uptake of a fluorescent marker in plant cells is sensitive to brefeldin a and wortmannin
TRANSCRIPT

The Plant Cell, Vol. 14, 71–86, January 2002, www.plantcell.org © 2002 American Society of Plant Biologists
Uptake of a Fluorescent Marker in Plant Cells Is Sensitive to Brefeldin A and Wortmannin
Neil Emans,
a,1
Sabine Zimmermann,
a
and Rainer Fischer
a,b
a
Institute for Molecular Biotechnology, Biology VII, Rheinisch-Westfälische Technische Hochschule Aachen, Worringerweg 1, D-52074 Aachen, Germany
b
Fraunhofer Institute for Molecular Biology and Applied Ecology, Grafschaft, Auf dem Aberg 1, D-57392 Schmallenberg, Germany
We assessed FM1-43 [
N
-(3-triethylammoniumpropyl)-4-(4-[dibutylamino]styryl)pyridinium dibromide] as a fluorescentendocytosis marker in intact, walled plant cells. At 4
�
C, FM1-43 stained the plasma membrane, and after 30 to 120 minof incubation at 26
�
C, FM1-43 labeled cytoplasmic vesicles and then the vacuole. Fluorimetric quantitation demon-strated dye uptake temperature sensitivity (
�
65% reduction at 16
�
C,
�
90% at 4
�
C). FM1-43 uptake in suspension cellswas stimulated more than twofold by brefeldin A and inhibited
�
0.4-fold by wortmannin. FM1-43 delivery to the vacuolewas largely inhibited by brefeldin A, although overall uptake was stimulated, and brefeldin A treatment caused the ac-cumulation of large prevacuolar endosomal vesicles heavily labeled with FM1-43. Three-dimensional time lapse imag-ing revealed that FM1-43–labeled vacuoles and vesicles are highly dynamic. Thus, FM1-43 serves as a fluorescentmarker for imaging and quantifying membrane endocytosis in intact plant cells.
INTRODUCTION
Endocytosis in animal cells begins with the budding of in-vaginated coated pits from the plasma membrane intocoated vesicles, which then uncoat and fuse with the en-docytic organelles (Gruenberg et al., 1989; Gruenberg andMaxfield, 1995). By this action, both membrane proteinsand fluid phase molecules are sequestered from the cell ex-terior into intracellular organelles, from where they can recy-cle to the plasma membrane or be transported to otherdestinations, such as the lysosomes. Thus, endocytosisserves to balance membrane secretion and to remove,transport, or degrade receptor ligand complexes from thecell surface (Goldstein et al., 1985). Although clathrin coat–dependent endocytosis is involved in receptor internaliza-tion, clathrin-independent endocytosis pathways also con-tribute to internalization. These include uptake via non-clathrin-coated caveolae (Anderson, 1998), macropinocyto-sis (Swanson, 1989; Hewlett et al., 1994), and phagocytosis(Aderem and Underhill, 1999).
Our understanding of animal cell endocytosis was ad-vanced by the use of fluorescent and electron-opaquemarkers that can be internalized (Salzman and Maxfield,1988, 1989; Griffiths et al., 1989). The use of fluid phase mark-
ers has permitted the development of assays that reconsti-tute endocytic vesicle fusion (Braell, 1987, 1992; Gruenberget al., 1989; Emans et al., 1995; Emans and Verkman, 1996)and led to the identification of many proteins that catalyzeand control fusion, such as the small GTPase rab5 (Gorvelet al., 1991) and its effector proteins (Christoforidis et al.,1999a, 1999b; McBride et al., 1999).
In plants, there is evidence that endocytosis is likely to occur(Low and Chandra, 1994; for review, see Battey et al., 1999;Marcote et al., 2000). Endocytosis is postulated to counterbal-ance membrane secretion (Samuels and Bialputra, 1990) andpermit cell volume to respond to changes in osmolality (Thiel etal., 1998; Kubitscheck et al., 2000). However, the study of plantcell endocytosis has been hampered by the scarcity of mark-ers that can be used in the presence of the plant cell wall(Low and Chandra, 1994; Buchanan et al., 2000).
The cell wall acts as a semipermeable barrier to the use oftracer molecules that could be used to label the fluid phaseof the plant endocytic pathway, essentially excluding pro-teins
�
25 kD (Carpita et al., 1979). As a result, the majorityof the evidence for endocytosis in plants has come from theuse of plant cell protoplasts (Fowke et al., 1991), in whichthe cell wall is removed enzymatically together with somecell surface proteins (Fowke and Gamborg, 1980), and thereare uncertainties regarding how closely protoplast mem-brane traffic may resemble that of intact cells.
Plants express much of the endocytic machinery identi-fied in animal cells, such as coated vesicles (Robinson et al.,1998; Pimpl et al., 2000) and clathrin heavy chains (Coleman
1
To whom correspondence should be addressed. E-mail [email protected]; fax 49-241-871062.Article, publication date, and citation information can be found atwww.plantcell.org/cgi/doi/10.1105/tpc.010339.

72 The Plant Cell
et al., 1987; Blackbourn and Jackson, 1996),
�
-adaptins(Holstein et al., 1994), and small
ras
-like
rab
family GTPases,which include homologs of
rab5
, which is known to beinvolved in animal cell endocytosis (Anuntalabhochai et al.,1991; Terryn et al., 1992; Borg et al., 1997). Arguments havebeen put forward that the general machinery used in en-docytosis is conserved from animals to plants, althoughspecific endocytic proteins may have different functionsfrom one kingdom to another (Marcote et al., 2000).
Using protoplasts and electron microscopy, clathrin-coated pits and clathrin-coated vesicles similar to those foundin animals have been observed (Emons and Traas, 1986;Galway et al., 1993), and their number increases with cellgrowth (Samuels and Bialputra, 1990). Endocytosed mark-ers are proposed to enter plant cells via coated pit invagina-tion and budding into the cytoplasm (Marcote et al., 2000).Electrophysiological studies indicate that early endocytic vesi-cles have a diameter between 70 and 100 nm (Thiel et al.,1998), and ultrastructural analysis has shown that the en-docytic pathway, as in animal and yeast cells, consists of aseries of structurally distinct organelles (Galway et al., 1993).
The markers used to follow plant endocytosis have in-cluded heavy metal salts, lucifer yellow, and insoluble mem-brane markers (reviewed by Fowke et al., 1991; Oparka etal., 1991). The disadvantages are that these dyes may notbe entirely cell impermeant and may enter by nonendocyticroutes (O’Drisscoll et al., 1991), as has been shown for fluo-rescein isothiocyanate (Cole et al., 1990) and lucifer yellow(Wright and Oparka, 1989; Roszak and Rambour, 1997), orthat the marker can be observed only by electron micros-copy in fixed protoplasts, such as for cationized ferritin(Fowke et al., 1991; Galway et al., 1993).
A fluorescent dye that can be used to quantify endocyto-sis in intact, walled living cells would have several advan-tages. It would permit the imaging of the organelles involvedin plant endocytosis in vivo. Additionally, it would permitcharacterization of the rate and extent of plant endocytosisin both single cells and a cell suspension. Furthermore,pharmacological agents could be screened for effects onboth endocytosis and the destination of a marker. Such afluorescent marker would aid in the characterization of themolecular machinery involved in plant cell endocytosis(Marcote et al., 2000).
The use of the styryl FM dyes as markers for neuronalendocytosis and exocytosis (Betz and Bewick, 1992; Betzet al., 1996; Henkel et al., 1996; Cochilla et al., 1999) andendocytosis in yeast (Vida and Emr, 1995), amoebae(Heuser et al., 1993), fungi (Fischer-Parton et al., 2000), and
Fu-cus
zygotes (Belanger and Quatrano, 2000a, 2000b) promptedus to evaluate using the amphipathic styryl dye FM1-43 [
N
-(3-triethylammoniumpropyl)-4-(4-[dibutylamino]styryl)pyridin-ium dibromide] to follow endocytosis in walled plant cells.The rationale was that the dye is sufficiently small to crossthe cell wall (
�
1 kD) and integrate into the plasma mem-brane or be internalized directly. If labeled fluorescentplasma membrane were internalized into cells, it could be
identified within cells after the marker remaining at theplasma membrane was removed by washing. To date,studies on the uptake of FM dyes in plants have usedmaize protoplasts (Carroll et al., 1998), marine algal cells(Battey et al., 1999; Belanger and Quatrano, 2000a,2000b), and
Vicia faba
guard cell protoplasts (Kubitschecket al., 2000), but there are few data on the use of FM1-43as a marker to analyze the morphology of the endocyticorganelles, the temperature dependence of its uptake,and the dynamics of endocytosis in intact, walled plantcells.
Here, we demonstrate that FM1-43 was internalized intointact, walled tobacco Bright Yellow 2 (BY-2) suspensioncells, from which it was transported in a time- and tempera-ture-dependent manner to internal vesicles and then to thevacuole. Endocytic compartments, including the vacuole,could be imaged in living cells, and dye uptake could bequantified by cuvette fluorimetry in a large cell population.The use of FM1-43 as a marker for intact plant cell endocy-tosis revealed that brefeldin A (BFA) stimulated endocyto-sis but wortmannin inhibited endocytosis. FM1-43 alsowas suitable for imaging vacuole and vacuolar strand dy-namics in real time by time lapse and four-dimensional (4D)imaging.
RESULTS
FM1-43 Plasma Membrane Labeling and Internalization
Our intent was to evaluate fluorescent markers suitable forfollowing endocytosis in real time in intact, walled plantcells. The prerequisites we selected for a marker for en-docytosis in plant cells were that it would be water soluble,readily cross the cell wall, have a high quantum yield, be cellimpermeable, be insensitive to pH, and be readily imagedwith conventional or confocal microscopy. After screening anumber of possible fluorophores and fluorophore conju-gates, such as low molecular weight polar dyes or rho-damine dextran conjugates (
�
6 kD) theoretically capable ofcrossing the cell wall, we identified FM1-43 as a suitablemarker for plasma membrane labeling. Other styryl dyes(FM4-64 and FM3-25) were found to be less suitable be-cause of their low brightness in cuvette studies compared withthat of FM1-43 (FM4-64) and because of difficulties in eithercomprehensively excluding that they were delivered to themitochondria (FM4-64) or with dye removal after labeling(FM3-25).
FM1-43 is a styryl chromophore that is readily soluble inwater but essentially nonfluorescent until bound to mem-branes. It then undergoes a 50- to 100-fold enhancement influorescence quantum yield as the hydrophobic portion ofthe dye dissolves into the outer leaflet of the membrane(Schote and Seelig, 1998). However, two positively charged

Fluorescent Marker for Plant Endocytosis 73
quarternary ammonium groups prevent the dye from cross-ing the bilayer, and it remains as an outer leaflet membrane-impermeant marker (Betz and Bewick, 1992; Schote andSeelig, 1998; Cochilla et al., 1999). The ready solubility ofFM1-43 in physiological buffers permits its use as a markerof endocytosis in neurons and animal cells, in which en-docytosed FM1-43 can be distinguished from plasma mem-brane labeling that can be washed out readily with excessmarker-free buffer (Betz et al., 1992).
We selected the tobacco BY-2 suspension line as amodel plant cell line because of its rapid growth under con-trolled conditions, simple morphology, and very low autoflu-orescence. Tobacco BY-2 cells possess simple plastids butdo not develop chloroplasts when grown on a simple sugar-rich medium in the dark (Nagata et al., 1992). To assessFM1-43 as a marker for BY-2 cell internalization, BY-2 cellswere incubated at 4
�
C with 20
�
M FM1-43 for 15 min andimaged by confocal sectioning, which revealed that the dyewas restricted to the plasma membrane (Figure 1A).
We confirmed that FM1-43 was membrane bound andnot intercalated into the cell wall matrix by imaging and fluo-rimetry. FM1-43 staining was observed only on the surfaceof living cells (Figure 1A) and never in cell wall fragments orcell wall ghosts, and it was seen only on the periphery ofBY-2 cells, where the cytoplasm had detached from the cellwall, but not on the cell wall itself (data not shown). The lowbinding to the plant cell wall was to be expected given thatthe fluorophore is enhanced in fluorescence only by bindingto membranes and is unlikely to be fluorescent when boundto the cell wall matrix. Membrane incorporation was con-firmed by fluorimetry; the emission spectra of FM1-43showed an
�
60-nm blue shift when added to plant cellscompared with the emission spectra in solution, as ex-pected for membrane binding (Molecular Probes, Eugene,OR) (Betz and Bewick, 1992; Betz et al., 1992; Schote andSeelig, 1998) (Figure 2B).
After insertion of FM1-43 into the plasma membrane at4
�
C, labeled BY-2 cells were incubated at 26
�
C, the opti-mal growth temperature (Nagata et al., 1992), for 30, 60, or120 min. At each time point, the cells were washed in ice-cold marker-free growth medium, which efficiently re-moved any plasma membrane FM1-43 from control cellskept at 4
�
C (data not shown). After 30 min of incubation at26
�
C, the FM1-43 distribution changed to label internalpunctate vesicles (Figure 1B, arrows). This was strikingwhen series of confocal sections were used to construct apseudo three-dimensional (3D) projection image (Figure1B, right). After incubation for 60 to 120 min, the pattern ofstaining shifted from cytoplasmic vesicles toward one thatresembled the vacuole (Figure 1D, arrows). Projection im-ages showed that FM1-43 was delivered to the vacuoleand that the morphology of the labeled compartmentschanged relative to the time of dye internalization (Figures1A to 1D, right). Therefore, FM1-43 satisfied the prerequi-sites for an internalized marker in that it crossed the cellwall, was internalized into cells, and revealed endocytic or-
ganelle morphology with stable, bright fluorescence at rel-atively low concentrations.
FM1-43 Internalization Is Temperature Dependent
An ideal fluorescent marker for endocytosis would be usefulfor quantitating uptake in a large population of cells by cu-vette fluorimetry. This would allow the temperature depen-dence of uptake and the effects of pharmacological agents,known to affect endocytosis in animal cells or postulated toact in plant cells, to be measured in a large population of in-tact, walled plant cells.
To determine if FM1-43 would be suitable as a fluorimet-ric marker and to analyze the temperature dependence ofFM1-43 uptake, a time-course endocytosis experiment wasperformed. BY-2 cells were labeled with 20
�
M FM1-43 at4, 16, or 26
�
C, and at 30, 60, or 120 min after incubation be-gan, an aliquot of cells was withdrawn and washed exten-sively to remove plasma membrane dye. Dye internalizationthen was quantitated by cuvette fluorimetry. Cell-associatedfluorescence was detected readily in a dilute cell suspen-sion (
�
5
�
10
4
cells/mL) and gave a robust signal (Figures2A and 2B), whereas cell autofluorescence was negligible(
�
0.1% of the labeled control). Internalization of FM1-43was maximal over the range of temperatures studied, at26
�
C, and cell-associated fluorescence increased with a sin-gle exponential time course (Figure 2). Dye uptake wasstrongly temperature dependent and was inhibited by
�
65% at 16
�
C and by
�
90% at 4
�
C after a 120-min incuba-tion (Figure 2A). Therefore, FM1-43 uptake satisfied the pre-requisite that it should show temperature dependence, ahallmark of endocytosis in animal cells.
FM1-43–Labeled Vesicular Staining Can Be Chased into the Vacuole
A prerequisite for identifying endocytosis is that it is medi-ated by vesicle traffic (Gruenberg and Maxfield, 1995). Thispredicts that the morphology of fluorescent labeling with anendocytic marker will change as the marker transits fromone compartment to another through membrane traffic.Therefore, we determined if the marker FM1-43 could be in-ternalized into a vesicle population and if the marker thencould be chased into the vacuole.
After a 30-min pulse of internalization, FM1-43 wasfound in cytoplasmic vesicles (Figure 3A). When these cellswere washed free of FM1-43 and further incubated inmarker-free medium at 26
�
C, FM1-43 staining shifted tothe vacuole and cytoplasmic vesicular staining diminishedwithin 60 min (Figure 3B). This is in agreement with the ob-served time course of dye internalization (Figures 1 and 2).Interestingly, vesicular profiles were stained after longerchase times (
�
120 min), which may indicate that the

74 The Plant Cell
Figure 1. Internalization of FM1-43 into Plant Suspension Cells.
Confocal sections of living BY-2 cells labeled with FM1-43 at 4�C ([A], arrows indicate plasma membrane) and after incubation at 26�C for 30min ([B], arrows indicate vesicles), 60 min (C), or 120 min ([D], arrows indicate vacuolar membrane).(A) Left, FM1-43 confocal section; middle, transmission image; right, overlay image.(B) to (D) Left, FM1-43 confocal section; middle, overlay with transmission image; right, projection image of an �30-�m confocal Z series tra-versing a BY-2 cell.n, nucleus; v, vacuole. Bar � 20 �m.

Fluorescent Marker for Plant Endocytosis 75
marker being delivered to a compartment connected to thevacuole by vesicle traffic (Figure 3C, arrow).
FM1-43 Colocalizes with a Fluid Phase Vacuolar Marker
We found that the highly polar dye Alexa 568 hydrazide wasdelivered to the vacuole over time at 26
�
C in intact walledtobacco BY-2 cells. This occurs presumably either byendocytosis, because it is a polar charged fluorophore andstaining was not visible in the cytoplasm after prolonged up-take (Figure 4A, left), or by a nonendocytic transport mecha-nism similar to that observed for other small solublefluorophores, such as fluorescein isothiocyanate (Cole et al.,1990) and lucifer yellow (Wright and Oparka, 1989; Roszakand Rambour, 1997).
To determine if FM1-43 was delivered to the same com-partment as was the fluid phase dye, cells were labeled withthe Alexa 568 dye and then washed and labeled with FM1-43as described for Figure 1. There was little colocalization be-tween the dyes when the FM1-43 was restricted to theplasma membrane (Figure 4B), but as FM1-43 uptake pro-gressed, FM1-43 colocalized with the vacuolar Alexa dye(Figures 4C and 4D).
FM1-43 Uptake Is Stimulated by BFA and Inhibitedby Wortmannin
FM1-43 is a stable fluorophore with a high quantum yieldand molecular extinction coefficient compared with manyother fluorophores. It can be detected easily in low dilutionsof cells by cuvette fluorimetry after internalization, with ahigh signal-to-noise ratio (Figure 2). As shown in Figure 5,we took advantage of these properties to investigate the ef-fects of two pharmacological agents, Brefeldin A (BFA) andwortmannin, on endocytosis. BFA is known to affect mem-brane traffic in the animal cell endocytic and secretory path-ways (Lippincott-Schwartz et al., 1989, 1991) and to affectthe plant Golgi (Satiat-Jeunemaitre and Hawes, 1992), actingin part through its actions on adenosine diphosphate ribosy-lation factor (ARF) nucleotide exchange (Orci et al., 1991;Donaldson et al., 1992; Morinaga et al., 1996). Wortmanninis a specific inhibitor of the phosphatidylinositol (PI) 3-kinasethat has been shown to block endocytosis in baby hamsterkidney cells (Clague et al., 1995). Wortmannin also has beenshown to be active in plant cells, where it inhibits proteinsorting to the vacuole at micromolar concentrations throughaction on both the PI 3- and PI 4-kinases (Matsuoka et al.,1995).
Initially, suspension cells were pretreated with 100
�
g/mLBFA or 33
�
M wortmannin for 30 min, FM1-43 was internal-ized for 60 min at 26
�
C, and cell-associated fluorescencewas quantitated after extensive washing by centrifugation.As shown in Figure 5, pretreatment of cells with 100
�
g/mLBFA significantly stimulated endocytosis compared with
control cells incubated at 26
�
C. BFA did not permeabilizethe cells directly, because cells pretreated with BFA at 26
�
Cand held on ice during FM1-43 labeling had similar cell-associated fluorescence to control cells held at 4
�
C (Figure5A). After subtraction of the background 4
�
C signal, the av-erage stimulation of endocytosis by 100
�
g/mL BFA overthe control was
�
136% (four experiments;
n
�
13) or 2.4-fold.Pretreatment of the cells with 33
�
M wortmannin inhibitedendocytosis by
�
40% (two experiments;
n
�
8) (Figure 5A)
Figure 2. FM Dye Internalization Is Time and Temperature Depen-dent.
(A) BY-2 cells were labeled with 20 �M FM1-43 at 4�C and then in-cubated at 4�C (diamonds), 16�C (squares), or 26�C (triangles) in thecontinuous presence of FM1-43. Cell-associated fluorescence wasquantitated using 488-nm excitation and 580-nm emission with a16-nm bandpass. Data were normalized to the control values at thestart of the incubation and are averages SD for three representativeexperiments.(B) Emission spectra of FM1-43 in medium or in cells labeled with 20�M FM1-43 for 60 min at 26�C and then washed three times by cen-trifugation with ice-cold BY-2 medium. Spectra are averages of trip-licate measurements.

76 The Plant Cell
Figure 3. FM1-43 Labeling Can Be Chased to the Vacuole.
Confocal sections of BY-2 cells labeled with FM1-43 using a pulsechase protocol.(A) BY-2 cells labeled for 30 min at 26�C with FM1-43 and washed.Left, FM1-43 image; right, projection image of a series of confocal Zsections.(B) BY-2 cells labeled for 30 min at 26�C with FM1-43, washed, andchased for 60 min in marker-free medium. Left, FM1-43 image; right,projection image.(C) BY-2 cells labeled for 30 min at 26�C with FM1-43, washed, andchased for 120 min in marker-free medium. Left, FM1-43 image;right, projection image. The arrow indicates a vesicle. Bar � 20 �m.
or 0.4-fold over the untreated control incubated at 26
�
C.Wortmannin pretreatment did not permeabilize the cells di-rectly, because cells pretreated with 33
�
M wortmannin andthen held on ice during FM1-43 labeling and endocytosisdid not have appreciably greater cell-associated fluores-cence than did untreated control cells held on ice (Figure5A). Investigation of the concentration dependence of BFAstimulation revealed that a significant effect was not de-tected at 10
�
g/mL (Figure 5B).Quantitative confocal imaging of cells labeled with FM1-43
across a time course demonstrated that cell-associatedfluorescence was increased significantly in the presence ofBFA (Figure 5C, right) compared with that in untreated con-trol cells (Figure 5C, left), as expected from the fluorimetricmeasurements (Figures 5A and 5B). However, the morphol-ogy of the endocytic pathway was altered dramatically byBFA treatment. After 60 min of uptake, the majority of FM1-43labeling in BFA-treated cells was found in an accumulationof large vesicles (Figure 5C), which were observed onlyrarely in control cells (Figure 1C). These vesicles presumablyare prevacuolar because they were labeled before FM1-43reached the vacuole. Furthermore, after 120 min of uptake un-der BFA treatment, prevacuolar vesicle labeling increased,but there was an inhibition of vacuolar delivery of FM1-43. Incontrol cells, vacuoles were labeled clearly after 120 min(Figure 5C, left), but under BFA treatment, vacuolar labelingwas minor and the majority of FM1-43 was accumulated inprevacuolar vesicles. Thus, although BFA treatment in-creased the overall uptake of FM1-43, dye delivery to thevacuole was inhibited and labeling accumulated in largeprevacuolar compartments.
In contrast, BFA had no significant effect on plastids labeledwith stroma-targeted green fluorescent protein in suspensioncells or on plastid dynamics (data not shown). This indicatedthat it did not disturb the structure of an unrelated compartment(plastids) under our experimental conditions. BFA is known toaffect the secretory pathway in plants (Satiat-Jeunemaitre andHawes, 1992; Driouich et al., 1993) and the secretory and en-docytic pathways in animal cells (Lippincott-Schwartz et al.,1989, 1991), Therefore, the effect we observed on the en-docytic pathway is unlikely to be nonspecific toxicity; rather, itseffect on endocytosis is analogous to effects on the secretorypathway (i.e., disruption of vesicle traffic).
FM1-43–Labeled Vacuoles Are Highly Dynamic
We used the stable fluorescent signal of FM1-43 in the vac-uole to observe vacuolar dynamics using time lapse confo-cal microscopy and 4D imaging (3D time lapse imaging).BY-2 cells were pulse labeled with FM1-43 for 120 min at26
�
C, washed on ice to remove plasma membrane staining,and then chased in marker-free medium.
Imaging the cells using a relatively deep confocal Z sec-tion showed that FM1-43 staining lined the vacuolar mem-brane, presumably after delivery via endocytic vesicle fusion/
membrane traffic (Figures 4 and 6B). We observed that thevacuole was strikingly dynamic and that vesicle movementthrough transvacuolar strands could be observed in realtime in living cells by time lapse confocal imaging (Figure6A). We observed a collection of FM1-43–labeled vesiclesmoving through strands as a bolus that arose from the pe-
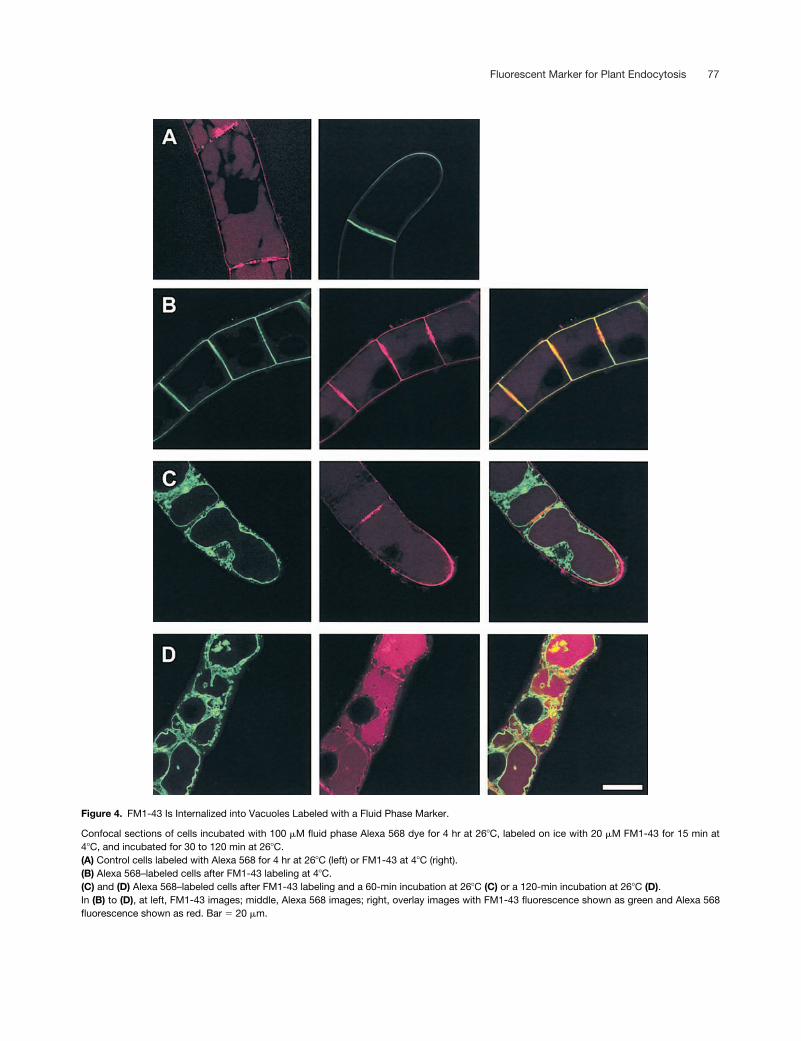
Fluorescent Marker for Plant Endocytosis 77
Figure 4. FM1-43 Is Internalized into Vacuoles Labeled with a Fluid Phase Marker.
Confocal sections of cells incubated with 100 �M fluid phase Alexa 568 dye for 4 hr at 26�C, labeled on ice with 20 �M FM1-43 for 15 min at4�C, and incubated for 30 to 120 min at 26�C.(A) Control cells labeled with Alexa 568 for 4 hr at 26�C (left) or FM1-43 at 4�C (right).(B) Alexa 568–labeled cells after FM1-43 labeling at 4�C.(C) and (D) Alexa 568–labeled cells after FM1-43 labeling and a 60-min incubation at 26�C (C) or a 120-min incubation at 26�C (D).In (B) to (D), at left, FM1-43 images; middle, Alexa 568 images; right, overlay images with FM1-43 fluorescence shown as green and Alexa 568fluorescence shown as red. Bar � 20 �m.

78 The Plant Cell
Figure 5. FM Dye Uptake Is Stimulated by BFA and Inhibited by Wortmannin.
Cell-associated FM1-43 fluorescence was quantitated after a 60-min internalization of the dye and normalized to control cell labeling at 4�C.(A) Cells were pretreated with BFA (100 �g/mL), wortmannin (33 �M), or solvent.(B) Cells were pretreated with 100, 50, or 10 �g/mL BFA or solvent.(C) Confocal images of BY-2 cells labeled with FM1-43 at 4�C and then incubated at 26�C for 30, 60, and 120 min. Left, control; right, plus 100�M BFA. 4�C panels are of a single confocal slice, and 26�C panels are projection images of an �30-�m confocal Z series traversing a BY-2 cell.All image acquisition parameters were kept constant for imaging control and BFA-treated cells at each time point.In (A) and (B), data shown are means SD and are representative of at least two experiments, each containing four individual measurements.Bars � 15 �m.

Fluorescent Marker for Plant Endocytosis 79
ripheral cytoplasm. The bolus of vesicles expanded thestrand as it passed through at a velocity of �0.5 �m/sec(Figure 6A, arrows). To be certain that discrete strandspassing through the vacuolar space were imaged, transvac-uolar strands were identified by serial Z sectioning beforetime lapse imaging was initiated. This enabled strands to bedistinguished in a confocal section from invaginations of thevacuolar wall into the vacuolar space or large bridges of cy-toplasm extending from the nucleus to the wall. The trans-vacuolar strands were mobile, and the stalk of the strandsoften migrated along the wall of the vacuole. The strands re-mained rigid during movement, and we also observed bothends of the strands moving at similar velocities across thevacuole in concert with vesicle movement through thestrand (data not shown).
It was challenging to image the movement of transvacu-olar strands, because they were rarely orientated so thatthey could be imaged in a single confocal Z section overtime. Therefore, we selected 4D imaging to observe vacu-olar dynamics because it captured movement within a large3D volume of a cell. For 4D imaging, a broad lateral sectionthrough a cell was defined and then imaged in four to eight�1-�m-deep consecutive confocal Z sections. The sectionsthen were used to create a 3D projection image on image pro-cessing (Figure 6B, 1). To create a 4D imaging series, the 3Dseries was acquired every 30 or 60 sec (Figure 6B, 1 to 9). EachZ series was image processed to create a projection image.
The 4D imaging series shown in Figure 6B shows a recon-structed �7-�m section through the middle of a chain ofBY-2 cells, which was acquired as a series of consecutive Zsections. Each panel shows a 3D projection image of thetime lapse series of Z sections, but in contrast to the projec-tion images shown above (Figure 1, right), the projection im-age does not show the entire cell but an �7-�m lateralsection, and images were captured over time. The nucleusis located in the middle of the image and is surrounded by ahalo of labeled inner vacuolar membrane, vesicles, broadbridges of cytoplasm (Figure 6B, 1, closed arrows), and thintransvacuolar strands (Figure 6B, 1, arrowheads). 4D imagingrevealed that strands fuse laterally to create broader strands(Figure 6B, 2 to 5, arrowheads) and that vesicles, presum-ably endocytic because they were labeled with FM1-43, movedin three dimensions within the transvacuolar strands (Figure6B, 4 to 9, open arrows). The fusion of two strands did notappear to involve lateral movement of the strands, but vacu-olar membrane moved along both strands from between thenuclear base of the strands toward the side of the vacuole,so that the strands were converted into a sheet of mem-brane stretching from the nucleus to the vacuolar wall (Fig-ure 6B, 2 to 5, arrowheads).
Vesicles moved along and within strands, and this wasseen most readily in 4D imaging movies of vacuolar dynam-ics (Figure 6B). Vesicles at the base of the fused vacuolarstrands described above appeared to move in three dimen-sions, and this was independent of strand movement (Fig-ure 6B, 4 to 9, open arrows).
DISCUSSION
In this study, the unique features of an amphipathic styryldye, FM1-43, were exploited as a noninvasive marker to re-veal the morphology, uptake kinetics, and ultrastructural dy-namics of the endocytic pathway in living plant cells.
Here, we have characterized FM1-43 as a fluorescentmarker for membrane uptake in intact, walled plant cells. Thescarcity of such a marker has hampered investigations intoplant cell endocytosis (Buchanan et al., 2000). Our intentwas to determine whether FM1-43 satisfied the prerequi-sites of a marker for endocytosis. Although the literature onthe use of FM1-43 in neurobiology is extensive (Betz et al.,1996; Cochilla et al., 1999), there have been few studies onits use in plants and, to our knowledge, no examination of itsuse in intact plant suspension cells. The uptake of FM dyeswas shown in V. faba guard cell protoplasts (Kubitscheck etal., 2000), maize root cap protoplasts (Carroll et al., 1998),and zygotes from marine alga (Battey et al., 1999; Belangerand Quatrano, 2000a). These studies prompted us to exam-ine whether FM1-43 was suitable for tobacco suspensioncells, which are walled, in contrast to protoplasts, and canbe cultured easily as a large, synchronized population(Nagata et al., 1992).
The prerequisites defined for a fluorescent marker formembrane endocytosis in plant cells were similar to thosefor animal cell endocytosis (Salzman and Maxfield, 1988,1989): that the probe be nontoxic, demonstrate a robustfluorescence in the membrane, is able to cross the plant cellwall, and is internalized into cells by a route that is stronglytemperature dependent. Our results indicate that FM1-43fulfills many of these prerequisites.
When cells were incubated at 4�C with FM1-43, confocalimaging demonstrated that staining was restricted to a pat-tern consistent with the plasma membrane. Plasma mem-brane incorporation of the dye was likely because nofluorescent labeling was observed in the cell walls or cellwall fragments. Incubation of the cells at 26�C, the optimalculture temperature for BY-2 cells, gave a cytoplasmic vesi-cle staining pattern after 30 min. Incubation at 26�C for 60 or120 min shifted the dye localization to a pattern consistentwith vacuolar labeling. Cationized ferritin is delivered to thevacuole after 60 min of exposure (Fowke et al., 1991), whichis consistent with the data presented here. Ferritin delivery ismediated by multivesicular bodies, which are similar to thoseobserved in animal cells that mediate early to late endosometransport along microtubules (Gruenberg et al., 1989) andwhich fuse with the vacuole. It is possible that FM1-43 is de-livered by a similar vesicular carrier. To confirm this specula-tion, further analysis is necessary using techniques, such asphotooxidation, that permit FM1-43 localization by electronmicroscopy (Nishikawa and Sasaki, 1996).
Because the FM1-43 crossed the cell wall and plasmamembrane–associated FM1-43 could be removed by washing,the intracellular sequestration of the dye could be followed by

80 The Plant Cell
Figure 6. Time Lapse and 4D Imaging of Vacuolar Dynamics in Fluorescently Labeled BY-2 Cells.
BY-2 cells were labeled with 20 �M FM1-43 for 120 min at 26�C and then washed and imaged.(A) Time lapse imaging of a single confocal section (�1 �m) showing vesicle movement through a transvacuolar strand. Time scale is shown inseconds. Arrows indicate vesicles and membrane boluses. Bar � 20 �m.(B) 4D imaging of labeled cells showing strand and vesicle movement. Z series (�7 �m deep) of approximately six confocal sections were ac-quired at 1-min intervals. 3D images were reconstructed for each time point and then used to create a 4D imaging movie. The montage showsnine images taken at 1-min intervals. Closed arrows, bridge of cytoplasm; arrowheads, transvacuolar strand; open arrows, vesicles within astrand. Bar � 20 �m.

Fluorescent Marker for Plant Endocytosis 81
measuring cell-associated fluorescence. Dye sequestrationwas time and temperature dependent: it was reduced by�65% by a temperature shift from 26 to 16�C and by �90%at 4�C. Uptake at 26�C followed a simple exponential timecourse, and the fluorescence emission spectra of seques-tered FM1-43 showed a blue shift (peak emission , 560 nm)compared with that of the dye in solution in water (peak emis-sion , 620 nm), consistent with the intercalation of the dyeinto the membrane (Schote and Seelig, 1998). Therefore,FM1-43 was internalized into intact cells by means of a time-and temperature-dependent mechanism by which a signifi-cant fraction of the dye remained bound to membranes.
We speculate that FM1-43 is taken up by endocytic vesi-cles at the plasma membrane of tobacco suspension cellsbecause dye uptake experiments show temperature-depen-dent traffic of the dye from the plasma membrane throughendomembrane compartments to the vacuole. This is sup-ported by the evidence that FM dyes can be used to studyendocytosis in many experimental organisms, including yeast,Dictyostelium, invertebrates, rat hippocampal and frog mo-tor neurons, starfish oocytes, and transgenic mouse–derivedhippocampal cell lines (reviewed by Betz et al., 1996; Cochillaet al., 1999).
However, alternative pathways may contribute to plantcell labeling; for example, the direct internalization of lipidsfrom the plasma membrane to the endoplasmic reticulum(ER) has been shown for soybean cells (Grabski et al., 1993),but the extent of this activity for specific lipids or fluorescentlipid analogs is unclear. We observed little evidence of thedirect delivery of FM1-43 from the plasma membrane to theER, based on the morphology and dynamics of the labeledcompartments and the temperature sensitivity of dye uptake.Even after long-term chase after cell labeling (16 to 48 hr),FM1-43 was found only in the vacuole, and there was no ev-idence of ER staining in intact cells. Although it has beendemonstrated that fluorescent analogs of phosphatidylcho-line or phosphatidylethanolamine are transported from theyeast plasma membrane by a nonendocytic transbilayermechanism to internal organelles, FM4-64, a dye closely re-lated to FM1-43, enters yeast by an endocytic mechanismrequiring increased temperature (Grant et al., 2001). This in-dicates that members of the FM family of dyes are not takenup by a nonendocytic pathway and so are not available fordelivery to the ER from the plasma membrane via the cyto-plasm. In addition, FM1-43 has a positive electric chargethat prevents it from crossing the membrane, so it remainspartitioned in the outer lipid leaflet of the animal cell plasmamembrane (Sulzer and Holtzman, 1989; Schote and Seelig,1998). Thus, we speculate that FM1-43 does not enter plantcells via the nonendocytic pathway observed for the phos-phatidylcholine analog (Grabski et al., 1993) but serves as amarker for temperature-dependent membrane internaliza-tion from the plasma membrane via vesicle traffic, as is wellestablished for FM4-64 in yeast (Vida and Emr, 1995).
Recently, a biochemical marker for receptor-mediated en-docytosis was developed for rice suspension cells based on
the specific binding and internalization of biotin-labeled pro-teins (Bahaji et al., 2001). FM1-43 has some advantagesover protein markers in that it is a membrane marker whoseuptake can be both imaged and easily quantified in living in-tact cells. We propose that FM1-43 will complement studieson receptor-mediated endocytosis by providing a marker forthe default pathway of membrane endocytosis.
Interestingly, Belanger and Quatrano (2000a) analyzed thedistribution and uptake of FM4-64 in fertilized Fucus zy-gotes. It was demonstrated that FM4-64 can be delivered toperinuclear endomembrane organelles that may include thevacuole, Golgi, or ER, and then to the newly forming cellplate. They proposed that the initial uptake of FM4-64 ismediated by endocytosis at the plasma membrane, fromwhich label is trafficked to the perinuclear region. We be-lieve that this finding supports our proposal that FM1-43 isendocytosed into plant cells under our conditions, but wespeculate that the membrane traffic pathways in BY-2 cellsare dissimilar to those in Fucus zygotes and that membranetrafficking is specialized along the developmental pathwaysrequired for zygote development.
A feature of FM1-43 as an endocytosis marker is that itcan be used with intact plant cells, in which it crosses thecell wall and implants into the plasma membrane. This per-mits analysis and imaging of the effects of pharmacologicalagents and plant signaling compounds on a defined stageof membrane traffic. BFA strongly stimulated temperature-dependent endocytosis of the marker, whereas wortmanninacted as an inhibitor. BFA stimulated endocytosis in BY-2cells at 50 to 100 �g/mL, a concentration that is within therange used for plants and plant suspension cells (Satiat-Jeunemaitre and Hawes, 1992; Gomez and Chrispeels,1993; Boevink et al., 1998; Wee et al., 1998; Mullen et al.,1999). Interestingly, Belanger and Quatrano (2000a) demon-strated that although BFA inhibited the asymmetric growthof Fucus zygotes and the formation of the cell plate, uptakeof FM4-64 continued but labeled membrane was not deliv-ered to the perinuclear region.
Because membrane traffic generally is conserved fromyeast to animals, we speculate that BFA exerts its action inplants by mechanisms analogous to those in animal cells. Inanimal cells, one of the major effects of BFA is the break-down of the Golgi through its inhibitory action on the gua-nine nucleotide exchange factor (GEF) for ARF1 (reviewedby Chavrier and Goud, 1999) and the restriction of vesiclebudding events from organelles. Several ARF GEFs in ani-mals and yeast are BFA sensitive, including yeast Sec7p;however, the BFA target in plant cells is uncharacterized.Whether an ARF is associated with the plant endocyticpathway also is unclear, but in animal and yeast cells thereis clear involvement of ARFs in endocytosis. Mutation ofARF1 in yeast inhibits endocytic transport to the vacuole(Gaynor et al., 1998), and expression of ARF6 mutants innonpolarized animal cells, in which the wild-type protein is lo-calized to endosomes and the plasma membrane, disruptsendocytosis (D’Souza-Schorey et al., 1998). Furthermore,

82 The Plant Cell
ARF6 is found on the apical surface of polarized Madin-Darby canine kidney cells, in which overexpression of theARF6-Q67L mutant, predicted to be locked in the GTP-bound form, stimulates apical endocytosis (Altschuler et al.,1999). The Arabidopsis protein GNOM is a membrane-asso-ciated ARF GEF involved in the polar localization of theauxin efflux carrier PIN1 (Steinmann et al., 1999). The GEFactivities of GNOM and PIN1 localization are BFA sensitive.It is attractive to speculate that GNOM or similar plant ARFGEFs regulate vesicle trafficking at the plant cell plasma mem-brane and that the stimulation of dye uptake by BFA isthrough its action on an ARF GEF involved in endocytosis.
We believe that the stimulation of FM1-43 uptake uponBFA treatment is based on an increase in membrane inter-nalization that may be coupled to decreased recycling andan inhibition of transport to the vacuole. Prevacuolar endo-somal vesicles accumulate and appear to enlarge underBFA treatment (Figure 5C), and this may be related to BFAdisrupting budding from an endosomal compartment andhence dye recycling to the plasma membrane, or to a fu-sion/sorting step in which these vesicles would be con-sumed. Furthermore, if budding is reduced along therecycling endocytic pathway, it would lead to a net increasein internalized FM1-43 by affecting a step that would lead todye loss from the cell (i.e., production of membrane des-tined for recycling).
A BFA-sensitive step is likely to be found between theprevacuolar endosomal compartment and the vacuole.Confocal imaging revealed that the majority of FM1-43 la-beling was found in the large prevacuolar endosomes andthat there was an inhibition of dye delivery to the vacuoleupon BFA treatment (Figure 5C). Therefore, BFA may act ata transport step from this prevacuolar compartment to thevacuole, and the prevacuolar compartment may accumulateas a consequence of inhibited transport.
BFA affects ER-to-Golgi transport in plants, in which itselectively blocks anterograde but not retrograde trans-port (Boevink et al., 1998). Transport and sorting of pro-teins to the vacuole also show differential sensitivity toBFA, phytohemagglutinin transport via the Golgi to thevacuole is blocked, and transport of tonoplast intrinsicprotein is unaffected (Gomez and Chrispeels, 1993). Im-portantly, BFA prevents coat protein (AtArf1p andcoatomer) recruitment to the Golgi membrane in vitro, afundamental stage in non-clathrin-coated vesicle budding(Pimpl et al., 2000). Thus, the effects we observed in en-docytosis may relate to coat protein recruitment at a stageof endocytosis responsible for dye loss from the cell—other steps of endocytosis are unaffected because theydo not possess a BFA-sensitive component—by analogywith retrograde Golgi-to-ER transport and vacuolar deliv-ery of tonoplast intrinsic protein.
Wortmannin is a PI 3-kinase inhibitor that inhibits endocy-tosis (Clague et al., 1995; Li et al., 1995) and receptor traf-ficking in animal cells (Shpetner et al., 1996). Although theconcentrations used here are higher than those used in ani-
mal cells, they are in the range known to affect protein sort-ing to the vacuole in tobacco BY-2 cells, in which it inhibitsPI 3-phosphate, PI 4-phosphate, and phospholipid synthe-sis (Matsuoka et al., 1995). Our data provide evidence thatendocytosis in BY-2 cells has a wortmannin-sensitive com-ponent, but further investigation is required to characterizethe mechanism by which it acts.
The robust fluorescence of FM1-43 in the vacuole permit-ted imaging of endocytic vesicle traffic through transvacu-olar strands and the dynamics of the strands themselves.We often observed vesicles moving as a bolus through vac-uolar strands and the movement and reorganization of thestrands, as has been observed with green fluorescent pro-tein fusion proteins targeted to the vacuole (Cutler et al.,2000). 4D imaging of living FM1-43 cells gave dramatic im-ages of the vacuole and vacuolar strand dynamics. Strandscould be observed to fuse laterally along their length, like azipper closing, and move through three dimensions.
We have demonstrated the utility of FM1-43 as a fluores-cent marker for resolving the kinetics of endocytosis mor-phology and the dynamics of the endocytic pathway in plantcells. Using FM1-43 as a marker for plant endocytosis in in-tact plant cells revealed BFA and wortmannin sensitivity.Further work will focus on identifying the protein targets thatare affected by BFA and wortmannin treatment and thatmay regulate plant cell endocytosis.
METHODS
Tobacco Suspension Cultures
Tobacco (Nicotiana tabacum cv Bright Yellow 2 [BY-2]) cells werecultured in Murashige and Skoog (1962) basal salt with minimal or-ganics (BY-2 medium: 0.47% [w/v] Murashige and Skoog salts, 0.15�g/mL thiamine, 0.02 �g/mL 2,4-D, 2 mg/mL KH2PO4, and 3% [w/v]sucrose, pH 5.2) in an orbital shaker (New Brunswick Scientific,Nürtingen, Germany) at 180 rpm and 26�C in the dark. All experi-ments with BY-2 cells were performed using cells after a 5% (v/v) di-lution and 3 days of subcultivation.
Chemicals, Media, and Fluorophores
All fluorophores were purchased from Molecular Probes. All otherfine chemicals were supplied by Sigma-Aldrich (Deisenhofen,Germany). FM1-43 [N-(3-triethylammoniumpropyl)-4-(4-[dibutyl-amino]styryl)pyridinium dibromide] was kept as a 20 mM stock solu-tion in DMSO. Alexa 568 hydrazide was a 50 mM stock in sterilewater. Brefeldin A (BFA) was kept as a 50 mg/mL stock in methanol,and wortmannin was kept as a 10 mM stock in DMSO at �20�C.
Plant Cell Confocal Imaging
BY-2 cells were imaged with a TCS-SP spectral confocal micro-scope equipped with argon ion, krypton, and helium-neon lasers

Fluorescent Marker for Plant Endocytosis 83
(Leica, Heidelberg, Germany). Images were acquired with a 1.2 nu-merical aperture �63 oil immersion PLAN-APO objective. FM1-43 wasexcited with the 488-nm argon laser line, and confocal sectionswere collected using a 510- to 580-nm emission setting. Alexa 568 wasexcited with the 568-nm krypton laser line, and confocal imageswere collected using 600- to 620-nm emission.
Plant Cell FM1-43 Labeling
Four milliliters of a 3-day-old BY-2 culture was placed on ice for 15min and then supplemented with 20 �M FM1-43. After 15 min of in-cubation on ice, the cells were transferred to an orbital shaker at26�C and 180 rpm. After 30, 60, and 120 min at 26�C, a 1-mL aliquotwas removed and washed three times by centrifugation (at 1600g,4�C, for 5 min) in 15 mL of marker-free ice-cold BY-2 medium. Thepellet was resuspended in 1 mL of BY-2 medium, and the cells wereimaged.
FM1-43 and Alexa 568 Double Labeling
Four milliliters of a BY-2 culture was placed on ice for 15 min andthen supplemented with 100 �M Alexa 568 hydrazide. The cells weretransferred to a 26�C orbital shaker for 4 hr. BY-2 cells were chilledby the addition of ice-cold BY-2 medium and washed three times bycentrifugation (at 1600g, 4�C, for 5 min) in 50 mL of ice-cold BY-2medium. The cell pellet was resuspended in 4 mL of ice-cold BY-2medium, and an aliquot was imaged. The Alexa 568–labeled cell sus-pension was supplemented with 20 �M FM1-43 and incubated for 15min on ice and then imaged. Cells then were transferred to a 26�Corbital shaker incubator. After 30, 60, and 120 min at 26�C, a 1-mLaliquot was removed and washed three times by centrifugation (at1600g, 4�C, for 5 min) in 15 mL of marker-free ice-cold BY-2 me-dium. The cells were resuspended in 1 mL of BY-2 medium and im-aged. Confocal FM1-43 and Alexa 568 images were acquiredserially and overlaid using Metamorph (Universal Imaging, WestChester, PA).
Plant Cell Fluorimetry Measurements
Four milliliters of a 3-day-old BY-2 culture was placed on ice for 15min and then supplemented with 20 �M FM1-43. After 15 min of in-cubation on ice, the cells were transferred to a 26�C orbital shaker in-cubator at 180 rpm. After 30, 60, and 120 min at 26�C, a 1-mL aliquotwas removed and washed three times by centrifugation (at 1600g,4�C, for 5 min) in marker-free ice-cold BY-2 medium. Cell-associatedfluorescence was quantitated by fluorimetry using an SLM AmincoBowman AB-2 fluorospectrometer (Polytec, Waldbronn, Germany)and a 1-cm path length polymethylacrylate cuvette. FM1-43 was ex-cited at 488 nm (16-nm bandpass), and emission was detected at580 nm (16-nm bandpass). For each time course, the cell-associatedfluorescence was normalized to the average fluorescent signal of thecells at the start of the incubation. In single time point measure-ments, cell-associated fluorescence was normalized to the cell-associated fluorescence of control cells incubated at 4�C. Excita-tion and emission spectra were acquired using a 5-nm/sec scanspeed, and the results of three measurements were averaged (4-nmbandpass).
Time Lapse Imaging
Four milliliters of a BY-2 cell culture was labeled for 120 min with 20�M FM1-43 at 26�C, washed three times by centrifugation in ice-cold BY-2 medium (at 1600g, 4�C, for 5 min), and then resuspendedin 4 mL of marker-free BY-2 medium. BY-2 cells were incubated at26�C in a shaker incubator at 180 rpm and imaged. Confocal XYsections (�2 �m in Z) were acquired at four frames per minute for10 min.
Four-Dimensional Imaging
Cells were labeled as for time lapse imaging and imaged. A lateralconfocal Z series, typically 6 to 8 �m, was defined by Z sectioning.This Z series then was acquired automatically every 30 or 60 sec forup to 25 min. The sequential series then was image processed usingcustom journals to generate a series.
Software and Image Processing
Images were acquired as eight-bit multi-image TIF files using theLeica spectral confocal TCSNT software, version 1.6.58, running un-der Windows NT on a 450-mHz Pentium II personal computerequipped with 384 megabytes of random access memory. Imageoverlay was achieved either using the host Leica software after par-allel confocal image acquisition or with the Universal Imaging Meta-morph 4.5 software after serial image acquisition. Multi-image TIFfiles of confocal time lapse images were montaged and labeled inMetamorph before export as *.TIF files. Image files were archived onCD-ROMs media. For presentation, individual files were importedinto Adobe PhotoShop (Mountain View, CA) running under WindowsNT and saved as 24-bit CMYK files. For four-dimensional (4D) imag-ing, custom Metamorph journals were used to automatically extracteach series from the multi-image TIF confocal files and generate in-dividual projection images using Metamorph.
ACKNOWLEDGMENTS
We thank Prof. Anne-Marie Lambert (Institut de Biologie Moléculairedes Plantes, Strasbourg, France) for providing the starter culture ofthe tobacco BY-2 cell line and Prof. Dr. Alan Slusarenko and Dr. YoramAltschuler for helpful discussions. We thank Dr. Flora Schuster forculturing the BY-2 cells and Norbert Gulde of Visitron Instruments forsupport with the Universal Imaging software. We are indebted toJürgen Drossard for originally suggesting BY-2 as a model systemand to the Fraunhofer Gesellschaft for providing the imaging suite.
Received August 9, 2001; accepted October 2, 2001.
REFERENCES
Aderem, A., and Underhill, D. (1999). Mechanisms of phagocytosisin macrophages. Annu. Rev. Immunol. 17, 593–623.

84 The Plant Cell
Altschuler, Y., Liu, S., Katz, L., Tang, K., Hardy, S., Brodsky, F.,Apodaca, G., and Mostov, K. (1999). ADP-ribosylation factor 6and endocytosis at the apical surface of Madin-Darby canine kid-ney cells. J. Cell Biol. 147, 7–12.
Anderson, R.G. (1998). The caveolae membrane system. Annu.Rev. Biochem. 67, 199–225.
Anuntalabhochai, S., Terryn, N., Van Montagu, M., and Inze, D.(1991). Molecular characterization of an Arabidopsis thalianacDNA encoding a small GTP-binding protein, Rha1. Plant J. 1,167–174.
Bahaji, A., Cornejo, M.J., Ortiz-Zapater, E., Contreras, I., andAniento, F. (2001). Uptake of endocytic markers by rice cells: Vari-ations related to the growth phase. Eur. J. Cell Biol. 80, 178–186.
Battey, N., James, N., Greenland, A., and Brownlee, C. (1999).Exocytosis and endocytosis. Plant Cell 11, 643–659.
Belanger, K.D., and Quatrano, R.S. (2000a). Membrane recyclingoccurs during asymmetric tip growth and cell plate formation inFucus distichus zygotes. Protoplasma 212, 24–37.
Belanger, K.D., and Quatrano, R.S. (2000b). Polarity: The role oflocalized secretion. Curr. Opin. Plant Biol. 3, 67–72.
Betz, W.J., and Bewick, G.S. (1992). Optical analysis of synapticvesicle recycling at the frog neuromuscular junction. Science 255,200–203.
Betz, W.J., Mao, F., and Bewick, G.S. (1992). Activity-dependentfluorescent staining and destaining of living vertebrate motornerve terminals. J. Neurosci. 12, 363–375.
Betz, W.J., Mao, F., and Smith, C.B. (1996). Imaging exocytosisand endocytosis. Curr. Opin. Neurobiol. 6, 365–371.
Blackbourn, H.D., and Jackson, A.P. (1996). Plant clathrin heavychain: Sequence analysis and restricted localisation in growingpollen tubes. J. Cell Sci. 109, 777–786.
Boevink, P., Oparka, K., Santa Cruz, S., Martin, B., Betteridge,A., and Hawes, C. (1998). Stacks on tracks: The plant Golgiapparatus traffics on an actin/ER network. Plant J. 15, 441–447.
Borg, S., Brandstrup, B., Jensen, T.J., and Poulsen, C. (1997).Identification of new protein species among 33 different smallGTP-binding proteins encoded by cDNAs from Lotus japonicus,and expression of corresponding mRNAs in developing root nod-ules. Plant J. 11, 237–250.
Braell, W.A. (1987). Fusion between endocytic vesicles in a cell-freesystem. Proc. Natl. Acad. Sci. USA 84, 1137–1141.
Braell, W.A. (1992). Detection of endocytic vesicle fusion in vitro,using assay based on avidin-biotin association reaction. MethodsEnzymol. 219, 12–21.
Buchanan, B., Gruissem, W., and Jones, R. (2000). Biochemistryand Molecular Biology of Plants. (Rockville, MD: American Societyof Plant Physiologists).
Carpita, N., Sabularse, D., Montezinos, D., and Delmer, D.P.(1979). Determination of the pore size of cell walls of living plantcells. Science 205, 1144–1147.
Carroll, A., Moyen, C., Van Kesteren, P., Tooke, F., Battey, N.,and Brownlee, C. (1998). Ca2�, annexins, and GTP modulateexocytosis from maize root cap protoplasts. Plant Cell 10, 1267–1276.
Chavrier, P., and Goud, B. (1999). The role of ARF and Rab GTPasesin membrane transport. Curr. Opin. Cell Biol. 11, 466–475.
Christoforidis, S., McBride, H.M., Burgoyne, R.D., and Zerial, M.(1999a). The Rab5 effector EEA1 is a core component of endo-some docking. Nature 397, 621–625.
Christoforidis, S., Miaczynska, M., Ashman, K., Wilm, M., Zhao,L., Yip, S.C., Waterfield, M.D., Backer, J.M., and Zerial, M.(1999b). Phosphatidylinositol-3-OH kinases are Rab5 effectors.Nat. Cell Biol. 1, 249–252.
Clague, M.J., Thorpe, C., and Jones, A.T. (1995). Phosphatidyli-nositol 3-kinase regulation of fluid phase endocytosis. FEBS Lett.367, 272–274.
Cochilla, A.J., Angleson, J.K., and Betz, W.J. (1999). Monitoringsecretory membrane with FM1-43 fluorescence. Annu. Rev. Neu-rosci. 22, 1–10.
Cole, L., Coleman, J., Evans, D., and Hawes, C. (1990). Internal-ization of fluorescein isothiocyanate and fluorescein isothiocyan-ate-dextran by suspension-cultured plant cells. J. Cell Sci. 96,721–730.
Coleman, J., Evans, D., Hawes, C., Horsley, D., and Cole, L.(1987). Structure and molecular organization of higher plantcoated vesicles. J. Cell Sci. 88, 35–45.
Cutler, S.R., Ehrhardt, D.W., Griffitts, J.S., and Somerville, C.R.(2000). Random GFP::cDNA fusions enable visualization of sub-cellular structures in cells of Arabidopsis at a high frequency.Proc. Natl. Acad. Sci. USA 97, 3718–3723.
Donaldson, J.G., Finazzi, D., and Klausner, R.D. (1992). BrefeldinA inhibits Golgi membrane–catalysed exchange of guanine nucle-otide onto ARF protein. Nature 360, 350–352.
Driouich, A., Zhang, G.F., and Staehelin, L.A. (1993). Effect ofbrefeldin A on the structure of the Golgi apparatus and on thesynthesis and secretion of proteins and polysaccharides insycamore maple (Acer pseudoplatanus) suspension-culturedcells. Plant Physiol. 101, 1363–1373.
D’Souza-Schorey, C., van Donselaar, E., Hsu, V.W., Yang, C.,Stahl, P.D., and Peters, P.J. (1998). ARF6 targets recycling vesi-cles to the plasma membrane: Insights from an ultrastructuralinvestigation. J. Cell Biol. 140, 603–616.
Emans, N., and Verkman, A.S. (1996). Real-time fluorescence mea-surement of cell-free endosome fusion: Regulation by secondmessengers. Biophys. J. 71, 487–494.
Emans, N., Biwersi, J., and Verkman, A.S. (1995). Imaging ofendosome fusion in BHK fibroblasts based on a novel fluorimetricavidin-biotin binding assay. Biophys. J. 69, 716–728.
Emons, A.M.C., and Traas, J.A. (1986). Coated pits and coatedvesicles on the plasma membrane of plant cells. Eur. J. Cell Biol.41, 57–64.
Fischer-Parton, S., Parton, R.M., Hickey, P.C., Dijksterhuis, J.,Atkinson, H.A., and Read, N.D. (2000). Confocal microscopy ofFM4-64 as a tool for analysing endocytosis and vesicle traffickingin living fungal hyphae. J. Microsc. 198, 246–259.
Fowke, L., and Gamborg, O. (1980). Applications of protoplasts inthe study of plant cells. Int. Rev. Cytol. 68, 9–51.
Fowke, L., Tanchak, M., and Galway, M. (1991). Ultrastructuralcytology of the endocytotic pathway in plants. In Endocytosis,Exocytosis and Vesicle Traffic in Plants, C. Hawes, J. Coleman,and D. Evans, eds (Cambridge, UK: Cambridge University Press),pp. 15–40.

Fluorescent Marker for Plant Endocytosis 85
Galway, M., Rennie, P., and Fowke, L. (1993). Ultrastructure of theendocytic pathway in glutaraldehyde fixed and high-pressure fro-zen/freeze-substituted protoplasts of white spruce (Picea glauca).J. Cell Sci. 106, 847–858.
Gaynor, E.C., Chen, C.Y., Emr, S.D., and Graham, T.R. (1998).ARF is required for maintenance of yeast Golgi and endosomestructure and function. Mol. Biol. Cell 9, 653–670.
Goldstein, J.L., Brown, M.S., Anderson, R.G., Russell, D.W., andSchneider, W.J. (1985). Receptor-mediated endocytosis: Con-cepts emerging from the LDL receptor system. Annu. Rev. CellBiol. 1, 1–39.
Gomez, L., and Chrispeels, M. (1993). Tonoplast and soluble vacu-olar proteins are targeted by different mechanisms. Plant Cell 5,1113–1124.
Gorvel, J.P., Chavrier, P., Zerial, M., and Gruenberg, J. (1991).rab5 controls early endosome fusion in vitro. Cell 64, 915–925.
Grabski, S., de Feijter, A., and Schindler, M. (1993). Endoplasmicreticulum forms a dynamic continuum for lipid diffusion betweencontiguous soybean root cells. Plant Cell 5, 25–38.
Grant, A.M., Hanson, P.K., Malone, L., and Nichols, J.W. (2001).NBD-labeled phosphatidylcholine and phosphatidylethanolamineare internalized by transbilayer transport across the yeast plasmamembrane. Traffic 2, 37–50.
Griffiths, G., Back, R., and Marsh, M. (1989). A quantitative analy-sis of the endocytic pathway in baby hamster kidney cells. J. CellBiol. 109, 2703–2720.
Gruenberg, J., and Maxfield, F.R. (1995). Membrane transport inthe endocytic pathway. Curr. Opin. Cell Biol. 7, 552–563.
Gruenberg, J., Griffiths, G., and Howell, K.E. (1989). Characteriza-tion of the early endosome and putative endocytic carrier vesiclesin vivo and with an assay of vesicle fusion in vitro. J. Cell Biol. 108,1301–1316.
Henkel, A.W., Lubke, J., and Betz, W.J. (1996). FM1-43 dye ultra-structural localization in and release from frog motor nerve termi-nals. Proc. Natl. Acad. Sci. USA 93, 1918–1923.
Heuser, J., Zhu, Q., and Clarke, M. (1993). Proton pumps populatethe contractile vacuoles of Dictyostelium amoebae. J. Cell Biol.121, 1311–1327.
Hewlett, L.J., Prescott, A.R., and Watts, C. (1994). The coated pitand macropinocytic pathways serve distinct endosome popula-tions. J. Cell Biol. 124, 689–703.
Holstein, S.E., Drucker, M., and Robinson, D.G. (1994). Identifica-tion of a beta-type adaptin in plant clathrin-coated vesicles. J. CellSci. 107, 945–953.
Kubitscheck, U., Homann, U., and Thiel, G. (2000). Osmoticallyevoked shrinking of guard-cell protoplasts causes vesicularretrieval of plasma membrane into the cytoplasm. Planta 210,423–431.
Li, G., D’Souza-Schorey, C., Barbieri, M.A., Roberts, R.L., Klippel,A., Williams, L.T., and Stahl, P.D. (1995). Evidence for phos-phatidylinositol 3-kinase as a regulator of endocytosis via acti-vation of Rab5. Proc. Natl. Acad. Sci. USA 92, 10207–10211.
Lippincott-Schwartz, J., Yuan, L.C., Bonifacino, J.S., and Klausner,R.D. (1989). Rapid redistribution of Golgi proteins into the ER incells treated with brefeldin A: Evidence for membrane cyclingfrom Golgi to ER. Cell 56, 801–813.
Lippincott-Schwartz, J., Yuan, L., Tipper, C., Amherdt, M., Orci,L., and Klausner, R.D. (1991). Brefeldin A’s effects on endo-somes, lysosomes, and the TGN suggest a general mechanismfor regulating organelle structure and membrane traffic. Cell 67,601–616.
Low, P., and Chandra, S. (1994). Endocytosis in plants. Annu. Rev.Plant Physiol. Plant Mol. Biol. 45, 609–631.
Marcote, M., Gu, F., Gruenberg, J., and Aniento, F. (2000). Mem-brane transport in the endocytic pathway: Animal versus plantcells. Protoplasma 210, 123–132.
Matsuoka, K., Bassham, D., Raikhel, N., and Nakamura, K.(1995). Different sensitivity to wortmannin of two vacuolar sortingsignals indicates the presence of distinct sorting machineries intobacco cells. J. Cell Biol. 130, 1307–1318.
McBride, H.M., Rybin, V., Murphy, C., Giner, A., Teasdale, R., andZerial, M. (1999). Oligomeric complexes link Rab5 effectors withNSF and drive membrane fusion via interactions between EEA1and syntaxin 13. Cell 98, 377–386.
Morinaga, N., Tsai, S.C., Moss, J., and Vaughan, M. (1996). Isola-tion of a brefeldin A–inhibited guanine nucleotide-exchange pro-tein for ADP ribosylation factor (ARF) 1 and ARF3 that contains aSec7-like domain. Proc. Natl. Acad. Sci. USA 93, 12856–12860.
Mullen, R.T., Lisenbee, C.S., Miernyk, J.A., and Trelease, R.N.(1999). Peroxisomal membrane ascorbate peroxidase is sorted toa membranous network that resembles a subdomain of the endo-plasmic reticulum. Plant Cell 11, 2167–2185.
Nagata, T., Nemoto, Y., and Hasezawa, S. (1992). Tobacco BY-2cell line as the “HeLa” cell in the cell biology of higher plants. Int.Rev. Cytol. 132, 1–30.
Nishikawa, S., and Sasaki, F. (1996). Internalization of styryl dyeFM1-43 in the hair cells of lateral line organs in Xenopus larvae. J.Histochem. Cytochem. 44, 733–741.
O’Drisscoll, D., Wolson, G., and Steer, M. (1991). Lucifer yellowand fluorescein isothiocyanate uptake by cells or Morinda citrifoliain suspension cultures is not confined to the endocytic pathway.J. Cell Sci. 100, 237–241.
Oparka, K., Cole, L., Wright, K., Hawes, C., Evans, D., andColeman, J.O.D. (1991). Fluid phase endocytosis and the subcel-lular distribution of fluorescent probes in plant cells. In Endocy-tosis, Exocytosis and Vesicle Traffic in Plants, C. Hawes, J.Coleman, and D. Evans, eds (Cambridge, UK: Cambridge Univer-sity Press), pp. 81–102.
Orci, L., Tagaya, M., Amherdt, M., Perrelet, A., Donaldson, J.G.,Lippincott-Schwartz, J., Klausner, R.D., and Rothman, J.E.(1991). Brefeldin A, a drug that blocks secretion, prevents theassembly of non-clathrin-coated buds on Golgi cisternae. Cell 64,1183–1195.
Pimpl, P., Movafeghi, A., Coughlan, S., Denecke, J., Hillmer, S.,and Robinson, D.G. (2000). In situ localization and in vitro induc-tion of plant COPI-coated vesicles. Plant Cell 12, 2219–2236.
Robinson, D.G., Hinz, G., and Holstein, S.E. (1998). The molecularcharacterization of transport vesicles. Plant Mol. Biol. 38, 49–76.
Roszak, R., and Rambour, S. (1997). Uptake of lucifer yellow byplant cells in the presence of endocytotic inhibitors. Protoplasma199, 198–215.
Salzman, N.H., and Maxfield, F.R. (1988). Intracellular fusion of

86 The Plant Cell
sequentially formed endocytic compartments. J. Cell Biol. 106,1083–1091.
Salzman, N.H., and Maxfield, F.R. (1989). Fusion accessibility ofendocytic compartments along the recycling and lysosomalendocytic pathways in intact cells. J. Cell Biol. 109, 2097–2104.
Samuels, A., and Bialputra, T. (1990). Endocytosis in elongatingroot cells. J. Cell Sci. 97, 157–165.
Satiat-Jeunemaitre, B., and Hawes, C. (1992). Redistribution of aGolgi glycoprotein in plant cells treated with brefeldin A. J. CellSci. 103, 1152–1166.
Schote, U., and Seelig, J. (1998). Interaction of the neuronal markerdye FM1-43 with lipid membranes: Thermodynamics and lipidordering. Biochim. Biophys. Acta 1415, 135–146.
Shpetner, H., Joly, M., Hartley, D., and Corvera, S. (1996). Potentialsites of PI-3 kinase function in the endocytic pathway revealed bythe PI-3 kinase inhibitor, wortmannin. J. Cell Biol. 132, 595–605.
Steinmann, T., Geldner, N., Grebe, M., Mangold, S., Jackson,C.L., Paris, S., Galweiler, L., Palme, K., and Jurgens, G. (1999).Coordinated polar localization of auxin efflux carrier PIN1 byGNOM ARF GEF. Science 286, 316–318.
Sulzer, D., and Holtzman, E. (1989). Acidification and endosome-like compartments in the presynaptic terminals of frog retinal pho-toreceptors. J. Neurocytol. 18, 529–540.
Swanson, J.A. (1989). Phorbol esters stimulate macropinocytosisand solute flow through macrophages. J. Cell Sci. 94, 135–142.
Terryn, N., Anuntalabhochai, S., Van Montagu, M., and Inze, D.(1992). Analysis of a Nicotiana plumbaginifolia cDNA encoding anovel small GTP-binding protein. FEBS Lett. 299, 287–290.
Thiel, G., Kreft, M., and Zorec, R. (1998). Unitary endocytic andexocytic events in Zea mays L. coleoptile protoplasts. Plant J. 13,117–120.
Vida, T.A., and Emr, S.D. (1995). A new vital stain for visualizingvacuolar membrane dynamics and endocytosis in yeast. J. CellBiol. 128, 779–792.
Wee, E.G., Sherrier, D.J., Prime, T.A., and Dupree, P. (1998). Tar-geting of active sialyltransferase to the plant Golgi apparatus.Plant Cell 10, 1759–1768.
Wright, K., and Oparka, K. (1989). Uptake of lucifer yellow CH intoplant cell protoplasts: A quantitative assessment of fluid phaseendocytosis. Planta 179, 257–264.

DOI 10.1105/tpc.010339 2002;14;71-86Plant Cell
Neil Emans, Sabine Zimmermann and Rainer FischerUptake of a Fluorescent Marker in Plant Cells Is Sensitive to Brefeldin A and Wortmannin
This information is current as of April 11, 2019
References /content/14/1/71.full.html#ref-list-1
This article cites 77 articles, 37 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists