root and leaf traits, water use and drought tolerance of maize genotypes
TRANSCRIPT

Biologia 68/6: 1123—1127, 2013Section BotanyDOI: 10.2478/s11756-013-0258-3
Root and leaf traits, water use and drought toleranceof maize genotypes**
Krisztina R. Végh
Institute for Soil Science and Agricultural Chemistry, Center of Agricultural Research Hungarian Academy of Sciences,Herman O. 15, H-1022 Budapest, Hungary; e-mail: [email protected]
Abstract: The main components of drought tolerance of six maize genotypes were studied to evaluate crop performancein water limiting environments: (1) the postponement of dehydration by reduced transpiration rate (TR) and an increasedefficiency of water acquisition from soil; (2) the tolerance of dehydration by effective physiological water use. The aim was todescribe the genotype dependent response to drought in leaf and root traits and water relations using data from controlledenvironment and field experiments, and using dynamic simulation by the Swedish Coup model. High genetic variation wasdetected in the root density, acquisition efficiency and water use among the genotypes. The female parent lines had thegreatest TR with the smallest dry matter accumulation in water deficiency, whereas hybrids could acquire more water fromdryer soil while maintaining a lower TR. Hybrid Mv 444 increased water potential more strongly in leaves than hybridNorma. The postponement of dehydration was observed for Norma, while more tolerance to dehydration characterized Mv444. Simulation was an effective tool for testing hypotheses considering water acquisition efficiency and for summarizingthe results of the measurements in a formalized structure; it helped to quantify the dynamics of water availability and theimpact of drought on the growth of the maize genotypes.
Key words: maize genotypes; leaf and root traits; water deficit; water potential; CoupModel
Introduction
Drought stress in crops evolves from an insufficient sup-ply of water in the soil and increased evaporative de-mand of the aerial environment. Increased temperatureand/or decreased humidity can have a large impacton evaporative demand, prompting concerns about theimpact of climate change on crop productivity. There-fore the management of water resources will become ofgreater importance to plant production in the futureincluding the selection of crops with greater droughttolerance. Understanding genetics by environment in-teractions is a substantial challenge in identifying traitsthat define the underlying mechanisms of the droughttolerance of crops.Some crops cope well under conditions where soil
generally imparts water stress (Yanagawa & Fujimaki2013). Huzsvai & Rajkai (2009) demonstrated theadaptability of maize plants to water deficit. It is ev-ident, that how the dimensions of water deficiency in-teract with genotype properties determines if the effectsof drought are irreversible to growth and therefore cropyield.Water flow in the soil-plant-atmosphere continuum
is extremely complex, varying both spatially and tem-porally, influenced by the historic background of thesoil (Lichner et al. 2012; Márton 2008). Crop simula-
tion models that capture the physiological processes ofgrowth and predict yield are suitable for study of geno-type by environment interactions and are more accurateand more widely applicable provided that appropriateparameter values are defined.The aim of this study was to describe the genotype
dependent response to drought in leaf and root traitsand water relations using data from controlled environ-ment and field experiments, and using dynamic simu-lation by the CoupModel (Jansson & Karlberg 2003).There are several strands to the research: (1) Postpone-ment of dehydration by reduced loss of transpired waterand increased efficiency of water acquisition from soil.(2) Dehydration tolerance: more effective physiologicalwater use by lowering water potential in the tissues andby osmotic adjustment.The dynamic simulation was applied in order to
designate the major ecophysiological and morpholog-ical components and their combinations and analyzetheir role in the genotype dependent responses to wa-ter deficiency in relation to changes in the soil and aerialenvironment.
Methods
Six maize genotypes with known water use efficiency (Végh& Szundy 2004) were selected: two single cross hybrids,
*Corresponding author, **Special Section on Biohydrology, guest-editors Ľubomír Lichner & Kálmán Rajkai
c©2013 Institute of Botany, Slovak Academy of Sciences

1124 K.R. Végh
Table 1. Water potential in the leaves (at transpiration rates 0.5–1.0 g g−1 d−1 (LT) and 1.0–1.5 g g−1 d−1 (NT) in the DAS 42-50period), relative leaf water deficit (RWD) and available water storage (AWS) in drought. Standard errors are given in parenthesis.
Leaf water potential (MPa) RWD AWSGenotype
LT NT (g m−2) (g)
DAS 42 DAS 43 DAS 45 DAS 46 DAS 49 DAS 50 DAS 46 DAS 51
444-B –0.91 –0.91 –1.03 –0.88 –0.93 –1.01 –1.25 31.3 (0.99) 12.29N-B –0.68 –0.76 –0.78 –0.90 –0.98 –0.96 –1.06 15.7 (3.93) 8.27N-A –1.16 –1.34 –1.18 –1.19 –0.97 –1.10 –1.62 41.6 (2.64) 15.08444-A –0.79 –0.84 –1.00 –0.93 –1.12 –0.95 –2.25 59.1 (2.59) 16.15Norma –0.67 –0.86 –0.90 –0.78 –0.78 –0.82 –1.22 10.9 (7.06) 0.05Mv 444 –1.09 –1.09 –1.03 –1.09 –1.01 –0.91 –2.20 13.5 (0.15) 5.55
Norma and Mv 444 and their parent lines, N-A, N-B, 444-Aand 444-B.
In controlled environment transpiration rate (TR), leafwater potential and relative water deficit (RWD) in leaveswere evaluated in the genotypes subjected to an increasingwater deficit. Pot size (16 cm high, 6.5 cm diameter) androoting medium (500 cm3 sand) was chosen for the rapiddevelopment of water stress and to allow the accurate mea-surability of the available water storage (AWS). The pots,arranged in a randomized block design, with three repli-cations, were weighed daily to determine soil AWS, plantwater uptake and TR, then irrigated with a complete nutri-ent solution. The drying treatment was imposed from the16th (V2 stage) to the 52nd day after sowing (DAS). Waterpotential and osmotic potential values were determined us-ing Wescor psychrometer (Wescor Inc. Logan, Utah, USA)on leaf discs and, at harvest, on secondary root samples.The RWD (g cm−2) was calculated as the mass differenceto saturated leaf per unit leaf area.
A three-year field experiment was conducted for fieldmeasurements and to create the dataset for the simulation ofwater dynamics in soil-plant-atmosphere system. The samesix genotypes were grown in a nutrient rich loam soil in arandomised block design with four replications in Marton-vásár, Hungary. Particle-size distribution, soil bulk density,saturated conductivity and water retention curve used toevaluate soil hydraulic properties were measured. Meteoro-logical variables (global radiation, air temperature, relativeair humidity, precipitation) were measured on a daily basisin the experiment. On 5–7 occasions in the growing seasonsplant samples were collected and the vertical distribution ofsoil water content was detected by a BR 150 capacitancemoisture probe (RISSAC, Budapest, Hungary) in 10 cm in-tervals down to 160 cm (Végh & Szundy 2009). The verticaldistribution of root length density was observed in 5 cmdiameter 10 cm long soil cores taken from in-row and inter-row sites down to 100 cm depth at anthesis. Root length incores was determined by image analysis (Delta-T Ltd. Cam-bridge, UK). Specific root density (SRD) was calculated bydividing root dry mass by root length (mg m−1).
Data were analyzed by ANOVA to evaluate differ-ences between treatments. Soil water effects for genotypesin the different water treatments were analyzed by two-way ANOVA for all traits separately, with genotypes anddrought treatments as the main factors. Linear correlationwas used to represent the relationship between predictedand measured parameters.
The CoupModel was used for simulating the waterflows in the maize stands with the experimental data forthe given genotypes. Model outputs were fitted on the depth
distribution of volumetric moisture content of the soil mea-sured by the BR150 probes. The model parameter set finelytuned on the soil moisture and crop data was then usedto simulate water dynamics for the genotypes. Simulationswere considered acceptable when differences to measuredvalues were less than 5%. The data and results of the fieldexperiment applied in the simulation were: (1) meteorolog-ical data set, (2) physical properties of the layers defined inthe soil profile, (3) maximum LAI values, (4) root growthand depth distribution in drying soil. The results of thepot experiment applied were: (1) vapor pressure deficit onthe canopy level and set of stomatal conductance for hybridNorma, (2) decrease of the value of critical pressure headcontrolling potential water uptake for Norma, (3) increaseof the parameter value that controls the slope of potentialwater uptake as the function of the pressure head for hybridMv 444.
Results and discussion
During the three years of our field experiment (1998-1999-2000), in-season precipitation, as related to the50-year average (277 mm = 100%), was 145.9%, 144.9%and 51.3% for year1, year2 and year3 respectively. Inyear3 the growing season was dry except some lightrain showers at anthesis.In year1 and year2 the maximum leaf area (LA)
per plant varied between 0.35 m2 and 0.55 m2 for theparent lines and 0.70 m2 to 0.75 m2 for the hybrids. Inthe dry year3, the maximum LA of all six genotypesdid not exceed 0.5 m2 per plant. In the pot experi-ment in controlled environment the transpiration rate(TR) ranged between 0.25 and 2.2 g water g−1 of plantfresh mass d−1. The dynamics of both the AWS in rootmedium and the TR differed significantly for the parentlines compared to the hybrids (p < 0.05). Maternal lines(N-A and 444-A) had the highest TR and accumulatedthe least biomass (p < 0.05), while hybrids transpiredat lower rates with a larger LA and less relative waterdeficit (RWD) (p < 0.01) among the genotypes. Thegreater RWD in the parent lines was caused by the lessefficient root acquisition of water. Hybrid Norma hadthe greatest biomass accumulation and maintained asimilar TR and RWD of Mv 444, but with lower AWSas shown in Table 1. This reflects more efficient wateruse.
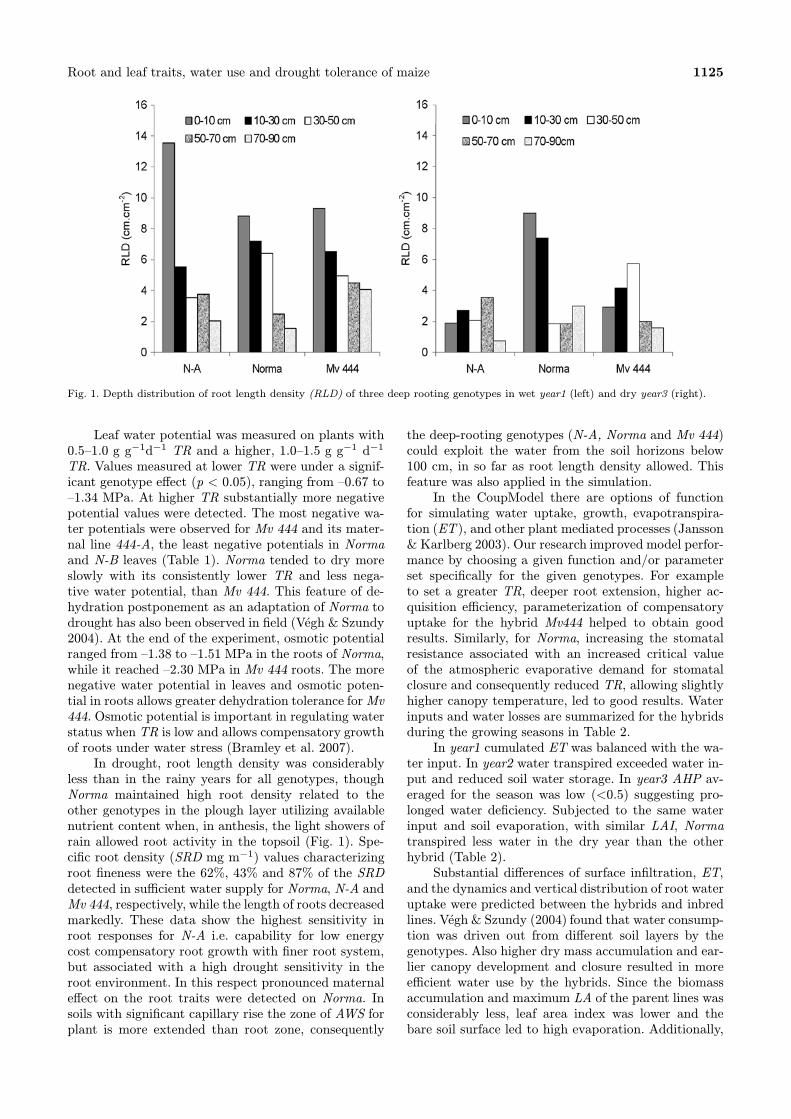
Root and leaf traits, water use and drought tolerance of maize 1125
Fig. 1. Depth distribution of root length density (RLD) of three deep rooting genotypes in wet year1 (left) and dry year3 (right).
Leaf water potential was measured on plants with0.5–1.0 g g−1d−1 TR and a higher, 1.0–1.5 g g−1 d−1
TR. Values measured at lower TR were under a signif-icant genotype effect (p < 0.05), ranging from –0.67 to–1.34 MPa. At higher TR substantially more negativepotential values were detected. The most negative wa-ter potentials were observed for Mv 444 and its mater-nal line 444-A, the least negative potentials in Normaand N-B leaves (Table 1). Norma tended to dry moreslowly with its consistently lower TR and less nega-tive water potential, than Mv 444. This feature of de-hydration postponement as an adaptation of Norma todrought has also been observed in field (Végh & Szundy2004). At the end of the experiment, osmotic potentialranged from –1.38 to –1.51 MPa in the roots of Norma,while it reached –2.30 MPa in Mv 444 roots. The morenegative water potential in leaves and osmotic poten-tial in roots allows greater dehydration tolerance forMv444. Osmotic potential is important in regulating waterstatus when TR is low and allows compensatory growthof roots under water stress (Bramley et al. 2007).In drought, root length density was considerably
less than in the rainy years for all genotypes, thoughNorma maintained high root density related to theother genotypes in the plough layer utilizing availablenutrient content when, in anthesis, the light showers ofrain allowed root activity in the topsoil (Fig. 1). Spe-cific root density (SRD mg m−1) values characterizingroot fineness were the 62%, 43% and 87% of the SRDdetected in sufficient water supply for Norma, N-A andMv 444, respectively, while the length of roots decreasedmarkedly. These data show the highest sensitivity inroot responses for N-A i.e. capability for low energycost compensatory root growth with finer root system,but associated with a high drought sensitivity in theroot environment. In this respect pronounced maternaleffect on the root traits were detected on Norma. Insoils with significant capillary rise the zone of AWS forplant is more extended than root zone, consequently
the deep-rooting genotypes (N-A, Norma and Mv 444)could exploit the water from the soil horizons below100 cm, in so far as root length density allowed. Thisfeature was also applied in the simulation.In the CoupModel there are options of function
for simulating water uptake, growth, evapotranspira-tion (ET ), and other plant mediated processes (Jansson& Karlberg 2003). Our research improved model perfor-mance by choosing a given function and/or parameterset specifically for the given genotypes. For exampleto set a greater TR, deeper root extension, higher ac-quisition efficiency, parameterization of compensatoryuptake for the hybrid Mv444 helped to obtain goodresults. Similarly, for Norma, increasing the stomatalresistance associated with an increased critical valueof the atmospheric evaporative demand for stomatalclosure and consequently reduced TR, allowing slightlyhigher canopy temperature, led to good results. Waterinputs and water losses are summarized for the hybridsduring the growing seasons in Table 2.In year1 cumulated ET was balanced with the wa-
ter input. In year2 water transpired exceeded water in-put and reduced soil water storage. In year3 AHP av-eraged for the season was low (<0.5) suggesting pro-longed water deficiency. Subjected to the same waterinput and soil evaporation, with similar LAI, Normatranspired less water in the dry year than the otherhybrid (Table 2).Substantial differences of surface infiltration, ET,
and the dynamics and vertical distribution of root wateruptake were predicted between the hybrids and inbredlines. Végh & Szundy (2004) found that water consump-tion was driven out from different soil layers by thegenotypes. Also higher dry mass accumulation and ear-lier canopy development and closure resulted in moreefficient water use by the hybrids. Since the biomassaccumulation and maximum LA of the parent lines wasconsiderably less, leaf area index was lower and thebare soil surface led to high evaporation. Additionally,

1126 K.R. Végh
Table 2. Predicted total seasonal water input and output, the ratio of actual to potential ET (agro-hydropotential index AHP) andmaximum leaf area index (LAI) for the hybrids.
Hybrids Precipitation Infiltration Evapotranspiration AHP Evaporation LAI(mm) (mm) (mm) (–) (mm) (–)
Norma Year1 482 482 484 0.81 138 3.3Year2 443 430 553 0.78 124 3.6Year3 173 157 236 0.41 97 2.7
Mv 444 Year1 482 482 501 0.88 143 3.3Year2 443 429 576 0.81 122 3.6Year3 173 159 313 0.49 98 2.5
microclimate must have been much dryer than insidethe closed stand of the hybrids, which resulted in muchlower soil water conductivity thus reduced infiltration(Homolák et al. 2009). Stomatal resistance may increaseas a consequence of the dryer microclimate (Anda 2008)reducing TR. Accordingly the productive component ofwater loss, transpiration, related to soil evaporation de-creased considerably for the parent lines.In year3 the crop was dependent mainly on stored
water in the soil profile. Ong et al. (1996) suggestedthat the main limitation on the rate of water supply toa plant growing on stored water in the soil profile is therate at which roots proliferate in soil. Nevertheless, apossible consequence may be that if a rapidly growingroot system exploits deeper horizons to supply the fullET demand early in the season, the crop may remainwithout water later (Passioura 1988). Slower depletionof soil water prior to anthesis may benefit the crop whilegrowing on stored water. Subjected to the same waterinput and soil evaporation Norma transpired less waterin the dry year than the other hybrid (Table 2). Thisgrowth strategy performed by Norma was a smaller up-take rate/hydraulic conductivity of roots and leaveswith lower TR. Although slightly lower grain yieldswere obtained in good years than for Mv 444 (Végh &Szundy 2004) this strategy may allow crop to surviveand grow in longer drought periods.There has been a vast amount of research on phys-
iological water use efficiency in connection with tran-spiration efficiency, leaf water relations, efficient har-vest index etc. but limited research deals with wateracquisition efficiency i.e. root physiology and regula-tion despite the substantial development of the discov-ery of root aquaporins (Bramley et al. 2007) and alsohydraulic lift (Liste & White 2008). The exciting ques-tions connected to the efficiency of water acquisition ofplants raise the issue of its physiological basis in roots,as well as in the root-shoot connections, which is farfrom clear.
Conclusions
Interactions of soil water supply and plant activity ledto significantly different rooting pattern along the soilprofile for the six genotypes. The genotypes differed incompensatory root growth and deep root extension thatwere observed in limited water conditions. One of the
hybrids (Mv 444) depleted soil water steadily and muchstrongly while another hybrid (Norma) consumed lesswater from the soil. The parent lines with slower growthrate and higher soil evaporation loss from the bare soilsurface utilized soil water with lower efficiency relatedto the hybrids.The evaluation of the principal mechanisms of tol-
erance of the two hybrids to drought stress suggests astrategy of postponement the dehydration by Normawhilst the toleration of a certain gradient of dehydra-tion by Mv 444.Simulation modeling was an effective tool in con-
trolling hypothesis and summarizing the results of themeasurements in a formalized structure; it helped toquantify the dynamics of water availability and the im-pact of drought on the growth of the maize genotypes.For a more comprehensive characterizing of the ac-
quisition strategy of the hybrids with high water use ef-ficiency and good performance additional studies haveto be done primarily on root physiological functions andfine morphology in the future. The regulation of wateruptake has not been clear so far; and, both the possibil-ity of a switched on high active uptake and an off chancefunctioning hydraulic lift have to be investigated.
Acknowledgements
Special thanks to Dr. Tamás Szundy (CAR, Martonvásár,Hungary) for providing the genotypes and the conditionsof field experiment and Prof. Paul D. Hallett (University ofAberdeen, Aberdeen, UK) for his valuable comments on themanuscript.
References
Anda A. 2008. Air temperature and humidity within a maizestand with sub-dosage irrigation. Novénytermelés 37: 69–84.(in Hungarian)
Bramley H., Turner D.W., Tyerman S.D. & Turner N.C. 2007.Water flow in the roots of crop species. The influence of rootstructure, aquaporin activity and waterlogging. Adv. Agron.96: 133–196.
Homolák M., Capuliak J., Pichler V. & Lichner L. 2009. Esti-mating hydraulic conductivity of a sandy soil under differentplant covers using minidisk infiltrometer and a dye tracer ex-periment. Biologia 64: 600–605.
Huzsvai L. & Rajkai K. 2009. Modeling of plant adaptation toclimatic drought induced water deficit. Biologia 64: 536–541.

Root and leaf traits, water use and drought tolerance of maize 1127
Jansson P.-E. & Karlberg L. 2003. Coupled heat and mass trans-fer model for soil-plant- atmosphere systems. http://www2.lwr.kth.se/CoupModel/index.html
Lichner L., Holko L., Zhukova N., Schacht K., Rajkai K., Fodor N.& Sandor R. 2012. Plants and biological soil crust influencethe hydrophysical parameters and water flow in an aeoliansandy soil. J. Hydrol. Hydromech. 60: 309–318.
Liste H.-H. & White J.C. 2008. Plant hydraulic lift of soil water –implications for crop production and land restoration. PlantSoil 313: 1–17.
Márton L. 2008. Long-term study of precipitation and fertiliza-tion interactions on winter wheat in Nyirlugos field trial be-tween 1973 and 1990. Cereal Res. Commun. 36: 511–522.
Ong C.K., Black C.R., Marshall F.M. & Corlett J.E. 1996. Prin-ciples of resource capture and utilization of light and water,pp. 74–158. In: Ong C.K & Huxley P. (eds), Tree – CropInteractions, Wallingford: CAB Intern.
Passioura J.B. 1988. Water transport in and to roots. Ann. Rev.Plant Physiol. Plant Mol. Biol. 39: 245–265.
Végh K.R. & Szundy T. 2004. Water supply and water use effi-ciency of maize genotypes. Agrokémia és Talajtan 53: 35–55.(in Hungarian)
Yanagawa A. & Fujimaki H. 2013. Tolerances of Canola todrought and salinity stresses in terms of parameter values ofroot water uptake model. J. Hydrol. Hydromech. 61: 73–80.
Received January 23, 2013Accepted June 3, 2013