aerotactic response azospirillum brasilense · five strains of azospirillum brasilense and two of...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, Nov. 1982, P. 643-6490021-9193/82/110643-07$02.00/0Copyright 6 1982, American Society for Microbiology
Vol. 152, No. 2
Aerotactic Response of Azospirillum brasilenseRINA BARAK, ISRAEL NUR, YAACOV OKON,* AND YIGAL HENIS
Department ofPlant Pathology and Microbiology, Faculty ofAgriculture, The Hebrew University ofJerusalem, Rehovot 76 100, Israel
Received 18 June 1982/Accepted 28 July 1982
Five strains of Azospirillum brasilense and two of Azospirillum spp., fromIsrael, responded to self-created and preformed oxygen gradients by formingaerotactic bands in capillary tubes and actively moving toward a specific zonewith low dissolved oxygen. Increasing the oxygen concentration in capillariescontaining phosphate buffer increased the number of attracted bacteria anddecreased band velocity. High 02 concentrations and H202 temporarily repulsedthe bacteria, causing the formation of a bacterial arc around the capillary mouth.There was no band formation under anaerobic conditions, although the bacteriaremained highly motile. Exogenous energy sources were unnecessary for aero-taxis in Azospirillum spp. The addition of oxidizable substrates to the capillaryslightly enhanced aerotaxis, possibly by accelerating 02 consumption. Aerotacticband formation was affected by pH, bacterial concentration and age, incubationtime, and respiratory inhibitors, but not by the lack of combined nitrogen in thegrowth medium. It is proposed that aerotaxis plays a role in the capacity ofAzospirillum spp. to reach an environment suitable for N2 fixation.
Attraction toward oxygen (aerotaxis), firstdemonstrated by Beijerinck (6), seems to exist ina wide variety of bacteria (5). Nevertheless, incontrast to extensive studies of chemotaxis,relatively few reports on aerotaxis have beenpublished since the turn of the century (1, 2, 4, 5,7, 12, 18). The phenomenon was examined most-ly by microscopic and macroscopic observationsand recently by temporal and spatial gradientassays (12).
Reports on the nitrogen-fixing bacteria, Azo-spirillum spp., indicate that this organism mighthave an aerotactic response. Nur et al. (14) andOkon et al. (16) found that Azospirillum cells,growing in test tubes in a semisolid medium,developed a narrow growth pellicle below thesurface. Increasing the agar concentrationcaused the pellicle to move closer to the air-water interface, thus indicating that Azospiril-lum cells seek low oxygen levels for growth. Apreliminary study of chemotaxis in Azospirillumbrasilense, with the capillary assay technique(3), showed attraction to solutions without oxi-dizable substrates, containing either phosphatebuffer or saline and also to distilled water (16).These findings suggest that Azospirillum cellsare attracted to the oxygen dissolved in water.We studied the aerotactic activity of A. brasi-
lense under various growth and incubation con-ditions by using microscopic observations andquantitative measurements in capillaries.
MATERIALS AND METHODSBacterial strains. A. brasilense ATCC 29729 (Cd),
isolated from roots of Cynodon dactylon (10), wasused in most experiments. It was compared withstrains Sp7, Spl3, Sp5le, Sp8l (9) and Cd-1 and Cd-3,isolated in Israel (14).Media and growth conditions. Batch cultures were
grown in 100-mi Erlenmeyer flasks, each containing 20ml of synthetic malate medium supplemented with0.05% NH4Cl and 0.01% yeast extract (17). The flaskswere inoculated with bacteria from a 24-h-old slantculture and incubated in a shaking bath (100 rpm) for20 h at 30°C. Unless otherwise stated, bacteria used inaerotaxis tests were grown in batch culture.Continuous cultures of Azospirillum spp. were
grown at 30°C in a New Brunswick chemostat (modelC 30) containing synthetic malate medium supplement-ed with 2 g of malic acid per liter. Dissolved 02 in thegrowth medium was measured with an autoclavablegalvanic type electrode (DO-81).Taxis medium, used for washing the cells and for
aerotaxis measurements, contained 60 mM potassiumbuffer (pH 6.8) supplemented with 100 FM Na-EDTA.
Aerotaxis asy. Azospirillum cultures, grown to anabsorbance of 0.1 to 0.4 at 420 nm (1 x 10' to 3.5 x 108colony-forming units per ml) were used. Before someexperiments, bacteria were centrifuged twice at 3,000x g for 10 min and resuspended in the taxis medium tothe initial turbidity. The aerotaxis experiments werecarried out by using the Adler capillary assay (3), withcapillary tubes (1 Jl; 0.2 mm internal diameter; Modu-lohm I/S. Copenhagen, Denmark) filled with the taxismedium. Unless otherwise stated, all experimentswere carried out for 45 min at 30°C in four replicates.
643
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

644 BARAK ET AL.
The number of bacteria accumulated in the capillarywas determined by diluting its contents in saline andplating portions on solid N-free synthetic medium (17).Band velocity was measured by placing a micrometricruler into the ocular piece of a microscope.Anaerobic aerotaxis assay. One vaccine bottle (21
ml) filled with 5 ml of a washed Azospirillum suspen-sion suspended in taxis medium and another filled with5 ml of the taxis medium were sealed with rubberstoppers and flushed for 30 min with O2-free nitrogen.Anaerobic conditions were confirmed by an oxygenelectrode. A hypodermic syringe (1 ml) was used totransfer the anaerobic suspension, under a continuousstream of nitrogen, into a 25-,lI capillary (ModulohmVS). The anaerobic taxis medium (dissolved 02, 0) wassimilarly transferred into a 1-,ul capillary. Both capil-laries were sealed with Cello-Seal (Fisher ScientificCo., Pittsburgh, Pa.). The open end of the small (1-lI)capillary was then inserted into the larger (25-pul) one,and the connection point was sealed with cello-seal.The same assay technique was used for the control,with an aerobic bacterial suspension and taxis medium(dissolved 02, 120 pM). The number of bacteria accu-mulated in the capillary was determined as describedfor the aerotaxis assay. No differences in bacterianumber in the capillary or band velocity and shapecould be detected when the modified method, underaerobic conditions, was compared with the Adlerassay.
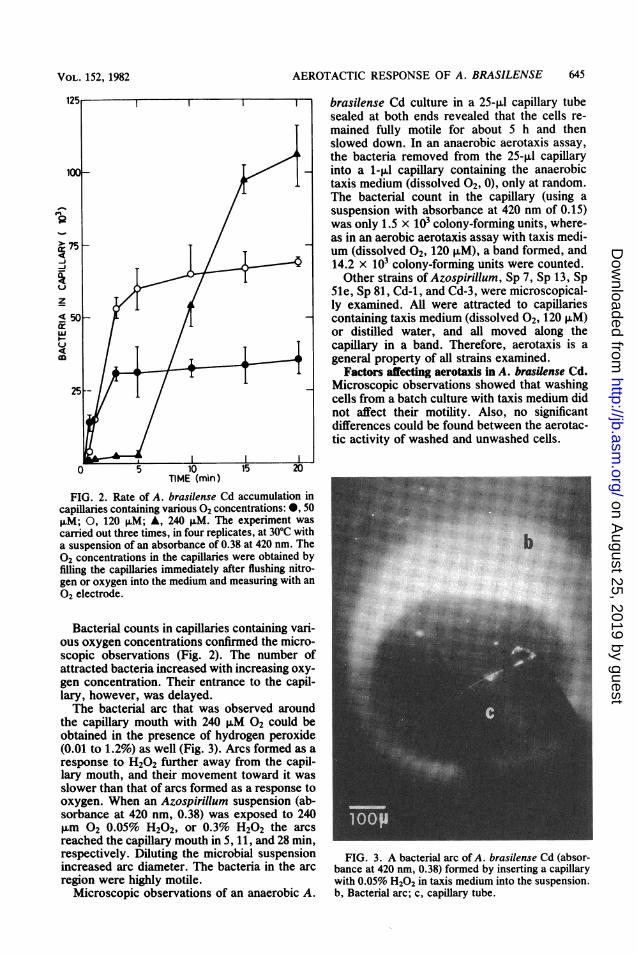
FIG. 1. An aerotactic band formed in an A. brasi-lense Cd suspension located in a chamber on a micro-scope slide (x250). ab, Aerotactic band; ag, aggre-gates; e, edge of the cover slip.
Oxygen uptake rate. The uptake rate of 02 in thebacterial suspension was measured by using an M 53chamber and an electrode (Yellow Spring InstrumentsCo., Yellow Springs, Ohio) and a U 125-M Unicorder(Japan). The reaction mixture contained 1 ml of potas-sium phosphate buffer (pH 7; final concentration, 60mM), 1 ml of MgSO4 * 7H20 (final concentration, 20mM), 0.2 ml of oxidizable substrate (final concentra-tion, 40 mM), and 1 ml of bacterial suspension. Mea-surements were carried out at 30°C for 10 min. Theuptake rates were expressed as microliters of 02 takenup per hour per milligram of bacterial dry weight.
RESULTSResponse of A. brasilense to dissolved oxygen
concentration. Microscopic observations of anA. brasilense Cd suspension placed in a micro-scopic chamber (3) revealed that in 1 to 4 min thebacteria migrated into a specific region, 0.5 to 2mm from the open edge of the chamber, forminga dense band parallel to it (Fig. 1). The bacteriathat did not arrive at the band aggregated. Theformation of aggregates was much faster on theside of the band open to the air than inside thesuspension. A similar band formed some dis-tance away from trapped air bubbles in thesuspension as well as in suspension smeared ona microscope slide under a cover slip. Flushingoxygen over the chamber caused the cells toform a new band farther away from the edge.Flushing nitrogen, on the other hand, caused anaccumulation of bacteria on the edge. About 5min after the oxygen or nitrogen flow was turnedoff, the band moved to its initial location.The response of A. brasilense Cd to dissolved
oxygen concentration was studied by using theAdler capillary assay (3). Capillaries were filledwith the taxis medium which had been flushedwith oxygen or nitrogen and measured with an02 electrode, thus obtaining three different con-centrations of dissolved oxygen. Each capillarywas then inserted into a microscopic chamberfilled with the Azospirillum suspension (absor-bance at 420 nm, 0.38). Microscopic observationshowed that exposing the culture to 50 ,uM 02caused a diffuse band of Azospirillum cells toenter the capillary after a lag period of 0.5 min atan initial velocity of 500 pum/min, which de-creased to 200 p.m/min within 30 min. The bandwas very diffuse at times, dispersing after a fewminutes. With 120 p.M 02, a large, dense aero-tactic band entered the capillary after a 1.5-minlag period at an initial velocity of 215 p.m/min,which decreased to 100 p.m/min after 30 min.Upon exposure to 240 p.M 02, the cells werefirst repulsed away from the capillary mouth,forming a bacterial arc around it. They thenmoved toward the capillary mouth, entering it ina very large band after a 5-min lag period. Theinitial velocity of 125 p.m/min decreased to 100p.m/min within 30 min.
J. BACTERIOL.
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
AEROTACTIC RESPONSE OF A. BRASILENSE 645
brasilense Cd culture in a 25-,ul capillary tubesealed at both ends revealed that the cells re-mained fully motile for about 5 h and thenslowed down. In an anaerobic aerotaxis assay,the bacteria removed from the 25-,ul capillaryinto a 1-,ul capillary containing the anaerobictaxis medium (dissolved 02, 0), only at random.The bacterial count in the capillary (using asuspension with absorbance at 420 nm of 0.15)was only 1.5 x 103 colony-forming units, where-as in an aerobic aerotaxis assay with taxis medi-um (dissolved 02, 120 F±M), a band formed, and14.2 x 103 colony-forming units were counted.Other strains of Azospirillum, Sp 7, Sp 13, Sp
51e, Sp 81, Cd-1, and Cd-3, were microscopical-ly examined. All were attracted to capillariescontaining taxis medium (dissolved 02, 120 ,uM)or distilled water, and all moved along thecapillary in a band. Therefore, aerotaxis is ageneral property of all strains examined.
Factors affecting aerotaxis in A. brasilense Cd.Microscopic observations showed that washingcells from a batch culture with taxis medium didnot affect their motility. Also, no significantdifferences could be found between the aerotac-tic activity of washed and unwashed cells.
TIME (min)
FIG. 2. Rate of A. brasilense Cd accumulation incapillaries containing various 02 concentrations: 0, 50,uM; 0, 120 ,uM; A, 240 ILM. The experiment wascarried out three times, in four replicates, at 30°C witha suspension of an absorbance of 0.38 at 420 nm. The02 concentrations in the capillaries were obtained byfilling the capillaries immediately after flushing nitro-gen or oxygen into the medium and measuring with an02 electrode.
Bacterial counts in capillaries containing vari-ous oxygen concentrations confirmed the micro-scopic observations (Fig. 2). The number ofattracted bacteria increased with increasing oxy-gen concentration. Their entrance to the capil-lary, however, was delayed.The bacterial arc that was observed around
the capillary mouth with 240 ,uM 02 could beobtained in the presence of hydrogen peroxide(0.01 to 1.2%) as well (Fig. 3). Arcs formed as aresponse to H202 further away from the capil-lary mouth, and their movement toward it wasslower than that of arcs formed as a response tooxygen. When an Azospirillum suspension (ab-sorbance at 420 nm, 0.38) was exposed to 240,um 02 0.05% H202, or 0.3% H202 the arcsreached the capillary mouth in 5, 11, and 28 min,respectively. Diluting the microbial suspensionincreased arc diameter. The bacteria in the arcregion were highly motile.
Microscopic observations of an anaerobic A.
FIG. 3. A bacterial arc of A. brasilense Cd (absor-bance at 420 nm, 0.38) formed by inserting a capillarywith 0.05% H202 in taxis medium into the suspension.b, Bacterial arc; c, capillary tube.
VOL. 152, 1982
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
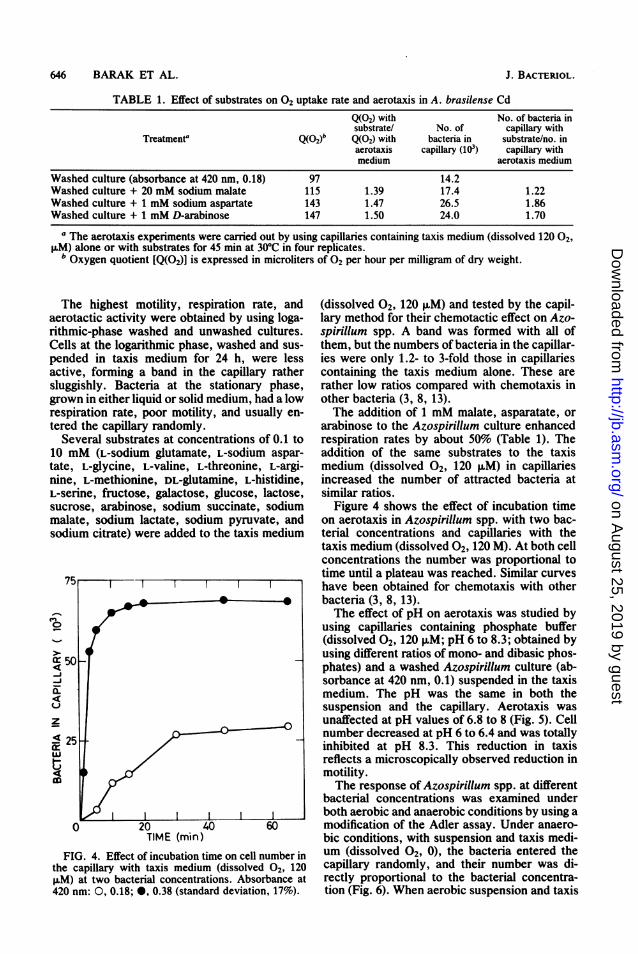
TABLE 1. Effect of substrates on 02 uptake rate and aerotaxis in A. brasilense Cd
Q(02) with No. of bacteria insubstrate/ No. of capillary with
Treatmenta Q(02)b Q(02) with bacteria in substrate/no. inaerotaxis capillary (103) capillary withmedium aerotaxis medium
Washed culture (absorbance at 420 nm, 0.18) 97 14.2Washed culture + 20 mM sodium malate 115 1.39 17.4 1.22Washed culture + 1 mM sodium aspartate 143 1.47 26.5 1.86Washed culture + 1 mM D-arabinose 147 1.50 24.0 1.70
a The aerotaxis experiments were carried out by using capillaries containing taxis medium (dissolved 120 02,,uM) alone or with substrates for 45 min at 30°C in four replicates.
b Oxygen quotient [Q(02)] is expressed in microliters of 02 per hour per milligram of dry weight.
The highest motility, respiration rate, andaerotactic activity were obtained by using loga-rithmic-phase washed and unwashed cultures.Cells at the logarithmic phase, washed and sus-pended in taxis medium for 24 h, were lessactive, forming a band in the capillary rathersluggishly. Bacteria at the stationary phase,grown in either liquid or solid medium, had a lowrespiration rate, poor motility, and usually en-tered the capillary randomly.
Several substrates at concentrations of 0.1 to10 mM (L-sodium glutamate, L-sodium aspar-tate, L-glycine, L-valine, L-threonine, L-argi-nine, L-methionine, DL-glutamine, L-histidine,L-serine, fructose, galactose, glucose, lactose,sucrose, arabinose, sodium succinate, sodiummalate, sodium lactate, sodium pyruvate, andsodium citrate) were added to the taxis medium
75 l l
cr _o-J-ja.(.)z
w
0 20 40 60TIME (min)
FIG. 4. Effect of incubation time on cell number inthe capillary with taxis medium (dissolved 02, 120,uM) at two bacterial concentrations. Absorbance at420 nm: 0, 0.18; 0, 0.38 (standard deviation, 17%).
(dissolved 02, 120 ,uM) and tested by the capil-lary method for their chemotactic effect on Azo-spirillum spp. A band was formed with all ofthem, but the numbers of bacteria in the capillar-ies were only 1.2- to 3-fold those in capillariescontaining the taxis medium alone. These arerather low ratios compared with chemotaxis inother bacteria (3, 8, 13).The addition of 1 mM malate, asparatate, or
arabinose to the Azospirillum culture enhancedrespiration rates by about 50% (Table 1). Theaddition of the same substrates to the taxismedium (dissolved 2, 120 ,uM) in capillariesincreased the number of attracted bacteria atsimilar ratios.
Figure 4 shows the effect of incubation timeon aerotaxis in Azospirillum spp. with two bac-terial concentrations and capillaries with thetaxis medium (dissolved 02, 120 M). At both cellconcentrations the number was proportional totime until a plateau was reached. Similar curveshave been obtained for chemotaxis with otherbacteria (3, 8, 13).The effect of pH on aerotaxis was studied by
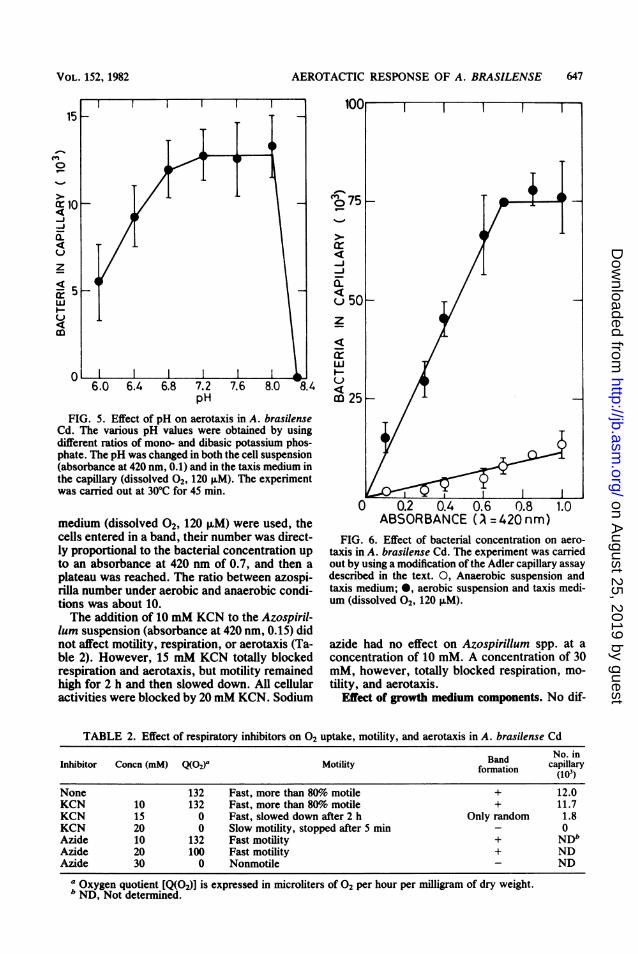
using capillaries containing phosphate buffer(dissolved 2, 120 ,uM; pH 6 to 8.3; obtained byusing different ratios of mono- and dibasic phos-phates) and a washed Azospirillum culture (ab-sorbance at 420 nm, 0.1) suspended in the taxismedium. The pH was the same in both thesuspension and the capillary. Aerotaxis wasunaffected at pH values of 6.8 to 8 (Fig. 5). Cellnumber decreased at pH 6 to 6.4 and was totallyinhibited at pH 8.3. This reduction in taxisreflects a microscopically observed reduction inmotility.The response of Azospirillum spp. at different
bacterial concentrations was examined underboth aerobic and anaerobic conditions by using amodification of the Adler assay. Under anaero-bic conditions, with suspension and taxis medi-um (dissolved 2, 0), the bacteria entered thecapillary randomly, and their number was di-rectly proportional to the bacterial concentra-tion (Fig. 6). When aerobic suspension and taxis
646 BARAK ET AL. J. BACTERIOL.
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
AEROTACTIC RESPONSE OF A. BRASILENSE 647
104t-J
z
w
co
06.0 6.4 6.8 7.2 7.6 8.0 8.4
pH
FIG. 5. Effect of pH on aerotaxis in A. brasilenseCd. The various pH values were obtained by usingdifferent ratios of mono- and dibasic potassium phos-phate. The pH was changed in both the cell suspension(absorbance at 420 nm, 0.1) and in the taxis medium inthe capillary (dissolved 02, 120 ,uM). The experimentwas carried out at 30°C for 45 min.
medium (dissolved 02, 120 ,uM) were used, thecells entered in a band, their number was direct-ly proportional to the bacterial concentration upto an absorbance at 420 nm of 0.7, and then aplateau was reached. The ratio between azospi-rilla number under aerobic and anaerobic condi-tions was about 10.The addition of 10 mM KCN to the Azospiril-
lum suspension (absorbance at 420 nm, 0.15) didnot affect motility, respiration, or aerotaxis (Ta-ble 2). However, 15 mM KCN totally blockedrespiration and aerotaxis, but motility remainedhigh for 2 h and then slowed down. All cellularactivities were blocked by 20 mM KCN. Sodium
0.2 0.4 0.6 0.8ABSORBANCE (A = 420 nm)
FIG. 6. Effect of bacterial concentration on aero-taxis in A. brasilense Cd. The experiment was carriedout by using a modification of the Adler capillary assaydescribed in the text. 0, Anaerobic suspension andtaxis medium; 0, aerobic suspension and taxis medi-um (dissolved 02, 120 ,uM).
azide had no effect on Azospirillum spp. at aconcentration of 10 mM. A concentration of 30mM, however, totally blocked respiration, mo-tility, and aerotaxis.
Effect of growth medium components. No dif-
TABLE 2. Effect of respiratory inhibitors on 02 uptake, motility, and aerotaxis in A. brasilense Cd
Band ~No. inInhibitor Concn (mM) Q(02)a Motility Ban capillaryformation (1O3)None 132 Fast, more than 80%o motile + 12.0KCN 10 132 Fast, more than 80%o motile + 11.7KCN 15 0 Fast, slowed down after 2 h Only random 1.8KCN 20 0 Slow motility, stopped after 5 min - 0Azide 10 132 Fast motility + NDbAzide 20 100 Fast motility + NDAzide 30 0 Nonmotile - ND
a Oxygen quotient [Q(02)] is expressed in microliters of 02 per hour per milligram of dry weight.b ND, Not determined.
VOL. 152, 1982
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

648 BARAK ET AL.
ferences in band shape and velocity could beobserved in aerotaxis tests of A. brasilense Cdpreviously grown in test tubes containing eitherN-free semisolid medium (17) or the same medi-um supplemented with NH4Cl. Both microscop-ic observations and bacterial counts showed thatexchanging malate in the growth medium forpyruvate, lactate, or succinate did not affectaerotaxis.
Cells taken from a continuous culture of A.brasilense Cd, growing under a dissolved oxy-gen concentration of 0.06 to 0.14 atm, respondedto oxygen in a manner similar to cells from abatch culture, by forming only one band in thecapillary. Azospirillum cultures growing at adissolved oxygen concentration of 0.03 atm re-sponded much faster to oxygen flushed into thesuspension or to H202 and formed two bands incapillaries containing the taxis medium (dis-solved 02, 120 ,uM). The second band movedslowly for some minutes and then stopped butdid not disperse. The reason for the appearanceof two bands is not clear; it might be that thethreshold response of the cells to oxygen waslower because they were adapted to a low oxy-gen concentration, enabling them to respond tothe residual oxygen behind the first band.
Bacteria grown in the chemostat at a dissolvedoxygen concentration of 0.17 to 0.18 atm be-haved similarly to bacteria from batch culturesfor the first 10 min after sampling, but no re-sponse toward oxygen could be detected after-ward. The oxygen uptake rate of the culture waszero, but despite the absence of respiratoryactivity, the cells remained highly motile forseveral hours. Respiration could be restored bythe addition of malate or succinate. Respiratoryactivity in washed bacteria from batch cultures,on the other hand, persisted for more than 24 h.The reason for this behavior of Azospirillumspp. under these conditions is as yet unknown.
DISCUSSIONThe rapid migration of azospirilla in a micro-
scopic chamber to a specific distance away fromthe air-water interface and the movement of theband under 02 or N2 flow show that the bacteriaactively seek microaerobic conditions. Nur et al.(14) reported that Azospirillum spp. inoculatedinto semisolid medium in test tubes did indeedprefer low 02 levels for growth and multiplica-tion. The dissolved 02 in the band area is veryclose to zero (I. Nur, Y. Okon, and Y. Henis,unpublished data). Another means of reachingmicroaerobic conditions is the formation of ag-gregates composed of the bacteria that did notarrive at the band. Okon et al. (17) and Doberei-ner and Day (9) reported that Azospirillum spp.required microaerobic conditions for N2 fixa-tion. The experiments in this work indicate that
the bacteria preferred low 02 even when thegrowth medium was supplemented with com-bined nitrogen and that this 02 level was reachedby aerotaxis.
Increasing the 02 concentration in capillariescontaining taxis medium delayed band forma-tion, reduced band speed, and increased thenumber of attracted bacteria. Also, no band wasformed under anaerobic conditions. Similar che-motactic responses of other bacteria to varyingattractant concentrations have been reported (3,11, 13). This supports our argument that thedissolved oxygen is the attractant.The formation of a bacterial arc around the
mouth of a capillary containing H202 seems tobe a repulsion from high levels of oxygen re-leased by the catalase activity of the bacteria,rather than from the peroxide itself, as similarbehavior was observed with high levels of oxy-gen. Similar chemotactic responses to high con-centrations of attractants have been reported inEscherichia coli (3).
Aerotaxis in Azospirillum spp. required highmotility and a high respiration rate, thus depend-ing strongly on factors affecting these activities:age of the culture, pH, and respiration inhibi-tors. The aerotactic activity of the cells was alsoaffected by incubation conditions, time, and bac-terial concentration, but it was unaffected byexchanging the carbon source in the growthmedium or by removing the combined nitrogenfrom it. KCN has been reported to inhibit aero-taxis in Salmonella typhimurium (18) and Spiril-lum volutans (7) although the effective dosevaried with the organism. Azide inhibited oxy-gen uptake in Salmonella sp. (18) but did notprevent aerotaxis. The KCN effect confirms thesuggestion of Taylor et al. (18) and Laszlo andTaylor (12) that the oxygen receptor for aero-taxis is the terminal oxidase.An exogenous source of energy was unneces-
sary for aerotaxis. Most bacteria examined byBaracchini and Sherris (5) and S. volutans (7)required oxidizable substrates for aerotaxis.Azospirillum spp., however, were capable ofutilizing an intracellular energy source, probablypoly-,-hydroxy butyrate. Okon et al. (15) founda poly-,3-hydroxy butyrate content of up to 30%of the dry weight of Azospirillum cells andshowed ,B-hydroxy butyrate dehydrogenase ac-tivity. The ability of Azospirillum spp. to re-spond in the absence of substrates allowed thisstudy of aerotaxis without masking the phenom-enon by chemotactic agents.The results presented in Table 1 suggest that
the accelerated respiration rates caused by thesubstrates led to increased oxygen gradientswhich in turn increased the number of attractedbacteria. However, it may also be a chemotacticeffect, partly masked by the aerotactic response
J. BACTERIOL.
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

AEROTACTIC RESPONSE OF A. BRASILENSE
triggered by the use of an endogenous source ofenergy. Nevertheless, chemotaxis in Azospiril-lum spp. could not be measured by the capillaryassay but could be examined by other methodsin which the aerotactic response was avoided(R. Barak, I. Nur, and Y. Okon, unpublishedresults).
Although respiration appears to be essentialfor aerotaxis, motility also occurred in the ab-sence of oxygen uptake. This conclusion isbased on the following observations. (i) KCN ata concentration high enough to totally blockrespiration (15 mM) did not affect cell motilityfor 2 h. (ii) In the anaerobic aerotaxis assay,bacteria preserved their motility for at least 5 h.(iii) In cells grown under high levels of 02 in thechemostat, respiration was blocked when thecells were drawn from the chemostat, whereasmotility was unaffected. Similar behavior wasobserved in S. volutans (7).
In this work, aerotactic activity was measuredfor the first time by using the quantitative capil-lary assay (3). The curves obtained by using thistechnique for the effect of pH, incubation time,and bacterial concentration on aerotaxis wereanalogous to those reported for the chemotacticresponse of other bacteria to organic compounds(3, 11, 13, 19).The anaerobic aerotaxis assay allowed a quan-
titative comparison between the numbers ofbacteria entering the capillary randomly andthose attracted to oxygen. This may be usefulfor the study of oxygen response in other orga-nisms with similar properties.The data presented in this work indicate that,
as in other bacteria (5), aerotaxis plays a role inthe capacity of Azospirillum spp. to reach anenvironment suitable for growth. It seems to bespecifically useful for the rhizosphere bacteria ofthe genus Azospirillum under N2-fixing condi-tions, when microaerobic conditions are essen-tial.
ACKNOWLEDGMENTSThis research was supported by grant 2476/81 from the
USA-Israel Binational Scientific Foundation, and grant 1-254-80 from the USA-Israel Binational Agriculture Research andDevelopment Fund (BARD).
LITERATURE CITED1. Adler, J. 1966. Chemotaxis in bacteria. Science 153:708-
716.2. Adler, J. 1966. Effect of amino acids and oxygen on
chemotaxis in Escherichia coli. J. Bacteriol. 92:121-129.3. Adler, J. 1973. A method for measuring chemotaxis and
use of the method to determine optimum conditions forchemotaxis by Escherichia coli. J. Gen. Microbiol. 74:77-91.
4. Armstrong, J. B., J. Adler, and M. M. Dahl. 1967. Non-chemotactic mutants of Escherichia coli. J. Bacteriol.93:390-398.
5. Barcchll, O., and J. C. Sherrls. 1959. The chemotacticeffect of oxygen on bacteria. J. Pathol. Bacteriol. 77:565-574.
6. BeUerlnck, M. W. 1893. Ueber Atmungfiguren beweg-licher Bakterien. Zentralbl. Bakteriol. Parasitenkd. Infe-kionskr. Hyg. Abt. 1 Orig. 14:827-845.
7. Caraway, B. H., and N. K. Krieg. 1974. Aerotaxis inSpirillum volutans. Can. J. Microbiol. 20:1367-1374.
8. Chet, I., Y. Zilbersteln, and Y. Henis. 1973. Chemotaxis ofPseudomonas lachrymans to plant extracts and to waterdroplets collected from leaf surfaces of resistant andsusceptible plants. Physiol. Plant Pathol. 3:473-479.
9. Dobereiner, J., and J. M. Day. 1975. Associative symbio-sis in tropical grasses: characterization of microorganismsand dinitrogen-fixing sites, p. 518-538. In W. E. Newtonand C. J. Nyman (ed.), Proceedings of the 1st Internation-al Symposium on Nitrogen Fixation. Washington StateUniversity Press, Pullman.
10. Eskew, D. L., D. D. Focht, and I. P. Ting. 1977. Nitrogenfixation, denitrification, and pleomorphic growth in ahighly pigmented Spirillum lipoferum. Appl. Environ.Microbiol. 34:582-585.
11. LaMarre, A. G., S. C. Straley, and S. F. Conti. 1977.Chemotaxis toward amino acids by Bdellovibrio bacterio-vorus. J. Bacteriol. 131:201-207.
12. Laszlo, D. J., and B. L. Taylor. 1981. Aerotaxis in Salmo-nella typhimurium: role of electron transport. J. Bacteriol.145:990-1001.
13. Moulton, R. C., and T. C. Montde. 1979. Chemotaxis byPseudomonas aeruginosa. J. Bacteriol. 137:274-280.
14. Nur, I., Y. Okon, and Y. Henas. 1980. Comparative studiesof nitrogen fixing bacteria associated with grasses in Israelwith Azospirillum brasilense. Can. J. Microbiol. 26:714-718.
15. Okon, Y., S. L. Albrecht, and R. H. Burris. 1976. Carbonand ammonia metabolism of Spirillum lipoferum. J. Bac-teriol. 128:592-597.
16. Okon, Y., L. Cakmakci, I. Nur, and I. Chet. 1980.Aerotaxis and chemotaxis of Azospirillum brasilense.Microb. Ecol. 6:277-280.
17. Okon, Y., J. P. Houchins, S. L. Albrecht, and R. H.Burris. 1977. Growth of Spirillum lipoferum at constantpartial pressures of oxygen, and the properties of itsnitrogenase in cell-free extracts. J. Gen. Microbiol. 98:87-93.
18. Taylor, B. L., J. B. MiWer, H. M. Warrick, and D. E.Koshland, Jr. 1979. Electron acceptor taxis and blue lighteffect on bacterial chemotaxis. J. Bacteriol. 140:567-573.
19. van der Drift, C., J. Dulverman, H. Bexkens, and A.Krljnen. 1975. Chemotaxis of a motile Streptococcustoward sugars and amino acids. J. Bacteriol. 124:1142-1147.
VOL. 152, 1982 649
on August 25, 2019 by guest
http://jb.asm.org/
Dow
nloaded from