stereological estimation of the surface area and oxygen diffusing
TRANSCRIPT

Stereological Estimation of the Surface Area andOxygen Diffusing Capacity of the Respiratory Stomachof the Air-breathing Armored Catfish Pterygoplichthysanisitsi (Teleostei: Loricariidae)
Andre Luis da Cruz, Ana Carolina Elias Pedretti, and Marisa Narciso Fernandes*
Department of Physiological Sciences, Federal University of Sao Carlos, Caixa Postal 676,13565-905 Sao Carlos, SP, Brazil
ABSTRACT The stomach of Pterygoplichthys anisitsihas a thin, translucent wall and a simple squamous epi-thelium with an underlying dense capillary network. Inthe cardiac and pyloric regions, most cells have shortmicrovilli distributed throughout the cell surface andtheir edges are characterized by short, densely packedmicrovilli. The mucosal layer of the stomach has twotypes of pavement epithelial cells that are similar tothose in the aerial respiratory organs. Type 1 pavementepithelial cells, resembling the Type I pneumocyte inmammal lungs, are flat, with a large nucleus, andextend a thin sheet of cytoplasm on the underlying capil-lary. Type 2 cells, resembling the Type II pneumocyte,possess numerous mitochondria, a well-developed Golgicomplex, rough endoplasmic reticulum, and numerouslamellar bodies in different stages of maturation. Thegastric glands, distributed throughout the mucosal layer,also have several cells with many lamellar bodies. Thetotal volume (air 1 tissue), tissue, and air capacity ofthe stomach when inflated, increase along with bodymass. The surface-to-tissue-volume ratio of stomachvaries from 108 cm21 in the smallest fish (0.084 kg) to59 cm21 in the largest fish (0.60 kg). The total stomachsurface area shows a low correlation to body mass.Nevertheless, the body-mass-specific surface area variedfrom 281.40 cm2 kg21 in the smallest fish to 68.08 cm2
kg21 in the largest fish, indicating a negative correlationto body mass (b 5 20.76). The arithmetic mean barrierthickness between air and blood was 1.52 6 0.07 lm,whereas the harmonic mean thickness (sh) of the diffusionbarrier ranged from 0.40 to 0.74 lm. The anatomical diffu-sion factor (ADF 5 cm2 lm21 kg21) and the morphologicalO2 diffusion capacity (DmorpholO2 5 cm3 min21 mmHg21
kg21) are higher in the smallest specimen and lower inthe largest one. In conclusion, the structure and morpho-metric data of P. anisitsi stomach indicate that this organis adapted for oxygen uptake from air. J. Morphol.270:601–614, 2009. � 2008 Wiley-Liss, Inc.
KEY WORDS: air-breathing; stomach; surface area;barrier thickness; oxygen diffusion capacity
INTRODUCTION
Many fish use accessory organs to breathe airduring some period or phase of their life to supplytheir oxygen needs, although the gills are the
main gas exchange organs in teleost species. Atleast 49 families of teleost fish are known to haveair-breathing species (Graham, 1997), most ofwhich live in stagnant or shallow tropical and sub-tropical waters, employing this mode of respirationwhen exposed to unfavorable conditions for aquaticrespiration or in response to increased oxygenrequirements.
Lungfish (Neoceratodus, Lepidosiren) and basalactinopterygeans such as Protoptherus use lungs(Maina and Maloiy, 1985; Kemp, 1987; Moraeset al., 2005) and a variety of teleosts such as Ara-paima (Sawaya, 1946), Gymnotus (Liem, 1989),Hoplerythrinus (Kramer, 1978; Mattias et al.,1996), and Piabucina (Graham, 1997) use theswim-bladder for air respiration. The wide varietyof accessory air-breathing organs found among thehigher teleosts reflects the loss of the pneumaticduct during evolution (Graham, 1997).
In the latter group, most accessory air-breathingorgans are located in the region of the head andconsist of the buccal and pharynx epithelia, as inElectrophorus, Monopterus, Blennius, and Gobios,pharyngeal pouches in Channa and Monopterusand modified branchial and opercular surfaces inClarias and Anabas (Munshi, 1985). However,some air-breathing structures are located in thedigestive tract (Carter and Beadle, 1931; Geeand Graham, 1978; Silva et al., 1997; Satora,1998; Podkowa and Goniakowska-Witalinska,1998, 2002).
Respiratory structures along the digestive tractimply regional modifications of this tract for oxy-
Contract grant sponsors: FAPESP, CNPq, and CAPES.
*Correspondence to: Marisa Narciso Fernandes, Department ofPhysiological Sciences, Federal University of Sao Carlos, CaixaPostal 676, 13565-905 Sao Carlos, SP, Brazil.E-mail: [email protected]
Published online 23 December 2008 inWiley InterScience (www.interscience.wiley.com)DOI: 10.1002/jmor.10708
JOURNAL OF MORPHOLOGY 270:601–614 (2009)
� 2008 WILEY-LISS, INC.

gen uptake from air, including high vasculariza-tion and a thin diffusion distance between theexternal and internal medium. In the early 20thcentury, Carter and Beadle (1931) reported insome loricariid species a transparent, richly vascu-larized stomach that was permanently filled withair. Crawford (1974) observed the use of the esoph-agus as an air-breathing organ in Dalia pectorisand Podkowa and Goniakowska-Witalinska (2002)described the ultrastructure of the posterior intes-tine of Corydoras, emphasizing its adaptation toair breathing.
Although the functional morphologicalapproach, together with morphometric data, are aprerequisite for understanding the function of agiven organ, most morphofunctional studies of theaccessory respiratory organs in air-breathing fishare so far restricted to some amphibious mudskip-pers (Biswas et al., 1981), Indian fishes (Hugheset al., 1974a,b; Munshi, 1985), and lungfishes(Hughes and Weibel, 1976; Maina and Maloy,1985; Moraes et al., 2005). Recently, histologicaland electron microscopical studies revealed a cap-illary network present in the lamina propria ofthe mucosal layer of the stomach, subjacent tothe epithelium (Silva et al., 1997; Satora, 1998;Satora and Winnicki, 2000; Oliveira et al., 2001;Podkowa and Goniakowska-Witalinska, 2003).Satora and Winnicki (2000) described the ultra-structure of Ancistrus multispinnis stomach,showing a dense network of capillaries, and Pod-kowa and Goniakowska-Witalinska (2003) deter-mined the capillary density per 100 lm of the mu-cosal epithelium length and the thickness of theair-blood barrier in the stomach of Hypostomusplecostomus. However, the effective respiratorysurface area and oxygen diffusion capacity haveyet to be determined.
The main purpose of the present study was toapply stereological principles to estimate the respi-ratory surface area, the air-blood barrier thicknessand the morphometric oxygen diffusion capacity ofthe stomach of the air-breathing loricariid fishPterygoplichthys anisitsi. Pterygoplichthys anisitsiis an armored catfish occurring in South Americanfreshwaters and is found throughout the Uruguay,Paraguay, and Parana River basins where itinhabits water bodies characterized by low oxygenconcentrations. In normoxia, this species does notbreathe air during the day, but at night itincreases its activity and usually swims to the sur-face. Under hypoxia, P. anisitsi breathes air evenduring the daytime (Cruz, 2007). Its stomach isusually full of air and sometimes contains smallfood particles. Preliminary observations underlight microscopy showed a single epithelial cell inthe mucous membrane of the stomach and a highdensity of capillaries close to this membrane (Oli-veira et al., 2001), suggesting an adaptation to airbreathing.
MATERIALS AND METHODSAnimals
Five armored catfish Pterygoplichthys anisitsi Eigenmannand Kennedy [body mass (MB) 5 0.084–0.600 kg; total length(LT) 5 24–39 cm] were obtained from the Aquaculture Center ofthe Sao Paulo State University (CAUNESP) at Jaboticabal, SaoPaulo, Brazil. In the laboratory, the fish were kept in a 1000 Laquarium with continuous water flow and aeration at 23–258Cand were fed with commercial fish food and Lactuta sativaleaves.
Anesthesia, Fixation, Tissue Sampling, andProcessing
The fish were irreversibly anesthetized with 0.1% benzo-caine1 (Sinth) and the stomach was exposed through a ventralabdominal incision. The stomach was prefixed in situ with a fix-ative solution containing 2.5% glutaraldehyde in 0.1 M phos-phate buffer pH 7.4, 300 mOsm, at 48C. The air volume insideits lumen was measured and replaced with the same volume offixative solution, slowly introduced into the stomach using athin polyethylene tube through the buccal cavity and esophagusto permit replacement of air bubbles and avoid tissue stretchingor distention. After tying off the esophagus and duodenum, thestomach was removed and immersed in the fixative solution forcomplete fixation at 48C.
The stomach was then opened ventrally through an antero-posterior incision and the major anteroposterior and transverselengths were measured. Tissue was taken from the open stom-ach according to systematic random sampling (Fig. 1a) using asquare-lattice grid. The sampling sites were chosen by countingthe number (N) of points lying over the open stomach andchoosing every 6th/N point to sampling tissue for light and ev-ery 7th/N point for electron microscopy and marked by a pinprick. The first sampling site was chosen at random from thefirst six potential sites. To obtain vertical sections (VUR) anduse the same section to estimate the fraction volume of stom-ach tissue, as well as surface density and total surface area,the external surface of the open stomach (serosa side) wasconsidered the horizontal plane. The embedding procedureswere designed so that the sections from each sample weredone in a sequential 1368 rotation relative to the previousone, with the first one sectioned at random between 0 and 368(Fig. 1b,c).
The samples for light microscopy (LM) were dehydrated, em-bedded in Historesin1 (Leica, Heidelberg, Germany) and sec-tions of 3-lm thickness were stained with toluidine blue andfuchsin. Additional samples from cardiac, corpus, and pyloricregions of stomach were dehydrated, embedded in paraffin andsections of 5-lm thickness were stained with periodic acid Schiffand Alcian blue pH 2.5.
The samples for transmission electron microscopy (TEM)analysis were postfixed in 1% osmium tetroxide in the samebuffer as before, block-stained using uranyl acetate, dehydratedin an ascending concentration of acetone, and embedded inAraldite 6005 Resin (Ladd, Burlington, VT). Semithin sections(0.5 lm) were stained with toluidine blue and the ultrathin sec-tions (60 nm) were placed on 200-mesh copper grids, stainedwith uranyl acetate and lead citrate and viewed in a JEOLJEM 100 CX TEM at 80 kV.
For scanning electron microscopy (SEM) analysis, the stom-achs of three other specimens were fixed as described above,ventrally opened through a median anteroposterior incision andsamples from the cardiac, blind-sac (corpus), and pyloric por-tions were removed, dehydrated through ascending concentra-tions of ethanol, soaked in two successive baths of 1,1,1,3,3,3-hexamethyldisilazane (Aldrich), and air dried. The sampleswere glued with silver paint onto the specimen stub, coatedwith 99% gold (Degussa S.A) in a vacuum sputter and exam-ined under a ZEISS-DSM 940A SEM at 25 kV.
602 A.L. DA CRUZ ET AL.
Journal of Morphology

Density Volume, Surface-to-Volume Ratio,and Surface Area of Stomach
All the LM data were obtained with an Olympus BX51 lightmicroscope, the images were captured with a JVC-TKC1380digital video camera and analyzed using version 2.00 of theCAST System software (Olympus Denmark S/A).The reference volume of stomach tissue (VSt) was calculated
based on the gross surface area of opened stomach by pointcount of an isotropic test system superimposed on the mucosaand the stomach wall thickness obtained from vertical sections.The differential tissue volumes of stomach were estimated
using the same sections that were used for the stomach surfacearea (SSt) at a final magnification of 15003. The differentialvolumes or density volume [VV (structure, reference)] of themain stomach (St) tissues (mucosa, submucosa, muscularis,serosa) were estimated by point count of an isotropic test-sys-
tem, as described by Howard and Reed (1998), and was givenas VV (structure,Stomach) 5 (PS 100)/PSt, where PS was thenumber of points on the analyzed structure or tissue (S) of thestomach and PSt was the total number of points on the stomach(St) (Fig. 2a). The density volumes of mucosa components (epi-thelium, capillaries, gastric gland) were estimated using thesame methodology, but at higher magnification (38003).
The stomach surface area (SSt) was estimated from its respec-tive surface-area-to-volume ratios (Sv), multiplied by the previ-ously determined stomach volume as SSt 5 Sv VSt, according toHoward and Reed (1998). The surface area-to-volume ratio (Sv)was estimated using a cycloid arch test system (Fig. 2a) as
Svðstructure; referenceÞ ¼ 2Pn
i¼1Ii
L; where Ii was the number of
intersections of the test lines with the surface of mucosal sur-face of stomach and L was the length of test lines falling withinthe reference space: mucosa profile. L was estimated by multi-
Fig. 1. Diagram of the sample scheme of stomach tissue of Pterygoplichthys anisitsi. (a) Systematic random samples from openstomach superimposed by a point square grid to obtain vertical uniform random samples. The tissue bottom is the serosa side (s) ofstomach and was fixed as the horizontal plane (HP) and the vertical axis (VA) is indicated. m, mucosa. (b) Each tissue sample wassequentially rotated on the vertical axis (VA) and embedded. (c) Vertical section obtained of embedded tissue. m, mucosa; s, serosa.Arrow indicates the vertical axis (VA).
Fig. 2. (a) A cycloid arc test array superimposed on the microscopic image of a light micrograph in vertical section of the stom-ach of Pterygoplichthys anisitsi for the evaluation of tissue differential volumes and the surface-area-to-volume ratio. The arrowindicates the vertical axis of measurements. M, mucosa side; S, serosa side of stomach. (b) A cycloid arc test array superimposedon the microscopic image of TEM micrograph in vertical section of the stomach of P. anisitsi for the measurements of barrier thick-ness. For illustration, the intersection of the cycloid with the surface of mucosal layer of stomach was the starting point of mea-surement of barrier thickness and the line direction were the direction-number between 1 and 97 (sine-weighted line) obtainedfrom a random number table (white line).
STOMACH OF THE AIR-BREATHING ARMORED CATFISH 603
Journal of Morphology

plying the point count for the reference space by the test linelength per point.
Estimation of Air-Blood Barrier Thickness
Ultrathin sections from five stomach samples per animalwere used to estimate the air-blood barrier thickness of thestomach. Subsampling of ultrathin stomach sections was basedon the systematic quadrant procedure. Starting at a randomposition in the top left-hand corner of the TEM grid, five quad-rants of ultrathin section were sampled from each grid and tis-sues were photographed at a magnification of 3,3003. The har-monic mean barrier thickness (s) was measured with a logarith-mic ruler and sine-weighted lines generated on the VURsections (Gundersen et al., 1988). A new angle was selected foreach sampled quadrant.Negative film was viewed in a light box superimposed with a
cycloid arch test system and a sine-weighted line to score pointcounts on the barrier (epithelial and endothelial cells and basallamina) and air-blood barrier thickness (Fig. 2b). Each intersec-tion of cycloid arc with surface of mucosal layer of stomach wasthe point to measure air-blood barrier distance and the linedirection were the direction-number between 1 and 97 obtainedfrom a random number table. The harmonic mean barrier thick-ness (Weibel and Knight, 1964) was calculated for each animalas sh 5 2/3lh, where lh is the harmonic mean of the lengths.
Anatomical Diffusion Factor andMorphometric Diffusion Capacity
The anatomical diffusion factor (ADF) was estimated directlyfrom the stereological data, dividing the respiratory surface areain relation to body mass (SR/MB) by the harmonic mean barrierthickness (sh) (Perry, 1989). The morphometric diffusion capacity(Dmorphol) was calculated as the product of the ADF and Krogh’sdiffusion coefficient (K) of the respective cell layers (epithelium,basal membrane, and endothelium) of the diffusion barrier.Since the value of K depends on the specific gas (O2 or CO2),
the temperature and tissue type, the K value for rat lungs wasused for the epithelium and endothelium, and the K value forfrog connective tissue (Bartels, 1971) was used for basal lamina.The relative volume of the epithelial and endothelial cells wasestimated separately from that of basement membrane by pointcounting, and each element was multiplied by the appropriateK value. This weighted numerical ratio yielded a K value for ox-ygen (KO2
) in the air-blood barrier of stomach. All values werecorrected to 258C.
Statistics
Descriptive statistics were drawn up with Microsoft’s Excelspreadsheet software. The data are presented as the mean val-ues accompanied by the respective standard errors (SEM). Sincethe standard deviation of a harmonic mean is not defined, theSEM was calculated from the arithmetic mean of the diffusiondistance of air-blood barrier of stomach.Regression analyses were made to seek possible significant cor-
relations, which were calculated by the least-squares method,and the correlation coefficient (r2) was estimated to determinethe goodness of fit. The data were log transformed and the powerequation: log Y 5 log a 1b log X was used to calculate the scal-ing exponent (b), resulting in the allometric equation.
RESULTSStomach Morphology
Pterygoplichthys anisitsi has a short esophagusthat opens into a large U-shaped stomach with athin, translucent wall (Fig. 3a). The stomach,
highly vascularized and expanded with air bubbles(Fig. 3a,b), occupies most of the volume of the coe-lomic cavity and was found empty in the all speci-mens dissected, although the intestine was full ofmaterial. Continuous to the stomach there is along, thin, transparent intestine located ventrallyto the stomach and forming several loops.
Figure 3c shows the four layers of the stomach:(1) the serosa, the outermost layer, which is char-acterized by a single layer of squamous epithelialcells with underlying loose connective tissue con-taining blood vessels; (2) the muscularis, composedof circular smooth muscles and discontinuous thickbundles of smooth muscles with longitudinal andoblique orientation among small amounts of con-nective tissue; (3) the submucosa, consisting ofconnective tissue and blood vessels of differentdiameters; (4) the mucosa, composed of the laminapropria containing small blood vessels, a singlelayer of flat epithelial cells whose apical surfacesare in contact with the stomach lumen, and thebasement membrane with numerous underlyingblood capillaries (Fig. 3d). Gastric glands arelocated in the mucous layer (Fig. 3c,d).
A SEM analysis of the mucosal surface of Ptery-goplichthys anisitsi stomach, under low magnifica-tion, showed irregular mucosal folds in the cardiacregion, longitudinal folds in the corpus region andan almost total absence of folds in the pyloricregion (Fig. 4a,c, and e). Under high magnification,the epithelial pavement cells of the mucosal stom-ach are visibly polygonal in shape. In the cardiacand pyloric regions, most of the cells have shortmicrovilli irregularly distributed throughout thecell surface and the boundaries of these cells arecharacterized by highly dense short microvilli;some cells show a smooth cell surface, in whichcase the cell boundary is characterized by short,continuous microridges (Fig. 4b,f). In the corpusregion, the cell surfaces are smooth and the celllimits are not clearly visible (Fig. 4c). Numerousgastric gland openings are distributed among cellsin all the stomach regions, but the frequency ofgastric glands is lower in the corpus region (Fig.4c,d).
The pavement epithelial cells of the stomach’smucosal layer are flat and have large nuclei, gen-erally located between adjacent capillaries (Fig.5a) or, rarely, elongated and located on the capilla-ries immediately below the basal lamina (Fig. 5b).The first cell type spreads a thin sheet of cyto-plasm on the underlying capillary thus resemblingthe Type I pneumocyte of the mammalian lung(Fig. 5a,c). The latter cell type possesses numerousmitochondria, a well-developed Golgi complex,rough endoplasmic reticulum, and free ribosomes(Fig. 6a). Some cells contain numerous lamellarbodies in different stages of maturation thereforeresembling the Type II pneumocyte (Fig. 6b,c) andnumerous vesicles close to the cellular membrane
604 A.L. DA CRUZ ET AL.
Journal of Morphology

(Fig. 6c). Some lamellar bodies have a centralregion composed of highly electron-dense amor-phous material from which emerges a system ofconcentric membranes (Fig. 6b,c).
The junctional complex between two pavementepithelial cells shows a tortuous boundary becauseof interdigitations of each cell with its neighbor,and is characterized by a short zonula occludensand zonula adherens, generally with only one des-mosome (Figs. 5b,c, and 6c). The basal lamina con-tacts the endothelial cells of capillaries, except inthe regions where the gastric glands are present.Endothelial cells have an elongated heterochro-matic nucleus protruding into the capillary lumen(Fig. 7a). Adjacent endothelial cells may overlap,showing short infolding and usually one desmo-
some was identified in the cell junctions. Pores ofdifferent diameters are visible in the endotheliumlining the apical and lateral parts of capillaries(Fig. 5c). The air-blood barrier that separates thegastric lumen from capillaries consists of threelayers, the cytoplasmic extension from epithelialcells, the basal lamina, and the sheet of endothe-lial cells (Figs. 5c and 6a,b).
Bundles of collagen fibers extending in differentdirections fill up the space between cells, bloodvessels, and capillaries and the gastric glands thatconstitute the mucosa and submucosa (Fig. 7a);collagen fibers are found among tissues that con-stitute the other layers of stomach wall.
The gastric glands are regularly distributedthroughout the mucosa, being more frequent in
Fig. 3. (a) Ventral view of the stomach of Pterygoplichthys anisitsi full of air showing its anatomic regions: cardia, corpus, andpylorus. E, esophagus; I, intestine. Scale bar in cm. (b) Detail of highly vascularized stomach wall. Note large (arrowhead) andsmall (*) blood vessels. Scale bar in mm. (c) Cross section of stomach wall showing the four stomach layers, the serosa (1), muscu-laris (2), submucosa (3), and mucosa (4). Note gastric glands (G) in the mucosal layer. m, muscles. Scale bar in lm. (d) Mucosaland submucosal layers of stomach wall showing capillaries (*) underlying the epithelial cells of mucosal layer and the gastricglands (G). Small blood vessels (v) and connective tissue (c) are in the submucosal layer. Scale bar in lm.
STOMACH OF THE AIR-BREATHING ARMORED CATFISH 605
Journal of Morphology

the cardiac and pyloric regions, and may have around or pyramidal shape. No morphological dif-ferences were found among the gastric glands fromthe three stomach regions. Although they are con-tinuous with the epithelium, these glands nestle inthe lamina propria and are surrounded by collagen
fibers (Fig. 7a). The glands have 8–12 cells andeach cell has a heterochromatic basal nucleus, asmall number of cytoplasm organelles, and the api-cal region of these cells has numerous smallvesicles; several cells have numerous lamellarbodies (Fig. 7a,b). The junctional complex between
Fig. 4. Stomach regions of Pterygoplichthys anisitsi. SEM. Low magnification of cardiac (a), corpus (c), and pyloric (e) regions.The arrowheads indicate the opening of gastric glands. Scale bar in lm. High magnification of the epithelium of the mucous layershowing the pavement cell surface architecture from the cardiac (b), corpus (d), and pyloric (f) regions of stomach. Note the cellsurface with short microvilli (*) in the (b) and (f) and a smooth surface in (d) (*). The arrowhead indicates the gastric gland openingin each stomach region and the double arrowhead the epithelial cell limits. Scale bar in lm.
606 A.L. DA CRUZ ET AL.
Journal of Morphology

Fig. 5. Stomach mucosa of Pterygoplichthys anisitsi. TEM. (a) Pavement epithelial cell showing the cell nucleus between two ad-jacent blood capillaries. BL, basal lamina; C, capillary; EnC, endothelial cell; Er, erythrocyte; n, nucleus; PVC, pavement epithelialcell. (b) Pavement epithelial cell showing the cell nucleus on the blood capillary. Note the junctional complex (arrow) between pave-ment cells. BL, basal lamina; C, capillary; EnC, endothelial cell; m, mitochondria; n, nucleus; PVC, pavement epithelial cell. (c)Pavement epithelial cell showing the junctional complex (arrow) between cells and microvilli (double arrowheads) in the apical cellsurface and the endothelial cell containing numerous pores (arrowheads). BL, basal lamina; C, capillary; EnC, endothelial cell; L,leukocyte; PVC, pavement epithelial cell. Scale bar in lm.
STOMACH OF THE AIR-BREATHING ARMORED CATFISH 607
Journal of Morphology

these cells is characterized by intense interdigita-tion and several desmosomes. The negative stain-ing with PAS and Alcian blue indicated that theseglands do not produce acid and neutral mucopoly-saccharides. Rare endocrine cells are found to-gether with the gastric glands and are character-ized by electron-lucent cytoplasm with electron-dense vesicles (Fig. 7b,c).
Stomach Morphometry
The mean anteroposterior and transversallength of stomach of Pterygoplichthys anisitsi in0.084–0.60 kg-fish were 6.5 6 0.87 and 5.03 60.61 cm, respectively. Of the 20.245 6 4.747 cm3
which corresponds to the stomach’s total mean vol-ume (air 1 tissue), only 0.425 6 0.080 cm3 repre-sents the stomach tissue, and the total stomachvolume to body mass ratio is 85.21 6 14.09 cm3
kg21. The volume of stomach tissue, air volumecapacity and the total volume (air 1 tissue) ofinflated stomach (Table 1) increased with bodymass, whereas the specific stomach volume (VSt
M�1B ) decreased according to the power equation V
5 aMbB, where V is the tissue (ts,St), air (air,St), or
total (St) volumes of stomach, MB is the body
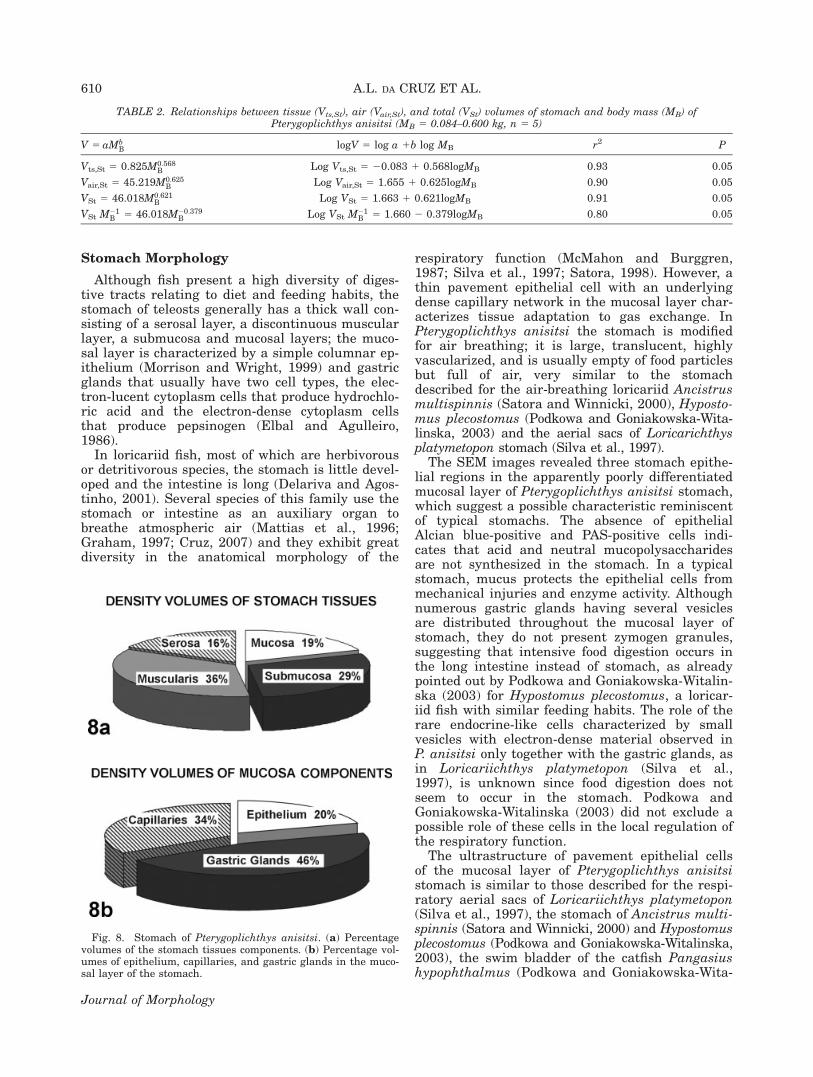
mass, a is the value for 1 g fish and b is the scal-ing or mass coefficient (Table 2). In the log form:log V 5 log a 1 b logMB, the stomach volumedescribes a straight line where a is the interceptand b is the slope (Table 2). The percentage of vol-ume densities of stomach tissue layers of P. ani-sitsi is depicted in Figure 8a. The muscle layersrepresent the major volume density of stomach tis-sue, followed by the submucosa, mucosa andserosa layers. Considering only the mucosa to-gether with the lamina propria, the capillaries andthe gastric glands represent 34 and 46%, respec-tively, of the volume of this layer (Fig. 8b).
The surface-to-tissue-volume ratio of Pterygo-plichthys anisitsi stomach varies from 108 cm21 inthe smallest fish (0.084 kg) to 59 cm21 in the larg-est fish (0.60 kg). The mean total stomach surface(SSt) (Table 3) increased with body mass, butshowed a low correlation with body mass (SSt 542.01 M0:23
B , r2 5 0.33). Nevertheless, the specificsurface area (SSt M�1
B ) varied from 281.40 cm2
kg21 in the smallest fish to 68.08 cm2 kg21 in thelargest one, showing a negative correlation with bodymass (SSt M
�1B 5 42.02 M�0:76
B , r2 5 0.84, P < 0.05).The mean value of barrier thickness between air
and blood (arithmetic mean) was 1.52 6 0.07 lm,
Fig. 6. Stomach mucosa of Pterygoplichthys anisitsi showing pavement epithelial cell resembling the Type II pneumocyte. TEM.(a) Cytoplasm organelles in the pavement epithelial cell. BL, basal lamina; EnC, endothelial cell; Er, erythrocyte; g, Golgi complex,m, mitochondria. (b) Lamellar bodies (*) with concentric membrane configuration. BL, basal lamina; C, capillary; EnC, endothelialcell. (c) Numerous vesicles (arrowhead) close to the apical surface of cell membrane. Note the presence of lamellar body (*) andshort microvilli (double arrowheads) on the cell limit. m, mitochondria. Scale bar in lm.
608 A.L. DA CRUZ ET AL.
Journal of Morphology

whereas the harmonic mean thickness (sh) of thediffusion barrier ranged from 0.40 to 0.74 lm. Thespecific anatomical diffusion factor (ADF 5 SSt s�1
h
M�1B ) and the morphological O2 diffusion capacity
were higher in the smallest specimen and lower inthe largest one (Table 3).
DISCUSSION
This is the first morphometric study of the stom-ach of an air-breathing fish, with special emphasison the respiratory function. Our results show themorphological adaptation of Pterygoplichthys ani-sitsi stomach to gas exchange. We estimated therespiratory surface area of this organ and the mor-phometric diffusion capacity for oxygen.
Fig. 7. (a) Gastric gland opened (arrowhead) in the mucosal layer of stomach of Pterygoplichthys anisitsi. Note the gastric glandorganization, nucleus position, and numerous lamellar bodies (*) in these cells. Arrow indicates collagen fibers. C, capillary; EnC,endothelial cell; Er, erythrocyte; n, nucleus. (b) Part of gastric gland (G) in the submucosal layer of stomach showing endocrine cell(*). (c) High magnification of endocrine cell showing electron-dense bodies (arrowheads) distributed in the cytoplasm and close tothe cell membrane. EndC, endocrine cell. Scale bar in lm.
TABLE 1. Range and mean (6SEM) of the body mass andlength, the air (Vair,St), tissue (Vts,St), and total (VTotal,St) volumeof stomach and total specific stomach volume (VTotal,St M
�1B ) of
Pterygoplichthys anisitsi (n 5 5)
Range Mean SEM
Body mass (kg) 0.084–0.600 0.285 0.096Body length (cm) 23–42 31.8 3.39Vair,St (cm
3) 11.3–35.0 19.320 4.692Vts,St (cm
3) 0.225–0.611 0.425 0.080VSt (cm
3) 11.519–35.611 20.245 4.747VSt M
�1B (cm3 kg21) 137.13–59.35 85.21 14.09
STOMACH OF THE AIR-BREATHING ARMORED CATFISH 609
Journal of Morphology
Stomach Morphology
Although fish present a high diversity of diges-tive tracts relating to diet and feeding habits, thestomach of teleosts generally has a thick wall con-sisting of a serosal layer, a discontinuous muscularlayer, a submucosa and mucosal layers; the muco-sal layer is characterized by a simple columnar ep-ithelium (Morrison and Wright, 1999) and gastricglands that usually have two cell types, the elec-tron-lucent cytoplasm cells that produce hydrochlo-ric acid and the electron-dense cytoplasm cellsthat produce pepsinogen (Elbal and Agulleiro,1986).
In loricariid fish, most of which are herbivorousor detritivorous species, the stomach is little devel-oped and the intestine is long (Delariva and Agos-tinho, 2001). Several species of this family use thestomach or intestine as an auxiliary organ tobreathe atmospheric air (Mattias et al., 1996;Graham, 1997; Cruz, 2007) and they exhibit greatdiversity in the anatomical morphology of the
respiratory function (McMahon and Burggren,1987; Silva et al., 1997; Satora, 1998). However, athin pavement epithelial cell with an underlyingdense capillary network in the mucosal layer char-acterizes tissue adaptation to gas exchange. InPterygoplichthys anisitsi the stomach is modifiedfor air breathing; it is large, translucent, highlyvascularized, and is usually empty of food particlesbut full of air, very similar to the stomachdescribed for the air-breathing loricariid Ancistrusmultispinnis (Satora and Winnicki, 2000), Hyposto-mus plecostomus (Podkowa and Goniakowska-Wita-linska, 2003) and the aerial sacs of Loricarichthysplatymetopon stomach (Silva et al., 1997).
The SEM images revealed three stomach epithe-lial regions in the apparently poorly differentiatedmucosal layer of Pterygoplichthys anisitsi stomach,which suggest a possible characteristic reminiscentof typical stomachs. The absence of epithelialAlcian blue-positive and PAS-positive cells indi-cates that acid and neutral mucopolysaccharidesare not synthesized in the stomach. In a typicalstomach, mucus protects the epithelial cells frommechanical injuries and enzyme activity. Althoughnumerous gastric glands having several vesiclesare distributed throughout the mucosal layer ofstomach, they do not present zymogen granules,suggesting that intensive food digestion occurs inthe long intestine instead of stomach, as alreadypointed out by Podkowa and Goniakowska-Witalin-ska (2003) for Hypostomus plecostomus, a loricar-iid fish with similar feeding habits. The role of therare endocrine-like cells characterized by smallvesicles with electron-dense material observed inP. anisitsi only together with the gastric glands, asin Loricariichthys platymetopon (Silva et al.,1997), is unknown since food digestion does notseem to occur in the stomach. Podkowa andGoniakowska-Witalinska (2003) did not exclude apossible role of these cells in the local regulation ofthe respiratory function.
The ultrastructure of pavement epithelial cellsof the mucosal layer of Pterygoplichthys anisitsistomach is similar to those described for the respi-ratory aerial sacs of Loricariichthys platymetopon(Silva et al., 1997), the stomach of Ancistrus multi-spinnis (Satora and Winnicki, 2000) and Hypostomusplecostomus (Podkowa and Goniakowska-Witalinska,2003), the swim bladder of the catfish Pangasiushypophthalmus (Podkowa and Goniakowska-Wita-
TABLE 2. Relationships between tissue (Vts,St), air (Vair,St), and total (VSt) volumes of stomach and body mass (MB) ofPterygoplichthys anisitsi (MB 5 0.084–0.600 kg, n 5 5)
V 5 aMbB logV 5 log a 1b log MB r2 P
Vts,St 5 0.825M0:568B Log Vts,St 5 20.083 1 0.568logMB 0.93 0.05
Vair,St 5 45.219M0:625B Log Vair,St 5 1.655 1 0.625logMB 0.90 0.05
VSt 5 46.018M0:621B Log VSt 5 1.663 1 0.621logMB 0.91 0.05
VSt M�1B 5 46.018M�0:379
B Log VSt M�1B 5 1.660 2 0.379logMB 0.80 0.05
Fig. 8. Stomach of Pterygoplichthys anisitsi. (a) Percentagevolumes of the stomach tissues components. (b) Percentage vol-umes of epithelium, capillaries, and gastric glands in the muco-sal layer of the stomach.
610 A.L. DA CRUZ ET AL.
Journal of Morphology

linska, 1998), and the lungfish lungs (Maina andMaloiy, 1985; Moraes et al., 2005). Most of the epi-thelial cells of P. anisitsi stomach resemble theType I pneumocyte of mammal lungs and some ofthem present characteristics of the Type II pneu-mocyte, exhibiting lamellar bodies in their cyto-plasm. Lamellar bodies have been described in epi-thelial cells of the air-breathing organs of fish suchas the stomach (Silva et al., 1997; Satora, 1998),intestine (Podkowa and Goniakowska-Witalinska,2002), and swim bladder (Podkowa and Goniakow-ska-Witalinska, 1998) without distinction amongepithelial cell types. This finding opens a questionabout the derivative characteristic of amniotes asthese cell types have been described in the lungepithelium of higher vertebrates. The lung is a de-rivative of the gut (foregut endoderm) and the la-mellar bodies have been described as the site ofsurfactant accumulation and are highly morpho-logically and biochemically conserved throughoutthe radiation of air-breathing organs among verte-brates (Daniels et al., 2004). Further biochemicaland physiology studies are needed to establishsimilarity, beyond considering only morphologicalcharacteristics. In P. anisitsi lamellar bodies arealso found distributed in the cytoplasm of gastricgland cells.
Lamellar bodies have been described as the siteof surfactant accumulation (Daniels et al., 2004)that is discharged by exocytosis into the respira-tory organ lumen. The thin film of this substanceover the luminal surface of the respiratory organreduces the forces of attraction between the watermolecules at the surface film, thereby decreasingthe surface tension and favoring oxygen absorption(Orgeig and Daniels, 2001). Furthermore, this sub-stance may be involved in the antimicrobialdefense mechanisms of respiratory organs (Danielset al., 1995; Rubio et al., 1996) and prevent epithe-lial desiccation and oxidative damage of epithelialcells (Daniels and Orgeig, 2001).
The structure of the air-blood diffusion barrierof respiratory epithelium of the stomach of Ptery-goplichthys anisitsi consisted of the same struc-tures as the respiratory epithelium of the lungs ofother vertebrates. The triple membrane model (ep-ithelium, basal membrane, and endothelium) of
the diffusion barrier characterizes the air-bloodbarrier of respiratory organs of vertebrates (Mainaand West, 2005) and the accessory organs for airbreathing in fish. Most endothelial cells of thecapillaries underlying the basal membrane may beclassified as continuous and are characteristic ofthe respiratory organs; fenestrated capillaries thatare characteristic capillaries of the digestive tractare also observed in the stomach of P. anisitsi asin Hypostomus (Podkowa and Goniakowska-Wita-linska, 2003) and in the respiratory regions of theintestine of Corydoras (Podkowa and Goniakow-ska-Witalinska, 2002). However, independent ofthe type of capillary underlying the mucosal epi-thelium of the stomach, the distance between airand blood is similar to the distance in the lungs ofaerial vertebrates, indicating that the stomach isthe auxiliary organ for breathing air in P. anisitsi.
Stomach Morphometry
The air volume of stomach (Vair,St) of Pterygo-plichthys anisitsi between 11.3 and 35.0 cm3 forfish weighing from 84 to 600 g and with 23 to42 cm length is congruent with that reported byGee (1976) for other loricariid fish, and also showsa positive correlation between body mass and air(Vair,St), tissue (Vts,St) and total (VSt) stomach vol-ume. A negative correlation between body massand specific total volume (VSt M
�1B ), as expected, is
consistent with most biological parameters in ani-mals.
The low percentage volume of the mucosal andserosal layers of stomach and the high volumedensity of the capillary underlying the epitheliumin the mucosal layer are similar to those found inthe tissue volumes of the respiratory organs ofother animals. In lungs, the volume of parenchy-mal tissue specialized in gas exchange (respiratoryepithelium) occupies a low volume density (Burriet al., 2003) and the capillary density is higherthan the density of epithelium (Burri et al., 2003;Moraes et al., 2005).
The potential efficiency of the Pterygoplichthysanisitsi stomach as a respiratory organ is highconsidering the simplicity of its structure. In theSouth American lungfish Lepidosiren paradoxa
TABLE 3. Range and mean values (6SEM) of the surface to volume ratio (SvSt), the surface area (SSt), specific surface area ofstomach (SSt M
�1B ), arithmetic mean of the air-blood diffusion barrier (sarith), harmonic mean (sh), anatomic diffusion factor (ADF)
and morphometric diffusion capacity for O2 (DmorpholO2) of stomach of Pterygoplichthys anisitsi (n 5 5)
Range Mean SEM
Sv,St (cm21) 58.62–107.79 77.14 8.33
SSt (cm2) 19.80–40.85 30.76 4.15
SSt M�1B (cm2 kg21) 68.08–281.40 157.21 45.34
sarith (lm) 1.36–1.82 1.52 0.07sh (lm) 0.40–0.74 — —ADF (cm2 lm21 kg21) 103.15–703.57 333.95 102.06DmorpholO2 (cm3 min21 mmHg21 kg21) 0.122–0.0487 0.0874 0.0291
STOMACH OF THE AIR-BREATHING ARMORED CATFISH 611
Journal of Morphology

(Moraes et al., 2005) and some reptiles, birds andmammals (Perry, 1989) that depend exclusively onthe pulmonary structure to obtain the oxygenneeds for metabolism, the potential respiratorysurface corresponds to higher values. Comparedwith the exclusively air-breathing animals, ourresults are evidence that P. anisitsi depends on thepotential respiratory surface of the stomach foraerial respiration only under adverse conditions inwhich aquatic respiration does not supply itsneeds. The 1.5 times higher Sv of stomach of smallfish and the decreasing specific surface area withincreasing body mass suggest that small animalswould be capable of obtaining greater specific oxy-gen uptake from atmospheric air than larger speci-mens.
The arithmetic (sarith) and harmonic (sh) mean ofthe thickness of the air-blood diffusion barrier ofPterygoplichthys anisitsi stomach lies in the rangeof most air-breathing organs of fish such as the ar-borescent organ of Clarias batrachus (Munshi,1985), the air sac of Heteropneustes fossilis(Hughes et al., 1974a) and Monopterus cuchia(Munshi et al., 1989), the suprabranchial cham-bers of Channa punctatus (Hakin et al., 1978), C.striatus and C. gachua (Munshi, 1985), the swimbladder of Pangasius hypophthalmus (Podkowaand Goniakowska-Witalinska, 1998), and the stom-ach of Hypostumus plecostomus (Podkowa andGoniakowska-Witalinska, 2003), and is lower thanthe lung of the South American lungfish, Lepidosi-ren paradoxa (Hughes and Weibel, 1976; Moraeset al., 2005) (Table 4). The low values of the diffu-sion barrier of respiratory organs evidence thepreservation of the barrier model in differentgroups of animals that breathe air to allow for anefficient gas exchange process, as emphasized byMaina and West (2005).
The ADF, which is the ratio of the respiratorysurface area to the harmonic mean of the diffusionbarrier (Perry, 1989), represents the anatomicalcomponent of the diffusion capacity of a respira-tory tissue, excluding the blood. The ADF of thestomach of Pterygoplichthys anisitsi is smallerthan that of the lungs of most vertebrates, whichincrease from amphibians to mammals (Perry,1992), but similar to those of the lungfish (Hughesand Weibel, 1976; Moraes et al., 2005). The stom-ach of P. anisitsi has a low specific stomach surfacearea (SSt M�1
B ) but its air-blood barrier is thinnerthan that of the lung of Lepidosiren paradoxa(Moraes et al., 2005) (Table 4).
The Dmorphol of tissue barrier of stomach of Pter-ygoplichthys anisitsi is higher than the air-breath-ing organs of most air-breathing fish but lowerthan the lung of lungfish (Table 4). Dmorphol 5K(S/t), where K is the Krogh specific diffusion con-stant for O2 or CO2, S is the respiratory surfacearea, and t is the harmonic mean of the diffusionbarrier thickness, represents the maximum diffu-sion capacity of a given respiratory organ underideal conditions of ventilation/perfusion in theentire diffusion barrier. On the other hand, Dphysiol
5 VO2/(PaO2 2 PeO2), where VO2 is the oxygenconsumption rate, PaO2 is the arterial partial oxy-gen tension, and PeO2 is the venous partial oxygentension, represents the diffusion capacity underspecific physiological conditions, considering thedifferences in the ventilation/perfusion ratio andthe heterogeneity of diffusion membrane barrier.The Dmorphol is usually much higher than the esti-mated Dphysiol, which approximates the Dmorphol
only during extreme exercise or in combinationwith hypoxia and/or hypercapnia (Scotto et al.,1987; Weibel, 1999). The Dmorphol of the stomach ofP. anisitsi showed a structural and functional simi-
TABLE 4. Specific surface area (SSt M�1B ), harmonic mean of the water-blood diffusion barrier (sh) and morphometric diffusion
capacity for O2 (DmorpholO2) of the air-breathing organs in fish
Animal Air-breathing organSSt M
�1B
(cm2 kg21)sh(lm)
DmorpholO2
(cm3 min21 mmHg21 kg21)
Clarias batrachus (Munshi, 1985) SuprabranchialChambers (SBC)
155 — 0.0420
Gill fans 34 — 0.0005Arborescent organ 287 0.55 0.0773
Heteropneustes fossilis (Hughes et al, 1974a) Air sac 310 1.605 0.0288Monopterus cuchia (Hughes et al., 1974b) Air sac 48 0.44 0.0165Channa punctatus (Hakin et al., 1978) SBC 392 0.780 0.0753Channa striatus (Munshi, 1985) SBC 231 1.359 0.0254Channa gachua (Munshi, 1985) SBC 195 0.56 0.0524Pangasius hypophthalmus (Padkova and
Gonialowska-Witalinska, 1998)Swimbladder — 0.7 —
Hypostumus plecostomus (Padkova andGonialowska-Witalinska, 2003)
Stomach — 0.86 —
P. anisitsi (present study) Stomach 157 0.40–0.74 0.0874Protopterus aethiopcus (Maina and Maloiy, 1985) Lungs 14000 0.37 13.035Lepidosiren paradoxa (Hughes and Weibel, 1976) Lungs 850 0.86 0.3Lepidosiren paradoxa (Moraes et al., 2005) Lungs 664 1.38 0.110
612 A.L. DA CRUZ ET AL.
Journal of Morphology

larity in the characteristics of respiratory tissuesspecialized in using atmospheric air for gasexchange (very thin diffusion barrier), and a pat-tern for ectothermic animals which present low ac-tivity (low respiratory surface area).
In conclusion, the structure and morphometricdata of Pterygoplichthys anisitsi stomach demon-strate the adaptation of this organ to gas exchangefrom air. Moreover, considering that this species isa continuous but non-obligatory air-breathing fish(Cruz, 2007) that uses its stomach as an auxiliaryorgan in response to hypoxia because of extremelypoor dissolved oxygen in water, and that atmos-pheric air contains 35 times more oxygen than air-saturated freshwater, the morphological and mor-phometric properties of the stomach of P. anisitsienable this species to live in severely hypoxic envi-ronments, since it can obtain O2 from air to supplyits metabolic needs.
ACKNOWLEDGMENTS
The authors thank the Aquaculture Center ofthe Sao Paulo State University, Jaboticabal cam-pus, for providing the fish. They also thank theanonymous referees for their comments and sug-gestions and A.L. Cruz and A.C.E. Pedrettiacknowledges CAPES and CNPq, respectively, forawarding scholarships.
LITERATURE CITED
Bartels H. 1971. Diffusion coefficients and Krogh’s diffusionconstants. In: Altman PL, Dittmer DS, editors. Respirationand Circulation. Biological Handbooks. Bethesda: FASEB. pp21–22.
Biswas N, Ojha J, Munshi JSD. 1981. Morphometrics of the re-spiratory organs of an estuarine goby, Boleophthalmus bod-daerti. J Ichthyol 27:316–326.
Burri PH, Haeni B, Tschanz AS, Makanya AN. 2003. Morphom-etry and allometry of the postnatal marsupial lung develop-ment: An ultrastructural study. Resp Physiol Neurobiol 138:309–324.
Carter GS, Beadle LC. 1931. The fauna of the swamps of Para-guayan Chaco in relation to its environment. II. Respiratoryadaptations in the fishes. J Linn Soc London (Zool) 37:327–368.
Crawford RH. 1974. Structure of an air-breathing organ andthe swim bladder in the Alaskan blackfish, Dallia pectorallisBean. Can J Zool 52:1221–1225.
Cruz AL. 2007. O comportamento respiratorio e a cascata de O2
no cascudo de respiracao bimodal Pterygoplichthys anisitsiEigenmann e Kennedy, 1903 (Teleostei, Loricariidae). PhDThesis. Sao Carlos: Universidade Federal de Sao Carlos. 161 p.
Daniels CB, Orgeig S. 2001. The comparative biology of pulmo-nary surfactant: Past, present and future. Comp BiochemPhysiol A 129:9–36.
Daniels CB, Orgeig S, Smits AW. 1995. The evolution of the ver-tebrate pulmonary surfactant. Physiol Zool 68:539–566.
Daniels CB, Orgeig S, Sullivan LC, Ling N, Bennett MB,Schurch S, Val AL, Brauner CJ. 2004. The origin and evolutionof the surfactant system in fish: Insights into the evolution oflungs and swim bladders. Physiol Biochem Zool 77:732–749.
Delariva RL, Agostinho AA. 2001. Relationship between mor-phology and diets of six neotropical loricariids. J Fish Biol58:832–847.
Elbal MT, Agullero B. 1986. A histochemical and ultrastructuralstudy of the gut of Sparus auratus (Teleostei) J SubmicroscCytol 18:335–347.
Gee JH. 1976. Buoyancy and aerial respiration: Factors influ-encing the evolution of reduced swim-bladder volume in someCentral American catfishes (Trichomycteridae, Callichthyidae,Loricariidae, Astroblepidae). Can J Zool 54:1030–1037.
Gee JH, Graham JB. 1978. Respiratory and hydrostatic func-tions of the intestine of the catfishes Hoplosternum thoraca-tum and Brochis splendens (Callichthyidae). J Exp Biol 74:1–16.
Graham JB. 1997. Air-Breathing Fishes: Evolution, Diversity,and Adaptation. San Diego: Academic Press. 299 p.
Gundersen HJG, Bendtsen TF, Korbo L. 1988. Some new simpleand efficient stereological methods and their use in pathologi-cal research and diagnosis. Acta Pathol Microbiol ImmunolScand 96:379–394.
Hakin A, Munshi JSD, Hughes GM. 1978. Morphometrics ofthe respiratory organs of the Indian green snake-headed fish,Channa punctata. J Zool (London) 184:519–543.
Howard CV, Reed MG. 1998. Unbiased Stereology: Three-Dimensional Measurement in Microscopy. Oxford: Bios Scie-tific Publishers. 246 p.
Hughes GM, Weibel ER. 1976. Morphometry of fish lungs. In:Hughes GM, editor. Respiration of Amphibious Vertebrates.London: Academic Press. pp 213–231.
Hughes GM, Singh BR, Guha G, Dube SC, Munshi JSD. 1974a.Respiratory surface areas of an air-breathing siluroid fishHeteropneustes fossilis in relation to body size. J Zool(London) 172:215–232.
Hughes GM, Singh BR, Thakur RN, Munshi JSD. 1974b. Areasof the air-breathing surfaces of Amphipnous cuchia (Ham).Proc Indian Nat Sci Acad 40:379–392.
Kemp NE. 1987. The biology of the Australian lungfish, Neocer-atodus forsteri. In: Bemis WE, Burggren WW, Kemp NE, edi-tors. The Biology and Evolution of Lungfishes. New York:Alan R. Liss. pp 181–198.
Kramer DL. 1978. Ventilation of the respiratory gas bladder inHoplerythrinus unitaeniatus (Pisces, Characoidei, Erythrini-dae). Can J Zool 56:931–938.
Liem KF. 1989. Respiratory gas bladder in teleosts: Functionalconservatism and morphological diversity. Am Zool 29:333–352.
Maina JN, Maloiy GMO. 1985. The morphometry of the lung ofthe African lungfish (Protopterus aethiopicus): Its structural-functional correlations. Proc R Soc Lond Biol Sci 224:399–420.
Maina JN, West JB. 2005. Thin and strong! The bioengineeringdilemma in the structural and functional design of the blood-gas barrier. Physiol Rev 85:811–844.
Mattias AT, Moron SE, Fernandes MN. 1996. Aquatic respira-tion during hypoxia of the facultative air-breathing Hoplery-thrinus unitaeniatus. A comparison with the water-breathingHoplias malabaricus. In: Val AL, Almeida-Val VM, RandallDJ, editors. Physiology and Biochemistry of the Fishes of theAmazon. Manaus: INPA. pp 203–209.
McMahon BR, Burggren WW. 1987. Respiratory physiology ofintestinal air breathing in the teleost fish Misgurnus anguilli-caudatus. J Exp Biol 133:371–393.
Moraes MFPG, Holler S, Costa OTF, Glass ML, Fernandes MN,Perry SF. 2005. Morphometric comparison of the respiratoryorgans of the South American lungfish Lepidosiren paradoxa(Dipnoi). Physiol Biochem Zool 78:546–559.
Morrison CM, Wright JR Jr. 1999. A study of the histology ofthe digestive tract of the Nile tilapia. J Fish Biol 54:597–606.
Munshi JSD. 1985. The structure, function and evolution of theaccessory respiratory organs of air-breathing fishes of India.Fortsch Zool 30:353–366.
Munshi JSD, Hughes GM, Gehr P, Weibel ER. 1989. Structureof the air-breathing organs of a swamp mud eel Monopteruscuchia. Japan J Ichthyol 35:453–465.
Oliveira C, Taboga SR, Smarra ALS, Bonilla-Rodriguez GO.2001. Microscopical aspects of acessory air breathing through
STOMACH OF THE AIR-BREATHING ARMORED CATFISH 613
Journal of Morphology

a modified stomach in the armoured catfish Liposarcus ani-sitsi (Siluriformes. Loricariidae). Cytobios 105:153–162.
Orgeig S, Daniels CB. 2001. The roles of cholesterol in pulmo-nary surfactant: Insights from comparative and evolutionarystudies. Comp Biochem Physiol A 129:75–89.
Perry SF. 1989. Structure and function of the reptilian respira-tory system. In: Wood SC, editor. Comparative PulmonaryPhysiology, Current Concepts. New York: Marcel Dekker.pp 193–236.
Perry SF. 1992. Morphometry of vertebrate gills and lungs: Acritical review. In: Egginton S, Ross HF, editors. Modelling ofOxygen Transport from Environment to Cell. SEB SeminarSeries 51. Cambridge: Cambridge University Press. pp 57–77.
Podkowa D, Goniakowska-Witalinska L. 1998. The structure ofthe airbladder of the catfish Pangasius hypophthalmus Rob-erts and Vidthayanon 1991, (previously P. sutchi Fowler1937). Folia Biol 46:189–196.
Podkowa D, Goniakowska-Witalinska L. 2002. Adaptations tothe air breathing in the posterior intestine of the catfish (Cor-ydoras aeneus, Callichthyidae): A histological and ultrastruc-tural study. Folia Biol 50:69–82.
Podkowa D, Goniakowska-Witalinska L. 2003. Morphology ofthe air-breathing stomach of the catfish Hypostomus plecosto-mus. J Morphol 257:147–163.
Rubio S, Chailley-Heu B, Ducroc R, Bourbon JR. 1996. Anti-body against pulmonary surfactant protein A recognizes pro-
teins in intestine and swim bladder of the freshwater fish,carp. Biochem Biophys Res Commun 225:901–906.
Satora L. 1998. Histological and ultrastructural study of thestomach of the air-breathing Ancistrus multispinnis (Siluri-formes, Teleostei). Can J Zool 76:83–86.
Satora L, Winnicki A. 2000. Stomach as an additional respira-tory organ, as exemplified by Ancistrus multispinnis (Couvieret Valenciennes, 1937). Siluriformes, Teleostei. Acta IchtPiscat 30:73–79.
Sawaya P. 1946. Sobre a biologia de alguns peixes de respiracaoaerea (Lepidosiren paradoxa Fitzinger e Arapaima gigas Cuv-ier). Bol Fac Fil Cienc Let Univ Sao Paulo 11:255–286.
Scotto P, Ichinose Y, Patane L, Meyer M, Piiper J. 1987. Alveo-lar-capillary diffusion of oxygen in dogs exercising in hypoxia.Resp Physiol 68:1–10.
Silva JM, Hernandez-Blazquez FJ, Julio HF Jr. 1997. A newaccesory respiratory organ in fishes: Morphology of the respi-ratory purses of Loricariichthys platymetopon (Pisces, Lori-cariidae). Ann Sci Natur Zool 18:93–103.
Weibel ER. 1999. Understanding the limitation of O2 supplythrough comparative physiology. Resp Physiol 118:85–93.
Weibel ER, Knight BW. 1964. A morphometric study on thethickness of the pulmonary air-blood barrier. J Cell Biol21:367–384.
614 A.L. DA CRUZ ET AL.
Journal of Morphology