream migration. These issues return for other ecological groups
and in particular for diadromous fish and are treated in this chapter.
4.3.4. Flow preference guilds
The conservation goals for these guilds are incorporated in freshwater resident species conservation
goals. Indeed freshwater species are rheophilic, eurytopic or limnophylic. We consider the latter group
less relevant for the estuary. In addition most op rheophilic A species are diadromous. As a summary
we repeat the main goal and essential habitat needs:
The main goal is the presence of a sustainable population of freshwater species including rheophilic A,
B and eurytopic species.
Rheophilic and eurytopic species need the ecological connectivity along longitudinal and transversal
river gradients which is translated in a good DO concentration and absence of barriers.
Accessible floodplains assure recruitment possibilities for eurytopic species.
Sheltered diversified intertidal habitat surfaces and subtidal areas with a diverse food supply in the
freshwater estuary are essential as nursery and feeding grounds for eurytopic and rheophilic species.
4.3.5. Reproductive special demands guild
We define special demands for the different groups within the salinity preference guild. The
importance of estuaries for recruitment of marine species decreases upstream along the decreasing
salinity gradient (Elliott et al., 1990). Reproductive habitats of marine fishes are generally located

HARBASINS – Conservation goals 25
offshore so estuarine conservation goals will specifically target younger life history stages. And those
will be similar to estuarine resident species demands. Most estuarine resident species, including partial
residents that live intertidally as adults during the breeding season, spawn within the intertidal zone.
Estuarine residents often produce demersal eggs since pelagic eggs and larvae cannot withstand the
occurring currents and wash-out events. For example, smelt (Osmerus eperlanus) will produce its eggs
in areas less liable to wash-out thus ensuring its young are retained in the estuary. Other species
migrate into the intertidal to spawn during high water levels. The benefits of intertidal spawning
follow from the increase of the embryonic development rate when emergent due to the food
availability and from temporal and spatial refuge for adult spawners and embryos. Success of
recruitment is affected by habitat factors while many larvae behave as habitat generalists and so
shelter and associated food availability will still be important. Sheltered microhabitats (e.g. tidepools)
will provide refuge from predators or competitive species but are subjected to high DO, salinity and
temperature fluctuations. Thus environmental variation will result in different species having
successful recruitment at different times. This is important for the coexistence and persistence of
assemblages. The upstream spawners consist of freshwater and diadromous species and will have
specific demands.
Based on the present knowledge of special reproductive demands we suggest:
CG10: On an estuary and habitat level offspring from diadromous, estuarine resident and freshwater
species should be present.
This presence is assured thanks to following habitat needs:
HN11: On a regional and basin-wide scale good water quality (DO concentration), a lack of physical
barriers and ecological connectivity should permit unconstrained movements for diadromous species
between the spawning grounds and adult habitat.
HN12: Within the estuary a variety of undisturbed macrohabitats should be present along the
longitudinal and transversal gradient. This includes in the mainstream sheltered habitats and
floodplains while upstream accessible spawning habitats such as sand beds, habitats with gravel and/or
stone bottom with clear and oxygenated water should be present.

HARBASINS – Conservation goals 26
4.3.6. Trophic guilds
Hydrographic regime and site specificity/substratum are factors controlling feeding controlling (Elliott
et al., 2002). Tidal elevation influences population size in fish. There is a relationship between the size
of the fish and the depth: as the body length increases, the depth of the water that the fish inhabits
increases (Gibson et al., 1995). Changes in tidal elevation by sea level rise or dredging activities may
increase the time for feeding but reduce the area and hence the carrying capacity. Salinity and DO
concentrations may also affect the feeding behaviour of fish. Intertidal areas are well defined as
juvenile fish feeding areas (Costa & Elliott, 1991). Mud and sandflats are important nursery areas for
plaice (Lockwood, 1972) as well as feeding areas for sea bass and flounder (Elliott & Taylor, 1989a,
b). The importance of mud and sandflats has been described above (see Elliott et al., 1998). Though in
general fish in estuaries are opportunistic (Breine et al., 2007b) food should be available and different
levels in the food web should be present. It may be assumed that the abundance and distribution of fish
feeding in the estuary is related to the quantity of food available in the intertidal and subtidal areas.
Though in general smaller in surface the intertidal area is the most important food source. Due to land-
claim the benthos community decreases and the fish communities in terms of species richness,
abundance and biomass reduce. Not only has the reduced the food supply diminished the estuaries’
value as nursery area, but also the loss of habitat.
We suggest following goals:
CG11: Basin-wide and within the estuary all species forming an undisturbed trophic web should be
present. This includes individuals from the groups represented in Table A.
CG12: According to the habitat type different species representing different links within the trophic
chain are present.
We therefore suggest following habitat needs:
HN13: Basin-wide and a variety of site substratum should be present along a longitudinal gradient.
HN14: The estuaries should have an undisturbed hydrographic regime.
HN15: The presence of shallow, low dynamic habitats providing a high and continuous supply of food
should be protected for estuarine resident and marine juvenile migrants. The availability of sheltered
diverse intertidal habitat surfaces and subtidal areas with a diverse food supply in the freshwater
estuary are essential feeding grounds for the freshwater resident species.

HARBASINS – Conservation goals 27
4.3.7. Stratum guilds
The stratum in which a fish species occur can be more or less sensitive to human impact. The most
widespread mechanism employed in the maintenance of navigable channels is physical dredging of
sediments from the bed of the channel for subsequent disposal elsewhere in the estuary, in coastal
waters or to land. At the disposal sites, fine-grained sediment may smother the bed and kill or cause a
change in the benthic community affecting benthic and demersal species. Clay particles will remain in
suspension for a longer time and high loads of suspended solids are harmful to pelagic fish (Bruton,
1985).
The goals we propose are corresponding to the more general goals or were already mentioned in one
of the described guilds. In general an undisturbed stratum should be considered as the ultimate habitat
need. As already mentioned dredging can not be stopped and therefore we suggest the following:
HN16: Within the ecosystem fish the ecological connectivity should allow fish to avoid effects.
HN17: Undisturbed habitat should be present.
These two habitat needs are also generic CG.
4.3.8. Tolerance guilds
Conservation goals for species tolerance to oxygen deficiency have been treated within the general
goals. Habitat sensitivity concerns the impact of fragmentation on species which depends on their
tolerance, the needed habitat, their dispersion ability (swim and jump capacities) and reproduction
capacities. Species adapted to a specific habitat type are generally less adaptive and therefore more
impacted by fragmentation e.g. rheophilic species (obligate). Eurytopic species (e.g. bream and rudd)
are less sensitive to fragmentation, while those needing a large habitat (eel, bleak, wells catfish,…) are
more sensitive. Species that do not migrate (brook lamprey, spined loach,…) are more sensitive than
species with a higher dispersion capacity (eel). Species having a high fecundity can easily re-colonize
new habitats. Species with parental care or nest builders having less offspring are more sensitive to
fragmentation Kroes et al. (2006). Habitat needs to correct fragmentation are similar as those for
diadromous species i.e. longitudinal and lateral migration should be possible within the estuary.
Habitat sensitivity deals also with special criteria such as presence of water vegetation for phytophilic
species (obligate or partial) or sensitivity to dredging. Conservation goals dealing with these criteria
have already been described and concern the presence of upstream undisturbed vegetated habitats and
floodplains.

HARBASINS – Conservation goals 28
4.4. Habitat status
4.4.1. Introduction
The progressive loss of estuarine areas remains a serious threat to the preservation of estuarine
biotopes and the integrity of the estuaries as a whole. Canalisation has resulted in an inhibition of
natural dynamics of sediment transport and water movements (Cattrijsse et al., 2002).
Observed loss of estuarine areas is most pronounced in intertidal areas, and the effects of that loss are
disproportionally high (Elliott & Taylor, 1989a, McLusky et al., 1992). Davidson et al. (1992) referred
to the process of gradual loss of shallow habitats as the estuarine squeeze. In the upper part of the
estuarine squeeze the high water line has shifted downshore through land-claim, building of sea
defences and the construction of docks. The driving forces of the lower part of the estuarine squeeze
are the extraction of sediments and the construction of barrages as well as sea level rise are. The
combined effort of these processes results in the loss of intertidal areas, the relative increase in deeper
subtidal areas and the consequent narrowing of the estuarine channel. Information on 14 European
estuaries showed that land-claim, channel management and barrages/impoundments are the most
important mechanisms resulting in disturbances on the estuarine fish communities ranging from loss
of species diversity over complete change from estuarine to a freshwater assemblage to the
disappearance of whole communities (Cattrijsse et al., 2002). Under natural conditions, erosion and
sedimentation are more or less in balance, and the natural loss of a habitat is likely to be mitigated by
sedimentation at a distant site. Even floods or storm-surges that may destroy complete habitats are
likely to be remedied. There is a natural drift of the main tidal channel, but in the Schelde it has ceased
due to consolidation of its banks. Due to these natural changes in geomorphology fish communities
will slowly change to a species composition that is better adapted to the new situation created by the
geomorphological changes. However, land-claim will decrease the benthic community and, if any of
the resources required by fish (space and food) are limiting, as a result also the fish communities in
terms of species number, abundance and biomass will reduce. Reduction of the food supply and the
loss of habitat reduce the estuaries’ value as nursery area and thus its carrying capacity. McLusky et
al. (1992) indicate the effects of loss of intertidal area on bird and fish populations. A similar effect is
observed with dredging although few reported quantitative effects on fish assemblage are available.
Due to the presence of a port and other artificial structures such as channel stabilisation with dykes an
increased flushing effect is created. This leads to segregation of the tidal currents.In turn an increase in
the ebb-related effects in the main channel and the flood current predominates in the adjacent
channels. The building of docks, wharves and jetties may induce loss of intertidal area or soft sediment
although they may create an artificial hard substratum which attracts a rocky shore community and the
associated fish fauna.

HARBASINS – Conservation goals 29
4.4.2. Habitat loss in the Schelde
Van Braeckel et al. (2006) assessed three human impact types on the River Schelde estuary. The
ecological changes during the past century were analysed using a hierarchical ecotope approach.
Changes in hydrodynamic and morphological parameters were analysed over the past century while an
overview of anthropogenic interventions and natural changes with potential effects on habitat acreage
and quality in the Zeeschelde and its tidal tributaries was given for past two centuries.
For the tidal regime Van Braeckel et al. (2006) analysed characteristics of the ten year mean high
water (MHW), mean low water (MLW), tidal asymmetry and the tidal amplitude. Temporal and
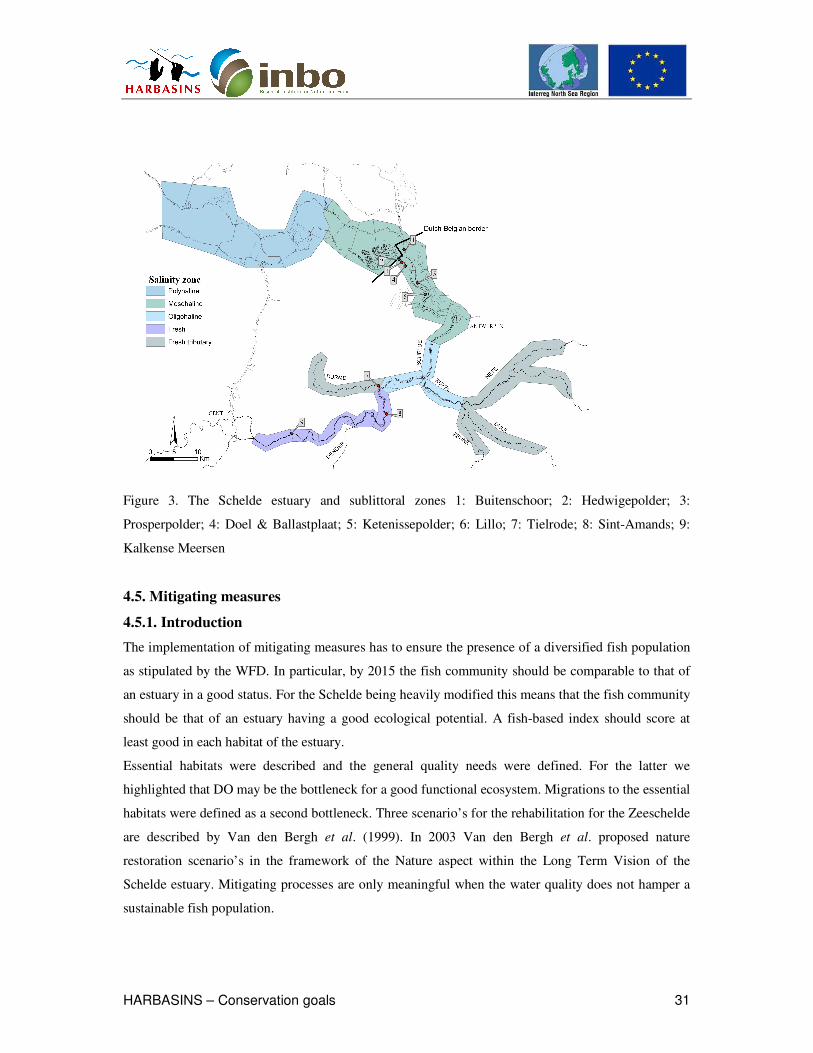
spatial patterns of these parameters were examined within different salinity zones in the Schelde, the
Durme and the Rupel (See Fig. 3). A gradual increase was observed for MHW in the whole Schelde
estuary while the MLW and tidal asymmetry showed a more irregular pattern. These are also more
sensitive to anthropogenic activities such as dredging and deepening in the navigation channel. MHW
and MLW changes resulted in a high increase of tidal amplitude in the Schelde and Rupel. The
observed changes can be explained by an analysis of the morphological evolutions in the estuary. Here
the authors indicated channel straightening, reduced flooding area and upstream shifts of the main
flood zones as the most important changes. Canalisation has a direct impact on the river ecosystem
e.g. by reducing its connectivity with its valley. In the fresh water zone with short retention time the
channel was shortened with 22% (10,5km). Consequently sinuosity declined and the river is now
classified as straight instead of meandering. The accompanying dike normalisations induced a loss of
95% or 826 ha of the former flooding area in this zone. In the tributaries canalisations equally resulted
in a significant reduction of connectivity with the surrounding valley. The Durme shortened with 12%
(2,5km) of its length compared to 1850. The disruption between the Rupel and its tidal brook the Vliet
caused the loss of one third of its tidal area. Fish migration between river and valley was hampered by
loss of natural flooding zones by blocking many tidal brooks. The largest reduction of storage width in
the Zeeschelde occurred in the mesohaline zone by the embankment of some vast polders (Fig. 3
Prosperpolder, Hedwigepolder, Groot Buitenschoor and Ketenisseschor). As mentioned, huge alluvial
and flooding areas such as ‘Kalkense Meersen’ were lost in the fresh water zone with short retention
time. A reduction of storage width was also found near Tielrode and St. Amands. The decline of
flooding area along the Durme from 1955 till 1990 resulted in an overall loss of 87,4% or 730 ha of
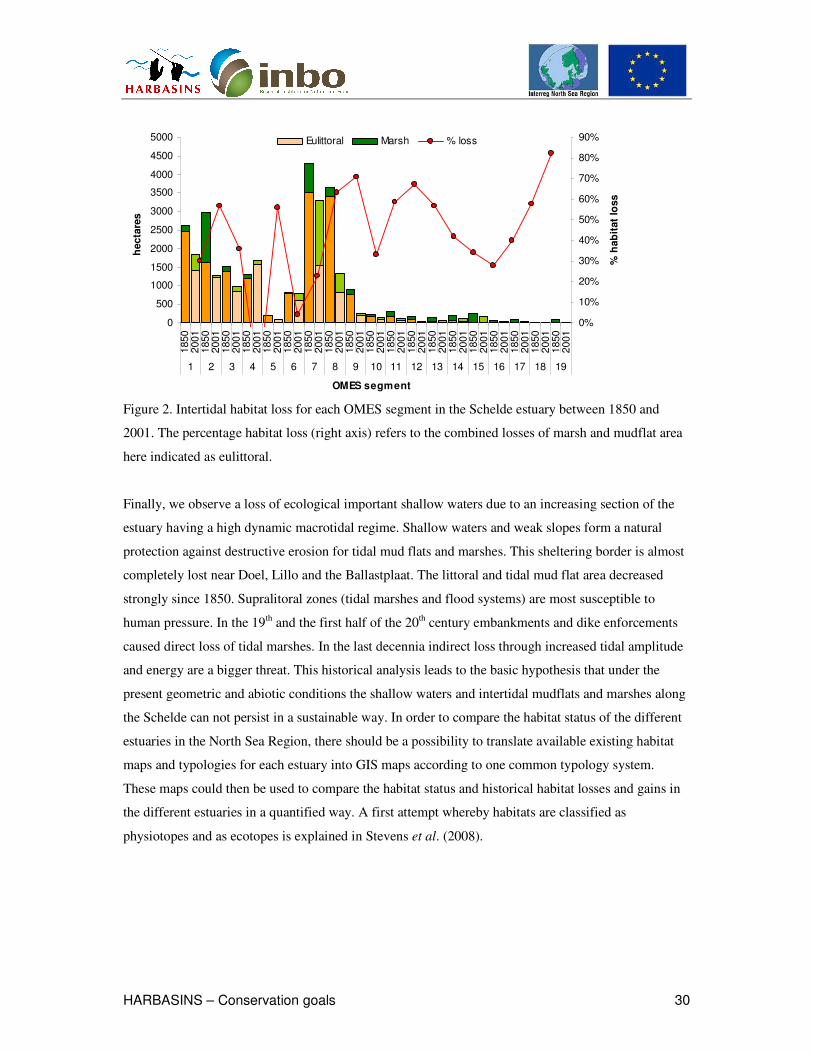
the adjacent flood zones compared to 1850. Figure 2 illustrates for each Omes segment the percentage
intertidal area loss (marches and mudflats) due to land claim for the last two centuries.
HARBASINS – Conservation goals 30
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1850
2001
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
OMES segment
hec
tare
s
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
% h
ab
itat
loss
Eulittoral Marsh % loss
Figure 2. Intertidal habitat loss for each OMES segment in the Schelde estuary between 1850 and
2001. The percentage habitat loss (right axis) refers to the combined losses of marsh and mudflat area
here indicated as eulittoral.
Finally, we observe a loss of ecological important shallow waters due to an increasing section of the
estuary having a high dynamic macrotidal regime. Shallow waters and weak slopes form a natural
protection against destructive erosion for tidal mud flats and marshes. This sheltering border is almost
completely lost near Doel, Lillo and the Ballastplaat. The littoral and tidal mud flat area decreased
strongly since 1850. Supralitoral zones (tidal marshes and flood systems) are most susceptible to
human pressure. In the 19th and the first half of the 20th century embankments and dike enforcements
caused direct loss of tidal marshes. In the last decennia indirect loss through increased tidal amplitude
and energy are a bigger threat. This historical analysis leads to the basic hypothesis that under the
present geometric and abiotic conditions the shallow waters and intertidal mudflats and marshes along
the Schelde can not persist in a sustainable way. In order to compare the habitat status of the different
estuaries in the North Sea Region, there should be a possibility to translate available existing habitat
maps and typologies for each estuary into GIS maps according to one common typology system.
These maps could then be used to compare the habitat status and historical habitat losses and gains in
the different estuaries in a quantified way. A first attempt whereby habitats are classified as
physiotopes and as ecotopes is explained in Stevens et al. (2008).
HARBASINS – Conservation goals 31
Figure 3. The Schelde estuary and sublittoral zones 1: Buitenschoor; 2: Hedwigepolder; 3:
Prosperpolder; 4: Doel & Ballastplaat; 5: Ketenissepolder; 6: Lillo; 7: Tielrode; 8: Sint-Amands; 9:
Kalkense Meersen
4.5. Mitigating measures
4.5.1. Introduction
The implementation of mitigating measures has to ensure the presence of a diversified fish population
as stipulated by the WFD. In particular, by 2015 the fish community should be comparable to that of
an estuary in a good status. For the Schelde being heavily modified this means that the fish community
should be that of an estuary having a good ecological potential. A fish-based index should score at
least good in each habitat of the estuary.
Essential habitats were described and the general quality needs were defined. For the latter we
highlighted that DO may be the bottleneck for a good functional ecosystem. Migrations to the essential
habitats were defined as a second bottleneck. Three scenario’s for the rehabilitation for the Zeeschelde
are described by Van den Bergh et al. (1999). In 2003 Van den Bergh et al. proposed nature
restoration scenario’s in the framework of the Nature aspect within the Long Term Vision of the
Schelde estuary. Mitigating processes are only meaningful when the water quality does not hamper a
sustainable fish population.

HARBASINS – Conservation goals 32
4.5.2. Mitigating processes in the Schelde
To reduce the tidal energy Van den Bergh et al. (2003) suggest to increase the stream profile by
adding shallow intertidal habitats downstream Dendermonde Temse. To avoid strong discharge
flushes retention areas upstream should be created. These restoration practices may result in an
increased DO in the estuary. Intertidal, shallow habitats, inland water treatment and floodplains are
essential to reinstate physical and chemical processes and hence increase the estuary’s self-purification
capacity and filtering capacity that sustains water quality (Van den Bergh et al., 2005, Lotze et al.,
2006). Aeration is the most important oxygen source in an estuary and can be improved by the
creation of tidal areas since their surface-to-volume area is high. Thus, estuarine functions and
carrying capacity are probably enhanced by increasing the surface of estuarine habitat. Adriaensen et
al. (2005) and Couderé et al. (2005) suggest that for the Zeeschelde an additional mudflat area of at
least 500ha will ensure a sustainable fish population. The same authors state that there is a need of
1500ha extra marsh surface to avoid silicon limitations which hamper the diatoms. As a second effect,
the creation of space will reduce the velocity and the associated effects such as erosion, turbidity and
sedimentation. Pas et al. (1998) state that Tielrodebroek, a flood controlling area of about 90ha nearby
the mouth of the River Durme, functions as a spawning and nursery area for certain freshwater
species. The Lippenbroek is a recently created flood control area situated in the freshwater part of the
Schelde-estuary. This pilot project assesses the possibility to combine the concept of a flood control
area (FCA) with nature development and the functions of intertidal areas, by use of simple sluice
constructions to introduce a controlled reduced tide (CRT). The creation of shallow area’s will be
beneficial for fish since these can be used as nursery and spawning places, shelter and resting areas
and feeding grounds. Floodplains have a higher productivity than the main channel and they provide
habitat complexity and habitat quality (Grift, 2001). Grift (2001) observed that fish rapidly colonized
newly created floodplain water bodies in the river Rhine. As such reconnected oxbow lakes have a
beneficial value for the riverine fish community since they provide a habitat that is better suitable for
0-group fish than the main stream. In addition they form important spawning and nursing areas for the
rheophilic species. Restoring interaction between the river and its floodplains is essential.
Physical and chemical barriers preventing this migration should be removed or bypassed. Solving fish
migration problems may not be a formal fish passage, but a natural solution such as restoration of old
meanders or the creation of a nature-like bypass channel. The basic steps to create a fish passages are
explained comprehensively by Kroes et al. (2006).
Upstream river rehabilitation and mitigation measures should improve the lateral connectivity of main
channels to ecotone complexity beneficial for fish (Angermeier & Karr, 1983; Sindilariu et al., 2006).
Turbidity should be reduced by construction of sedimentation areas such as wetlands where the flow is

HARBASINS – Conservation goals 33
reduced and this will also reduce sedimentation on the spawning grounds and increase their
availability. These wetlands increase the retention time in the Schelde.
5. Conclusions
An extensive literature study has allowed the definition of essential habitat requirements for fish
species both at general and guild levels. Some of the described conservation goals are applicable in
different guilds hence the importance of having CG which go across the different guilds. The proposed
conservation goals are qualitative and so further research is needed to provide quantitative
conservation goals. A crucial step constitutes the definition of stressors or impacts for each particular
estuary and the subsequent assessment of these impacts. Impacts comprise land-claim, recreation,
fishery, industrial and harbour activities including navigation and dredging, presence of barriers to
migration, aquaculture, coastal defences and so forth. Possible threats to estuarine habitats are listed
by McLusky & Elliott (2005) or Kennish (2004). Also the DPSIR approach is considered as a useful
tool to classify impacts. A report on impacts is in progress. Impacts were assessed according to Aubry
& Elliott (2006). In addition the carrying capacity of the different habitat types within the different
estuary zones should be calculated. Finally a fish-based evaluation system should be developed to
assess the status of the estuary and to evaluate the impact of the implementation of mitigating
measures.
References
Aarts, B.G.W. & P.H. Nienhuis, 2003. Fish zonations and guilds as the basis for assessment of
ecological integrity of large rivers. Hydrobiologia. 500: 157-178.
Adriaensen, F., Van Damme, S., Van den Bergh, E., Van Hove, D., Brys, R., Cox, T., Jacobs, S.,
Konings, P., Maes, J., Maris, T., Mertens, W., Nachtergale, L., Struyf, E., Van Braeckel, A. & P.
Meire, 2005. Instandhoudingsdoelstellingen Schelde-estuarium, Universiteit Antwerpen, Rapport
Ecobe 05-R82, Antwerpen. 252 pp. + annexes
Angermeier, P.L. & J. Karr, 1983. Relationship between woody debris and fish habitat in a small
warmwater stream. Transactions of the American Fisheries Society. 113: 716-726.
Aprahamian, M.W., 1988. The biology of the twaite shad Alosa fallax fallax (Lacépède) in the Severn
Estuary. Journal of Fish Biology. 33A: 141–152.
Aprahamian, M.W. & C.D. Aprahamian, 2001. The influence of water temperature and flow on year-
class strength of twaite shad (Alosa fallax) from the River Severn, England. Bulletin Français de la
Pêche et de la Pisciculture. 362/363: 953–972

HARBASINS – Conservation goals 34
Aubry, A. & M. Elliott, 2006. The use of environmental integrative indicators to assess seabed
disturbance in estuaries and coasts: application to the Humber estuary, UK. Marine Pollution Bulletin.
53: 175-185.
Balon, E.K., 1975. Reproductive guilds of fishes: a proposal and definition. Journal Fisheries Research
Board Canada. 32: 821-864.
Balon, E.K., 1981. Additions and amendments to the classification of reproductive styles in fishes.
Environ. Biol. Fish. 6: 377-389.
Batzer, D.P., Pusateri, C.R. & R. Vetter, 2000. Impacts of fish predation on marsh invertebrates: direct
and indirect effects. Wetlands 20: 307-312.
Beck, M., Heck JR., Able, K.W., Childers, D.L., Eggleston, D.B., Gillnaders, B.M., Halpern, B.,
Hays, C.G., Hoshino, K., Minello, T.J., Orth, R.J., Sheridan, P.F. & M.P. Weinstein, 2001. The
Identification, Conservation, and Management of Estuarine and Marine Nurseries for Fish and
Invertebrates. Bioscience. 51 (8): 633-641.
Belpaire, C., Smolders, R., Vanden Auweele, I., Ercken, D., Breine, J., Van Thuyne, G. & F. Ollevier,
2000. An Index of Biotic Integrity characterizing fish populations and the ecological quality of
Flandrian water bodies. Hydrobiologia. 434: 17-33.
Berrebi dit Thomas, R., Belliard, J. & P. Boët, 1998. Caractéristiques des peuplements piscicoles
sensibles aux altérations du milieu dans les cours d'eau du bassin de la Seine. Bull. Fr. Pêche
Pisciculture. 348: 47-64.
Blaber, S.J.M. & T.G. Blaber, 1980. Factors affecting the distribution of juvenile estuarine and inshore
fishes. Journal of Fish Biology. 17: 143-162.
Borja, A. & M. Elliott, 2007. What does ‘good ecological potential’ mean, within the European Water
Framework Directive? Marine Pollution Bulletin. 54: 1559-1564.
Borja, A., Franco, J., Valencia, V., Bald, J., Muxika, I., Belzunce, M. J., & O. Soluan, 2004.
Implementation of the European water framework directive from the Basque country (northern Spain):
a methodological approach. Marine Pollution Bulletin. 48: 209–218.
Breine, J.J., Goethals, P., Simoens, I., Ercken, D, Van Liefferinghe, C., Verhaegen, G., Belpaire, C.,
De Pauw, N., Meire, P. & F. Ollevier, 2001. De visindex als instrument voor het meten van de
biotische integriteit van de Vlaamse binnenwateren. Instituut voor Bosbouw en Wildbeheer,
Groenendaal. Eindverslag van project VLINA 9901, studie uitgevoerd voor rekening van de Vlaamse
Gemeenschap binnen het kader van het Vlaams Impulsprogramma Natuurontwikkeling. 173 pp. +
bijlagen.
Breine, J.J., Maes, J., Quataert, P., Van den Bergh, E., Simoens, I., Van Thuyne, G. & C. Belpaire,
2007a. A fish-based assessment tool for the ecological quality of the brackish Schelde estuary in
Flanders (Belgium). Hydrobiologia. 575: 141-159

HARBASINS – Conservation goals 35
Breine, J., Simoens, I., Goethals, P., Quataert, P., Ercken, D., Van Liefferinghe, C. & C. Belpaire,
2004. A fish-based index of biotic integrity for upstream brooks in Flanders (Belgium).
Hydrobiologia. 522: 133-148.
Breine, J., Simoens, I., Haidvogl, G., Melcher, A., Pont, D. & S. Schmutz, 2005. Manual for the
application of the European Fish Index - EFI. A fish-based method to assess the ecological status of
European rivers in support of the Water Framework Directive. Version 1.1, January 2005. 81 pp.
Breine, J., Stevens, M., Simoens, I. & E. Van den Bergh, 2007b. A reference list of fish species for a
heavily modified estuary and its tributaries: the River Scheldt. INBO.R.2008.4. 25 pp.
Bruslé, J. & J-P. Quignard, 2001. Biologie des poisons d’eau douce européens. Technique et
documentation. 625 pp.
Bruton, M.H., 1985. The effects of suspensoids on fish. Hydrobiologia. 152: 221-241.
Capra, H., Breil, P. & Y. Souchon, 1995. A new tool to interpret magnitude and duration of fish
habitat variations. Regulated rivers, Research and Management. 10: 281-289.
Bulger, A.J., Hayden, B.P., Monaco, M.E., Nelson, D.M. & M.G. McCormick-Ray, 1993. Biology-
based estuarine salinity zones derived from a multivariate analysis. Estuaries. 16: 311-322.
Cattrijse, A., Codling, I., Conides, A., Duhamel, S., Gibson, R.N., Hostens, K., Mathieson, S. & D.S.
McLusky, 2002. Estuarine development/habitat restoration and re-creation and their role in estuarine
management for the benefit of aquatic resources. In Elliott, M. & K.L. Hemingway (Ed.), 2002. Fishes
in estuaries, Blackwell Science, London. 636 pp., 266-321.
Cites, 2006. Commission Staff Working Document: Commission Services' assessment of CITES CoP
ENV 721 WTO 274. 13 pp.
Chow, V.T. (1973). Open-channel hydraulics. Civil Engineering Series. Mcgraw-hill: Auckland. ISBN
0-07-Y85906-X. 680 pp.
Coates, S.A., Colclough, S.R., Robson, M. & T.D. Harrison, 2004. Development of an estuarine
classification scheme for the water framework directive. Phase 1&2– Transitional Fish Component.
Environment Agency R&D Technical Report E1–131/TR, ISBN 1 84432 262 9. 42 pp. + appendices.
Costa, M.J., Cabral, H.N., Drake, P., Economou, A.N., Fernandez-Delgado, C., Gordo, L., Marchand,
J. & R. Thiel, 2002. Recruitment and production of commercial species in estuaries. In Elliott, M. &
K. Hemingway (Ed.), 2002. Fishes in estuaries, Blackwell Science, London. 636 pp., 54-123.
Costa, M.J. & M. Elliott, 1991. Fish usage and feeding in two industrialised estuaries- The Tagus,
Portugal, and the Forth, Scotland. In: (Elliott, M. & J.-P. Ducrotoy, Eds.), Estuaries and Coasts:
Spatial and Temporal Intercomparisons. Olsen & Olsen, Fredensborg, Denmark. 289-297.
Copp, G.H., 1989. The habitat diversity and fish reproductive function of floodplain ecosystems.
Environmental Biology of Fishes. 26: 1-27.

HARBASINS – Conservation goals 36
Couderé, K., Vincke, J., Nachtergaele, L., Van den Bergh, E., Dauwe, W., Bulckaen, D. & J.
Gauderis, 2005. Geactualiseerd Sigmaplan voor veiligheid en natuurlijkheid in het bekken van de
Zeeschelde: synthesenota. Waterwegen & Zeekanaal NV: Antwerpen, Belgium. II. 74 pp.
Davidson, N.C., Laffoley, D.d’A., Doody, J.P., Way, J.S., Gordon, J., Key, R., Pienkowsky, M.W.,
Mitchell, R. & K.L. Duff, 1991. Nature Conservation and Estuaries in Great Britain. Joint Nature
Conservation Committee, Peterborough.
Deegan, L. A., Finn, J.T., Ayvazian, S.G., Ryder-Kiefer, C.A.& J. Buonaccorsi, 1997. Development
and validation of an Estuarine Biotic Integrity Index. Estuaries. 20: 601–617.
De Nie, H.W., 1996. Atlas van de Nederlandse zoetwatervissen. Media Pub.- III. 151 pp.
Elliott, M. & F. Dewailly, 1995. The structure and components of European estuarine fish
assemblages. Netherlands Journal of Aquatic Ecology. 29: 397-417.
Elliott, M. & K.L. Hemingway, 2002. In Elliott, M. & K.L. Hemingway (Eds), Fishes in estuaries.
Blackwell Science, Oxford. 577-579.
Elliott, M., Hemingway, K.L., Costello, M.J., Duhamel, S., Hostens, K., Labropoulou, M., Marshall,
S. & H. Winkler, 2002. Links between fish and other trophic levels. In Elliott, M. & K. Hemingway
(Ed.), 2002. Fishes in estuaries, Blackwell Science, London.636 pp. 124-216
Elliott, M., Nedwell, S., Jones, N.V., Read, S.J., Cutts, N.D. & K.L. Hemingway, 1998. Volume II-
Intertidal sand and mudflats & subtidal mobile sandbanks: an overview of dynamic and sensitivity
characteristics for conservation management of marine SACs. Institute of Estuarine & Coastal Studies,
University of Hull (Report for and prepare by Scottish Association for Marine science (SAMS) for the
UK Marine SACs project).
Elliott, M., O’Reilly, M.G. & C.J.L. Taylor, 1990. The Forth estuary: a nursery and overwintering area
for the North Sea fishes. Hydrobiologia. 195: 89-103.
Elliott, M. & C.J.L. Taylor, 1989a. The structure and functioning of an estuarine/marine fish
community in the Forth estuary, Scotland. In Proceedings of the 21st European Marine Biology
Symposium Gdánsk, 14-19 September 1986. Polish Academy of Sciences, Institute of Oceanology,
Warsaw. 227-240.
Elliott, M. & C.J.L. Taylor, 1989b. The production ecology of the subtidal benthos of the Forth
estuary, Scotland. Scientia Marina. 53: 531-541.
Elliott, M., Whitfield, A.K., Potter, I.C., Blaber, S.J.M., Cyrus, D.P., Nordlie, F.G. & T.D. Harrison,
2007. The guild approach to categorizing estuarine fish assemblages: a global review. Fish and
fisheries. 8: 241-268.
Erichsen Jones, J.R., 1964. Fish and river pollution. London, Butterworths. 203 pp.

HARBASINS – Conservation goals 37
EU Water Framework Directive, 2000. Directive of the European parliament and of the council
2000/60/EC establishing a framework for community action in the field of water policy. Official
Journal of the European Communities 22.12.2000 L 327/1.
Fairbridge, R. 1980. The estuary, its definition and geodynamic cycle. In Olaussen, E. & I. Cato (Eds).
Chemistry and biochemistry of estuaries. Wiley, NY. 1-35.
Gibson, R.N., Yin, M.C. & L. Robb, 1995. The behavioural basis of predator-prey size relationships
between shrimp (Crangon crangon) and juvenile plaice (Pleuronectus platessa). Journal of the Marine
Biological Association of the UK. 75: 337-349.
Gerking, S.D., 1994. Feeding ecology of fish. Academic Press. ISBN 0-12-280780-4. 461 pp.
Gerstmeier, R. & T. Romig, 1998. Zoetwatervissen van Europa. Troin. 368 pp.
Goffaux, D., Roset, N., Breine, J.J., de Leeuw, J., Oberdorff, T. & P. Kestemont, 2001. A biotic index
of fish integrity (IBIP) to evaluate the ecological quality of lotic ecosystems - Application to the
Meuse River Basin. Final report: Life97 / ENV / B / 000419. 171 pp.
Grandmottet, J.P., 1983. Principales exigences des téléostéens dulcicoles vis à vis de l'habitat
aquatique. Annales Scientifiques de l'Université de Franche-Comté. 4: 3-32.
Grenouillet, G., Pont, D. & J.M. Olivier, 2000. Habitat occupancy patterns of juvenile fishes in a large
lowland river: interaction with macrophytes. Arch. Hydrobiol. 149: 307-326.
Grift, R. 2001. How fish benefit from floodplain restoration along the lower river Rhine. PhD thesis
Wageningen. 205 pp.
Harrison, T.D., Cooper, J.A. & A.E.L. Ramm, 2000. State of South African estuaries.
Geomorphology, ichthyofauna, water quality and aesthetics. State of the Environment Series Report
No. 2. Department of Environmental Affairs and Tourism, Pretoria. 127 pp.
Harrison, T.D. & A.K. Whitfield, 2004. A multimetric index to assess the environmental condition of
estuaries. Journal of Fish Biology. 65: 683–710.
Hoffmann, M. & P. Meire, 1979. De oevers langs de Zeeschelde: inventarisatie van de huidige
oeverstructuren. Water. 95: 131-137.
Horrevoets, A.C., Savenije, H.G., Schuurman, J.N. & S. Graas, 2004. The influence of river discharge
on tidal damping in alluvial estuaries. Journal of hydrology. 294 (4): 213-228.
Huet, M., 1949. Appréciation de la valeur piscicole des eaux douces. Publication du Ministère de
l'Agriculture, Administration des Eaux et Forêts, Groenendaal. 55 pp.
Hughes, J.E., Deegan, L.A., Weaver, M.J. & J. E. Costa, 2002. Regional application of an index of
estuarine biotic integrity based on fish communities. Estuaries. 25: 250–263.
Hughes, R.M. & T. Oberdorff, 1999. Applications of IBI Concepts and Metrics to Water Outside the
United States and Canada. In Simon, T. P. (ed.), Assessing the Sustainability and Biological Integrity
of Water Resources Using Fish Communities. CRC Press LLC, Washington DC. 62–74.

HARBASINS – Conservation goals 38
ICES (2006). Report of the 2006 session of the joint EIFAC/ICES Working Group on Eels, 3. ICES
CM 2006/ACFM:16. EIFAC Occasional Paper No. 38: 352 pp.
Jager, Z. & J. Kranenbarg, 2004. Referenties en maatlatten: Achtergronddocument vissen. OVB,
Nieuwegein. 144 pp.
Junk, W.J., Bayley, P.B. & R.E. Sparks, 1989. The flood pulse concept in river-floodplain systems. In
Dodge D.E. (Ed) Proceedings of the international large river symposium. Can. Spec. Publ. Fish.
Aquat. Sci. 106: 110-127.
Kennish, M.J., 2004. In Kennish, M. J. (editor). 2004. Estuarine Research, Monitoring, and Resource
Protection. CRC Press, Boca Raton, Florida. 297 pp.
Kroes, M.J., Gough, P., Schollema, P.P. & H. Wanningen, 2006. From sea to source; Practical
guidance for restoration of fish migration in European rivers. 119 pp.
Kroes, J. G. & S. Monden, (Ed.) 2005. Vismigratie: een handboek voor herstel in Vlaanderen en
Nederland. Aminal, Afdeling Water: Brussel. 207 pp.
Kruk, A. 2007. Role of habitat degradation in determining fish distribution and abundance along the
lowland Warta River, Poland. J. Appl. Ichthyol. 23: 9-18.
Léger, L., 1945. Economie biologique et productivité de nos rivières à cyprinides. Bull. français de
Pisciculture. 139: 49-69.
Limburg, K., 1996. Modelling the ecological constraints on growth and movement of juvenile
American shad (Alosa sapidissima) in the Hudson river estuary. Estuaries. 19, 794–813.
Lobb, D.M. & D.L., Orth, 1991. Habitat use by an assemblage of fish in a large warmwater stream.
Transactions of the American Fisheries Society. 120: 65- 78.
Lockwood, S., 1972. An ecological survey of an O-Group plaice (Pleuronectus platessa, L.)
population, Filey Bay, Yorkshire. PhD Thesis, University of East Anglia, UK, unpublished.
Lotze, H.K., Lenihan, H.S., Bourque, B.J., Bradbury, R.H., Cooke, R.G., Kay, M.C., Kidwell, S.M.,
Kirby, M.X., Petersons, C.H. & J.B.C. Jackson, 2006. Depletion, degradation, and recovery potential
of estuaries and coastal seas. Science. 312: 1806-1809.
Lucas, M.C. & E. Baras, 2001. Migration of freshwater fishes. Blackwell Science Ltd, Oxford UK.
420 pp.
Maes, J., 2000. The structure of the fish community of the Zeeschelde estuary. Laboratory of Aquatic
Ecology, Catholic University Leuven, PhD thesis. 143 pp.
Maes, J., Geysen, B., Stevens, M., Ollevier, F., Breine, J. & C. Belpaire, 2005. Opvolging van het
visbestand van de Zeeschelde: resultaten voor 2004. IBW.Wb.V.R.2005.151. 40 pp.
Maes, J., Stevens, M. & J. Breine 2007a. Poor water quality constrains the distribution and movements
of twaite shad Alosa fallax fallax (Lacépède, 1803) in the watershed of river Scheldt. Hydrobiologia

HARBASINS – Conservation goals 39
Maes, J., Stevens, M. & J. Breine 2007b. Modelling the migration opportunities of diadromous fish
species along a gradient of dissolved oxygen concentration in a European tidal watershed. Estuarine
Coastal and Shelf Science. 75: 151-162.
Maitland, P.S., 2000. Guide to freshwater fish of Britain and Europe. Hamlyn, London 256 pp.
Maitland, P.S. & T.W. Hatton-Ellis, 2003. Ecology of the Allis and Twaite Shad. Conserving Natura
2000 Rivers Ecology Series No. 3. English Nature, Peterborough.
Mann, R.H.K., 1996. Environmental requirements of European non-salmonid fish in rivers.
Hydrobiologia. 323: 223-235.
Mathieson, S., Cattrijsse, A., Costa, M.J., Drake, P., Elliott, M., Gardner, J. & J. Marchand, 2000. Fish
assemblages of European tidal marshes: a comparison based on species, families and functional guilds.
Marine Ecology Progress Series. 204: 225-242.
McLusky, D.S., Bryant, D.M. & M. Elliott, 1992. The impact of land-claim on the invertebrates, fish
and birds of the Forth estuary. Aquatic conservation: marine and Freshwater Progress Series. 11: 173-
179.
McLusky, D.S. & M. Elliott, 2005. The estuarine ecosystem; ecology threats and Management.
Oxford University press NY. 214 pp.
Milner, N., 1984. Biological 4: Fish. In Lewis G. & G. Williams. Rivers and wildlife handbook. A
guide to practises witch further the conservation of wildlife on rivers. RSPB, London. 51-55.
Moy, Y.C., 2004. Development and evaluation of an estuarine biotic integrity index for South
Carolina tidal creeks. Thesis submitted and approved to obtain the Master of Science degree in marine
biology at the Graduate School, college of Charleston. 321 pp.
Muus, B.J., Nielsen, J.G., Dahlstrøm, P. & B.O. Nyström, 1999. Zeevissen van Noord- en West-
Europa. Schuyt & Co. 338 pp.
Nijsen, H. & S.J De Groot, 1987. De vissen van Nederland Kon. Ned. Nathist. Ver. III, 224 pp.
OVB, 1988. Cursus vissoorten. Organisatie ter verbetering van de binnenvisserij (OVB). Nieuwegein
(Nl). 914 pp.
OVB, 1997. OVB Berichten, De visstand in diepe wateren, III. 86-104.
Pas, J., Peeters, B., Maes, J., Vlietinck, K., Pauwels, F. & F. Ollevier, 1998. Opvolging van het
visbestand in de Zeeschelde en de bijhorende overstromingsgebieden. Studierapport in opdracht van
AMINAL. 94 pp. + annexes.
Pauly, D., 1982. On the interrelationships between natural mortality, growth parameters, and mean
environmental temperature in 175 fish stocks. Journal du Conseil international d’Exploration de la
Mer. 39: 175–192.

HARBASINS – Conservation goals 40
Pihl, L., Cattrijse, A., Codling, I., Mathieson, S., McLusky, D.S. & C. Roberts, 2002. Habitat use by
fishes in estuaries and other brackish areas. In Fishes in Estuaries. Elliott, M. and Hemmingway, K.L.
(Eds). Blackwell Science, Oxford. 10-53.
Phillipart, J.C. & M. Vranken, 1983a. Animaux menacés en Wallonie protégeons nos poissons,
Gembloux. 206 pp.
Phillipart, J.C. & M. Vranken, 1983b. Atlas des poissons de Wallonie. Distribution, écologie,
éthologie, pêche, conservation. Cagiers d'Ethologie Appliquée. Vol.3 Suppl. 1-2. Université de Liège.
395 pp.
Pollux, B.J.A., Korsi, A., Verberk, W.C.E.P., Pollux, P.M.J. & G. van der Velde, 2006. Reproduction,
growth, and migration of fishes in a regulated lowland tributary: potential recruitment to the river
Meuse. Hydrobiologia. 565: 105-120.
Quak, J., 1994. De visstand in stromende wateren. In vismigratie, visgeleiding en vispassages in
Nederland (Ed. J.P. Raat), O.V.B., Nieuwegein, Nederland, 59-84.
Reincke, H., Lischke, P. & J. Schindler, 1992. Vorstellung der Ergebnisse der Arbeitsgruppe ‛Mess-
und Untersuchungsprogramme’ der Internationalen Kommission zum Schutz der Elbe. Proceedings of
the 4th Magdeburger Gewässerschutzseminars zur Situation der Elbe, 275-282.
Reitsma, J.M., 1992. Habitat- en corridorfunctie van oevers voor fauna. Zoogdieren, vogels,
amfibieën, vissen, insekten. Rijkswaterstaat, Dienst Weg- en Waterbouwkunde Delft. 17 pp.
Riemersma, P., 2000. Vismigratie in Noord-Holland. Technische nota. Organisatie ter verbetering van
de Binnenvisserij, Nieuwegein. 106 pp. + bijlagen.
Rijnsdorp, A.D., Van Beek, F.A., Flatman, S., Millner, R.M., Riley, J.D., Giret, M. & R. De Clerck,
R., 1992. Recruitment of sole stocks, Solea solea L. in the Northeast Atlantic. Netherlands
Journal of Sea Research. 29: 173-192.
Roule, L., 1927. Les poisons et le monde vivant des eaux. Tome II. La vie et l’action. Librairie
Delagrave, Paris. 327 pp.
Root, R.B., 1967. The niche exploitation pattern of the blue-gray gnatcacher. Ecol. Monogr. 37: 317-
350.
Robinet, T. & E. Feunteun, 2002. Sublethal effects of exposure to chemical compounds: A cause for
the decline in atlantic eels? Ecotoxicology. 11: 265-277.
Salas, F., Nete, J.M., Borja, A. & J.C. Marques, 2004. Evaluation of the applicability of a marine
biotic index to characterize the status of estuarine ecosystems: the case of Mondego estuary (Portugal).
Ecological Indicators. 4: 215–225.
Schiemer, F. & H. Waidbacher, 1999. ‘Strategies for conservation of the Danubian fish fauna’, in
Boon, P.J., Calow, P. and Petts, G.E. (Eds), River Conservation and Management, John Wiley,
Chichester. 364–384.

HARBASINS – Conservation goals 41
Scholten, M., Wirtz, C. & R. Thiel, 2003. The modular habitat model (MHM) for the ide, Leuciscus
idus (L.)- a new method to predict the suitability of inshore habitats for fish. J. Appl. Ichthyol. 19:
315-329.
Schouten, W.J. & J. Quak, 1994. De Visstand in de Stromende Rijkswateren. RIZA/OVB VO 1993-
01. Organisation for the Improvement of Inland Fisheries, Nieuwegein/Institute for Inland Water
Management and Waste Water Treatment, Lelystad. 123 pp. + annexes
Scholz, U., 1986. Sauerstoffmangelresistenz von Küstenfischen unter dem Einfluss von
Parasitenbefall. Diplomarbeit, Universität, Kiel.
Sindilariu, P., Freyhof, J. & C. Wolter, 2006. Habitat use of juvenile fish in the lower Danube and the
Danube Delta: implications for ecotone connectivity. Hydrobiologia. 571: 51-61
Stevens, M., Van den Bergh, E., Simoens, I. & J. Breine, 2008. A physiotope system for classification
of estuarine habitats. 14 pp.
Thiel, R. & I.C. Potter, 2001. The ichthyofaunal composition of the Elbe estuary: an analysis in space
and time. Mar. Biol. 138: 603-616.
Turnpenny, A.W.H., Clough, S.C., Holden, S.D.J., Bridges, M., Bird, H. O'Keeffe, N.J., Johnson, D.,
Edmonds, M. & C. Hinks, 2004. Thames Tideway Strategy: Experimental studies on the dissolved
oxygen requirements on fish. Babtie Aquatic Report, Thames Water Utilities. 137 pp.
Turnpenny, A., Coughlan, J. & K. Liney, 2006. Review of temperature and dissolved oxygen effects
on fish in transitional waters. Jacobs Babtie. 81 pp.
Van Aelbroeck, F., 1910. Poissons des eaux douces et saumâtres dans leur habitat. Imp. Scient.
Charles Bulens (Ed.), Brux. 166 pp.
Van Braeckel, A., Piesschaert, F. & E. Van den Bergh, 2006. Historical analysis of the Zeeschelde and
tidal tributaries. 19th century till present. (in Dutch) INBO.R.2006.29. 185 pp.
Vandelannoote, A., Yseboodt, R., Bruylants, B., Verheyen, R., Belpaire, C., Van Thuyne, G.,
Denayer, B., Beyens, J., De Charleroy, D., Coeck, J., Maes, J. & P. Vandenabeele, 1998. Atlas van de
Vlaamse beek- en riviervissen. Water Energik Vlario, Wijnegem. 303 pp.
Vanden Auweele, I., 1995, Ontwikkeling van een visindex als biologische evaluatiemethode voor de
kwaliteit van de Vlaamse waterlopen en getoetst aan het Dijle- en Demerbekken. Thesis. 85 pp. +
bijlagen
Van den Bergh, E., Meire, P., Hoffman, M. & T. Ysebaert, 1999. Natuurherstelplan Zeeschelde: drie
mogelijke inrichtingsvarianten. Rapport Instituut voor Natuurbehoud 99/18, Brussel. 164 pp. +
bijlagen.
Van den Bergh, E., Van Damme, S., Graveland, J., de Jong, D.J., Baten, I. & P. Meire, 2003.
Studierapport natuurontwikkelingsmaatregelen ten behoeve van de Ontwikkelingsschets 2010 voor het
Schelde-estuarium. Werkdocument/RIKZ/OS/2003.825x. 93 pp.

HARBASINS – Conservation goals 42
Van den Bergh, E., van Damme, S., Graveland, J., de Jong, D., Baten, I. & P. Meire, 2005. Ecological
rehabilitation of the Schelde estuary (The Netherlands-Belgium; northwest Europe): linking ecology,
safety against floods, and accessibility for port development. In: Restoration ecology. 13(1): 204-214.
van Emmerik, W.A.M., 2003. Indeling van de vissoorten van de Nederlandse binnenwateren in
ecologische gilden en in hoofdgroepen. Organisatie ter Verbetering van de Binnenvisserij,
Nieuwegein. OVB Onderzoeksrapport 00160. 73 pp. + 2 bijlagen
Vanote, R.L., Minshall, G.W., Cummins, K.W., Sedell, J.R., & C.E. Cushing, 1980. The river
continuum concept. Can. J. Fish. Aquat. Sci. 37: 130-137.
Venice system, 1959. The final resolution of the symposium on the classification of brackish waters.
Archo Oceanogr. Limnol. 11 (suppl): 243–248.
Verneaux, J., 1981. Les poissons et la qualité des cours d'eau. Annales Scientifiques de l'Université de
Franche-Comté. 2: 33-41.
Ward, J.V., 1989. Riverine-wetland interactions. Freshwater Wetlands and Wildlife. In: DOE
Symposium Series. R.R. Sharitz and J.W. Gibbons (Eds). Oak Ridge, TN. 385-400.
Ward, J.V., Tockner, K. & F. Schiemer, 1999. Biodiversity of floodplain river ecosystems: ecotones
and connectivity. Regulated Rivers Research and Management. 15: 125-139.
Whitfield, A.K. & M. Elliott, 2002. Fishes as indicators of environmental and ecological changes
within Estuaries – a review of progress and some suggestions for the future. Journal of Fish Biology.
61: 229–250.
Williot, P., 1991. Biologie et productions de l'esturgeon. 1ère éd. CEMAGREF-DICOVA. 51 pp.
Wolfstein, K. & E. Van den Bergh, 2004. Conceptueel beschrijving van een onderzoeksvoorstel in het
kader “Lange Termijn Visie, Onderzoek & Monitoring Schelde-estuarium, onderdeel natuurlijkheid”
Wootton, R.J., 1990. Ecology of teleost fishes. Fish and fisheries series 1., Chapman & Hall, London.
404 pp.
Wootton, R.J., 1992. Fish ecology. Blackie & Son Ltd., Glasgow. 212 pp.
Zander, C.D., Nieder, J. & K. Martin, 1999. Vertical distribution patters. In Horn, M.H., Martin,
K.L.M. & M.A. Chotkowski (Ed) Intertidal Fishes Life in two worlds. Academic press London. 399
pp.
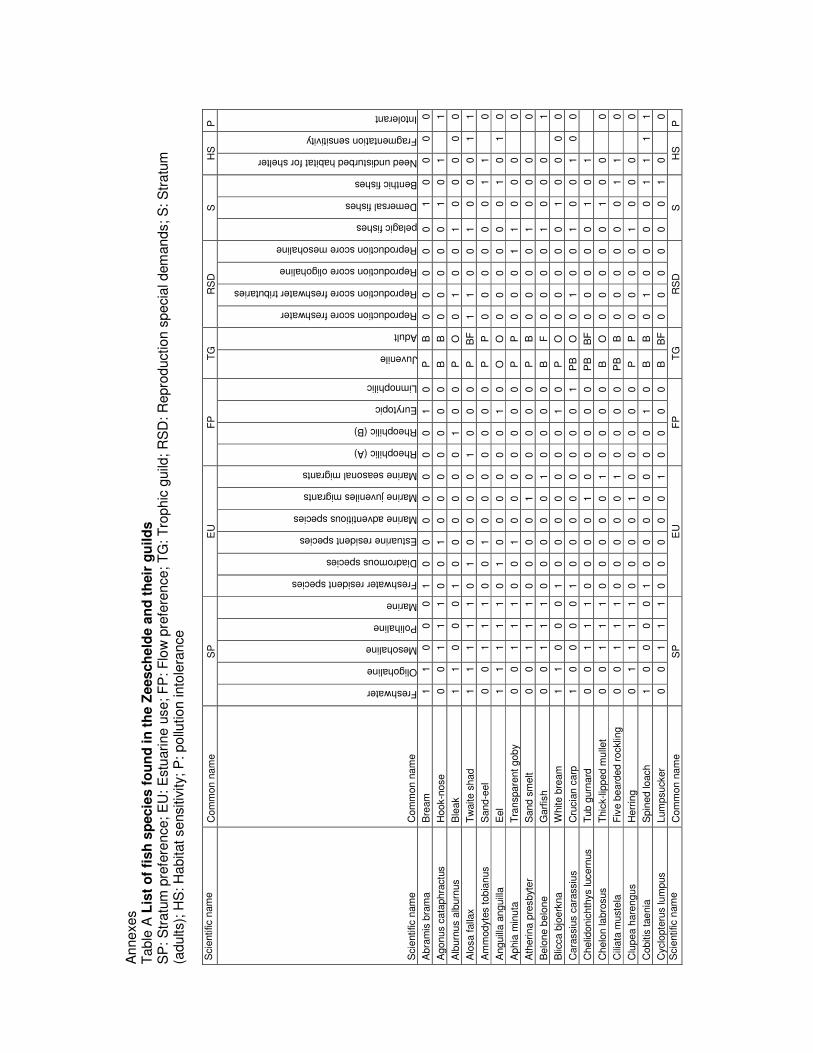
Annexes
Table
A L
ist
of
fish
sp
ecie
s f
ou
nd
in
th
e Z
eesch
eld
e a
nd
th
eir
gu
ild
s
SP
: S
tratu
m p
refe
rence; E
U:
Estu
arine u
se;
FP
: F
low
pre
fere
nce; T
G:
Tro
phic
guild
; R
SD
: R
epro
duction s
pecia
l dem
ands; S
: S
tratu
m
(adults);
HS
: H
abitat sensitiv
ity;
P:
pollu
tion into
lera
nce
Scie
ntific n
am
e
Com
mon n
am
e
SP
E
U
FP
T
G
RS
D
S
HS
P
Scie
ntific n
am
e
Com
mon n
am
e
Freshwater
Oligohaline
Mesohaline
Polihaline
Marine
Freshwater resident species
Diadromous species
Estuarine resident species
Marine adventitious species
Marine juveniles migrants
Marine seasonal migrants
Rheophilic (A)
Rheophilic (B)
Eurytopic
Limnophilic
Juvenile
Adult
Reproduction score freshwater
Reproduction score freshwater tributaries
Reproduction score oligohaline
Reproduction score mesohaline
pelagic fishes
Demersal fishes
Benthic fishes
Need undisturbed habitat for shelter
Fragmentation sensitivity
Intolerant
Abra
mis
bra
ma
Bre
am
1
1
0
0
0
1
0
0
0
0
0
0
0
1
0
P
B
0
0
0
0
0
1
0
0
0
0
Agonus c
ata
phra
ctu
s
Hook-n
ose
0
0
1
1
1
0
0
1
0
0
0
0
0
0
0
B
B
0
0
0
0
0
1
0
1
1
Alb
urn
us a
lburn
us
Ble
ak
1
1
0
0
0
1
0
0
0
0
0
0
1
0
0
P
O
0
1
0
0
1
0
0
0
0
0
Alo
sa f
alla
x
Tw
aite s
had
1
1
1
1
1
0
1
0
0
0
0
1
0
0
0
P
BF
1
1
0
0
1
0
0
0
1
1
Am
modyte
s tobia
nus
Sand-e
el
0
0
1
1
1
0
0
1
0
0
0
0
0
0
0
P
P
0
0
0
0
0
0
1
1
0
Anguill
a a
nguill
a
Eel
1
1
1
1
1
0
1
0
0
0
0
0
0
1
0
O
O
0
0
0
0
0
0
1
0
1
0
Aphia
min
uta
T
ranspare
nt goby
0
0
1
1
1
0
0
1
0
0
0
0
0
0
0
P
P
0
0
0
1
1
0
0
0
0
Ath
erina p
resbyt
er
Sand s
melt
0
0
1
1
1
0
0
0
0
1
0
0
0
0
0
P
B
0
0
0
0
1
0
0
0
0
Belo
ne b
elo
ne
Garf
ish
0
0
1
1
1
0
0
0
0
0
1
0
0
0
0
B
F
0
0
0
0
1
0
0
0
1
Blic
ca b
joerk
na
White b
ream
1
1
0
0
0
1
0
0
0
0
0
0
0
1
0
P
O
0
0
0
0
0
1
0
0
0
0
Cara
ssiu
s c
ara
ssiu
s
Cru
cia
n c
arp
1
0
0
0
0
1
0
0
0
0
0
0
0
0
1
PB
O
0
1
0
0
1
0
0
1
0
0
Chelid
onic
hth
ys lucern
us
Tub g
urn
ard
0
0
1
1
1
0
0
0
0
1
0
0
0
0
0
PB
B
F
0
0
0
0
0
1
0
1
Chelo
n labro
sus
Thic
k-lip
ped m
ulle
t 0
0
1
1
1
0
0
0
0
0
1
0
0
0
0
B
O
0
0
0
0
0
1
0
0
0
Cili
ata
muste
la
Fiv
e b
eard
ed r
ocklin
g
0
0
1
1
1
0
0
0
0
0
1
0
0
0
0
PB
B
0
0
0
0
0
0
1
1
0
Clu
pea h
are
ngus
Herr
ing
0
1
1
1
1
0
0
0
0
1
0
0
0
0
0
P
P
0
0
0
0
1
0
0
0
0
Cobitis
taenia
S
pin
ed loach
1
0
0
0
0
1
0
0
0
0
0
0
0
1
0
B
B
0
1
0
0
0
0
1
1
1
1
Cyclo
pte
rus lum
pus
Lum
psucker
0
0
1
1
1
0
0
0
0
0
1
0
0
0
0
B
BF
0
0
0
0
0
0
1
0
0
Scie
ntific n
am
e
Com
mon n
am
e
SP
E
U
FP
T
G
RS
D
S
HS
P

HA
RB
AS
INS
– C
onserv
ation g
oals
44
Scie
ntific n
am
e
Com
mon n
am
e
Freshwater
Oligohaline
Mesohaline
Polihaline
Marine
Freshwater resident species
Diadromous species
Estuarine resident species
Marine adventitious species
Marine juveniles migrants
Marine seasonal migrants
Rheophilic (A)
Rheophilic (B)
Eurytopic
Limnophilic
Juvenile
Adult
Reproduction score freshwater
Reproduction score freshwater tributaries
Reproduction score oligohaline
Reproduction score mesohaline
pelagic fishes
Demersal fishes
Benthic fishes
Need undisturbed habitat for shelter
Fragmentation sensitivity
Intolerant
Dic
entr
arc
hus labra
x
Sea B
ass
0
1
1
1
1
0
0
0
0
1
0
0
0
0
0
PB
B
F
0
0
0
0
0
1
0
0
0
Engra
ulis
encra
sic
olu
s
Anchovy
0
0
1
1
1
0
0
0
0
0
1
0
0
0
0
PB
P
B
0
0
0
0
0
1
0
0
0
Esox luciu
s
Pik
e
1
1
0
0
0
1
0
0
0
0
0
0
0
1
0
PB
V
F
1
1
0
0
0
1
0
1
1
1
Gadus m
orh
ua
Cod
0
0
1
1
1
0
0
0
0
1
0
0
0
0
0
PB
O
0
0
0
0
0
1
0
0
1
Gaste
roste
us a
cule
atu
s
Thre
e-s
pin
ed s
tickle
back
1
1
1
1
1
1
1
0
0
0
0
0
0
1
0
PB
B
1
1
0
0
1
0
0
1
1
0
Gobio
gobio
G
udgeon
1
0
0
0
0
1
0
0
0
0
0
0
1
0
0
B
B
0
0
0
0
0
0
1
0
0
0
Gym
nocephalu
s c
ern
uus
Ruff
e
1
1
1
1
0
1
0
0
0
0
0
0
0
1
0
PB
B
0
0
0
0
0
0
1
1
0
0
Lam
petr
a f
luvia
tilis
R
iver
lam
pre
y
1
1
1
1
1
0
1
0
0
0
0
1
0
0
0
B
F
1
1
0
0
0
0
1
1
1
1
Leucis
cus c
ephalu
s
Euro
pean c
hub
1
1
0
0
0
1
0
0
0
0
0
0
1
0
0
PB
O
0
1
0
0
1
0
0
0
1
0
Leucis
cus idus
Ide
1
1
0
0
0
1
0
0
0
0
0
0
1
0
0
PB
B
F
0
1
0
0
1
0
0
0
1
1
Leucis
cus leucis
cus
Dace
1
1
0
0
0
1
0
0
0
0
0
1
0
0
0
B
B
1
1
0
0
1
0
0
0
1
1
Lim
anda lim
anda
Dab
0
0
1
1
1
0
0
0
0
1
0
0
0
0
0
B
BF
0
0
0
0
0
0
1
1
1
Lip
aris lip
aris
Sea s
nail
0
0
1
1
1
0
0
1
0
0
0
0
0
0
0
B
B
0
0
0
0
0
0
1
1
1
Liz
a r
am
ado
Thin
lip m
ulle
t 1
1
1
1
1
0
1
0
0
0
0
0
1
0
0
PP
B
O
0
0
0
0
1
0
0
0
1
1
Lota
lota
B
urb
ot
1
1
1
0
0
1
0
0
0
0
0
0
1
0
0
B
F
0
1
0
0
0
0
1
0
1
1
Merlangiu
s m
erlangus
Whitin
g
0
0
1
1
1
0
0
0
0
1
0
0
0
0
0
B
BF
0
0
0
0
0
1
0
1
0
Mis
gurn
us f
ossili
s
Wheath
erf
ish
1
1
0
0
0
1
0
0
0
0
0
0
0
0
1
B
B
0
1
0
0
0
0
1
1
0
0
Myoxocephalu
s s
corp
ius
Bull
rout
0
1
1
1
1
0
0
1
0
0
0
0
0
0
0
B
BF
0
0
0
0
0
0
1
0
0
Osm
eru
s e
perlanus
Sm
elt
1
1
1
1
1
0
1
0
0
0
0
0
1
0
0
B
BF
1
1
0
0
1
0
0
0
1
1
Perc
a flu
via
tilis
P
erc
h
1
1
1
0
0
1
0
0
0
0
0
0
0
1
0
B
BF
0
0
0
0
1
0
0
0
0
0
Petr
om
yzo
n m
arinus
Sea lam
pre
y
1
1
1
1
1
0
1
0
0
0
0
1
0
0
0
B
F
1
1
1
1
0
1
0
1
1
1
Pholis
gunnellu
s
Butt
erf
ish
0
0
1
1
1
0
0
1
0
0
0
0
0
0
0
PB
B
0
0
0
0
0
0
1
1
Scie
ntific n
am
e
Com
mon n
am
e
SP
E
U
FP
T
G
RS
D
S
HS
P

HA
RB
AS
INS
– C
onserv
ation g
oals
45
Scie
ntific n
am
e
Com
mon n
am
e
Freshwater
Oligohaline
Mesohaline
Polihaline
Marine
Freshwater resident species
Diadromous species
Estuarine resident species
Marine adventitious species
Marine juveniles migrants
Marine seasonal migrants
Rheophilic (A)
Rheophilic (B)
Eurytopic
Limnophilic
Juvenile
Adult
Reproduction score freshwater
Reproduction score freshwater tributaries
Reproduction score oligohaline
Reproduction score mesohaline
pelagic fishes
Demersal fishes
Benthic fishes
Need undisturbed habitat for shelter
Fragmentation sensitivity
Intolerant
Pla
tichth
ys fle
sus
Flo
under
1
1
1
1
1
0
0
1
0
0
0
0
0
1
0
PB
B
F
0
0
0
0
0
0
1
1
1
0
Ple
uro
necte
s p
late
ssa
Pla
ice
0
0
1
1
1
0
0
0
0
1
0
0
0
0
0
B
B
0
0
0
0
0
0
1
1
Pom
ato
schis
tus lozanoi
Lozano's
goby
0
0
1
1
1
0
0
0
1
0
0
0
0
0
0
PB
B
0
0
0
0
0
0
1
0
Pom
ato
schis
tus m
icro
ps
Com
mon g
oby
0
1
1
1
1
0
0
1
0
0
0
0
1
0
0
PB
B
0
0
1
1
0
0
1
1
Pom
ato
schis
tus m
inutu
s
Sand g
oby
0
1
1
1
1
0
0
1
0
0
0
0
1
0
0
PB
B
0
0
1
1
0
0
1
1
Psett
a m
axim
a
Turb
ot
0
0
1
1
1
0
0
0
0
1
0
0
0
0
0
PB
F
0
0
0
0
0
0
1
1
Pungitiu
s p
ungitiu
s
Nin
e-s
pin
ed s
tickle
back
1
1
1
0
0
1
0
0
0
0
0
0
0
1
0
PB
B
1
1
0
0
0
1
0
1
1
0
Rhodeus s
ericeus
Bitte
rlin
g
1
1
0
0
0
1
0
0
0
0
0
0
0
0
1
PP
B
P
0
1
0
0
0
0
1
1
1
0
Rutilu
s r
utilu
s
Roach
1
1
1
0
0
1
0
0
0
0
0
0
0
1
0
O
O
0
0
0
0
1
0
0
0
0
0
Salm
o tru
tta
Bro
wn tro
ut
1
1
1
1
1
0
1
0
0
0
0
1
0
0
0
B
BF
0
1
0
0
1
0
0
0
1
1
Sander
lucio
perc
a
Zander
1
1
1
0
0
1
0
0
0
0
0
0
0
1
0
PB
F
0
0
0
0
0
1
0
0
0
0
Scard
iniu
s e
ryth
rophth
alm
us
Rudd
1
1
0
0
0
1
0
0
0
0
0
0
0
0
1
O
O
0
0
0
0
1
0
0
1
0
0
Scophth
alm
us r
hom
bus
Brill
0
0
1
1
1
0
0
0
1
0
0
0
0
0
0
PB
B
F
0
0
0
0
0
0
1
1
Silu
rus g
lanis
W
els
catfis
h
1
1
0
0
0
1
0
0
0
0
0
0
0
1
0
PB
V
F
1
1
0
0
0
0
1
0
1
0
Sole
a s
ole
a
Sole
0
1
1
1
1
0
0
0
0
1
0
0
0
0
0
PB
B
0
0
0
0
0
0
1
1
1
Spra
ttus s
pra
ttus
Spra
t 0
1
1
1
1
0
0
0
0
0
1
0
0
0
0
P
P
0
0
0
0
1
0
0
0
Syngnath
us a
cus
Gre
ate
r pip
efish
0
1
1
1
1
0
0
1
0
0
0
0
0
0
0
PB
B
F
0
0
0
1
0
0
1
1
1
Syngnath
us r
oste
llatu
s
Nils
son's
pip
efish
0
1
1
1
1
0
0
1
0
0
0
0
0
0
0
P
B
0
0
0
1
0
0
1
1
1
Tin
ca t
inca
Tench
1
0
0
0
0
1
0
0
0
0
0
0
0
0
1
PP
B
0
1
0
0
0
1
0
1
0
0
Trisopte
rus luscus
Pouting
0
1
1
1
1
0
0
0
0
1
0
0
0
0
0
B
BF
0
0
0
0
0
1
0
0
Zoarc
es v
ivip
aru
s
Viv
iparo
us b
lenny
0
1
1
1
1
0
0
1
0
0
0
0
0
0
0
PB
B
0
0
0
1
0
0
1
1

Table B: Essential fish habitats in relation to ecological guilds
MJ ER AN CA RA RB EU LIM
FRESHWATER TIDAL AREA
subtidal channel X X X X X
intertidal sandbanks X X
intertidal mudflat X X X
intertidal creeks X X X X X
intertidal marsh X X X X X
floodplain X X X X
BRACKISHWATER TIDAL AREA
subtidal channel X X X X
intertidal sandbanks X X
intertidal mudflat X X
intertidal creeks X X X X
intertidal marsh X X X X
MARINE TIDAL AREA
subtidal channel X X X X
intertidal sandbanks X X
intertidal mudflat X X
intertidal creeks X X X X
intertidal marsh X X X X
Mussel beds X X
Eelgrass beds X X
MJ: marine juvenile migrants; MA: marine adventitious species; MS: marine seasonal migrants; ER:
estuarine resident species; AN: anadromous species; CA: catadromous species; RA: rheophilic A
(obligate) species; RB: rheophilic B; EU: eurytopic species and LIM: limnophilic species.