characterization of desirable and undesirable lactobacilli from cheese in fermented milk
TRANSCRIPT

Lebensm.-Wiss. u.-Technol., 32, 433}439 (1999)
Characterization of Desirable and UndesirableLactobacilli from Cheese in Fermented Milk
M. A. Drake*, Y. KaraguK l-YuK ceer, X. Q. Chen and K. R. Cadwallader
Department of Food Science & Technology, Southeast Dairy Foods Research Center, Mississippi State University,MS 39762-9805 (U.S.A.)
(Received August 21, 1998; accepted June 6, 1999)
Aroma volatiles, acid production and proteolytic activity in the fermented milk of xve lactobacillus cheese adjuncts were studied. Thelactobacillus strains were previously characterized as desirable or undesirable adjuncts in cheese. The xve lactobacilli varied widely involatile production, acid production and proteolytic activity. The presence of a lactococcus did not enhance volatile production orproteolytic activity (P(0.05). Methyl ketones, 2-heptanone and 2-nonanone, were produced in large quantities by the desirable cheeseadjuncts (P(0.05). Proteolytic activity was also higher in the two desirable cheese adjuncts (P(0)05). Proteolytic activity was alsohigher in the two desirable cheese adjuncts (P(0.05). Methanethiol was produced by one desirable and one undesirable adjunct. It wasnot possible to cluster the lactobacilli using principal components analysis of volatile production and proteolytic activity as desirable orundesirable cheese adjuncts.
( 1999 Academic Press
Keywords: cheese #avour; adjunct starters; lactobacilli; #avour volatiles; proteolysis
Introduction
Starter cultures are responsible for the #avours found infermented dairy products. Volatile and nonvolatile #a-vour compounds are produced by various enzymes andmetabolic activities (1). The role of lactic acid bacteria incheese #avour development in particular has been thefocus of much research (2}4). Both proteolytic enzymesand key volatile production are thought to be importantin the development of desirable cheese #avours.Much research has focused on #avour components of thecheese or on the comparative e!ects of di!erent adjunctstarts (5}7). Recent research has addressed the total vol-atile production in milk of various mesophilic and ther-mophilic lactobacilli on aroma impact (8), but no e!orthas been made to relate the results to the role that theindividual lactobacilli play in dairy products. Little pub-lished research has focused on volatile and proteolyticactivity of cheese adjunct starters in milk.Curd slurries have been proposed as an alternative totime-consuming cheese studies for screening and evalu-ation of starter cultures and starter adjuncts (9}11).A disadvantage to curd slurries, however, is the preven-tion of contamination by other microorganisms (12). Theuse of sterile or heat-treated fermented milk for cultureevaluation would be a quicker alternative to curd slurriesand would provide less opportunity for contamination.
*To whom correspondence should be addressed. Fax: 601-325-8727;E-mail: [email protected]
0023-6438/99/070433#07 $30.00/0( 1999 Academic Press All ar
43
The objective of this study was to examine the volatileproduction and proteolytic activity of "ve characterizedadjunct lactobacilli in fermented milk with and withoutthe presence of starter lactococci and to determinewhether any relationships among the adjunct starterscould be determined based on these data.
Materials and Methods
ChemicalsStandard reference compounds identi"ed in Table 1 wereobtained from the following commercial sources:no. 1 (Mallinckrodt Inc., Paris, KY, U.S.A.); nos 2, 4, 5,8}11, 13}20 (Aldrich Chemical Co., St. Louis, MO,U.S.A.); no. 3 (Spectrum Chemical Mfg. Corp., Gardena,CA, U.S.A.); nos 6, 12 (Sigma Chemical Co., St. Louis,MO, U.S.A.) and no. 7 (J.T. Baker Chemical Co., Phillip-sburg, NJ, U.S.A.). The internal standard (2-methyl-3-heptanone) was purchased from Aldrich Chemical Co.Microbiological media were purchased from BBL BectonDickenson (Cockeysville, MD, U.S.A.). Reconstitutedskim milk (100 g/L) was autoclaved at 121 3C for 8 minand used for all experiments.
Cultures¸actococcus lactis ssp lactis MM410 (Rhodia, Inc.Madison, WI, U.S.A.) was grown in Elliker's broth at30 3C. ¸actobacillus casei LC301, ¸actobacillus paracaseiL71, and ¸actobacillus pentosus L23 were obtained from
Article No. fstl.1999.0572ticles available online at http://www.idealibrary.com on
3

Tab
le1
Con
cent
ration
sof
sele
cted
vola
tile
com
poun
dsin
bas
em
ilkan
dfe
rmen
ted
milk
sde
term
ined
by
DH
S-G
C-M
S
Conce
ntr
atio
n(k
g/L)
MM
410#
MM
410#
MM
410#
MM
410#
MM
410#
No.
Com
pound
RI
Bas
em
ilkM
M41
0L71
LH
212
LC
301
CN
RZ32
L23
LC
301
L71
L23
LH
212
CN
RZ32
1A
ceta
ldeh
yde
(60
08.
40c
273.
11a
171.
36abc
83.6
5bc
9.16
c30
.56c
35.0
4c11
8.00
abc
162.
57ab
c22
7.85
ab13
3.00
abc
79.5
5bc
2M
ethan
ethio
l(
600
0.80
b1.
05b
157.
99a
0.19
b1.
39b
29.0
1ab
0.75
b5.
36b
8.00
b1.
65b
4.23
b0.
75b
3A
ceto
ne
(60
016
4.53
a6.
91d
76.4
1abcd
8.18
d12
6.09
ab11
1.62
abc
44.6
4bcd
61.1
4bcd
78.9
5abcd
13.2
7cd
112.
64ab
c55
.67b
cd
4D
imet
hyl
sulp
hid
e(
600
0.87
b1.
05b
0.49
b0.
76b
0.47
b2.
93ab
0.26
b1.
63ab
4.65
a0.
42b
0.31
b0.
14b
5D
iace
tyl
620
13.1
7b76
.53b
97.0
0b7.
60b
1132
.08a
3.23
b0.
23b
792.
64a
230.
20b
64.7
8b69
.98b
29.7
4b6
2-Buta
none
625
1.24
a0.
30cd
0.61
bcd
0.11
d1.
11ab
0.9a
bc0.
38cd
0.66
abcd
0.49
cd0.
45cd
0.90
abc
0.68
abcd
7Eth
ylac
etat
e63
50.
38a
3.56
a4.
56a
2.12
a4.
57a
15.6
6a0.
65a
7.83
a4.
61a
3.88
a2.
69a
4.58
a
83-
Met
hyl
buta
nal
668
0.96
c1.
71bc
5.31
a2.
14bc
1.72
bc3.
70ab
1.48
bc3.
02bc
1.69
bc1.
82bc
1.95
bc1.
96bc
92-
Met
hyl
buta
nal
675
8.05
c16
.38a
bc41
.94a
b17
.86a
bc16
.79a
bc42
.93a
10.4
0c21
.39a
bc14
.55b
c13
.61c
16.7
8abc
16.1
7abc
102-
Pen
tanon
e69
51.
12b
1.07
b1.
49b
5.14
b5.
00b
16.9
3a0.
62b
4.42
b0.
60b
0.49
b5.
86b
5.06
b
11Pen
tanal
704
13.6
6ab
26.3
5ab
30.5
9ab
10.7
1b18
.75a
b10
.67b
8.47
b51
.35a
21.3
8ab
17.9
5ab
22.9
1ab
16.6
6ab
12D
imet
hyl
disu
lphi
de
749
1.75
b1.
09b
2.66
b1.
12b
2.28
b2.
21b
1.11
b2.
58b
1.17
b2.
24b
5.88
a1.
04b
132-
Hex
anone
791
0.36
ab0.
81a
0.40
ab0.
12b
0.29
ab0.
11b
0.14
b0.
35ab
0.58
ab0.
60ab
0.31
ab0.
35ab
14H
exan
al80
34.
14b
13.7
7a6.
12b
3.66
b3.
55b
2.33
b2.
73b
4.11
b3.
88b
8.88
ab5.
65b
4.60
b
152-
Hep
tanone
889
3.60
c3.
24c
1.84
c15
2.81
b11
.47c
272.
78a
2.69
c7.
93c
3.71
c3.
88c
95.4
5b10
8.35
b
16H
epta
nal
901
3.91
b21
.47a
7.20
b5.
39b
3.09
b4.
75b
3.19
b4.
35b
4.89
b10
.28b
6.13
b5.
71b
17O
ctan
al10
005.
67b
125.
39a
18.2
4b12
.91b
7.94
b11
.93b
10.5
0b11
.95b
12.7
5b44
.24b
16.1
4b10
.99b
182-
Nona
none
1087
1.41
d1.
64d
4.42
d13
6.75
b22
.83c
d27
1.90
a2.
92d
27.5
0cd
2.53
d3.
65d
86.0
7bc
69.6
9cd
19N
ona
nal
1102
16.6
6a58
5.81
a97
.33a
62.5
7a30
.23a
70.5
0a40
.57a
67.1
5a46
.44a
196.
93a
68.9
5a48
.59a
20D
ecan
al12
0817
.14b
725.
97a
78.5
3b91
.42b
31.4
7b77
.15b
29.1
5b70
.82b
87.0
0b19
4.88
ab12
1.21
b48
.02b
R.I."
Ret
ention
inde
xon
DB-5
ms
colu
mn
Mea
ns
follo
wed
bydi!
eren
tle
tter
sin
each
row
are
sign
i"ca
ntly
di!
eren
tat
the
0.05
leve
lM
M41
0,¸ac
toco
ccus
lact
isss
pla
ctis;C
NR
Z32
,¸ac
toba
cillu
she
lvet
icus
;LH
212,
¸ac
toba
cillu
she
lvet
icus
;LC
301,
¸ac
toba
cilu
sca
sei;
L23
,¸ac
toba
cillu
spe
ntos
usan
dL71
,¸ac
toba
cillu
spa
raca
sei
lwt/vol. 32 (1999) No. 7
434

lwt/vol. 32 (1999) No. 7
J. Broadbent (Utah State University, Logan, UT, U.S.A.).¸actobacillus helveticus CNRZ32 was obtained from J.Steele (University of Wisconsin, Madison, WI, U.S.A.).¸actobacillus helveticus LH212 was obtained from Rho-dia, Inc. (Madison, WI, U.S.A.). The lactobacilli selectedhad been previously characterized in cheese. CNRZ32and LH212 are considered adjuncts that promote desir-able #avours in cheese, while LC301 is an adjunct thatpromotes poor #avour quality in cheese (13). AdjunctsL23 and L71 were isolated from poor quality (unclean-#avoured) Cheddar cheese. The lactobacilli were grownin MRS broth at 373C. All strains were frozen in300 mL/L glycerol and stored at !70 3C until use.Stock cultures were thawed and grown in their respectivemedia at their respective temperatures (37 3C for lac-tobacilli and 30 3C for MM410) for two transfers prior toinoculation into milk for testing. A 10 mL/L inoculum ofa 16 h broth culture was transferred into a sterile recon-stituted skim milk blank for all experiments. For indi-vidual strain analysis, skim milk cultures were incubatedat the respective temperatures. For analysis of lac-tobacilli with ¸. lactis ssp lactis MM410, milk cultureswere incubated at 30 3C. Duplicates of inoculated skimmilk were prepared for all analyses. Acid production ofeach strain was evaluated by pH and titratable acidity.
ProteolysisFollowing inoculation into skim milk, proteolysis wasdetermined every 8 h through a 48 h incubation. Pro-teolysis was monitored using a lithium ninhydrin assay.The assay was conducted by modifying the procedure ofPearce et al. (14). The sample size was increased to 10 gand the sodium citrate volume was increased to 100 mLto facilitate analysis. Uninoculated milks served as thecontrols. Duplicate analyses of each timepoint were per-formed from duplicate skim milk samples. Absorbanceswere converted to moles amino groups per mole casein asdescribed by Pearce et al. (14) and expressed as thedi!erence between the sample and the control (*NH2).
Bacterial counts of fermented milksLactococci and lactobacilli were enumerated at eachtimepoint by dilution pourplating in triplicate. Lac-tococci were counted following incubation for 48 h at30 3C on Elliker's agar with overlay. Lactobacilli werecounted following incubation for 48 h at 37 3C on MRSagar with overlay When lactococci and lactobacilli wereboth added to milks, the lactobacilli were enumerated byincubation for 48 h at 37 3C on LBS agar with overlay.The lactococci counts were estimated by counting colo-nies formed after incubation for 72 h at 30 3C on Elliker'sagar with overlay and subtracting the number of lac-tobacilli from the LBS counts.
Sensory evaluationAroma-pro"ling of the milks following 24 h fermentationwas conducted by seven faculty and sta! memberswho were experienced in both sensory evaluation and
43
aroma-pro"ling. Sensory evaluation of aroma was con-ducted using a modi"ed Flavor Pro"leTM technique (15).Each panelist was provided with 50 mL of the milk in a150-mL glass beaker covered with aluminum foil. Thepanelists sni!ed the sample headspace and discussed anddetermined primary aroma attributes of each milk. Theintensity of each attribute, including overall aroma inten-sity was also determined as mild, medium or intense. Thisprocess was repeated twice to determine a representativeconsensus aroma-pro"le for each strain of milk.
Dynamic headspace sampling gas chromatography massspectroscopyA Tekmar 3000 Purge and Trap Concentrator/Cryofocusing Module (Tekmar, Co., Cinncinnati, OH,U.S.A.) coupled with an HP5890 series II GC/5972 massselective detector (MSD, Hewlett-Packard, Co., PaloAlto, CA, U.S.A.) was employed for dynamic headspacesampling gas chromatography mass spectrometry (DHS-GC-MS). Separations were performed on fused silicacapillary column (DB}5 ms, 60 m length]0.25 mmi.d.]0.25 km; J & W Scienti"c, Folsom, CA, U.S.A.). Thecarrier gas was helium at a constant #ow of 0.96 mL/min.The oven temperature was programmed from 40 to2003C at a rate of 63C/min with initial and "nal holdtimes of 5 and 60 min, respectively. MSD conditions wereas follows: the capillary direct interface temperature was2803C; the ionization energy was 70 eV; the mass rangewas 33}50 a.m.u.; the EM voltage was Atune#200 V;and the scan rate was 2.2 scan/s.A 5 mL aliquot of milk in a 25-mL standard purge tubewas prepurged with helium (40 mL/min) for 2 min andthe sample was then preheated to 60 3C for 5 min. Thevolatiles were swept from the sample headspace at 60 3Cfor 20 min on to a Tenax TA trap (part no. 12-0083-303,Tekmar, Co.) maintained at 0 3C. After sampling, the trapwas dry-purged for 5 min and the volatiles were desorbed(1803C for 1 min) and subsequently cryofocused(!120 3C) on to a 15 cm section of a 0.53 mm i.d. deac-tivated fused silica capillary column for GC-MS. Thetransfer lines and valves were maintained at a temper-ature of 150 3C and the trap pressure control was set at20 kPa. Helium #ow during thermal desorption of theTenax trap (20 mL/min) and the cryofocusing trap(1.4 mL/min) was controlled by the split/splitless elec-tronic pressure control pneumatics of the GC. Betweeneach analysis, the system was purged and the Tenax TAtrap was subsequently baked at 225 3C for 15 min.
Quantitative analysis by DHS-GC-MSThe milk blanks or fermented milks were added to a100-mL volumetric #ask and spiked with 10 kL of internalstandard solution (50 kL/mL) using a syringe. The solu-tions were shaken vigorously for 5 min and then analysedin duplicate by DHS-GC-MS. Calibration was accomp-lished by addition of 0 (blank) or 10 kL from threeconcentration levels (level 1"stock; level 2"1/5 dilu-tion of stock; level 3"1/25 dilution of stock). The stan-dard stock solution contained 1351 kg of no. 1, 313 kg of
5
lwt/vol. 32 (1999) No. 7
no. 2, 1210 kg of no. 3, 78.2 kg of no. 4, 1495 kg of no. 5,29.1 kg of no. 6, 283 kg of no. 7, 128 kg of no. 8, 32.9 kg ofno. 9, 393 kg of no. 10, 185 kg of no. 11, 168 kg of 12,21.6 kg of no. 13, 92.5 kg of no. 14, 2948 kg of no. 15,179 kg of no. 16, 532 kg of no. 17, 1655 kg of no. 18,1617 kg of no. 19 and 854 kg of no. 20 per L in methanoladded to 100 mL aliquots of milk prior to analysis. Thesolutions for internal standardization were prepared byspiking three 100 mL aliquots of milk with 10 kL ofinternal standard solution and then spiking the respect-ive milks with 10 kL of the appropriate dilution of stan-dard stock solution.
Statistical analysisThe data were analysed using Sigma Plot (SPSS, Inc.,Chicago, IL, U.S.A.) and SAS (1988). The signi"cancewas established at P(0.05. An analysis of variance withDuncan's multiple range test was used to determine sig-ni"cant di!erences between means of headspace volatilesand proteolysis data. A principal components analysis(PCA) was calculated to determine whether culturescould be clustered or grouped based on volatile produc-tion, proteolysis and acid production.
Results and Discussion
From more than 70 volatile peaks, 20 compounds wereselected for quanti"cation further study (Table 1). Thesecompounds were selected on the basis of their concentra-tion in the milks and on perceived importance in cheese#avour (16). All the compounds studied had been pre-viously found in fermented milk (8, 17). The volatile pro-"les di!ered widely among the lactobacilli. LH212 andCNRZ32 produced large amounts of the methyl ketones,2-heptanone and 2-nonanone. Although the signi"canceof methyl ketones in Cheddar cheese #avour has not been
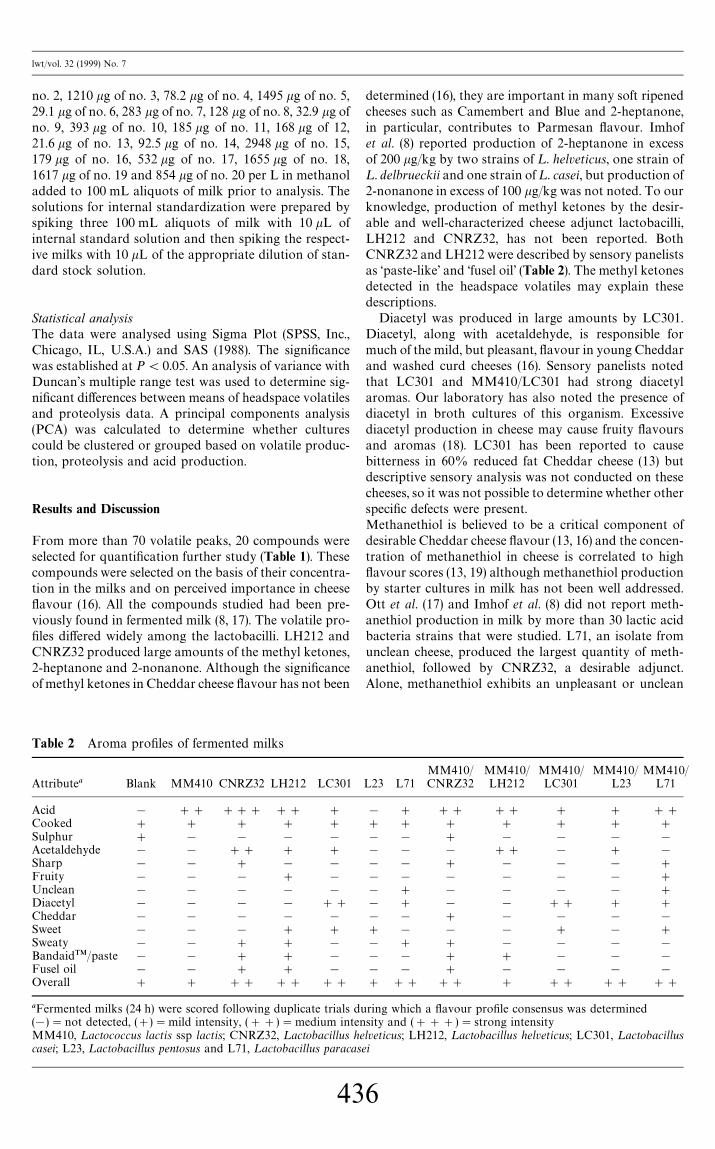
Table 2 Aroma pro"les of fermented milks
Attributea Blank MM410 CNRZ32 LH212 LC301
Acid ! ## ### ## #
Cooked # # # # #
Sulphur # ! ! ! !
Acetaldehyde ! ! ## # #
Sharp ! ! # ! !
Fruity ! ! ! # !
Unclean ! ! ! ! !
Diacetyl ! ! ! ! ##
Cheddar ! ! ! ! !
Sweet ! ! ! # #
Sweaty ! ! # # !
BandaidTM/paste ! ! # # !
Fusel oil ! ! # # !
Overall # # ## ## ##
aFermented milks (24 h) were scored following duplicate trials du(!)"not detected, (#)"mild intensity, (##)"medium intenMM410, ¸actococcus lactis ssp lactis; CNRZ32, ¸actobacillus hecasei; L23, ¸actobacillus pentosus and L71, ¸actobacillus paracas
43
determined (16), they are important in many soft ripenedcheeses such as Camembert and Blue and 2-heptanone,in particular, contributes to Parmesan #avour. Imhofet al. (8) reported production of 2-heptanone in excessof 200 kg/kg by two strains of ¸. helveticus, one strain of¸. delbrueckii and one strain of ¸. casei, but production of2-nonanone in excess of 100 kg/kg was not noted. To ourknowledge, production of methyl ketones by the desir-able and well-characterized cheese adjunct lactobacilli,LH212 and CNRZ32, has not been reported. BothCNRZ32 and LH212 were described by sensory panelistsas &paste-like' and &fusel oil' (Table 2). The methyl ketonesdetected in the headspace volatiles may explain thesedescriptions.
Diacetyl was produced in large amounts by LC301.Diacetyl, along with acetaldehyde, is responsible formuch of the mild, but pleasant, #avour in young Cheddarand washed curd cheeses (16). Sensory panelists notedthat LC301 and MM410/LC301 had strong diacetylaromas. Our laboratory has also noted the presence ofdiacetyl in broth cultures of this organism. Excessivediacetyl production in cheese may cause fruity #avoursand aromas (18). LC301 has been reported to causebitterness in 60% reduced fat Cheddar cheese (13) butdescriptive sensory analysis was not conducted on thesecheeses, so it was not possible to determine whether otherspeci"c defects were present.Methanethiol is believed to be a critical component ofdesirable Cheddar cheese #avour (13, 16) and the concen-tration of methanethiol in cheese is correlated to high#avour scores (13, 19) although methanethiol productionby starter cultures in milk has not been well addressed.Ott et al. (17) and Imhof et al. (8) did not report meth-anethiol production in milk by more than 30 lactic acidbacteria strains that were studied. L71, an isolate fromunclean cheese, produced the largest quantity of meth-anethiol, followed by CNRZ32, a desirable adjunct.Alone, methanethiol exhibits an unpleasant or unclean
L23 L71MM410/CNRZ32
MM410/LH212
MM410/LC301
MM410/L23
MM410/L71
! # ## ## # # ##
# # # # # # #
! ! # ! ! ! !
! ! ! ## ! # !
! ! # ! ! ! #
! ! ! ! ! ! #
! # ! ! ! ! #
! # ! ! ## # #
! ! # ! ! ! !
# ! ! ! # ! #
! # # ! ! ! !
! ! # # ! ! !
! ! # ! ! ! !
# ## ## # ## ## ##
ring which a #avour pro"le consensus was determinedsity and (###)"strong intensitylveticus; LH212, ¸actobacillus helveticus; LC301, ¸actobacillusei
6
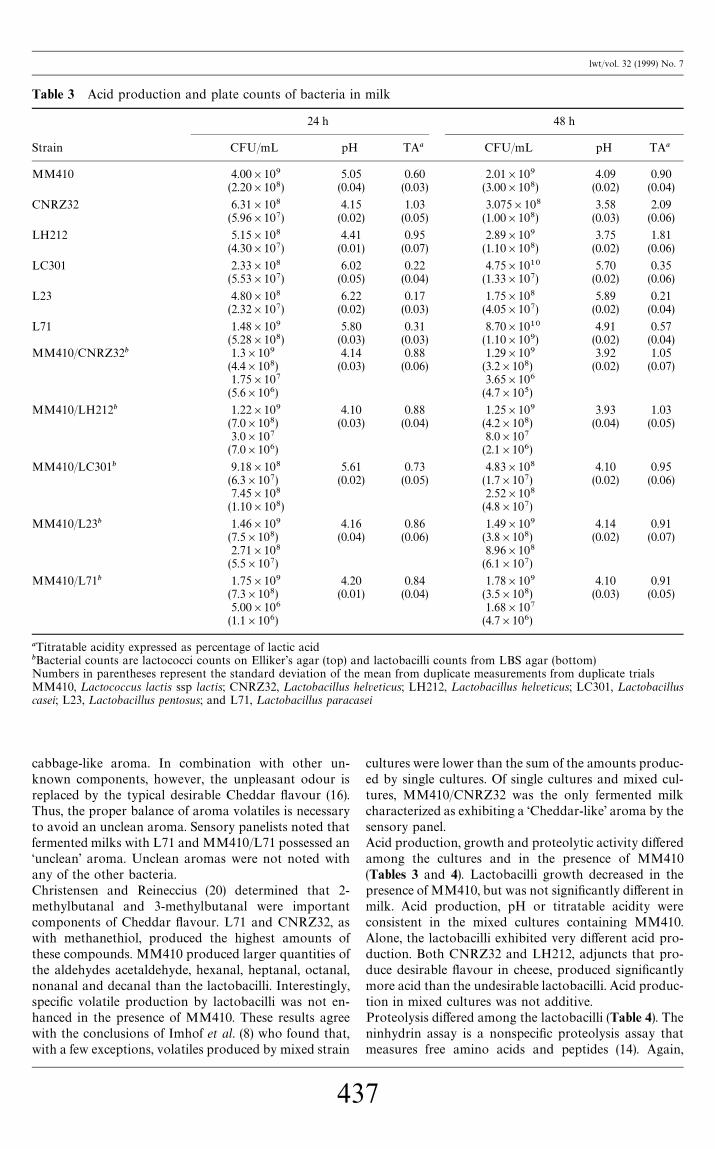
Table 3 Acid production and plate counts of bacteria in milk
24 h 48 h
Strain CFU/mL pH TAa CFU/mL pH TAa
MM410 4.00]109 5.05 0.60 2.01]109 4.09 0.90(2.20]108) (0.04) (0.03) (3.00]108) (0.02) (0.04)
CNRZ32 6.31]108 4.15 1.03 3.075]108 3.58 2.09(5.96]107) (0.02) (0.05) (1.00]108) (0.03) (0.06)
LH212 5.15]108 4.41 0.95 2.89]109 3.75 1.81(4.30]107) (0.01) (0.07) (1.10]108) (0.02) (0.06)
LC301 2.33]108 6.02 0.22 4.75]1010 5.70 0.35(5.53]107) (0.05) (0.04) (1.33]107) (0.02) (0.06)
L23 4.80]108 6.22 0.17 1.75]108 5.89 0.21(2.32]107) (0.02) (0.03) (4.05]107) (0.02) (0.04)
L71 1.48]109 5.80 0.31 8.70]1010 4.91 0.57(5.28]108) (0.03) (0.03) (1.10]109) (0.02) (0.04)
MM410/CNRZ32b 1.3]109 4.14 0.88 1.29]109 3.92 1.05(4.4]108) (0.03) (0.06) (3.2]108) (0.02) (0.07)1.75]107 3.65]106(5.6]106) (4.7]105)
MM410/LH212b 1.22]109 4.10 0.88 1.25]109 3.93 1.03(7.0]108) (0.03) (0.04) (4.2]108) (0.04) (0.05)3.0]107 8.0]107(7.0]106) (2.1]106)
MM410/LC301b 9.18]108 5.61 0.73 4.83]108 4.10 0.95(6.3]107) (0.02) (0.05) (1.7]107) (0.02) (0.06)7.45]108 2.52]108(1.10]108) (4.8]107)
MM410/L23b 1.46]109 4.16 0.86 1.49]109 4.14 0.91(7.5]108) (0.04) (0.06) (3.8]108) (0.02) (0.07)2.71]108 8.96]108(5.5]107) (6.1]107)
MM410/L71b 1.75]109 4.20 0.84 1.78]109 4.10 0.91(7.3]108) (0.01) (0.04) (3.5]108) (0.03) (0.05)5.00]106 1.68]107(1.1]106) (4.7]106)
aTitratable acidity expressed as percentage of lactic acidbBacterial counts are lactococci counts on Elliker's agar (top) and lactobacilli counts from LBS agar (bottom)Numbers in parentheses represent the standard deviation of the mean from duplicate measurements from duplicate trialsMM410, ¸actococcus lactis ssp lactis; CNRZ32, ¸actobacillus helveticus; LH212, ¸actobacillus helveticus; LC301, ¸actobacilluscasei; L23, ¸actobacillus pentosus; and L71, ¸actobacillus paracasei
lwt/vol. 32 (1999) No. 7
cabbage-like aroma. In combination with other un-known components, however, the unpleasant odour isreplaced by the typical desirable Cheddar #avour (16).Thus, the proper balance of aroma volatiles is necessaryto avoid an unclean aroma. Sensory panelists noted thatfermented milks with L71 and MM410/L71 possessed an&unclean' aroma. Unclean aromas were not noted withany of the other bacteria.Christensen and Reineccius (20) determined that 2-methylbutanal and 3-methylbutanal were importantcomponents of Cheddar #avour. L71 and CNRZ32, aswith methanethiol, produced the highest amounts ofthese compounds. MM410 produced larger quantities ofthe aldehydes acetaldehyde, hexanal, heptanal, octanal,nonanal and decanal than the lactobacilli. Interestingly,speci"c volatile production by lactobacilli was not en-hanced in the presence of MM410. These results agreewith the conclusions of Imhof et al. (8) who found that,with a few exceptions, volatiles produced by mixed strain
43
cultures were lower than the sum of the amounts produc-ed by single cultures. Of single cultures and mixed cul-tures, MM410/CNRZ32 was the only fermented milkcharacterized as exhibiting a &Cheddar-like' aroma by thesensory panel.Acid production, growth and proteolytic activity di!eredamong the cultures and in the presence of MM410(Tables 3 and 4). Lactobacilli growth decreased in thepresence of MM410, but was not signi"cantly di!erent inmilk. Acid production, pH or titratable acidity wereconsistent in the mixed cultures containing MM410.Alone, the lactobacilli exhibited very di!erent acid pro-duction. Both CNRZ32 and LH212, adjuncts that pro-duce desirable #avour in cheese, produced signi"cantlymore acid than the undesirable lactobacilli. Acid produc-tion in mixed cultures was not additive.Proteolysis di!ered among the lactobacilli (Table 4). Theninhydrin assay is a nonspeci"c proteolysis assay thatmeasures free amino acids and peptides (14). Again,
7

Table 4 Mean proteolysis of strains in milk
Strain 24 h 48 h
MM410 0.345e$0.08 1.085e$0.11CNRZ32 1.07b$0.10 2.49c$0.13LH212 1.16b$0.09 5.54a$0.13LC301 0.605c$0.07 0.661f$0.13L23 0.220e$0.09 0.448f$0.10L71 0.870c$0.05 1.00c$0.08MM410/CNRZ32 1.81a$0.11 2.62c$0.08MM410/LH212 1.12b$0.07 3.02b$0.12MM410/LC301 0.210e$0.05 1.67d$0.08MM410/L23 0.224e$0.14 0.950e$0.11MM410/L71 0.580d$0.07 1.52d$0.13
Di!erence from a milk blank (control) expressed as molesamino group per mole caseinThe means followed by di!erent letters in each column aresigni"cantly di!erent at P40.05The mean values are from four replicates (duplicate assays fromduplicate trials)MM410, ¸actococcus lactis ssp lactis; CNRZ32, ¸actobacillushelveticus; LH212, ¸actobacillus helveticus; LC301, ¸actobacil-lus casei; L23, ¸actobacillus pentosus; and L71, ¸actobacillusparacasei
lwt/vol. 32 (1999) No. 7
CNRZ32 and LH212, the desirable adjuncts, exhibitedsigni"cantly higher protease activity than the other lac-tobacilli. Proteolysis was not enhanced in mixed cultures.Proteinases and peptidases are important, but little isknown about the role of individual cellular enzymes ortheir overall potentially complex role in #avour develop-ment. Lynch et al. (21) studied lactobacillus strains iso-lated from strong-#avoured cheese and concluded thatadjunct lactobacilli produced peptides that contributedto and intensi"ed cheese #avour. Drake et al. (7) studiedan adjunct lactobacillus that improved reduced fat Ched-dar cheese and reported that rates of proteolysis wereaccelerated in these cheeses compared to a control. WhileWeimer et al. (13) concluded that aminopeptidase activ-ity of adjunct lactobacilli was not correlated with im-proved cheese #avour, evidence from other studies (7, 22)suggested that overall proteolytic activity may be a ne-cessary trait for a desirable cheese adjunct.Overall, both desirable adjuncts, CNRZ32 and LH212,produced larger amounts of methyl ketones and exhib-ited greater proteolytic activity than the undesirablecheese adjuncts, LC301, L71 and L23. There were noapparent similarities among the undesirable cheese ad-juncts. The lack of a relationship among undesirableadjuncts is not surprising as poorly #avoured cheese maybe due to a variety of volatile organic compounds. It wasnot possible to group desirable and undesirable adjunctlactobacilli by principal components analysis based onvolatile production, acid production or proteolytic activ-ity in milk (data not shown). These results are similarto those of Imhof et al. (8) who observed that lactic acidbacteria could not be grouped by species based onvolatile production.Speci"c proteinase activity and volatile production canbe a!ected by salt and pH (13). Volatiles and proteolyticactivity more indicative of behaviour in cheese may beproduced in milk with an acidic pH and added salt(pH 5.5, 50 g/L salt). Studies of these cultures under
43
cheese-like conditions in milk may provide additionalinformation that could rapidly identify key character-istics of desirable and undesirable lactobacilli without theneed for cheese slurry studies.
Acknowledgements
This research was funded by the Mississippi State Agri-culture and Forestry Experiment Station (MAFES) atMississippi State University. This paper is journal articlenumber 9524 of MAFES, Mississippi State University.
References
1 ROMERO, D. Bacteria as potential sources of #avour metab-olites. Food ¹echnology, 46, 122}126 (1992)
2 AMANTEA, G. F., SKURA, B. J. AND NAKAI, S. Culture e!ecton ripening characteristics and rheological behavior ofCheddar cheese. Journal of Food Science, 51, 912}918 (1986)
3 STEEL, J. L. AND UNLU, G. Impact of lactic acid bacteria oncheese #avor development. Food ¹echnology, 46, 128}136(1992)
4 FOX, P. F., LAW, J., MCSWEENEY, P. L. H. AND WALLACE, J.Biochemistry of cheese ripening. In: FOX, P. F. (Ed.), Cheese:Chemistry, Physics, and Microbiology, <ol. 1. New York:Chapman & Hall, pp. 103}146 (1993)
5 BARLOW, I., LLOYD, G. T., RAMSHAW, E. H., MILLER, A. J.,MCCABE, G. P. AND MCCABE, L. Correlations and changesin #avour and chemical parameters of Cheddar cheesesduring maturation. Australian Journal of Dairy ¹echnology,44, 7}18 (1989)
6 DACREMONT, C. AND VICKERS, Z. Concept matching tech-nique for assessing the importance of volatile compoundsfor cheese aroma. Journal of Food Science, 59, 981}985(1994)
7 DRAKE, M. A., BOYLSTON, T. D., SPENCE, K. D. AND
SWANSON, B. G. Improvement of sensory quality of reducedfat Cheddar cheese by a ¸actobacillus adjunct. Food Re-search International, 30, 35}40 (1997)
8 IMHOF, R., GLATTLI, H. AND BOSSET, J. O. Volatile organiccompounds produced by thermophilic and mesophilicsingle dairy starter cultures. ¸ebensmittel-=issenschaft und-¹echnologie, 28, 78}86 (1995)
9 WOODILL, J. D.;se of adjunct cultures and elevated ripeningtemperatures in reduced fat Edam cheese manufacture.Masters thesis, Mississippi State University, MS, U.S.A.(1998)
10 JENSEN, K. N. ;se of adjunct cultures in reduced fatJarlsberg-type cheese. Masters thesis, Mississippi StateUniversity, MS, U.S.A. (1998)
11 MUEHLENKAMP, M. R. AND WARTHESON, J. J. The in#u-ence of non-starter lactobacilli on proteolysis in Cheddarslurries and cheeses during ripening. Abstract 81ADSA}ASAS Joint Meeting, Denver, CO, U.S.A. July27}31 (1998)
12 LAW, B. A. Microbiology and Biochemistry of Cheese andFermented Milk. London, U.K.: Chapman and Hall (1997)
13 WEIMER, B., DIAS, B., UMMADI, M., BROADBENT, J., BREN-
NAND, C., JAEGI, J., JOHNSON, M., MILANI, F., STEELE, J.and SISSON, D. V. In#uence of NaCl and pH on intracellu-lar enzymes that in#uence Cheddar cheese ripening. ¸ait,77, 383}398 (1997)
14 PEARCE, K. N., KARAHALIOS, D. AND FRIEDMAN, M. Nin-hydrin assay for proteolysis in ripening cheese. Journal ofFood Science, 53, 432}435, 438 (1988)
15 LAWLESS, H. T. AND HEYMANN, H. Descriptive analysis. In:Sensory Evaluation of Food. New York, U.S.A.: Chapmanand Hall, pp. 341}372 (1997)
8

lwt/vol. 32 (1999) No. 7
16 URBACH, G. The #avour of milk and dairy products: II.Cheese: contribution of volatile compounds. InternationalJournal of Dairy ¹echnology, 50, 79}89 (1997)
17 OTT, A., FAY, L. B. AND CHAINTREAU, A. Determination andorigin of the aroma impact compounds of yogurt #avor.Journal of Agricultural Food Chemistry, 45, 850}858 (1997)
18 BODYFELT, F. W., TOBIAS, J. AND TROUT, G. M. Sensoryevaluation of cheese. Ch. 8. In: ¹he Sensory Evaluation ofDairy Products. New York: U.S.A.: AVI Van NostrandReinhold, pp. 300}376 (1988)
19 DIMAS, A., URBACH, G. E. AND MILLER, A. J. Changes in#avor and volatiles of the full-fat and low-fat cheesesduring maturation. International Dairy Journal, 6,981}995 (1996).
43
20 CHRISTENSEN, K. R. AND REINECCIUS, G. A. Aroma extractdilution analysis of aged Cheddar. Journal of Food Science,60, 218}220 (1995)
21 LYNCH, C. M., MCSWEENEY, P. L. H., FOX, P. F., COGAN,T. M. AND DRINAN, F. D. Contribution of starter lactococciand non-starter lactobacilli to proteolysis in Cheddarcheese with a controlled micro#ora. ¸ait, 77, 441}459(1997)
22 JOHNSON, J. A. C., ETZEL, M. R., CHEN, C. M. AND
JOHNSON, M. E. Accelerated ripening of reduced-fat Ched-dar cheese using four attenuated ¸actobacillus helveticusCNRZ-32 adjuncts. Journal of Dairy Science, 78, 769}776(1995)
9