enhancement of monoclonal antibody dependent cell mediated cytotoxicity by il2 and gm-csf
TRANSCRIPT

British Journal of Haefnatology. 1989. 73, 468-474
Enhancement of monoclonal antibody dependent cell mediated cytotoxicity by IL2 and GM-CSF
ALESSANDRA c. BIANCHI,* HELEN E. HESLOP, PAUL V E Y S , MARION M A C E Y , ~ MARK HOLLAND, H. GRANT PRENTICE A N D MALCOLM K. BRENNER Department of Haematology, Royal Free Hospital, Pond Street, Hampstead, London
Received 29 March 1989: accepted for publication 2 August 1989
Summary. Rodent monoclonal antibodies (MAb) directed against cells of the immune system may be used in vivo for applications including conditioning prior to marrow trans- plantation and treatment of lymphoid malignancies. Although some MAb may lyse targets by complement fixation, MAb dependent cell mediated cytotoxicity (MAb- DCC) appears to be a n important additional effector mecha- nism. We have investigated the cellular basis of the phenome- non and the response of the effector cells to recombinant cytokines in an attempt to maximize the efficacy of MAb-DCC and thereby increase the therapeutic potency of MAbs. Blood mononuclear cells (PBM) coated with CAMPATH 1 G (pan lymphocyte reactive rat IgG2b) MAb were used as targets (T) and autologous lymphocytes or granulocytes were used as effector (E) cells. We studied function in normal donors as well as patients 1 week before and 3-6 weeks after bone marrow transplantation (BMT). In the absence of CAMPATH 1 G, specific 5’Cr release from autologous PBM was < 1% in all groups, even after pre-incubation of the effector cells with granulocyte-macrophage, colony stimulating factor (GM-
The infusion of monoclonal antibodies (MAb) directed against immune system cells has been used as part of conditioning regimens prior to bone marrow transplantation (BMT)(Fischeretnl, 1986:Mackinnonetal, 1988; Prentice& Brenner, 1989) to abrogate graft rejection or graft-versus- host disease that may complicate the procedure (Martin et aZ, 19 88; Beelen et al, 1988) and for the treatment of lymphoma and lymphocytic leukaemia (Blanche et al, 1988; Dyer et al, 1988; Hale et nl, 1988). Other applications seem likely to follow, but almost all require the destruction or irreversible inactivation of the target cells. Although complement mediated lysis is one of the most important ways in which MAb destroy target cells in vitro. this mechanism appears less relevant iri vivo (Chassoux rt al. 1988). IgM MAb fix human
* Present address: Cattedra d’Ematologia. [Jniversity La SapienAa. Rome.
$ Department of Haematology. London Hospital Medical College. London E l LRB.
Correspondence: Dr M. K. Brenner, Department of Haematology, Royal Free Hospital. Pond Street, Hampstead. London NW3 2QG.
CSF) or interleukin-2 (IL2). In the presence 0 1 S pg/ml of MAb, and at an E:T ratio of S O : 1, lymphocytes from normal donors induced a low level (6.5%) of j’Cr release from auto PBM rising to 9% after pre-incubation of effector cells with ILZ ( P = 0.02 1. Granulocytes had greater activity inducing 10% jLCK release (range 2-23X) which rose to 21.6% with GM-CSF (range 12-48%) (P<0031) . Pre-BMT. killing by lymphocytes and granulocytes was not significantly different from normal. and responded to IL2 and GM-CSF. In contrast. granulocyte killing after BMT was significantly impaired (”Cr release 3%) and showed no rise with GM-CSF. Killing by lymphocytes, however, remained normal, as did their IL2 response. Loss of granulocyte mediated MAb-DCC coincided with significant post-BMT impairment of oxidative metab- olism: expression of Fc receptors I1 and 111. however, was normal. Optimum therapeutic effect of MAb-DCC is likely to be achieved when MAb are given together with appropriate cytokines. the choice of which will depend upon the clinical circumstances.
complement well, but produce only a short-lived reduction in circulating lymphocyte numbers implying they have poor ir i vivo tissue penetration (Dyer rt ul, 1989). IgG antibodies have better tissue penetration but a varied and unpredictable capacity to fix sufficient human complement to induce complete lysis of the target cell population (Chassoux et al. 1988; Bindon et al, 1988; Dyer et al, 1989). Nonetheless these IgG rodent MAb (and one humanized derivative) have been used with significant clinical success (Dyer ot nl, 1989; Hale et d. 19 88); in vivo they may produce cell destruction not only by C’ mediated lysis but also by antibody dependent cell mediated cytolysis (ADCC) (Chassoux et 01, 1988). This effector function is partly mediated by fixed tissue macro- phages but may also be produced by Fc receptor positive mononuclear cells (MNC) and by granulocytes (Dyer et nl. 1988). The relative contribution of these two cell types in v~vo is likely to depend both 011 the recruiting antibody and on the clinical setting in which such antibodies are used: in many of the clinical states described above, mononuclear cell and/or granulocyte numbers and function may be abnormal (Livnat
468

MAb Dependent Cell Mediated Cytotoxicity 469
et al. 1980; Clarke et al. 1976; Ichinos et al, 1986; Sosa et ( 1 1 , 1980: Kooney Pt al. 1986).
Analysts of the relative contribution of MNC and granulo- cytes to monoclonal antibody dependent cell mediated cytotoxictty (MAb-DCC) in different clinical conditions has therapeutic implications. as recombinant ( r ) cytokines able to enhance the cytotoxic function of each effector population are available for clinical use. For example, rIL2 has been shown to augment the cytotoxic function of both the lymphoid and monocytoid components of mononuclear cells (Braakman et a/. 1986: Malkovsky et al, 1987). while rGM- CSF promotes neutrophil killing (Perussia et al, 1987). Infusion of appropriate cytokines could thus potentiallv increase the efficacy of MAb given in vivo.
To investigate the contribution of MNC and granulocytes to MAb-DCC we have obtained mononuclear cells and neutrophils from normal individuals and from patients after Chemotherapy or after BMT. To assess the impact of rIL2 and rGM-CSF on the phenomenon. the effector cells were treated with each of these cytokines. Autologous lymphocytes were used as target cells and were coated with the monoclonal antibody CAMPATH 1 G (Hale e f al. 1988); this pan- lymphocyte rat IgG 2b MAb (and its humanized derivative) are used therapeutically in BMT and haematological malig- nancy (Dyer et a1, 1989; Hale et aI, 1988).
METHODS
Patient r~hnracteristics. Thirteen normal donors and 14 patients undergoing bone marrow transplantation were studied. Six patients underwent autologous and eight alloge- neic lymphocyte depleted bone marrow transplantation for haematological malignancies. Patients having bone marrow transplantation were conditioned with cyclophosphamide 1 2 0 mg/kg followed by single fraction total body irradiation (TBI) 750 Gy prior to cryopreserved autologous or HLA identical sibling marrow reinfusion (Prentice et 01, 1984) . For allogeneic transplants donor marrow had been lymphocyte depleted using CAMPATH IM antibody and fresh autologous serum as source of complement (Waldmann et a / , 1984) . All grafts were self sustaining and no patient was on any immunosuppressive agent a t the time of study. In patients undergoing bone marrow transplantation, studies were performed 1-2 weeks before conditoning for BMT, which was 4-10 weeks from their last course of chemotherapy. They were then re-studied 3-6 weeks following transplantation when the total white cell count had reached 1 x 10"/1.
Statisticcil analyses. Within group comparisons were made using paired t testing, while between group studies used analysis of variance (Kubycrete ANOVA program).
Prepamtion of purified neutrophils and lymphocytes. Dextran 80 was used to sediment 60-100 ml of venous blood. Leucocyte rich plasma was aspirated. washed once in RPMI 1640 (Flow Labs) and layered onto a discontinuous Percoll gradient. consisting of Percoll diluted 60% and 80% v/v with KPMT/jc)/, fetal calf serum (FCS). After spinning for 2 0 min at 400 g, cells from the upper interface-representing the mononuclear cell population-and cells from the lower interface-representing the polymorphonuclear granulo-
cytes-were aspirated and washed twice in RPMI, before resuspending in RPMI/10% FCS. The mononuclear cell preparation contained < 1% granulocytes, while the granu- locyte fraction contained < 1% cells positive for monocyte or lymphoid markers (CD5. CD19).
Cytokines. Recombinant IL2 and GM-CSF, expressed in Escherichia coli. were the gift of Glaxo-IMB, Geneva. Specific activity of r-IL2 was 1 . 7 x 10" u/mg protein and of r-GM-CSF 1 .0 x lox u/mg. Products contained no detectable endotoxin.
Treatment of the ejiector cells with reroriibinant c!/tokines. Granulocytes or mononuclear cells at a concentration of 2.5 x IOh/ml were incubated for 2 h a t 3 7°C in 5% C02 with or without granulocyte macrophage-colony stimulating fac- tor 12 50 PM. or IL2 200 or 500 u/ml. They were washed and resuspended in RPMI/lO% FCS at a final concentration of 5 x 1 O6/in1 before use in the "chromium release assay.
Tnrgets. 3-5 x lo6 mononuclear cells were pelleted and resuspended in a 200 pI of FCS, 7.4-1 1.1 MBq of sodium chromate (j'Cr) (Amersham International) were added and the cells incubated for 2 h a t 3 7°C in 5% CO?. After washing, the cells were resuspended in 100 p1 of medium. CAMPATH IgG 2b (CAMPATH 1G. Dr H. Waldmann. University of Cambridge) was added to produce a final concentration between 1 and 2 0 pg/pl. and the cells were incubated at 4°C for 30 min. After a final wash in RPMI, cells were resus- pended at 5 x 10'/ml for use in the 5'Cr release assay.
C;yfotoxicity assay. The jlCr release cytotoxicity assay was carried out in 96-well V-bottom microwell tissue culture plates (Nunc, Gibco) by adding 5 x 1 O4 target cells treated or untreated with CAMPATH 1 G in 0.1 ml of medium followed by either mononuclear cells or granulocytes (after incubation in medium alone or with the different cytokines as described) suspended in 0.1 ml of medium. Dilutions of effector cells were made to give four different effector to target ratios and each ratio was set up in triplicate. Values for maximum and spontaneous release were obtained by substituting Triton and KPMt/10% FCS respectively for the effector cell suspen- sion. After 4 h incubation at 3 7°C. in humidified 5% CO?. 100 111 of cell supernatant was aspirated for isotope counting. Per cent specific cytotoxicity was calculated from the formula:
x 100 experimental release - spontaneous release
maximum release - spontaneous release
Measurenienf of nrutrophil Fc receptors. 3-6 weeks after BMT, venous blood was taken from patients and from normal controls into 2 ml 5% Na EDTA. Leucocyte rich plasma was obtained by dextran sedimentation for 30 tnin at room temperature. The supernatant was washed twice in cold phosphate buffered saline (PBS)/O.2% bovine serum albumin (BSA) and the leucocytes resuspended in cold PBS/0.2o/, BSA at a cell concentration of 2 x 10h/ml. 100 pl aliquots were labelled separately with anti-human FcRII (FITC IV3. Medare) and anti-human FcRIII (Leu IIa. Becton Dickinson) with appropriate irrelevant antibodies as controls. All label- ling was undertaken at 4°C. Neutrophil membrane fluores- cence was measured using flow cytometric analysis using parameters previously described (Veys et al, 1989) .
Chentilurninesccnce ( C L ) . Granulocytes were separated on a discontinuous Percoll gradient as previously described (Hes-
470 Alessandra C. Bianchi et a1
^^ M-ADCC Normals T ri 20 MNC 6 Granulocytes 16 - 16 s - 14 : 12
3 6
10 t?
6 4 2 0
MNC-/+ IL2 Granulocytes -/+ CSF Cell type
El UAbtCK
22 20 16 16 - 14 s - 12
% p 10 E 8 0 6
4 2 0
- ..
E:T ra t io
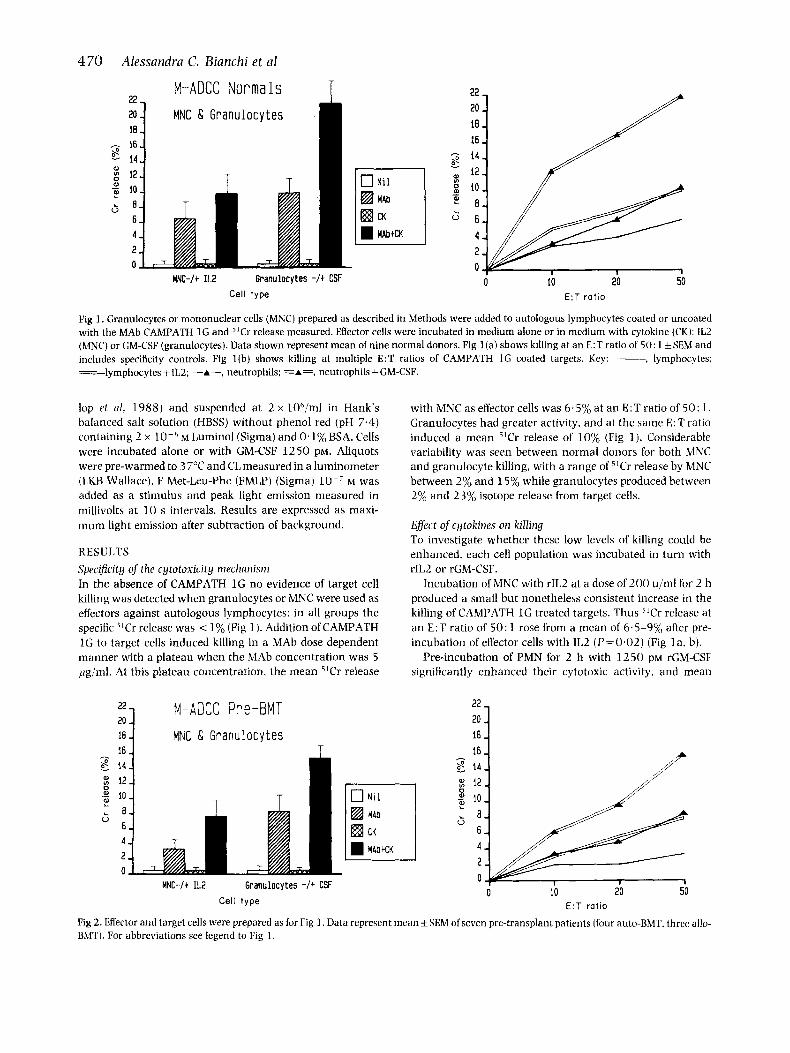
Pig 1 . Granulocytes or mononuclear cells (MNC) prepared as described in Methods were added to autologous lymphocytes coated or uncoated with the MAb CAMPATH 1 G and ’lCr release measured. Effector cells were incubated in medium alone or in medium with cytokine (CK): IL2 (MNC) or GM-CSF (granulocytes). Data shown represent mean of nine normal donors. Fig 1 (a) shows killing at an E:T ratio of 50: 1 +SEM and includes specificity controls. Fig l (b ) shows killing at multiple E:T ratios of CAMPATH 1 G coated targets. Key: ___ , lymphocytes: =lymphocytes+ IL2: -A-, neutrophils; =A=, neutrophils +GM-CSF.
lop rt r i l , 1988) and suspended at 2 x 10h/ml in Hank’s balanced salt solution (HBSS) without phenol red (pH 7.4) containing 2 x lo-‘ M Luminol (Sigma) and 0.1% BSA. Cells were incubated alone or with GM-CSF 1250 PM. Aliquots were pre-warmed to 3 7°C and CL measured in a luminometer (LKB Wallace). F Met-Leu-Phe (FMLP) (Sigma) lo-’ M was added as a stimulus and peak light emission measured in millivolts at 10 s intervals. Results are expressed as maxi- mum light emission after subtraction of background.
RESULTS Specijcity of’ the cytotoxicity nirclianisrn In the absence of CAMPATH 1G no evidence of target cell killing was detected when granulocytes or MNC were used as effectors against autologous lymphocytes: in all groups the specific ”Cr release was < 1% (Fig 1 ). Addition of CAMPATH 1C; to target cells induced killing in a MAb dose dependent manner with a plateau when the MAb concentration was 5 pg/ml. At this plateau concentration, the mean “Cr release
M-ADCC Pre-BMT :: 1 6 4 MNC 6 Granulocytes
with MNC as effector cells was 6.5% at an E:T ratio of 50: 1. Granulocytes had greater activity, and at the same E : T ratio induced a mean ilCr release of 10% (Fig 1 ) . Considerable variability was seen between normal donors for both MNC and granulocyte killing, with a range of 51Cr release by MNC between 2% and 15% while granulocytes produced between 2% and 2 3% isotope release from target cells.
Eflect of cytokines on killing To investigate whether these low levels of killing could be enhanced, each cell population was incubated in turn with rIL2 or rGM-CSF.
Incubation of MNC with rIL2 at a dose of200 u/ml for 2 h produced a small but nonetheless consistent increase in the killing of CAMPATH 1 G treated targets. Thus j’Cr release at a n E:T ratio of 50: 1 rose from a mean of 6.5-9% after pre- incubation of effector cells with IL2 (P=0.02) (Fig 1 a, b).
Pre-incubation of PMN for 2 h with 1250 PM rGM-CSF significantly enhanced their cytotoxic activity, and mean
22
16 I “I 14
16
2 12 * 10 Y L 8
6 4 2 0
,-.. g 14
0 0
HAbtCK
0 10 20 50 Granulocytes -/+ CSF MNC-/t ILZ
Cell type E:T ra t io
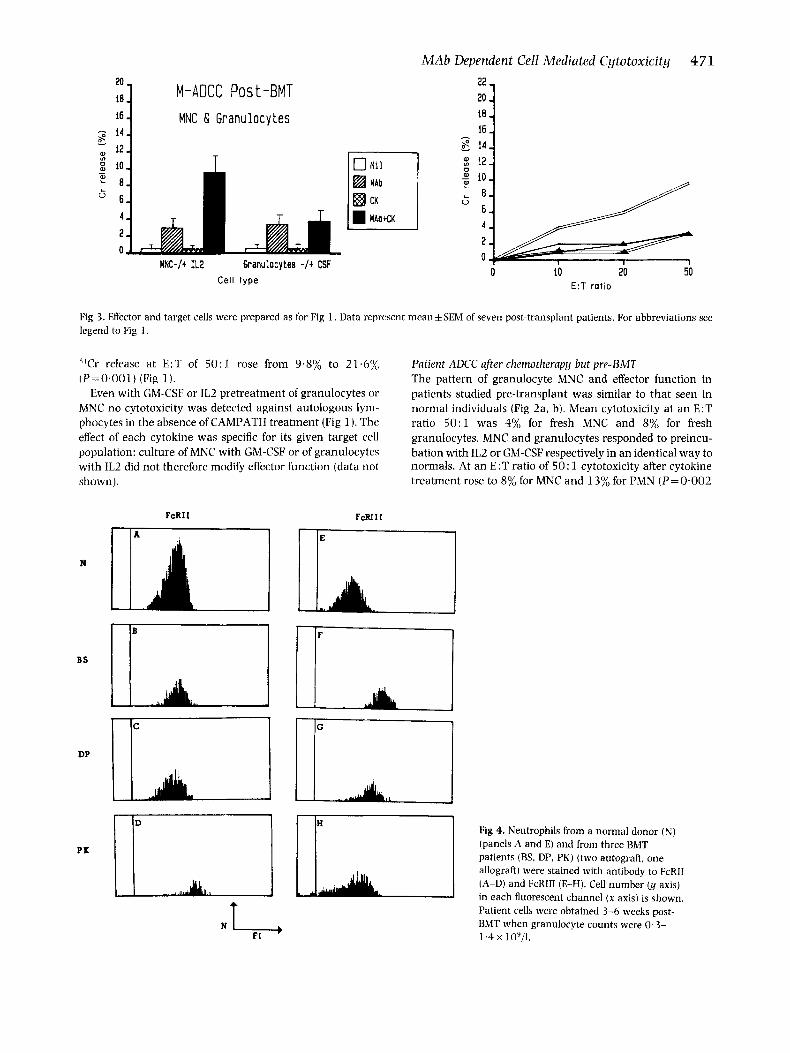
Fig 2. Effector and target cells were prepared as for Fig 1. Data represent mean & SEM of seven pre-transplant patients (four auto-BMT. three allo- BMT). For abbreviations see legend to Fig 1.
20 M-ADCC POS t -BMT MNC 6 Granulocytes - 14 s - 12
% r
HNC-/+ IL2 Granulocytes -/t CSF Cell type
MAb Dependent Cell Mediated Cytotoxicity 471 22
16
!!!!A WAbtCK
0 10 20 50 E:T ratio
Fig 3 . Effector and target cells were prepared as for Fig 1. Data represent rneanf SEM of seven post-transplant patients. For abbreviations see legend to Fig 1 .
"Cr release at E:T of 50: l rose from 9.8% to 21.6%, (P=O.OOl) (Fig 1).
Even with GM-CSF or IL2 pretreatment of granulocytes or MNC no cytotoxicity was detected against autologous lym- phocytes in the absence of CAMPATH treatment (Fig 1 ). The effect of each cytokine was specific for its given target cell population: culture of MNC with GM-CSF or of granulocytes with IL2 did not therefore modify effector function (data not shown).
I
BS
DP
PK
FcRII
t
Patient ADCC after chemotherapy but pre-BMT The pattern of granulocyte MNC and effector function in patients studied pre-transplant was similar to that seen in normal individuals (Fig 2a, b). Mean cytotoxicity at an E:T ratio 5 0 : l was 4% for fresh MNC and 8% for fresh granulocytes. MNC and granulocytes responded to preincu- bation with IL2 or GM-CSF respectively in an identical way to normals. At an E:T ratio of 50: 1 cytotoxicity after cytokine treatment rose to 8% for MNC and 13% for PMN (P=0.002
FcRl I I
7 L
H I Fig Neutrophils from a normal donor (N)
I (panels A and E) and from three BMT patients (BS. DP. PK) (two autograft. one
I Il.lr I allograft) were stained with antibody to FcRII
I (A-D) and FcRIII (E-H). Cell number (y axis) in each fluorescent channel ( x axis) is shown.
"+ Patient cells were obtained 3-6 weeks post- BMT when granulocyte counts were 0.3- 1.4 x 10'11.
472 Alessandra C. Bianchi et a1
1 1 2 0
1 0 0
8 0 > E - 6 0 -I 0
-
4 0
2 0
0
Time ( m i d
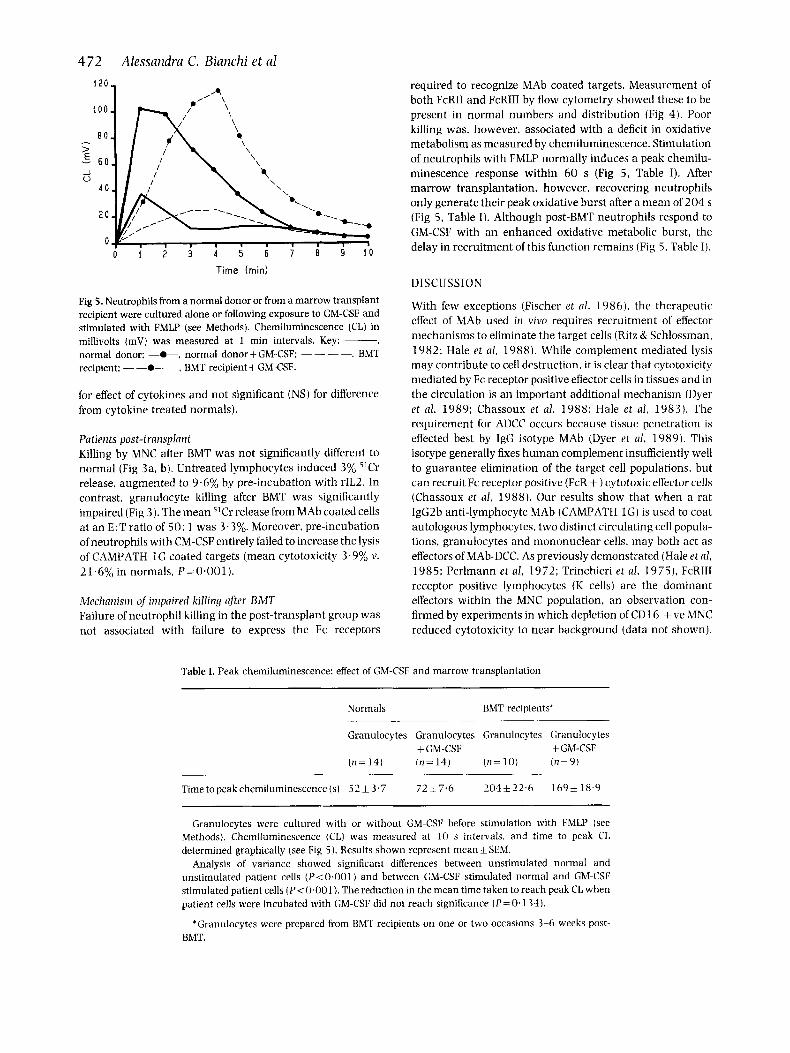
required to recognize MAb coated targets. Measurement of both FcRII and FcRIII by flow cytometry showed these to be present in normal numbers and distribution (Fig 4). Poor killing was, however, associated with a deficit in oxidative metabolism as measured by chemiluminescence. Stimulation of neutrophils with FMLP normally induces a peak chemilu- minescence response within 6 0 s (Fig 5, Table I). After marrow transplantation, however, recovering neutrophils only generate their peak oxidative burst after a mean of 204 s (Fig 5, Table I). Although post-BMT neutrophils respond to GM-CSF with a n enhanced oxidative metabolic burst. the delay in recruitment of this function remains (Fig 5. Table I).
Fig 5. Neutrophils from a normal donor or from a marrow transplant recipient were cultured alone or following exposure to GM-CSF and stimulated with FMLP (see Methods). Chemiluminescence (CL) in millivolts (mV) was measured at 1 min intervals. Key: -, normal donor; -0-, normal donor -t GM-CSF: - - - -. BMT recipient: - -0- -, BMT recipient + GM-CSF.
for effect of cytokines and not significant (NS) for difference from cytokine treated normals).
Patierits post-trarisplant Killing by MNC after BMT was not significantly different to normal (Fig 3a, b). Untreated lymphocytes induced 374 51Cr release. augmented to 9.6% by pre-incubation with rIL2. In contrast. granulocyte killing after BMT was significantly impaired (Fig 3 ) . The mean jlCr release from MAb coated cells at an E:T ratio of 50: 1 was 3 .3%. Moreover, pre-incubation of neutrophils with CM-CSF entirely failed to increase the lysis of CAMPATH 1 G coated targets (mean cytotoxicity 3.9% v. 21.6% in normals, P=O.OOl) .
Mechanism of inipoired killing ofter BMT Failure of neutrophil killing in the post-transplant group was not associated with failure to express the Fc receptors
DISCUSSION
With few exceptions (Fischer et ol. 1986), the therapeutic effect of MAb used iri vivo requires recruitment of effector mechanisms to eliminate the target cells (Kitz L? Schlossman, 1982: Hale et a / . 1988). While complement mediated lysis may contribute to cell destruction, it is clear that cytotoxicity mediated by Fc receptor positive effector cells in tissues and in the circulation is an important additional mechanism (Dyer et ( 7 1 . 1989: Chassoux et al. 1988: Hale et ul. 1983). The requirement for ADCC occurs because tissue penetration is effected best by IgG isotype MAb (Dyer et al, 1989). This isotype generally fixes human complement insufficiently well to guarantee elimination of the target cell populations, but can recruit Fc receptor positive (FcR + ) cytotoxic effector cells (Chassoux et al. 1988). Our results show that when a rat IgG2b anti-lymphocyte MAb (CAMPATH 1 G ) is used to coat autologous lymphocytes, two distinct circulating cell popula- tions. granulocytes and mononuclear cells, may both act as effectors of MAb-DCC. As previously demonstrated (Hale et al. 1985; Perlmann e t 01, 1972; Trinchieri et nl, 1975). FcRlII receptor positive lymphocytes (K cells) are the dominant effectors within the MNC population, an observation con- firmed by experiments in which depletion of CD16 f v e MNC reduced cytotoxicity to near background (data not shown).
Table I. Peak chemiluminescence: effect of GM-CSF and marrow transplantation
Normals BMT recipients'
Granulocytes Granulocytes Granulocytes Granulocytes + GM-CSF + GM-CSF
(11=14) (r7=74) ln=10) ( n = 9 )
Time topeakchemiluminescence(s) 5 2 1 3.7 72 & 7.6 204f22 .6 l 6 9 k 1S.9
Granulocytes were cultured with or without GM-CSF before stimulation with FMLP (see Methods). Chemiluminescence (CL) was measured at 10 s intervals. and time to peak CL determined graphically (see Fig 5). Results shown represent mean f SEM.
Analysis of variance showed significant differences between unstimulated normal and unstimulated patient cells (P< 0,001) and between GM-CSF stimulated normal and GM-CSF stimulated patient cells ( P < 0.001 ). The reduction in the mean time taken to reach peak CI, when patient cells were incubated with GM-CSF did not reach significance (P=0,134).
*Granulocytes were prepared from BMT recipients on one or two occasions 3-6 weeks post- BMT.

MAb Dependent Cell Mediated Cgtotoxicity 473 cell killing, perhaps by allowing increased time for the target cell to repair oxidative damage.
Whatever mechanism underlies reduced granulocyte cyto- toxicity. our observations illustrate that the choice of cyto- kine(s) to enhance MAb-DCC will be influenced by the clinical setting in which they are to be used. Moreover, while these data refer only to CAMPATH l G , the observations are likely also to apply to the iri vivo use of other rodent MAb with more restricted target cell specificity. Appropriately chosen cyto- kines should also enhance killing when hybrid humanized MAb variants (Hale et al. 1988) are used, since these MAb too would be predicted to recruit identical cellular effector mechanisms with the same cytokine responsiveness.
But while these results confirm that MAb-DCC can contribute to target cell elimination, levels of killing measured by j'Cr release in a 4 h assay are low (Fig 1). In an attempt to boost cytotoxicity, and thereby promote MAb potency. we investi- gated the action of two cytokines which have proved effective in promoting ADCC mediated by conventional polyclonal antisera. We studied GM-CSF which can enhance granulo- cyte ADCC (Weisbart r t nl, 1986, 1987: Gasson et al, 1984; Fabian rt 01, 1987: Lopez e t d . 1986) and IL2 which has been reported to increase killing by both the lymphocyte and monocyte components of blood MNC (Braakman et nl, 1986: Malkovsky et al. 1987). Pre-incubation of effector lympho- cytes with IL2 enhanced mononuclear cell killing of MAb coated targets while incubation of neutrophils with GM-CSF increased granulocyte killing (Fig 1). These effects were specific; ILL had no action on granulocytes while GM-CSF did not modify MNC mediated lysis. neither did it induce killing of MAb uncoated target lymphocytes. Other cytokines includ- ing yIFN, TNF and M-CSF (Shalaby et a/. 198 5: Perussia et a!, 1987; Heslop et n1, 1988) may also promote granulocyte/ MNC killing in normal individuals, and the effect of cytokine combinations on MAb-DCC remains to be evaluated.
The ability of cytokines to promote granulocyte and MNC MAb dependent killing in normal donors may have little relevance in the clinical setting in which the MAb are likely to be used. CAMPATH l G , for example, has been used as part of the conditioning regimen prior to marrow transplantation and for the treatment of GvHD following the procedure. MAb- DCC and the response to cytokines may be abnormal in both patient groups, pre-transplant because of the effects of chemotherapy (Ichinos et nl. 1986; Livnat r t a] . 1980) and post-transplant because of the delay in restitution of normal granulocyte and MNC function and phenotype (Clarke et al. 1976: Sosa rt nl, 1980; Brenner et al. 1987; Reittie et n1. 1989). I n fact. our results show that MAb-DCC mediated by both MNC and granulocytes are normal pre-BMT and can be enhanced by cytokine exposure. In contrast, patterns of killing post-BMT are abnormal. The MNC population kill normally and cytotoxicity is increased by IL2. However. attempts to enhance ADCC by infusion of IL2 may be of limited value after conventional allogeneic BMT since IL2 itself can increase the severity of GvHD (Malkovsky rt 01. 1986) by its action on alloreactive T cells, and could probably only be used after T cell depleted BMT. Enhancement of granulocyte cytotoxicity by GM-CSF would be more readily applicable to all allografts. The data in Fig 3 , however, illustrate clearly that after BMT granulocyte killing is poor and cannot be restored by GM-CSF. At least two events are required for granulocyte killing of MAb coated target cells. The effector cell must bind to its target via its FcRII and I11 receptors (Shen et d. 1988) and a n oxidative burst must be generated (Fabian et ul, 1987). After BMT circulating neutrophils may be functionally immature, and although Fc receptor I1 and 111 expression is normal, recruitment of oxidative metabolism-as measured by chemilumines- cence-is delayed (Fig 5). GM-CSF increases peak oxidative metabolism, but it does not reduce the time taken to generate the burst, so that maximal activity is substantially delayed. We suggest that this recruitment delay compromises target
ACKNOWLEDGMENTS
This work was supported by the Wellcome Trust and the [Jniversity La Sapienza (Rome). We would like to thank Dr Phillip Diamond for his help with isotope measurements. Mrs Joyce Reittie for technical assistance, Drs H. Waldmann and G. Hale for their provision of CAMPATH 1 G and Mrs Megan Evans for word processing the manuscript.
REFERENCES
Beelen. D.W., Graeven. LJ.. Schulz. G., Grosse-Wilde. H.. Doxiadis. I., Schaefer. L1.W.. Quabeck. K.. Sayer, H. & Schmidt, C.G. (1988) HLA-partially matched marrow transplantation with a nionoclon- al antibody (BMAO31) against the T cell receptor. First results of a phase 1/11 trial. Onkologic. 11, 56-58.
Bindon. C.I.. Hale, G.. Bruggemann. M. & Waldmann, H. (1988) Human monoclonal IgG isotypes differ in complement activation function at the level of C4 as well as Clq. loiirnal of Experimental Medicine. 168, 127-142.
Blanche. S., Le Deist, F.. Veber. F.. Lenoir, G.. Fischer, A.M.. Brochier. I.. Boucheix. C.. Delaage. M., Griscelli. C. & Fischer. A. (1988) Treatment of severe Epstein-Barr virus-induced polyclonal B- lymphocyte proliferation by anti-B cell monoclonal antibodies. Two cases after HLA-mismatched bone marrow transplantation. Aniinls of lriteriinl Medicine, 108, 199-203.
Braakman, E.. Van Tunen. A,. Meager. A . & Lucas. C.J. (1986) ILL and WN gamma-enhanced natural cytotoxic activity: analysis of the role of different lymphoid subsets and implications for activation routes. Cellular lmn ludogy . 99, 476-488.
Brenner. M.K.. Vyakarnam, A,. Reittie, J.E.. Wimperis. J.Z., Grob, J.Ph., Hofirand, A.V. & Prentice. H.G. (1987) Human large granular lymphocytes induce immunoglobulin synthesis after bone marrow transplantation. Eriropenn Journal oflmmunology, 17, 43-47.
Chassoux. D.M.. Linares-Crux, L.G., Bazinb, H. & Stanislawski. M. (1988) K-cell-mediated cytotoxicity induced with rat monoclonal antibodies. I. Antibodies of various isotypes differ in their ability to induce cytotoxicity mediated by rat and human effectors. Ifnniu-
Clark, R.A.. Johnson, F.L.. Klebanoff, S J . & Thomas, E.D. (1976) Defective neutrophil chemotaxis in bone marrow transplant patients. /uurnnl of Clinical Irivcstigatiori. 58, 22-3 1.
Dyer, M.J.S., Hale. G.. Hayhoe, F.C.J.& Waldrnann. H. (1988) Effects of monoclonal antibodies of the Campath series in vivo: require- ments for effective antibody therapy. (Abstract). British Joournal of Hnenintologg, 69. 119.
Dyer. M.J.S., Hale, G.. Hayhoe. F.G.J. 6: Waldmann. H. (19891 Effects
rlolog!j. 65, 613-628.

474 Alessandra C. Bianchi et a1 of CAMPATH-1 antibodies in patients with lymphoid malignan- cies: influence of antibody isotype. Blood. 73, 1431-1439.
Fabian, I.. Baldwin, G.C. & Golde. D.W. (1987) Biosynthetic granulocyte-macrophage colony-stimulating factor enhances neutrophil cytotoxicity toward human leukemia cells. Leukerttia. 1,
Fischer, A,. Blanche. S., Veber, F., Delaage. M.. Mawas, C.. Griscelli. C., Le Deist, F.. Lopez. M.. Olive, D. &Janossy. G. (1986) Prevention of graft failure by an anti-HLFA-1 monoclonal antibody in HLA- mismatched bonemarrow transplantation. Lancet. ii, 1058-1061.
Gasson. J.C., Weishart. R.H., Kaufman. S.E.. Clark. S.C.. Hewick. R.M.. Wong. G.G. & Colde. D.W. (1984) Purified human granulo- cyte-macrophage colony stimulating factor: direct action on neutrophils. Scietice, 226, 1339-1 342.
Hale, G.. Clark, M. & Waldmann. H. ( 1 985) Therapeutic potential of rat monoclonal antibodies: Isotype specificity of antibody-depen- dent cell-mediated cytotoxicity with human lymphocytes. /ournu1 of Imrniinolog~~. 134, 3056.
Hale, G.. Dyer, M.J.S.. Clark. M.R.. Phillips. I.M., Marcus, R.. Reichmann. L.. Winter, G. & Waldrnan. H. (1988) Remission induction in non-Hodgkin's lymphoma with reshaped human monoclonal antibody Campath-1H. Lancet, ii, 1394-1 399.
Hale. G.. Hoang. T.. Prospero. T.. Watt. S.M. & Waldmann. H. (1983) Removal of T cells from some marrow for transplantation: comparison of rat monoclonal anti-lymphocyte antibodies of different isotypes. Molecular Biology and Medicine, I , 305-3 19.
Heslop. H.E., Price, G.M., Prentice. H.G.. Cordingley. F.T., Webster, A.D.B.. Hofforand. A.V. & Brenner. M.K. (1988) In vitro analysis of the interactions of recombinant IL2 with regenerating lymphoid and myeloid cells after allogeneic hone marrow transplantation. /oirrtiul of Irnrnurtoloyg, 140, 3461-3466.
Ichinos. Y.. Hara, N.. Motohiro. A,. Noge, S.. Ohta. M. & Yagawa. K. ( 1 986) Influence of chemotherapy on superoxide anion generat- ing activity of polymorphonuclear leukocytes in patients with lung cancer. Canrer. 58, 1663-1 667.
Livnat. S.. Seigneuret. M.. Storb, R. & Prentice, R.L. (1980) Analysis of cytotoxic effector cell function in patients with leukemia or aplastic anemia before and after marrow transplantation. /ourrial of b?~nrurtolog~j. 124, 481-490.
Lopez. A.F., Williamson, D.]., Gamble, J.R.. Begley, C.G.. Harlan. J.M.. Klehanoff, S.J.. Waltersdorph. A,. Wong. G., Clark, S.C. & Vadas. M.A. ( 1986) Recombinant human granulocyte-macrophage col- ony-stimulating factor stimulates in vitro mature human neutro- phi1 and eosinophil function, surface receptor expression and survival. lourrinl qf Clinical Iiiv&igutiori, 78, 1220-1228.
Mackinnon. S..]ames. N.D., Apper1ey.J.F.. Arthur. C.K.. Sikora. K. & Goldman, J.M. (1988) Addition of total lymphoid irradiation (TLI) and chemotherapy assures engraftment in T cell depleted (TCD) donor marrow in allogeneic hone marrow transplant (BMT) for leukaemia. (Abstract). British /ournu1 of Haematology. 69, 188.
Malkovsky. M.. Brenner, M.K., Hunt, R.. Rastan, S., Dore, C.. Brown, S.. North, M.E., Asherson, G.L.. Prentice, H.G., Medawar. P.B. (1986) T cell depletion of allogeneic bone marrow prevents acceleration of graft-versus-host disease induced by exogenous interleukin 1. Cellirlar lntmiinology. 103. 476-48 1.
Malkovsky, M.. Loveland. B.. North, M.. Asherson. G.L., Gao. L.. Ward, P. & Fiers, W. (1987) Recombinant interleukin-2 directly augments the cytotoxicity of human monocytes. Nature. 325,
Martin. P.J., Hansen. J.A., Anasetti. C., Zutter. M., Durnam, D.. Storb. R. & Thomas, E.D. (1988) Treatment of acute graft-versus-host disease with anti-CD3 monoclonal antibodies. American lourrial of Kidlie!) Disense. 11, 149-1 52.
613-617.
262-265.
Perlmann, P.. Perlmann. H. & Wigzell. H. (1972) Lymphocyte mediated cytotoxicity in vitro: induction and inhibition by humoral antibody and nature of effector cells. Trnttsplnritatiori Reviews. 13, 9 1 .
Perussia. B.. Kohayashi, M., Rossi, M.E.. Anegon. 1. & Trinchieri. C. ( 1 987) Immune interferon enhances functional properties of human granulocytes. Role of Fc receptors and effect of lympho- toxin, tumour necrosis factor. and granulocyte-macrophage col- ony stimulating factor. Joiirrial of Inirnirnoloyy. 138, 765-774.
Prentice. H.G.. Blacklock, H.A.. Janossy. G.. Gilmore, M.I.M.L., Price- Jones. L.. Trejdosiewicz, L.K.. Skeggs. D.B.L.. Panjwani, D.. Ball. s.. Graphakos. S.. Patterson, J. & Hoarand. A.V. (1984) Depletion of T lymphocytes in donor marrow prevents significant graft versus host disease in matched allogeneic leukaemic marrow transplant recipients. Lancet, i, 472-476.
Prentice. H.G. &Brenner.M.K. (1989)DonormarrowTcelldepletion for prevention of GvHD without loss of the GvL effect. Current results in acute myelohlastic leukaemia and future directions. Brvir~ Marrow Transplantation: Current Coritrovrrsiils (ed. by R . P. Gale. R. E. Champlin). p. 11 7. Alan R. Liss. New York.
Reittie, J.E.. Cottlieb. D., Heslop, H.E.. Leger. 0.. Drexler. HC.. Hazlehurst. G.. Hofforand. A.V.. Prentice. H.G. & Brenner. M.K. (1989) Endogenously generated activated killer cells circulate after autologous and allogeneic marrow transplantation but not after chemotherapy. Blood, 73, 1351-1358.
Rita. J. & Schlossman. S.F. (1982) Utilization of monoclonal anti- bodies in the treatment of leukemia and lymphoma. Blood. 59. 1- 1 1 .
Rooney. C.M.. Wimperis. J.Z.. Brenner. M.K.. Patterson. J.. Hoff- brand. A.V. & Prentice. H.G. (1986) Natural killer cell activity following T-cell depleted allogeneic bone marrow transplantation. British Joirrrial of Hnernntology. 62, 41 3-420.
Shalaby. M.R.. Aggarwal. B.B.. Rinderknecht. E.. Suedersky, L.P.. Finkle. B.S. & Palladino. MA. (1985) Activation of human polymorphonuclear neutrophil functions by gamma interferon and TNF. joiirrial ofZr?iniunology. 135. 2069-2073.
Shen. L., Guyre. P.M. & Fanger, M.W. (1987) Polymorphonuclear leukocyte function triggered through the high affinity Fc receptor for monomeric IgG. Jourrial ofbnmitnology. 139, 5 34-538.
Sosa. R.. Weiden, P.L.. Storb. R.. Syrotuck, J . &Thomas, E.D. ( 1980) Granulocyte function in human allogeneic marrow graft reci- pients. Experinteiital Hernatology. 8, 1183-1 189.
Trinchieri, G.. Banmann. P., de March. M. & Tokes, Z. (1975) Antibody-dependent cell-mediated cytotoxicity in humans. I. Characterization of the effector cell. /oirrnnl of In i i~ i i~ t io log~~, 1 IS, 249.
Veys. P.A.. Gutteridge. C.N., Macey. M.. Ord. 1. & Newland, A.C. (1989) Detection of granulocyte antibodies using flow cytometric analysis of leukocyte immunofluorescence. Vox Sangirinis, 56.42- 47.
Waldmann. H., Polliak. A,. Hale, G., Or, R., Cividalli. G., Weiss, L.. Weshler. Z., Samuel, S., Manor. D.. Brautbar. C.. Rachmilewita. E.A. & Slavin. S. ( 1 984) Elimination of graft versus host disease by in vitro depletion of alloreactive lymphocytes with a monoclonal rat anti-human antibody (CAMPATH-I ). Lnricet. ii, 48 3-486.
Weishart. R.H.. Golde, D.W. & Gasson. 1.C. (1986) Biosynthetic human GM-CSF modulates the number and affinity of neutrophil F-Met- Leu-€'he receptors. /ourrial of Initnunofogy, 137, 3584- 3587.
Weishart. R.H.. Kwan. L.U.Z., Golde. D.W. & Gasson, J.C. 1198;) Human GM-CSF primes neutrophils for enhanced oxidative metabolism in response to the major physiological chemoattrac- tants. Blood. 69. 18-2 1.