Özet - hoşgeldinizacikarsiv.ankara.edu.tr/browse/1599/2231.pdf · taq polimeraz enzim...
TRANSCRIPT

ÖZET
Doktora Tezi
FARKLI TRİTİCALE HATLARININ MORFOLOJİK VE DNA MARKÖRLERİYLE GENETİK
KARAKTERİZASYONU
Mehmet ATAK
Ankara Üniversitesi Fen Bilimleri Enstitüsü
Tarla Bitkileri Anabilim Dalı
Danışman: Prof. Dr. Cemalettin Yaşar ÇİFTÇİ
Bu çalışma; 2000-2001 yıllarında A.Ü. Ziraat Fakültesi, Araştırma Uygulama Çiftliği deneme tarlalarında 2 yıl süreyle ve Tarla Bitkileri Bölümü, biyoteknoloji labaratuvarında yürütülmüştür. Denemelerde, 25 adet tritikale çeşit/hattı verim ve verim öğeleri yönünden incelenmiş ve ayrıca RAPD-PCR tekniği ile genetik karakterizasyonu belirlenmeye çalışılmıştır. İki yılın ortalaması olarak; çiçeklenme tarihi 27.00-35.87 gün arasında değişmiş olup, en erkenci hat ZF 12 ve en geççi hat ise LAD 388’dir. En kısa bitki boyuna sahip tritikale hattı MT 1 (109.6 cm), en uzun tritikale hattı ise ZF 3 (144.1 cm) olmuştur. En fazla kardeşlenme gösteren tritikale hattı 5.27 adet /bitki ile ZF 15, en az kardeşlenme gösteren tritikale hattı ise 4. 31 adet/bitki ile JGS olmuştur. En uzun başak uzunluğuna sahip tritikale hattı CWT (107.9 mm), en kısa başak uzunluğuna sahip tritikale hattı ise ZF 7 (85.24 mm) olmuştur. Tritikale hatlarının ortalama başakçık sayısı 19.42 adet/başak (ZF 12) ile 27.05 adet/başak (ZF 3) arasında değişmiştir. En yüksek başakta tane sayısına sahip tritikale hattı LAD 388, en düşük başakta tane sayısına sahip tritikale hattı ise ZF 12 olmuştur. Başakta tane ağırlığı yönünden en yüksek değer ZF 8 tritikale hattında, en düşük başakta tane ağırlığı ise ZF 12 tritikale hattından elde edilmiştir. Bin tane ağırlığı yönünden 43.62 g ile ZF 16 tritikale hattı en yüksek değeri, 32.45 g ile LAD 388 tritikale hattı ise en düşük değeri vermiştir. En yüksek tane verimine sahip tritikale hattı ZF 16 (5920.9 kg/ha) olup, bunu sırasıyla ZF 15 tritikale hattı ve Tatlıcak-97 tritikale çeşidi izlemiştir. En düşük tane verimine sahip tritikale hattı ise ZF 12 (4750.3 kg/ha) olmuştur. Hasat indeksi yönünden tritikale hatları % 29.30 ile 36.37 arasında değişen değerler göstermişlerdir. Protein oranı yönünden, tritikale hatlarının ortalama olarak % 11.76 (ZF 6) ile 14.37 (ZF 3) arasında değişen değerler gösterdiği saptanmıştır. RAPD-PCR sonuçlarına göre; kullanılan primerlerden 15 tanesi amplifikasyon göstermiş ve % 61.6 oranında polimorfizm elde edilmiştir. Primer başına ortalama 4 adet RAPD ürünü oluşmuş, bunlardan 2.46’sı polimorfik bulunmuştur. Tritikale çeşit ve hatları başlıca 3 grupta toplanmıştır. Birinci grupta yer alan ZF 2 hattı diğer tüm çeşit ve hatlarla 0.16 oranında farklılık göstermiş, LO 51 hattı ile diğer tüm hatlarlarla 0.15 oranında farklı olarak saptanmıştır. Diğer hatlar 3. grupta yer almıştır. ZF 8–GTAD, ZF 6- ZF 7, ZF 16–JGS hatları birbirine genetik olarak en yakın hatlar olarak bulunmuştur. Tatlıcak-97 tritikale çeşidine en yakın hattın ise ZF 1 olduğu saptanmıştır. Tritikale hatları arasındaki genetik uzaklık 0.034 ile 0.457 arasında değişme göstermiştir. RAPD-PCR yönteminin tritikale genotiplerinin genetik karakterizasyonunda kullanılabileceği belirlenmiştir. 2004, 106 sayfa ANAHTAR KELİMELER: Tritikale, Polimeraz zincir reaksiyonu (PCR), Rasgele çoğaltılmış DNA farklılığı (RAPD)
i

ABSTARACT
Ph.D. Thesis
THE GENETIC DNA MARKER AND MORPHOLOGICAL CHARACTERIZATION OF
DIFFERENT TRITICALE LINES
Mehmet ATAK
Ankara Üniversity Graduate School of Natural Science and Applied Sciences
Department of Agronomy
Supervisor:Prof. Dr. Cemalettin Yaşar ÇİFTÇİ
The study was carried out at the Research Farm and Biotechnology Laboratory of the Department of Field Crops, Faculty of Agriculture, University of Ankara during 2000-2001 for a period of two years. Twenty five lines and one cultivar of triticale were evaluated in the field experiment for yield and some yield components. Moreover, RAPD-PCR technique was used for the genetic characterization of the genotypes. Mean values of two years showed that anthesis date among the genotypes had range of 27.00-35.87 days with earliest anthesis date in ZF 12 and the latest anthesis date in LAD 388. The shortest genotype was MT 1 (109.6 cm) and the longest genotype was ZF 3 (144.1 cm). The highest number of 5.27 tillers/plant were recorded on ZF 15 and the least number 4.31 tiller/plant were recorded on JGS. The longest spike (107.9 mm) was found on CWT and the smallest spike (85.24 mm) was found ZF 7. Mean number of 19.42 (ZF 12) spikilets/spike to 27.05 (ZF 3) spikilets/spike were observed among the genotypes. The highest and the least number of seeds/spike were observed on LAD 388 and ZF 12, respectively. On the basis of the seed weight, the heaviest seeds were found on ZF 8 and the lightest seeds were recorded on ZF 12. Whereas, on the basis of 1000 kernel weight, the largest value of 43.62 g was observed from line ZF 16 and the least value of 32.45 g was observed from line LAD 388. The highest yield of 5920.9 kg/ha was observed from line ZF 16 followed closely by line ZF 15 and cultivar Tatlıcak-97. The lowest yield was obtained from line ZF 12 (4750.3 kg/ha). A harvest index of 29.30-36.37 % was noted among lines. Mean protein of 11.76 % (ZF 6) to 14.37 % (ZF 3) was recorded among the evaluated lines and cultivar. On the basis of RAPD-PCR analysis, 15 primers showed amplification producing 61.6 % polymorphism, an average of 4 RAPD band were detected and 2.46 of them were polymorphic, constituting 3 main groups of triticale lines. ZF 2 in the first group showed variation of approximately 0.16 from other lines and cultivar. Similarly LO 51 of in the second group showed variation of 0.15 from other genotypes. Lines ZF 8- GTAD, ZF 6- ZF 7, ZF 16-JGS were very similar and showed themselves in the third group. The closest relative of Tatlıcak-97 was found to be ZF 1. A genetic variation of 0.034-0.457 was recorded among evaluated lines and cultivar. It was found satisfactory that RAPD-PCR could be effectively used to judge genetic characterization among the triticale genotypes. 2004, 106 pages Key Words: Triticale, Polymerase chain reaction (PCR), Random amplified polymorphic DNA (RAPD)
ii

TEŞEKKÜR
Çalışmam süresince tüm olanak ve bilgilerini sağlayan, tezin gelişimini titizlikle
inceleyen ve yöneten sayın hocam Prof. Dr. Cemalettin Y. ÇİFTÇİ’ye, denemede
kullanılan bitki materyalini sağlayan ve yardımlarını esirgemeyen sayın Prof. Dr. Saime
ÜNVER’e, araştırmanın yürütülmesinde laboratuar ve bilimsel alt yapı imkanlarını
sağlayan, araştırmanın her aşamasında desteklerini esirgemeyen sayın Prof. Dr.
Sebahattin ÖZCAN’a, tez çalışmamın tüm aşamalarında değerli katkılarından
yararlandığım Sayın Prof. Dr. Mehmet KILINÇ’ a, çalışmalarımda bana bilimsel destek
ve yardımda bulunan Yard. Doç.Dr. Muharrem KAYA, laboratuar çalışmalarının her
aşamasında yardımcı olan Araş.Gör. İskender PARMAKSIZ, Dr. Serkan URANBEY,
Dr. Halid Mahmood KHAWAR, Araş. Gör. M. Demir KAYA, Araş. Gör. Arif İPEK,
Dr. Ali ERGÜL, Dr. Selma GÜLBİTTİ, Sn. Muhammed KAYA ve Araş. Gör.
Ercüment Osman SARIHAN’a, araştırmanın tarla denemelerini yürüttüğüm sırada
imkanlarından yararlandığım A.Ü. Ziraat Fakültesi Araştırma Uygulama Çiftliği
Müdürü sayın Prof. Dr. Hatice DUMANOĞLU olmak üzere Ziraat Mühendisleri Şaban
KOÇ, Taşkın EROL, Kamil KARA ve öğrenci arkadaşlara, Tarım ve Köyişleri
Bakanlığı Tarımsal Araştırmalar Genel Müdürlüğü, Tarla Bitkileri Merkez Araştırma
Enstitüsü çalışanları; Zir. Yüksek Mühendisi Selami YAZAR ve ekibine teşekkürlerimi
sunarım.
Ayrıca çalışmalarım sırasında bana maddi manevi desteğini sunan, sabrından dolayı
hayran olduğum biricik eşim Esra ATAK başta olmak üzere tüm aileme teşekkürü bir
borç bilirim.
Mehmet ATAK
Ankara, Eylül 2004
iii

İÇİNDEKİLER
i ii iii vi vii
ÖZET.................................................................................................................. ABSTRACT....................................................................................................... TEŞEKKÜR ...................................................................................................... SİMGELER DİZİNİ........................................................................................... ŞEKİLLER DİZİNİ............................................................................................ ÇİZELGELER DİZİNİ....................................................................................... vii 1. GİRİŞ......................................................................................................... 1 2. KAYNAK ÖZETLERİ............................................................................. 8 2.1. Morfolojik Özelliklerin Değerlendirilmesi.............................................. 8 2.2. DNA İzolasyonu ve RAPD PCR Tekniği............................................... 16 3. MATERYAL ve YÖNTEM..................................................................... 24 3.1 Materyal.................................................................................................... 24 3.2. Araştırma Yeri ve Özellikleri.................................................................. 24 3.2.1. İklim özellikleri................................................................................... 25 3.2.2. Toprak özellikleri ................................................................................ 27 3.3. Yöntem .................................................................................................. 27 3.3.1. Tarla denemesi .................................................................................... 27 3.3.2. Laboratuvar çalışmaları....................................................................... 27 3.3.2.1. DNA izolasyonu ............................................................................... 28 3.3.2.2. DNA miktarı ve temizliği ................................................................. 29 3.3.2.3. DNA konsantrasyonunun optimizasyonu ......................................... 30 3.3.2.4. MgCl2 konsantrasyonunun optimizasyonu ....................................... 30 3.3.2.5. Primer konsantrasyonunun optimizasyonu ....................................... 30 3.3.2.6. dNTP konsantrasyonunun optimizasyonu ....................................... 32 3.3.2.7. Taq Polimeraz enzim konsantrasyonunun optimizasyonu ………... 33 3.3.2.8. Diğer reaksiyon bileşenleri .............................................................. 33 3.4. PCR Koşulları ve RAPD Reaksiyonları ................................................. 33 3.5. Agaroz Jel Elektroforez .......................................................................... 34 3.6. Verilerin Elde Edilmesi ......................................................................... 35 3.7. Verilerin Değerlendirilmesi ................................................................... 36 4. ARAŞTIRMA BULGULARI VE TARTIŞMA ……………………… 37 4.1. Tarla Deneme Bulguları ve Tartışma ..................................................... 37 4.1.1. Çiçeklenme tarihi (1.Yıl) ..................................................................... 37 4.1.2. Çiçeklenme tarihi (2.Yıl) ..................................................................... 38 4.1.3. Bitki boyu (1.Yıl) ................................................................................ 40 4.1.4. Bitki boyu (2.Yıl) ................................................................................ 42 4.1.5. Fertil kardeş sayısı (1.Yıl) .................................................................. 44 4.1.6. Fertil kardeş sayısı (2.Yıl) .................................................................. 45 4.1.7. Başak uzunluğu (1.Yıl) ....................................................................... 48 4.1.8. Başak uzunluğu (2.Yıl) ....................................................................... 49 4.1.9. Başakta başakçık sayısı (1.Yıl) ........................................................... 51 4.1.10. Başakta başakçık sayısı (2.Yıl) ......................................................... 52 4.1.11. Başakta tane sayısı (1.Yıl) ................................................................ 54
iv

4.1.12. Başakta tane sayısı (2.Yıl) ................................................................. 56 4.1.13. Başakta tane ağırlığı (1.Yıl) ............................................................... 58 4.1.14. Başakta tane ağırlığı (2.Yıl) .............................................................. 59 4.1.15. Bin tane ağırlığı (1.Yıl) ..................................................................... 62 4.1.16. Bin tane ağırlığı (2.Yıl) ..................................................................... 63 4.1.17. Tane verimi (1.yıl) ............................................................................ 65 4.1.18. Tane verimi (2.yıl) ............................................................................ 66 4.1.19. Hasat indeksi (1.Yıl) ......................................................................... 69 4.1.20. Hasat indeksi (2.Yıl) ......................................................................... 70 4.1.21. Protein oranı (1.Yıl) .......................................................................... 72 4.1.22. Protein oranı (2.Yıl) .......................................................................... 74 4.2. Laboratuvar Çalışması Bulguları ve Tartışma ....................................... 78 4.2.1. DNA izolasyonunun optimizasyonu ................................................... 78 4.2.2. DNA konsantrasyonunun optimizasyonu ........................................... 80 4.2.3. MgCl2 konsantrasyonunun optimizasyonu .......................................... 80 4.2.4. Primer konsantrasyonunun optimizasyonu .......................................... 81 4.2.5. dNTP konsantrasyonunun optimizasyonu ........................................... 82 4.2.6. Taq Polimeraz enzim konsantrasyonunun optimizasyonu .................. 82 4.2.7. Diğer reaksiyon bileşenleri .................................................................. 82 4.2.8. PCR koşulları ....................................................................................... 83 4.2.9. Agaroz jel elektroforez ........................................................................ 84 4.2.10. RAPD polimorfizmi ve genetik mesafenin hesaplanması ................. 85 5. SONUÇ ..................................................................................................... 97
100 KAYNAKLAR .................................................................................................. Ö ÖZGEÇMİŞ ...................................................................................................... 106
v

SİMGELER DİZİNİ
S.D. Serbestlik derecesi K.T. Kareler toplamı K.O. Kareler ortalaması V.K. Varyasyon kaynakları g Gram µg Mikrogram L Litre mM Milimolar µl Mikrolitre µM Mikromolar nm Nanometre ng Nanogram nM Nanomolar DNA Deoksiribonükleik asit EDTA Etilendiamintetra asetik asit KCl Potasyum klorür M Molar Mg Magnezyum MgCl2 Magnezyum klorür pmol Pikamol PCR Polimeraz zincir reaksiyonu RAPD Random amlified polymorphic DNA (Rastgele çoğaltımış polimorfik DNA) RFLP Resrtriction fragment lenght polymorphism (kesilmiş parça uzunluğu
polimorfizmi) SDS Sodyum dedosil sülfat SSR Simple sequence repeats- mikrosatelit (Basit dizi tekrarları) STS Sequence tagget site-DNA dizisi bilinen Tris Tris (hidroksimetil) amino metan TBE Tris-borik asit- EDTA TE Tris-EDTA UV Ultra viole UPGMA Unweigted pair group method- ağırlıklı olmayan çift grup metodu Kb Kilo baz bp Base pair (Baz çifti) A Adenin G Guanin T Timin C Sitozin B Baz dNTP Deoksiribonükleosid trifosfat Taq Taq polimeraz enzimi U Ünite
vi

ŞEKİLLER DİZİNİ
Şekil 3.1. Tarla denemesin genel görünüşü ............................................................ 26 Şekil 4.1. İzole edilen genomik DNA’ların jel elektroforez görüntüsü ................ 78 Şekil 4.2. MA 7 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ...... 87 Şekil 4.3. A00 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ........ 88 Şekil 4.4. A01 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ........ 88 Şekil 4.5. A02 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ........ 88 Şekil 4.6. A03 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ........ 89 Şekil 4.7. A04 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ........ 89 Şekil 4.8. A06 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ........ 89 Şekil 4.9. MA 1 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ...... 90 Şekil 4.10. MA 3 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ...... 90 Şekil 4.11. MA 8 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ...... 90 Şekil 4.12. MA 9 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ...... 91 Şekil 4.13. MA 10 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri … 91 Şekil 4.14. OD 8 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ....... 91 Şekil 4.15. OPC 08 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri ... 92 Şekil 4.16. RAPD 1 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri .. 92 Şekil 4.17. B 379 primeri ile tritikale çeşit ve hatlarının RAPD-PCR profilleri …. 92 Şekil 4.18. PCR-RAPD yöntemine göre tritikale çeşit ve hatlarının genetik
benzerlik dendogramı ............................................................................. 95
vii

ÇİZELGELER DİZİNİ
Çizelge 3.1. Araştırma yerine ilişkin iklim verileri ........................................... 25 Çizelge 3.2. Araştırma yerine ilişkin toprak analiz sonuçları ............................ 27 Çizelge 3.3. RAPD analizinde kullanılan primerler, baz dizilişleri, GC (%)
oranları ve Tm sıcaklıkları (oC) ..................................................... 31
Çizelge 4.1. Farklı tritikale hatlarında çiçeklenme tarihine ilişkin varyans analizi .............................................................................................
37
Çizelge 4.2. Farklı tritikale hatlarında çiçeklenme tarihi ortalamaları (1 mayıstan sonra geçen gün sayısı) ..................................................
38
Çizelge 4.3. Farklı tritikale hatlarında çiçeklenme tarihine ilişkin varyans analizi ............................................................................................
38
Çizelge 4.4. Farklı tritikale hatlarında çiçeklenme tarihi ortalamaları (1 mayıstan sonra geçen gün sayısı) ..................................................
39
Çizelge 4.5. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama çiçeklenme tarihle (1 mayıstan sonra geçen gün sayısı) ................
40
Çizelge 4.6. Farklı tritikale hatlarında bitki boyuna ilişkin varyans analizi…... 41 Çizelge 4.7. Farklı tritikale hatlarında bitki boyu ortalamaları (cm) ................. 41 Çizelge 4.8. Farklı tritikale hatlarında bitki boyuna ilişkin varyans analizi…... 42 Çizelge 4.9. Farklı tritikale hatlarında bitki boyu ortalamaları (cm) ................. 42 Çizelge 4.10. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama bitki
boyu (cm)………………………………………………………… 43
Çizelge 4.11. Farklı tritikale hatlarında fertil kardeş sayısına ilişkin varyans analizi .............................................................................................
44
Çizelge 4.12. Farklı tritikale hatlarında fertil kardeş sayısı ortalamaları (adet/bitki) .....................................................................................
45
Çizelge 4.13. Farklı tritikale hatlarında fertil kardeş sayısına ilişkin varyans analizi .............................................................................................
45
Çizelge 4.14. Farklı tritikale hatlarında fertil kardeş sayısı ortalamaları (adet/bitki) .....................................................................................
46
Çizelge 4.15. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve fertil kardeş sayısı (adet/bitki) ...........................................................................
47
Çizelge 4.16. Farklı tritikale hatlarında başak uzunluğuna ilişkin varyans analizi ............................................................................................
48
Çizelge 4.17. Farklı tritikale hatlarında başak uzunluğu ortalamaları (mm) ....... 48 Çizelge 4.18. Farklı tritikale hatlarında başak uzunluğuna ilişkin varyans
analizi ............................................................................................ 49
Çizelge 4.19. Farklı tritikale hatlarında başak uzunluğu ortalamaları (mm)…… 49 Çizelge 4.20. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama başak
uzunluğu (mm) .............................................................................. 50
Çizelge 4.21. Farklı tritikale hatlarında başakta başakçık sayısına ilişkin varyans analizi ..............................................................................
51
Çizelge 4.22. Farklı tritikale hatlarında başakta başakçık sayısı ortalamaları (adet/başak) ....................................................................................
52
viii

Çizelge 4.23. Farklı tritikale hatlarında başakta başakçık sayısına ilişkin varyans analizi ...............................................................................
52
Çizelge 4.24. Farklı tritikale hatlarında başakta başakçık sayısı ortalamaları (adet/başak) ...................................................................................
53
Çizelge 4.25. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama başakta başakçık sayısı (adet/başak) .............................................
54
Çizelge 4.26. Farklı tritikale hatlarında başakta tane sayısına ilişkin varyans analizi ............................................................................................
55
Çizelge 4.27. Farklı tritikale hatlarında başakta tane sayısı ortalamaları (adet/başak) ...................................................................................
55
Çizelge 4.28. Farklı tritikale hatlarında başakta tane sayısına ilişkin varyans analizi ............................................................................................
56
Çizelge 4.29. Farklı tritikale hatlarında başakta tane sayısı ortalamaları (adet/başak) ..............................................................................
56
Çizelge 4.30. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama başakta tane sayısı (adet/başak) ....................................................
57
Çizelge 4.31. Farklı tritikale hatlarında başakta tane ağırlığına ilişkin varyans analizi ............................................................................................
58
Çizelge 4.32. Farklı tritikale hatlarında başakta tane ağırlığı ortalamaları (g/başak) ........................................................................................
59
Çizelge 4.33. Farklı tritikale hatlarında başakta tane ağırlığına ilişkin varyans analizi ............................................................................................
59
Çizelge 4.34. Farklı tritikale hatlarında başakta tane ağırlığı ortalamaları (g/başak) ........................................................................................
60
Çizelge 4.35. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama başakta tane ağırlığı (g/başak) .......................................................
61
Çizelge 4.36. Farklı tritikale hatlarında bin tane ağırlığına ilişkin varyans analizi .............................................................................................
62
Çizelge 4.37. Farklı tritikale hatlarında bin tane ağırlığıortalamaları (g) ............ 62 Çizelge 4.38. Farklı tritikale hatlarında bin tane ağırlığına ilişkin varyans
analizi ............................................................................................. 63
Çizelge 4.39. Farklı tritikale hatlarında bin tane ağırlığı ortalamaları (g)........ 64 Çizelge 4.40. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama bin
tane ağırlığı(g) ............................................................................... 65
Çizelge 4.41. Farklı tritikale hatlarında tane verimine ilişkin varyans analizi……………………………………………………………..
65
Çizelge 4.42. Farklı tritikale hatlarında tane verimi ortalamaları (kg/da) ........... 66 Çizelge 4.43. Farklı tritikale hatlarında tane verimine ilişkin varyans
analizi…………………………………………………………….. 66
Çizelge 4.44. Farklı tritikale hatlarında tane verimi ortalamaları (kg/da) ........... 67 Çizelge 4.45. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama tane
verimi(kg/da) ................................................................................. 68
Çizelge 4.46. Farklı tritikale hatlarında hasat indeksine ilişkin varyans analizi... 69 Çizelge 4.47. Farklı tritikale hatlarında hasat indeksi ortalamaları (%) .............. 70 Çizelge 4.48. Farklı tritikale hatlarında hasat indeksine ilişkin varyans analiz… 70 Çizelge 4.49. Farklı tritikale hatlarında hasat indeksi ortalamaları (%)………... 71 Çizelge 4.50. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama hasat
indeksi (%) ................................................................................... 72
ix

Çizelge 4.51. Farklı tritikale hatlarında protein oranına ilişkin varyans analizi……………………………………………………………..
73
Çizelge 4.52. Farklı tritikale hatlarında protein oranı ortalamaları (%) .............. 73 Çizelge 4.53. Farklı tritikale hatlarında protein oranına ilişkin varyans
analizi…………………………………………………………….. 74
Çizelge 4.54. Farklı tritikale hatlarında protein oranı ortalamaları (%) .............. 74 Çizelge 4.55. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama
protein oranı (%) ............................................................................ 75
Çizelge 4.56. Tritikale çeşit ve hatlarında spektrometrede ölçülen DNA miktarları .......................................................................................
79
Çizelge 4.57. PCR optimizasyonunu için test edilen parametreler ve elde edilen sonuçlar ..............................................................................
83
Çizelge 4.58. RAPD analizinde kullanılan polimorfik ve monomorfik bant veren pimerler ...............................................................................
86
Çizelge 4.59. Yirmibeş tritikale genotipi arasındaki genetik uzaklık matrisi (Nei 1972, Nei 1978) ....................................................................
93
x

1.GİRİŞ
Yetersiz ve dengesiz beslenme, günümüzde tüm dünya ülkelerinin önemle üzerinde
durduğu konuların başında gelmektedir. Özellikle az gelişmiş ya da gelişmekte olan
ülkelerde beslenme sorunları nedeniyle binlerce insan ölmekte veya bedensel ya da
zihinsel hastalıklara yakalanmaktadır. Beslenme sorunlarının çözülebilmesi amacıyla,
artan nüfusa bağlı olarak gıda maddelerinin de yeterli miktar ve kalitede üretilmesi
gereklidir. Hayvansal kaynaklı gıda ürünlerinin pahalı olması ve uzun süre
depolanmalarının zorluğu nedeniyle bitkisel kaynaklı gıda maddelerinin üretimi ön
plana çıkmaktadır. Tüm dünyada olduğu gibi ülkemizde de tahıl ve tahıla dayalı ürünler
insan beslenmesinde önemli bir yer tutmaktadır. İnsan beslenmesinde kullanılan
proteinin, dünyada % 52’si, ülkemizde ise % 80’i; karbonhidrat ve enerjinin ise
dünyada % 50’si, ülkemizde % 60’ı tahıllardan karşılanmaktadır (Kün 1996). Bu
nedenle dünya üzerinde tahıllar, geniş alanlarda ekimi ve üretimi yapılan ürün grubunu
oluşturmaktadırlar.
Dünyada en fazla ekimi ve üretimi yapılan tahıllar arasında buğday, mısır, çeltik, arpa,
ve çavdar yer almaktadır. Son yıllarda bunların dışında buğday ile çavdarın
melezlenerek kromozom sayısının iki katına çıkartılması sonucu elde edilen ve Tritikale
(xTriticosecale Wittmack) adı verilen cins önem kazanmaya başlamıştır. Dünya nüfusu
hızla artmakta, bazı bölgelerde beslenme sorunları yaşanmaktadır. Tritikale hızla artan
dünya nüfusunun beslenme sorunlarını çözmede alternatif bir tahıl olarak önem
kazanmıştır. İlk buğday x çavdar melezinin 1875 yılında İskoçya’da Wilson tarafından
yapıldığı kabul edilmektedir. Ancak, ilk buğday x çavdar melezleri kısır olup, fertil olan
ilk buğday x çavdar melezi 1891 yılında Almanya’da Rimpau tarafından
gerçekleştirilmiştir. 1915’li yıllarda doğal tritikale hibritlerine rastlanılmıştır. Tritikale
ismine ilk kez 1935 yılında Almanya’da basılan yazılı kaynaklarda rastlanılmaktadır. İlk
tritikale çeşidi (1966) Avrupa’da tescil edilmiş ve bunu Kuzey Amerika’da tescil edilen
‘Kosner’ isimli tritikale çeşidi izlemiştir. Tritikale AABBRR ya da AABBDDRR
genomik formülüne sahip, buğday ve çavdar türlerinin melezlenmesi sonucu elde edilen
yapay bir tahıl cinsidir (Briggle 1969).
1

Tritikaleler oktoploid ya da tetraploid olmasına karşın, tritikale çeşitlerinin çoğu
hekzaploid’dir (Lukaszewski ve Gustafson 1987). Buğday (AABB) ve çavdar (RR)
melezlenmesi sonucu elde edilen hekzaploid tritikaleler birincil hekzaploid tritikale
(AABBRR) olarak adlandırılırlar. Hekzaploid tritikalelerin melezlenmesi ya da
hekzaploid buğday x tritikale melezleri ve oktoploid tritikaleler ise ikincil hekzaploid
tritikale (AABBRRDD) olarak adlandırılır. İkincil tritikalelerin avantajı, ekmeklik
buğdaydan D genomunun katılması sonucu genetik çeşitliliğinin artmış olmasıdır.
Tritikalenin yazlık ve kışlık olarak yetiştirilen formları mevcut olup, buğdaya oranla
daha az kardeşlenme özelliğinde, daha uzun boylu, ve daha büyük başaklı olduğu
bildirilmektedir. Tritikale çeşitleri genelde kılçıklıdır, ancak son yıllarda kılçıksız
tritikale formları da geliştirilmiştir (Briggle 1969, Stallknecht vd 1996).
Buğday x çavdar melezinin amfidiploidi olan tritikalede ilk araştırmalar daha çok
taksonomi ve evrim çalışmalarını içermiştir. Daha sonra verim ve hektolitre ağırlığının
yükseltilmesinin yanında bitki boyunun kısaltılması çalışmalarına öncelik verilmiş ve
günümüz tritikale çeşitlerinde bu özelliklerin iyileştirilmesiyle ekim alanlarında önemli
bir artış sağlanmıştır.
Dünyada tritikale insan gıdası ve hayvan yemi olarak değişik şekillerde kullanılmakta
ve her geçen gün ekim alanları artmaktadır. Son verilere göre Dünyada 3.1 milyon ha
ekim alanına, 10.2 milyon ton üretime ve 3300 kg /ha verime sahip olan tritikale’nin en
fazla tarımının yapıldığı ülkeler; Polonya, Almanya, Avustralya, Çin ve Fransa’dır
(Anonymous 2003). Bugün üretimi yapılan tritikale çeşitlerinin tamamına yakını
makarnalık buğday ile çavdarın melezlenmesi sonucu elde edilen çeşitlerdir. Diğer
bitkilerde olduğu gibi tritikale ıslahında da amaç, daha verimli ve daha kaliteli yeni
çeşitlerin elde edilmesidir.
Bilindiği gibi verim üzerinde çeşidin genotipi kadar bölgeden bölgeye değişen çevre
koşulları da etkili olmaktadır. Çeşitler, değişik çevre koşullarında farklı tepkiler
gösterebilmektedir. Bu amaçla ıslah edilen üstün hat ya da çeşitlerin denemeye alınarak
bölge koşullarına uyum sağlayan ve yüksek verimli olanların belirlenmesi
gerekmektedir.
2

Tritikale, çavdarın iklim ve toprak özellikleri yönünden fazla seçici olmayan özelliği ile
hastalık ve zararlılara dayanıklılığını; buğdayın yüksek verim ve kalitesini içeren bir
cinstir. Tritikale; serin iklim tahıllarından buğday ve arpaya göre olumsuz koşullara
(iklim ve toprak) daha fazla dayanmakta ve stres koşullarında da olsa belirli bir verime
ulaşabilmektedir. Almanya’da yapılan bir araştırma sonuçlarına göre tritikale buğday ve
arpaya oranla daha düşük sıcaklıklara dayanabilmektedir (Sachs vd 1999). Tritikale,
bitki boyunun uzun olmasına karşın sapının sağlam ve yatmaya dayanıklı olması,
buğdaya göre daha az kardeşlenmesi ve başakçık dış kavuzlarının çavdardaki gibi tüylü
olması ile dikkati çekmektedir. Marjinal alanların değerlendirilmesinde öncelikli
bitkinin tritikale olduğu ve yeni çeşitlerin geliştirilmesiyle ekim alanı ve üretiminde
önemli artışların sağlanacağı belirtilmektedir (Müntzing 1989, Mergoum vd 1992, Kün
1996).
Tritikale, özellikle buğday tarımına uygun olmayan toprak derinliği az, çorak ve kışları
çok sert geçen bölgelerde buğdaydan daha verimli olabilmektedir. Tritikale diğer serin
iklim tahıllarına göre topraktan daha iyi yararlanabilmekte ve değişen çevre
koşullarında daha stabil durumda bulunmaktadır. Özellikle, marjinal alanların
değerlendirilmesinde ve artan yem açığının kapatılmasında tritikale önemli bir alternatif
bitki olarak karşımıza çıkmaktadır. Son yıllarda yapılan araştırmalara göre, tritikalenin
yem verimi, kuru madde oranı, lif içeriği, hazım olma derecesi gibi hayvan beslemede
önemli özellikler yönünden diğer tahıllara eşdeğer ya da daha üstün olduğu
saptanmıştır. Protein oranı ve amino asit içeriği ile amino asit dengesi buğdaya göre
daha iyi durumdadır (Fernandez-Figares 2000). Ayrıca tanelerinin fosfor, mangan,
demir ve bakır içeriği de oldukça yüksektir. Bu nedenle tritikale hayvan beslenmesinde
arpa, yulaf ve buğdayın yerine kolaylıkla kullanabilecektir. Özellikle domuz ve kanatlı
hayvanların beslenmesinde yem rasyonlarına katılması önerilmektedir (Çiftçi vd 2003).
Tritikale unu, buğday ve çavdar ununa katılarak, ekmek, pasta ve bisküvi yapımında
kullanılmaktadır. Tritikalede tane kırışıklığı, hektolitre ağırlığının ve un randımanının
düşük olması nedeniyle unu buğday ununa belli oranlarda (% 50-70) karıştırılarak
kullanılmasına karşın, son yıllarda geliştirilen yeni çeşitlerde % 100 tritikale unu ile
3

ekmek yapılabilmektedir. Üretici ve tüketicinin isteklerine uygun yeni çeşit ve
hatlarının geliştirilmesi ile tritikale ekim alanları her geçen gün artmaktadır.
Ülkemizde üretim izinli olarak ilk kez Bakırçay tritikale çeşidi yetiştirilmeye başlamış,
daha sonra Tatlıcak 97, Tacettinbey, Presto, Karma 2000, Melez 2001 ve MİKHAM
2002 tritikale çeşitleri tescil edilmiştir. Adana, Konya, Tokat ve Sivas gibi illerimizde
yetiştirilmeye başlanan ve ekim alanları giderek artan tritikale de sağlıklı istatistiki
veriler bulunmamaktadır.
Ülkemizde hayvanlarımızın ihtiyacı olan kaba yemin karşılanmasında, meralarımızın
aşırı otlatılmasının önlenmesinde ve erozyon kontrolünde önemli katkıları olabilecek bir
bitki olan tritikalenin alım garantisinin olmaması, çeşit geliştirme ve tohumluk
sorunlarının bulunması ve çiftçinin yeterince bitkiyi tanımaması nedeniyle tarımı
istenilen düzeyde gelişememiştir. Tritikale tarımının yaygınlaştırılması gerekmektedir.
Son yıllarda bitki ıslahçıları tarafından kullanım amaçlarına uygun, tarımsal özellikleri
üstün genotipler elde edilmiş olmasına rağmen çalışmalar hızla devam etmektedir. Bu
amaçla, çeşit geliştirme çalışmalarında bilinen klasik ıslah çalışmalarının yanında bu
çalışmaları hızlandıracak, başarı şansını artıracak ve onları daha etkin kılacak moleküler
tekniklerin kullanılması büyük önem taşımaktadır.
Bitkilerde üstün bireylerin belirlenmesinde, morfolojik özellikleri yansıtan agronomik
verilerin yanında, daha kesin tanılara dayalı yöntemler son yıllarda hızla kullanıma
girmiştir. Abiyotik (sıcaklık vb.) ve biyotik (virüs vb.) koşullardan etkilenmeyen ve
direkt olarak karakterlerin orijinini temel alan moleküler markörler (protein ve DNA
markörler) birçok bitki türünde tanımlama amacıyla kullanılmaktadır. Genetik
markörler, kalıtım şekilleri morfolojik (çiçek rengi gibi), biyokimyasal (izoenzimler
gibi) ve DNA düzeyinde (moleküler markörler) izlenebilen karakterlerdir. Bu
karakterlerin markör (işaret) olarak isimlendirilmesinin nedeni, çalışılan organizmadaki
ilgilenilen diğer özelliklerin genetiği hakkında dolaylı olarak ta olsa bilgi
verebilmeleridir. Moleküler markörler DNA’nın aktif bölgeleri (genler) veya herhangi
4

bir genetik kodlama fonksiyonuna sahip olmayan DNA parçaları şeklinde olabilir
(Yıldırım ve Kandemir 2001).
Moleküler markörler; bitki organizmalarında detaylı fiziksel ve genetik kromozom
haritalarının hazırlanmasında, bitkilerde istenilen özelliğin dolaylı olarak seleksiyon
yaparak seçilmesi yoluyla klasik ıslah metodlarının başarı şansını artırmada yaygın
olarak kullanılmaktadır. Bu kullanımlarının dışında, moleküler markörler, bitkilerde gen
kaynaklarının özelliklerinin belirlenmesinde, genetik incelemelerde, transgenik
bitkilerin belirlenmesi gibi bir çok alanda kullanım alanı bulmaktadır (Gupta vd 1999).
DNA markörler, melezlemeye dayalı (RFLP), PCR’a dayalı (RAPD, SSR, ALFP, STS
vb.) ve dizi analizlari olmak üzere başıca 3 grupta incelebilmektedir (Gupta vd 1999).
PCR (Polimeraz Zincir Reaksiyonu) DNA polimeraz enziminin kullanılmasıyla yapay
şartlarda DNA üretilmesini ifade etmektedir (Seçilmiş DNA parçasının çok sayıda
kopyasını yapan enzimatik bir metodtur). Bu yapay üretim için genelde 6-25 nükleotid
uzunluğunda başlatıcı DNA’lar (primerler) kullanılmakta, DNA üretim işlemi birbirini
izleyen bir seri ve çok özel işlemler sonucu sağlanmaktadır. Önce 95 °C civarında bir
sıcaklık kullanımıyla DNA iplikçiklerinin açılması sonucu DNA tek iplikçik haline
gelmekte, sonra 30-60 ° C civarında bir sıcaklıkta başlatıcı DNA’ nın kalıp DNA’ ya
bağlanması, daha sonra da 72 °C de DNA üretimi sağlanmış olmaktadır. Bu devrelerin
her biri 1-2 dakika kadar sürmekte, bu 3 devre istenildiği kadar tekrarlanabilmekte ve
istenilen miktarda DNA üretimi yapılabilmektedir (Maniatis vd 1982, Kumar 1989).
Kleppe vd (1971), tarafından PCR’ın teorik temelleriyle ilgili olarak ilk makaleler
yayınlanmıştır. Ancak, bu (Kary Mullis vd 1980)’de Genomik DNA’dan çok sayıda
tek kopya genlerin çoğaltılması ile ilgili çalışmalarına kadar bilim çevrelerince pek
dikkati çekmemiştir. PCR’ ın ilk uygulamasında kullanılan enzimin denatürasyon adımı
süresince inaktif hala dönüşmesi nedeniyle her döngü süresince E. coli DNA
polimerazın klenow fragmentlarinden eklemek gerekiyordu. Daha sonra Thermus
aquaticus’dan ısıya dayanıklı Taq DNA polimeraz enziminin elde edilmesiyle bu sorun
ortadan kaldırılmış ve PCR çalışmalarının otomasyonu sağlanmıştır. Bu sayede Taq
5

DNA polimerazın yapışma (annealing) ve uzaması (extension) için yüksek sıcaklığın
kullanımına olanak sağlanmıştır (Kumar 1989).
PCR’ ın gelişimi ve özel DNA parçalarının çoğaltılması yolu ile DNA polimorfizmi
belirlenebilmektedir. Protein veya DNA markörlerine dayanan haritalar ise ‘moleküler
haritalar’ olarak adlandırılmaktadır (Reiter vd 1993). Bireyler arasındaki genetik uzaklık
bir çok tarla bitkisinde belirlenmeye çalışılmıştır. DNA dizilişindeki polimorfizmi
belirlemek amacıyla RFLP (Kesilmiş Çoğaltılmış Polimorfik DNA), AFLP (Çoğaltılmış
Parça Uzunluğu Farklılığı), SSR (Basit Dizi Tekrarları), STS (Dizisi Etiketlenmiş
Alanlar) ve RAPD (Rastgele Çoğaltılmış DNA Farklılığı) gibi teknikler geliştirilmiştir
(Gupta vd 1999).
PCR’a dayalı DNA markörler günümüzde ön plana çıkmıştır. Bunlardan RAPD
(Rastgele Çoğatılmış DNA Farklılığı) bugün en çok kullanılan yöntemler arasındadır.
RAPD markörleri spesifik (özel) primerleri ve genomik DNA’ yı kullanan PCR’ ın
amplifiye olmuş (çoğatılımış) ürünleridir (Williams vd 1990). Bu teknikte 6-10
nükleotid uzunluğundaki başlatıcı DNA’ lar kullanılarak genom üzerinde rasgele
bölgelerin DNA amplifikasyonu (çoğaltılması) gerçekleştirilmektedir. Reaksiyon
şartlarının spesifik (özel) olmaması, rasgele çoğaltıma izin vermektedir. DNA parçaları
bir agaroz jel üzerinde elektroforesis’e tabi tutulduğunda bazı parçaların bazı
genotiplerde üretilip bazı genotiplerde üretilmediği gözlenmektedir. Bu işlem açılan bir
populasyonda yapıldığında ebeveynlere (anaçlara) ait üretim motiflerine bakarak
döllerin genotip analizleri gerçekleştirilebilir (Yıldırım ve Kandemir 2001).
Diğer PCR esaslı teknikler gibi RAPD tekniği de haritalama ve karakterizasyon
çalışmalarında daha az zamana, çalışmaya ve gidere gereksinim duyulması nedeniyle
daha çok tercih edilmektedir. Özellikle RAPD yöntemi çok az bir DNA miktarına
gereksinim duyulmasından ötürü tahılların yanında bir çok bitki türünde ümitvar
sonuçlar vermiştir (Williams vd 1990, Devos ve Gale 1992).
RAPD ürünleri için bireyler arasında ortaya çıkan bir çok varyasyon (farklılık) DNA
üzerindeki baz çiftlerinin yer değiştirmesi ya da artışı/azalışı nedeniyledir. RAPD
6

markörleri taksonomik sınıflandırmada, sistematik ilişkilerin belirlenmesinde,
populasyon genetiği çalışmalarında, türler arası melezlemelerde ve anaçların
belirlenebilmesinde kullanılabilmektedir (Fahima vd 1999).
PCR’ a dayalı moleküler markörler, zamanı kısalttığı, moleküler haritalamada gerekli iş
gücü ve harcamayı azalttığı için günümüzde geniş ölçüde kullanılmaktadır (Gupta vd
1999). Rastgele dizilişi belirlemek amacıyla tek DNA primeri kullanımını içeren
RAPD tekniğinin tahılları da içeren bir çok bitki grubunda kullanım olanağı
bulunmaktadır (Williams vd 1990). RAPD tekniği tahıllarda; genetik haritaların
hazırlanmasında, gen işaretlemede, çeşit belirlemede, populasyonlarda ve türlerdeki
varyasyonun ortaya çıkartılmasında, türler, alt türler ve çeşitler arasındaki filogenetik
ilişkilerin çalışılması gibi bir çok alanda kullanılmaktadır (Gupta vd 1999).
Kültürü yapılan bitkiler içerisinde genetik uzaklık, çeşitler arasında ortalama genetik
farklılığın belirlenmesini sağlar. Bir tür içerisindeki çeşitler arasındaki genetik ilişkinin
belirlenmesi, yapılacak melezlemeler için uygun ana ve babanın seçiminde ve oluşacak
döllerin performanslarını tahmin etmede kullanışlı olduğu belirlenmiştir (Frei vd 1986).
Bitki ıslahı programlarında ebeveynlerin benzerliği hakkındaki bilgi, genetik olarak
uniform ya da çok benzer olan bireylerin melezlemede kullanılmasını önleyecek ve
böylece uzun süreli seleksiyonlardan elde edilecek kazançlar tehlikeye atılmayacak,
genetik kaynaklar korunacaktır (Sun vd 2003).
Bu çalışmada, bazı tritikale hatlarının morfolojik özelliklerinin belirlenmesine ve DNA
markörleri ile genetik karakterizasyonu yapılmasına çalışılmıştır. Bu amaçla, tritikale
hatlarında çiçeklenme tarihi, bitki boyu, fertil kardeş sayısı, başak uzunluğu, başakta
başakçık sayısı, başakta tane sayısı, başakta tane ağırlığı, hasat indeksi, birim alan
verimi ve protein oranı gibi agronomik özellikler incelenmiştir. Ayrıca RAPD-PCR
tekniği kullanılarak hatlar arasında genetik benzerlik ya da farklılıkların belirlenmesine
çalışılmıştır. Tritikale hatlarının (morfolojik görüntü olarak birbirine benzer) morfolojik
ve genetik karakterizasyonunun yapılması bu hatların bitki ıslahı (yeni çeşitlerin
geliştirilmesi) amacıyla daha etkin olarak kullanılmasında faydalı olacaktır.
7

2. KAYNAK ÖZETLERİ 2.1. Morfolojik Özelliklerin Değerlendirilmesi Dünyada insan ve hayvan beslenmesinde önemli bir yere sahip olan tritikale, ülkemizde
halkımız tarafından tam olarak bilinmediği ve yeterli çeşit bulunmadığı için gereği
kadar yetiştirilememektedir, ancak ülkemizin değişik bölgelerinde tritikale yetiştirilme
olanağı bulunmaktadır. Tritikale konusunda yapılmış yerli ve yabancı çalışmalar
aşağıda özetlenmiştir.
Sapra vd (1973), buğday ve çavdar melezi olan tritikalenin kuru ot veriminin buğday,
arpa ve çavdarla aynı olduğunu, ancak tritikalenin Amerika Birleşik Devletlerinin
Güneydoğu eyaletlerinde ilkbaharda biçildikten sonra yazlık buğdaya oranla daha hızlı
gelişme gösterdiğini bildirmektedirler.
Szigat ve Müller (1975), Macaristan koşullarında yaptıkları çalışmada; tritikaleden
çevre koşullarına göre değişmekle birlikte, buğday ve çavdara oranla % 29-49 daha
fazla tane verimi elde ettiklerini vurgulamışlardır.
Krolow (1977), Almanya’da yaptığı çalışmasında; tritikale, buğday ve çavdarın parsel
verimlerini karşılaştırmıştır. Hekzaploid tritikalede 780 g/m2, oktoploid tritikalede 610
g/m2, ekmeklik buğdayda 880 g/m2 ve diploid çavdarda 885 g/m2 parsel verimi elde
ettiğini belirtmiştir.
Demir vd (1979), Ege bölgesi lokasyonlarında ve Diyarbakır yöresinde yürüttükleri
denemelerde, tritikale hatlarının ekmeklik ve makarnalık buğdaylardan sırasıyla (% 5-
44) ve (% 5-71) daha fazla verim elde ettiklerini bildirmişlerdir.
Baier vd (1980), Güney Brezilya’nın asitli topraklarında, kumlu topraklarda, yüksek
yerlerde ve kurak bölgelerde diğer tahıllara göre tritikalenin en uygun ve erkenci bir
tahıl olduğunu belirtmişlerdir.
8

Demir vd (1986), 1982-86 yılları arasında CIMMYT’den sağlanan iki tritikale hattı,
Cumhuriyet-75 ekmeklik, Gediz–75 makarnalık buğday çeşitleri ile Bakırçay tritikale
çeşidini materyal olarak kullandıkları çalışmalarında; hektolitre ağırlığı buğday
çeşitlerine göre düşük olan tritikalenin diğer özellikler (verim, bin tane ağırlığı ve bitki
boyu) yönünden buğdaylara yakın ve üstün değerler gösterdiğini bildirmişlerdir.
Genç vd (1987), 1985-1987 yıllarında Çukurova koşullarında iki ayrı deneme olarak
yürüttükleri ve tritikalenin verim ve verim öğelerini inceledikleri çalışmalarında; 25
tritikale hattı ile Cumhuriyet-75 ekmeklik ve Gediz-75 makarnalık buğday çeşitlerini
kullanmışlardır. Deneme I ve Deneme II de sırasıyla; tritikale hatlarında, başakta tane
sayısını 37.9-50.7 adet ve 32.3-51.3 adet, başak tane verimini 1.49-2.15 ve 1.61-2.30 g,
bin tane ağırlığını 35.9-49.4 ve 36.6-48.5 g, dekara tane verimini 540-667 kg/da ve 576
- 673 kg/da ile en düşük ve en yüksek değerler olarak elde ettiklerini vurgulamışlardır.
Genç vd (1988), Çukurova koşullarında; Orso, Cumhuriyet-75 ekmeklik, Balcalı-85
makarnalık buğday çeşitleri, iki sıralı Kaya ve altı sıralı Gen arpa çeşitleri ile CIMMYT
kökenli Mapache ve ICARDA kökenli N.Ic Bulk 1-81 tritikale hatlarını materyal olarak
kullandıkları çalışmalarında; Tritikale hatlarında 122.59-126.93 cm ile en uzun bitki
boyu, 49.00-52.99 adet başakta tane sayısı, 1.70-2.20 g başakta tane ağırlığı, 34.99-
40.45 g bin tane ağırlığı ve 432.60-447.60 kg/da tane verimi elde ettiklerini
bildirmişlerdir. Araştırıcılar N.Ic. Bulk 1-81 tritikale hattının diğer tahıl cinslerine göre
% 8 daha fazla verim sağladığının ve kurak koşullarda buğdayla rekabet etme şansının
yüksek olduğunu açıklamışlardır.
Yağbasanlar vd (1988), Çukurova koşullarında N. Ic. Bulk 1-81 tritikale çeşidinde altı
farklı azot dozunun (0, 4, 8, 12, 16 ve 20 kg/da) ve dört değişik tohumluk miktarının
(10, 15, 20 ve 25 kg/da) etkisini araştırmışlardır. Bitki boyu, başakta tane sayısı, başakta
tane ağırlığı, bin tane ağırlığı, hektolitre ağırlığı ve tane veriminde azot dozları ve
tohumluk miktarları arasında önemli farklılıklar belirlediklerini, en uygun azot dozunun
12 kg/da ve tohumluk miktarının 20 kg/da olduğunu bildirmişlerdir.
9

Maurice (1989), tritikale üzerinde yapılan çalışmaların büyük bir kısmının hayvan
beslenmesindeki öneminin oluşturduğunu, agronomik özellikler üzerindeki çalışmaların
daha az olduğunu, özellikle kanatlı hayvanların ve domuzların beslenmesinde yem
rasyonuna tritikalenin katılmasının olumlu sonuçlar verdiğini belirtmiştir.
Müntzing (1989), tritikalenin alkali topraklarda iyi yetiştiğini, buğdaya göre alkali ve
kireçli topraklarda daha güçlü bir çıkış gösterdiğini, bakır, çinko ve manganez eksikliği
gösteren ve bor fazlalılığı olan topraklarda buğdaya göre daha yüksek verim sağladığını
bildirmiştir.
Ülger vd (1989), CIMMYT ve ICARDA’dan sağlanan 540 tritikale hattından seçilen 46
genotipi materyal olarak kullanarak 3 yıl süreyle yürütmüş oldukları çalışmada, Merino-
S-JLO tritikale hattından başakta tane sayısını 47.5 adet, başakta tane ağırlığını 2.34 g,
bin tane ağırlığını 48.2 g ile en yüksek değerleri elde ettiklerini açıklamışlardır.
Yağbasanlar vd (1989), ICARDA ve CIMMYT’den sağlanan Beagle, Palouse,
Mapache, Bacum, Siskiyou ve N.Ic Bulk 1-81 tritikale hatlarını, Orso ve Cumhuriyet –
75 ekmeklik buğday çeşitleri ile denemeye aldıklarını, tritikale hatlarındaki 3 yıllık bitki
boyu ortalamalarının 108-126 cm, başak uzunluğunun 8.4-13.2 cm, başakta başakçık
sayısının 20.2-33.6 adet, başakta tane sayısının 44.4-51.9 adet, başakta tane ağırlığının
1.52-2.02 g, bin tane ağırlığının 34.0-42.4 g ve tane veriminin 281-434 kg/da arasında
değiştiğini, bin tane ağırlığı dışında ele alınan diğer özelliklerde ekmeklik ve
makarnalık buğday çeşitlerine göre bazı tritikale hatlarında daha yüksek değerler elde
ettiklerini bildirmişlerdir.
Yağbasanlar vd (1990), 1987-88 yıllarında Çukurova ve Şanlıurfa koşullarında
yürütülen denemelerde; 13 tritikale hattı ile Orso ve Balcalı-85 buğday çeşitleri
kullanılmıştır. Çukurova koşullarındaki denemede; tritikale hatlarında bitki boyu 111.0-
154.3 cm, başakta tane sayısı 45.9-58.1 adet, başakta tane ağırlığı 1.84-2.89 g, bin tane
ağırlığı 37.1-51.6 g, tane verimi 527.1-678.3 kg/da olarak belirlerken, Şanlıurfa
koşullarında sırasıyla; 110.0-139.8 cm, 52.3-68.7 adet, 1.99-2.39 g, 30.0-36.8 g, 285.0-
10

389.0 kg/da olarak belirlediklerini, incelenen özelliklerde Orso ekmeklik buğday
çeşidinin daha düşük değerler gösterdiğini vurgulamışlardır.
Yağbasanlar (1991), 1988-89 yılında Çukurova koşullarında CIMMYT’den sağlanan
beş tritikale çeşidinde yabancı döllenme ve tohum tutma oranını belirlemek amacıyla
yürüttüğü çalışmada, tohum tutma oranını % 20.69 olarak saptadığını, bu oranın hatlar
arasında % 17.92-22.40 olarak değiştiğini bildirmiştir.
Mergoum vd (1992), tritikalenin dünyada tahıl yetiştirilen yarı kurak ve kurak bölgeler
için önemli bir bitki olduğunu, bu bölgelerde tritikalenin buğdaya göre daha yüksek
verim verdiğini ve gelecekte tritikalede kalitenin iyileştirilmesiyle daha geniş alanlara
yayılabileceğini vurgulamışlardır.
Milovanoviç (1993), dokuz hekzaploid tritikale çeşidini buğday ve çavdarla
karşılaştıran araştırıcı, iki yıl süreyle yaptığı deneme sonucunda tritikale hatlarının
buğday ve çavdara göre daha yüksek tane verimi verdiğini belirtmiştir.
Feil ve Fossati (1995), yaptıkları bir araştırmada; tritikale de tane veriminin 5020-6710
kg/ha 1000 tane ağırlığının 31.1-41.7 g, hektolitre ağırlığının 70.9-57.5 kg, tanelerinde
ortalama protein miktarının 96- 122 g kg –1, P miktarının 3.61-4.33 g kg –1, K miktarının
4.35-5.46 g kg –1, Mg miktarının 1.20-1.46 g kg –1, Ca miktarının 350-498 µg g-1, Fe
miktarının 28.1-35.6 µg g-1, ve Zn miktarının 20.8–31.3 µg g-1 olduğunu
bildirmişleridir. Araştırıcılar daha uygun iklim koşulları ve verimli çeşitlerin
kullanılmasıyla verim artışının sağlanabileceğini ancak, protein oranı ve mineral madde
konsantrasyonlarının azalacağını bildirmişlerdir.
Horlein ve Valentine (1995), tritikalenin ekmeklik kalitesinin, ekmeklik buğdaya göre
daha düşük olmasına rağmen, tritikale tanelerinin mutlak gerekli amino asitler
yönünden oldukça zengin olmasının tritikaleyi buğdaya oranla daha değerli bir besin
kaynağı yaptığını vurgulamıştır.
11

Varughese vd (1996), çevre koşullarının ve yetiştirme tekniklerinin bütün bitkilerde
olduğu gibi tritikalede de verim ve verim öğeleri üzerinde etkili olduğunu ve tritikale de
verimin 250-970 kg/da arasında değiştiğini belirtmişlerdir.
Gill (1996), Polonya’da yaptığı çalışmasında, kışlık ve yazlık olarak yetiştirilen
tritikalede tanelerin kimyasal bileşenlerini ortaya koymaya çalışmıştır. Araştırıcı yazlık
olarak yetiştirilen tritikale tanelerinden elde edilen unların, kışlık olarak yetiştirilen
tritikale tanelerinden elde edilen unlara oranla daha yüksek protein içeriğinin olduğunu
bildirmiştir.
Sencer vd (1997), Tokat-Artova koşullarında yürüttükleri çalışmada; 15 tritikale hattı ile
12 buğday çeşidi ve bir çavdar populasyonunda verim ve verim öğelerini incelediklerini
ve tritikale hatlarında başakta tane sayısını 35.6–44.0 adet, başak tane verimini 1.1-1.6
g, bin tane ağırlığını 29.9-38.9 g, dekara tane verimini ise 164.9-363.6 kg/da arasında
saptadıklarını bildirmişlerdir.
Kınacı (1998), toprak ve iklim koşullarının üretimi zorlaştırdığı alanlarda, buğday ve
çavdara göre daha yüksek verim sağlayan tritikalenin dünyada ekim alanlarının hızla
arttığını, ülkemizde yem hammaddesinin sorunlarının çözümünde tritikalenin önemli bir
yere sahip olacağını belirtmiştir.
Bağcı vd (1999), farklı 2 lokasyonda ve sulu koşullarda yürütülen tritikale geliştirme
çalışmalarında; 1996-97 yılında Tatlıcak- 97 tritikale çeşidin de 555-652 kg/da, BDMT-
19’da 561 - 731 kg/da, Gerek-79 ekmeklik buğday çeşidinde 402-553 kg/da tane verimi
elde ettiklerini bildirmişlerdir.
Taşyürek vd (1999a), Sivas- Şarkışla koşullarında Tatlıcak-97 tritikale çeşidine 5 farklı
azotlu gübre dozu uygulayarak 4 lokasyonda yürüttükleri çalışmada; başakta tane
sayısını lokasyonların ortalaması olarak; 35.9-39.8 adet, bin tane ağırlığını 35.6-37.6 g,
hasat indeksini % 25.3-30.6 ve dekara tane verimini 252-460 kg/da arasında
belirlediklerini, ayrıca inceledikleri özelliklerde 8 kg/da azot uygulamasının en yüksek
değeri verdiğini vurgulamıştır.
12

Taşyürek vd (1999b), Sivas- Şarkışla koşullarında yapılan çalışmada; dört ekmeklik, iki
makarnalık, dört arpa çeşidi ve Tatlıcak-97 tritikale çeşidini materyal olarak
kullanmışlar, başakta tane sayısı, başak tane verimi, sap verimi ve tane verimi yönünden
en yüksek ortalamaların Tatlıcak-97 çeşidinden elde edildiğini belirtmişlerdir. Ayrıca,
Tatlıcak-97 tritikale çeşidinde başakta tane sayısının 41.9 adet, başak tane veriminin
1.73 g, bin tane ağırlığının 37.7 g, hektolitre ağırlığının 77.2 kg, sap veriminin 1700
kg/da, hasat indeksinin % 31.3 ve tane veriminin 531.7 kg/da olarak saptamışlardır.
Yağbasanlar vd (1999), Çukurova’nın kıraç koşullarında 1996-1998 yılları arasında
yürüttükleri çalışmada; Fahad-1 tiriticale hattına 12, 16 ve 20 kg/da azotlu gübre dozları
ile 350, 450 ve 550 tohum/m2 ekim sıklığı uyguladıklarını ve en yüksek bin tane
ağırlığını 16 kg/da azot 450 tohum/m2 ekim sıklığından, en düşük bin tane ağırlığını ise
12 kg/da ve 550 tohum/m2 uygulamasından elde ettiklerini, ayrıca iki yılın ortalaması
olarak; bin tane ağırlığını 47.3 g, hektolitre ağırlığını 72.9 kg ve tane verimini 429.7
kg/da olarak saptadıklarını belirtmişlerdir.
Ünver (1999), Ankara koşullarında 1996-1997 yıllarında yürüttüğü çalışmasında; Tarla
Bitkileri Merkez Araştırma Enstitüsü’nce CIMMYT’ten sağlanan on yedi adet tritikale
ıslah hattı ile çeşit adayı (Tatlıcak-97) materyal olarak kullanmıştır. İki yıl
ortalamalarına göre; bitki boyunun 103.20-123.69 cm, bitkide kardeş sayısının 2.77-
3.95 adet, başak uzunluğunun 10.23-13.35 cm, başakta tane sayısının 41.35-55.13 adet,
başak tane veriminin 1.71-2.34 g, hasat indeksinin % 21.68-31.51, tane veriminin
206.25-340.00 kg/da ve bin tane ağırlığının 43.76-53.90 g arasında değiştiğini
bildirmiştir.
Fernandez-Figares vd (2000), Akdeniz çevresinde kurak koşullarda tritikale
tanelerinden 300 kg/ha ve sulanan koşullarda ise 900 kg/ha ham protein elde
edilebileceğini, tritikalenin bu yönüyle enerji ve protein kaynağı olarak önemli bir yere
sahip olduğunu belirtmişlerdir. Araştırıcılar ayrıca, buğday ve arpa tanelerinin düşük
oranda lysine yanında methionine ve threonine gibi diğer amino asit içermelerine
karşın, tritikale tanelerinin yüksek seviyelerde lysine, kolayca hazmolabilir amino asit
içerdiğini bildirmektedirler.
13

Wesolowski ve Gregorczyk (2000), Polonya’da 1987-91 yılları arasında Bolero kışlık
tritikale çeşidini kullanarak yaptıkları ekim nöbeti çalışmalarında; bezelyeden sonra
ekilen tritikalede 5.74 t /ha, patatesten sonra ekilen tritikaleden 5.49 t/ha tane verimi
aldıklarını, tritikalede tane veriminin, ekim nöbetine kışlık tahıl alındığında (buğday ya
da çavdar) önemli ölçüde azaldığını bildirmektedirler.
Koc vd (2000), Polonya’da yaptıkları 3 yıllık tarla deneme sonuçlarına göre, yazlık
tritikale Maja çeşidine, 0-120 kg /ha arasında değişen N dozu uygulamışlar ve protein
oranının azot dozundaki artışa paralel olarak % 0.3–1.4 oranında artış gösterdiğini
belirlemişlerdir. Araştırıcılar en yüksek tane veriminin (4.83 t/ha) ve en yüksek protein
oranının (% 12) 100 kg /ha N dozundan elde edildiğini belirtmişlerdir.
Çöplü (2001), Bursa koşullarında iki yıl süreyle yürüttüğü çalışmasında, dokuz tritikale
hattında verim ve verim öğelerini, mayoz bölünmede kromozom eşleşmelerini ve yarım
diallel melezlemede kalıtım derecelerini incelemiştir. Araştırıcı, bitki boyunu 109.20-
125.25 cm, başak uzunluğunu 12.45-15.55 cm, başakta başakçık sayısını 25.40-28.85
adet, başakta tane sayısını 62.93-78.95 adet, başakta tane ağırlığını 2.92-4.12 g, bin tane
ağırlığını 46.34-62.03 g ve tane verimini 744.6-960.3 kg/da arasında saptamıştır.
Çengel (2001), Ankara, Haymana koşullarında 24 tritikale hattı ve 1 adet tescilli çeşidi
kullanarak yaptığı çalışmada; kontrol ve ıslah hatlarında, bitki boyunun 88.94-128.41
cm, fertil kardeş sayısının 2.10-2.63, başak uzunluğunun 8.58-11.77 cm, başakta
başakçık sayısının 22.77-29.63 adet, başakta tane sayısının 32.20-44.07 adet, başakta
tane ağırlığının 1.24-2.08 g, birim alan tane veriminin 644.50-857.58 kg/da ve bin tane
ağırlığının 33. 83- 49.53 g arasında değiştiğini vurgulamıştır.
Doxastakis vd (2002), Tritikalenin besin değeri üzerinde yapılan son 20 yıllık araştırma
sonuçlarına göre, tritikalenin besin değerinin buğdaya oranla daha yüksek olmasına
rağmen, kül içeriğinin daha yüksek, un randımanın daha düşük oluşu, kabarmasının
daha az olması ve ekmeğinin sert oluşu (fazla kabarmaması) gibi nedenlerden dolayı
ekmek yapımında yaygın olarak kullanılamadığını bildirmişlerdir. Ekmeklik buğdayla
karşılaştırıldığında, tritikalenin daha düşük gluten içerdiği, gluteninin daha elastiki
14

olduğu ve bu nedenle kabarmasının buğday ekmeğine oranla daha az olduğu araştırıcılar
tarafından vurgulanmıştır. Tritikalenin ekmek yapımında kullanımının, daha çok
ekmeklik buğdayla karıştırılma şeklinde olduğu ve bu şekilde kullanıldığında daha
geniş bir kullanım alanına sahip olacaktır. Nitekim yapılan bir çalışmada buğday ununa
% 5 ve % 10 oranında tritikale ununun katılması ile ağırlık, hacim, yapı ve görünüş
olarak kabul edilebilir nitelikte ekmek yapılabildiği bildirilmektedir.
Rodriges vd (2002), yoğun tarımsal girdi, özelikle de fazla azotlu gübre kullanımının
olduğu Batı Avrupa ülkelerinde Endüstri bitkilerinin yetiştirilmesinden sonra toprakta
kalan fazla azotun yıkanarak çevreye ve yer altı sularına zarar vermesini önlemek
amacıyla tritikalenin azot tutucu bitki olarak kullanıldığını bildirmektedirler.
Araştırıcılar, tritikalenin toprakta bulunan fazla miktardaki azotu bünyesinde tutarak ve
mineralizasyonuna neden olarak kirletici etkisini ortadan kaldırdığını belirtmrktrdirler.
Çiftçi vd (2003), yaptıkları araştırmada; tritikale de ham protein mitarını 135 g/kg, kül
miktarını 20.8 g/kg lif içeriğini 34.6 g/kg olduğunu bildirmektedirler. Aynı araştırıcılar,
mısır da 84 g/kg protein içeriği, 11.6 g/kg kül miktarı, 26.5 g/kg lif içeriği ve buğday da
protein içeriğini 120.4 g/kg, kül miktarını 17.1 g/kg ve lif içeriğinin 33.2 g/kg olarak
bildirmişlerdir. Bu yönüyle kanatlı hayvanların beslenmesinde tritikalenin buğday ve
mısıra alternatif tane yemi olarak kullanılabileceğini bildirmişlerdir.
Perez vd (2003), bazı tritikale çeşitlerinden elde edilen unların özellikle kraker yapımına
son derece uygun olduğu bildirilmektedirler. Araştırıcılar, tritikalenin bu özelliği ile
özellikle beyaz teneli yumuşak buğdayların yetişme olanağının olmadığı ülkelerde
kraker yapımında kullanılabilecek alternatif bir tahıl olarak kullanılabileceğini
vurgulamışlardır.
15

2.2. DNA İzolasyonu ve RAPD- PCR Tekniği
PCR, seçilmiş bir DNA parçasının çok sayıda kopyasını yapan enzimatik bir yöntem
olup, bu çoğaltma işleminde, iki oligonükleotid, ısıya dayanıklı DNA polimeraz, 4 tane
deoksiribonükleosid trifosfat, kalıp DNA ve buffer kullanılmaktadır. PCR’ın işleyişi şu
şekildedir; Çift zincirli kalıp DNA ısıtılınca, komplementer (tamamlayan) zincirleri
tutan iyonik bağlar zayıflamakta ve DNA tek zincirli duruma geçmeye başlamaktadır.
Bu olaya denatürasyon denemekte, genelikle denatürasyon için 94 °C sıcaklık
uygulanmaktadır. Denatürasyonun ardından ortam yavaş yavaş soğutulmakta ve bu
esnada oligonükleotidler tek zincirli DNA’daki komplementeri (tamamlayan) olan
bölgeye bağlanmaktadır. DNA sentezi bu noktada başlamaktadır. Primer yapışması
(annealing) denilen bu aşamada, primer tek zincirli DNA’nın 3’ucundan
bağlanmaktadır. Oligonükleotidlere primer adı verilmektedir. Çünkü DNA sentezini
primerler hazırlamaktadır. Daha sonra ortamın ısısı yükseltilmekte, bu aşamada enzim,
hedef DNA parçasının sentezini yapmaktadır. Bu safhaya, uzama (extension)
denmektedir. DNA polimeraz senteze, çift zincirin 3’ucundan başlamakta ve yeni DNA
5’-3’ yönünde sentezlenmektedir. Taq polimeraz 72 °C sıcaklıkta iyi çalıştığından, bu
sıcaklık derecesi de uzama için seçilmektedir. Isıtma ve soğutma işlemleri pek çok kez
tekrarlanarak özel bölgenin çok sayıda kopyası elde edilebilmektedir (Maniatis vd 1982,
Yıldırım ve Kandemir 2001).
RAPD-PCR motodu PCR’a dayalı önemli metodlardan birisi olup, tesadüfi bir dizilişe
sahip 10 bazlık primerleri kullanarak in vitro koşullarda enzimatik yollarla DNA’nın
çoğaltılması yöntemine dayanır. RAPD tekniği bir PCR tekniği olmakla birlikte klasik
PCR’dan ayrı olarak tek bir primer kullanmakta ve bu primer PCR primerlerine göre
daha az sayıda nükleotid (baz) içermektedir (Williams vd 1990).
Williams vd (1990), rastgele çoğaltılan DNA farklılığı yönteminin (RAPD markers),
PCR ile uygun primer ve genomik DNA kullanılması yolu ile istenilen DNA parçasının
kolayca amplifikasyonunun (çoğaltımının) yapılabileceğini bildirmiştir. Araştırıcılar,
RAPD-PCR tekniğinin DNA’nin in vitro şartlarında çoğaltılmasına dayalı bir teknik
olduğunu ve son yıllarda bitki tür ve çeşitlerinin moleküler düzeyde tanımlanmasında
16

önemli bir rol oynadığını bildirmişlerdir. Bu teknik kullanılarak son yıllarda türlerin ve
çeşitlerin moleküler düzeyde tanımlanmasında önemli mesafeler alınmıştır.
Devos ve Gale (1992), buğdayda genetik markör sistemi olarak RAPD markörlerinin
kullanımını değerlendirmişlerdir. RAPD yöntemine DNA, Mg, Taq polimeraz
konsantrasyonu ve denatürasyon sıcaklığının etkili olduğunu bildirmişlerdir. RAPD
ürünlerinin dominant davranışının, buğdayda linkage (bağlılık) haritalarının
oluşturulması için genetik markör olarak kullanımını sınırladığını, bununla birlikte
genotiplerin analizinde yararlanılabileceğini açıklamışlardır. Araştırıcılar, RAPD
tekniğinin buğdayda kullanılmasında ilk önceleri az düzeydeki çoğalma ve düşük
polimorfizmden dolayı sorunlarla karşılaştıklarını bildirmişlerdir. Bu sorunun buğdayın
geniş genomik yapıya ve yüksek orandaki tekrarlanan DNA karakteristliğine sahip
olmasından kaynaklandığı vurgulanmıştır. Bu sorunu aşmak amacıyla bir çok yaklaşım
denenmiş ve RAPD tekniğinin diğer bazı tekniklerle (Gradient gel- elektrophosis ve
DNA nın restriction enzimleri ile ön ayrıştırılaması gibi) birleştirilmesi sonucu bu sorun
ortadan kaldırılmıştır.
Skroch vd (1992), iki genotipin RAPD markörleri ile karşılaştırılmasının, her genotip
için RAPD markörlerinin var olduğu (1) veya olmadığına (0) göre yapıldığını ve markör
amplifikasyonlarının varlığı veya yokluğu için genotip karşılaştırması yapılırken
sonuçların karşılaştırma türüne göre değişebildiğini bildirmişlerdir. Her iki gotipte de
RAPD markörlerinin olması, bu lokusta yüksek düzeyde sekans (baz dizilişi)
benzerliğinin olduğuna işaret etmektedir. Bir genotipte markör olup ta diğerinde
olamaması durumunda iki genotipte sekans (baz dizilişi) farkı olduğu kesindir. Üçüncü
bir olasılık her iki genotipte de amplifikasyon olmamasıdır. Bu sonuç sekans benzerliği
hakkında bir şey söylemez. Amplifikasyon yokluğu gösteren genotip için rastgele
lokuslarda yüksek oranda sekans bilgisi vardır. Böylece amplifikasyon yokluğu (0,0)
karşılaştırmaları benzerlik kanıtıdır ve (1,1) karşılaştırmalarına neredeyse eşdeğer bilgi
sağlar.
Weeden vd (1992), farklı primer konsantrasyonlarının değişik sayıda bant oluşumuna
neden olduğunu, düşük primer konsantrasyonu, büyük fragmentleri (1500-3000 bp)
17

çoğaltma eğiliminde iken, primer konsantrasyonunun artması ile bu fragmentler
kaybolmakta ve yerlerini daha küçük (200-400 bp) fragmentlere bıraktığını
vurgulamaktadırlar. Bununla birlikte diğer araştırıcılar dikkatli optimize edilmiş
koşullar ve PCR’ ın sıkı bir şekilde kontrol edilmesiyle RAPD’ in buğdayda
kullanılması olanağının olduğunu bildirmektedirler (Joshi ve Nguyen 1993). Bundan
sonraki bir çok çalışmada RAPD markörleri kullanılarak diploid (Joshi ve Nguyen
1993), hekzaploid (He vd 1992) buğday türlerindeki genetik farklılık, buğdayda
hastalıklara dayanıklı genlerin işaretlenmesi (Fahima vd 1995, Sun vd 1997), buğday ve
tritikalede böceklere dayanıklılık genlerinin işaretlenmesi (Dweiket vd 1994, Fritz vd
1999), tritikalede depo proteinlerinin belirlenmesi (Igrejas vd 1999) gibi çalışmalar
başarıyla sonuçlanmıştır.
Smith vd (1994), bazı buğday ve tritikale çeşitlerinde glutenin allellerini
belirleyebilmek amacıyla PCR DNA markörlerini kullanmışlardır. Araştırıcılar,
optimum PCR koşulları için 100 µl’lik hacimde 1mM MgCl2, 0.4 mM dNTP’s, 0.1 mM
primer, 800 ng genomik DNA ve 4 U Tag Polimeraz kullanmışlardır.
Pammi vd (1994), sorgumda tekrarlanabilir RAPD markörlerinin amplifikasyonu için
primer, DNA miktarı, Taq polimeraz, MgCl2 ve yapışma (annealing) sıcaklığını test
etmişlerdir. Araştırıcılar primerin artan veya azalan konsantrasyonlarının amplifiye olan
ürünlerin verimlerini azalttığını ve optimum 3-6 µM primer yoğunluğunun yeterli
olduğunu saptamışlardır. DNA’nın ise yüksek konsantrasyonları (20-50 ng/20 µl)
dışında amplifiye olmuş ürünlerin profili ve veriminde bir etkisi olmamıştır. Taq
polimerazın 15µ’ lik reaksiyon karışımında 0.38 U olarak kullanılmasıyla iyi sonuçların
alındığı tespit edilmiştir. Annealing (yapışma) sıcaklığının 36 °C de iken az
amplifikasyon belirlenirken, 48 °C de optimum sonuçlar almışlardır. Araştırıcılar
MgCl2’ ün düşük ve yüksek konsantrasyonlarının amplifiye olmuş DNA oluşumunu
azalttığını ve optimum 25 mM MgCl2’ın iyi sonuçlar verdiğini bulmuşlardır. En son
olarak PCR döngüleri analiz edilmiş ve 25–30 döngü arasında hızlı bir
amplifikasyonun, 30 ve 35 döngü arasında ise daha düşük amplifikasyon olduğunu
bildirmişlerdir.
18

Pancholi (1995), RAPD’in en büyük avantajının, hedef DNA dizilerinin önceden
bilinmesine gereksinim olmaması olduğunu, ancak kalıp DNA konsantrasyonunun
başarı için çok kritik bir faktör olduğunu bildirmiştir. Kalıp DNA’nın kalitesi amlifiye
olmuş ürünlerin çoğaltılması ve güvenirliğine etkili olup, güvenilir bir DNA
replikasyonu için Taq polimerazın önemli olduğunu ve magnezyuma bir divalent katyon
kaynağı olarak ihtiyaç duyduğunu bildirmiştir. Optimum Mg konsantrasyonunun her
PCR tekniği için ayarlanması gerektiğini, çok az Mg konsantrasyonunun amplifikasyon
verimini azaltırken çok yüksek miktardaki Mg konsantrasyonunun ise spesifik olmayan
bağlanmalara neden olduğunu vurgulamıştır.
Weising vd (1995), RAPD-PCR yönteminde DNA bantlarının üretilebilirliği ve
belirginliği, izole edilen genomik DNA’nin saflığına ve kırılıp kırılmamasına göre
değiştiğini vurgulamışlardır. Bu nedenle, DNA izolasyonunun çok önemli olduğu ve
DNA’nın temizliğini azaltacak ve degredasyonunu (bozulma) sağlayacak etmenlerin
ortadan kaldırılması gerektiği bildirilmiştir. Araştırıcılar, RAPD-PCR yönteminde
DAN’nın az miktarda parçalanmasının PCR ürünlerinin değişmesine ve DNA’daki
yüksek parçalanmanın ise amplifikasyonun gerçekleşememesine neden olacağını
belirtmişlerdir. PCR reaksiyonunun bir çok bileşeni içermesi ve PCR işleminin farklı
basamaklarda gerçekleşmesi nedeniyle RAPD tekniğinin optimizasyonunun zaman
alabileceği, PCR aletinin markası, primerler, yapışma sıcaklığı ve magnezyumun
konsantrasyonunun bant görüntülerine etki eden temel faktörler olduğu araştırıcılar
tarafından ayrıca vurgulanmıştır.
Penner (1996), RAPD markörlerinin taksonomik belirlemelerde, sistematik ilişkilerin
ortaya konmasında, populasyon genetiği çalışmalarında, türlerin hibridizasyonunda ve
akrabalık ilişkilerinin belirlenmesinde kullanılabileceğini belirtmiştir. Araştırıcı, RAPD-
PCR tekniğinin PCR’a dayalı olarak tesadüfi 10 bazlık primerleri kullanarak in vitro
şartlarda enzimatik yolla DNA’nın çoğaltılması esasına dayandığını bildirmektedir.
Ayrıca, PCR optimizasyon koşullarının önemli olduğu, reaksiyona giren maddelerin
konsantarsyonlarındaki değişikliklerin PCR ürünlerinin üretilebilirliğini önemli
derecede etkilendiği belirtilmiş, her bir reaksiyon için 2 U’den fazla Tag polimeraz ve
19

2 mM’ın üzerinde MgCl2 kullanımının istenmeyen bantların oluşumuna neden
olabileceği araştırıcı tarafından vurgulanmıştır.
Myburg vd (1997), ekmeklik buğdayda RAPD tekniği kullanarak yaptıkları
çalışmalarda düşük seviyede polimorfizm (farklılık) belirlendiğini, ancak buna rağmen,
son zamanlarda yapılan ekmeklik buğday, tritikale ve çavdar ve 5 Afrika kökenli çeşit
ile 5 Rusya kökenli ekmeklik buğday çeşidinde 29 RAPD markörü kullanılarak parmak
izi analizi yapıldığını bildirmektedirler. Bu çalışmalar sonucunda çeşide, türe ve
genoma özel makörler RAPD analiziyle belirlenmiştir.
De Bustos vd (1998), Arpa’da yaptıkları çalışmada, PCR reaksiyonu için 25 µl’lik mix
karışımında; uygun genomik DNA yoğunluğunu 12.5 ng ve primer yoğunluğunu 7.5 ng
olarak tespit etmişlerdir. Araştırıcılar, 2.5 µl buffer ve MgCl2, 100 mM dNTP’s ve 0.5 U
Tag polimeraz kullanarak RAPD yapmışlardır.
Li vd (1999), Israil de PCR RAPD tekniğini kullanarak 118 tescilli yabani emmer
buğdayı (Triticum dicoccoides) bireylerlerinde genetik çeşitliliği araştırmışlar ve DNA
polimorfizmi bulmuşlardır. Araştırıcılar bu DNA polimorfizminin mikro klimatik
stresten kaynaklandığını belirtmişlerdir.
Brunel vd (1999), Çavdarda bulunan 2R kromozomunun biotik ve abiotik streslere
dayanıklılık genlerini taşıdığını ve bunun buğday ıslahında kullanılabileceğini
bildirmektedirler. Bu genlerin ıslah amaçlı kullanılabilmesi için 2R kromozomu ile
buğdayda bulunan 2B kromozomu arasında translokasyon (yer değiştirme)
gerçekleştirebilmek amacıyla RAPD markörlerini bir araç olarak kullanmışlardır.
Merkezi translokasyonlu 2RS.2BL ve 2BS.2RL homozigot buğday hatları 489 primer
ile denenmiş ve 65 adet net çoğatılabilen polimorfizm gözlenmiştir. Bu markörlerin
buğday x çavdar melezlerinde translokaskonların kırılma noktalarının belirlenmesinde
kullanılabileceğini bildirmişlerdir.
20

Fritz vd (1999), tritikalede Rus buğday afidine dayanıklı hatları belirlemek için
yürüttükleri çalışmada, RAPD markörlerini kullanmışlar ve dayanıklı hatları
belirlemişlerdir.
Fahima vd (1999), Israil’de yaptıkları çalışmada RAPD DNA’lardaki genetik çeşitliği
110 adet tetraploid buğdayların genitörü olan yabani buğdaylarda (Triticum diccoides )
test etmişler ve İsrail’den toplanan buğdaylardaki yüksek oranda genetik çeşitliliği
RAPD markörleri ile ortaya koymuşlardır. Kullanılan 10 primerde 59 tane polimorfik
locus ve 11 tane monomorfik locus amplifiye (çoğalma) olmuştur. Araştırıcılar RAPD
analizinin Türkiye ve İsrail’in değişik coğrafik bölgelerinden toplanan Triticum
dicoccoides genotiplerinin ayrımasında oldukça etkili olduğu sonucuna varmışlardır.
Araştırıcılar ayrıca, yabani T. dicoccoides formlarının genetik farklılığının tahmin
edilmesinde ve melezleme için uygun anaçların belirlenmesinde, yabani formlardan
alınacak önemli karakterlerin saptanmasında kullanılacak haritalamaya yarayışlı uygun
populasyonların belirlenmesinde RAPD markörlerinin gerekli olduğu sonucuna
varmışlardır.
Sivolap vd (1999), ekmeklik buğday çeşitlerindeki polimorfizmi ortaya koymak için
yaptıkları çalışmada, RAPD ve SSR belirleyici sistemlerini kullanmışlar, RAPD’in çeşit
ayrımında en güçlü belirleyici sistemlerinden birisi olduğunu, RAPD belirleyicilerine
dayalı dendogramların büyük oranda pedigri verilerini doğruladığını ve sonuç olarak,
iki belirleyici sisteminin buğday genetik ve ıslah çalışmalarında birlikte kullanılmasını
önermişlerdir.
Şençiçek (2000), Susam da yaptığı çalışmada, PCR reaksiyonu için genomik DNA
yoğunluğunu 2, 3.5, 7 ve 13 ng / 25 µl olarak test etmiş ve uygun DNA yoğunluğunu 7
ng /25 µl olarak belirlemiştir. Yapılan çalışmada diğer PCR bileşenleri ise 25 µl’lik
karışımda; 3mM MgCl2, 25 pmol primer, 200 µM dNTP, 1 U Tag DNA, 2.5 µl buffer
olarak bulunmuştur.
Freitas vd (2000), RAPD belirleyicileriyle 14 Brezilya ekmeklik buğday çeşidindeki
genetik varyasyonu belirlemek amacıyla 10 baz dizinlik rasgele 50 primeri taramışlar;
21

48 primerden toplam 256 tekrarlanabilir DNA amplifikasyon ürünü elde etmişler, bunun
% 33’ünün ise polimorfik olduğunu, yapılan genetik yakınlık hesapları ve
dendogramlara göre düşük farklılıklara karşın (ortalama genetik yakınlık % 27) atalarını
yansıtan iki grup çeşidin tanımlandığını belirtmişlerdir.
Szucs vd (2000), farklı kökenlere sahip 23 makarnalık buğday çeşidinde genetik
çeşitliliği belirlemek amacıyla RAPD ve RFLP belirleyici sistemlerini kullandıklarını,
kullanılan 47 RFLP probe endlonukleaz kombinasyonun 13’ünde polimorfizm oranı %
28 iken, 16 RAPD primerinin 8’inde % 50 olduğunu, benzerlik değerlerinin RFLP için
0.04 ile 0.14 arasında iken, RAPD için 0.03 ile 0.18 arasında bulunduğunu, bu
çalışmayla makarnalık çeşitlerin oransal olarak genetik tabanlarının dar da olsa
polimorfizmi başarıyla ortaya konulduğunu ve bu sonuçların ıslah programlarının
düzenlenmesi ve farklı agronomik özelliklerle bağlantılı olan DNA belirleyicilerinin
haritalanması için gerekli makarnalık buğday melezlerinin yapılmasında
kullanılabileceğini bildirmişlerdir.
Malgorzata vd (2002), tritikale ve buğday çeşitlerinde genetik çeşitliliği araştıdıkları
çalışmalarında; 20 µl’lik reaksiyon karışımında 20 ng genomik DNA, 1.0-1.2 µM
primer, 200 µM MgCl2, 1.0 U Tag polimeraz ve yeterli miktarda buffer kullanmışlardır.
Araştırıcılar, RAPD primerleri ile düşük seviyede polimorfizm elde etmişlerdir.
Amiour vd (2002), Avrupa’da yetiştirilen tritikale çeşitlerinin depo proteinleri üzerine
yaptıkları bir çalışmada; PCR reaksiyonu için 20 µl’lik reaksiyon karışımında 80 ng
genomik DNA, 200 µM her bir deoxynucleotide, 0.4 U Tag-DNA polymerase, 0.5 µM
primer ve 1.5 mM MgCl2 içeren 1X aplifikasyon tamponu kullanmışlardır. Araştırıcılar
PCR reaksiyonu için, 94 °C’de 5 dakika ön uzatma, 94 °C’de 30 sn 30 çevrim ve 72
°C’de 5 dakika son uzatma süreleri ile primerlere göre değişen sıcaklıklarda Tm
ısılarını kullanmışlardır. Araştırıcılar tritikale çeşitlerinin depo proteinleri yönünden
yüksek oranda polimorfizm gösterdiklerini vurgulamışlardır.
Sun vd (2003), yaptıkları çalışmada, RAPD markörlerini, Fusarium (solgunluk) a
dayanıklılık yönünden çeşitlilik gösteren 35 yazlık buğday çeşidi ve hattında genetik
22

farklılığı ortaya koymak amacıyla kullanmışlardır. Araştırıcılar, RAPD markörleri ile
hatlar arasındaki ve hastalık ırkları arasındaki genetik benzerlikleri ortaya koymaya
çalışmışlardır. Fusariuma dayanıklı ve dayanıksız hatların DNA bantları kullanılarak
primerler test edilmiş ve denenen 160 primerden 17 tanesi polimorfik ve yeniden elde
edilebilen bantlar verdiğini bildirilmişlerdir. RAPD verilerine göre hesaplanan genetik
benzerlik 0.64–0.98 arasında değişme göstermiştir. RAPD mörkörleri ve Fusairuma
dayanıklılık indeksi hesaplaması sonucunda 3 adet RAPD mörkörü belirlenmiştir.
Belirlenen bu 3 markörün Fusairuma dayanıklı hatlara özel olduğu bildirilmektedir.
Kuleung vd (2003), DNA’ ya dayalı moleküler markörlerin; genomik haritaların
çıkartılmasında, DNA parmakizi analizlerinin yapılmasında, populasyon yapısının
belirlenmesinde ve genetik çeşitliliğin ortaya konmasında oldukça güçlü bir araç
olduğunu bildirmektedirler. Araştırıcılar yaptıkları bir çalışmada kullandıkları 176 adet
SSR marköründen 96 tanesinin (85 buğday ve 11 çavdar markörünün) tritikalede
amplifikasyon gösterdiğini ortaya koymuşlardır. Buğday markörleri, buğday, çavdar ve
tritikalede sırası ile ortalama 2.6, 2.7 ve 2.4 adet polimorfik DNA bandı vermiştir.
Tritikalede aplifiye olmuş markörlerin baz dizilişleri ve büyüklükleri verici türlerle
(çavdar ve buğday) benzer olması, bu markörün aynı locustan geldiğini, ve bu allelik
primer bağlanma bölgelerinin melezleme sırasında korunduğu sonucunu ortaya
koymaktadır.
Tams vd (2004), şu ana kadar yapılan çalışmalarda tritikale genotiplerinde çoğrafik
orijin olarak genetik çeşitliliği gruplamanın mümkün olmadığını, ABD, Kanada ve
Meksika’dan toplanan tritikale genotiplerinin tam ve değişmiş, Avrupa’dan toplanan
genotiplerin ise yazlık ve kışlık şeklinde gruplandırıldığını bildirmişlerdir. Araştırıcılar,
tür içerisindeki genetik çeşitlilik hakkında bilgi sahip olmanın hat ve hibrit ıslah
programlarında kullanılacak ana ve babanın seçiminde önemli olduğunu vurgulamışlar
ve buğday ve çavdar SSR markörlerini kullanarak Avrupa’da yetiştirilen 128 adet
tritikale genotipi arasında genetik çeşitliği araştırmışlar; ortalama polimorfizm oranının
0.54, ortalama genetik uzaklığın ise 0.33 olduğunu belirlemişlerdir.
23

4. ARAŞTIRMA BULGULARI VE TARTIŞMA
4.1. Tarla Deneme Bulguları ve Tartışma
Bu araştırma; 2000 ve 2001 yıllarında bazı tritikale hatlarının, çiçeklenme tarihi, bitki
boyu, bitkide kardeş sayısı, başak uzunluğu, başakçık sayısı, başakta tane sayısı, başakta
tane ağırlığı, bin tane ağırlığı, hasat indeksi, birim alan tane verimi ve protein oranı
üzerine etkilerini belirlemek amacıyla 2 yıl süreyle yürütülmüştür. Ele alınan özelliklere
ilişkin veriler ve bu verilerin değerlendirilmesi yıllara göre ayrı ayrı başlıklar halinde
açıklanmıştır (yıllar birleştirilerek yapılan analizinde yıllar önemli çıkmıştır).
4.1.1. Çiçeklenme tarihi (1. Yıl)
Bazı tritikale hatlarında çiçeklenme tarihine ilişkin verilerle yapılan varyans analiz
sonuçları Çizelge 4.1.’de verilmiştir.
Çizelge 4.1. Farklı tritikale hatlarında çiçeklenme tarihlerine ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 1550.990 Bloklar 3 29.070 9.690 Hatlar 24 1355.740 56.489 ** Hata 72 166.180 2.308 C.V. 4.90
**) 0.01 düzeyinde önemli
Çizelge 4.1.’de görüldüğü gibi, çiçeklenme tarihi yönünden tritikale hatları arasındaki
farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.2.’de gösterilmiştir.b
Çizelge 4.2.’de de görüldüğü gibi, tritikale hatları arasında çiçeklenme tarihi yönünden
en yüksek değer 39.50 gün ile LAD 490 tritikale hattından elde edilmiş, LAD 490,
LAD388 ve CWT hatları arasında istatistiki yönden önemli bir farklılık gözlenmemiş
olup, aynı grupta yer almışlardır. En düşük değer ise 25.75 gün ile ZF 12 tritikale
hattında saptanmıştır. Bu verilere göre LAD 490, LAD 388 ve CWT tritikale hatlarının
geççi, ZF 12, ZF 13, ZF 11, ZF 10, ZF 15, ZF 14 ve ZF 9 tritikale hatlarının ise erkenci
hatlar olduğu söylenebilir.
37

Çizelge 4.2. Farklı tritikale hatlarında çiçeklenme tarihi ortalamaları (1 Mayıstan sonra geçen gün sayısı)
Hatlar Ortalama LAD 490 39.50 a 1* LAD 388 39.00 a 1 CWT 38.00 a 12 BDMT 35.75 b 23 LO 51 33.00 c 34 JGS 32.50 cd 45 TATLICAK-97 32.25 cde 4-6 MT 1 31.75 c-f 4-7 ZF 3 31.50 c-g 4-7 ZF 6 31.50 c-g 4-7 GTAD 31.25 c-g 4-8 ZF 5 31.00 c-g 4-9 ZF 4 30.75 d-h 4-10 ZF 1 30.25 d-ı 4-11 ZF 2 29.75 e-k 4-12 ZF 7 29.50 f-k 5-12 ZF 8 29.00 g-l 6-13 ZF 16 28.50 h-l 7-13 ZF 9 28.00 i-m 8-13 ZF 14 27.75 j-m 9-13 ZF 15 27.50 j-m 10-13 ZF 10 27.25 klm 11-13 ZF 11 27.00 lm 11-13 ZF 13 26.75 lm 12-13 ZF 12 25.75 m 13
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir 4.1.2. Çiçeklenme tarihi (2. Yıl) Bazı tritikale hatlarında çiçeklenme tarihine ilişkin verilerle yapılan varyans analiz sonuçları Çizelge 4.3.’de verilmiştir. Çizelge 4.3. Farklı tritikale hatlarında çiçeklenme tarihlerine ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 389.040 Bloklar 3 10.160 3.387 Hatlar 24 300.040 12.502 ** Hata 72 78.840 1.095 C.V. 3.42
**) 0.01 düzeyinde önemli Çizelge 4.3.’de görüldüğü gibi, çiçeklenme tarihi yönünden tritikale hatları arasındaki
farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.4.’de özetlenmiştir.
38

Çizelge 4.4. Farklı tritikale hatlarında çiçeklenme tarihi ortalamaları (1 Mayıstan sonraki geçen gün sayısı)
Hatlar Ortalama LO 51 33.50 a 1* GTAD 33.25 a 12 BDMT 32.75 abc 1-3 LAD 388 32.75 abc 1-3 MT 1 32.50 a-d 1-3 CWT 32.00 a-d 1-3 JGS 31.75 a-e 1-4 LAD 490 31.75 b-e 1-5 ZF 3 31.50 b-e 1-5 ZF 6 31.25 c-f 1-5 ZF 5 31.00 c-f 1-6 TATLICAK-97 30.75 d-g 2-7 ZF 4 30.75 e-h 3-8 ZF 2 30.00 e-h 3-9 ZF 1 30.00 f-ı 4-9 ZF 7 29.50 g-j 4-9 ZF 11 29.25 h-k 5-9 ZF 16 29.00 ijk 6-9 ZF 14 29.00 ijk 7-9 ZF 8 28.75 ijk 7-9 ZF 10 28.75 ijk 7-9 ZF 9 28.50 ijk 8-9 ZF 12 28.25 jk 9 ZF 13 28.00 jk 9 ZF 15 27.75 k 9
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir Çizelge 4.4.’de görüldüğü gibi çiçeklenme tarihi yönünden 2. yılda da önemli
farklılıklar saptanmıştır. En yüksek çiçeklenme tarihi 33.50 gün ile LO 51 tritikale
hattında elde edilmiş, GTAD, BDMT, LAD 388, MT 1, CWT ve JGS hatları da
çiçeklenme tarihi yönünden LO 51 çeşidi ile aynı grupta yer almıştır. En düşük
çiçeklenme tarihi ise 27.75 gün ile ZF 15 tritikale hattında belirlenmiştir; çiçeklenme
tarihi yönünden ZF 15, ZF 13, ZF 12, ZF 9, ZF 10, ZF 8, ZF 14, ZF16 ve ZF 11
tritikale hatları arasında istatistiki yönden bir fark görülmemiştir.
Genel olarak araştırmanın ilk yılında erkenci ve geççi olarak belirlenen hatlar ikinci
yılda da çok az bir değişiklik olsa bile aynı grupta yer almışlardır. Bu farklılıklar
araştırmanın ikinci yılındaki iklim faktörlerinden kaynaklandığı söylenebilir. Farklı
tritikale hatlarında birinci yıl, ikinci yıl ve ortalama çiçeklenme tarihleri Çizelge 4.5.’te
verilmiştir.
39

Çizelge 4.5. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama çiçeklenme tarihleri (1 mayıstan sonra geçen gün sayısı)
Hatlar 1. Yıl 2.Yıl Ortalama TATLICAK-97 32.25 31.00 31.63 ZF 1 30.25 30.00 30.13 ZF 2 29.75 30.75 30.25 ZF 3 31.50 31.75 31.63 ZF 4 30.75 30.75 30.75 ZF 5 31.00 31.25 31.13 ZF 6 31.50 31.50 31.50 ZF 7 29.50 30.00 29.75 ZF 8 29.00 29.00 29.00 ZF 9 28.00 28.50 28.25 ZF 10 27.25 28.75 28.00 ZF 11 27.00 29.50 28.25 ZF 12 25.75 28.25 27.00 ZF 13 26.75 28.00 27.37 ZF 14 27.75 29.00 28.37 ZF 15 27.50 27.75 27.62 ZF 16 28.50 29.25 28.87 LAD 490 39.50 31.75 35.62 LAD 388 39.00 32.75 35.87 JGS 32.50 32.00 32.25 BDMT 35.75 32.75 34.25 CWT 38.00 32.50 35.25 GTAD 31.25 33.25 32.25 MT 1 31.75 32.50 32.12 LO 51 33.00 33.50 33.25
Çizelgede görüldüğü gibi iki yılın ortalaması olarak en erkenci tritikale hattı ZF 12
olup, bunu ZF 13, ZF 15 ve ZF 10 tritikale hatları izlemiştir. En geççi tritikale hattı ise
LAD 388 olup, bunu sırasıyla LAD 490, CWT, BDMT ve LO 51 tritikale hatları
izlemiştir.
4.1.3. Bitki boyu (1. Yıl) Bazı tritikale hatlarında bitki boyuna ilişkin verilerle yapılan varyans analiz sonuçları
Çizelge 4.6.’da verilmiştir. Çizelge 4.6.’da görüldüğü gibi, bitki boyu yönünden tritikale
hatları arasındaki farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların
önem düzeyini belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.7.’de
gösterilmiştir.
40

Çizelge 4.6. Farklı tritikale hatlarında bitki boyuna ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 6564.561 Bloklar 3 473.343 157.781 Hatlar 24 4909.824 204.576 ** Hata 72 1181.395 16.408 C.V. 3.35
**) 0.01 düzeyinde önemli Çizelge 4.7. Farklı tritikale hatlarında bitki boyu ortalamaları (cm)
Hatlar Ortalama ZF 3 148.9 a 1* ZF 1 126.1 b 2 ZF 15 125.8 b 2 ZF 4 124.3 bc 23 ZF 6 123.6 bc 23 JGS 123.3 bc 23 ZF 2 123.1 bc 23 ZF 12 123.0 bc 23 ZF 7 122.2 bcd 2-4 ZF 13 122.1 bcd 2-4 TATLICAK-97 121.9 bcd 2-4 ZF 8 121.6 bcd 2-5 CWT 120.8 bcd 2-5 BDMT 119.4 b-e 2-6 ZF 7 119.2 b-e 2-6 LAD 490 118.6 cde 2-6 ZF 16 118.4 cde 2-6 LO 51 118.1 cde 2-6 ZF 14 117.8 c-f 2-6 ZF 9 115.9 def 3-6 GTAD 115.8 def 3-6 ZF 10 113.8 ef 4-6 MT 1 113.4 ef 4-6 LAD 388 112.7 ef 56 ZF 11 111.3 f 6
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir. Çizelge 4.7.’de görüldüğü gibi bitki boyu yönünden önemli farklılıklar oluşmuş, bitki
boyu ortalamaları 111.3-148.9 cm arasında değişen değerler göstermiştir. En uzun bitki
boyu ortalaması 148.9 cm ile ZF 3 tritikale hattından, en kısa bitki boyu ortalaması
111.3 cm ile ZF 11 tritikale hattından elde edilmiştir. ZF 11, LAD 388, MT 1, ZF 10,
GTAD, ZF 9 ve ZF14 tritikale hatları arasında bitki boyu yönünden istatistiki olarak bir
fark görülmemiştir. Diğer tritikale hatları bitki boyu yönünden bu iki değer arasında yer
almıştır.
41

4.1.4. Bitki boyu (2.Yıl) Bazı tritikale hatlarında bitki boyuna ilişkin verilerle yapılan varyans analiz sonuçları
Çizelge 4.8.’de verilmiştir.
Çizelge 4.8. Farklı tritikale hatlarında bitki boyuna ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 6596.198 Bloklar 3 166.399 55.466 Hatlar 24 4821.820 200.909 ** Hata 72 1607.978 22.333 C.V. 4.18
**) 0.01 düzeyinde önemli Çizelge 4.8.’de görüldüğü gibi denemenin ikinci yılında da, bitki boyu yönünden tritikale
hatları arasındaki farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem
düzeyini belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.9.’da
gösterilmiştir.
Çizelge 4.9. Farklı tritikale hatlarında bitki boyu ortalamaları (cm)
Hatlar Ortalama ZF 3 139.3 a 1* ZF 4 121.8 b 2 ZF 1 120.1 bc 23 ZF 2 119.3 bcd 2-4 TATLICAK-97 116.9 b-e 2-5 ZF 6 116.9 b-e 2-5 GTAD 115.5 b-f 2-6 JGS 114.7 b-f 2-6 ZF 9 113.8 c-g 2-6 ZF 7 113.3 c-g 2-6 ZF 8 112.6 c-g 2-6 LAD 490 111.8 d-g 2-6 BDMT 111.6 d-g 2-6 ZF 12 110.7 efg 3-6 ZF 14 109.9 efg 3-6 ZF 13 109.0 efg 4-6 ZF 11 108.7 fg 56 CWT 108.1 fg 56 ZF 15 108.1 fg 56 ZF 16 108.0 fg 56 ZF 9 107.9 fg 56 ZF 10 107.7 fg 56 LAD 388 107.6 fg 56 LO 51 106.4 g 56 MT 1 105.8 g 6
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir
42

Çizelgede görüldüğü gibi 2. yılda da bitki boyu yönünden en uzun bitki boyu 139.3 cm
ile ZF 3 tritikale hattından, en kısa bitki boyu ise 105. 8 cm ile MT 1 tritikale hattından
elde edilmiştir.
Farklı tritikale hatlarından elde edilen birinci yıl, ikinci yıl ve ortalama bitki boyu
değerleri Çizelge 4.10’da özetlenmiştir. Çizelgede görüldüğü gibi bitki boyu yönünden
iki yılın ortalaması olarak tritikalede bitki boyu 109.6-144.1 cm arasında değişmekte
olup, en uzun bitki boyuna sahip tritikale hattı ZF 3, en kısa bitki boyuna sahip tritikale
hattı ise MT 1 olmuştur.
Çizelge 4.10. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama bitki boyu (cm)
Hatlar 1. Yıl 2.Yıl Ortalama TATLICAK-97 121.9 116.9 119.4 ZF 1 126.1 120.1 123.1 ZF 2 123.1 119.3 121.2 ZF 3 148.9 139.3 144.1 ZF 4 124.3 121.8 123.0 ZF 5 119.2 113.8 116.5 ZF 6 123.6 116.9 120.2 ZF 7 122.2 113.3 112.7 ZF 8 121.6 112.6 117.1 ZF 9 115.9 107.9 111.9 ZF 10 113.8 107.7 110.7 ZF 11 111.3 108.7 110.0 ZF 12 123.0 110.7 116.8 ZF 13 122.1 109.0 115.5 ZF 14 117.8 109.9 113.8 ZF 15 125.8 108.1 116.9 ZF 16 118.4 108.0 113.2 LAD 490 118.6 111.8 115.2 LAD 388 112.7 107.6 110.1 JGS 123.3 114.7 119.0 BDMT 119.4 111.6 115.5 CWT 120.8 108.1 114.4 GTAD 115.8 115.5 115.6 MT 1 113.4 105.8 109.6 LO 51 118.1 106.4 112.2
Tritikale hatlarına ilişkin bitki boyu ortalamalarında görüldüğü gibi; tritikale oldukça
uzun boylu bir serin iklim tahıl cinsidir. Bitki boyu, iklim ve toprak koşullarından
etkilenen bir özellik olmasının yanında genetik yapının da etkisi altında olan bir
karakterdir. Araştırmanın ikinci yılında tritikale hatlarına ait bitki boyu ortalamalarının
43

daha kısa oluşunun, araştırmanın ikinci yılında yağış miktarının daha az ortalama
sıcaklığın daha fazla olmasından kaynaklandığı söylenebilir.
Bitki boyuna ilişkin sonuçlarımız; Ankara koşullarında iki yıl süreyle denemeye alınan
18 tritikale hattı ve kontrolde bitki boyunu 130.3–123.9 cm olarak belirlediğini bildiren
Ünver (1999), Çukurova koşullarında bitki boyunu ortalamasını 122.5-126.9 cm olarak
saptayan Genç vd (1998), Çukurova koşullarında 3 yılın ortalaması olarak 108-126 cm
bitki boyu belirlediklerini bildiren Yağbasanlar vd (1989)’un, Şanlıurfa koşullarında
110.0-139.8 cm bitki boyu ortalaması elde eden Yağbasanlar vd (1990)’un, Bursa
koşullarında 109.20–125.25 cm bitki boyu ortalaması saptadığını bildiren Çöplü
(2001)’in ve Ankara Haymana koşullarında bitki boyu ortalamasını 88.9-128.41 cm
bulduğunu bildiren Çengel (2001)’in bulgularıyla uyum göstermektedir.
4.1.5. Fertil kardeş sayısı (1. Yıl) Bazı tritikale hatlarında fertil kardeş sayısına ilişkin verilerle yapılan varyans analiz
sonuçları Çizelge 4.11.’de verilmiştir.
Çizelge 4.11. Farklı tritikale hatlarında fertil kardeş sayısına ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 50.292 Bloklar 3 1.447 0.482 Hatlar 24 12.737 0.531 * Hata 72 36.108 0.502 C.V. 12.94
*) 0.05 düzeyinde önemli Çizelge 4.11.’de görüldüğü gibi, fertil kardeş sayısı yönünden tritikale hatları arasındaki
farklılıklar 0.05 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.12’de verilmiştir.
Çizelge 4.12.’de görüldüğü gibi, tritikale hatları arasında bitkide fertil kardeş sayısı
ortalamaları 6.22-4.90 adet/bitki arasında değişmiştir. En yüksek fertil kardeş sayısı ZF
1 (6.22) tritikale hattında belirlenirken, en düşük fertil kardeş sayısı ZF 10 (4.90)
tritikale hattından elde edilmiştir.
44

Çizelge 4.12. Farklı tritikale hatlarında fertil kardeş sayısı ortalamaları (adet/bitki)
Hatlar Ortalama ZF 1 6.22 a* ZF 16 6.15 ab ZF 15 5.92 abc BDMT 5.80 abc ZF 6 5.77 abc ZF 12 5.70 abc LAD 490 5.65 abc ZF 8 5.65 abc ZF 4 5.62 abc TATLICAK-97 5.62 abc LAD 388 5.62 abc LO 51 5.50 abc ZF 11 5.47 abc ZF 13 5.45 abc GTAD 5.40 abc ZF 2 5.40 abc ZF 5 5.40 abc ZF 14 5.37 abc ZF 9 5.17 abc JGS 5.15 abc MT 1 5.02 abc ZF 3 5.00 abc CWT 4.95 abc ZF 7 4.90 c ZF 10 4.90 c
*) Harfler 0.05 düzeyinde farklı grupları göstermektedir
4.1.6. Fertil kardeş sayısı (2. Yıl)
Bazı tritikale hatlarında fertil kardeş sayısına ilişkin verilerle yapılan varyans analiz
sonuçları Çizelge 4.13.’de verilmiştir.
Çizelge 4.13. Farklı tritikale hatlarında fertil kardeş sayısına ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 28.684 Bloklar 3 1.869 0.623 Hatlar 24 11.384 0.474 ** Hata 72 15.431 0.214 C.V. 11.18
**) 0.01 düzeyinde önemli Çizelge 4.13.’de görüldüğü gibi, fertil kardeş sayısı yönünden tritikale hatları arasındaki
farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.14.’de gösterilmiştir.
45

Çizelge 4.14.’de görüldüğü gibi; fertil kardeş sayısı yönünden araştırmanın 2. yılında
farklı gruplar oluşmuştur. En yüksek fertil kardeş sayısı LO 51 tritikale hattında
saptanırken en düşük kardeş sayısı ise JGS tritikale hattında bulunmuştur.
Araştırmanın 2. yılında ortalama fertil kardeş sayısının 1. yıl ortalamalarına göre daha
düşük çıkması, denemenin 2. yılındaki iklim şartlarından kaynaklandığı söylenebilir.
Özellikle yıllık ortalama yağışın ikinci yılda daha az olması ve erken ilkbaharda görülen
yağış yetersizliği nedeniyle tritikale hatları daha az kardeşlenme eğilimi göstermişlerdir.
Çizelge 4.14. Farklı tritikale hatlarında fertil kardeş sayısı ortalamaları (adet/bitki)
Hatlar Ortalama LO 51 4.950 a 1* ZF 10 4.675 ab 12 ZF 15 4.625 abc 12 ZF 13 4.475 a-d 1-3 ZF 4 4.450 a-d 1-3 ZF 6 4.430 a-d 1-3 ZF 7 4.300 a-d 1-3 ZF 5 4.275 a-d 1-3 TATLICAK- 97 4.275 a-d 1-3 GTAD 4.275 a-d 1-3 MT 1 4.225 a-e 1-3 ZF 8 4.175 a-e 1-3 ZF 9 4.175 a-e 1-3 ZF 12 4.150 b-e 1-3 ZF 16 4.125 b-e 1-3 ZF 14 3.975 b-e 1-3 ZF 11 3.950 b-e 1-3 BDMT 3.950 b-e 1-3 CWT 3.850 cde 23 ZF 1 3.850 cde 23 ZF 2 3.775 de 23 LAD 388 3.775 de 23 ZF 3 3.375 de 23 LAD 490 3.675 de 23 JGS 3.475 e 3
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama fertil kardeş sayıları Çizelge
4.15’te verilmiştir. Çizelgede görüldüğü gibi fertil kardeş sayıları yönünden tritikale
hatları arasında önemli farklılıklar gözlenmezken en fazla kardeşlenme eğilimi gösteren
tritikale hattı ZF 15, en az kardeşlenme gösteren tritikale hatları ise JGS ve CWT’ dir.
46

Ortalama fertil kardeş sayısına ilişkin bulgularımız; iki yıl ortalaması olarak 2.77–3.95
adet/bitki fertil kardeş sayısı belirleyen Ünver (1999)’un bulgularıyla uyum
göstermektedir. Ancak, Ankara koşullarında ortalama fertil kardeş sayısını 2.10–2.63
adet arasında değiştiğini bildiren Çengel (2001)’in bulgularıyla uyum
göstermemektedir. Bu farklılıkların iklim ve toprak koşullarının farklı oluşu ve
yetiştirme tekniklerindeki farklılıktan kaynaklandığı söylenebilir.
Çizelge 4.15. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama fertil kardeş sayısı
Hatlar 1. Yıl 2.Yıl Ortalama TATLICAK-97 5.62 4.27 4.95 ZF 1 6.22 3.85 5.03 ZF 2 5.40 3.77 4.59 ZF 3 5.00 3.37 4.18 ZF 4 5.62 4.45 5.04 ZF 5 5.40 4.27 4.84 ZF 6 5.77 4.43 5.10 ZF 7 4.90 4.30 4.60 ZF 8 5.65 4.17 4.91 ZF 9 5.17 4.17 4.67 ZF 10 4.90 4.67 4.79 ZF 11 5.47 3.95 4.71 ZF 12 5.70 4.15 4.93 ZF 13 5.45 4.47 4.96 ZF 14 5.37 3.97 4.67 ZF 15 5.92 4.62 5.27 ZF 16 6.15 4.12 5.14 LAD 490 5.65 3.67 4.66 LAD 388 5.62 3.77 4.69 JGS 5.15 3.47 4.31 BDMT 5.80 3.95 4.88 CWT 4.95 3.85 4.40 GTAD 5.40 4.27 4.84 MT 1 5.02 4.22 4.62 LO 51 5.50 4.95 5.23
Kardeşlenme üzerine etkili faktörler arasında; ekim sıklığı, ekim zamanı, ekim derinliği
gibi yetiştirme tekniklerinin yanında iklim ve toprak özellikleri bulunmakla birlikte,
kullanılan çeşitlerin genetik özelliği de olabilir. Tritikale, arpa ve buğdaya göre daha az
kardeşlenme özelliği gösteren bir serin iklim tahıl cinsidir (Kün 1996).
47
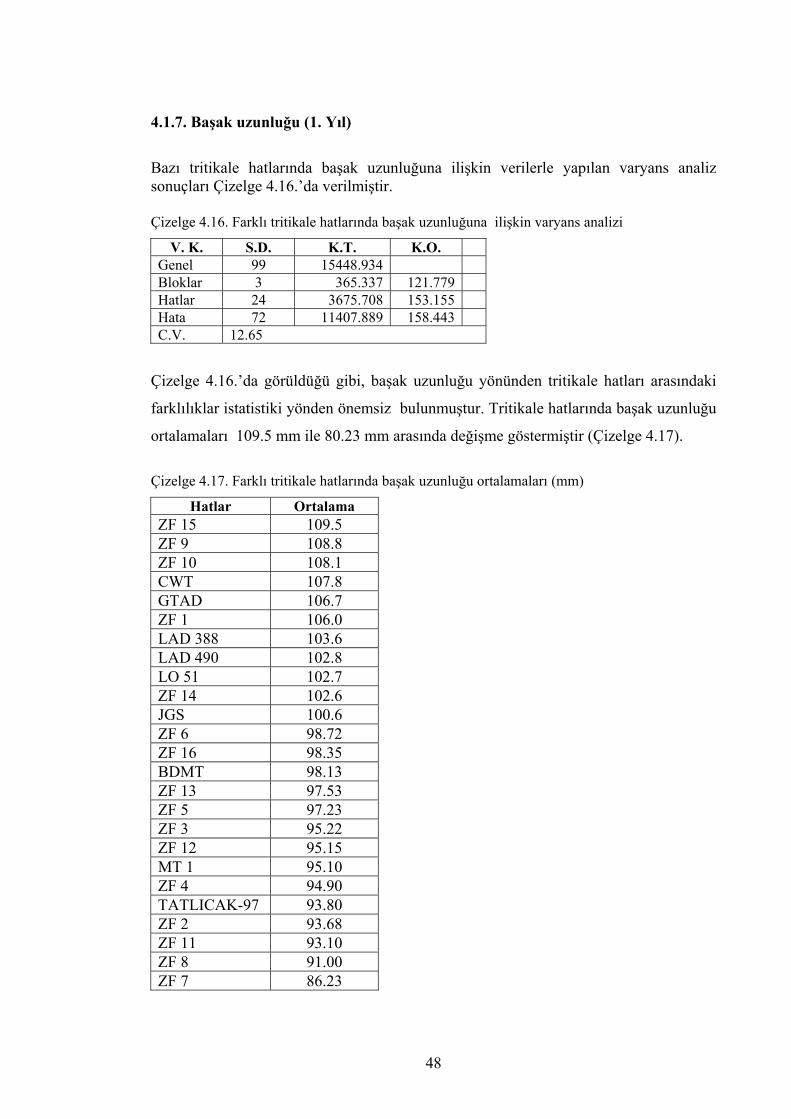
4.1.7. Başak uzunluğu (1. Yıl)
Bazı tritikale hatlarında başak uzunluğuna ilişkin verilerle yapılan varyans analiz sonuçları Çizelge 4.16.’da verilmiştir. Çizelge 4.16. Farklı tritikale hatlarında başak uzunluğuna ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 15448.934 Bloklar 3 365.337 121.779 Hatlar 24 3675.708 153.155 Hata 72 11407.889 158.443 C.V. 12.65
Çizelge 4.16.’da görüldüğü gibi, başak uzunluğu yönünden tritikale hatları arasındaki
farklılıklar istatistiki yönden önemsiz bulunmuştur. Tritikale hatlarında başak uzunluğu
ortalamaları 109.5 mm ile 80.23 mm arasında değişme göstermiştir (Çizelge 4.17).
Çizelge 4.17. Farklı tritikale hatlarında başak uzunluğu ortalamaları (mm)
Hatlar Ortalama ZF 15 109.5 ZF 9 108.8 ZF 10 108.1 CWT 107.8 GTAD 106.7 ZF 1 106.0 LAD 388 103.6 LAD 490 102.8 LO 51 102.7 ZF 14 102.6 JGS 100.6 ZF 6 98.72 ZF 16 98.35 BDMT 98.13 ZF 13 97.53 ZF 5 97.23 ZF 3 95.22 ZF 12 95.15 MT 1 95.10 ZF 4 94.90 TATLICAK-97 93.80 ZF 2 93.68 ZF 11 93.10 ZF 8 91.00 ZF 7 86.23
48
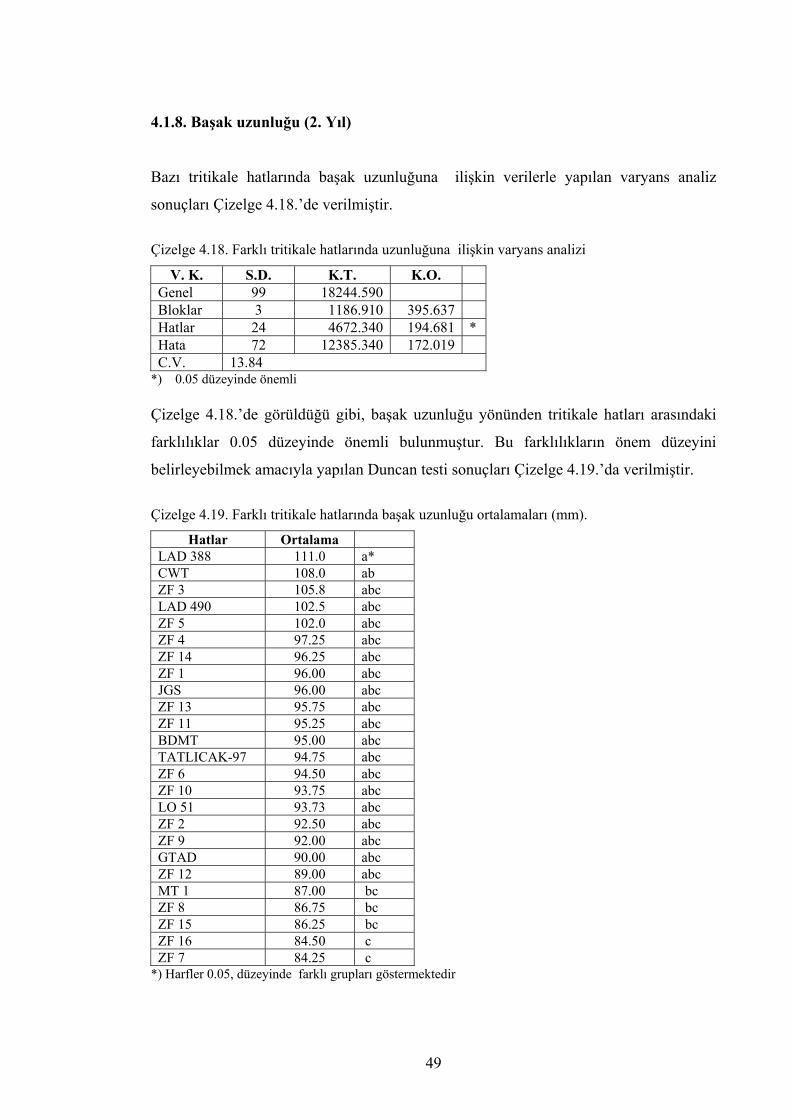
4.1.8. Başak uzunluğu (2. Yıl) Bazı tritikale hatlarında başak uzunluğuna ilişkin verilerle yapılan varyans analiz
sonuçları Çizelge 4.18.’de verilmiştir.
Çizelge 4.18. Farklı tritikale hatlarında uzunluğuna ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 18244.590 Bloklar 3 1186.910 395.637 Hatlar 24 4672.340 194.681 * Hata 72 12385.340 172.019 C.V. 13.84
*) 0.05 düzeyinde önemli Çizelge 4.18.’de görüldüğü gibi, başak uzunluğu yönünden tritikale hatları arasındaki
farklılıklar 0.05 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.19.’da verilmiştir.
Çizelge 4.19. Farklı tritikale hatlarında başak uzunluğu ortalamaları (mm).
Hatlar Ortalama LAD 388 111.0 a* CWT 108.0 ab ZF 3 105.8 abc LAD 490 102.5 abc ZF 5 102.0 abc ZF 4 97.25 abc ZF 14 96.25 abc ZF 1 96.00 abc JGS 96.00 abc ZF 13 95.75 abc ZF 11 95.25 abc BDMT 95.00 abc TATLICAK-97 94.75 abc ZF 6 94.50 abc ZF 10 93.75 abc LO 51 93.73 abc ZF 2 92.50 abc ZF 9 92.00 abc GTAD 90.00 abc ZF 12 89.00 abc MT 1 87.00 bc ZF 8 86.75 bc ZF 15 86.25 bc ZF 16 84.50 c ZF 7 84.25 c
*) Harfler 0.05, düzeyinde farklı grupları göstermektedir
49

Çizelge 4.19.’da görüldüğü gibi; araştırmanın 2. yılında ortalama başak uzunluğu 111.0
mm ile 84.25 mm arasında değişmiş olup, en yüksek başak uzunluğu 111.0 mm ile LAD
388 tritikale hattından, en düşük başak uzunluğu ise ZF 7 tritikale hattından elde
edilmiştir. Araştırmanın ikinci yılında başak uzunlukları tritikale hatlarında genelde
azalma göstermiştir.
Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama başak uzunlukları Çizelge
4.20’de verilmiştir.
Çizelge 4.20. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama başak uzunluğu (mm)
Hatlar 1. Yıl 2.Yıl OrtalamaTATLICAK-97 93.80 94.75 94.28ZF 1 106.0 96.00 101.00ZF 2 93.68 92.50 93.09ZF 3 95.22 105.8 100.51ZF 4 94.90 97.25 96.08ZF 5 97.23 102.0 99.62ZF 6 98.72 94.50 96.61ZF 7 86.23 84.25 85.24ZF 8 91.00 86.75 88.89ZF 9 108.80 92.00 100.40ZF 10 108.10 93.75 100.93ZF 11 93.10 95.25 94.18ZF 12 95.15 89.00 92.08ZF 13 97.53 95.75 96.64ZF 14 102.60 96.25 99.43ZF 15 109.50 86.25 97.88ZF 16 98.35 84.50 91.43LAD 490 102.80 102.5 102.65LAD 388 103.60 111.0 107.30JGS 100.60 96.00 98.30BDMT 98.13 95.00 95.57CWT 107.80 108.0 107.90GTAD 106.70 90.00 98.35MT 1 95.10 87.00 91.05LO 51 102.70 93.75 98.23
Çizelgede görüldüğü gibi en uzun başak uzunluğuna sahip tritikale hattı CWT en kısa
başak uzunluğuna sahip tritikale hattı ise ZF 7 dir. Diğer çeşit ve hatlar bu iki değer
arasında yer almıştır.
Tritikale hatlarında belirlenen başak uzunluğu ortalamalarına ilişkin sonuçlarımız;
Çukurova koşullarında tritikale çeşit ve hatlarında başak uzunluğunu 3 yılın ortalaması
50

olarak 8.4-13.2 cm arasında değiştiğini belirten Yağbasanlar (1989)’un, Ankara
koşullarında farklı tritikale ıslah hatlarında 2 yıl ortalaması olarak başak uzunluğunu
10.23-13.35 cm arasında belirleyen Ünver (1999)’un, Ankara, Haymana koşullarında
tritikale hatları arasında ortalama başak uzunluğunu 8.58-11.77 cm olarak belirleyen
Çelgel (2001) ve Bursa koşullarında tritikale hatlarında başak uzunluğu ortalamalarının
12.45-15.55 cm arasında değiştiğini bildiren Çöplü (2001)’in bulgularıyla benzerlik
göstermektedir.
4.1.9. Başakta başakçık sayısı (1. Yıl) Bazı tritikale hatlarında başakçık sayısına ilişkin verilerle yapılan varyans analiz
sonuçları Çizelge 4.21.’de verilmiştir.
Çizelge 4.21. Farklı tritikale hatlarında başakçık sayısına ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 477.689 Bloklar 3 11.322 3.774 Hatlar 24 225.129 9.380 ** Hata 72 241.239 3.351 C.V. 8.20
**) 0.01 düzeyinde önemli Çizelge 4.21.’de görüldüğü gibi, başakçık sayısı yönünden tritikale hatları arasındaki
farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.22,’de gösterilmiştir.
Çizelge 4.22’de özetlendiği gibi; tritikale hatları arasında ortalama başakta başakçık
sayısı 24.88-19.63 arasında değişmiştir. Başakta başakçık sayısı yönünden en yüksek
değer CWT (24.88 adet) tritikale hattından, en düşük değer ise ZF 12 (19.63 adet)
tritikale hattından elde edilmiştir.
51
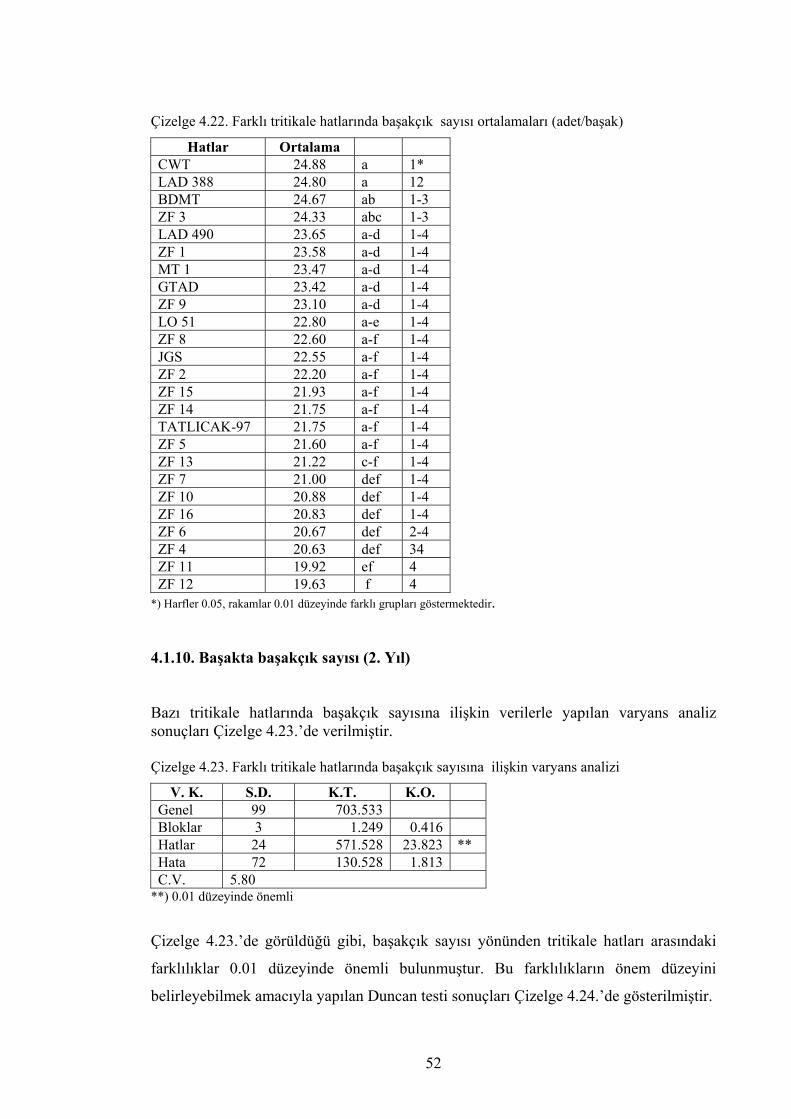
Çizelge 4.22. Farklı tritikale hatlarında başakçık sayısı ortalamaları (adet/başak)
Hatlar Ortalama CWT 24.88 a 1* LAD 388 24.80 a 12 BDMT 24.67 ab 1-3 ZF 3 24.33 abc 1-3 LAD 490 23.65 a-d 1-4 ZF 1 23.58 a-d 1-4 MT 1 23.47 a-d 1-4 GTAD 23.42 a-d 1-4 ZF 9 23.10 a-d 1-4 LO 51 22.80 a-e 1-4 ZF 8 22.60 a-f 1-4 JGS 22.55 a-f 1-4 ZF 2 22.20 a-f 1-4 ZF 15 21.93 a-f 1-4 ZF 14 21.75 a-f 1-4 TATLICAK-97 21.75 a-f 1-4 ZF 5 21.60 a-f 1-4 ZF 13 21.22 c-f 1-4 ZF 7 21.00 def 1-4 ZF 10 20.88 def 1-4 ZF 16 20.83 def 1-4 ZF 6 20.67 def 2-4 ZF 4 20.63 def 34 ZF 11 19.92 ef 4 ZF 12 19.63 f 4
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir. 4.1.10. Başakta başakçık sayısı (2. Yıl) Bazı tritikale hatlarında başakçık sayısına ilişkin verilerle yapılan varyans analiz sonuçları Çizelge 4.23.’de verilmiştir. Çizelge 4.23. Farklı tritikale hatlarında başakçık sayısına ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 703.533 Bloklar 3 1.249 0.416 Hatlar 24 571.528 23.823 ** Hata 72 130.528 1.813 C.V. 5.80
**) 0.01 düzeyinde önemli
Çizelge 4.23.’de görüldüğü gibi, başakçık sayısı yönünden tritikale hatları arasındaki
farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.24.’de gösterilmiştir.
52

Çizelge 4.24. Farklı tritikale hatlarında başakçık sayısı ortalamaları (adet/başak)
Hatlar Ortalama ZF 3 29.77 a 1* LAD 388 26.42 b 2 BDMT 25.35 bc 23 CWT 25.30 bc 23 TATLICAK-97 25.15 bc 234 ZF 5 24.98 bcd 234 LAD 490 24.83 bcd 2-5 LO 51 24.20 cde 2-6 ZF 2 24.15 c-f 2-6 ZF 6 24.13 c-f 2-6 JGS 23.77 c-g 2-6 ZF 4 23.73 c-g 2-7 MT 1 23.42 c-h 3-7 ZF 1 23.23 c-h 3-7 GTAD 22.75 d-ı 3-8 ZF 8 22.55 e-ı 3-8 ZF 14 22.33 e-j 4-8 ZF 11 21.92 f-k 5-9 ZF 7 21.73 g-k 6-10 ZF 13 21.40 h-k 6-10 ZF 9 20.88 i-l 7-10 ZF 16 20.25 jkl 8-10 ZF 10 20.00 kl 8-10 ZF 12 19.20 l 910 ZF 15 18.90 l 10
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir Çizelge 4.24’de özetlendiği gibi araştırmanın ikinci yılında tritikale hatları arasında
ortalama başakçık sayısı 29.77-18.90 adet arasında değişmiş olup, en yüksek değer ZF 3
tritikale hattından ve en düşük değer ise ZF 15 tritikale hattından elde edilmiştir.
Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama başakçık sayıları Çizelge
4.25.’te verilmiştir.
Çizelge 4.25.’te özetlendiği gibi en yüksek başakta başakçık sayısına sahip hat ZF 3
olup, bunu LAD 388, CWT ve BDMT tritikale hatları izlemiştir. En düşük başakta
başakçık sayısı ise sırası ile ZF 12, ZF 15 ve ZF 16 tritikale hatlarından elde edilmiştir.
53

Çizelge 4.25. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama başakçık sayısı (adet/başak)
Hatlar 1. Yıl 2.Yıl Ortalama TATLICAK-97 21.75 25.15 23.45 ZF 1 23.58 23.23 23.41 ZF 2 22.20 24.15 23.16 ZF 3 24.33 29.77 27.05 ZF 4 20.63 23.73 22.18 ZF 5 21.60 24.98 23.29 ZF 6 20.67 24.13 22.40 ZF 7 21.00 21.73 21.37 ZF 8 22.60 22.55 22.76 ZF 9 23.10 20.88 21.99 ZF 10 20.88 20.00 20.44 ZF 11 19.92 21.92 20.92 ZF 12 19.63 19.20 19.42 ZF 13 21.22 21.40 21.31 ZF 14 21.75 22.33 22.04 ZF 15 21.93 18.90 20.42 ZF 16 20.83 20.25 20.54 LAD 490 23.65 24.83 24.24 LAD 388 24.80 26.42 25.61 JGS 22.55 23.77 23.16 BDMT 24.67 25.35 25.01 CWT 24.88 25.30 25.09 GTAD 23.42 22.75 23.09 MT 1 23.47 23.42 23.45 LO 51 22.80 24.20 23.50
Başakta başakçık sayısına ilişkin bulgularımız; Yağbasanlar vd (1989)’un Çukurova
koşullarında yaptığı araştırmada 20.2-33.6 adet başak belirlediği başakta başakçık
sayısına ilişkin değerler, Bursa koşullarında denemeye alınan tritikale hatlarında 25.40-
28.85 adet başakta başakçık sayısı bulan Çöplü (2001) ve Ankara, Haymana
koşullarında yaptığı çalışmada ortalama başakçık sayısının 29.63-22.77 adet arasında
değiştiğini bildiren Çengel (2001)’in bulgularıyla uyumludur. Genel olarak başak
uzunluğu yüksek olan tritikale hatlarında başakçık sayısının da yüksek olduğu
görülmektedir.
4.1.11. Başakta tane sayısı (1. Yıl) Bazı tritikale hatlarında başakta tane sayısına ilişkin verilerle yapılan varyans analiz
sonuçları Çizelge 4.26.’da verilmiştir.
54

Çizelge 4.26. Farklı tritikale hatlarında başakta tane sayısına ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 4265.210 Bloklar 3 217.635 72.545 Hatlar 24 1683.377 70.141 ** Hata 72 2364.197 32.836 C.V. 12.13
**) 0.01 düzeyinde önemli Çizelge 4.26.’da görüldüğü gibi, başakta tane sayısı yönünden tritikale hatları arasındaki
farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.27.’de gösterilmiştir.
Çizelge 4.27. Farklı tritikale hatlarında başakta tane sayısı ortalamaları (adet/başak)
Hatlar Ortalama GTAD 54.55 a 1* LAD 388 53.35 ab 12 ZF 8 53.15 ab 12 BDMT 52.15 abc 12 LO 51 51.78 abc 12 JGS 51.65 abc 12 LAD 490 50.78 a-d 12 ZF 9 50.03 a-e 12 MT 1 49.28 a-e 12 CWT 48.47 a-e 12 ZF 3 48.25 a-e 12 ZF 7 47.90 a-e 12 ZF 15 47.35 a-e 12 ZF 1 46.70 a-e 12 ZF 5 46.30 a-e 12 ZF 6 45.72 a-e 12 ZF 10 43.80 b-e 12 ZF 2 43.78 b-e 12 ZF 14 43.67 b-e 12 ZF 12 43.20 cde 12 ZF 16 42.97 cde 12 ZF 4 42.90 cde 12 TATLICAK-97 41.42 de 2 ZF 13 41.33 de 2 ZF 11 40.80 e 2
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir Çizelgede görüldüğü gibi; başakta tane sayısı ortalamaları 54.55-40.80 adet/başak
arasında değişmiş olup, başakta tane sayısı yönünden en yüksek değer GTAD tritikale
hattından, en düşük değer ise ZF 11 tritikale hattından elde edilmiştir.
55

4.1.12. Başakta tane sayısı (2. Yıl) Bazı tritikale hatlarında başakta tane sayısına ilişkin verilerle yapılan varyans analiz
sonuçları Çizelge 4.28.’de verilmiştir.
Çizelge 4.28. Farklı tritikale hatlarında başakta tane sayısına ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 3662.260 Bloklar 3 47.580 15.860 Hatlar 24 2652.365 110.515 ** Hata 72 962.315 13.365 C.V. 8.16
**) 0.01 düzeyinde önemli Çizelge 4.28.’de görüldüğü gibi, başakta tane sayısı yönünden tritikale hatları arasındaki
farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.29.’da gösterilmiştir.
Çizelge 4.29. Farklı tritikale hatlarında başakta tane sayısı ortalamaları (adet/başak)
Hatlar Ortalama LAD 388 54.58 a 1* ZF 3 53.08 ab 12 ZF 2 51.45 abc 123 JGS 50.55 a-d 1-4 ZF 4 50.22 a-d 1-5 ZF 8 48.58 b-e 1-6 LO 51 48.10 b-f 1-6 ZF 5 47.35 b-g 1-6 CWT 45.90 c-h 2-7 BDMT 45.83 c-h 2-7 ZF 13 45.75 c-h 2-7 LAD 490 45.65 c-h 2-7 ZF 6 45.13 d-ı 2-7 TATLICAK-97 44.70 d-j 3-7 ZF 1 44.58 d-ı 3-7 ZF 7 43.20 e-j 4-8 ZF 14 43.15 e-j 4-8 ZF 13 42.30 f-j 5-8 GTAD 41.55 g-j 678 ZF 16 41.50 g-j 678 ZF 9 41.13 hıj 678 MT 1 39.13 ıjk 789 ZF 10 38.80 jk 789 ZF 12 35.47 kl 89 ZF 15 32.35 l 9
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir.
56

Çizelgede görüldüğü gibi, başakta tane sayıları ortalamaları yönünden tritikale hatları
arasında araştırmanın 2. yalında da farklı gruplar oluşmuştur. En yüksek başakta tane
sayısı ortalamaları 54.58 adet ile LAD 388 tritikale hattından elde edilmiş, bunu
sırasıyla 53.08, 51.45, 50.55 ve 50.22 adet ile ZF 3, ZF 2, JGS ve ZF 4 tritikale hatları
izlemiştir. En düşük başakta tane sayısı ortalaması ise 32.35 adet ile ZF 15 hattında
saptanmıştır.
Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama başakta tane sayıları Çizelge
4.30’da verilmiştir.
Çizelge 4.30. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama başakta tane sayısı
(adet/başak)
Hatlar 1. Yıl 2.Yıl Ortalama TATLICAK-97 41.42 44.70 43.06 ZF 1 46.70 44.58 45.64 ZF 2 43.78 51.45 47.62 ZF 3 48.25 53.08 50.67 ZF 4 42.90 50.22 46.50 ZF 5 46.30 47.35 46.83 ZF 6 45.72 45.13 45.43 ZF 7 47.90 43.20 45.55 ZF 8 53.15 48.58 50.87 ZF 9 50.03 41.13 45.58 ZF 10 43.80 38.80 41.30 ZF 11 40.80 42.30 41.55 ZF 12 43.20 35.47 39.34 ZF 13 41.33 45.75 43.54 ZF 14 43.67 43.15 43.41 ZF 15 47.35 32.35 39.85 ZF 16 42.97 41.50 42.24 LAD 490 50.78 45.65 48.22 LAD 388 53.35 54.58 53.97 JGS 51.65 50.55 51.10 BDMT 52.15 45.83 48.99 CWT 48.47 45.90 47.19 GTAD 54.55 41.55 48.05 MT 1 49.28 39.13 44.21 LO 51 51.78 48.10 49.94
Çizelgede görüldüğü gibi en yüksek başakta tane sayısına sahip tritikale hattı LAD 388
olup, bunu sırasıyla JGS, ZF 8 ve ZF 3 tritikale hatları izlemiştir. En düşük başakta tane
sayısı ise ZF 12 tritikale hattından elde edilmiş olup, bunu sırasıyla ZF 10 ve ZF 15
tritikale hatları izlemiştir. Başakta tane sayısı ortalamalarına ilişkin elde edilen
57

sonuçlarımız; Çukurova koşullarında yapılan çalışmada tritikale hatlarında başakta tane
sayısının birinci denemede 37.9-50.7 adet/başak, ikinci denemede 32.3-51.3 adet/başak
arasında değiştiğini bildiren Genç vd (1987)’nin, Tokat koşullarında yürütülen
çalışmada başakta tane sayısını 35.6-44.0 adet/başak arasında belirlediklerini bildiren
Sencer vd (1987)’nin, Cukurova koşullarında tritikale hatlarında başakta tane sayısının
49.00-52.99 adet/başak olarak belirleyen Genç vd (1988) ve Haymana koşullarında
ortalama başakta tane sayısını 44.07-32.20 adet olarak belirleyen Çengel (2001)’in
bulgularıyla benzerlik göstermektedir. Ancak, Bursa koşullarında yürütülen çalışmada
farklı tritikale hatlarında 62.93-78.95 adet başakta tane sayısını belirlediğini bildiren
Çöplü (2001)’in bulgularıyla uyum göstermemektedir. Bunun nedeni, başakta tane
sayısının; başak uzunluğu, başakta başakçık sayısıyla ilişkili ve önemli genetik özellik
olmasının yanında, farklı ekolojik ve farklı tritikale hatlarının kullanılmasının bu
farklılıkların oluşabileceği düşünülebilir.
4.1.13. Başakta tane ağırlığı (1. Yıl) Bazı tritikale hatlarında başakta tane ağırlığına ilişkin verilerle yapılan varyans analiz
sonuçları Çizelge 4.31.’de verilmiştir. Çizelge 4.31.’de görüldüğü gibi, başakta tane
ağırlığı yönünden tritikale hatları arasındaki farklılıklar 0.01 düzeyinde önemli
bulunmuştur. Bu farklılıkların önem düzeyini belirlemek amacıyla yapılan Duncan testi
sonuçlarına ilişkin ortalamalar Çizelge 4.32.’de gösterilmiştir.
Çizelge 4.31. Farklı tritikale hatlarında başakta tane ağırlığına ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 8.010 Bloklar 3 0.051 0.017 Hatlar 24 4.373 0.182 ** Hata 72 3.586 0.050 C.V. 11.29
**) 0.01 düzeyinde önemli
Çizelgede görüldüğü gibi başakta tane ağırlığı yönünden tritikale hatları oldukça farklı
gruplarda yer almıştır. Ortalama başakta tane ağırlığı araştırmanın ilk yılında, 2.510-
1.463 g arasında değişmiştir. Başakta tane ağırlığı yönünden en yüksek değer 2.510 g
ile ZF 8 tritikale hattından elde edilmiş olup, bunu sırasıyla 2.328 g, 2.205 g ve 2.197 g
58

ile ZF 9, JGS ve LAD 388 tritikale hatları izlemiştir. Başakta tane ağırlığı yönünden en
düşük değer ise 1.163 g ile Tatlıcak-97 tritikale hattından elde edilmiş olup, bunu
sırasıyla ZF 12 (1.717 g), ZF 6 (1.733 g) ve ZF 10 (1.749 g) tritikale hatları izlemiştir.
Çizelge 4.32. Farklı tritikale hatlarında başakta tane ağırlığı ortalamaları (g/başak)
Hatlar Ortalama ZF 8 2.510 a 1* ZF 9 2.328 ab 12 JGS 2.205 abc 123 LAD 388 2.197 abc 123 GTAD 2.125 bcd 123 ZF 3 2.108 b-e 123 LO 51 2.100 b-f 123 CWT 2.075 b-f 123 ZF 7 2.033 b-f 123 ZF 15 2.003 b-f 23 ZF 14 1.988 b-f 23 LAD 490 1.977 b-f 23 BDMT 1.965 b-f 23 ZF 1 1.952 b-f 23 ZF 2 1.946 c-f 234 ZF 5 1.938 c-f 234 ZF 16 1.913 c-f 234 MT I 1.880 c-f 234 ZF 4 1.850 c-f 234 ZF 13 1.840 c-f 234 ZF 11 1.840 c-f 234 ZF 10 1.740 d-g 34 ZF 6 1.733 efg 34 ZF 12 1.717 fg 34 TATLICAK -97 1.463 g 4
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir 4.1.14. Başakta tane ağırlığı (2. Yıl) Bazı tritikale hatlarında başakta tane ağırlığına ilişkin verilerle yapılan varyans analiz sonuçları Çizelge 4.33.’de verilmiştir. Çizelge 4.33. Farklı tritikale hatlarında başakta tane ağırlığına ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 6.423 Bloklar 3 0.063 0.021 Hatlar 24 4.364 0.182 ** Hata 72 1.997 0.028 C.V. 9.77
**) 0.01 düzeyinde önemli
59

Çizelge 4.33.’de görüldüğü gibi, başakta tane ağılığı yönünden tritikale hatları
arasındaki farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem
düzeyini belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.34.’de
gösterilmiştir.
Çizelge 4.34. Farklı tritikale hatlarında başakta tane ağırlığı ortalamaları (g/başak)
Hatlar Ortalama ZF 3 2.220 a 1* ZF 2 2.098 ab 12 ZF 8 2.005 abc 123 ZF 1 1.900 bcd 1-4 ZF 5 1.892 bcd 1-4 ZF 4 1.868 bcd 234 ZF 13 1.832 c-f 2-5 ZF 7 1.770 c-g 2-5 ZF 11 1.773 d-g 3-6 JGS 1.720 d-h 3-7 ZF 14 1.717 d-h 3-7 LAD 388 1.697 d-h 3-7 TATLICAK-97 1.685 d-h 3-7 LO 51 1.670 d-h 3-7 LAD 490 1.638 e-h 3-7 ZF 6 1.625 f-ı 4-7 ZF 9 1.605 f-j 4-7 ZF 16 1.587 g-k 4-7 CWT 1.577 g-k 4-7 BDMT 1.575 g-k 4-7 GTAD 1.553 g-k 4-7 ZF 10 1.485 h-k 567 MT 1 1.392 ijk 67 ZF 15 1.388 jk 67 ZF 12 1.358 k 7
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir Çizelgede görüldüğü gibi araştırmanın ikinci yılında ortalama başakta tane ağırlığı
2.220–1.358 g arasında değişmiştir. En yüksek başakta tane ağırlığı ZF 3 tritikale
hattından, en düşük başakta tane ağırlığı ise ZF 12 tritikale hattından elde edilmiştir.
Araştırmanın ikinci yılında başakta tane ağırlığının birinci yıla oranla daha düşük oluşu;
araştırmanın ikinci yılında yağışın daha az oluşu ve buna bağlı olarak; başak boyu,
başakta başakçık sayısı ve tane sayısının daha az olmasıyla açıklanabilir.
Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama başakta tane ağırlıkları
aşağıdaki çizelge 4.35’de verilmiştir.
60

Çizelge 4.35. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama başakta tane ağırlığı (g/başak)
Hatlar 1. Yıl 2.Yıl OrtalamaTATLICAK-97 1.463 1.685 1.57 ZF 1 1.952 1.900 1.93 ZF 2 1.946 2.098 2.02 ZF 3 2.108 2.220 2.16 ZF 4 1.850 1.868 1.86 ZF 5 1.938 1.892 1.92 ZF 6 1.733 1.625 1.68 ZF 7 2.033 1.770 1.90 ZF 8 2.510 2.005 2.26 ZF 9 2.328 1.605 1.97 ZF 10 1.740 1.485 1.61 ZF 11 1.840 1.733 1.79 ZF 12 1.717 1.358 1.54 ZF 13 1.840 1.832 1.84 ZF 14 1.988 1.717 1.89 ZF 15 2.003 1.388 1.70 ZF 16 1.913 1.587 1.75 LAD 490 1.977 1.638 1.81 LAD 388 2.197 1.697 1.95 JGS 2.205 1.720 1.96 BDMT 1.965 1.575 1.77 CWT 2.075 1.577 1.83 GTAD 2.125 1.553 1.84 MT 1 1.880 1.392 1.64 LO 51 2.100 1.670 1.89
Çizelgede görüldüğü gibi başakta tane ağırlığı yönünden en yüksek değer ZF 8 tritikale
hattında elde edilirken bunu sırasıyla ZF 3 ve ZF 2 tritikale hatları izlemiştir. En düşük
başakta tane ağırlığı ise ZF 12 tritikale hattından elde edilmiş olup bunu sırasıyla
Tatlıcak-97, ZF 10 ve ZF 6 tritikale hattı izlemiştir.
Tritikale hatlarında başakta tane ağırlığına ilişkin bulgularımız; Genç vd (1987)’nin,
Ülger vd (1989)’un, Yağbasanlar (1989)’un, Ünver (1999)’un ve Çengel (2001)’in
bulgularıyla benzerlik göstermektedir. Araştırıcılar; farklı koşullarda ve farklı tritikale
hatlarıyla yaptıkları araştırmalarda başakta tane ağırlığını 1.10-2.40 g arasında
değiştiğini bildirmektedir. Çöplü (2001)’in elde ettiği sonuçlar (2.92-4.12 g), diğer
araştırıcılar ve bizim sonuçlarımıza göre oldukça yüksek bulunmuştur. Bu farklılığın
değişik yetiştirme koşulları ve kullanılan çeşit farklılığından meydana geldiği
söylenebilir.
61

4.1.15. Bin tane ağırlığı (1. Yıl)
Bazı tritikale hatlarında bin tane ağırlığına ilişkin verilerle yapılan varyans analiz
sonuçları Çizelge 4.36.’da verilmiştir. Çizelge 4.36.’da görüldüğü gibi, bin tane ağırlığı
yönünden tritikale hatları arasındaki farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu
farklılıkların önem düzeyini belirleyebilmek amacıyla yapılan Duncan testi sonuçları
Çizelge 4.37.’de verilmiştir.
Çizelge 4.36. Farklı tritikale hatlarında bin tane ağırlığına ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 1865.092 Bloklar 3 8.306 2.769 Hatlar 24 1581.165 65.882 ** Hata 72 275.620 3.828 C.V. 4.76
**) 0.01 düzeyinde önemli Çizelge 4.37. Farklı tritikale hatlarında bin tane ağırlığı ortalamaları (g)
Hatlar Ortalama ZF 2 48.34 a 1* ZF 16 47.79 ab 12 ZF 1 46.79 abc 1-3 ZF 7 45.27 bcd 1-4 ZF 12 45.16 bcd 1-4 ZF 11 44.29 cde 1-5 ZF 9 44.02 c-f 2-5 ZF 10 43.45 d-g 3-6 LO 51 43.08 d-g 3-7 ZF 3 42.23 d-h 4-7 ZF 14 41.99 e-h 4-8 ZF 8 41.89 e-h 4-8 ZF 4 41.50 e-h 4-8 ZF 15 40.91 f-ı 5-9 JGS 40.67 ghı 5-9 ZF 13 40.38 ghı 5-10 MT 1 39.22 hıj 6-11 CWT 39.13 hıj 7-12 ZF 5 37.97 ıjk 8-12 ZF 6 37.26 jk 9-13 LAD 388 36.86 jk 9-13 GTAD 36.47 jk 10-13 LAD 490 35.32 kl 11-13 TATLICAK -97 35.02 kl 12-13 BDMT 33.46 l 13
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir
62

Çizelge 4.37.’de görüldüğü gibi; tritikale hatlarının ortalama bin tane ağırlığı 48. 34-
33.46 g arasında değişmiştir. Bin tane ağırlığı yönünden en yüksek değer ZF 2 tritikale
hattından elde edilmiş olup, bunu sırasıyla ZF 16 (47.79 g) ve ZF 1 (46.79g) tritikale
hatları izlemiştir. En düşük değer ise BDMT tritikale hattından elde edilmiş olup, bunu
Tatlıcak-97 (35.02 g) ve LAD 490 (34.32 g) tritikale hatları izlemiştir.
4.1.16. Bin tane ağırlığı (2. Yıl)
Bazı tritikale hatlarında bin tane ağırlığına ilişkin verilerle yapılan varyans analiz
sonuçları Çizelge 4.38.’de verilmiştir.
Çizelge 4.38. Farklı tritikale hatlarında bin tane ağırlığına ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 1414.188 Bloklar 3 41.297 13.766 Hatlar 24 1067.145 11.464 ** Hata 72 305.786 4.246 C.V. 5.18
**) 0.01 düzeyinde önemli Çizelge 4.38.’de görüldüğü gibi, bin tane ağırlığı yönünden tritikale hatları arasındaki
farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.39.’da gösterilmiştir.
Çizelgede görüldüğü gibi; araştırmanın 2. yılında tritikale hatlarının ortalama bin tane
ağırlığı 42.81-28.03 g arasında değişmiştir. En yüksek bin tane ağırlığı ZF 3 tritikale
hattından elde edilmiş olup, bunu ZF 16 (39.46 g) tritikale hattı izlemiştir. En düşük
değer ise LAD 388 tritikale hattından elde edilmiş olup, bunu LO 51 (30.87 g) tritikale
hattı izlemiştir. Araştırmanın 2. yılında ortalama bin tane ağırlının daha düşük oluşu, 2.
yılda yağışın daha az oluşu, bitkilerin yeterince gelişememesi ve tanelerini
dolduramamalarıyla açıklanabilir.
63

Çizelge 4.39. Farklı tritikale hatlarında bin tane ağırlığı ortalamaları (g)
Hatlar Ortalama ZF 3 42.81 a 1* ZF 16 39.46 ab 12 ZF 1 39.03 bc 123 ZF 6 38.65 bc 234 ZF 7 38.59 bc 234 ZF 2 38.17 bcd 234 ZF 8 38.14 bcd 234 ZF 12 37.46 b-e 2-5 LAD 490 37.31 b-e 2-5 ZF 13 36.89 b-f 2-6 ZF 5 36.62 b-f 2-6 ZF 9 35.96 c-g 2-7 ZF 11 35.65 c-h 2-7 ZF 4 35.63 c-h 2-7 ZF 15 34.79 d-ı 3-8 ZF 14 34.73 d-ı 3-8 ZF 10 34.12 e-j 4-8 TATLICAK-97 33.53 f-j 5-8 CWT 32.83 g-j 6-8 JGS 32.55 hıj 6-8 MT 1 31.93 ıj 789 BDMT 31.92 ıj 789 GTAD 31.56 ıj 789 LO 51 30.87 jk 89 LAD 388 28.03 k 9
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama bin tane ağırlıkları Çizelge
4.40.’da verilmiştir.
Çizelge 4.40.’da görüldüğü gibi bin tane ağırlığı yönünden en yüksek değer ZF 16
tritikale hattından elde edilmiş olup, bu hattı ZF 2 ve ZF 1 tritikale hatları izlemiştir. Bin
tane ağırlıkları 34.27 (TATLICAK-97)-43.62 (ZF 16) g arasında değişme göstermiştir.
Bin tane ağırlığına ilişkin elde edilen sonuçlarımız; Genç vd (1988), Yağbasanlar vd
(1989), Ünver (1999), Çöplü (2001) ve Çengel (2001)’in bulgularıyla uyum
göstermektedir.
64

Çizelge 4.40. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama bin tane ağırlığı (g)
Hatlar 1. Yıl 2.Yıl OrtalamaTATLICAK-97 35.020 33.525 34.27 ZF 1 46.795 39.030 42.91 ZF 2 48.340 38.167 43.25 ZF 3 42.233 42.810 42.52 ZF 4 41.502 35.635 38.57 ZF 5 37.975 36.620 37.30 ZF 6 37.255 38.648 37.95 ZF 7 45.273 38.587 41.93 ZF 8 41.892 38.135 40.01 ZF 9 44.023 35.955 39.99 ZF 10 43.448 34.120 38.78 ZF 11 44.288 35.655 39.97 ZF 12 45.160 37.460 41.31 ZF 13 40.383 36.887 38.64 ZF 14 41.993 34.732 38.36 ZF 15 40.098 34.792 37.45 ZF 16 47.788 39.460 43.62 LAD 490 35.322 37.310 36.32 LAD 388 36.863 28.030 32.45 JGS 40.672 32.548 36.61 BDMT 33.455 31.920 32.69 CWT 39.127 32.825 35.98 GTAD 36.472 31.555 34.01 MT 1 39.222 31.930 35.58 LO 51 43.080 30.870 36.98
4.1.17. Tane verimi (1. Yıl) Bazı tritikale hatlarında tane verimine ilişkin verilerle yapılan varyans analiz sonuçları Çizelge 4.41.’de verilmiştir. Çizelge 4.41. Farklı tritikale hatlarında tane verimine ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 233583.627 Bloklar 3 697.788 232.596 Hatlar 24 179353.945 7473.081 ** Hata 72 53531.894 743.499 C.V. 3.93
**) 0.01 düzeyinde önemli Çizelge 4.41.’de görüldüğü gibi, tane verimi yönünden tritikale hatları arasındaki
farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.42.’de gösterilmiştir.
65

Çizelge 4.42. Farklı tritikale hatlarında tane verimi ortalamaları (kg/da)
Hatlar Ortalama ZF 9 764.1 a 1* ZF 16 760.9 ab 1 L0 51 758.2 ab 1 ZF 15 740.3 abc 12 GTAD 733.6 a-d 123 ZF 5 720.9 a-e 1-4 LAD 388 717.7 b-e 1-5 ZF 10 712.8 c-f 1-5 ZF 11 712.4 c-f 1-5 ZF 13 711.5 c-f 1-5 CWT 706.3 c-f 1-6 MT 1 695.8 c-g 2-7 BDMT 692.7 d-h 2-7 JGS 689.7 d-h 2-7 ZF 4 687.4 e-ı 2-7 TATLICAK-97 684.3 e-j 2-7 ZF 3 678.3 e-j 3-7 ZF 7 670.4 f-j 4-7 ZF 2 667.3 f-j 4-7 ZF 14 660.3 g-j 567 LAD 490 659.3 g-j 567 ZF 8 649.8 hıj 67 ZF 1 643.5 ıj 7 ZF 6 641.3 j 7 ZF 12 576.1 k 8
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir Çizelgede görüldüğü gibi, en yüksek tane verimi ortalamasına 764.1 kg/da ile ZF 9 tritikale
hattı sahip olurken, bunu 760.9 kg/da ile ZF 16, 758.2 kg/da ile LO 51, 740.3 kg/da ile ZF
15, 733.6 kg/da ile GTAD ve 720.9 kg/da ile ZF 5 tritikale hatları izlemiştir. En düşük tane
verimi ortalaması ise 576.1 kg /da ile ZF 12 tritikale hattından elde edilmiştir.
4.1.18. Tane verimi (2. Yıl) Bazı tritikale hatlarında tane verimine ilişkin verilerle yapılan varyans analiz sonuçları
Çizelge 4.43.’de verilmiştir.
Çizelge 4.43. Farklı tritikale hatlarından elde edilen tane verimine ilişkin varyans analizi V. K. S.D. K.T. K.O.
Genel 99 118667.898 Bloklar 3 2726.172 908.724 Hatlar 24 74744.147 3114.339 ** Hata 72 41197.580 572.189 C.V. 5.81
**) 0.01 düzeyinde önemli
66

Çizelge 4.43.’de görüldüğü gibi, tane verimi yönünden tritikale hatları arasındaki
farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.44.’de gösterilmiştir.
Çizelge 4.44. Farklı tritikale hatlarında tane verimi ortalamaları (kg/da)
Hatlar Ortalama TATLICAK-97 472.9 a 1* ZF 2 470.8 a 1 ZF 1 450.6 ab 12 ZF 4 439.2 abc 1-3 ZF 15 426.7 bcd 1-4 JGS 425.6 b-e 1-5 ZF 16 425.0 b-e 1-5 ZF 5 424.6 b-e 1-5 ZF 7 422.1 b-f 1-5 ZF 6 421.3 b-f 1-5 LAD 490 419.4 b-g 1-5 MT I 414.6 b-g 1-5 GTAD 406.7 b-h 2-5 ZF 14 405.0 b-h 2-5 CWT 402.5 c-h 2-5 ZF 13 401.3 c-h 2-5 ZF 8 398.8 c-h 2-5 ZF 10 393.8 c-h 2-5 LAD 388 392.9 c-h 2-5 BDMT 392.5 c-h 2-5 ZF 11 383.5 d-h 3-5 ZF 9 379.4 e-h 3-5 LO 51 376.8 fh 45 ZF 12 374.6 gh 45 ZF 3 364.4 h 5
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir. Çizelge 4.44.’de görüldüğü gibi; araştırmanın 2. yılında ortalama tane verimi 472.9-
364.4 kg /da arasında değişen değerler göstermiştir. En yüksek tane verimi Tatlıcak-97
tritikale çeşidinden, en düşük tane verimi ZF 3 tritikale hattından elde edilmiştir.
Araştırmanın ikinci yılına ait tane verimi ortalamaları 1. yıla oranla düşük çıkmıştır.
Denemenin 2. yılında bölgede görülen yağış miktarının az oluşu tane verimini azaltıcı
başlıca faktör olarak söylenebilir. Ayrıca denemenin ilk yılında tane verimi yönünden
iyi performans gösteren tritikale hatları, araştırmanın ikinci yılında aynı performansı
gösterememişlerdir. Bunun nedeni, bazı tritikale hatlarının yağış azlığından
(kuraklıktan) az, bazılarının ise daha fazla etkilenmiş olması olarak düşünülebilir (Yıl x
çeşit interaksiyonu).
67

Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama tane verimleri Çizelge 4.45’te
verilmiştir.
Çizelge 4. 45. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama tane verimi (kg/da)
Hatlar 1. Yıl 2.Yıl Ortalama TATLICAK-97 684.2 472.9 578.6 ZF 1 643.5 450.6 547.0 ZF 2 667.3 470.8 569.0 ZF 3 678.3 364.4 521.4 ZF 4 687.4 439.2 563.3 ZF 5 720.9 424.6 572.6 ZF 6 641.3 421.3 531.3 ZF 7 670.4 422.1 546.3 ZF 8 649.7 398.8 524.2 ZF 9 764.0 379.4 571.7 ZF 10 712.7 393.8 552.9 ZF 11 712.3 383.5 547.9 ZF 12 576.1 374.6 475.3 ZF 13 711.4 401.3 555.7 ZF 14 660.2 405.0 532.6 ZF 15 740.3 426.7 583.5 ZF 16 760.9 425.0 592.9 LAD 490 659.2 419.4 539.3 LAD 388 717.6 392.9 555.3 JGS 689.6 425.6 557.6 BDMT 692.7 392.5 542.7 CWT 706.2 402.5 554.6 GTAD 733.6 406.7 570.1 MT 1 695.7 414.6 555.2 LO 51 758.1 376.9 567.4
Çizelgede görüldüğü gibi iki yılın ortalaması olarak en yüksek tane verimi ZF 16
tritikale hattından elde edilmiş olup, bunu sırasıyla ZF 15, Tatlıcak-97 tritikale çeşidini
ve ZF 5 tritikale hattı izlemiştir. En düşük tane verimi ise ZF 12 tritikale hattından elde
edilmiştir. Çizelgede görüldüğü gibi, bazı tritikale hatlarının (ZF 15 ve ZF 16) verimleri
2 yılın ortalaması olarak kontrol çeşidini geçmiştir, bazıları ise yakın değerler
göstermiştir (ZF 12, ZF 5, ZF 9 ve GTAD). Bu hatlar daha sonra yapılacak ıslah
çalışmalarında, yüksek verim potansiyeli yönünden çeşit adayı olarak önerilebilir.
Tritikale hatlarında belirlenen verim değerleri; Almanya koşullarında yürüttüğü
çalışmada tritikalede tane verimini 610-780 kg/da olarak bildiren Krolow (1977),
Çukurova koşullarında tane verimini 540-667 kg/da olarak belirten Genç vd (1987),
68

Polonya koşullarında triticale verimini 483 kg/da olarak bildiren Koc vd (2000), Bursa
koşullarında yürütülen çalışmada tritikale hatlarında 744.6-960.3 kg/da verim elde
ettiğini belirten Çöplü (2001) ve Ankara, Haymana koşullarında yürüttüğü çalışmada
857.5–644.5 kg /da tane verimi elde ettiğini belirten Çengel (2001)’in sonuçlarıyla
benzerlik göstermektedir. Ancak tane veriminne ilişkin bulgularımız; Tokat Artova
koşullarında tritikalede tane verimini 164-369 kg/da olarak bildiren Sencer (1997) ve
Sivas-Şarkışla koşullarında tritikalede tane verimini 252-460 kg/da olarak bildiren
Taşyürek vd (1999a)’un sonuçlarıyla benzerlik göstermemiştir. Belirlenen bu farklılığın
farklı iklim ve toprak koşullarından kaynaklandığı söylenebilir.
Tane verimini etkileyen pek çok faktör bulunmaktadır. İklim ve toprak özelliklerinin
yanında yetiştirme tekniklerinin etkisi altında değişen verim, kantitatif bir karakter olup,
pek çok sayıda genin etkisi altındadır. Çalışmamızda elde edilen bulgulara göre; tane
veriminin bazı tritikale hatları için, iklim faktörlerinden son derece etkilendiği
söylenebilir.
4.1.19. Hasat indeksi (1. Yıl)
Bazı tritikale hatlarında hasat indeksine ilişkin verilerle yapılan varyans analiz sonuçları
Çizelge 4.46.’da verilmiştir.
Çizelge 4.46. Farklı tritikale hatlarında hasat indeksine ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 605.606 Bloklar 3 5.391 1.797 Hatlar 24 216.808 9.034 * Hata 72 383.407 5.325 C.V. 6.44
*) 0.05 düzeyinde önemli Çizelge 4.46.’da görüldüğü gibi, hasat indeksi yönünden tritikale hatları arasındaki
farklılıklar 0.05 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.47.’de gösterilmiştir.
Çizelgede görüldüğü gibi araştırmanın ilk yılında ortalama hasat indeksi % 39.65–
28.40 arasında değişmiştir. Hasat indeksi yönünden en yüksek değer ZF 16 tritikale
hattında belirlenirken, en düşük hasat indeksi ZF 3 tritikale hattından elde edilmiştir.
69

Çizelge 4.47. Farklı tritikale hatlarında hasat indeksi ortalamaları (%)
Hatlar Ortalama ZF 16 39.65 a GTAD 37.70 ab ZF 9 37.20 ab ZF 4 36.97 abc ZF 8 36.57 abc ZF 5 36.42 a-d LAD 388 35.42 a-d ZF 11 35.02 a-d ZF 14 35.00 a-d BDMT 34.97 a-d ZF 2 34.82 a-e LO 51 34.45 a-e ZF 13 34.35 a-e ZF 10 34.32 a-e ZF 7 34.32 a-e TATLICAK_97 33.72 a-e LAD 490 33.65 a-e MT 1 33.47 a-e ZF 1 33.35 a-e JGS 33.02 b-e ZF 6 32.62 b-e ZF 12 31.90 b-e CWT 30.55 cde ZF 15 29.97 de ZF 3 28.40 e
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir 4.1.20. Hasat İndeksi (2. Yıl) Bazı tritikale hatlarında hasat indeksine ilişkin verilerle (açı değerleri) yapılan varyans
analiz sonuçları Çizelge 4.48.’de verilmiştir.
Çizelge 4.48. Farklı tritikale hatlarında hasat indeksine ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 610.767 Bloklar 3 3.466 1.155 Hatlar 24 216.139 9.006 Hata 72 391.163 5.433 C.V. 6.86
Çizelge 4.48.’de görüldüğü gibi, hasat indeksi yönünden tritikale hatları arasındaki
farklılıklar istatistiki yönden önemsiz bulunmuştur. Tritikale hatlarının hasat indeksi
yönünden almış olduğu ortalama (gerçek değerler) değerler Çizelge 4.49.’da
gösterilmiştir.
70

Çizelge 4.49. Farklı tritikale hatlarında hasat indeksi ortalamaları (%)
Hatlar Ortalama ZF 2 36.15 ZF 13 34.07 GTAD 33.97 ZF 7 34.20 ZF 16 33.10 ZF 11 33.42 LAD 388 33.02 MT 1 32.87 LO 51 32.80 BDMT 31.85 CWT 31.60 TATLICAK-97 31.77 ZF 10 31.70 ZF 15 31.42 ZF 9 30.92 JGS 30.60 ZF 4 30.52 ZF 3 30.20 ZF 6 30.35 ZF 8 30.12 ZF 5 28.45 ZF 14 28.10 ZF 12 27.87 LAD 490 27.65 ZF 1 26.17
Çizelgede görüldüğü gibi araştırmanın 2. yılında hasat indeksi ortalamaları % 36.15-
26.17 arasında değişme göstermiştir. Hasat indeksi yönünden en yüksek ortalama %
36.15 ile ZF 2 tritikale hattından, en düşük ortalama hasat indeksi % 26.17 ile ZF 1
tritikale hattından elde edilmiştir. Ancak araştırmanın ikinci yılında tritikale hatları
hasat indeksi yönünden istatistiki olarak aynı grupta yer almışlardır.
Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama hasat indeksi değerleri
Çizelge 4.50’de verilmiştir.
Çizelge 4.50’de görüldüğü gibi en yüksek hasat indeksine sahip tritikale hattı ZF 16
(%36.37), en düşük hasat indeksine sahip hat ise ZF 3 (% 29.30) tür.
71

Hasat indeksi ortalamaları incelendiğinde tritikalenin genel olarak diğer serin iklim
tahıllarına oranla daha düşük hasat indeksine sahip olduğu söylenebilir. Bu sonuç, bitki
boyunun tritikalede diğer serin iklim tahıllarına oranla daha fazla olması ile
açıklanabilir. Araştırma bulgularımız, tritikalede hasat indeksini % 25.3-30.6 olarak
bildiren Taşyürek vd 1999a ve % 21.68-31.51 olarak bulan Ünver 1999’un sonuçlarıyla
uyum göstermektedir.
Çizelge 4.50. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama hasat indeksi (%)
Hatlar 1. Yıl 2.Yıl OrtalamaTATLICAK-97 33.72 31.57 32.64 ZF 1 33.35 26.17 29.76 ZF 2 34.82 36.15 35.48 ZF 3 28.40 30.20 29.30 ZF 4 36.97 30.52 33.74 ZF 5 36.42 28.45 32.43 ZF 6 32.62 30.35 31.48 ZF 7 34.32 34.20 34.26 ZF 8 36.57 30.12 33.34 ZF 9 37.20 30.92 34.06 ZF 10 34.32 31.70 33.01 ZF 11 35.02 33.42 34.22 ZF 12 31.90 27.87 29.88 ZF 13 34.35 34.07 34.21 ZF 14 35.00 28.10 31.55 ZF 15 29.97 31.42 30.69 ZF 16 39.65 33.10 36.37 LAD 490 33.65 27.65 30.65 LAD 388 35.42 33.02 34.22 JGS 33.02 30.60 31.81 BDMT 34.97 31.85 33.41 CWT 30.55 31.60 31.07 GTAD 37.70 33.97 35.83 MT 1 33.47 32.87 33.17 LO 51 34.45 32.80 33.62
4.1.21. Protein oranı (1. Yıl) Bazı tritikale hatlarında protein oranına ilişkin (açı değerleri) verilerle yapılan varyans
analiz sonuçları Çizelge 4.51.’de verilmiştir. Çizelge 4.51.’de görüldüğü gibi, protein
oranı yönünden tritikale hatları arasındaki farklılıklar 0.01 düzeyinde önemli
bulunmuştur. Bu farklılıkların önem düzeyini belirleyebilmek amacıyla yapılan Duncan
testi sonuçları Çizelge 4.52.’de gösterilmiştir
72

Çizelge 4.51. Farklı tritikale hatlarında protein oranına ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 58.739 Bloklar 3 0.383 0.128 Hatlar 24 37.767 1.574 ** Hata 72 20.589 0.286 C.V. 2.51
**) 0.01 düzeyinde önemli
Çizelge 4.52’de görüldüğü gibi tritikale hatları protein oranı yönünden farklılık
göstermiştir. En yüksek protein oranı % 14.33 ile ZF 3 tritikale hattından elde edilirken,
en düşük protein oranı ise % 11.31 ile JGS tritikale hattından elde edilmiştir (Gerçek
değerler).
Çizelge 4.52. Farklı tritikale hatlarında protein oranı ortalamaları (%)
Hatlar Ortalama ZF 3 14.33 a 1* BDMT 14.19 ab 1 ZF 1 14.25 ab 1 ZF 14 13.81 abc 12 LO 51 13.77 abc 123 ZF 16 13.74 abc 123 LAD 388 13.58 a-d 123 MT 1 13.77 a-d 123 ZF 9 13.79 a-d 123 ZF 15 13.54 a-e 123 CWT 13.45 a-e 123 ZF 8 13.66 a-e 123 ZF 7 13.25 b-f 123 ZF 13 13.25 b-f 123 ZF 2 12.88 c-f 123 ZF 12 12.93 c-f 123 ZF 6 13.00 c-f 123 ZF 5 12.98 c-f 1-4 ZF 11 12.98 c-f 1-4 LAD 490 12.54 def 234 ZF 10 12.65 def 234 TATLICAK-97 12.54 efg 2-5 ZF 4 12.31 fg 345 GTAD 11.69 gh 45 JGS 11.31 h 5
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir
73

4.1.22. Protein oranı (2. Yıl) Bazı tritikale hatlarında protein oranına ilişkin verilerle yapılan varyans analiz
sonuçları Çizelge 4.53.’de verilmiştir.
Çizelge 4.53. Farklı tritikale hatlarında protein oranına ilişkin varyans analizi
V. K. S.D. K.T. K.O. Genel 99 131.076 Bloklar 3 0.675 0.225 Hatlar 24 113.435 4.726 ** Hata 72 16.965 0.236 C.V. 2.32
**) 0.01 düzeyinde önemli Çizelge 4.53.’de görüldüğü gibi, protein oranı yönünden tritikale hatları arasındaki
farklılıklar 0.01 düzeyinde önemli bulunmuştur. Bu farklılıkların önem düzeyini
belirleyebilmek amacıyla yapılan Duncan testi sonuçları Çizelge 4.54’de gösterilmiştir.
Çizelge 4.54. Farklı tritikale hatlarında protein oranı ortalamaları (%)
Hatlar Ortalama ZF 13 14.77 a 1* ZF 3 14.44 ab 12 ZF 16 14.29 abc 1-3 ZF 9 14.21 abc 1-4 ZF 12 14.10 a-d 1-4 ZF 4 14.04 a-d 1-4 ZF 15 13.78 b-e 1-4 LAD 388 13.62 b-f 1-5 ZF 8 13.51 c-g 2-5 ZF 1 13.29 d-h 3-6 ZF 5 12.83 e-ı 3-6 ZF 11 13.18 e-ı 3-6 JGS 12.91 e-ı 3-6 ZF 2 12.83 f-ı 456 ZF 14 12.63 ghı 567 ZF 10 12.62 gl 567 ZF 7 12.23 hı 678 GTAD 12.35 hıj 678 MT I 12.06 hıj 678 LAD 490 11.74 ıj 78 TATLICAK-97 11.28 k 8 CWT 11.19 k 8 BDMT 11.14 k 8 ZF 6 10.52 k 8 LO 51 10.06 l 9
*) Harfler 0.05, rakamlar 0.01 düzeyinde farklı grupları göstermektedir
74

Çizelgede görüldüğü gibi; protein oranı yönünden tritikale hatları arasında oldukça
farklı gruplar oluşmuştur. Tritikale hatları arasında ortalama protein oranı 2. yılda; %
14.77-10.06 arasında değişme göstermiştir. En yüksek protein oranı ZF 13 tritikale
hattında saptanırken, en düşük protein oranı LO 51 tritikale hattından elde edilmiştir.
Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama protein oranları Çizelge
4.55’te verilmiştir.
Çizelge 4.55. Farklı tritikale hatlarında birinci yıl, ikinci yıl ve ortalama protein oranı
Hatlar 1. Yıl 2.Yıl Ortalama TATLICAK-97 12.54 11.27 11.90 ZF 1 14.25 13.29 13.77 ZF 2 12.88 12.83 12.85 ZF 3 14.33 14.44 14.37 ZF 4 12.31 14.04 13.17 ZF 5 12.98 12.83 12.90 ZF 6 13.00 10.52 11.76 ZF 7 13.25 12.23 12.74 ZF 8 13.66 13.51 13.58 ZF 9 13.79 14.21 14.00 ZF 10 12.65 12.62 12.63 ZF 11 12.98 13.18 13.08 ZF 12 12.93 14.10 13.51 ZF 13 13.25 14.77 14.01 ZF 14 13.81 12.62 13.21 ZF 15 13.54 13.77 13.65 ZF 16 13.74 14.29 14.01 LAD 490 12.54 11.73 12.14 LAD 388 13.58 13.62 13.60 JGS 11.31 12.91 12.11 BDMT 14.19 11.14 12.66 CWT 13.45 11.19 12.32 GTAD 11.69 12.35 12.02 MT 1 13.77 12.06 12.92 LO 51 13.77 10.06 11.91
Çizelgede görüldüğü gibi protein oranı yönünden en yüksek değer ZF 3 tritikale
hattından elde edilmiş olup, bunu ZF 16, ZF 13 ve ZF 9 tritikale hattı izlemiştir. Protein
oranı yönünden en düşük değer ise ZF 6 tritikale hattından elde edilmiştir.
Protein oranına ilişkin bulgularımız Tritikalede protein oranını % 9.6-12.2 olarak
belirten Feil ve Fossati (1995)’in ve % 12 olarak bildiren Koc vd (2000)’in bulgularıyla
75

benzerlik göstermektedir. Kün (1996) ve Çiftçi vd (2003) tritikalede protein oranının
diğer serin iklim tahıl cinslerine göre daha yüksek olabileceğini bildirmektedirler.
Doktora çalışmasının tarla deneme sonuçları topluca değerlendirildiğinde; Ankara
koşullarında 24 tritikale ıslah hattı ve kontrol çeşidi (Tatlıcak-97) kullanılan çalışmada,
verim ve verim öğelerinin incelenen bazı özellikler yönünden (1.yıl başak uzunluğu ve
2.yıl hasat indeksi dışında) önemli farklılıklar belirlenmiştir. İki yılın ortalaması olarak
elde edilen sonuçlar aşağıda özetlenmiştir.
Çiçeklenme tarihi; en erken çiçeklenme tarihine sahip olan hat ZF 12 (27.00 gün), en
geç çiçeklenme tarihine sahip hat ise LAD 388 (35.87 gün)’dir. Diğer hatlar bu iki değer
arasında yer almıştır.
Bitki boyu; en kısa bitki boyuna sahip tritikale hattı MT 1 (109.6 cm), en uzun tritikale
hattı ise ZF 3 (144.1 cm) dir.
Fertil kardeş sayısı; en fazla kardeşlenme gösteren tritikale hattı 5.27 adet/bitki ile ZF
15, en az kardeşlenme gösteren tritikale hattı ise 4. 31 adet/bitki ile JGS olmuştur.
Başak uzunluğu; en uzun başak uzunluğuna sahip tritikale hattı CWT (107.9 mm) olup,
bunu LAD 388 (107.3 mm) hattı izlemiştir. En kısa başak uzunluğuna sahip tritikale
hattı ise ZF 7 (85.24 mm) olmuştur. Diğer tritikale hatları bu iki değer arasında başak
uzunluğuna sahip olmuştur.
Başakçık sayısı; tritikale hatlarının ortalama başakçık sayısı 19.42 adet/başak (ZF 12)
ile 27. 05 adet /başak (ZF 3) arasında değişmiştir.
Başakta tane sayısı; en yüksek başakta tane sayısına sahip tritikale hattı LAD 388, en
düşük başakta tane sayısına sahip tritikale hattı ise ZF 12 olmuştur.
Başakta tane ağırlığı; yönünden en yüksek değer ZF 8 tritikale hattında elde edilirken,
en düşük başakta tane ağırlığı ise ZF 12 tritikale hattından elde edilmiştir.
76

Bin tane ağırlığı; bin tane ağırlığı yönünden en yüksek değer 43.62 g ile ZF 16 tritikale
hattından elde edilmiştir. En düşük değer ise 32.45 g ile LAD 388 tritikale hattından
elde edilmiştir.
Tane verimi; en yüksek tane verimine sahip tritikale hattı ZF 16 (592.9 kg/da) olup, bu
sırasıyla ZF 15 tritikale hattı ve Tatlıcak-97 tritikale çeşidi izlemiştir. En düşük tane
verimine sahip tritikale hattı ise ZF 12 (475.3 kg/da) olmuştur.
Hasat indeksi; hasat indeksi yönünden tritikale hatları % 29.30 ile 36.37 arasında
değişen değerler göstermişlerdir.
Protein oranı; Yapılan protein analizine göre tritikale hatlarının ortalama olarak % 11.76
(ZF 6) ile 14.37 (ZF 3) arasında değişen değerler gösterdiği saptanmıştır. Tritikale
hatlarında incelenen özellikler yönüyle geniş bir varyasyon belirlenmiştir.
Tritikale çeşit ve hatlarında incelenen morfolojik karakterler yönüyle geniş bir
varyasyonun olduğu görülmektedir. Bu varyasyonların yapılacak ıslah çalışmalarında
kullanılma olanağı bulunmaktadır. Tritikale çeşit ve hatlarında incelenen özellikler
yönüyle ortaya çıkan farklılıkların genetik farklılıktan kaynaklandığı söylenebilir.
Yıllara göre genotiplerin incelenen özellikler yönüyle farklı performans göstermesinin
iklim faktörlerinden kaynaklandığı söylenebilir. Morfolojik görüntü olarak birbirine
benzer tritikale genotiplerinin incelenen morfolojik özellikler (verim ve verim kriterleri)
yönüyle farklı olduğu saptanmıştır.
77

4.2. Laboratuvar Çalışması Bulguları ve Tartışma
Doktora çalışmasının bu bölümünde; RAPD-PCR tekniği kullanılarak bazı tritikale
hatlarının genetik karakterizasyonu belirlenmeye çalışılmıştır. RAPD-PCR tekniğinden
önce PCR optimizasyonu çalışmaları yapılmıştır.
4.2.1. DNA izolasyonunun optimizasyonu
Literatüre dayalı olarak tritikalede uygulanabilecek DNA izolasyon yöntemleri
araştırılmış ve farklı yöntemler denenmiştir. Ön çalışmalar sonucunda DNA yoğunluğu
ve temizliği yönünden en uygun olarak Ma ve Sorrells (1995)’den modifiye edilen
DNA izolasyon yöntemi bulunmuştur. Bu yönteme göre yapılan DNA izolasyonu
sonucu elde edilen DNA’ların jel elektroforez görüntüleri Şekil 4.1’de, spektrometre ile
yapılan ölçüm değerleri ise Çizelge 4.56’da verilmiştir. İzole edilen genomik
DNA’lardan 2 µl ve 7 µl yükleme tamponu ile Agaroz jele yüklenmiş ve DNA’lar 20-30
dakika elektroforezis yapılmıştır. Şekil 4.1’de görüldüğü gibi ZF 3 ve ZF 4 tritikale
hatlarına ait DNA örneklerinin çok yoğun, ZF 14, ZF 15, ZF 16, JGS, CWT ve MT 1
hatlarına ait DNA örneklerinin yoğun olduğu gözlenirken, ZF 1, ZF 8, LAD 388 ve LO
51 hatlarına ait DNA örneklerin yoğunluğunun ise az olduğu gözlenmektedir.
Şekil 4.1. İzole edilen genomik DNA’ların jel elektroforez görüntüsü
78

Çizelge 4.56. Tritikale çeşit ve hatlarında spektrometrede ölçülen DNA miktarları
Hatlar DNA (µg/ml) A 260/280 A 230/260 A 3201 Tatlıcak-97 480 1.83 2.69 0.000 2 ZF 1 124 1.73 2.20 0.003 3 ZF 2 295 1.77 2.65 0.004 4 ZF 3 756 1.71 - 0.001 5 ZF 4 364 1.84 3.43 0.001 6 ZF 5 219 1.81 3.22 0.004 7 ZF 6 462 1.73 5.15 0.004 8 ZF 7 178 1.80 2.02 0.006 9 ZF 8 131 1.77 4.06 0.007 10 ZF 9 145 1.70 19.00 0.005 11 ZF 10 149 1.55 - 0.008 12 ZF 11 376 1.60 3.90 0.014 13 ZF 12 219 1.65 4.70 0.007 14 ZF 13 1002 1.80 2.90 0.006 15 ZF 14 1385 1.50 4.16 0.009 16 ZF 15 400 1.60 9.00 0.001 17 ZF 16 1750 2.00 4.04 0.001 18 GTAD 410 1.50 - 0.008 19 JGS 1800 1.40 3.97 0.021 20 BDMT 1100 2.15 1.60 0.000 21 CWT 500 1.48 2.14 0.015 22 MT 1 1860 1.42 2.15 0.013 23 LAD 490 1350 1.51 2.20 0.012 24 LAD 388 160 1.73 2.30 0.013 25 LO 51 170 1.70 2.43 0.012
Çizelge 4.56’da görüldüğü gibi, tritikale çeşit ve hatlarında genomik DNA
miktarlarının 124–1860 µg/ml arasında değişmiştir. DNA miktarı 1860 µg/ml ile MT 1
tritikale çeşidinde en yüksek, 124 µg/ml ile ZF 1 tritikale hattında en düşük olarak
bulunmuştur. DNA’ nın protein saflığı (A 260/280 ) JGS hattında 1.40 ile en az, ZF 16
hattında ise 2.00 ile en yüksek olarak bulunmuştur. DNA’nın temizliği için ise A260/280
ve A 260/230 değerlerine bakılmıştır. Çünkü DNA 260, protein 280 ve karbonhidratlar da
230 nm dalga boylarında pik (en yüksek değer) yapmaktadır. Temiz bir DNA’ da
A260/280 oranı 1.80 ile 2.00 arasında; A 260/230 oranı ise 2.00 den büyük; A 320 ise 0’ a
yakın olmalıdır. Nitekim araştırıcılar, RAPD yönteminde ne kadar saf ve yüksek
molekül ağırlıklı DNA izole edilebilirse bantların belirginliğinin ve üretilebilirliğinin o
derece arttığını belirtmektedirler (Weising vd 1995, Penner 1996).
79

Yapılan DNA ön izolasyon çalışmalarında tritikale bitkisinde, yaprakların alım
zamanının da DNA kalitesi ve yoğunluğu açısından önemli olduğu gözlenmiştir. DNA
izolasyonunda büyüme periyodu başlangıcında genç sürgün ve yapraklarının
kullanılması elde edilen DNA yoğunluğu ve temizliği yönünden iyi sonuç vermiştir.
4.2.2. DNA konsantrasyonunun optimizasyonu
Kalıp DNA konsantrasyonu amplifikasyonun başarısı için çok önemli bir faktördür.
Tritikale yapraklarından izole edilen DNA miktarı, UV-spektrometrede 260 nm dalga
boyunda absorbsiyon değerinden faydalanılarak hesaplanmıştır.
DNA miktarı (µg/ml) =Absorbbans değeri (OD 260) x 50 Seyreltme Katsayısı
Yukarıdaki formüle göre hesaplanan tüm tritikale çeşit ve hatlarına ait DNA miktarları
124-1860 µg/ml arasında değişme göstermiştir. Tekrarlanabilen RAPD profillerinin elde
edilmesi için reaksiyon karışımında bulunması gereken DNA miktarının
belirlenebilmesi ve reaksiyon kullanımı için her bir DNA örneği 50 ng/µl olacak
şekilde TE buffer ile seyreltilmiştir. RAPD-PCR çalışmalarında kalıp DNA
konsantrasyonu; Susamda 7 ng/25 µl (Şençiçek 2000), Arpada 12.5 ng/25 µl (De Bustos
1998), Sorgumda 20-50 ng/20 µl (Pammi vd 1994 ), buğdayda 20 ng/20 µl (Fahima vd
1999) ve tritikalede ise 80 ng/20 µl (Amiour vd 2002) olarak belirtilmektedir.
Çalışmamızda, ön optimizasyon çalışmaları boyunca, 1, 1.5, 2.0, 2.5, 3.0, 3.5, 4.0 ve 5.0
µl (50ng/µl) DNA konsantrasyonları denenmiş ve RAPD-PCR reaksiyonlarında
tritikale hatları için uygun olan DNA miktarının 2 µl DNA /25µl olduğu saptanmıştır.
25 µl’ lik PCR tüplerinde toplam 100 ng DNA olacak şekilde karışımlar (mix)
hazırlanmış ve RAPD reaksiyonlarında kullanılmıştır (Çizelge 4.57).
4.2.3. MgCl2 konsantrasyonunun optimizasyonu
Magnezyum klorür konsantrasyonu primer yapışmasına (annealing), enzim aktivitesine,
ürünün belirginliğine ve primer-dimer oluşumlarına etki etmektedir (Devos ve Gale
1992, Pancholi 1995). Düşük konsantrasyonlu MgCl2 kullanıldığı zaman genellikle
80

düşük verimli ürün veya hiç ürün oluşmazken, yüksek konsantrasyonlu Mg iyonu
kullanıldığında ise istenmeyen ürünlerin oluşmasına neden olmaktadır. Çalışmamızda
elde edilen sonuçlar bu araştırıcıların sonuçlarıyla benzerlik göstermiştir. Çalışmada
düşük Mg konsantrasyonlarında PCR ürünleri çok silik görüntü vermiş, yüksek Mg
konsantrasyonlarında ise Agaroz jel üzerinde tüm DNA kulvarına yayılan (smear) bir
görüntü oluşmuştur. PCR ön optimizasyon çalışmalarında; 1.8, 2.0, 2.5, 3.0, 3.5, 4.0,
4.5, ve 6.0 µl/25 µl (25 mM MgCl2) konsantrasyonları denenmiş, elde edilen sonuçlara
göre 2.5–3.5 µl/25 µl MgCl2 (25 mM) konsantrasyonun görülebilir RAPD bantları
verdiği saptanmıştır. Ancak, çalışmamızda PCR reaksiyonları için 2.5 µl/25 µl MgCl2
konsantrasyonunun kullanılmıştır (Çizelge 4.57).
Yapılan diğer bazı PCR-RAPD çalışmalarında uygun MgCl2 konsantrasyonu;
Sorgumda 25 mM/20 µl (Pammi vd 1994), arpada 50 mM/25µl (De Bustos vd 1988),
tritikale ve buğdayda 200 µM/20µl (Malgorzata vd 2002) ve tritikalede 1.5 mM olarak
bildirilmektedir.
4.2.4. Primer konsantrasyonunun optimizasyonu
Primer konsantrasyonu yüksek olduğu durumlarda kalıp DNA’dan bağımsız primer-
dimerlerinin ve spesifik (özel) olmayan ürünlerin oluşmasına neden olmaktadır
(Weeden vd 1992, Pancholi 1995). Yapılan bazı çalışmalarda tritikalede PARD için
uygun primer konsantrasyonu 0.5-1.2 µM/20µl olarak belirtilmektedir (Malgorzata vd
2002, Amiour vd 2002).
Çalışmada 10, 12 ve 18 nükleotidlik 60 farklı primer kullanılmıştır. Çalışmamızda,
tritikale çeşit ve hatları için uygun primer yoğunluğunu tespit etmek amacıyla 50
ng/µl’lik stoktan 1.2.3.4 µl kullanılarak denemeler yapılmış ve uygun primer yoğunluğu
4µl olarak tespit edilmiştir (Çizelge 4.57). Primerlerin Tm sıcaklığının 2-3 °C
üzerindeki sıcaklıklar amplifikasyon ürünlerinin daha net görünmesini sağlamıştır.
RAPD yönteminde primerlerin DNA bantlarının üretilebilirliğini önemli derecede
etkilediği söylenebilir. DNA bantlarının üretilebilirliğini artırmak amacıyla her bir
primer için ayrı ayrı optimizasyonun yapılması önerilebilir.
81

4.2.5. dNTP konsantrasyonunun optimizasyonu
Deoksinukleotid trifosfat (dNTP) konsantrasyonunun düşük miktarda kullanımı PCR’ ın
güvenirliğini artırmaktadır. Düşük dNTP konsantrasyonu nükleotidlerin yanlış
bağlanma olasılığını azaltmaktadır (Pancholi 1995). Malgorzata vd (2002) tritikalede
RAPD için uygun dNTP yoğunluğunu 200 µM/20 µl olarak bildirmektedir.
Çalışmamızda 1, 1.5, 2 ve 2.5 µl dNTP konsantrasyonları denenmiş ve uygun dNTP
konsantrasyonu 2 µl/25µl olarak saptanmıştır (Çizelge 4. 57).
4.2.6. Taq Polimeraz enzim konsantrasyonunun optimizasyonu
Taq DNA polimeraz 25 µl’lik reaksiyon karışımında 0.25 ve 0.6 unite olacak şekilde
önerilmektedir. Bununla birlikte Taq polimeraz miktarı kalıp DNA’ ya veya primerlere
göre değişmektedir. Reaksiyon karışımında eğer enzim miktarı çok yüksek olursa
spesifik olmayan yan ürünler birikirken, düşük Taq konsantrasyonunda istenen
ürünlerün miktarı yetersiz olmaktadır (Devos and Gale 1992). Çalışmada ön denemeler
sonucunda uygun Taq polimeraz konsantrasyonu 1 U/25µl olarak belirlenmiştir
(Çizelge 4. 57).
4.2.7. Diğer reaksiyon bileşenleri
Çalışmada kullanılan 10X buffer (Tampon) Taq polimeraz enzimi ile birlikte gelmiş
olup (Promega), 50 mM Tris-HCI (pH 8.0), 100 mM KCl, 0.1 mM EDTA, 1 mM DTT,
% 50 glcerol ve % 1 TritonX-100 içermektedir. Reaksiyonda 10X buffer’dan her PCR
tüpü için 2.0 µl olacak şekilde kullanılması iyi sonuç vermiştir (Çizelge 4.57).
Çalışmamızda öptimizasyon çalışmalarında test edilen parametreler ve elde edilen
sonuçlar toplu olarak Çizelge 4. 57’de özetlenmiştir.
82

Çizelge 4.57. PCR optimizasyonu için test edilen parametreler ve elde edilen sonuçlar
PCR için reaksiyon karışımı
Test edilen reaksiyon bileşeni
Bileşen konsantrasyonu (µl / 25 µl)
Sonuç
MgCl2 ( 2.5) Primer (4 µl) dNTP (2µl) Taq ( 0.3 µl) 10 x buffer ( 2.5µl)
DNA
1.0 1.5 2.0 2.5 3.0 3.5 4.0 5.0
Amplifikasyon yok Amplifikasyon yok Amplifikasyon çok iyi Belirgin olmayan bant Çok silik bant Amplifikasyon yok Amplifikasyon yok Amplifikasyon yok
DNA (2.0 µl) Primer (4 µl) dNTP (2.0µl) Taq (0.1, 0.3 µl) 10 x buffer ( 2.5µl)
MgCl2
1.8 2.0 2.5 3.0 3.5 4.0 4.5 6.0
Amplifikasyon yok Silik bant oluşumu Belirgin bantlar Belirgin bantlar Belirgin ancak smerli Semearli bant oluşumu Semearli bant oluşumu Amplifikasyon yok
DNA ( 2.0 µl) MgCl2 (2.5µl) dNTP (2.0µl) Taq ( 0.3 µl) 10 x buffer ( 2.5µl)
Primer
1.0 2.0 3.0 4.0
Amplifikasyon yok Amplifikasyon zayıf Amplifikasyon zayıf Amplifikasyon çok iyi
DNA (2.0 µl) MgCl2 (2.5 µl) Primer (4 µl) Taq ( 0.3 µl) 10 x buffer ( 2.5µl)
dNTPs
1.0 1.5 2.0 2.5
Amplifikasyon yok Amplifikasyon yok Amplifikasyon iyi Amplifikasyon zayıf
DNA (2.0 µl) MgCl2 (2.5 µl) Primer (4 µl) 10 x buffer ( 2.5µl) dNTP (2.0 µl)
Taq polimeraz
0.15 (0.5 U) 0.30 (1.0 U) 0.45 (1.5 U)
Amplifikasyon yok Amplifikasyon iyi Silik bant oluşumu
4.2. 8. PCR koşulları
PCR reaksiyonunda çift zincirli DNA 90-95 °C arasındaki sıcaklıkta denatüre
olmaktadır. Bu nedenle, çalışmada 94 °C’de 30 saniye denatürasyon süresi iyi sonuç
vermiştir. Zincirleri ayrılan DNA’ lara primerin komplamenteri (bileşeni) olan bölgeye
bağlanma sıcaklığı olan 37-60 °C düşürülmesi gerekmektedir (Plumbi vd 1991).
Çalışmada primerlerin Tm ısılarına göre değişen sıcaklıklarda bağlanma sıcaklığı
uygulanmıştır (28-64 °C). Genelde primerlerin Tm ısılarının 2-3 °C üzeri sıcaklıklar
amplifikasyon yönünden olumlu sonuçlar vermiştir. Bu sıcaklığı takiben bağlanan
83

primerlerin Taq polimeraz ile çoğaltılması için 70-75 °C arasında bekletilmesi
gerekmektedir (Palumbi vd 1991). 72 °C’ de 1 dakika 45 saniye ekstension (uzama)
sıcaklık ve süresi verilmiştir. Bu döngüler 35 kez tekrarlanış ve en son 72 °C’ de 8
dakika son uzama süresi verilmiştir. PCR ürünleri gerektiğinde 4 °C’de buzdolabında
saklanmış ve daha sonra agaroz jel elektroforez de koşturulmuştur.
RAPD reaksiyonları 0.2 ml’lik ince cidarlı tüplerde gerçekleştirilmiştir. RAPD
reaksiyonları her bir tüp içinde 25 µl’ye ayarlanmıştır. Yapılan ön optimizasyon
çalışmaları sonucunda; 25 µl’lik reaksiyon tüplerine, 2 µl genomik DNA (50ng/µl),
2.5 µl MgCl2, 2.5 µl 10 X Taq polimeraz buffer, 4 µl primer, 2 µl dNTPs, 0.3 µl Taq
polimeraz (1U) ve 11.7 µl distile su olacak şekilde karışım hazırlanmış ve PCR
reaksiyonu gerçekleştirilmiştir (Çizelge 4.57).
4.2.9. Agaroz jel elektroforez
PCR ürünleri % 2’lik Agaroz jel (Sea Kem LE) kullanılarak elektroforezis yapılmıştır.
Agaroz yoğunluğunun artırılması bantların sayısı ve görünebilirliğini artırmıştır. Bu
nedenle PCR ürünleri %’2 lik Agaroz da yürütülmesi daha net görüntülerin elde
edilmesini sağlamıştır. PCR ürünlerini jelde yürütmek için 20 µl DNA örneğine 5 µl
yükleme tamponu ilave edilerek yükleme yapılmıştır.
PCR ürünleri, 100 V’ta 2-3 saat süreyle yürütülmüştür. Koşturma süresinin az ya da çok
fazla tutulması RAPD ürünlerinin görünebilirliğini olumsuz yönde etkilediğinden 2.5
saatlik koşturma görüntüleme açısından tatminkar sonuçlar vermiştir. RAPD profilleri
fotoğraf çekimleri için UVP UV/ White Darkroom marka karanlık oda sistemi
kullanılmıştır.
Agaroz jeller hazırlanırken özellikle yüksek konsantrasyonlarda jellerdeki kırılmaları
önlemek amacıyla gerekli titizlik gösterilmiştir. Jellerin çok iyi şekilde erimeleri
sağlanmış, sıcakken dökülen jeller de daha net görüntüler elde edilmiştir. Jel
hazırlanırken yapılan hata RAPD batlarının görüntülenmesinde sorunlar çıkarmıştır.
84

4. 10. RAPD polimorfizmi ve genetik mesafenin hesaplanması
RAPD-PCR için reaksiyon koşullarının optimize edilmesinden sonra, seçilen bir DNA
örneği ile PCR yapılarak, tritikaleye özel (amplifikasyon veren) primerler belirlenmeye
çalışılmıştır. Kullanılan bu primerlerden hepsi amplifikasyon göstermemiş olup,
primerlerden amplifikasyon gösterenler Çizelge 4.58’de özetlenmiştir. Amplifikasyon
gösteren primerlerin belirlenmesinden sonra, amplifikasyon gösteren primerler
kullanılarak polimorfizm oluşturmak amacıyla tüm tritikale çeşit ve hatlarına ait DNA
örnekleri ile PCR yapılmıştır.
PCR sonucu oluşan RAPD-DNA batlarının jel elektroforez görütüleri dikkate alınarak
değerlendirilmiştir. Bantların okunması ve değerlendirilmesinde, net olarak oluşan
DNA bantları değerlendirmeye alınırken, oluşan silik yada smear’li (kırık) bantlar
değerlendirilme dışı bırakılmıştır. Kullanılan 60 primerden 15 tanesi (% 25) tritikale
genotiplerinde amplifikasyon göstermiştir. Monomorfik ve polimorfik bant üreten 15
primer ile yapılan tritikale çeşit ve hatlarının RAPD-PCR profilleri Şekil 4.3 - 4.17’ de
verilmiştir.
Amplifikasyon gösteren 15 primerden 10 tanesi değişik oranlarda polimorfik (%75)
bant oluştururken, 5 tanesi monomorfik (% 25) bant oluşumu sağlamıştır (Çizelge 4.58).
Çizelge 4.58’de verildiği gibi en fazla polimorfizm gösteren primerler MA 10 ve OPC
08 primerleri olmuştur. Amplifikasyon veren 15 primerden toplam 60 adet RAPD-DNA
bantı oluşmuş, bunların 37 tanesi polimorfiktir. Tritikale çeşit ve hatlarında aplifikasyon
gösteren primerlerin polimorfizm oranı % 61.6 olarak saptanmıştır (Çizelge 4.58).
Primer başına ortalama 4 adet RAPD ürünü oluşurken, 2.46 adet polimorfik RAPD
DNA bandı oluşmuştur. Tritikale çeşit ve hatlarında az sayıda RAPD-DNA bandı
oluşurken, oluşan bantlarda polimorfizm oranının yüksek olduğu görülmektedir.
Çalışmamızda elde edilen primer başına ortalama polimorfik bant sayısı; buğday SSR
primerlerinin ortalama olarak sırasıyla buğdayda 2.6, çavdarda 2.7 ve tritikalede 2.4
polimorfik bant verdiğini bildiren, Kuleung vd (2003) bulgularıyla uyumludur.
85

Çizelge 4.58. RAPD analizinde kullanılan polimorfik ve monomorfik bant veren primerler
Primer Adı
Polimorfik Band Sayısı
Toplam Band sayısı
% Polimorfik Band Sayısı
A00 3 5 60.0 A01 4 6 66.6 A02 4 5 80.0 A03 0 4 - A04 6 7 85.7 A06 2 3 66.6 B 379 0 3 - MA 1 0 2 - MA3 3 4 75.0 MA 8 3 4 75.0 MA9 0 1 - MA 10 5 5 100 OD 8 0 3 - OPC 08 4 4 100 RAPD1 3 4 75.0 Toplam 37 60 61.6
Bulgularımız, Israil’de yabani buğdaylar (Triticum dicoccoides) üzerinde yaptıkları
çalışmada kullanılan 10 adet RAPD primerinin 59 adet elde edilebilir DNA bandı
verdiğini ve oluşan bu bantlardan 48 tanesinin polimorfik (% 81) ve 11 tanesinin
monomorfik (% 19) DNA bantları olduğunu bildiren Fahima vd (1999)’un sonuçlarıyla
benzerlik göstermektedir. Ancak, Freitas vd (2000), Brezilyada yaptıkları çalışmada
ekmeklik buğdayda polimorfizm oranını %33, Szuch vd (2000) makarnalık buğdayda
polimorfizm oranını %28 ve Malgorzata vd (2002), tritikalede yaptıkları çalışmada
RAPD primerleri ile düşük seviyelerde polimorfizm elde ettiklerini, Tams vd (2004),
Avrupa yetiştirilen tritikale çeşitleri arasındaki polimorfizm oranını 0.54 olduğunu
bildirmektedirler.
Çalışmamızda, kullanılan primerlerde polimorfizm oranının yüksek olması, kullanılan
primerlerin daha önceki çalışmalar ışığında seçilmiş olması ve RAPD-PCR koşullarının
iyi optimize edilmiş olmasıyla açıklanabilir. Bulgularımız, RAPD-PCR tekniğinin
tritikalede genetik farklılığı ortaya koymada bir araç olarak kullanılabileceğini ortaya
koymuştur.
86

Kullanılan bazı primerler tritikale çeşit ve hatlarında hiç amplifikasyon göstermemiştir.
Aplifikasyon göstermeyen primerlere örnek olarak MA 7 primeri ile tritikale çeşit ve
hatlarının RAPD-PCR band görüntüsü Şekil 4.2’de verilmiştir.
Şekil 4.2. MA 7 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri Şekil 4.2’ de görüldüğü gibi MA 7 primeri tritikale çeşit ve hatlarında amplifikasyon
göstermediği için tüm DNA’ lar yükleme kuyucuklarından çıkamamış ve genomik
DNA olarak yükleme kuyularında kalmıştır.
A00 primeri, yapılan RAPD PCR sonucunda 5 farklı bant oluşmuş, bant büyüklükleri
350-800 bp arasında değişme göstermiştir (Şekil 4.3). A01 primeri bant büyüklükleri
450-1200 bp arasında değişen 6 farklı bant vermiştir (Şekil 4.4). A 02 primeri 450-1300
bp arasında değişen 5 farklı bant vermiştir (Şekil 4.5). A03 primeri 370, 450, 750 ve
1500 bp uzunluğunda 4 adet monomorfik bant oluşturmuştur (Şekil 4.6). A04 primeri
250-1400 bp arasında değişen 7 farklı bant vermiştir (Şekil 4.7). A06 primeri
amplifikasyon sonucu, 3 farklı bant vermiş bant büyüklükleri 450-750 bp arasında
değişme göstermiştir (Şekil 4.8). MA 1 primeri 400 ve 750 bp uzunluğunda 2 adet
monomorfik bant oluşturmuştur (Şekil 4.9). MA 3 primeri ile 250- 600 bp arasında
değişen 4 farklı DNA bantı oluşmuştur (Şekil 4.10). MA 8 primeri ile yapılan PCR
sonuçlarına göre büyüklükleri 700-1100 arasında değişen DNA bantları oluşmuştur
(Şekil 4.11). MA 9 primeri 1000 bp uzunluğunda 1 adet monomorfik bant
oluşturmuştur (Şekil 4.12). MA 10 primeri uzunlukları 450-2800 bp arasında değişen 5
farklı amplifikasyon ürünü DNA oluşturmuştur (Şekil 4.13). OD 8 primeri 500, 600 ve
800 bp uzunluğunda 3 adet monomorfik bant oluşturmuştur (Şekil 4.14). OPC 08
primeri uzunlukları 850-1300 bp arasında değişen 4 farklı amplifikasyon ürünü DNA
oluşturmuştur (Şekil 4.15). RAPD 1 primeri ile yapılan PCR sonucu 600-1100 bp
87

arasında değişen 4 farklı amplifikasyon ürünü oluşmuştur (Şekil 4.16). B 379 primeri
370, 750 ve 1350 bp uzunluğunda 3 adet monomorfik bant oluşturmuştur (Şekil 4.17).
Bantların büyüklükleri markörler kullanılarak belirlenmiştir. Şekillerden de anlaşılacağı
üzere, tritikalede RAPD-PCR sonucu az sayıda amplifikasyon ürünü DNA bantı
oluşmuştur, ancak oluşan polimorfik bant yüzdesi yüksektir.
Şekil 4.3. A00 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
Şekil 4.4. A01 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
Şekil 4.5. A02 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
88

Şekil 4.6. A03 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
Şekil 4.7. A04 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
Şekil 4.8. A06 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
89

Şekil 4.9. MA 1 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
Şekil 4.10. MA 3 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
Şekil 4.11. MA 8 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
90

Şekil 4.12. MA 9 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
Şekil 4.13. MA 10 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
Şekil 4.14. OD 8 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
91

92
Şekil 4.15. OPC 08 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
Şekil 4.16. RAPD1 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri
Şekil 4.17. B379 primeri ile tritikale çeşit ve hatlarının RAPD-PCR band profilleri Tritikale çeşit ve hatları arasında hesaplanan (POPGENE Version 1.31) genetik uzaklık
matrisi değerleri Çizelge 4.59’da verilmiştir.

93
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25Tat-97 0 ZF 1 0.105 0
0 0
0
ZF 2 0.310 0.203 0ZF 3 0.244 0.163 0.244 0ZF 4 0.203 0.087 0.288 0.143 0ZF 5 0.203 0.087 0.244 0.182 0.069 0ZF 6 0.182 0.105 0.266 0.163 0.124 0.087 0ZF 7 0.182 0.105 0.266 0.163 0.163 0.124 0.034 ZF 8 0.223 0.105 0.266 0.163 0.163 0.124 0.069 0.034 ZF 9 0.223 0.223 0.457 0.288 0.163 0.163 0.143 0.143 0.182 0ZF 10 0.244 0.203 0.381 0.266 0.182 0.182 0.163 0.163 0.163 0.203 0ZF 11 0.163 0.124 0.333 0.182 0.105 0.105 0.087 0.124 0.124 0.163 0.069 0ZF 12 0.183 0.105 0.310 0.163 0.124 0.124 0.105 0.069 0.069 0.143 0.087 0.051 0 ZF 13 0.244 0.124 0.333 0.182 0.105 0.105 0.124 0.087 0.087 0.087 0.143 0.105 0.051 0 ZF 14 0.203 0.124 0.333 0.182 0.182 0.182 0.124 0.087 0.051 0.203 0.143 0.105 0.051 0.105 0ZF 15 0.203 0.124 0.288 0.105 0.143 0.143 0.087 0.051 0.051 0.203 0.143 0.105 0.051 0.105 0.069 0ZF 16 0.223 0.105 0.266 0.163 0.124 0.087 0.034 0.034 0.034 0.143 0.124 0.087 0.069 0.087 0.087 0.051 0 GTAD
0.266 0.143 0.310 0.203 0.163 0.124 0.069 0.069 0.034 0.182 0.163 0.124 0.105 0.124 0.051 0.087 0.034
JGS 0.223 0.105 0.266 0.203 0.124 0.051 0.069 0.069 0.069 0.143 0.124 0.087 0.069 0.087 0.124 0.087 0.034 0.069 0BDMT
0.310 0.182 0.310 0.203 0.163 0.163 0.143 0.105 0.069 0.223 0.203 0.163 0.105 0.124 0.087 0.087 0.105 0.069 0.105 0
CWT 0.266 0.266 0.357 0.288 0.244 0.203 0.182 0.223 0.182 0.223 0.244 0.163 0.182 0.244 0.163 0.203 0.182 0.143 0.182 0.143 0 MT 1 0.203 0.124 0.203 0.143 0.143 0.143 0.124 0.124 0.124 0.203 0.182 0.143 0.124 0.143 0.182 0.143 0.124 0.163 0.124 0.163 0.203 0 LA490 0.288 0.203 0.333 0.310 0.266 0.182 0.203 0.163 0.124 0.244 0.266 0.223 0.163 0.182 0.143 0.182 0.163 0.124 0.124 0.087 0.124 0.223 0 LA380 0.357 0.223 0.406 0.288 0.203 0.244 0.223 0.182 0.143 0.223 0.244 0.203 0.143 0.124 0.124 0.163 0.182 0.143 0.182 0.069 0.182 0.244 0.087 0 LO 51 0.431 0.288 0.431 0.357 0.223 0.310 0.333 0.333 0.288 0.381 0.357 0.310 0.288 0.310 0.266 0.310 0.288 0.244 0.288 0.203 0.288 0.357 0.266 0.203 0
Çizelge 4. 59. Yirmibeş tritikale genotipi arasındaki genetik uzaklık matrisi ( Nei 1972, Nei 1978)

Çizelgede görüldüğü gibi, tüm genotipler arasındaki genetik uzaklık değerleri 0.034 ile
0.457 arasında değişme göstermiştir. Genel ortalama olarak tüm çeşit ve hatlar 0.172
oranında farklı bulunmuştur (Çizelge 4.59). En yüksek genetik uzaklık 0.457 ile (ZF 9-
ZF 2) arasında gözlenirken; bunu 0.431 ile ZF 2 - LO 51, LO 51- Tatlıcak-97 hatları
izlemiş, en düşük genetik uzaklık 0.034 ile ZF 7 - ZF 6, ZF 8 - ZF 7, ZF16 - JGS, ZF
16 - ZF 6, ZF 16 - ZF 7, ZF 16 - ZF 8, GTAD - ZF 8 hatları arasında gerçekleşmiştir.Bu
hatlar getenik olarak birbirine en yakın hatlar olarak nitelendirilebilirler. Tatlıcak-97
tritikale çeşidine genetik olarak ZF 1 (0.105) ile en yakın, ZF 11 (0.163), ZF 6 ve ZF 7
(0.182) ile yakın hatlar olarak görülürken, LO 51 (0.431) hattı ise en uzak olarak
görülmektedir. Tatlıcak-97 çeşidine ZF 4, ZF 5, ZF 14, ZF 15 ve MT 1 (0.203) hatları
genetik olarak eşit uzaklıkta bulunmuştur.
Fahima vd (1999), yabani buğday populasyonları arasındaki genetik uzaklığın 0.017-
0.164 arasında değiştiğini, Freitas vd (2000), Brezilyada ekmeklik buğday çeşitleri
arasındaki genetik benzerliğin 0.27 olduğunu, Szucs vd (2000), yazlık buğday çeşitleri
arasındaki genetik benzerliğin 0.03-0.18 arasında olduğunu, Amiour vd (2002), tritikale
hatları arasındaki genetik uzaklığın 0.50-1.00 arasında değiştiğini, Sun vd (2003), yazlık
buğday çeşitlerinde RAPD verilerine göre hesaplanan genetik benzerliğin 0.64-0.98
arasında değiştiğini, Tams vd (2004), Avrupa tritikale genotipleri arasındaki genetik
uzaklığın ortalama olarak 0.33 olduğunu bildirmektedir.
Çalışmamızda kullanılan tritikale genotipleri arasındaki ortalama genetik uzaklığın
(0.172) düşük olduğu görülmektedir. Bu durum kullanılan hat ve çeşitlerin büyük bir
kısmının aynı coğrafik bölgeden gelmesiyle açıklanabilir. Ancak bazı hatlar arasındaki
yüksek genetik uzaklık değerleri göze çarpmaktadır.
POPGENE (Version 1.31) genetik benzerlik matrisi değerleri kullanılarak MEGA
bilgisayar programında UPGMA’ya göre kümeleme analizi yapılmış ve tritikale çeşit
ve hatlarının genetik benzerlik dendogramı oluşturulmuştur (Şekil 4.18).
94

ZF8
GTAD
ZF14
ZF12
ZF15
ZF6
ZF7
ZF16
JGS
ZF10
ZF11
ZF1
ZF4
ZF5
ZF9
ZF13
ZF3
MT1
CWT
LAD490
BDMT
LAD388
TATLICAK-97
LO51
ZF2
0.000.050.100.15 Şekil 4.18. PCR RAPD yöntemine göre tritikale çeşit ve hatlarının genetik benzerlik
dendogramı Şekil 4.18’de görüldüğü gibi tritikale çeşit ve hatları başlıca 3 grup (kol) oluşturmuştur.
Birinci kolda yer alan ZF 2 hattı diğer tüm çeşit ve hatlarla yaklaşık 0.16 oranında
farklılık göstermektedir. İkinci kolda yer alan LO 51 hattı ile diğer tüm hatlarlarla 0.15
oranında farklı olarak görülmektedir. Diğer tüm çeşit ve hatlar 3. grupta (kol) yer
almıştır. Üçüncü kol içerisinde de farklı küçük kolların olduğu gözlenmektedir. Üçüncü
kolda yer alan genotipler başlıca 2 alt kola ayrılmakta; 3 kolun 1. dalında 1 adet
(Tatlıcak-97), 2. dalında ise 22 adet genotip bulunmaktadır. Bu alt kollar içerisinde de
çeşitli alt alt kollar oluşmuştur. ZF 8-GTAD, ZF 6-ZF 7, ZF 16-JGS hatları birbirine
genetik olarak en yakın hatlar olarak göze çarpmaktadır.
95

Dikkatli optimize edilmiş PCR koşulları ile birlikte RAPD tekniği, diploid buğday
(Joshi ve Nguyen 1993) ve hekzaploid buğday (He vd 1992) türleri arasındaki genetik
farklılığı ortaya koymada başarılı bir şekilde kullanılmıştır. Fahima vd (1995), Sun vd
(1997) buğdayda hastalıklara dayanıklı genlerin belirlenmesinde, Dweiket vd (1994),
Fritz vd (1999) buğday ve tritikalede böceklere dayanıklı genlerin işaretlenmesinde,
Igrejas vd (1999) tritikalede depo proteinlerini belirlemede RAPD-PCR tekniğini
kullanmışlardır. Gupta vd (1999) ekmeklik buğdayda RAPD tekniğini kullanarak
yapılan çalışmalarda başlangıçta düşük seviyelerde polimorfizm elde edildiğini
bildirmektedir. Bu çalışma ile RAPD-PCR tekniği kullanılarak tritikale genotiplerinin
genetik karakterizasyonu yapılmıştır.
Son zamanlarda ekmeklik buğday, tritikale ve çavdar üzerine yapılan çalışmalarda ise
genoma, türe ve çeşide özel RAPD markörleri belirlenmiştir. Çalışmamızda kullanılan
primerlerin daha önceki yapılan çalışmalar ışığında seçilmiş olması ve RAPD-PCR
koşullarının iyi optimize edilmiş olması nedeniyle, kullanılan primerlerden % 61.6
oranında polimorfik DNA bantları elde edilebilmiştir. RAPD-PCR tekniğinin tritikale
çeşit ve hatlarında genetik benzerlik ve farklılığı ortaya çıkarmada kullanılabileceği
sonucuna varılmıştır. Araştırma bulguları tritikale çeşit ve hatları arasında genetik
olarak belli oranda uzaklığın (farklılığı) olduğunu ortaya koymuştur. Çalışma
sonucunda; tritikale çeşit ve hatlarının genetik olarak karakterize edilmesinde RAPD-
PCR tekniğinin kullanılabileceği sonucuna varılmıştır.
Budan sonraki çalışmalarda eğer bu hatlar kullanılarak melezleme yapılacak ise genetik
olarak birbirinden uzak hatların (ZF 2 ve ZF 9, ZF 2 ve LO 51, LO 51 ve Tatlıcak-97
vb.) melezlemede kullanılması, genetik olarak birbirine yakın hatların (ZF 7 ve ZF 6,
ZF 8 ve ZF 7, ZF 16 ve ZF 6, ZF 16 ve ZF 7 vb.) yeniden melezlenmemesi, açılan
generasyonlarda varyasyonu artıracak ve seleksiyonlarda üstün bireylerin seçilme
şansını artıracaktır. Gereksiz melezlemelerden kaçınılacağı için ıslah süresi kısaltacak
ve üstün genotiplerin seçilme şansını artıracaktır. Özellikle istenilen bir özellikte
heterosis gösteren genotiplerin seçiminde bu bilgiler faydalı olacaktır.
96

5. SONUÇ
Bitki ıslahı çalışmalarında bitkisel kaynakların morfolojik ve genetik olarak
tanımlanması bu gen kaynaklarının daha hızlı ve etkin şekilde kullanılmasına olanak
sağlamaktadır. Özellikle çalışılan bitki materyalinin morfolojik olarak benzerliği
kullanılan genotiplerin gerçekte farklı olup olmadığı konusunda kişileri yanılgılara
düşürmektedir. Bu durumda çalışılan genotiplerin değişik yöntemlerle karakterizasyona
tabi tutulmasını gerektirmektedir. Genetik çeşitliliğin belirlenmesinde, morfolojik,
biyokimyasal ve moleküler markörler yaygın olarak kullanılmaktadır. Moleküler
markörler DNA düzeyinde izlenebilen karakterlerdir. Bu karakterler markör olarak
isimlendirilirler ve PCR kullanınılarak suni şartlarda çoğaltılıp bireyler arasındaki DNA
polimorfizmini belirlemede yaygın olarak kullanılırlar.
Bu çalışmada; 25 adet tritikale çeşit/hattı verim ve bazı verim öğeleri yönünden
incelenmiş ve ayrıca RAPD-PCR tekniği ile genetik karakterizasyonu belirlenmeye
çalışılmıştır. Tritikale genotipleri morfolojik (Tarla denemesi) ve genetik (Laboratuvar
çalışması) karakterler olarak incelenmiştir. Ankara, Haymana koşullarında 24 tritikale
ıslah hattı ve kontrol çeşidi (Tatlıcak-97) kullanıldığı tarla çalışmasında, tritikale çeşit
ve hatlarının incelenen verim ve bazı verim öğeleri yönünden (1. yıl başak uzunluğu ve
2. yıl hasat indeksi dışında) önemli farklılıklar belirlenmiştir.
Tarla deneme sonuçlarına göre; çiçeklenme tarihi 27.00-35.87 gün arasında değişmiş
olup, en erkenci hat ZF 12 ve en geççi hat LAD 388’dir. En kısa bitki boyuna sahip
tritikale hattı MT 1 (109.6 cm), en uzun tritikale hattı ise ZF 3 (144.1 cm) olmuştur. En
fazla kardeşlenme gösteren tritikale hattı 5.27 adet/bitki ile ZF 15, en az kardeşlenme
gösteren tritikale hattı ise 4.31 adet/bitki ile JGS olmuştur. En uzun başak uzunluğuna
sahip tritikale hattı CWT (107.9 mm) olup, bunu LAD 388 (107.3 mm) hattı izlemiştir.
En kısa başak uzunluğuna sahip tritikale hattı ise ZF 7 (85.24 mm) olmuştur. Tritikale
hatlarının ortalama başakçık sayısı 19.42 adet/başak (ZF 12) ile 27. 05 adet/başak (ZF
3) arasında değişmiştir. En yüksek başakta tane sayısına sahip tritikale hattı LAD 388,
en düşük başakta tane sayısına sahip tritikale hattı ise ZF 12 olmuştur. Başakta tane
ağırlığı yönünden en yüksek değer ZF 8 tritikale hattında elde edilirken, en düşük
97

başakta tane ağırlığı ise ZF 12 tritikale hattından elde edilmiştir. Bin tane ağırlığı
yönünden en yüksek değer 43.62 g ile ZF 16 tritikale hattından elde edilmiştir. En
düşük değer ise 32.45 g ile LAD 388 tritikale hattından elde edilmiştir. En yüksek tane
verimine sahip tritikale hattı ZF 16 (592.9 kg/da) olup, bu sırasıyla ZF 15 tritikale hattı
ve Tatlıcak-97 tritikale çeşidi izlemiştir. En düşük tane verimine sahip tritikale hattı ise
ZF 12 (475.3 kg/da) olmuştur. Hasat indeksi yönünden tritikale hatları % 29.30 ile
36.37 arasında değişen değerler göstermişlerdir. Tritikale hatlarında protein oranı
ortalama olarak % 11.76 (ZF 6) ile 14.37 (ZF 3) arasında değişen değerler gösterdiği
saptanmıştır.
RAPD-PCR yöntemi kullanılarak genetik benzerlik ya da farklılığın araştırıldığı
labaratuvar çalışmasında; kullanılan primerler ile % 61.6 oranında polimorfik DNA
bantları elde edilmiştir. Polimorfizmin yüksek olması, tritikale çeşit ve hatlarının
karakterizasyonunda RAPD-PCR yönteminin kullanılabileceğini göstermektedir.
RAPD yönteminden elde edilen araştırma sonuçlarına göre tritikale hatlarında genetik
olarak belli oranda genetik farklılıklar saptanmıştır. Bazıları genetik olarak benzer,
bazıları ise genetik olarak farklı bulunmuştur. Tarla deneme sonuçlarına göre
morfolojik karakterler yönünden farklı çıkan hatlar RAPD-PCR yöntemine göre de
DNA düzeyinde farklı çıkmıştır.
Tritikale hatları arasındaki genetik uzaklık değerleri 0.034 ile 0.457 arasında değişme
göstermiştir. En yüksek genetik uzaklık 0.457 (ZF 9) ile ( ZF 2) arasında gözlenirken;
en düşük genetik uzaklık 0. 034 ile ZF 7 ve ZF 6, ZF 7 ve ZF 8, ZF 16 ve JGS, ZF
16 ve ZF 6, ZF 16 ve ZF 7, ZF 16 ve ZF 8, GTAD ve ZF8 hatları arasında
gerçekleşmiştir. Genel ortalama olarak tüm çeşit ve hatlar 0.172 oranında farklı
bulunmuştur.
RAPD PCR sonuçlarına göre hesaplanan genetik benzerlik matrisi değerleri
kullanılarak yapılan kümeleme analizi sonuçlarına göre; tritikale çeşit ve hatları başlıca
3 kolda (grupta) toplanmıştır. Birinci kolda yer alan ZF 2 hattı diğer tüm çeşit ve
hatlarla yaklaşık 0.16 oranında farklılık göstermiştir. LO 51 hattı ile diğer tüm
hatlarlarla 0.15 oranında farklı olarak görülmektedir. ZF 8–GTAD, ZF 6-ZF 7, ZF 16–
98

JGS hatları birbirine genetik olarak en yakın hatlar olarak bulunmuştur. Tatlıcak-97
tritikale çeşidine genetik olarak en yakın hattın ise ZF 1, en uzak hattın ise LO 51
olduğu belirlenmiştir.
Tritikale konusunda RAPD-PCR tekniğinin kullanılmasıyla ilgili (özellikle genetik
karakterizasyon çalışmaları ile ilgili) yerli ve yabancı literatürde yeterli kaynak
bulunamamaktadır. Bu çalışma, ülkemizde tritikale konusunda yapılan ilk moleküler
çalışma olup, bundan sonraki çalışmalara ışık tutacak niteliktedir. Yapılan çalışma ile
Tritikalede kullanılabilecek, DNA izolasyonu yöntemi ve RAPD-PCR koşulları
optimize edilmiştir. Bu bilgiler ileride yapılacak çalışmalarda yararlı olabilecektir. Bu
çalışma sonucuna göre; RAPD tekniğinin tritikale’de DNA polimorfizmini ortaya
çıkarmada ve genetik karakterizasyonda etkili şekilde kullanılabileceği söylenebilir.
Ortaya konan genetik farklılıklar yapılacak ıslah çalışmalarında, melezlemede
kullanılacak ebeveynlerin seçiminde yardımcı olacaktır. Melezlemede kullanılacak ana
ve babanın genetik özelliklerinin bilinmesi, oluşacak döllerin performanslarını tahmin
etmede, gereksiz melezleme ve geri melezlemeleri azaltmada, genetik olarak uniform
bireylerin melezlenmesini önlemede kullanılacaktır. Böylece, uzun süreli
seleksiyonların yapılması engellenecek ve kullanılacak genetik kaynakların
korunmasını sağlayacaktır.
.
99

KAYNAKLAR
Akbay, G. ve Ünver, S. 1986. Tokak 157/37 iki sıralı arpa (Hordeum vulgare L.) çeşidine uygulanan farklı EMS (Ethyl Methane Sulphonate) dozlarının M1 bitkilerinin bazı özellikleri üzerindeki etkileri. A.Ü. Ziraat Fakültesi Yıllığı, Cilt: 38, S: 151-163.
Amiour, N., Bouguennec, A. Marcoz, C. Sourdille, P. Bourgoin, M. Khelifi, D. and Branlard, G. 2002. Diversity of seven glutenin and secalin loci within triticale cultivars grown in Europa. Euphytica, 123; 295- 305.
Anonymous, 2003. http.//faostat.fao.org/faostat
Bağcı, S. A., Tutukçu, E. Çeri, S. ve Ekiz, H. 1999. Triticale: İnsan ve hayvan beslenmesi için geliştirilmiş alternatif bir bitki. Orta Anadolu’da Hububat Tarımının Sorunları ve Çözüm Yolları Sempozyumu. 8-11 Haziran, s: 126-132, Konya.
Baier, A.C., Dias, J.A. and Nedel, J.L. 1980. Triticale research. Annual Wheat Newsletter, 26: 46-47.
Briggle, L. W. 1969. Triticale- A Rewiev. Crop. Sci., Vol; 9, March- April, p.197-202.
Brunel, M.S, Lukaszewski, A. and Whitkus, R. 1999. Development of arm specific RAPD markers for rye chromosome 2r in wheat. Crop. Sci., 39; 1702-1706.
Ceyhan, E. 1999. konya ekolojik şartlarında farklı ekim zamanlarının yemeklik bezelye (Pisum sativum L.) çeşitlerinde verim, ve verim unsurları ile kalite üzerine etkileri. Selçuk Üniversitesi, Fen Bilimleri Enstitüsü, Yüksek Lisans Tezi (basılmamış), 92 s.Konya.
Çengel, A. 2001. Ankara koşullarında yetiştirilen bazı triticale hatlarının verim ve verim öğelerinin incelenmesi. Ankara Üniversitesi, Fen Bilimleri Enstitüsü Yüksek Lisans Tezi (basılmamış).
Çiftçi, İ., Yenice, E. and Eleroğlu, E. 2003. Use of triticale alone and in combination with wheat or maize: effect of diet type and enzyme supplementation on hen performance, egg quality, organ weights, intestinal viscosity and digestive system characteristics. Animal Feed Science and Technology, 105; 149- 161.
Çöplü, N., 2001. Bazı triticale genotiplerinin diallel melezlerinde kantitatif ve sitolojik analizler. T.C. Uludağ Üniversitesi, Fen Bilimleri Enstitüsü Doktora Tezi (basılmamış).
De Bustos, A., Casanova, C. Doler, D. and Jouve, N. 1998. RAPD variation in wild populations of four species of the genus Hordeum (Poaceae). Theor. Appl. Genet, 96; 101.111.
Demir, İ., Aydem, N., Korkut, K.Z. ve Şölen, P. 1979. Türkiye’de tritikale ıslahı çalışmaları. Bitki Islahı Simpozyumu. S:158-165. 22-25 Mayıs 1979, İzmir.
Demir, İ., Korkut, K.Z. Altınbaş, M. Akdemir, H. ve Dutlu, C. 1986. Yazlık tritikale ıslahı çalışmaları. Bitki Islahı Simpozyumu. S:131-138. 15-17 Ekim 1986, İzmir.
100

Devos, K. M., and Gale, M.D. 1992. The use of random amplified polymorphic DNA markers in wheat. Theor. Appl. Genet., 84;567 – 572.
Doxastakis, G., Zafiriadis, I. Irakli, M. Marlani, H. and Tananaki, C. 2002. Lupin, soya and triticale addition to wheat flour doughs and their effect on rheological properties. Food Chemisrty, 77; 219-227.
Düzgüneş, O., Kesici, T. Kavuncu, O. ve Gürbüz, F. 1987. Araştırma ve Deneme Metodları (İstatistik Metodları II) A.Ü. Ziraat Fakültesi Yayın No:1021, Ders Kitabı: 295, Ankara.
Dweiket, I., Ohm, H. Mackenzie, S. Patterson, F. Cambron, S. and Ratcliffe, R. 1994. Association of a DNA marker with hessian fly resistance gene H9 in wheat. Theor. Appl. Genet, 89; 964- 968.
Fahima, T., Grama, A. Korol, A. Turpeinen, T. and Nevo, E. 1995. Idendification of DNA markers linked to novel yellow rust resistance gene introgresed from triticum dicoccoides. Wheat News Lett, 41; 125.
Fahima, T., Sun, G.L. Beharav, A. Krugman, T. Beiles, A. and Nevo, E. 1999. RAPD polymorphism of wild emmer wheat populations, Triticum dicoccoides, in Isreal. Theor. Appl. Genet, 98; 434-447.
Feil, B., and Fossati, D. 1995. Mineral composition of triticale grains as related to grain yield and grain protein. Crop. Sci., 35; 1426-1431.
Fernandez-Figares, I., Marinetto, J. Royo, C. Ramos, J.M. and Garcia del Moral, L.F. 2000. Amino-acid composition and protein and carbohydrate accumulation in the grain of triticale grown under terminal water stress simulated by a senescing agent. Journal of Cereal Science, 32; 249-258.
Frei, O.M., Stuber, C.V. Goodman, M..M. 1986. Use of allozymes as genetic markers for predicting genetic performence in maize single cross hybrids. Crop. Sci, 26;37-42.
Freitas, L.B. de., Jerusalinsky, L. Bonatto, S.L. Salzano, F.M. and de Freitas, L.B. 2000. Extreme homogeneity among Brazilian wheat genotypes determined by RAPD markers. Pesquisa-Agropecuaria-Brasileira, 35; 11, 2255-2260; 24 ref.
Fritz, A.K., Caldwell, S. and Worrall. W.D. 1999. Molecular mapping of russian wheat aphid resistance from triticale accession pı 386156. Crop. Sci, 39; 1707-1710.
Genç, İ.1977. Tahıllarda tane veriminin fizyolojik ve morfolojik esasları. Ç.Ü. Ziraat Fakültesi Yıllığı 8, sayı: 1, Adana.
Genç, İ., Ülger, A.C., Yağbasanlar, T., Kırtok, Y. ve Topal, M. 1988. Çukurova koşullarında triticale, buğday ve arpanın verim ve verim öğeleri üzerinde kıyaslamalı bir araştırma. Ç.Ü. Ziraat Fak. Dergisi, Cilt:3, sayı: 2, s: 1-13, Adana.
Genç, İ., Yağbasanlar, T., Ülger, A. C. ve Kırtok, Y. 1987. Çukurova koşullarında triticalenin verim ve verim öğeleri üzerinde bir araştırma. Türkiye Tahıl Simpozyumu. 6-9 Ekim, s: 103- 114, Bursa.
Gil, Z. 1996. Chemical composition and biochemical properties of spring and winter triticales. Field Crops Abstaracts. 1996. Vol. 49; No. 6, page 486.
101

Gupta, P.K., Varshney, R.K. Sharma, P.C. and Ramesh, B. 1999. Reviev. Moleküler markers and their application in wheat breedind. Plant Breeding, 118; 369-390.
He, S., Ohm, H. Mackenzie, S. 1992. Detection of DNA sequences polymorphism among wheat varieties. Theor. Appl. Genet, 84; 573-578.
Hillis, D.M., and Moritz, C. 1990. Molecular systematics, Sinaur Associates, Inc. Publisher, p.217-220. Sunderland.
Horlein, A,. and Valentine, J. 1995. Triticale (x Triticosecale). In. Williams J.T. (ed) Cereals and pseudocereals. Chapman and Hall, New York, pp. 187-221.
Igrejas, G., Guedes-Pinto, H. Carnide, V. and Branland, G. 1999. Seed storage protein diversity in triticale varieties commonly grown in Portagual. Plant Breeding, 188; 303-306.
Joshi, C.P. and Nguyen, H.T. 1993. Application of random amplified polymorphic DNA technigues for the detection of polymorphism among wild and cultivated tetraploid wheat. Genome, 36; 602- 609.
Kaçar, B. 1972. Bitki ve Toprağın Kimyasal Analizleri. II. Bitki Analizleri. A.Ü. Ziraat Fakületesi yayınları.453. Ankara, s. 25-27.
Kadester, I.E. 1960. Zirai Kimya Tatbikatı Birinci Kitap. Yem Analizleri. A.Ü. Ziraat Fakültesi yay.113. Ders Kitabı: 40. s.51-70. Ankara.
Kınacı, E. 1998. Zor koşulların bitkisi tritikalenin yem olarak değeri ve geleceği. Hasat Dergisi, Yıl:14, Sayı:157 s:18-19.
Koc, J., Szymczyk, S. Domska, D. Wojtkowiak, K. and Wojnowskai, T. 2000. Protein amino acid and composition of spring triticale grain grown at different nitrogen fertilizer rates. Field Crops Abstracts, Vol; 53. No. 10 pp; 928.
Kuleung, C., Baenziger, P.S. and Dweikat, I. 2003. Transferability of SSR markers among wheat, rye and triticale. Theor Appl Genet; 108, 1147-1150.
Kumar, R. 1989. The tecnigue of polymerase chain reaction. A journal of Methods in Cell and Molecular Biology, 1; 133-152.
Kumar, S., Tamura, K. Jakobsen, I.B. and Nei, M. 2001. MEGA2: Molecular evolutionary genetics analysis software, Bioinformatics (submitted).
Kün, E. 1996. Tahıllar- I. A.Ü. Ziraat Fakültesi Yayın No:1451, Ders Kitabı:431, Ankara.
Li, Y.C., Fahima, T. Beiles, A. Korol, A.B. and Nevo, E. 1999. Microclimatic stress and adaptive DNA differentiation in wild emmer wheat, Triticum dicoccoides. Theor. Appl. Genet, 98; 873-883.
Lukaszewski, A.J., and Gustafson, J.P.D. 1987. Cytogenetics of triticale. Plant Breed. Rev, 5; 41-93.
Ma, Z-Q., and Sorrells, M.E. 1995. Genetic analysis of fertility restoration in whaet. Using restriction fragment lenght polymorphism. Crop Sci., 35;1137-1143.
102

Malgorzata, G., Wisniewska, I. and Rafalski, A. 2002. Semi-specific PCR evaluation of diversity among cultivars wheat and triticale. Cellular & Moleculer Biology Letters. Volume 7, pp;577-582.
Maniatis, T., Fritsch, E. and Sambrook, J. 1982. Molecular Cloning: A Laboratory Manual: Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
Maurice, D.V. 1989. Chemical composition and nutritive value of triticale (Florida 201) for broiler chickens. Applied Agricultural Research, 4; 243-247.
Mergoum, M., Ryan, J. Shroyer, J. P. and Monem, M. A. 1992. Potential for adapting triticale in Morocco. Journal of Natural Resources and Life Sciences Education, 21: 2, 137-141.
Milovanovic, M. 1993. Investigation of yield and technological tips of grain of intergenus hybrids triticale (Triticosecale wittmack). Rewiev of Research Work at the Faculty of Agriculture, Belgrade. 38: 2, 71-82.
Müntzing, A. 1989. Triticale results and problems. Advances in Plant Breeding. Supplement to Journal of Plant Breeding. Verlag Paul Parey. Berlin und Hamburg. 103 p.
Myburg, A.A., Botha, A.M. Wingfield, B.D. and Wilding, W.J.M. 1997. Idendification and genetic distance analysis of wheat cultivars using RAPD finger printing. Cereal Res. Commun, 25; 875-882.
Nei, N. 1972. Genetic distance between populations. Am.Nat.106: 283-292.
Nei, N. 1978. Estimation of average heterozygosity and genetic distance from a small number of induviduals. Genetics, 89; 583-590.
Palumbi, S., Martin A. Romana, S. McMillan, W.D. Stice, L. and Grabowski, G. 1991. The simple tool’s guide to PCR . University of Hawaii, sf.12.
Pammi, S., Schertz, K. Xu, G. Hart, G. and Mullet, J.E. 1994. Random amplified polimorfic DNA markers in sorgum. Theor. Appl. Genet, 89; 80-88.
Pancholi, N. 1995. Aspect of tissue culture in relation to banana improvemnet and germplasm conservation. The University of Reading, Department of Agricultural Botany, School of Plant Sciences, 301 pp.
Penner, G.A. 1996. RAPD analysis of plant genomes, In: Methods of genome analysis in plants. CRC Press. Inc, Boco Raton Florida, p. 251-268.
Perez, T.G., Alberto, E.L. Ribotta, P.D. Aguirre, A. Rubiolo, O. J. and Anon, M. 2003. Use of triticale flours in craker-making. European Food Research and Tegnology. Springer-Verlag.
Reiter, S.R., Young, M. and Scolnik, P.A. 1993. Genetic linkage of the arabidopsis genome: Methods for Mapping with Recombinant Inbreds and RAPDs. Methods in Arabidopsis Research, World Scientific Publishing, Singapore.
Rodriges., M.A, Countinho, J. and Martins, F. 2002. Efficacy and limitations of triticale as a nitgogen catch crop in mediterranean environments. European Journal of Agronomy. 17; 155-160.
103

Sachs, E., Bivour, W. and Krumbiegel, D. 1999. Damege to winter barley (Hordeum vulgare L.), winter wheat (Triticum aestivum L.) and winter triticale (Triticosecale Wittm.) caused by winter kill in Güterfelde/ Branderburg. 1999/97. Field Crops Abstaracts 1999. Vol.52, No.9, page 890.
Sapra, V.T., Sharma, G.C. Hughes, J. and Bradford, R.R. 1973. Triticale, a wheat – rye hybrid. J.Tenn. Acad. Sci. 48: 59-61.
Sencer, Ö., Gökmen, S. ve Sakin, M. A. 1997. Tokat Artova koşullarında triticale, buğday ve çavdarın verim ve verim unsurları üzerine bir araştırma. Türkiye II: Tarla Bitkileri Kongresi, 22-25 Eylül, s: 113-117, Samsun.
Sivolap, Yu. M., Chebotar, S.V. Topchieva, E.A. Korzun, V.N. and Totskiy, V.N. 1999. RAPD and SSRP analyses of molecular-genetic polymorphism in Triticum aestivum L. cultivars. Genetika-Moskova, 35;12, 1665-1673; 24 ref.
Skroch, P. W., Tivang, J. and Nienhuis, J. 1992. Analysis of genetic relationship using RAPD marker data. Applification of RAPD Technology to Plant Breeding. Joint Plant Breeding Symposia Series, Minneapolis, Minnesota.
Smith R.L., Schweder, M.E. and Barnett, R.D. 1994. Idendification of glutein alleles in wheat and triticale using PCR-generated DNA markers. Crop. Sci., 34; 1373-1378.
Stallknecht, G.F., Gilbertson, K.M. and Ranney, J.E. 1996. Alternative wheat cereals as food grains. Einkorn, emmer, spelt, kamut, and triticale. P. 156-170. In. J. Janick (ed.) Progress in new crops. ASHS Pressi Alexandria, VA.
Sun, G., Bond, M. Nas, H. Martin, R. and Dong, Z. 2003. RAPD polymorphism in spring wheat cultivars and lines with different level of Fusarium resistance. Theor Appl Genet, 106;1059-1067.
Szigat, G., and Müller, H.W. 1975. Results from testing and international triticale assortment, Field Crops Abstract, Vol;28, No: 8, Abst. No: 4339.
Szucs, P., Juhasz, A. Karsai, I. Lang, L.Veisz, O. and Bedo, Z. 2000. Use of molecular markers for studying genetic diversity in durum wheat (Triticum durum Desf.). Journal-of-Genetics-and-Breeding, 54; 1, 25-33; 44 ref.
Şençiçek, A.G.E. 2000. RARD markerlerini kullanarak Türk susam (Sesamum indicum L.) populasyonlarında genetic uzaklıkların analizi. T.C. Akdeniz Üniversitesi, Fen Bilimleri Enstitüsü Doktora Tezi. 102 s.(Basılmamış). Antalya.
Tams, S.H., Bauner, E., Oetter, G. ve Melchinger, A.E. 2004. Genetic diversity in European winter triticale determined witk SSR markers and coancestry coefficient. Theor Appl Genet, 108:1385-1391.
Taşyürek, T., Demir. M. ve Gökmen, S. 1999a. Sivas yöresinde triticalenin azotlu gübre isteği. Orta Anadolu’da Hububat Tarımının Sorunları ve Çözüm Yolları Sempozyumu. 8-11 Haziran, s: 259-265, Konya.
Taşyürek, T., Gökmen, S., Temirkaynak, V. ve Sakin, M. 1999b. Sivas- Şarkışla koşullarında buğday, arpa ve triticalenin verim ve verim unsurları üzerine bir araştırma. Orta Anadolu’da Hububat Tarımının Sorunları ve Çözüm Yolları Sempozyumu. 8-11 Haziran, s: 616-620, Konya.
104

Tosun, O. ve Yurtman, N. 1973. Ekmeklik buğdaylarda verime etkili başlıca morfolojik ve fizyolojik karakterler arasındaki ilişkiler. A.Ü. Ziraat Fakültesi Yıllığı, 23.
Ülger, A.C., Yağbasanlar, T. ve Genç, İ. 1989. Çukurova koşullarında seçilen yüksek verimli tritikale (Triticosecale Wittm.) hatlarının önemli tarımsal karakterleri üzerinde bir araştırma.TÜBİTAK Doğa Bilim Dergisi, Cilt:13, Sayı:3,10,13 s.
Ünver, S. 1999. Bazı tritikale hatlarında verim ve verim öğelerinin incelenmesi. Tarla Bitkileri Merkez Araştırma Enstitüsü Dergisi, (8), 1-2 S: 82-92, Ankara.
Ünver, S.1995. Buğdayda tohum iriliğinin verim ve verim öğeleri üzerine etkisi. TARM Yayın No: 1, 37 s. Ankara.
Varughese, G., Pfeıffer, W.H. and Pene, R.R.J. 1996. Specialty grains, Triticale. A succesful alternative crop. Cereal Food World. Vol. 41, No. 6, 1 pp. 474-482 part 2pp 635-645.
Weeden, N.F., Timmerman, G.M. Hemmat, M. Kneen, B.E. and Lodhi, M.A. 1992. Inheritance and Reliability of RAPD markers, Applification of RAPD Technology to Plant Breeding. Joint Plant Breeding Symposia Series, Minneapolis, Minnesota.
Weising, K.H., Nybom, W. K. and Meyer, W.1995. DNA fingerprinting In Plants and Fungi, CRC Press Inc., Boca Raton, Florida, p. 1-200.
Wesolowski, M. and Gregorczyk, K. 2000. Yield of winter triticale under different crops sequences and plant protection systems. Field Crops Abstracts. 2000. Vol. 10. page. 928.
Williams, J.G.K., Kubelik, A.R. Livak, K.J. Rafalski, J.A. and Tingery, S.V. 1990. DNA polymorhism amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 18:6531-6535.
Yağbasanlar, T., Genç, İ. ve Ülger, A.C. 1988. Çukurova koşullarında tritikalede farklı azot dozu ve tohumluk miktarının verim ve verim unsurlarına etkisi. Ç.Ü. Ziraat Fak. Dergisi, Cilt:3, sayı: 2 s: 23-35, Adana.
Yağbasanlar, T., Ülger, A.C. ve Genç, İ. 1989. Çukurova koşullarında bazı yabancı tritikale çeşitlerinin verim ve verim öğeleri üzerinde bir araştırma. TÜBİTAK Doğa Bilim Dergisi, Cilt:13, Sayı: 3, 1.9, 13 s.
Yağbasanlar, T., Çölkesen, M. ve Genç, İ. 1990. Çukurova ve Şanlıurfa koşullarında bazı tritikale hatlarının verim ve verim unsurları üzerinde bir araştırma. Ç.Ü. Ziraat Fak. Dergisi, 5, (2): 125-140. Adana.
Yağbasanlar, T. 1991. Triticale (X Triticosecale Wittmack)’de doğal yabancı döllenme ile tohum tutma oranı. Ç.Ü. Ziraat Fak. Dergisi, 6, (1):127-136.
Yağbasanlar, T., Genç, İ. Toklu, F. ve Özkan, H. 1999. Çukurova koşullarında Fahad-1 triticale hattına uygun yetiştirme tekniklerinin belirlenmesi üzerine bir araştırma. Türkiye 3. Tarla Bitkileri Kongresi, 15-18 Kasım. Cilt-1, s: 169-173, Adana.
Yıldırım, A. ve Kandemir, N. 2001. Genetik Markörler ve Analiz Metodları. BİTKİ BİYOTEKNOLOJİSİ. Genetik Mühendisliği ve Uygulamaları. (Ed.) Özcan, S. Gürel, E. Babaoğlu, M. S.Ü. Vakfı yayınları, s, 456. Konya.
105

ÖZGEÇMİŞ 1969 yılında Ankara’da doğdu. İlk, Orta ve Lise öğrenimini Ankara’da tamamladı.
1988 yılında Ankara Üniversitesi, Ziraat Fakültesi, Tarla Bitkileri Bölümünü kazanarak
Üniversite öğretimine başladı ve 1992 yılında aynı bölümden mezun oldu. 1993 yılında
A. Ü. Fen Bilimleri Enstitüsü, Tarla Bitkileri Anabilim dalında yüksek lisans yapmaya
başladı. Yüksek lisans eğitimine devam ederken, 1994 yılında Milli Eğitim Bakanlığı
Yurt dışı bursluluk sınavını kazanarak ABD’ye gitti 1995-1997 yıllarında University of
Nebraska- Lincoln’de dil ve Yüksek Lisans eğitimi yaptı.
1998 yılı Şubat ayında yurda dönerek, Hatay Mustafa Kemal Üniversitesi, Ziraat
Fakültesi, Tarla Bitkileri Bölümünde Araştırma Görevlisi olarak göreve başladı. Eylül
1999’da YÖK kanununun 35. maddesi gereğince Doktora eğitimi yapmak üzere A.Ü.
Fen Bilimleri Enstitüsü, Tarla Bitkileri Anabilim dalına atandı. Halen bu bölümde
doktora eğitimine devam etmektedir. Evli ve bir çocuk babasıdır.
106