nucleotide sequence, functional characterization and...
TRANSCRIPT

Molecular Microbiology (2003)
47
(6) 1545ndash1562
copy 2003 Blackwell Publishing Ltd
Blackwell Science LtdOxford UKMMIMolecular Microbiology0950-382XBlackwell Publishing Ltd 200347
615451562
Original Article
pFKN a virulence plasmid in P syringae pv maculicolaL Rohmer S Kjemtrup P Marchesini and J L Dangl
Accepted 28 November 2002 For correspondence E-maildanglemailuncedu Tel (
+
1) 919 962 5624 Fax (
+
1) 919 9621625
dagger
Present address Paradigm Genetics Inc 104 Alexander DrPO Box 14528 RTP NC 27709-4528 USA
Nucleotide sequence functional characterization and evolution of pFKN a virulence plasmid in
Pseudomonas syringae
pathovar
maculicola
Laurence Rohmer
1
Susanne Kjemtrup
1dagger
Patrizia Marchesini
1dagger
and Jeffery L Dangl
1ndash3
1
Department of Biology
2
Curriculum in Genetics and
3
Department of Microbiology and Immunology Coker Hall 108 CB3280 University of North Carolina at Chapel Hill Chapel Hill NC 27599-3280 USA
Summary
Pseudomonas syringae
pv
maculicola
strain M6 (
Psm
M6) carries the
avrRpm1
gene encoding a type IIIeffector on a 40 kb plasmid pFKN We hypothesizedthat this plasmid might carry additional genesrequired for pathogenesis on plants We report thesequence and features of pFKN In addition to
avrRpm1
pFKN carries an allele of another type IIIeffector termed
avrPphE
and a gene of unknownfunction (ORF8) expression of which is induced
inplanta
suggesting a role in the plantndashpathogen inter-action The region of pFKN carrying
avrRpm1
avrPphE
and ORF8 exhibits several features of patho-genicity islands (PAIs) Curing of pFKN (creating
Psm
M6C) caused a significant reduction in virulence on
Arabidopsis
leaves However complementation stud-ies using
Psm
M6C demonstrated an obviousvirulence function only for
avrRpm1
pFKN can inte-grate and excise from the chromosome of
Psm
M6 atlow frequency via homologous recombinationbetween identical sequence segments located on thechromosome and on pFKN These segments are partof two nearly identical transposons carrying
avrPphE
The
avrPphE
transposon was also detected in otherstrains of
P s
pv
maculicola
and in
P s tomato
strainDC3000 The
avrPphE
transposon was found insertedat different loci in different strains The analysis ofsequences surrounding the
avrPphE
transposoninsertion site in the chromosome of
Psm
M6 indicatesthat pFKN integrates into a PAI that encodes type III
effectors The integration of pFKN into this chromo-somal region may therefore be seen as an evolution-ary process determining the formation of a new PAIin the chromosome of
Psm
M6
Introduction
Pathogens require an arsenal of pathogenicity factors toinfect a host Genes important for pathogenicity are oftengrouped in regions of the chromosome or on plasmidsThese regions are referred to as pathogenicity islands(PAIs) (for a review see Hacker and Kaper 2000) PAIstypically carry virulence genes and sequences relatedto DNA mobility such as genes encoding integrases ortransposases and insertion sequences (IS) It is very likelythat PAIs are acquired through horizontal transfer (Hackerand Kaper 1999) This is suggested by the fact that theirG
+
C content often differs from that of the core genomeMoreover in many cases PAIs are flanked by tRNA genesSome tRNA loci are known to be integration targets forDNA of bacteriophages or plasmids (Hou 1999) The VPI(
Vibrio
pathogenicity island) for example results from theintegration of the bacteriophage VPI
F
(Karaolis
et al
1999) into the chromosome of
Vibrio cholerae
In othercases the PAI seems to have arisen from the integrationof a plasmid Indeed Rajakumar
et al
(1997) proposedthat a 99 kb chromosomal element in
Shigella flexneri
2aYSH6000 carrying a multiantibiotic resistance locus aroseafter the integration of an NR1-like plasmid
The mosaic-like structure of numerous PAIs suggeststhat they are generated by a multistep process This pro-cess may involve various mobile elements and multiplegenomic rearrangements For example the SPI2 (
Salmo-nella
PAI 2) PAI contains separate 25 kb and 145 kbregions that differ from each other in their G
+
C contenttheir codon usage the type of functions they encode andtheir respective distribution in other strains of
Salmonella
(Hensel
et al
1999) Similarly the comparison of fivelocus of enterocyte effacement (LEE) PAIs from threerabbit-specific strains of enteropathogenic
Escherichiacoli
(REPEC 8339 84110-1 and RDEC-1) and humanenteropathogenic or enterohaemorrhagic
E coli
strains(E234869 and EDL933) shows that the five PAIs origi-
1546
L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd
Molecular Microbiology
47
1545ndash1562
nated from the same source (Tauschek
et al
2002) How-ever each of them has evolved and diverged from thePAIs of the other strains through DNA insertions andpotential deletions (Tauschek
et al
2002) On someoccasions a PAI may excise from the chromosome Themechanism of excision is sometimes the same as themechanism of integration For example an integraseencoded on the HPI (high pathogenicity island) of
Yersiniapseudotuberculosis
and
Yersinia pestis
is responsible forthe integration of this PAI into its attachment site on thechromosome (Rakin
et al
2001) Rakin and coworkersshowed that when this integrase is expressed the HPIcould be excised
In the plant pathogen
Pseudomonas syringae
severalPAIs have been identified (Alarcon-Chaidez
et al
1999Jackson
et al
1999 2000 Alfano
et al
2000 Lu
et al
2002) They carry genes encoding the type III secretionsystem apparatus (called the
hrp
cluster) type III effectorproteins that serve as virulence factors or proteins catal-ysing the production of toxins Thus PAIs carry genesrepresenting the major components of
P syringae
patho-genicity The
hrp
cluster is necessary for pathogenicity(Lindgren
et al
1986 1988) and encodes a type III pilus(Jin and He 2001)
The
P syringae
type III secretion system delivers typeIII effector proteins to the plant cell (Cornelis and VanGijsegem 2000 Jin and He 2001 Li
et al
2002)Although some type III effectors have demonstrable rolesin virulence very little is known about their host targets(Nimchuk
et al
2001) Target processes affected by
Psyringae
type III effectors were described recently (Mackey
et al
2002 Van der Hoorn
et al
2002) Some type IIIeffectors are recognized inside the plant cell by the plant
R
gene system and these trigger a disease resistance-associated programmed cell death known as the hyper-sensitive response (HR) (Dangl and Jones 2001)Because the recognition of some type III effector proteinscauses the plant to be resistant to a particular pathogenstrain most of the genes encoding
P syringae
type IIIeffectors were originally named avirulence (
avr
) genesThe
P syringae hrp
PAI also encodes the HrpL alterna-tive sigma factor This factor regulates the expression ofboth the genes encoding the type III delivery pilus andthose encoding type III effectors (Xiao
et al
1994) Thepromoters of these genes contain a consensus
hrp
box(Innes
et al
1993) Hence many genes encoding puta-tive type III effectors have been identified recently usingsequence analysis methods on the whole genome of the
Pst
DC3000 strain and sequences of DNA fragments fromother pathovars (Jackson
et al
1999 Alfano
et al
2000Arnold
et al
2001 Boch
et al
2002 Fouts
et al
2002Petnicki-Ocwieja
et al
2002)The type III effector avrRpm1 (Ritter and Dangl 1995)
contributes to virulence on
Arabidopsis
plants that cannot
recognize it (host genotype
rpm1
) It is encoded on aplasmid in several
P syringae
pv
maculicola
(
Psm
)strains hereafter named pFKN Several
P syringae
plas-mids have been partially or fully characterized They oftencarry genes encoding virulence factors or functionsenabling the bacteria to survive in given environments (fora review see Vivian
et al
2001) We describe the analy-sis of the pFKN sequence and suggest that it carries atleast one PAI We investigated the mechanism by whichthe plasmid integrates and excises into and out of thechromosome of
Psm
M6 as well as the chromosomalregion involved in the process We also found that a pieceof the pFKN mosaic structure is a transposon carrying anallele of a second type III effector
avrPphE
Finally wedescribe the genomic context of this transposon in several
P syringae
strains Our work provides insights into theevolutionary process leading to the formation of new PAIsin
P syringae
and how they may be transferred fromstrain to strain
Results
Sequencing of pFKN
The sequence of pFKN was determined to an average of
ordf
31
yen
coverage using a shotgun approach The completenucleotide sequence of pFKN consists of 39 554 bp Itsoverall G
+
C content is 5353 Twenty-nine putative openreading frames (ORFs) were identified representing757 of the sequence To assign possible functions tothese ORFs we compared the sequence of each withavailable databases using
BLASTN
and
BLASTX
Twenty-seven ORFs had homology with genes present in thedatabases We searched the translated products of eachORF for known functional domains and motifs and couldassign a putative function to most of them (Table 1)
We organized the ORFs into various functional groups(Fig 1) The plasmid encodes two type III effectors(
avrRpm1
and an allele of
avrPphE
) Five non-contiguousregions on the plasmid may be mobile elements or rem-nants thereof Three other groups of genes were definedaccording to the putative function that they encode
Plasmid replication and maintenance
ORF1 shares highhomology with genes that encode the replication proteinsof plasmids belonging to the PT23A family in the P syrin-gae species (Sesma et al 1998) Homologous replicationproteins are found in other genera such as the replicationproteins from the ColE2 family in E coli (Sesma et al1998) The product of ORF10 shares similarity with ParAa protein involved in partitioning working in concert with aprotein encoded by ParB No ParB gene was found onpFKN Therefore it is not clear how the plasmid copies(around five per cell data not shown) are distributed effi-
pFKN a virulence plasmid in
P syringae
pv
maculicola 1547
copy 2003 Blackwell Publishing Ltd
Molecular Microbiology
47
1545ndash1562
ciently after cell division ORF28 may encode a proteininvolved in the transfer of pFKN The localization of theprotein encoded by ORF28 is predicted to be in the bac-terial periplasmic space It has three domains The N-terminus shows high homologies to VirB1 of Brucella suis(a mammalian pathogen) named after its homology toVirB1 of Agrobacterium tumefaciens The central regionshows homology to VirB4 of the same organism The C-terminus shares homology with different DNA-binding pro-teins (DNA nickase primase) However no other gene onthe plasmid seems to encode a function related to trans-fer It is highly unlikely that this unique protein would allow
transfer of pFKN Therefore ORF28 encodes an unknownfunction or could represent the remnant of a transfer sys-tem encoded on the plasmid before genomic rearrange-ments leading to its loss
Type III effectors
The promoter regions of two genes onthe plasmid contain an
hrp
box suggesting that they arepart of the type III regulon ORF2 is 80 identical to the
avrPphE
allele carried by
P s syringae
(
Psy
) (Alfanoet al 2000) The avrPphE gene had originally beenidentified in P s phaseolicola (Pph) and encodes aprotein delivered to the host cell (Mansfield et al 1994)
Table 1 ORFs and non-coding elements identified in Ps pv maculicola pFKN
a We identified 4 DNA fragments that were conserved between pFKN and the genome of Pst DC3000 (gt85 nucleotide identity) The ORFs thathave no homologues in the genome of Pst DC3000 are noted as lsquononersquo the fragments are numbered 1ndash4b The expression of each gene carried by the plasmid was assessed in KB and MM (hrp-inducing media) by RT-PCR and by using GFP-translationalfusions in planta for 7 of them (not shown) When the gene is expressed in one given condition the table exhibits lsquo+rsquo A lack of expression is markedby lsquondashrsquo NA not available
1548 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Since then several alleles have been identified in otherpathovars of P syringae (Fouts et al 2002 Guttmanet al 2002) They all contain an hrp box in ORF2 ofpFKN this is found 51 bp upstream of the ATG at nt 1561as 5cent-GGAACTGA-N13-CGACATA-3cent (Innes et al1993) Based on the homologies and the presence ofthe hrp box in the promoter we hypothesized that theAvrPphE-like protein encoded by ORF2 is also a type IIIeffector (see below) AvrRpm1 (numbered ORF5) is iden-tical to that defined in the strain Psm M2 (Dangl et al1992)
Mobile element-related genes ORF3 (TnpA) encodes aputative truncated transposase Importantly avrPphE andTnpA are located between nearly identical invertedrepeats of 66 bp (Fig 1) thus defining a probable trans-posable element remnant The sequences surroundingavrRpm1 (ORF4 and ORF6) are homologous to asequence identified in P s pisi by Arnold et al (2001)with 96ndash97 nucleotide identity The DNA fragment car-rying these sequences in P s pisi may also be a trans-poson or a remnant It is therefore likely that avrRpm1 isalso located on the remnant of a transposon
ORF13 and ORF14 are related to ORFs found in theprophage 933 L on the LEE of the enterohaemorrhagic Ecoli O157H7 L0015 and L0014 respectively These twoORFs are thought to belong to an IS element from theIS66 family primarily identified in Sinorhizobium melilotithat includes IS elements from animal and plant patho-gens These two genes are usually associated with a thirdone L0013 in the LEE that is absent in pFKN (the
sequence upstream of ORF14 does not contain any ORFand does not show homology to L0013) Moreover the 5centend of ORF14 is also missing Therefore the sequenceencoding ORF13 and ORF14 may represent the remnantof an IS element belonging to the IS66 family not previ-ously identified in P syringae
Transcription factors Four pFKN genes share homologywith transcriptional regulators Some of these may func-tion during interactions with eukaryotic hosts For exam-ple the closest homologue to ORF24 is a putativetranscriptional factor from V cholerae ORF24 is alsorelated to putative proteins of unknown function (along thewhole length of the protein) found in organisms as diverseas E coli (50 identity67 homology) Caenorhabditiselegans (4157) Homo sapiens (4258) Droso-phila melanogaster (4058) and Arabidopsis thaliana(2942) A domain characteristic of FAD-dependentoxidoreductase (in Pfam) is detected in ORF24 There-fore the function of the translation product of this ORFmay require electron transfer through FAD
The protein encoded by ORF27 is a member of thesmall RfaH family RfaH controls the transcription of E coliand Salmonella operons that direct the synthesis assem-bly and export of the lipopolysaccharide core exopolysac-charide F-conjugation pilus and haemolysin toxin (for areview see Santangelo and Roberts 2002) RfaH isa specific regulator of transcript elongation its lossincreases transcription polarity in these operons withoutaffecting initiation from the operon promoters In additionin one case the RfaH protein controls the transcription of
Fig 1 Graphical map of pFKN and its 29 iden-tified ORFs The proposed direction of tran-scription is indicated by arrows Red boxes indicate genes encoding for type III effectors A black arrow line at the 5cent end of any genes indicates a possible hrp box Genes related to those of mobile elements are represented by green boxes The avrPphE transposon and the IS801 homologue are framed Transposon repeats surrounding the avrPphE transposon are marked by green lines The avrRpm1 gene is surrounded by repeats similar to the repeats of the transposon Tn501 these repeats are also marked by green lines Purple boxes indi-cate genes with replication and partition func-tions Genes encoding for putative transporters are indicated by blue boxes The putative tran-scriptional regulators are indicated by orange boxes The grey boxes represent genes that do not fall into any of the categories listed The number of each ORF is noted in bold on the outside of the circle
pFKN a virulence plasmid in P syringae pv maculicola 1549
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
a single gene encoding an outer membrane haeminreceptor ChuA in pathogenic E coli as well as othergenes involved in pathogenicity of the strain (Nagy et al2001 2002) The promoter of the genes or operons con-trolled by RfaH contains a conserved sequence calledJUMPStart (Hobbs and Reeves 1994) No JUMPStartsequence was detected on pFKN
Transporters ORF18 encodes a probable transmem-brane protein that is highly homologous to sugar trans-porters One is the product of mae1 a gene of the yeastSchizosaccharomyces pombe encoding a proteininvolved in the uptake of L-malate succinate and malonicacid The product of ORF18 may provide the bacteria witha fitness advantage in certain conditions
Other putative functions encoded on the plasmid Othergenes may encode proteins providing an advantage to thebacteria during the infection process For example theproduct of ORF12 displays all the characteristics ofmethyl-accepting chemotaxis proteins The genes encod-ing the flagellar unit and the chemotaxis functions areoften organized in clusters which is not the case onpFKN In this particular case the flagellar unit may beprovided in trans
pFKN is composed of different fragments that indicate a multistep process leading to its formation
The G+C content of the pFKN ORFs is highly variablealong the plasmid (Table 2) Yet several contiguous genesets on the plasmid have similar G+C contents This prob-ably indicates that these genes were acquired togetherfrom an original source The G+C content might allow usto track the multistep acquisition process resulting inpFKN For example ORFs 7 8 and 10 share nearly iden-tical G+C content (505) but are separated by an ISelement with a significantly different G+C content (603)To determine the distribution of pFKN ORFs in otherpathovars and how this distribution is related to G+Ccontent by ORF group we performed sequence compar-isons with the nucleotide sequence of the completegenomes of P syringae pv tomato DC3000 (httptigr-blasttigrorgufmg) and P syringae pv syringae B728a(httpwwwjgidoegovJGI_microbialhtmlindexhtml)DNA fragments carrying from two to seven contiguouspFKN ORFs were detected in both genome sequences(85ndash100 identity) The four DNA fragments from P stomato DC3000 carrying linked pFKN ORFs were num-bered and added to Table 2 Of these Pst DC3000 frag-ment 2 was also detected in Psy B728a (98 identical topFKN) In addition sequences homologous to repA andORF13 were also detected in this Psy B728a Table 2shows that ORFs belonging to the same contiguous DNA
fragment have a similar G+C content For example genesbelonging to fragment 4 have a relatively high G+C con-tent Genes belonging to fragments 1 and 2 have similarG+C contents and genes in fragment 3 exhibit a lowerG+C content Thus the different fragments constitutingpFKN were probably acquired by the three genomes indifferent steps followed by subsequent sequence dele-tions andor rearrangements with respect to Psm M6 asthe entire collinear pFKN is not detected in either PstDC3000 or Psy B728A
Full virulence of PsmM6 requires pFKN and is essentially caused by avrRpm1
To determine the contribution to virulence of the plasmidpFKN we cured Psm M6 of pFKN (see Experimentalprocedures) to generate strain Psm M6C There was noobvious macroscopic difference in visible disease symp-toms after inoculation of either wild-type or Psm M6Conto rpm1-3 Arabidopsis leaves (unable to recognizeavrRpm1) At high levels of inoculation (5 yen 106 cfu ml-1)the inoculated leaves exhibited water soaking and tissuecollapse starting at 24 h after inoculation Inoculation at5 yen 105 cfu ml-1 led to chlorosis between 48 and 72 h afterinoculation Using this low-density inoculum we mea-sured pathogen growth over time in rpm1-3 leaves PsmM6C grew 10-fold less than the wild-type strain (Fig 2A)Therefore the cured strain is less virulent than the wild-type strain We performed complementation experimentsusing various pFKN subclones in Psm M6C to determinewhich pFKN ORFs would partially or fully restore viru-lence Only clones carrying avrRpm1 could restore fullvirulence (Fig 2A) This result is consistent with inser-tional mutation of avrRpm1 in the Psm M2 strain (Ritterand Dangl 1995) No other clone conferred enhancedvirulence (data not shown)
The role of avrPphE in virulence could not be assessedusing Psm M6C because a second copy of the gene ispresent on the chromosome of the cured strain [asassessed by polymerase chain reaction (PCR) amplifica-tion and sequencing of the PCR product see below] Wereplaced the avrPphE copy on the chromosome of PsmM6C with an omega fragment to create Psm M6CKO (seeExperimental procedures) Psm M6CKO grew similarly toPsm M6C on rpm1-3 plants (Fig 2B) suggesting thatavrPphE plays no major role in the virulence of Psm M6However our recent results indicate that expression ofavrPphE from the lac-nptII promoter in PsmM6CKO doesenhance virulence of PsmM6CKO suggesting that thequantity of AvrPphE protein delivered to the host cell maybe important in its function (J Shock J Chang Z Nim-chuk L Rohmer and J L Dangl unpublished) As avrP-phE was located in the vicinity of avrRpm1 on pFKN wetested the potential involvement of avrPphE in the viru-
1550 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
lence function of avrRpm1 We compared the growth ofPsm M6C and Psm M6CKO when each carried avrRpm1on a plasmid No difference in growth was observed(Fig 2B) indicating that avrPphE does not affectavrRpm1 virulence function
Identification of additional genes putatively involved in pathogenicity based on their expression pattern
The expression of known P syringae virulence factors isspecifically induced in planta Minimal media (MM) plus asugar source seems to mimic the plant apoplast leadingto the expression of the type III pilus genes and effectors
(Huynh et al 1989 Innes et al 1993) We compared theexpression of the pFKN ORFs in MM and in rich medium(KB) using reverse transcription RT-PCR methodsWhen the expression of an ORF was observed in bothmedia we assumed that the gene was constitutivelyexpressed Expression of a gene in MM but not in KB mayindicate a role for this gene during interaction with thehost Most of the pFKN genes are constitutivelyexpressed However the expression of both avrRpm1(Ritter and Dangl 1995) and avrPphE was detected inMM but not in KB Additionally ORF26 is expressed inMM but not in KB ORFs 14 and 24 were not expressedin either of the two conditions The expression of ORF8
Table 2 pFKN ORFs sorted by descending G+C content
a Some pFKN ORFs are contiguous on the plasmid and are collinear in this tableindicating that they possess a similar G+C content When this is the case an emptyrow separates them from the next ORF in the tableb The DNA fragments from the Pst DC3000 genome that contain sequenceshomologous to ORFs from pFKN were colour-coded (1 green 2 red 3 blue4 pink) The table indicates that fragment 4 has a relatively high G+C contentwhereas fragment 3 has a relatively low content
pFKN a virulence plasmid in P syringae pv maculicola 1551
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
and ORF11 could not be assessed using this techniquedespite many attempts
The six genes (avrRpm1 avrPphE ORF8 ORF11ORF14 and ORF24) for which no constitutive expressionwas observed by RT-PCR were selected for further studyWe constructed transcriptional fusions between the 5cent endof the genes of interest and a gene encoding green fluo-rescent protein (GFP) and chose ORF10 as a constitutivecontrol The fluorescence of PsmM6C carrying the tran-scriptional fusions was assessed in KB MM and in plantaby exposing the bacteria grown in these conditions to bluelight (488 nm) and observing them by fluorescencemicroscopy A strain carrying promoterless GFP did notexhibit any fluorescence in the conditions tested whereas
all cells carrying the ORF10ndashGFP fusion did Only theavrPphE avrRpm1 and ORF8 GFP fusions were active inplants (Table 1)
pFKN can integrate into the Psm M6 chromosome
One colony of Psm M6 derived during routine mainte-nance exhibited unusual features Although this straintriggered recognition (measured by HR) on RPM1 plantsand disease symptoms on rpm1 plants no plasmid couldbe isolated from it AvrRpm1 was still present in totalDNA as assayed by DNA blot (Fig 3A) We thereforeconcluded that avrRpm1 was now located on the PsmM6 chromosome To determine whether additional por-tions of pFKN were also present on the variance chromo-some total DNA from the variant Psm M6 was digestedwith EcoRI HindIII EcoRV SacI PvuII SmaI BglII andNsiI and probed with the DNA from different parts of thepFKN plasmid The DNA of Psm M6 containing pFKNwas used as a positive control This analysis revealed apolymorphism between the avrRpm1 gene in this strain(eg an 80 kb BglII fragment) compared with pFKN (egan 88 kb BglII fragment Fig 3A) All restriction frag-ments from pFKN were detected in the DNA of PsmM6integron Therefore pFKN has integrated into thechromosome For the purpose of clarity the strain bear-ing avrRpm1 in the chromosome will be referred to asPsm M6integron
We have on occasion isolated a replicon from indi-vidual colonies of Psm M6integron (see next section)The plasmid purified from these bacteria always exhib-ited a restriction profile identical to that of pFKNTherefore pFKN can excise from the chromosome andPsm M6integron can revert to Psm M6 Interestinglythe overall fitness of Psm M6 is affected by the integra-tion status of pFKN into the chromosome PsmM6integron grows in media (KB and MM) more slowlythan either the original Psm M6 strain or a revertantfrom Psm M6integron that is cured of the plasmid(Fig 3B) Because Psm M6 (with a pFKN replicon) andthe plasmid-cured derivative of Psm M6integron bothgrew equivalently we can conclude that it is the inte-gration of pFKN per se that results in this growthimpairment This is not uncommon as several plasmidreplicons are known to slow growth of the organismwhen integrated (see Discussion)
The pFKN integration site in Psm M6integron is the avrPphE transposon
We noticed from the DNA blot that the genomic environ-ment of avrRpm1 is different in Psm M6 and PsmM6integron This is also the case for both copies of avrP-phE (data not shown) suggesting that the pFKN integra-tion site is near avrRpm1 and avrPphE and that the
Fig 2 avrRpm1 encodes the determining virulence factor on pFKNA The growth of Psm M6C on rpm1-3 plants was compared with the growth of the wild-type Psm M6 strain and Psm M6C strains comple-mented with different fragments from pFKN The pFKN subclone carrying avrRpm1 restored full virulence to Psm M6C (the other subclones did not restore any virulence and are not shown) Leaves were hand inoculated with 105 cfu ml-1 and a titration of bacteria in the leaves was performed 3 days after inoculation Data points rep-resent the mean plusmn standard error of four samplesB The avrPphE gene was deleted from the chromosome of Psm M6C to generate Psm M6CKO To assess the virulence function of avrPphE or its influence on the virulence function of avrRpm1 the growth of Psm M6CKO on rpm1-3 plants was compared with the growth of Psm M6C Psm M6C + avrRpm1 and Psm M6CKO + avrRpm1 Data points represent the mean plusmn standard error of four samples
1552 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
chromosomal location into which the plasmid integrates isnear the chromosomal copy of avrPphE We first definedthat the chromosomal avrPphE is carried on the sametransposon found in the sequence of pFKN using PCRamplification on total DNA of Psm M6C with primersspecific to sequence on the avrPphE transposon(Table 5) The PCR products were sequenced and com-pared with the corresponding sequenced regions ofpFKN As shown in Fig 4A the chromosomal copy of thetransposon contains two sequences not present on pFKNOne 1244 bp sequence (region A) is located betweenavrPphE and the coding region of TnpA It carries an ORFwith a sequence that is nearly identical to various genesencoding resolvases of Tn3 transposons Additionally aportion of 661 bp of this region encodes the 5cent portion ofTnpA Another region of 271 bp unique to the chromo-somal copy is located within the coding region of thetransposase (region C) and is 3cent from a region shared byboth pFKN and chromosomal copies of TnpA (region B)The addition of regions A and C to the chromosomal copyresults in a complete TnpA ORF correcting the frameshiftat the 3cent end of TnpA carried on pFKN
TnpA avrPphE and the putative resolvase fragmentare located between nearly identical inverted repeats of66 bp on both the plasmid and the chromosome Theirorganization is reminiscent of Tn3-like transposons(Sherrat 1989) Moreover this TnpA is 81 identical tothose of the Tn21 subfamily of Tn3 elements (Diveret al 1983) As we suspected that the integration site isin the vicinity of avrPphE in the chromosome and theplasmid (ie in the two transposons) we analysed byPCR the two transposons in the genome of PsmM6integron Figure 4B shows that the unique regions Aand C are no longer located on the same transposonbut are instead separated by the entire plasmidsequence (Fig 4C) This indicated that the integrationsite of the plasmid is located in the region betweenregion A and region C ie within the homology providedby region B (Fig 4)
We used this sequence data to investigate the rateof excision occurring in Psm M6integron grown indifferent conditions by PCR The rate of excision wasestimated at around 10-4 (data not shown) No differencein rate of excision could be detected between bacteria
Fig 3 pFKN can integrate into the Psm M6 chromosomeA Total DNA from Psm M6 (lanes F) and Psm M6integron (lanes I) was digested with EcoRI HindIII EcoRV SacI PvuII SmaI BglII and NsiI and probed with various pFKN fragments Here the DNA blot was probed with the avrRpm1 ORF The two DNA samples exhibit a similar pattern except for a polymorphism (eg EcoRI BglII and SmaI) near the site of pFKN integra-tion into the chromosomeB The integration of the plasmid alters the overall fitness of Psm M6integron The growth of Psm M6integron in KB (empty symbols) and MM (filled symbols) was compared with the growth of Psm M6 and Psm M6C Bacteria were inoculated at 104 cfu ml-1 and cultures were titrated at 0 6 12 29 32 54 and 60 h after the inoculation Psm M6pFKN is indicated by squares Psm M6C is indicated by triangles and rhombi indicate Psm M6integron Data points represent the mean plusmn standard error of four samples (some error bars are not visible because too small) Psm M6pFKN and Psm M6C grew equally well in both minimal medium and KB Psm M6integron grows slower than Psm M6pFKN and Psm M6C in both media
pFKN a virulence plasmid in P syringae pv maculicola 1553
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
grown in KB MM and on resistant or susceptibleplants
The genomic context of avrPphE is different in various P syringae strains
We investigated the regions surrounding the avrPphEtransposon on the chromosome of Psm M6 (Fig 5) byTAIL-PCR The left border shows 97 nt identity to the 5centend of the insertion sequence ISPsy3 (AB063176) fromP syringae pv eriobotryae This element is interrupted bythe avrPphE transposon The translation product of theinterrupted ORF found on this border is homologous tothe IS801 transposase (79 identity and 86 homology)
and IS801 has been associated with other type III effec-tors in various P syringae pathovars (Kim et al 1998)On the right border the 200 bp sequence following theterminal repeat of the avrPphE transposon encodes aprotein homologous (81 nucleotide identity) to the 3centend of an unrelated putative transposase gene fromPseudomonas aeruginosa The next 1062 bp sequencedownstream shows high nucleotide homology tosequence identified in P syringae pv pisi (Arnold et al2001) This DNA fragment may be a transposon remnantand carries the avrPpiA1 allele of avrRpm1 from P syrin-gae pv pisi among other genes In the chromosome ofPsm M6 this sequence shows homology to the P s pisigenes ORFD (94 identity) encoding a putative trans-
Fig 4 The pFKN integration site is a second copy of the avrPphE transposonA The region in the Psm M6 chromosome that hybridizes to the avrPphE and TnpA probes was amplified and sequenced The sequence of the chromosomal region is 100 identical to the transposon in the plasmid but contains two additional regions (regions A and C) TnpA encodes a transposase and res a resolvase Based on homology the chromosomal avrPphE transposon could be complete and functional The avrPphE transposon carried on the plas-mid lacks the 5cent end of TnpA and res (region A) and the absence of region C results in the truncation and a frameshift (marked FS) of the gene encoding the transposase The primers used for PCR amplification (Fig 4B) are indi-cated by arrows and their names are in boldB To investigate where regions A and C were distributed after integration of the plasmid into the chromosome PCRs were performed on the DNA of Psm M6integron and Psm M6 using Larp3 and K110 and K110 and K113 (see Fig 4A) The product amplified from Psm M6integron is run in lane I The product ampli-fied from Psm M6 is run in lane F The product amplified from Psm M6C is run in lane C The difference in size observed between the PCR products from Psm M6integron and Psm M6 using Larp3 and K110 indicates that regions A and C are not carried on the same putative transposon in the chromosome of Psm M6integron (in contrast to the chromosome of Psm M6) The PCR products from Psm M6integron and Psm M6 using K113 and K110 show that neither region A nor region C has been lost in the chromosome of Psm M6integron The identical size of the PCR prod-ucts from Psm M6integron and Psm M6 using K113 and Larp2 indicates that region C is on the right border of pFKN when integrated into the chromosome The region A is on the left border as it is carried by the other putative transposonC After integration of pFKN into the chromo-some TnpA on the left border contains region A but not region C TnpA on the right border contains region C The primers used for the PCR amplification are indicated by arrows and their names are in bold (in Fig 4A)
1554 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
posase and ORF3 encoding a putative type III effector(100 identity) Thus this region in Psm M6 is a remnantof the putative transposon found in P s pisi
We detected the avrPphE transposon in other strains ofP syringae pv maculicola and in the sequence of the
genome of P s tomato DC3000 In Psm M2 Psm M8Psm M12 Psm m90 and Psm m10831 the borders arethe same as in Psm M6 (data not shown) On the otherhand they differ in Psm M5 and Psm M9 and these alldiffer from Pst DC3000 In Psm M5 (Fig 5) and Psm M9
Fig 5 Sequences surrounding the avrPphE transposon define four different genetic contexts The figure is a graphical representation of the four genomic environments in which the avrPphE transposon was detected The regions represented were characterized by sequence comparison using the BLASTX algorithm It is not known whether the avrPphE transposon is located in the chromosome or on a plasmid in the genome of Psm M5 and Pst DC3000
pFKN a virulence plasmid in P syringae pv maculicola 1555
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(not shown) the left border consists of the 5cent end of agene sharing 99 identity to the replicase of pPS10 aplasmid found in P syringae pv savastanoi (Nieto et al1992) This replicase is not related to that of pFKN (aPT23A-like plasmid see above) The right bordersequence is a remnant of a gene encoding a putative ABCtransporter (23 identity and 43 homology to a putativesugar uptake ABC transporter ATP-binding protein fromSinorhizobium meliloti)
In the genome of Pst DC3000 we looked at 30 kbsurrounding the transposon (Fig 5) The proteinsencoded on this 30 kb region share homology to eitherDNA mobility functions (transposases plasmid partition-ing proteins replicases) or type III effectors (identical toavrPpiB Cournoyer et al 1995) The region also containsother sequences often found associated with plasmidsand genes involved in pathogenicity such as rulA and rulB(Sesma et al 2000) Sequences directly neighbouringthe transposon on the left side are homologous to a geneencoding the transposase of the IS100 elements fromYersinia pestis Ralstonia solanacearum and Salmonellaenterica On the right-hand side of the avrPphE transpo-son the first homology is to a fragment of a putative ABCtransporter identical to the one identified in Psm M5 Thissuggests that this part of the two genomes is morerecently diverged than either is from Psm M6
No common sequence was identified in the four differ-ent environments surrounding the avrPphE transposonthat would help to identify the integration target of theavrPphE transposon although the proximal right bordersin Psm M5 and Pst DC3000 are nearly identical Thissuggests that the avrPphE transposon and this border-ing sequence were acquired together by these twostrains and that subsequent events at both the distalright flank and on the left border are responsible for thedifferences in genomic context In contrast the insertionsites of the avrPphE transposon in Psm M5 and PsmM6 are completely different In sum the features ofthese integration sites are consistent with the model thatPAIs evolve by repeated insertion and deletion intopre-existing PAIs (Hensel et al 1999 Tauschek et al2002)
Discussion
We sequenced pFKN an ordf40 kb plasmid that contributesto the virulence of Psm M6 The plasmid carries two typeIII effectors on remnants of transposons contiguous toORF8 itself neighbouring an element closely related toIS801 avrRpm1 encodes a known virulence factor andavrPphE a type III effector The expression of ORF8 isinduced in planta and therefore ORF8 may play a role inthe interaction with the host plant The portion of theplasmid carrying avrRpm1 avrPphE and ORF8 may thus
be considered as a PAI Interestingly the G+C content ofthe ORFs carried on this region are very heterogeneousand the three genes are flanked by sequences related tomobile elements and therefore are likely to have differentorigins This is consistent with the idea that the formationof PAIs requires a multistep process (Hensel et al 1999Alfano et al 2000 Tauschek et al 2002) It is also con-sistent with the findings of Kim et al (1998) showing thatmost of the known type III effectors are associated withthis type of sequence pFKN (and therefore the PAI that itcarries) sometimes integrates into and can excise fromthe chromosome as demonstrated by the discovery of thevariant of the strain Psm M6 We identified the region ofthe chromosome into which the integration took placeThis region is a transposon carrying a second copy ofavrPphE
Alleles of the avrPphE gene have been detected inmany P syringae pathovars (Mansfield et al 1994Alfano et al 2000 Fouts et al 2002 Guttman et al2002) suggesting that it provides an important virulencefunction on a variety of host plants Our findings are thefirst to associate avrPphE with a transposon In PsyB728a and in Pph 1302A for example avrPphE residesdirectly next to the hrp locus in a region called thelsquoexchangeable effector locusrsquo or EEL (Alfano et al 2000)and is not associated with a transposase In fact in PsyB728a avrPphE is predicted to be the third gene in anoperon controlled by the hrpK promoter (Alfano et al2000) This striking finding illustrates the dynamism ofPAIs
We determined by complementation assay that the par-tial loss of virulence in the strain cured of pFKN resultsfrom the absence of avrRpm1 The virulence function ofavrRpm1 has already been demonstrated in the strainPsm M2 (Ritter and Dangl 1995) Recently AvrRpm1 hasbeen demonstrated to interact with RIN4 a proteininvolved in the regulation of some of the plant defenceresponses (Mackey et al 2002) Mackey and coworkersproposed that the role of AvrRpm1 is to reduce the levelof plant defence response triggered by the attack of thepathogen This hypothesis might be consistent with thefact that only quantitative differences in growth wereobserved between Psm M6C and Psm M6 No differencein growth was observed between Psm M6C and PsmM6CKO (lacking avrPphE) Similar results were observedwith the avrPphE allele of P s phaseolicola (Stevenset al 1998) This observation could be explained byeither a functional redundancy with another gene in thegenome of the Psm M6 or the fact that the assay isinappropriate to assess the virulence function of this typeIII effector
The sequence and the integration mechanism of pFKNinto the Psm M6 chromosome provide insights on theevolutionary dynamics of the genomes of P syringae
1556 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
pathovars First the analysis of the G+C content of frag-ments of pFKN suggests that they may have differentorigins This hypothesis is further supported by the pres-ence of some defined fragments from pFKN in thegenomes of Pst DC3000 and Psy B728a The fragment 3(see Table 2) encodes one putative transcriptional regu-lator proteins predicted to be located in the bacterialinner or outer membrane by PSORT (inorganic pyrophos-phatase sugar transporter putative protein) and a puta-tive integraserecombinase homologous to ripX fromBacillus subtilis The presence of this latter gene sug-gests that fragment 3 may be the remnant of abacteriophage
The integration and excision mechanism is very proba-bly a homologous recombination in the 15 kb sequenceregion B of TnpA carried on the avrPphE transposon(Fig 6) It is not clear whether the integrationexcisionprocess relies on general recombination or on a site-specific recombination dependent on an integraserecombinase As indicated by the sequence of the twotransposons before and after integration of pFKN nonucleotide sequences were altered at the site ofexchange and not a single nucleotide was lost or gained(this excludes a mechanism of transpositional site-specificrecombination) General recombination requires a largeDNA region whereas the site-specific recombinationrequires as few as 240 bp for the lambda phage for exam-ple (Thompson and Landy 1989) The rate of generalrecombination is unknown for P syringae but has beenestimated at 10-3 in Pseudomonas tolaasii (and whenrecA is inactivated at 10-4 Sinha et al 2000) The rateof excision of pFKN has been estimated between 10-3 and10-4 and would therefore be consistent with a mechanismof general recombination Additionally the region B is
15 kb long enough to ensure general recombinationOlder reports describe the integration of pMMC7501 intothe chromosome of P s pv phaseolicola (Curiale andMills 1982) An imprecise excision of pMMC7501 led tothe formation of eight derivative plasmids all sharing thesame origin of replication (Szabo and Mills 1984a) Thisexcision occurred through recombination events betweensets of repeats in the chromosome exhibiting sequencehomology with IS elements (Szabo and Mills 1984bPoplawsky and Mills 1987)
The integration of a virulence gene into the chromo-some may be a first step towards its stabilization in thegenome Indeed lack of selection can lead to the stochas-tic loss of the plasmid and many steps in the pathogenlife cycle probably do not require the presence of virulencefunctions The loss of a plasmid carrying the type III effec-tor avrPpiB was observed in P syringae pv pisi (Bavageet al 1991) We propose that the integration of pFKNcould lead to the stabilization of avrRpm1 in the genomeof Psm M6 Interestingly a DNA segment of at least 42 kbcarrying avrPphB in P syringae pv phaseolicola canexcise from the chromosome but it is not maintained inthe strain as a plasmid This region may represent anancient plasmid integration event followed by successivegenomic rearrangements deleterious for plasmid mainte-nance (Jackson et al 2000) The fact that we can stillobserve the excision of pFKN from the chromosome ofPsm M6integron shows that the newly generated geneticunit is not rapidly stabilized in the genome and that furtherevents might be necessary to prevent excision Further-more the integration of pFKN into the chromosomeslightly impairs the fitness of Psm M6integron The pres-ence of the functional origin of replication of pFKN mayinterfere with the replication of the chromosome Integra-
Fig 6 The integration of pFKN may have occurred via homologous recombination between the regions B common to the two TnpA genes The TnpA genes are constituted of four DNA sequence regions (1) before the integration the chromosomal copy of TnpA contains all four regions (A B C and D) The TnpA on the plasmid is missing the 5cent end of the gene (region A) and a fragment towards the 3cent end (region C) which results in a frameshift (FS) After the homologous recombination between two identical regions (region B) in the coding region of the TnpA genes (2) the region A is found in the TnpA of the left border and region C in the TnpA of the right border (3)
pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

1546
L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd
Molecular Microbiology
47
1545ndash1562
nated from the same source (Tauschek
et al
2002) How-ever each of them has evolved and diverged from thePAIs of the other strains through DNA insertions andpotential deletions (Tauschek
et al
2002) On someoccasions a PAI may excise from the chromosome Themechanism of excision is sometimes the same as themechanism of integration For example an integraseencoded on the HPI (high pathogenicity island) of
Yersiniapseudotuberculosis
and
Yersinia pestis
is responsible forthe integration of this PAI into its attachment site on thechromosome (Rakin
et al
2001) Rakin and coworkersshowed that when this integrase is expressed the HPIcould be excised
In the plant pathogen
Pseudomonas syringae
severalPAIs have been identified (Alarcon-Chaidez
et al
1999Jackson
et al
1999 2000 Alfano
et al
2000 Lu
et al
2002) They carry genes encoding the type III secretionsystem apparatus (called the
hrp
cluster) type III effectorproteins that serve as virulence factors or proteins catal-ysing the production of toxins Thus PAIs carry genesrepresenting the major components of
P syringae
patho-genicity The
hrp
cluster is necessary for pathogenicity(Lindgren
et al
1986 1988) and encodes a type III pilus(Jin and He 2001)
The
P syringae
type III secretion system delivers typeIII effector proteins to the plant cell (Cornelis and VanGijsegem 2000 Jin and He 2001 Li
et al
2002)Although some type III effectors have demonstrable rolesin virulence very little is known about their host targets(Nimchuk
et al
2001) Target processes affected by
Psyringae
type III effectors were described recently (Mackey
et al
2002 Van der Hoorn
et al
2002) Some type IIIeffectors are recognized inside the plant cell by the plant
R
gene system and these trigger a disease resistance-associated programmed cell death known as the hyper-sensitive response (HR) (Dangl and Jones 2001)Because the recognition of some type III effector proteinscauses the plant to be resistant to a particular pathogenstrain most of the genes encoding
P syringae
type IIIeffectors were originally named avirulence (
avr
) genesThe
P syringae hrp
PAI also encodes the HrpL alterna-tive sigma factor This factor regulates the expression ofboth the genes encoding the type III delivery pilus andthose encoding type III effectors (Xiao
et al
1994) Thepromoters of these genes contain a consensus
hrp
box(Innes
et al
1993) Hence many genes encoding puta-tive type III effectors have been identified recently usingsequence analysis methods on the whole genome of the
Pst
DC3000 strain and sequences of DNA fragments fromother pathovars (Jackson
et al
1999 Alfano
et al
2000Arnold
et al
2001 Boch
et al
2002 Fouts
et al
2002Petnicki-Ocwieja
et al
2002)The type III effector avrRpm1 (Ritter and Dangl 1995)
contributes to virulence on
Arabidopsis
plants that cannot
recognize it (host genotype
rpm1
) It is encoded on aplasmid in several
P syringae
pv
maculicola
(
Psm
)strains hereafter named pFKN Several
P syringae
plas-mids have been partially or fully characterized They oftencarry genes encoding virulence factors or functionsenabling the bacteria to survive in given environments (fora review see Vivian
et al
2001) We describe the analy-sis of the pFKN sequence and suggest that it carries atleast one PAI We investigated the mechanism by whichthe plasmid integrates and excises into and out of thechromosome of
Psm
M6 as well as the chromosomalregion involved in the process We also found that a pieceof the pFKN mosaic structure is a transposon carrying anallele of a second type III effector
avrPphE
Finally wedescribe the genomic context of this transposon in several
P syringae
strains Our work provides insights into theevolutionary process leading to the formation of new PAIsin
P syringae
and how they may be transferred fromstrain to strain
Results
Sequencing of pFKN
The sequence of pFKN was determined to an average of
ordf
31
yen
coverage using a shotgun approach The completenucleotide sequence of pFKN consists of 39 554 bp Itsoverall G
+
C content is 5353 Twenty-nine putative openreading frames (ORFs) were identified representing757 of the sequence To assign possible functions tothese ORFs we compared the sequence of each withavailable databases using
BLASTN
and
BLASTX
Twenty-seven ORFs had homology with genes present in thedatabases We searched the translated products of eachORF for known functional domains and motifs and couldassign a putative function to most of them (Table 1)
We organized the ORFs into various functional groups(Fig 1) The plasmid encodes two type III effectors(
avrRpm1
and an allele of
avrPphE
) Five non-contiguousregions on the plasmid may be mobile elements or rem-nants thereof Three other groups of genes were definedaccording to the putative function that they encode
Plasmid replication and maintenance
ORF1 shares highhomology with genes that encode the replication proteinsof plasmids belonging to the PT23A family in the P syrin-gae species (Sesma et al 1998) Homologous replicationproteins are found in other genera such as the replicationproteins from the ColE2 family in E coli (Sesma et al1998) The product of ORF10 shares similarity with ParAa protein involved in partitioning working in concert with aprotein encoded by ParB No ParB gene was found onpFKN Therefore it is not clear how the plasmid copies(around five per cell data not shown) are distributed effi-
pFKN a virulence plasmid in
P syringae
pv
maculicola 1547
copy 2003 Blackwell Publishing Ltd
Molecular Microbiology
47
1545ndash1562
ciently after cell division ORF28 may encode a proteininvolved in the transfer of pFKN The localization of theprotein encoded by ORF28 is predicted to be in the bac-terial periplasmic space It has three domains The N-terminus shows high homologies to VirB1 of Brucella suis(a mammalian pathogen) named after its homology toVirB1 of Agrobacterium tumefaciens The central regionshows homology to VirB4 of the same organism The C-terminus shares homology with different DNA-binding pro-teins (DNA nickase primase) However no other gene onthe plasmid seems to encode a function related to trans-fer It is highly unlikely that this unique protein would allow
transfer of pFKN Therefore ORF28 encodes an unknownfunction or could represent the remnant of a transfer sys-tem encoded on the plasmid before genomic rearrange-ments leading to its loss
Type III effectors
The promoter regions of two genes onthe plasmid contain an
hrp
box suggesting that they arepart of the type III regulon ORF2 is 80 identical to the
avrPphE
allele carried by
P s syringae
(
Psy
) (Alfanoet al 2000) The avrPphE gene had originally beenidentified in P s phaseolicola (Pph) and encodes aprotein delivered to the host cell (Mansfield et al 1994)
Table 1 ORFs and non-coding elements identified in Ps pv maculicola pFKN
a We identified 4 DNA fragments that were conserved between pFKN and the genome of Pst DC3000 (gt85 nucleotide identity) The ORFs thathave no homologues in the genome of Pst DC3000 are noted as lsquononersquo the fragments are numbered 1ndash4b The expression of each gene carried by the plasmid was assessed in KB and MM (hrp-inducing media) by RT-PCR and by using GFP-translationalfusions in planta for 7 of them (not shown) When the gene is expressed in one given condition the table exhibits lsquo+rsquo A lack of expression is markedby lsquondashrsquo NA not available
1548 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Since then several alleles have been identified in otherpathovars of P syringae (Fouts et al 2002 Guttmanet al 2002) They all contain an hrp box in ORF2 ofpFKN this is found 51 bp upstream of the ATG at nt 1561as 5cent-GGAACTGA-N13-CGACATA-3cent (Innes et al1993) Based on the homologies and the presence ofthe hrp box in the promoter we hypothesized that theAvrPphE-like protein encoded by ORF2 is also a type IIIeffector (see below) AvrRpm1 (numbered ORF5) is iden-tical to that defined in the strain Psm M2 (Dangl et al1992)
Mobile element-related genes ORF3 (TnpA) encodes aputative truncated transposase Importantly avrPphE andTnpA are located between nearly identical invertedrepeats of 66 bp (Fig 1) thus defining a probable trans-posable element remnant The sequences surroundingavrRpm1 (ORF4 and ORF6) are homologous to asequence identified in P s pisi by Arnold et al (2001)with 96ndash97 nucleotide identity The DNA fragment car-rying these sequences in P s pisi may also be a trans-poson or a remnant It is therefore likely that avrRpm1 isalso located on the remnant of a transposon
ORF13 and ORF14 are related to ORFs found in theprophage 933 L on the LEE of the enterohaemorrhagic Ecoli O157H7 L0015 and L0014 respectively These twoORFs are thought to belong to an IS element from theIS66 family primarily identified in Sinorhizobium melilotithat includes IS elements from animal and plant patho-gens These two genes are usually associated with a thirdone L0013 in the LEE that is absent in pFKN (the
sequence upstream of ORF14 does not contain any ORFand does not show homology to L0013) Moreover the 5centend of ORF14 is also missing Therefore the sequenceencoding ORF13 and ORF14 may represent the remnantof an IS element belonging to the IS66 family not previ-ously identified in P syringae
Transcription factors Four pFKN genes share homologywith transcriptional regulators Some of these may func-tion during interactions with eukaryotic hosts For exam-ple the closest homologue to ORF24 is a putativetranscriptional factor from V cholerae ORF24 is alsorelated to putative proteins of unknown function (along thewhole length of the protein) found in organisms as diverseas E coli (50 identity67 homology) Caenorhabditiselegans (4157) Homo sapiens (4258) Droso-phila melanogaster (4058) and Arabidopsis thaliana(2942) A domain characteristic of FAD-dependentoxidoreductase (in Pfam) is detected in ORF24 There-fore the function of the translation product of this ORFmay require electron transfer through FAD
The protein encoded by ORF27 is a member of thesmall RfaH family RfaH controls the transcription of E coliand Salmonella operons that direct the synthesis assem-bly and export of the lipopolysaccharide core exopolysac-charide F-conjugation pilus and haemolysin toxin (for areview see Santangelo and Roberts 2002) RfaH isa specific regulator of transcript elongation its lossincreases transcription polarity in these operons withoutaffecting initiation from the operon promoters In additionin one case the RfaH protein controls the transcription of
Fig 1 Graphical map of pFKN and its 29 iden-tified ORFs The proposed direction of tran-scription is indicated by arrows Red boxes indicate genes encoding for type III effectors A black arrow line at the 5cent end of any genes indicates a possible hrp box Genes related to those of mobile elements are represented by green boxes The avrPphE transposon and the IS801 homologue are framed Transposon repeats surrounding the avrPphE transposon are marked by green lines The avrRpm1 gene is surrounded by repeats similar to the repeats of the transposon Tn501 these repeats are also marked by green lines Purple boxes indi-cate genes with replication and partition func-tions Genes encoding for putative transporters are indicated by blue boxes The putative tran-scriptional regulators are indicated by orange boxes The grey boxes represent genes that do not fall into any of the categories listed The number of each ORF is noted in bold on the outside of the circle
pFKN a virulence plasmid in P syringae pv maculicola 1549
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
a single gene encoding an outer membrane haeminreceptor ChuA in pathogenic E coli as well as othergenes involved in pathogenicity of the strain (Nagy et al2001 2002) The promoter of the genes or operons con-trolled by RfaH contains a conserved sequence calledJUMPStart (Hobbs and Reeves 1994) No JUMPStartsequence was detected on pFKN
Transporters ORF18 encodes a probable transmem-brane protein that is highly homologous to sugar trans-porters One is the product of mae1 a gene of the yeastSchizosaccharomyces pombe encoding a proteininvolved in the uptake of L-malate succinate and malonicacid The product of ORF18 may provide the bacteria witha fitness advantage in certain conditions
Other putative functions encoded on the plasmid Othergenes may encode proteins providing an advantage to thebacteria during the infection process For example theproduct of ORF12 displays all the characteristics ofmethyl-accepting chemotaxis proteins The genes encod-ing the flagellar unit and the chemotaxis functions areoften organized in clusters which is not the case onpFKN In this particular case the flagellar unit may beprovided in trans
pFKN is composed of different fragments that indicate a multistep process leading to its formation
The G+C content of the pFKN ORFs is highly variablealong the plasmid (Table 2) Yet several contiguous genesets on the plasmid have similar G+C contents This prob-ably indicates that these genes were acquired togetherfrom an original source The G+C content might allow usto track the multistep acquisition process resulting inpFKN For example ORFs 7 8 and 10 share nearly iden-tical G+C content (505) but are separated by an ISelement with a significantly different G+C content (603)To determine the distribution of pFKN ORFs in otherpathovars and how this distribution is related to G+Ccontent by ORF group we performed sequence compar-isons with the nucleotide sequence of the completegenomes of P syringae pv tomato DC3000 (httptigr-blasttigrorgufmg) and P syringae pv syringae B728a(httpwwwjgidoegovJGI_microbialhtmlindexhtml)DNA fragments carrying from two to seven contiguouspFKN ORFs were detected in both genome sequences(85ndash100 identity) The four DNA fragments from P stomato DC3000 carrying linked pFKN ORFs were num-bered and added to Table 2 Of these Pst DC3000 frag-ment 2 was also detected in Psy B728a (98 identical topFKN) In addition sequences homologous to repA andORF13 were also detected in this Psy B728a Table 2shows that ORFs belonging to the same contiguous DNA
fragment have a similar G+C content For example genesbelonging to fragment 4 have a relatively high G+C con-tent Genes belonging to fragments 1 and 2 have similarG+C contents and genes in fragment 3 exhibit a lowerG+C content Thus the different fragments constitutingpFKN were probably acquired by the three genomes indifferent steps followed by subsequent sequence dele-tions andor rearrangements with respect to Psm M6 asthe entire collinear pFKN is not detected in either PstDC3000 or Psy B728A
Full virulence of PsmM6 requires pFKN and is essentially caused by avrRpm1
To determine the contribution to virulence of the plasmidpFKN we cured Psm M6 of pFKN (see Experimentalprocedures) to generate strain Psm M6C There was noobvious macroscopic difference in visible disease symp-toms after inoculation of either wild-type or Psm M6Conto rpm1-3 Arabidopsis leaves (unable to recognizeavrRpm1) At high levels of inoculation (5 yen 106 cfu ml-1)the inoculated leaves exhibited water soaking and tissuecollapse starting at 24 h after inoculation Inoculation at5 yen 105 cfu ml-1 led to chlorosis between 48 and 72 h afterinoculation Using this low-density inoculum we mea-sured pathogen growth over time in rpm1-3 leaves PsmM6C grew 10-fold less than the wild-type strain (Fig 2A)Therefore the cured strain is less virulent than the wild-type strain We performed complementation experimentsusing various pFKN subclones in Psm M6C to determinewhich pFKN ORFs would partially or fully restore viru-lence Only clones carrying avrRpm1 could restore fullvirulence (Fig 2A) This result is consistent with inser-tional mutation of avrRpm1 in the Psm M2 strain (Ritterand Dangl 1995) No other clone conferred enhancedvirulence (data not shown)
The role of avrPphE in virulence could not be assessedusing Psm M6C because a second copy of the gene ispresent on the chromosome of the cured strain [asassessed by polymerase chain reaction (PCR) amplifica-tion and sequencing of the PCR product see below] Wereplaced the avrPphE copy on the chromosome of PsmM6C with an omega fragment to create Psm M6CKO (seeExperimental procedures) Psm M6CKO grew similarly toPsm M6C on rpm1-3 plants (Fig 2B) suggesting thatavrPphE plays no major role in the virulence of Psm M6However our recent results indicate that expression ofavrPphE from the lac-nptII promoter in PsmM6CKO doesenhance virulence of PsmM6CKO suggesting that thequantity of AvrPphE protein delivered to the host cell maybe important in its function (J Shock J Chang Z Nim-chuk L Rohmer and J L Dangl unpublished) As avrP-phE was located in the vicinity of avrRpm1 on pFKN wetested the potential involvement of avrPphE in the viru-
1550 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
lence function of avrRpm1 We compared the growth ofPsm M6C and Psm M6CKO when each carried avrRpm1on a plasmid No difference in growth was observed(Fig 2B) indicating that avrPphE does not affectavrRpm1 virulence function
Identification of additional genes putatively involved in pathogenicity based on their expression pattern
The expression of known P syringae virulence factors isspecifically induced in planta Minimal media (MM) plus asugar source seems to mimic the plant apoplast leadingto the expression of the type III pilus genes and effectors
(Huynh et al 1989 Innes et al 1993) We compared theexpression of the pFKN ORFs in MM and in rich medium(KB) using reverse transcription RT-PCR methodsWhen the expression of an ORF was observed in bothmedia we assumed that the gene was constitutivelyexpressed Expression of a gene in MM but not in KB mayindicate a role for this gene during interaction with thehost Most of the pFKN genes are constitutivelyexpressed However the expression of both avrRpm1(Ritter and Dangl 1995) and avrPphE was detected inMM but not in KB Additionally ORF26 is expressed inMM but not in KB ORFs 14 and 24 were not expressedin either of the two conditions The expression of ORF8
Table 2 pFKN ORFs sorted by descending G+C content
a Some pFKN ORFs are contiguous on the plasmid and are collinear in this tableindicating that they possess a similar G+C content When this is the case an emptyrow separates them from the next ORF in the tableb The DNA fragments from the Pst DC3000 genome that contain sequenceshomologous to ORFs from pFKN were colour-coded (1 green 2 red 3 blue4 pink) The table indicates that fragment 4 has a relatively high G+C contentwhereas fragment 3 has a relatively low content
pFKN a virulence plasmid in P syringae pv maculicola 1551
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
and ORF11 could not be assessed using this techniquedespite many attempts
The six genes (avrRpm1 avrPphE ORF8 ORF11ORF14 and ORF24) for which no constitutive expressionwas observed by RT-PCR were selected for further studyWe constructed transcriptional fusions between the 5cent endof the genes of interest and a gene encoding green fluo-rescent protein (GFP) and chose ORF10 as a constitutivecontrol The fluorescence of PsmM6C carrying the tran-scriptional fusions was assessed in KB MM and in plantaby exposing the bacteria grown in these conditions to bluelight (488 nm) and observing them by fluorescencemicroscopy A strain carrying promoterless GFP did notexhibit any fluorescence in the conditions tested whereas
all cells carrying the ORF10ndashGFP fusion did Only theavrPphE avrRpm1 and ORF8 GFP fusions were active inplants (Table 1)
pFKN can integrate into the Psm M6 chromosome
One colony of Psm M6 derived during routine mainte-nance exhibited unusual features Although this straintriggered recognition (measured by HR) on RPM1 plantsand disease symptoms on rpm1 plants no plasmid couldbe isolated from it AvrRpm1 was still present in totalDNA as assayed by DNA blot (Fig 3A) We thereforeconcluded that avrRpm1 was now located on the PsmM6 chromosome To determine whether additional por-tions of pFKN were also present on the variance chromo-some total DNA from the variant Psm M6 was digestedwith EcoRI HindIII EcoRV SacI PvuII SmaI BglII andNsiI and probed with the DNA from different parts of thepFKN plasmid The DNA of Psm M6 containing pFKNwas used as a positive control This analysis revealed apolymorphism between the avrRpm1 gene in this strain(eg an 80 kb BglII fragment) compared with pFKN (egan 88 kb BglII fragment Fig 3A) All restriction frag-ments from pFKN were detected in the DNA of PsmM6integron Therefore pFKN has integrated into thechromosome For the purpose of clarity the strain bear-ing avrRpm1 in the chromosome will be referred to asPsm M6integron
We have on occasion isolated a replicon from indi-vidual colonies of Psm M6integron (see next section)The plasmid purified from these bacteria always exhib-ited a restriction profile identical to that of pFKNTherefore pFKN can excise from the chromosome andPsm M6integron can revert to Psm M6 Interestinglythe overall fitness of Psm M6 is affected by the integra-tion status of pFKN into the chromosome PsmM6integron grows in media (KB and MM) more slowlythan either the original Psm M6 strain or a revertantfrom Psm M6integron that is cured of the plasmid(Fig 3B) Because Psm M6 (with a pFKN replicon) andthe plasmid-cured derivative of Psm M6integron bothgrew equivalently we can conclude that it is the inte-gration of pFKN per se that results in this growthimpairment This is not uncommon as several plasmidreplicons are known to slow growth of the organismwhen integrated (see Discussion)
The pFKN integration site in Psm M6integron is the avrPphE transposon
We noticed from the DNA blot that the genomic environ-ment of avrRpm1 is different in Psm M6 and PsmM6integron This is also the case for both copies of avrP-phE (data not shown) suggesting that the pFKN integra-tion site is near avrRpm1 and avrPphE and that the
Fig 2 avrRpm1 encodes the determining virulence factor on pFKNA The growth of Psm M6C on rpm1-3 plants was compared with the growth of the wild-type Psm M6 strain and Psm M6C strains comple-mented with different fragments from pFKN The pFKN subclone carrying avrRpm1 restored full virulence to Psm M6C (the other subclones did not restore any virulence and are not shown) Leaves were hand inoculated with 105 cfu ml-1 and a titration of bacteria in the leaves was performed 3 days after inoculation Data points rep-resent the mean plusmn standard error of four samplesB The avrPphE gene was deleted from the chromosome of Psm M6C to generate Psm M6CKO To assess the virulence function of avrPphE or its influence on the virulence function of avrRpm1 the growth of Psm M6CKO on rpm1-3 plants was compared with the growth of Psm M6C Psm M6C + avrRpm1 and Psm M6CKO + avrRpm1 Data points represent the mean plusmn standard error of four samples
1552 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
chromosomal location into which the plasmid integrates isnear the chromosomal copy of avrPphE We first definedthat the chromosomal avrPphE is carried on the sametransposon found in the sequence of pFKN using PCRamplification on total DNA of Psm M6C with primersspecific to sequence on the avrPphE transposon(Table 5) The PCR products were sequenced and com-pared with the corresponding sequenced regions ofpFKN As shown in Fig 4A the chromosomal copy of thetransposon contains two sequences not present on pFKNOne 1244 bp sequence (region A) is located betweenavrPphE and the coding region of TnpA It carries an ORFwith a sequence that is nearly identical to various genesencoding resolvases of Tn3 transposons Additionally aportion of 661 bp of this region encodes the 5cent portion ofTnpA Another region of 271 bp unique to the chromo-somal copy is located within the coding region of thetransposase (region C) and is 3cent from a region shared byboth pFKN and chromosomal copies of TnpA (region B)The addition of regions A and C to the chromosomal copyresults in a complete TnpA ORF correcting the frameshiftat the 3cent end of TnpA carried on pFKN
TnpA avrPphE and the putative resolvase fragmentare located between nearly identical inverted repeats of66 bp on both the plasmid and the chromosome Theirorganization is reminiscent of Tn3-like transposons(Sherrat 1989) Moreover this TnpA is 81 identical tothose of the Tn21 subfamily of Tn3 elements (Diveret al 1983) As we suspected that the integration site isin the vicinity of avrPphE in the chromosome and theplasmid (ie in the two transposons) we analysed byPCR the two transposons in the genome of PsmM6integron Figure 4B shows that the unique regions Aand C are no longer located on the same transposonbut are instead separated by the entire plasmidsequence (Fig 4C) This indicated that the integrationsite of the plasmid is located in the region betweenregion A and region C ie within the homology providedby region B (Fig 4)
We used this sequence data to investigate the rateof excision occurring in Psm M6integron grown indifferent conditions by PCR The rate of excision wasestimated at around 10-4 (data not shown) No differencein rate of excision could be detected between bacteria
Fig 3 pFKN can integrate into the Psm M6 chromosomeA Total DNA from Psm M6 (lanes F) and Psm M6integron (lanes I) was digested with EcoRI HindIII EcoRV SacI PvuII SmaI BglII and NsiI and probed with various pFKN fragments Here the DNA blot was probed with the avrRpm1 ORF The two DNA samples exhibit a similar pattern except for a polymorphism (eg EcoRI BglII and SmaI) near the site of pFKN integra-tion into the chromosomeB The integration of the plasmid alters the overall fitness of Psm M6integron The growth of Psm M6integron in KB (empty symbols) and MM (filled symbols) was compared with the growth of Psm M6 and Psm M6C Bacteria were inoculated at 104 cfu ml-1 and cultures were titrated at 0 6 12 29 32 54 and 60 h after the inoculation Psm M6pFKN is indicated by squares Psm M6C is indicated by triangles and rhombi indicate Psm M6integron Data points represent the mean plusmn standard error of four samples (some error bars are not visible because too small) Psm M6pFKN and Psm M6C grew equally well in both minimal medium and KB Psm M6integron grows slower than Psm M6pFKN and Psm M6C in both media
pFKN a virulence plasmid in P syringae pv maculicola 1553
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
grown in KB MM and on resistant or susceptibleplants
The genomic context of avrPphE is different in various P syringae strains
We investigated the regions surrounding the avrPphEtransposon on the chromosome of Psm M6 (Fig 5) byTAIL-PCR The left border shows 97 nt identity to the 5centend of the insertion sequence ISPsy3 (AB063176) fromP syringae pv eriobotryae This element is interrupted bythe avrPphE transposon The translation product of theinterrupted ORF found on this border is homologous tothe IS801 transposase (79 identity and 86 homology)
and IS801 has been associated with other type III effec-tors in various P syringae pathovars (Kim et al 1998)On the right border the 200 bp sequence following theterminal repeat of the avrPphE transposon encodes aprotein homologous (81 nucleotide identity) to the 3centend of an unrelated putative transposase gene fromPseudomonas aeruginosa The next 1062 bp sequencedownstream shows high nucleotide homology tosequence identified in P syringae pv pisi (Arnold et al2001) This DNA fragment may be a transposon remnantand carries the avrPpiA1 allele of avrRpm1 from P syrin-gae pv pisi among other genes In the chromosome ofPsm M6 this sequence shows homology to the P s pisigenes ORFD (94 identity) encoding a putative trans-
Fig 4 The pFKN integration site is a second copy of the avrPphE transposonA The region in the Psm M6 chromosome that hybridizes to the avrPphE and TnpA probes was amplified and sequenced The sequence of the chromosomal region is 100 identical to the transposon in the plasmid but contains two additional regions (regions A and C) TnpA encodes a transposase and res a resolvase Based on homology the chromosomal avrPphE transposon could be complete and functional The avrPphE transposon carried on the plas-mid lacks the 5cent end of TnpA and res (region A) and the absence of region C results in the truncation and a frameshift (marked FS) of the gene encoding the transposase The primers used for PCR amplification (Fig 4B) are indi-cated by arrows and their names are in boldB To investigate where regions A and C were distributed after integration of the plasmid into the chromosome PCRs were performed on the DNA of Psm M6integron and Psm M6 using Larp3 and K110 and K110 and K113 (see Fig 4A) The product amplified from Psm M6integron is run in lane I The product ampli-fied from Psm M6 is run in lane F The product amplified from Psm M6C is run in lane C The difference in size observed between the PCR products from Psm M6integron and Psm M6 using Larp3 and K110 indicates that regions A and C are not carried on the same putative transposon in the chromosome of Psm M6integron (in contrast to the chromosome of Psm M6) The PCR products from Psm M6integron and Psm M6 using K113 and K110 show that neither region A nor region C has been lost in the chromosome of Psm M6integron The identical size of the PCR prod-ucts from Psm M6integron and Psm M6 using K113 and Larp2 indicates that region C is on the right border of pFKN when integrated into the chromosome The region A is on the left border as it is carried by the other putative transposonC After integration of pFKN into the chromo-some TnpA on the left border contains region A but not region C TnpA on the right border contains region C The primers used for the PCR amplification are indicated by arrows and their names are in bold (in Fig 4A)
1554 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
posase and ORF3 encoding a putative type III effector(100 identity) Thus this region in Psm M6 is a remnantof the putative transposon found in P s pisi
We detected the avrPphE transposon in other strains ofP syringae pv maculicola and in the sequence of the
genome of P s tomato DC3000 In Psm M2 Psm M8Psm M12 Psm m90 and Psm m10831 the borders arethe same as in Psm M6 (data not shown) On the otherhand they differ in Psm M5 and Psm M9 and these alldiffer from Pst DC3000 In Psm M5 (Fig 5) and Psm M9
Fig 5 Sequences surrounding the avrPphE transposon define four different genetic contexts The figure is a graphical representation of the four genomic environments in which the avrPphE transposon was detected The regions represented were characterized by sequence comparison using the BLASTX algorithm It is not known whether the avrPphE transposon is located in the chromosome or on a plasmid in the genome of Psm M5 and Pst DC3000
pFKN a virulence plasmid in P syringae pv maculicola 1555
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(not shown) the left border consists of the 5cent end of agene sharing 99 identity to the replicase of pPS10 aplasmid found in P syringae pv savastanoi (Nieto et al1992) This replicase is not related to that of pFKN (aPT23A-like plasmid see above) The right bordersequence is a remnant of a gene encoding a putative ABCtransporter (23 identity and 43 homology to a putativesugar uptake ABC transporter ATP-binding protein fromSinorhizobium meliloti)
In the genome of Pst DC3000 we looked at 30 kbsurrounding the transposon (Fig 5) The proteinsencoded on this 30 kb region share homology to eitherDNA mobility functions (transposases plasmid partition-ing proteins replicases) or type III effectors (identical toavrPpiB Cournoyer et al 1995) The region also containsother sequences often found associated with plasmidsand genes involved in pathogenicity such as rulA and rulB(Sesma et al 2000) Sequences directly neighbouringthe transposon on the left side are homologous to a geneencoding the transposase of the IS100 elements fromYersinia pestis Ralstonia solanacearum and Salmonellaenterica On the right-hand side of the avrPphE transpo-son the first homology is to a fragment of a putative ABCtransporter identical to the one identified in Psm M5 Thissuggests that this part of the two genomes is morerecently diverged than either is from Psm M6
No common sequence was identified in the four differ-ent environments surrounding the avrPphE transposonthat would help to identify the integration target of theavrPphE transposon although the proximal right bordersin Psm M5 and Pst DC3000 are nearly identical Thissuggests that the avrPphE transposon and this border-ing sequence were acquired together by these twostrains and that subsequent events at both the distalright flank and on the left border are responsible for thedifferences in genomic context In contrast the insertionsites of the avrPphE transposon in Psm M5 and PsmM6 are completely different In sum the features ofthese integration sites are consistent with the model thatPAIs evolve by repeated insertion and deletion intopre-existing PAIs (Hensel et al 1999 Tauschek et al2002)
Discussion
We sequenced pFKN an ordf40 kb plasmid that contributesto the virulence of Psm M6 The plasmid carries two typeIII effectors on remnants of transposons contiguous toORF8 itself neighbouring an element closely related toIS801 avrRpm1 encodes a known virulence factor andavrPphE a type III effector The expression of ORF8 isinduced in planta and therefore ORF8 may play a role inthe interaction with the host plant The portion of theplasmid carrying avrRpm1 avrPphE and ORF8 may thus
be considered as a PAI Interestingly the G+C content ofthe ORFs carried on this region are very heterogeneousand the three genes are flanked by sequences related tomobile elements and therefore are likely to have differentorigins This is consistent with the idea that the formationof PAIs requires a multistep process (Hensel et al 1999Alfano et al 2000 Tauschek et al 2002) It is also con-sistent with the findings of Kim et al (1998) showing thatmost of the known type III effectors are associated withthis type of sequence pFKN (and therefore the PAI that itcarries) sometimes integrates into and can excise fromthe chromosome as demonstrated by the discovery of thevariant of the strain Psm M6 We identified the region ofthe chromosome into which the integration took placeThis region is a transposon carrying a second copy ofavrPphE
Alleles of the avrPphE gene have been detected inmany P syringae pathovars (Mansfield et al 1994Alfano et al 2000 Fouts et al 2002 Guttman et al2002) suggesting that it provides an important virulencefunction on a variety of host plants Our findings are thefirst to associate avrPphE with a transposon In PsyB728a and in Pph 1302A for example avrPphE residesdirectly next to the hrp locus in a region called thelsquoexchangeable effector locusrsquo or EEL (Alfano et al 2000)and is not associated with a transposase In fact in PsyB728a avrPphE is predicted to be the third gene in anoperon controlled by the hrpK promoter (Alfano et al2000) This striking finding illustrates the dynamism ofPAIs
We determined by complementation assay that the par-tial loss of virulence in the strain cured of pFKN resultsfrom the absence of avrRpm1 The virulence function ofavrRpm1 has already been demonstrated in the strainPsm M2 (Ritter and Dangl 1995) Recently AvrRpm1 hasbeen demonstrated to interact with RIN4 a proteininvolved in the regulation of some of the plant defenceresponses (Mackey et al 2002) Mackey and coworkersproposed that the role of AvrRpm1 is to reduce the levelof plant defence response triggered by the attack of thepathogen This hypothesis might be consistent with thefact that only quantitative differences in growth wereobserved between Psm M6C and Psm M6 No differencein growth was observed between Psm M6C and PsmM6CKO (lacking avrPphE) Similar results were observedwith the avrPphE allele of P s phaseolicola (Stevenset al 1998) This observation could be explained byeither a functional redundancy with another gene in thegenome of the Psm M6 or the fact that the assay isinappropriate to assess the virulence function of this typeIII effector
The sequence and the integration mechanism of pFKNinto the Psm M6 chromosome provide insights on theevolutionary dynamics of the genomes of P syringae
1556 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
pathovars First the analysis of the G+C content of frag-ments of pFKN suggests that they may have differentorigins This hypothesis is further supported by the pres-ence of some defined fragments from pFKN in thegenomes of Pst DC3000 and Psy B728a The fragment 3(see Table 2) encodes one putative transcriptional regu-lator proteins predicted to be located in the bacterialinner or outer membrane by PSORT (inorganic pyrophos-phatase sugar transporter putative protein) and a puta-tive integraserecombinase homologous to ripX fromBacillus subtilis The presence of this latter gene sug-gests that fragment 3 may be the remnant of abacteriophage
The integration and excision mechanism is very proba-bly a homologous recombination in the 15 kb sequenceregion B of TnpA carried on the avrPphE transposon(Fig 6) It is not clear whether the integrationexcisionprocess relies on general recombination or on a site-specific recombination dependent on an integraserecombinase As indicated by the sequence of the twotransposons before and after integration of pFKN nonucleotide sequences were altered at the site ofexchange and not a single nucleotide was lost or gained(this excludes a mechanism of transpositional site-specificrecombination) General recombination requires a largeDNA region whereas the site-specific recombinationrequires as few as 240 bp for the lambda phage for exam-ple (Thompson and Landy 1989) The rate of generalrecombination is unknown for P syringae but has beenestimated at 10-3 in Pseudomonas tolaasii (and whenrecA is inactivated at 10-4 Sinha et al 2000) The rateof excision of pFKN has been estimated between 10-3 and10-4 and would therefore be consistent with a mechanismof general recombination Additionally the region B is
15 kb long enough to ensure general recombinationOlder reports describe the integration of pMMC7501 intothe chromosome of P s pv phaseolicola (Curiale andMills 1982) An imprecise excision of pMMC7501 led tothe formation of eight derivative plasmids all sharing thesame origin of replication (Szabo and Mills 1984a) Thisexcision occurred through recombination events betweensets of repeats in the chromosome exhibiting sequencehomology with IS elements (Szabo and Mills 1984bPoplawsky and Mills 1987)
The integration of a virulence gene into the chromo-some may be a first step towards its stabilization in thegenome Indeed lack of selection can lead to the stochas-tic loss of the plasmid and many steps in the pathogenlife cycle probably do not require the presence of virulencefunctions The loss of a plasmid carrying the type III effec-tor avrPpiB was observed in P syringae pv pisi (Bavageet al 1991) We propose that the integration of pFKNcould lead to the stabilization of avrRpm1 in the genomeof Psm M6 Interestingly a DNA segment of at least 42 kbcarrying avrPphB in P syringae pv phaseolicola canexcise from the chromosome but it is not maintained inthe strain as a plasmid This region may represent anancient plasmid integration event followed by successivegenomic rearrangements deleterious for plasmid mainte-nance (Jackson et al 2000) The fact that we can stillobserve the excision of pFKN from the chromosome ofPsm M6integron shows that the newly generated geneticunit is not rapidly stabilized in the genome and that furtherevents might be necessary to prevent excision Further-more the integration of pFKN into the chromosomeslightly impairs the fitness of Psm M6integron The pres-ence of the functional origin of replication of pFKN mayinterfere with the replication of the chromosome Integra-
Fig 6 The integration of pFKN may have occurred via homologous recombination between the regions B common to the two TnpA genes The TnpA genes are constituted of four DNA sequence regions (1) before the integration the chromosomal copy of TnpA contains all four regions (A B C and D) The TnpA on the plasmid is missing the 5cent end of the gene (region A) and a fragment towards the 3cent end (region C) which results in a frameshift (FS) After the homologous recombination between two identical regions (region B) in the coding region of the TnpA genes (2) the region A is found in the TnpA of the left border and region C in the TnpA of the right border (3)
pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143
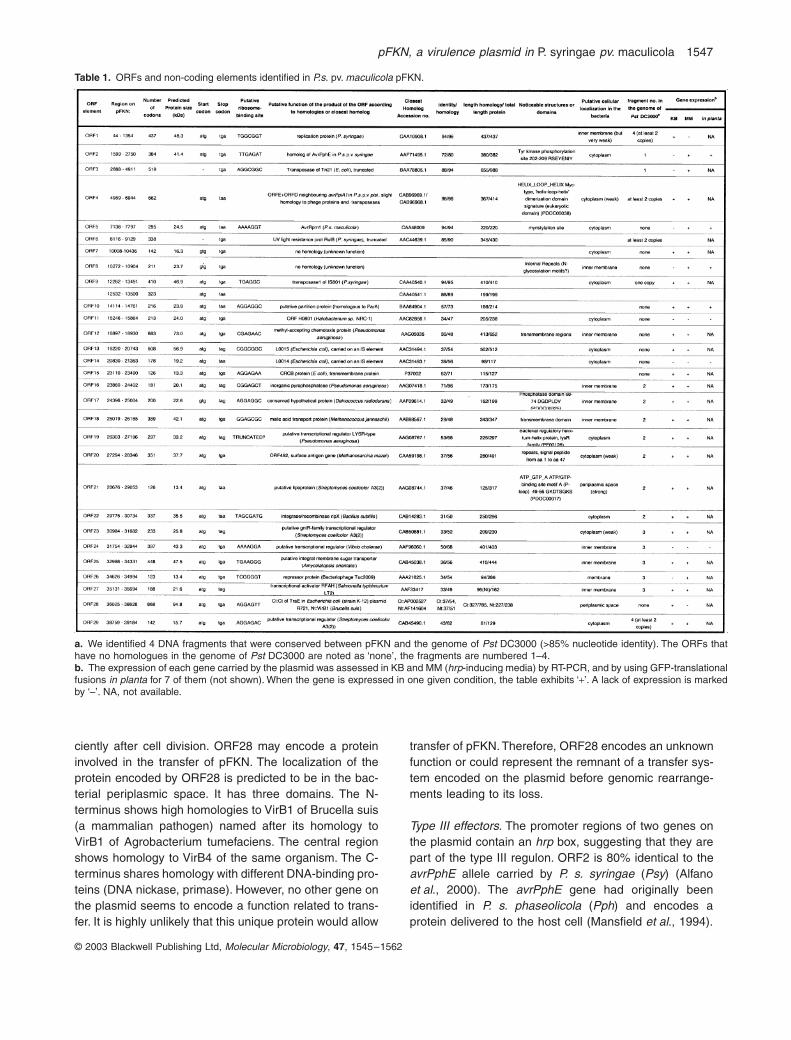
pFKN a virulence plasmid in
P syringae
pv
maculicola 1547
copy 2003 Blackwell Publishing Ltd
Molecular Microbiology
47
1545ndash1562
ciently after cell division ORF28 may encode a proteininvolved in the transfer of pFKN The localization of theprotein encoded by ORF28 is predicted to be in the bac-terial periplasmic space It has three domains The N-terminus shows high homologies to VirB1 of Brucella suis(a mammalian pathogen) named after its homology toVirB1 of Agrobacterium tumefaciens The central regionshows homology to VirB4 of the same organism The C-terminus shares homology with different DNA-binding pro-teins (DNA nickase primase) However no other gene onthe plasmid seems to encode a function related to trans-fer It is highly unlikely that this unique protein would allow
transfer of pFKN Therefore ORF28 encodes an unknownfunction or could represent the remnant of a transfer sys-tem encoded on the plasmid before genomic rearrange-ments leading to its loss
Type III effectors
The promoter regions of two genes onthe plasmid contain an
hrp
box suggesting that they arepart of the type III regulon ORF2 is 80 identical to the
avrPphE
allele carried by
P s syringae
(
Psy
) (Alfanoet al 2000) The avrPphE gene had originally beenidentified in P s phaseolicola (Pph) and encodes aprotein delivered to the host cell (Mansfield et al 1994)
Table 1 ORFs and non-coding elements identified in Ps pv maculicola pFKN
a We identified 4 DNA fragments that were conserved between pFKN and the genome of Pst DC3000 (gt85 nucleotide identity) The ORFs thathave no homologues in the genome of Pst DC3000 are noted as lsquononersquo the fragments are numbered 1ndash4b The expression of each gene carried by the plasmid was assessed in KB and MM (hrp-inducing media) by RT-PCR and by using GFP-translationalfusions in planta for 7 of them (not shown) When the gene is expressed in one given condition the table exhibits lsquo+rsquo A lack of expression is markedby lsquondashrsquo NA not available
1548 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Since then several alleles have been identified in otherpathovars of P syringae (Fouts et al 2002 Guttmanet al 2002) They all contain an hrp box in ORF2 ofpFKN this is found 51 bp upstream of the ATG at nt 1561as 5cent-GGAACTGA-N13-CGACATA-3cent (Innes et al1993) Based on the homologies and the presence ofthe hrp box in the promoter we hypothesized that theAvrPphE-like protein encoded by ORF2 is also a type IIIeffector (see below) AvrRpm1 (numbered ORF5) is iden-tical to that defined in the strain Psm M2 (Dangl et al1992)
Mobile element-related genes ORF3 (TnpA) encodes aputative truncated transposase Importantly avrPphE andTnpA are located between nearly identical invertedrepeats of 66 bp (Fig 1) thus defining a probable trans-posable element remnant The sequences surroundingavrRpm1 (ORF4 and ORF6) are homologous to asequence identified in P s pisi by Arnold et al (2001)with 96ndash97 nucleotide identity The DNA fragment car-rying these sequences in P s pisi may also be a trans-poson or a remnant It is therefore likely that avrRpm1 isalso located on the remnant of a transposon
ORF13 and ORF14 are related to ORFs found in theprophage 933 L on the LEE of the enterohaemorrhagic Ecoli O157H7 L0015 and L0014 respectively These twoORFs are thought to belong to an IS element from theIS66 family primarily identified in Sinorhizobium melilotithat includes IS elements from animal and plant patho-gens These two genes are usually associated with a thirdone L0013 in the LEE that is absent in pFKN (the
sequence upstream of ORF14 does not contain any ORFand does not show homology to L0013) Moreover the 5centend of ORF14 is also missing Therefore the sequenceencoding ORF13 and ORF14 may represent the remnantof an IS element belonging to the IS66 family not previ-ously identified in P syringae
Transcription factors Four pFKN genes share homologywith transcriptional regulators Some of these may func-tion during interactions with eukaryotic hosts For exam-ple the closest homologue to ORF24 is a putativetranscriptional factor from V cholerae ORF24 is alsorelated to putative proteins of unknown function (along thewhole length of the protein) found in organisms as diverseas E coli (50 identity67 homology) Caenorhabditiselegans (4157) Homo sapiens (4258) Droso-phila melanogaster (4058) and Arabidopsis thaliana(2942) A domain characteristic of FAD-dependentoxidoreductase (in Pfam) is detected in ORF24 There-fore the function of the translation product of this ORFmay require electron transfer through FAD
The protein encoded by ORF27 is a member of thesmall RfaH family RfaH controls the transcription of E coliand Salmonella operons that direct the synthesis assem-bly and export of the lipopolysaccharide core exopolysac-charide F-conjugation pilus and haemolysin toxin (for areview see Santangelo and Roberts 2002) RfaH isa specific regulator of transcript elongation its lossincreases transcription polarity in these operons withoutaffecting initiation from the operon promoters In additionin one case the RfaH protein controls the transcription of
Fig 1 Graphical map of pFKN and its 29 iden-tified ORFs The proposed direction of tran-scription is indicated by arrows Red boxes indicate genes encoding for type III effectors A black arrow line at the 5cent end of any genes indicates a possible hrp box Genes related to those of mobile elements are represented by green boxes The avrPphE transposon and the IS801 homologue are framed Transposon repeats surrounding the avrPphE transposon are marked by green lines The avrRpm1 gene is surrounded by repeats similar to the repeats of the transposon Tn501 these repeats are also marked by green lines Purple boxes indi-cate genes with replication and partition func-tions Genes encoding for putative transporters are indicated by blue boxes The putative tran-scriptional regulators are indicated by orange boxes The grey boxes represent genes that do not fall into any of the categories listed The number of each ORF is noted in bold on the outside of the circle
pFKN a virulence plasmid in P syringae pv maculicola 1549
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
a single gene encoding an outer membrane haeminreceptor ChuA in pathogenic E coli as well as othergenes involved in pathogenicity of the strain (Nagy et al2001 2002) The promoter of the genes or operons con-trolled by RfaH contains a conserved sequence calledJUMPStart (Hobbs and Reeves 1994) No JUMPStartsequence was detected on pFKN
Transporters ORF18 encodes a probable transmem-brane protein that is highly homologous to sugar trans-porters One is the product of mae1 a gene of the yeastSchizosaccharomyces pombe encoding a proteininvolved in the uptake of L-malate succinate and malonicacid The product of ORF18 may provide the bacteria witha fitness advantage in certain conditions
Other putative functions encoded on the plasmid Othergenes may encode proteins providing an advantage to thebacteria during the infection process For example theproduct of ORF12 displays all the characteristics ofmethyl-accepting chemotaxis proteins The genes encod-ing the flagellar unit and the chemotaxis functions areoften organized in clusters which is not the case onpFKN In this particular case the flagellar unit may beprovided in trans
pFKN is composed of different fragments that indicate a multistep process leading to its formation
The G+C content of the pFKN ORFs is highly variablealong the plasmid (Table 2) Yet several contiguous genesets on the plasmid have similar G+C contents This prob-ably indicates that these genes were acquired togetherfrom an original source The G+C content might allow usto track the multistep acquisition process resulting inpFKN For example ORFs 7 8 and 10 share nearly iden-tical G+C content (505) but are separated by an ISelement with a significantly different G+C content (603)To determine the distribution of pFKN ORFs in otherpathovars and how this distribution is related to G+Ccontent by ORF group we performed sequence compar-isons with the nucleotide sequence of the completegenomes of P syringae pv tomato DC3000 (httptigr-blasttigrorgufmg) and P syringae pv syringae B728a(httpwwwjgidoegovJGI_microbialhtmlindexhtml)DNA fragments carrying from two to seven contiguouspFKN ORFs were detected in both genome sequences(85ndash100 identity) The four DNA fragments from P stomato DC3000 carrying linked pFKN ORFs were num-bered and added to Table 2 Of these Pst DC3000 frag-ment 2 was also detected in Psy B728a (98 identical topFKN) In addition sequences homologous to repA andORF13 were also detected in this Psy B728a Table 2shows that ORFs belonging to the same contiguous DNA
fragment have a similar G+C content For example genesbelonging to fragment 4 have a relatively high G+C con-tent Genes belonging to fragments 1 and 2 have similarG+C contents and genes in fragment 3 exhibit a lowerG+C content Thus the different fragments constitutingpFKN were probably acquired by the three genomes indifferent steps followed by subsequent sequence dele-tions andor rearrangements with respect to Psm M6 asthe entire collinear pFKN is not detected in either PstDC3000 or Psy B728A
Full virulence of PsmM6 requires pFKN and is essentially caused by avrRpm1
To determine the contribution to virulence of the plasmidpFKN we cured Psm M6 of pFKN (see Experimentalprocedures) to generate strain Psm M6C There was noobvious macroscopic difference in visible disease symp-toms after inoculation of either wild-type or Psm M6Conto rpm1-3 Arabidopsis leaves (unable to recognizeavrRpm1) At high levels of inoculation (5 yen 106 cfu ml-1)the inoculated leaves exhibited water soaking and tissuecollapse starting at 24 h after inoculation Inoculation at5 yen 105 cfu ml-1 led to chlorosis between 48 and 72 h afterinoculation Using this low-density inoculum we mea-sured pathogen growth over time in rpm1-3 leaves PsmM6C grew 10-fold less than the wild-type strain (Fig 2A)Therefore the cured strain is less virulent than the wild-type strain We performed complementation experimentsusing various pFKN subclones in Psm M6C to determinewhich pFKN ORFs would partially or fully restore viru-lence Only clones carrying avrRpm1 could restore fullvirulence (Fig 2A) This result is consistent with inser-tional mutation of avrRpm1 in the Psm M2 strain (Ritterand Dangl 1995) No other clone conferred enhancedvirulence (data not shown)
The role of avrPphE in virulence could not be assessedusing Psm M6C because a second copy of the gene ispresent on the chromosome of the cured strain [asassessed by polymerase chain reaction (PCR) amplifica-tion and sequencing of the PCR product see below] Wereplaced the avrPphE copy on the chromosome of PsmM6C with an omega fragment to create Psm M6CKO (seeExperimental procedures) Psm M6CKO grew similarly toPsm M6C on rpm1-3 plants (Fig 2B) suggesting thatavrPphE plays no major role in the virulence of Psm M6However our recent results indicate that expression ofavrPphE from the lac-nptII promoter in PsmM6CKO doesenhance virulence of PsmM6CKO suggesting that thequantity of AvrPphE protein delivered to the host cell maybe important in its function (J Shock J Chang Z Nim-chuk L Rohmer and J L Dangl unpublished) As avrP-phE was located in the vicinity of avrRpm1 on pFKN wetested the potential involvement of avrPphE in the viru-
1550 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
lence function of avrRpm1 We compared the growth ofPsm M6C and Psm M6CKO when each carried avrRpm1on a plasmid No difference in growth was observed(Fig 2B) indicating that avrPphE does not affectavrRpm1 virulence function
Identification of additional genes putatively involved in pathogenicity based on their expression pattern
The expression of known P syringae virulence factors isspecifically induced in planta Minimal media (MM) plus asugar source seems to mimic the plant apoplast leadingto the expression of the type III pilus genes and effectors
(Huynh et al 1989 Innes et al 1993) We compared theexpression of the pFKN ORFs in MM and in rich medium(KB) using reverse transcription RT-PCR methodsWhen the expression of an ORF was observed in bothmedia we assumed that the gene was constitutivelyexpressed Expression of a gene in MM but not in KB mayindicate a role for this gene during interaction with thehost Most of the pFKN genes are constitutivelyexpressed However the expression of both avrRpm1(Ritter and Dangl 1995) and avrPphE was detected inMM but not in KB Additionally ORF26 is expressed inMM but not in KB ORFs 14 and 24 were not expressedin either of the two conditions The expression of ORF8
Table 2 pFKN ORFs sorted by descending G+C content
a Some pFKN ORFs are contiguous on the plasmid and are collinear in this tableindicating that they possess a similar G+C content When this is the case an emptyrow separates them from the next ORF in the tableb The DNA fragments from the Pst DC3000 genome that contain sequenceshomologous to ORFs from pFKN were colour-coded (1 green 2 red 3 blue4 pink) The table indicates that fragment 4 has a relatively high G+C contentwhereas fragment 3 has a relatively low content
pFKN a virulence plasmid in P syringae pv maculicola 1551
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
and ORF11 could not be assessed using this techniquedespite many attempts
The six genes (avrRpm1 avrPphE ORF8 ORF11ORF14 and ORF24) for which no constitutive expressionwas observed by RT-PCR were selected for further studyWe constructed transcriptional fusions between the 5cent endof the genes of interest and a gene encoding green fluo-rescent protein (GFP) and chose ORF10 as a constitutivecontrol The fluorescence of PsmM6C carrying the tran-scriptional fusions was assessed in KB MM and in plantaby exposing the bacteria grown in these conditions to bluelight (488 nm) and observing them by fluorescencemicroscopy A strain carrying promoterless GFP did notexhibit any fluorescence in the conditions tested whereas
all cells carrying the ORF10ndashGFP fusion did Only theavrPphE avrRpm1 and ORF8 GFP fusions were active inplants (Table 1)
pFKN can integrate into the Psm M6 chromosome
One colony of Psm M6 derived during routine mainte-nance exhibited unusual features Although this straintriggered recognition (measured by HR) on RPM1 plantsand disease symptoms on rpm1 plants no plasmid couldbe isolated from it AvrRpm1 was still present in totalDNA as assayed by DNA blot (Fig 3A) We thereforeconcluded that avrRpm1 was now located on the PsmM6 chromosome To determine whether additional por-tions of pFKN were also present on the variance chromo-some total DNA from the variant Psm M6 was digestedwith EcoRI HindIII EcoRV SacI PvuII SmaI BglII andNsiI and probed with the DNA from different parts of thepFKN plasmid The DNA of Psm M6 containing pFKNwas used as a positive control This analysis revealed apolymorphism between the avrRpm1 gene in this strain(eg an 80 kb BglII fragment) compared with pFKN (egan 88 kb BglII fragment Fig 3A) All restriction frag-ments from pFKN were detected in the DNA of PsmM6integron Therefore pFKN has integrated into thechromosome For the purpose of clarity the strain bear-ing avrRpm1 in the chromosome will be referred to asPsm M6integron
We have on occasion isolated a replicon from indi-vidual colonies of Psm M6integron (see next section)The plasmid purified from these bacteria always exhib-ited a restriction profile identical to that of pFKNTherefore pFKN can excise from the chromosome andPsm M6integron can revert to Psm M6 Interestinglythe overall fitness of Psm M6 is affected by the integra-tion status of pFKN into the chromosome PsmM6integron grows in media (KB and MM) more slowlythan either the original Psm M6 strain or a revertantfrom Psm M6integron that is cured of the plasmid(Fig 3B) Because Psm M6 (with a pFKN replicon) andthe plasmid-cured derivative of Psm M6integron bothgrew equivalently we can conclude that it is the inte-gration of pFKN per se that results in this growthimpairment This is not uncommon as several plasmidreplicons are known to slow growth of the organismwhen integrated (see Discussion)
The pFKN integration site in Psm M6integron is the avrPphE transposon
We noticed from the DNA blot that the genomic environ-ment of avrRpm1 is different in Psm M6 and PsmM6integron This is also the case for both copies of avrP-phE (data not shown) suggesting that the pFKN integra-tion site is near avrRpm1 and avrPphE and that the
Fig 2 avrRpm1 encodes the determining virulence factor on pFKNA The growth of Psm M6C on rpm1-3 plants was compared with the growth of the wild-type Psm M6 strain and Psm M6C strains comple-mented with different fragments from pFKN The pFKN subclone carrying avrRpm1 restored full virulence to Psm M6C (the other subclones did not restore any virulence and are not shown) Leaves were hand inoculated with 105 cfu ml-1 and a titration of bacteria in the leaves was performed 3 days after inoculation Data points rep-resent the mean plusmn standard error of four samplesB The avrPphE gene was deleted from the chromosome of Psm M6C to generate Psm M6CKO To assess the virulence function of avrPphE or its influence on the virulence function of avrRpm1 the growth of Psm M6CKO on rpm1-3 plants was compared with the growth of Psm M6C Psm M6C + avrRpm1 and Psm M6CKO + avrRpm1 Data points represent the mean plusmn standard error of four samples
1552 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
chromosomal location into which the plasmid integrates isnear the chromosomal copy of avrPphE We first definedthat the chromosomal avrPphE is carried on the sametransposon found in the sequence of pFKN using PCRamplification on total DNA of Psm M6C with primersspecific to sequence on the avrPphE transposon(Table 5) The PCR products were sequenced and com-pared with the corresponding sequenced regions ofpFKN As shown in Fig 4A the chromosomal copy of thetransposon contains two sequences not present on pFKNOne 1244 bp sequence (region A) is located betweenavrPphE and the coding region of TnpA It carries an ORFwith a sequence that is nearly identical to various genesencoding resolvases of Tn3 transposons Additionally aportion of 661 bp of this region encodes the 5cent portion ofTnpA Another region of 271 bp unique to the chromo-somal copy is located within the coding region of thetransposase (region C) and is 3cent from a region shared byboth pFKN and chromosomal copies of TnpA (region B)The addition of regions A and C to the chromosomal copyresults in a complete TnpA ORF correcting the frameshiftat the 3cent end of TnpA carried on pFKN
TnpA avrPphE and the putative resolvase fragmentare located between nearly identical inverted repeats of66 bp on both the plasmid and the chromosome Theirorganization is reminiscent of Tn3-like transposons(Sherrat 1989) Moreover this TnpA is 81 identical tothose of the Tn21 subfamily of Tn3 elements (Diveret al 1983) As we suspected that the integration site isin the vicinity of avrPphE in the chromosome and theplasmid (ie in the two transposons) we analysed byPCR the two transposons in the genome of PsmM6integron Figure 4B shows that the unique regions Aand C are no longer located on the same transposonbut are instead separated by the entire plasmidsequence (Fig 4C) This indicated that the integrationsite of the plasmid is located in the region betweenregion A and region C ie within the homology providedby region B (Fig 4)
We used this sequence data to investigate the rateof excision occurring in Psm M6integron grown indifferent conditions by PCR The rate of excision wasestimated at around 10-4 (data not shown) No differencein rate of excision could be detected between bacteria
Fig 3 pFKN can integrate into the Psm M6 chromosomeA Total DNA from Psm M6 (lanes F) and Psm M6integron (lanes I) was digested with EcoRI HindIII EcoRV SacI PvuII SmaI BglII and NsiI and probed with various pFKN fragments Here the DNA blot was probed with the avrRpm1 ORF The two DNA samples exhibit a similar pattern except for a polymorphism (eg EcoRI BglII and SmaI) near the site of pFKN integra-tion into the chromosomeB The integration of the plasmid alters the overall fitness of Psm M6integron The growth of Psm M6integron in KB (empty symbols) and MM (filled symbols) was compared with the growth of Psm M6 and Psm M6C Bacteria were inoculated at 104 cfu ml-1 and cultures were titrated at 0 6 12 29 32 54 and 60 h after the inoculation Psm M6pFKN is indicated by squares Psm M6C is indicated by triangles and rhombi indicate Psm M6integron Data points represent the mean plusmn standard error of four samples (some error bars are not visible because too small) Psm M6pFKN and Psm M6C grew equally well in both minimal medium and KB Psm M6integron grows slower than Psm M6pFKN and Psm M6C in both media
pFKN a virulence plasmid in P syringae pv maculicola 1553
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
grown in KB MM and on resistant or susceptibleplants
The genomic context of avrPphE is different in various P syringae strains
We investigated the regions surrounding the avrPphEtransposon on the chromosome of Psm M6 (Fig 5) byTAIL-PCR The left border shows 97 nt identity to the 5centend of the insertion sequence ISPsy3 (AB063176) fromP syringae pv eriobotryae This element is interrupted bythe avrPphE transposon The translation product of theinterrupted ORF found on this border is homologous tothe IS801 transposase (79 identity and 86 homology)
and IS801 has been associated with other type III effec-tors in various P syringae pathovars (Kim et al 1998)On the right border the 200 bp sequence following theterminal repeat of the avrPphE transposon encodes aprotein homologous (81 nucleotide identity) to the 3centend of an unrelated putative transposase gene fromPseudomonas aeruginosa The next 1062 bp sequencedownstream shows high nucleotide homology tosequence identified in P syringae pv pisi (Arnold et al2001) This DNA fragment may be a transposon remnantand carries the avrPpiA1 allele of avrRpm1 from P syrin-gae pv pisi among other genes In the chromosome ofPsm M6 this sequence shows homology to the P s pisigenes ORFD (94 identity) encoding a putative trans-
Fig 4 The pFKN integration site is a second copy of the avrPphE transposonA The region in the Psm M6 chromosome that hybridizes to the avrPphE and TnpA probes was amplified and sequenced The sequence of the chromosomal region is 100 identical to the transposon in the plasmid but contains two additional regions (regions A and C) TnpA encodes a transposase and res a resolvase Based on homology the chromosomal avrPphE transposon could be complete and functional The avrPphE transposon carried on the plas-mid lacks the 5cent end of TnpA and res (region A) and the absence of region C results in the truncation and a frameshift (marked FS) of the gene encoding the transposase The primers used for PCR amplification (Fig 4B) are indi-cated by arrows and their names are in boldB To investigate where regions A and C were distributed after integration of the plasmid into the chromosome PCRs were performed on the DNA of Psm M6integron and Psm M6 using Larp3 and K110 and K110 and K113 (see Fig 4A) The product amplified from Psm M6integron is run in lane I The product ampli-fied from Psm M6 is run in lane F The product amplified from Psm M6C is run in lane C The difference in size observed between the PCR products from Psm M6integron and Psm M6 using Larp3 and K110 indicates that regions A and C are not carried on the same putative transposon in the chromosome of Psm M6integron (in contrast to the chromosome of Psm M6) The PCR products from Psm M6integron and Psm M6 using K113 and K110 show that neither region A nor region C has been lost in the chromosome of Psm M6integron The identical size of the PCR prod-ucts from Psm M6integron and Psm M6 using K113 and Larp2 indicates that region C is on the right border of pFKN when integrated into the chromosome The region A is on the left border as it is carried by the other putative transposonC After integration of pFKN into the chromo-some TnpA on the left border contains region A but not region C TnpA on the right border contains region C The primers used for the PCR amplification are indicated by arrows and their names are in bold (in Fig 4A)
1554 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
posase and ORF3 encoding a putative type III effector(100 identity) Thus this region in Psm M6 is a remnantof the putative transposon found in P s pisi
We detected the avrPphE transposon in other strains ofP syringae pv maculicola and in the sequence of the
genome of P s tomato DC3000 In Psm M2 Psm M8Psm M12 Psm m90 and Psm m10831 the borders arethe same as in Psm M6 (data not shown) On the otherhand they differ in Psm M5 and Psm M9 and these alldiffer from Pst DC3000 In Psm M5 (Fig 5) and Psm M9
Fig 5 Sequences surrounding the avrPphE transposon define four different genetic contexts The figure is a graphical representation of the four genomic environments in which the avrPphE transposon was detected The regions represented were characterized by sequence comparison using the BLASTX algorithm It is not known whether the avrPphE transposon is located in the chromosome or on a plasmid in the genome of Psm M5 and Pst DC3000
pFKN a virulence plasmid in P syringae pv maculicola 1555
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(not shown) the left border consists of the 5cent end of agene sharing 99 identity to the replicase of pPS10 aplasmid found in P syringae pv savastanoi (Nieto et al1992) This replicase is not related to that of pFKN (aPT23A-like plasmid see above) The right bordersequence is a remnant of a gene encoding a putative ABCtransporter (23 identity and 43 homology to a putativesugar uptake ABC transporter ATP-binding protein fromSinorhizobium meliloti)
In the genome of Pst DC3000 we looked at 30 kbsurrounding the transposon (Fig 5) The proteinsencoded on this 30 kb region share homology to eitherDNA mobility functions (transposases plasmid partition-ing proteins replicases) or type III effectors (identical toavrPpiB Cournoyer et al 1995) The region also containsother sequences often found associated with plasmidsand genes involved in pathogenicity such as rulA and rulB(Sesma et al 2000) Sequences directly neighbouringthe transposon on the left side are homologous to a geneencoding the transposase of the IS100 elements fromYersinia pestis Ralstonia solanacearum and Salmonellaenterica On the right-hand side of the avrPphE transpo-son the first homology is to a fragment of a putative ABCtransporter identical to the one identified in Psm M5 Thissuggests that this part of the two genomes is morerecently diverged than either is from Psm M6
No common sequence was identified in the four differ-ent environments surrounding the avrPphE transposonthat would help to identify the integration target of theavrPphE transposon although the proximal right bordersin Psm M5 and Pst DC3000 are nearly identical Thissuggests that the avrPphE transposon and this border-ing sequence were acquired together by these twostrains and that subsequent events at both the distalright flank and on the left border are responsible for thedifferences in genomic context In contrast the insertionsites of the avrPphE transposon in Psm M5 and PsmM6 are completely different In sum the features ofthese integration sites are consistent with the model thatPAIs evolve by repeated insertion and deletion intopre-existing PAIs (Hensel et al 1999 Tauschek et al2002)
Discussion
We sequenced pFKN an ordf40 kb plasmid that contributesto the virulence of Psm M6 The plasmid carries two typeIII effectors on remnants of transposons contiguous toORF8 itself neighbouring an element closely related toIS801 avrRpm1 encodes a known virulence factor andavrPphE a type III effector The expression of ORF8 isinduced in planta and therefore ORF8 may play a role inthe interaction with the host plant The portion of theplasmid carrying avrRpm1 avrPphE and ORF8 may thus
be considered as a PAI Interestingly the G+C content ofthe ORFs carried on this region are very heterogeneousand the three genes are flanked by sequences related tomobile elements and therefore are likely to have differentorigins This is consistent with the idea that the formationof PAIs requires a multistep process (Hensel et al 1999Alfano et al 2000 Tauschek et al 2002) It is also con-sistent with the findings of Kim et al (1998) showing thatmost of the known type III effectors are associated withthis type of sequence pFKN (and therefore the PAI that itcarries) sometimes integrates into and can excise fromthe chromosome as demonstrated by the discovery of thevariant of the strain Psm M6 We identified the region ofthe chromosome into which the integration took placeThis region is a transposon carrying a second copy ofavrPphE
Alleles of the avrPphE gene have been detected inmany P syringae pathovars (Mansfield et al 1994Alfano et al 2000 Fouts et al 2002 Guttman et al2002) suggesting that it provides an important virulencefunction on a variety of host plants Our findings are thefirst to associate avrPphE with a transposon In PsyB728a and in Pph 1302A for example avrPphE residesdirectly next to the hrp locus in a region called thelsquoexchangeable effector locusrsquo or EEL (Alfano et al 2000)and is not associated with a transposase In fact in PsyB728a avrPphE is predicted to be the third gene in anoperon controlled by the hrpK promoter (Alfano et al2000) This striking finding illustrates the dynamism ofPAIs
We determined by complementation assay that the par-tial loss of virulence in the strain cured of pFKN resultsfrom the absence of avrRpm1 The virulence function ofavrRpm1 has already been demonstrated in the strainPsm M2 (Ritter and Dangl 1995) Recently AvrRpm1 hasbeen demonstrated to interact with RIN4 a proteininvolved in the regulation of some of the plant defenceresponses (Mackey et al 2002) Mackey and coworkersproposed that the role of AvrRpm1 is to reduce the levelof plant defence response triggered by the attack of thepathogen This hypothesis might be consistent with thefact that only quantitative differences in growth wereobserved between Psm M6C and Psm M6 No differencein growth was observed between Psm M6C and PsmM6CKO (lacking avrPphE) Similar results were observedwith the avrPphE allele of P s phaseolicola (Stevenset al 1998) This observation could be explained byeither a functional redundancy with another gene in thegenome of the Psm M6 or the fact that the assay isinappropriate to assess the virulence function of this typeIII effector
The sequence and the integration mechanism of pFKNinto the Psm M6 chromosome provide insights on theevolutionary dynamics of the genomes of P syringae
1556 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
pathovars First the analysis of the G+C content of frag-ments of pFKN suggests that they may have differentorigins This hypothesis is further supported by the pres-ence of some defined fragments from pFKN in thegenomes of Pst DC3000 and Psy B728a The fragment 3(see Table 2) encodes one putative transcriptional regu-lator proteins predicted to be located in the bacterialinner or outer membrane by PSORT (inorganic pyrophos-phatase sugar transporter putative protein) and a puta-tive integraserecombinase homologous to ripX fromBacillus subtilis The presence of this latter gene sug-gests that fragment 3 may be the remnant of abacteriophage
The integration and excision mechanism is very proba-bly a homologous recombination in the 15 kb sequenceregion B of TnpA carried on the avrPphE transposon(Fig 6) It is not clear whether the integrationexcisionprocess relies on general recombination or on a site-specific recombination dependent on an integraserecombinase As indicated by the sequence of the twotransposons before and after integration of pFKN nonucleotide sequences were altered at the site ofexchange and not a single nucleotide was lost or gained(this excludes a mechanism of transpositional site-specificrecombination) General recombination requires a largeDNA region whereas the site-specific recombinationrequires as few as 240 bp for the lambda phage for exam-ple (Thompson and Landy 1989) The rate of generalrecombination is unknown for P syringae but has beenestimated at 10-3 in Pseudomonas tolaasii (and whenrecA is inactivated at 10-4 Sinha et al 2000) The rateof excision of pFKN has been estimated between 10-3 and10-4 and would therefore be consistent with a mechanismof general recombination Additionally the region B is
15 kb long enough to ensure general recombinationOlder reports describe the integration of pMMC7501 intothe chromosome of P s pv phaseolicola (Curiale andMills 1982) An imprecise excision of pMMC7501 led tothe formation of eight derivative plasmids all sharing thesame origin of replication (Szabo and Mills 1984a) Thisexcision occurred through recombination events betweensets of repeats in the chromosome exhibiting sequencehomology with IS elements (Szabo and Mills 1984bPoplawsky and Mills 1987)
The integration of a virulence gene into the chromo-some may be a first step towards its stabilization in thegenome Indeed lack of selection can lead to the stochas-tic loss of the plasmid and many steps in the pathogenlife cycle probably do not require the presence of virulencefunctions The loss of a plasmid carrying the type III effec-tor avrPpiB was observed in P syringae pv pisi (Bavageet al 1991) We propose that the integration of pFKNcould lead to the stabilization of avrRpm1 in the genomeof Psm M6 Interestingly a DNA segment of at least 42 kbcarrying avrPphB in P syringae pv phaseolicola canexcise from the chromosome but it is not maintained inthe strain as a plasmid This region may represent anancient plasmid integration event followed by successivegenomic rearrangements deleterious for plasmid mainte-nance (Jackson et al 2000) The fact that we can stillobserve the excision of pFKN from the chromosome ofPsm M6integron shows that the newly generated geneticunit is not rapidly stabilized in the genome and that furtherevents might be necessary to prevent excision Further-more the integration of pFKN into the chromosomeslightly impairs the fitness of Psm M6integron The pres-ence of the functional origin of replication of pFKN mayinterfere with the replication of the chromosome Integra-
Fig 6 The integration of pFKN may have occurred via homologous recombination between the regions B common to the two TnpA genes The TnpA genes are constituted of four DNA sequence regions (1) before the integration the chromosomal copy of TnpA contains all four regions (A B C and D) The TnpA on the plasmid is missing the 5cent end of the gene (region A) and a fragment towards the 3cent end (region C) which results in a frameshift (FS) After the homologous recombination between two identical regions (region B) in the coding region of the TnpA genes (2) the region A is found in the TnpA of the left border and region C in the TnpA of the right border (3)
pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

1548 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Since then several alleles have been identified in otherpathovars of P syringae (Fouts et al 2002 Guttmanet al 2002) They all contain an hrp box in ORF2 ofpFKN this is found 51 bp upstream of the ATG at nt 1561as 5cent-GGAACTGA-N13-CGACATA-3cent (Innes et al1993) Based on the homologies and the presence ofthe hrp box in the promoter we hypothesized that theAvrPphE-like protein encoded by ORF2 is also a type IIIeffector (see below) AvrRpm1 (numbered ORF5) is iden-tical to that defined in the strain Psm M2 (Dangl et al1992)
Mobile element-related genes ORF3 (TnpA) encodes aputative truncated transposase Importantly avrPphE andTnpA are located between nearly identical invertedrepeats of 66 bp (Fig 1) thus defining a probable trans-posable element remnant The sequences surroundingavrRpm1 (ORF4 and ORF6) are homologous to asequence identified in P s pisi by Arnold et al (2001)with 96ndash97 nucleotide identity The DNA fragment car-rying these sequences in P s pisi may also be a trans-poson or a remnant It is therefore likely that avrRpm1 isalso located on the remnant of a transposon
ORF13 and ORF14 are related to ORFs found in theprophage 933 L on the LEE of the enterohaemorrhagic Ecoli O157H7 L0015 and L0014 respectively These twoORFs are thought to belong to an IS element from theIS66 family primarily identified in Sinorhizobium melilotithat includes IS elements from animal and plant patho-gens These two genes are usually associated with a thirdone L0013 in the LEE that is absent in pFKN (the
sequence upstream of ORF14 does not contain any ORFand does not show homology to L0013) Moreover the 5centend of ORF14 is also missing Therefore the sequenceencoding ORF13 and ORF14 may represent the remnantof an IS element belonging to the IS66 family not previ-ously identified in P syringae
Transcription factors Four pFKN genes share homologywith transcriptional regulators Some of these may func-tion during interactions with eukaryotic hosts For exam-ple the closest homologue to ORF24 is a putativetranscriptional factor from V cholerae ORF24 is alsorelated to putative proteins of unknown function (along thewhole length of the protein) found in organisms as diverseas E coli (50 identity67 homology) Caenorhabditiselegans (4157) Homo sapiens (4258) Droso-phila melanogaster (4058) and Arabidopsis thaliana(2942) A domain characteristic of FAD-dependentoxidoreductase (in Pfam) is detected in ORF24 There-fore the function of the translation product of this ORFmay require electron transfer through FAD
The protein encoded by ORF27 is a member of thesmall RfaH family RfaH controls the transcription of E coliand Salmonella operons that direct the synthesis assem-bly and export of the lipopolysaccharide core exopolysac-charide F-conjugation pilus and haemolysin toxin (for areview see Santangelo and Roberts 2002) RfaH isa specific regulator of transcript elongation its lossincreases transcription polarity in these operons withoutaffecting initiation from the operon promoters In additionin one case the RfaH protein controls the transcription of
Fig 1 Graphical map of pFKN and its 29 iden-tified ORFs The proposed direction of tran-scription is indicated by arrows Red boxes indicate genes encoding for type III effectors A black arrow line at the 5cent end of any genes indicates a possible hrp box Genes related to those of mobile elements are represented by green boxes The avrPphE transposon and the IS801 homologue are framed Transposon repeats surrounding the avrPphE transposon are marked by green lines The avrRpm1 gene is surrounded by repeats similar to the repeats of the transposon Tn501 these repeats are also marked by green lines Purple boxes indi-cate genes with replication and partition func-tions Genes encoding for putative transporters are indicated by blue boxes The putative tran-scriptional regulators are indicated by orange boxes The grey boxes represent genes that do not fall into any of the categories listed The number of each ORF is noted in bold on the outside of the circle
pFKN a virulence plasmid in P syringae pv maculicola 1549
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
a single gene encoding an outer membrane haeminreceptor ChuA in pathogenic E coli as well as othergenes involved in pathogenicity of the strain (Nagy et al2001 2002) The promoter of the genes or operons con-trolled by RfaH contains a conserved sequence calledJUMPStart (Hobbs and Reeves 1994) No JUMPStartsequence was detected on pFKN
Transporters ORF18 encodes a probable transmem-brane protein that is highly homologous to sugar trans-porters One is the product of mae1 a gene of the yeastSchizosaccharomyces pombe encoding a proteininvolved in the uptake of L-malate succinate and malonicacid The product of ORF18 may provide the bacteria witha fitness advantage in certain conditions
Other putative functions encoded on the plasmid Othergenes may encode proteins providing an advantage to thebacteria during the infection process For example theproduct of ORF12 displays all the characteristics ofmethyl-accepting chemotaxis proteins The genes encod-ing the flagellar unit and the chemotaxis functions areoften organized in clusters which is not the case onpFKN In this particular case the flagellar unit may beprovided in trans
pFKN is composed of different fragments that indicate a multistep process leading to its formation
The G+C content of the pFKN ORFs is highly variablealong the plasmid (Table 2) Yet several contiguous genesets on the plasmid have similar G+C contents This prob-ably indicates that these genes were acquired togetherfrom an original source The G+C content might allow usto track the multistep acquisition process resulting inpFKN For example ORFs 7 8 and 10 share nearly iden-tical G+C content (505) but are separated by an ISelement with a significantly different G+C content (603)To determine the distribution of pFKN ORFs in otherpathovars and how this distribution is related to G+Ccontent by ORF group we performed sequence compar-isons with the nucleotide sequence of the completegenomes of P syringae pv tomato DC3000 (httptigr-blasttigrorgufmg) and P syringae pv syringae B728a(httpwwwjgidoegovJGI_microbialhtmlindexhtml)DNA fragments carrying from two to seven contiguouspFKN ORFs were detected in both genome sequences(85ndash100 identity) The four DNA fragments from P stomato DC3000 carrying linked pFKN ORFs were num-bered and added to Table 2 Of these Pst DC3000 frag-ment 2 was also detected in Psy B728a (98 identical topFKN) In addition sequences homologous to repA andORF13 were also detected in this Psy B728a Table 2shows that ORFs belonging to the same contiguous DNA
fragment have a similar G+C content For example genesbelonging to fragment 4 have a relatively high G+C con-tent Genes belonging to fragments 1 and 2 have similarG+C contents and genes in fragment 3 exhibit a lowerG+C content Thus the different fragments constitutingpFKN were probably acquired by the three genomes indifferent steps followed by subsequent sequence dele-tions andor rearrangements with respect to Psm M6 asthe entire collinear pFKN is not detected in either PstDC3000 or Psy B728A
Full virulence of PsmM6 requires pFKN and is essentially caused by avrRpm1
To determine the contribution to virulence of the plasmidpFKN we cured Psm M6 of pFKN (see Experimentalprocedures) to generate strain Psm M6C There was noobvious macroscopic difference in visible disease symp-toms after inoculation of either wild-type or Psm M6Conto rpm1-3 Arabidopsis leaves (unable to recognizeavrRpm1) At high levels of inoculation (5 yen 106 cfu ml-1)the inoculated leaves exhibited water soaking and tissuecollapse starting at 24 h after inoculation Inoculation at5 yen 105 cfu ml-1 led to chlorosis between 48 and 72 h afterinoculation Using this low-density inoculum we mea-sured pathogen growth over time in rpm1-3 leaves PsmM6C grew 10-fold less than the wild-type strain (Fig 2A)Therefore the cured strain is less virulent than the wild-type strain We performed complementation experimentsusing various pFKN subclones in Psm M6C to determinewhich pFKN ORFs would partially or fully restore viru-lence Only clones carrying avrRpm1 could restore fullvirulence (Fig 2A) This result is consistent with inser-tional mutation of avrRpm1 in the Psm M2 strain (Ritterand Dangl 1995) No other clone conferred enhancedvirulence (data not shown)
The role of avrPphE in virulence could not be assessedusing Psm M6C because a second copy of the gene ispresent on the chromosome of the cured strain [asassessed by polymerase chain reaction (PCR) amplifica-tion and sequencing of the PCR product see below] Wereplaced the avrPphE copy on the chromosome of PsmM6C with an omega fragment to create Psm M6CKO (seeExperimental procedures) Psm M6CKO grew similarly toPsm M6C on rpm1-3 plants (Fig 2B) suggesting thatavrPphE plays no major role in the virulence of Psm M6However our recent results indicate that expression ofavrPphE from the lac-nptII promoter in PsmM6CKO doesenhance virulence of PsmM6CKO suggesting that thequantity of AvrPphE protein delivered to the host cell maybe important in its function (J Shock J Chang Z Nim-chuk L Rohmer and J L Dangl unpublished) As avrP-phE was located in the vicinity of avrRpm1 on pFKN wetested the potential involvement of avrPphE in the viru-
1550 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
lence function of avrRpm1 We compared the growth ofPsm M6C and Psm M6CKO when each carried avrRpm1on a plasmid No difference in growth was observed(Fig 2B) indicating that avrPphE does not affectavrRpm1 virulence function
Identification of additional genes putatively involved in pathogenicity based on their expression pattern
The expression of known P syringae virulence factors isspecifically induced in planta Minimal media (MM) plus asugar source seems to mimic the plant apoplast leadingto the expression of the type III pilus genes and effectors
(Huynh et al 1989 Innes et al 1993) We compared theexpression of the pFKN ORFs in MM and in rich medium(KB) using reverse transcription RT-PCR methodsWhen the expression of an ORF was observed in bothmedia we assumed that the gene was constitutivelyexpressed Expression of a gene in MM but not in KB mayindicate a role for this gene during interaction with thehost Most of the pFKN genes are constitutivelyexpressed However the expression of both avrRpm1(Ritter and Dangl 1995) and avrPphE was detected inMM but not in KB Additionally ORF26 is expressed inMM but not in KB ORFs 14 and 24 were not expressedin either of the two conditions The expression of ORF8
Table 2 pFKN ORFs sorted by descending G+C content
a Some pFKN ORFs are contiguous on the plasmid and are collinear in this tableindicating that they possess a similar G+C content When this is the case an emptyrow separates them from the next ORF in the tableb The DNA fragments from the Pst DC3000 genome that contain sequenceshomologous to ORFs from pFKN were colour-coded (1 green 2 red 3 blue4 pink) The table indicates that fragment 4 has a relatively high G+C contentwhereas fragment 3 has a relatively low content
pFKN a virulence plasmid in P syringae pv maculicola 1551
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
and ORF11 could not be assessed using this techniquedespite many attempts
The six genes (avrRpm1 avrPphE ORF8 ORF11ORF14 and ORF24) for which no constitutive expressionwas observed by RT-PCR were selected for further studyWe constructed transcriptional fusions between the 5cent endof the genes of interest and a gene encoding green fluo-rescent protein (GFP) and chose ORF10 as a constitutivecontrol The fluorescence of PsmM6C carrying the tran-scriptional fusions was assessed in KB MM and in plantaby exposing the bacteria grown in these conditions to bluelight (488 nm) and observing them by fluorescencemicroscopy A strain carrying promoterless GFP did notexhibit any fluorescence in the conditions tested whereas
all cells carrying the ORF10ndashGFP fusion did Only theavrPphE avrRpm1 and ORF8 GFP fusions were active inplants (Table 1)
pFKN can integrate into the Psm M6 chromosome
One colony of Psm M6 derived during routine mainte-nance exhibited unusual features Although this straintriggered recognition (measured by HR) on RPM1 plantsand disease symptoms on rpm1 plants no plasmid couldbe isolated from it AvrRpm1 was still present in totalDNA as assayed by DNA blot (Fig 3A) We thereforeconcluded that avrRpm1 was now located on the PsmM6 chromosome To determine whether additional por-tions of pFKN were also present on the variance chromo-some total DNA from the variant Psm M6 was digestedwith EcoRI HindIII EcoRV SacI PvuII SmaI BglII andNsiI and probed with the DNA from different parts of thepFKN plasmid The DNA of Psm M6 containing pFKNwas used as a positive control This analysis revealed apolymorphism between the avrRpm1 gene in this strain(eg an 80 kb BglII fragment) compared with pFKN (egan 88 kb BglII fragment Fig 3A) All restriction frag-ments from pFKN were detected in the DNA of PsmM6integron Therefore pFKN has integrated into thechromosome For the purpose of clarity the strain bear-ing avrRpm1 in the chromosome will be referred to asPsm M6integron
We have on occasion isolated a replicon from indi-vidual colonies of Psm M6integron (see next section)The plasmid purified from these bacteria always exhib-ited a restriction profile identical to that of pFKNTherefore pFKN can excise from the chromosome andPsm M6integron can revert to Psm M6 Interestinglythe overall fitness of Psm M6 is affected by the integra-tion status of pFKN into the chromosome PsmM6integron grows in media (KB and MM) more slowlythan either the original Psm M6 strain or a revertantfrom Psm M6integron that is cured of the plasmid(Fig 3B) Because Psm M6 (with a pFKN replicon) andthe plasmid-cured derivative of Psm M6integron bothgrew equivalently we can conclude that it is the inte-gration of pFKN per se that results in this growthimpairment This is not uncommon as several plasmidreplicons are known to slow growth of the organismwhen integrated (see Discussion)
The pFKN integration site in Psm M6integron is the avrPphE transposon
We noticed from the DNA blot that the genomic environ-ment of avrRpm1 is different in Psm M6 and PsmM6integron This is also the case for both copies of avrP-phE (data not shown) suggesting that the pFKN integra-tion site is near avrRpm1 and avrPphE and that the
Fig 2 avrRpm1 encodes the determining virulence factor on pFKNA The growth of Psm M6C on rpm1-3 plants was compared with the growth of the wild-type Psm M6 strain and Psm M6C strains comple-mented with different fragments from pFKN The pFKN subclone carrying avrRpm1 restored full virulence to Psm M6C (the other subclones did not restore any virulence and are not shown) Leaves were hand inoculated with 105 cfu ml-1 and a titration of bacteria in the leaves was performed 3 days after inoculation Data points rep-resent the mean plusmn standard error of four samplesB The avrPphE gene was deleted from the chromosome of Psm M6C to generate Psm M6CKO To assess the virulence function of avrPphE or its influence on the virulence function of avrRpm1 the growth of Psm M6CKO on rpm1-3 plants was compared with the growth of Psm M6C Psm M6C + avrRpm1 and Psm M6CKO + avrRpm1 Data points represent the mean plusmn standard error of four samples
1552 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
chromosomal location into which the plasmid integrates isnear the chromosomal copy of avrPphE We first definedthat the chromosomal avrPphE is carried on the sametransposon found in the sequence of pFKN using PCRamplification on total DNA of Psm M6C with primersspecific to sequence on the avrPphE transposon(Table 5) The PCR products were sequenced and com-pared with the corresponding sequenced regions ofpFKN As shown in Fig 4A the chromosomal copy of thetransposon contains two sequences not present on pFKNOne 1244 bp sequence (region A) is located betweenavrPphE and the coding region of TnpA It carries an ORFwith a sequence that is nearly identical to various genesencoding resolvases of Tn3 transposons Additionally aportion of 661 bp of this region encodes the 5cent portion ofTnpA Another region of 271 bp unique to the chromo-somal copy is located within the coding region of thetransposase (region C) and is 3cent from a region shared byboth pFKN and chromosomal copies of TnpA (region B)The addition of regions A and C to the chromosomal copyresults in a complete TnpA ORF correcting the frameshiftat the 3cent end of TnpA carried on pFKN
TnpA avrPphE and the putative resolvase fragmentare located between nearly identical inverted repeats of66 bp on both the plasmid and the chromosome Theirorganization is reminiscent of Tn3-like transposons(Sherrat 1989) Moreover this TnpA is 81 identical tothose of the Tn21 subfamily of Tn3 elements (Diveret al 1983) As we suspected that the integration site isin the vicinity of avrPphE in the chromosome and theplasmid (ie in the two transposons) we analysed byPCR the two transposons in the genome of PsmM6integron Figure 4B shows that the unique regions Aand C are no longer located on the same transposonbut are instead separated by the entire plasmidsequence (Fig 4C) This indicated that the integrationsite of the plasmid is located in the region betweenregion A and region C ie within the homology providedby region B (Fig 4)
We used this sequence data to investigate the rateof excision occurring in Psm M6integron grown indifferent conditions by PCR The rate of excision wasestimated at around 10-4 (data not shown) No differencein rate of excision could be detected between bacteria
Fig 3 pFKN can integrate into the Psm M6 chromosomeA Total DNA from Psm M6 (lanes F) and Psm M6integron (lanes I) was digested with EcoRI HindIII EcoRV SacI PvuII SmaI BglII and NsiI and probed with various pFKN fragments Here the DNA blot was probed with the avrRpm1 ORF The two DNA samples exhibit a similar pattern except for a polymorphism (eg EcoRI BglII and SmaI) near the site of pFKN integra-tion into the chromosomeB The integration of the plasmid alters the overall fitness of Psm M6integron The growth of Psm M6integron in KB (empty symbols) and MM (filled symbols) was compared with the growth of Psm M6 and Psm M6C Bacteria were inoculated at 104 cfu ml-1 and cultures were titrated at 0 6 12 29 32 54 and 60 h after the inoculation Psm M6pFKN is indicated by squares Psm M6C is indicated by triangles and rhombi indicate Psm M6integron Data points represent the mean plusmn standard error of four samples (some error bars are not visible because too small) Psm M6pFKN and Psm M6C grew equally well in both minimal medium and KB Psm M6integron grows slower than Psm M6pFKN and Psm M6C in both media
pFKN a virulence plasmid in P syringae pv maculicola 1553
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
grown in KB MM and on resistant or susceptibleplants
The genomic context of avrPphE is different in various P syringae strains
We investigated the regions surrounding the avrPphEtransposon on the chromosome of Psm M6 (Fig 5) byTAIL-PCR The left border shows 97 nt identity to the 5centend of the insertion sequence ISPsy3 (AB063176) fromP syringae pv eriobotryae This element is interrupted bythe avrPphE transposon The translation product of theinterrupted ORF found on this border is homologous tothe IS801 transposase (79 identity and 86 homology)
and IS801 has been associated with other type III effec-tors in various P syringae pathovars (Kim et al 1998)On the right border the 200 bp sequence following theterminal repeat of the avrPphE transposon encodes aprotein homologous (81 nucleotide identity) to the 3centend of an unrelated putative transposase gene fromPseudomonas aeruginosa The next 1062 bp sequencedownstream shows high nucleotide homology tosequence identified in P syringae pv pisi (Arnold et al2001) This DNA fragment may be a transposon remnantand carries the avrPpiA1 allele of avrRpm1 from P syrin-gae pv pisi among other genes In the chromosome ofPsm M6 this sequence shows homology to the P s pisigenes ORFD (94 identity) encoding a putative trans-
Fig 4 The pFKN integration site is a second copy of the avrPphE transposonA The region in the Psm M6 chromosome that hybridizes to the avrPphE and TnpA probes was amplified and sequenced The sequence of the chromosomal region is 100 identical to the transposon in the plasmid but contains two additional regions (regions A and C) TnpA encodes a transposase and res a resolvase Based on homology the chromosomal avrPphE transposon could be complete and functional The avrPphE transposon carried on the plas-mid lacks the 5cent end of TnpA and res (region A) and the absence of region C results in the truncation and a frameshift (marked FS) of the gene encoding the transposase The primers used for PCR amplification (Fig 4B) are indi-cated by arrows and their names are in boldB To investigate where regions A and C were distributed after integration of the plasmid into the chromosome PCRs were performed on the DNA of Psm M6integron and Psm M6 using Larp3 and K110 and K110 and K113 (see Fig 4A) The product amplified from Psm M6integron is run in lane I The product ampli-fied from Psm M6 is run in lane F The product amplified from Psm M6C is run in lane C The difference in size observed between the PCR products from Psm M6integron and Psm M6 using Larp3 and K110 indicates that regions A and C are not carried on the same putative transposon in the chromosome of Psm M6integron (in contrast to the chromosome of Psm M6) The PCR products from Psm M6integron and Psm M6 using K113 and K110 show that neither region A nor region C has been lost in the chromosome of Psm M6integron The identical size of the PCR prod-ucts from Psm M6integron and Psm M6 using K113 and Larp2 indicates that region C is on the right border of pFKN when integrated into the chromosome The region A is on the left border as it is carried by the other putative transposonC After integration of pFKN into the chromo-some TnpA on the left border contains region A but not region C TnpA on the right border contains region C The primers used for the PCR amplification are indicated by arrows and their names are in bold (in Fig 4A)
1554 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
posase and ORF3 encoding a putative type III effector(100 identity) Thus this region in Psm M6 is a remnantof the putative transposon found in P s pisi
We detected the avrPphE transposon in other strains ofP syringae pv maculicola and in the sequence of the
genome of P s tomato DC3000 In Psm M2 Psm M8Psm M12 Psm m90 and Psm m10831 the borders arethe same as in Psm M6 (data not shown) On the otherhand they differ in Psm M5 and Psm M9 and these alldiffer from Pst DC3000 In Psm M5 (Fig 5) and Psm M9
Fig 5 Sequences surrounding the avrPphE transposon define four different genetic contexts The figure is a graphical representation of the four genomic environments in which the avrPphE transposon was detected The regions represented were characterized by sequence comparison using the BLASTX algorithm It is not known whether the avrPphE transposon is located in the chromosome or on a plasmid in the genome of Psm M5 and Pst DC3000
pFKN a virulence plasmid in P syringae pv maculicola 1555
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(not shown) the left border consists of the 5cent end of agene sharing 99 identity to the replicase of pPS10 aplasmid found in P syringae pv savastanoi (Nieto et al1992) This replicase is not related to that of pFKN (aPT23A-like plasmid see above) The right bordersequence is a remnant of a gene encoding a putative ABCtransporter (23 identity and 43 homology to a putativesugar uptake ABC transporter ATP-binding protein fromSinorhizobium meliloti)
In the genome of Pst DC3000 we looked at 30 kbsurrounding the transposon (Fig 5) The proteinsencoded on this 30 kb region share homology to eitherDNA mobility functions (transposases plasmid partition-ing proteins replicases) or type III effectors (identical toavrPpiB Cournoyer et al 1995) The region also containsother sequences often found associated with plasmidsand genes involved in pathogenicity such as rulA and rulB(Sesma et al 2000) Sequences directly neighbouringthe transposon on the left side are homologous to a geneencoding the transposase of the IS100 elements fromYersinia pestis Ralstonia solanacearum and Salmonellaenterica On the right-hand side of the avrPphE transpo-son the first homology is to a fragment of a putative ABCtransporter identical to the one identified in Psm M5 Thissuggests that this part of the two genomes is morerecently diverged than either is from Psm M6
No common sequence was identified in the four differ-ent environments surrounding the avrPphE transposonthat would help to identify the integration target of theavrPphE transposon although the proximal right bordersin Psm M5 and Pst DC3000 are nearly identical Thissuggests that the avrPphE transposon and this border-ing sequence were acquired together by these twostrains and that subsequent events at both the distalright flank and on the left border are responsible for thedifferences in genomic context In contrast the insertionsites of the avrPphE transposon in Psm M5 and PsmM6 are completely different In sum the features ofthese integration sites are consistent with the model thatPAIs evolve by repeated insertion and deletion intopre-existing PAIs (Hensel et al 1999 Tauschek et al2002)
Discussion
We sequenced pFKN an ordf40 kb plasmid that contributesto the virulence of Psm M6 The plasmid carries two typeIII effectors on remnants of transposons contiguous toORF8 itself neighbouring an element closely related toIS801 avrRpm1 encodes a known virulence factor andavrPphE a type III effector The expression of ORF8 isinduced in planta and therefore ORF8 may play a role inthe interaction with the host plant The portion of theplasmid carrying avrRpm1 avrPphE and ORF8 may thus
be considered as a PAI Interestingly the G+C content ofthe ORFs carried on this region are very heterogeneousand the three genes are flanked by sequences related tomobile elements and therefore are likely to have differentorigins This is consistent with the idea that the formationof PAIs requires a multistep process (Hensel et al 1999Alfano et al 2000 Tauschek et al 2002) It is also con-sistent with the findings of Kim et al (1998) showing thatmost of the known type III effectors are associated withthis type of sequence pFKN (and therefore the PAI that itcarries) sometimes integrates into and can excise fromthe chromosome as demonstrated by the discovery of thevariant of the strain Psm M6 We identified the region ofthe chromosome into which the integration took placeThis region is a transposon carrying a second copy ofavrPphE
Alleles of the avrPphE gene have been detected inmany P syringae pathovars (Mansfield et al 1994Alfano et al 2000 Fouts et al 2002 Guttman et al2002) suggesting that it provides an important virulencefunction on a variety of host plants Our findings are thefirst to associate avrPphE with a transposon In PsyB728a and in Pph 1302A for example avrPphE residesdirectly next to the hrp locus in a region called thelsquoexchangeable effector locusrsquo or EEL (Alfano et al 2000)and is not associated with a transposase In fact in PsyB728a avrPphE is predicted to be the third gene in anoperon controlled by the hrpK promoter (Alfano et al2000) This striking finding illustrates the dynamism ofPAIs
We determined by complementation assay that the par-tial loss of virulence in the strain cured of pFKN resultsfrom the absence of avrRpm1 The virulence function ofavrRpm1 has already been demonstrated in the strainPsm M2 (Ritter and Dangl 1995) Recently AvrRpm1 hasbeen demonstrated to interact with RIN4 a proteininvolved in the regulation of some of the plant defenceresponses (Mackey et al 2002) Mackey and coworkersproposed that the role of AvrRpm1 is to reduce the levelof plant defence response triggered by the attack of thepathogen This hypothesis might be consistent with thefact that only quantitative differences in growth wereobserved between Psm M6C and Psm M6 No differencein growth was observed between Psm M6C and PsmM6CKO (lacking avrPphE) Similar results were observedwith the avrPphE allele of P s phaseolicola (Stevenset al 1998) This observation could be explained byeither a functional redundancy with another gene in thegenome of the Psm M6 or the fact that the assay isinappropriate to assess the virulence function of this typeIII effector
The sequence and the integration mechanism of pFKNinto the Psm M6 chromosome provide insights on theevolutionary dynamics of the genomes of P syringae
1556 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
pathovars First the analysis of the G+C content of frag-ments of pFKN suggests that they may have differentorigins This hypothesis is further supported by the pres-ence of some defined fragments from pFKN in thegenomes of Pst DC3000 and Psy B728a The fragment 3(see Table 2) encodes one putative transcriptional regu-lator proteins predicted to be located in the bacterialinner or outer membrane by PSORT (inorganic pyrophos-phatase sugar transporter putative protein) and a puta-tive integraserecombinase homologous to ripX fromBacillus subtilis The presence of this latter gene sug-gests that fragment 3 may be the remnant of abacteriophage
The integration and excision mechanism is very proba-bly a homologous recombination in the 15 kb sequenceregion B of TnpA carried on the avrPphE transposon(Fig 6) It is not clear whether the integrationexcisionprocess relies on general recombination or on a site-specific recombination dependent on an integraserecombinase As indicated by the sequence of the twotransposons before and after integration of pFKN nonucleotide sequences were altered at the site ofexchange and not a single nucleotide was lost or gained(this excludes a mechanism of transpositional site-specificrecombination) General recombination requires a largeDNA region whereas the site-specific recombinationrequires as few as 240 bp for the lambda phage for exam-ple (Thompson and Landy 1989) The rate of generalrecombination is unknown for P syringae but has beenestimated at 10-3 in Pseudomonas tolaasii (and whenrecA is inactivated at 10-4 Sinha et al 2000) The rateof excision of pFKN has been estimated between 10-3 and10-4 and would therefore be consistent with a mechanismof general recombination Additionally the region B is
15 kb long enough to ensure general recombinationOlder reports describe the integration of pMMC7501 intothe chromosome of P s pv phaseolicola (Curiale andMills 1982) An imprecise excision of pMMC7501 led tothe formation of eight derivative plasmids all sharing thesame origin of replication (Szabo and Mills 1984a) Thisexcision occurred through recombination events betweensets of repeats in the chromosome exhibiting sequencehomology with IS elements (Szabo and Mills 1984bPoplawsky and Mills 1987)
The integration of a virulence gene into the chromo-some may be a first step towards its stabilization in thegenome Indeed lack of selection can lead to the stochas-tic loss of the plasmid and many steps in the pathogenlife cycle probably do not require the presence of virulencefunctions The loss of a plasmid carrying the type III effec-tor avrPpiB was observed in P syringae pv pisi (Bavageet al 1991) We propose that the integration of pFKNcould lead to the stabilization of avrRpm1 in the genomeof Psm M6 Interestingly a DNA segment of at least 42 kbcarrying avrPphB in P syringae pv phaseolicola canexcise from the chromosome but it is not maintained inthe strain as a plasmid This region may represent anancient plasmid integration event followed by successivegenomic rearrangements deleterious for plasmid mainte-nance (Jackson et al 2000) The fact that we can stillobserve the excision of pFKN from the chromosome ofPsm M6integron shows that the newly generated geneticunit is not rapidly stabilized in the genome and that furtherevents might be necessary to prevent excision Further-more the integration of pFKN into the chromosomeslightly impairs the fitness of Psm M6integron The pres-ence of the functional origin of replication of pFKN mayinterfere with the replication of the chromosome Integra-
Fig 6 The integration of pFKN may have occurred via homologous recombination between the regions B common to the two TnpA genes The TnpA genes are constituted of four DNA sequence regions (1) before the integration the chromosomal copy of TnpA contains all four regions (A B C and D) The TnpA on the plasmid is missing the 5cent end of the gene (region A) and a fragment towards the 3cent end (region C) which results in a frameshift (FS) After the homologous recombination between two identical regions (region B) in the coding region of the TnpA genes (2) the region A is found in the TnpA of the left border and region C in the TnpA of the right border (3)
pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

pFKN a virulence plasmid in P syringae pv maculicola 1549
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
a single gene encoding an outer membrane haeminreceptor ChuA in pathogenic E coli as well as othergenes involved in pathogenicity of the strain (Nagy et al2001 2002) The promoter of the genes or operons con-trolled by RfaH contains a conserved sequence calledJUMPStart (Hobbs and Reeves 1994) No JUMPStartsequence was detected on pFKN
Transporters ORF18 encodes a probable transmem-brane protein that is highly homologous to sugar trans-porters One is the product of mae1 a gene of the yeastSchizosaccharomyces pombe encoding a proteininvolved in the uptake of L-malate succinate and malonicacid The product of ORF18 may provide the bacteria witha fitness advantage in certain conditions
Other putative functions encoded on the plasmid Othergenes may encode proteins providing an advantage to thebacteria during the infection process For example theproduct of ORF12 displays all the characteristics ofmethyl-accepting chemotaxis proteins The genes encod-ing the flagellar unit and the chemotaxis functions areoften organized in clusters which is not the case onpFKN In this particular case the flagellar unit may beprovided in trans
pFKN is composed of different fragments that indicate a multistep process leading to its formation
The G+C content of the pFKN ORFs is highly variablealong the plasmid (Table 2) Yet several contiguous genesets on the plasmid have similar G+C contents This prob-ably indicates that these genes were acquired togetherfrom an original source The G+C content might allow usto track the multistep acquisition process resulting inpFKN For example ORFs 7 8 and 10 share nearly iden-tical G+C content (505) but are separated by an ISelement with a significantly different G+C content (603)To determine the distribution of pFKN ORFs in otherpathovars and how this distribution is related to G+Ccontent by ORF group we performed sequence compar-isons with the nucleotide sequence of the completegenomes of P syringae pv tomato DC3000 (httptigr-blasttigrorgufmg) and P syringae pv syringae B728a(httpwwwjgidoegovJGI_microbialhtmlindexhtml)DNA fragments carrying from two to seven contiguouspFKN ORFs were detected in both genome sequences(85ndash100 identity) The four DNA fragments from P stomato DC3000 carrying linked pFKN ORFs were num-bered and added to Table 2 Of these Pst DC3000 frag-ment 2 was also detected in Psy B728a (98 identical topFKN) In addition sequences homologous to repA andORF13 were also detected in this Psy B728a Table 2shows that ORFs belonging to the same contiguous DNA
fragment have a similar G+C content For example genesbelonging to fragment 4 have a relatively high G+C con-tent Genes belonging to fragments 1 and 2 have similarG+C contents and genes in fragment 3 exhibit a lowerG+C content Thus the different fragments constitutingpFKN were probably acquired by the three genomes indifferent steps followed by subsequent sequence dele-tions andor rearrangements with respect to Psm M6 asthe entire collinear pFKN is not detected in either PstDC3000 or Psy B728A
Full virulence of PsmM6 requires pFKN and is essentially caused by avrRpm1
To determine the contribution to virulence of the plasmidpFKN we cured Psm M6 of pFKN (see Experimentalprocedures) to generate strain Psm M6C There was noobvious macroscopic difference in visible disease symp-toms after inoculation of either wild-type or Psm M6Conto rpm1-3 Arabidopsis leaves (unable to recognizeavrRpm1) At high levels of inoculation (5 yen 106 cfu ml-1)the inoculated leaves exhibited water soaking and tissuecollapse starting at 24 h after inoculation Inoculation at5 yen 105 cfu ml-1 led to chlorosis between 48 and 72 h afterinoculation Using this low-density inoculum we mea-sured pathogen growth over time in rpm1-3 leaves PsmM6C grew 10-fold less than the wild-type strain (Fig 2A)Therefore the cured strain is less virulent than the wild-type strain We performed complementation experimentsusing various pFKN subclones in Psm M6C to determinewhich pFKN ORFs would partially or fully restore viru-lence Only clones carrying avrRpm1 could restore fullvirulence (Fig 2A) This result is consistent with inser-tional mutation of avrRpm1 in the Psm M2 strain (Ritterand Dangl 1995) No other clone conferred enhancedvirulence (data not shown)
The role of avrPphE in virulence could not be assessedusing Psm M6C because a second copy of the gene ispresent on the chromosome of the cured strain [asassessed by polymerase chain reaction (PCR) amplifica-tion and sequencing of the PCR product see below] Wereplaced the avrPphE copy on the chromosome of PsmM6C with an omega fragment to create Psm M6CKO (seeExperimental procedures) Psm M6CKO grew similarly toPsm M6C on rpm1-3 plants (Fig 2B) suggesting thatavrPphE plays no major role in the virulence of Psm M6However our recent results indicate that expression ofavrPphE from the lac-nptII promoter in PsmM6CKO doesenhance virulence of PsmM6CKO suggesting that thequantity of AvrPphE protein delivered to the host cell maybe important in its function (J Shock J Chang Z Nim-chuk L Rohmer and J L Dangl unpublished) As avrP-phE was located in the vicinity of avrRpm1 on pFKN wetested the potential involvement of avrPphE in the viru-
1550 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
lence function of avrRpm1 We compared the growth ofPsm M6C and Psm M6CKO when each carried avrRpm1on a plasmid No difference in growth was observed(Fig 2B) indicating that avrPphE does not affectavrRpm1 virulence function
Identification of additional genes putatively involved in pathogenicity based on their expression pattern
The expression of known P syringae virulence factors isspecifically induced in planta Minimal media (MM) plus asugar source seems to mimic the plant apoplast leadingto the expression of the type III pilus genes and effectors
(Huynh et al 1989 Innes et al 1993) We compared theexpression of the pFKN ORFs in MM and in rich medium(KB) using reverse transcription RT-PCR methodsWhen the expression of an ORF was observed in bothmedia we assumed that the gene was constitutivelyexpressed Expression of a gene in MM but not in KB mayindicate a role for this gene during interaction with thehost Most of the pFKN genes are constitutivelyexpressed However the expression of both avrRpm1(Ritter and Dangl 1995) and avrPphE was detected inMM but not in KB Additionally ORF26 is expressed inMM but not in KB ORFs 14 and 24 were not expressedin either of the two conditions The expression of ORF8
Table 2 pFKN ORFs sorted by descending G+C content
a Some pFKN ORFs are contiguous on the plasmid and are collinear in this tableindicating that they possess a similar G+C content When this is the case an emptyrow separates them from the next ORF in the tableb The DNA fragments from the Pst DC3000 genome that contain sequenceshomologous to ORFs from pFKN were colour-coded (1 green 2 red 3 blue4 pink) The table indicates that fragment 4 has a relatively high G+C contentwhereas fragment 3 has a relatively low content
pFKN a virulence plasmid in P syringae pv maculicola 1551
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
and ORF11 could not be assessed using this techniquedespite many attempts
The six genes (avrRpm1 avrPphE ORF8 ORF11ORF14 and ORF24) for which no constitutive expressionwas observed by RT-PCR were selected for further studyWe constructed transcriptional fusions between the 5cent endof the genes of interest and a gene encoding green fluo-rescent protein (GFP) and chose ORF10 as a constitutivecontrol The fluorescence of PsmM6C carrying the tran-scriptional fusions was assessed in KB MM and in plantaby exposing the bacteria grown in these conditions to bluelight (488 nm) and observing them by fluorescencemicroscopy A strain carrying promoterless GFP did notexhibit any fluorescence in the conditions tested whereas
all cells carrying the ORF10ndashGFP fusion did Only theavrPphE avrRpm1 and ORF8 GFP fusions were active inplants (Table 1)
pFKN can integrate into the Psm M6 chromosome
One colony of Psm M6 derived during routine mainte-nance exhibited unusual features Although this straintriggered recognition (measured by HR) on RPM1 plantsand disease symptoms on rpm1 plants no plasmid couldbe isolated from it AvrRpm1 was still present in totalDNA as assayed by DNA blot (Fig 3A) We thereforeconcluded that avrRpm1 was now located on the PsmM6 chromosome To determine whether additional por-tions of pFKN were also present on the variance chromo-some total DNA from the variant Psm M6 was digestedwith EcoRI HindIII EcoRV SacI PvuII SmaI BglII andNsiI and probed with the DNA from different parts of thepFKN plasmid The DNA of Psm M6 containing pFKNwas used as a positive control This analysis revealed apolymorphism between the avrRpm1 gene in this strain(eg an 80 kb BglII fragment) compared with pFKN (egan 88 kb BglII fragment Fig 3A) All restriction frag-ments from pFKN were detected in the DNA of PsmM6integron Therefore pFKN has integrated into thechromosome For the purpose of clarity the strain bear-ing avrRpm1 in the chromosome will be referred to asPsm M6integron
We have on occasion isolated a replicon from indi-vidual colonies of Psm M6integron (see next section)The plasmid purified from these bacteria always exhib-ited a restriction profile identical to that of pFKNTherefore pFKN can excise from the chromosome andPsm M6integron can revert to Psm M6 Interestinglythe overall fitness of Psm M6 is affected by the integra-tion status of pFKN into the chromosome PsmM6integron grows in media (KB and MM) more slowlythan either the original Psm M6 strain or a revertantfrom Psm M6integron that is cured of the plasmid(Fig 3B) Because Psm M6 (with a pFKN replicon) andthe plasmid-cured derivative of Psm M6integron bothgrew equivalently we can conclude that it is the inte-gration of pFKN per se that results in this growthimpairment This is not uncommon as several plasmidreplicons are known to slow growth of the organismwhen integrated (see Discussion)
The pFKN integration site in Psm M6integron is the avrPphE transposon
We noticed from the DNA blot that the genomic environ-ment of avrRpm1 is different in Psm M6 and PsmM6integron This is also the case for both copies of avrP-phE (data not shown) suggesting that the pFKN integra-tion site is near avrRpm1 and avrPphE and that the
Fig 2 avrRpm1 encodes the determining virulence factor on pFKNA The growth of Psm M6C on rpm1-3 plants was compared with the growth of the wild-type Psm M6 strain and Psm M6C strains comple-mented with different fragments from pFKN The pFKN subclone carrying avrRpm1 restored full virulence to Psm M6C (the other subclones did not restore any virulence and are not shown) Leaves were hand inoculated with 105 cfu ml-1 and a titration of bacteria in the leaves was performed 3 days after inoculation Data points rep-resent the mean plusmn standard error of four samplesB The avrPphE gene was deleted from the chromosome of Psm M6C to generate Psm M6CKO To assess the virulence function of avrPphE or its influence on the virulence function of avrRpm1 the growth of Psm M6CKO on rpm1-3 plants was compared with the growth of Psm M6C Psm M6C + avrRpm1 and Psm M6CKO + avrRpm1 Data points represent the mean plusmn standard error of four samples
1552 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
chromosomal location into which the plasmid integrates isnear the chromosomal copy of avrPphE We first definedthat the chromosomal avrPphE is carried on the sametransposon found in the sequence of pFKN using PCRamplification on total DNA of Psm M6C with primersspecific to sequence on the avrPphE transposon(Table 5) The PCR products were sequenced and com-pared with the corresponding sequenced regions ofpFKN As shown in Fig 4A the chromosomal copy of thetransposon contains two sequences not present on pFKNOne 1244 bp sequence (region A) is located betweenavrPphE and the coding region of TnpA It carries an ORFwith a sequence that is nearly identical to various genesencoding resolvases of Tn3 transposons Additionally aportion of 661 bp of this region encodes the 5cent portion ofTnpA Another region of 271 bp unique to the chromo-somal copy is located within the coding region of thetransposase (region C) and is 3cent from a region shared byboth pFKN and chromosomal copies of TnpA (region B)The addition of regions A and C to the chromosomal copyresults in a complete TnpA ORF correcting the frameshiftat the 3cent end of TnpA carried on pFKN
TnpA avrPphE and the putative resolvase fragmentare located between nearly identical inverted repeats of66 bp on both the plasmid and the chromosome Theirorganization is reminiscent of Tn3-like transposons(Sherrat 1989) Moreover this TnpA is 81 identical tothose of the Tn21 subfamily of Tn3 elements (Diveret al 1983) As we suspected that the integration site isin the vicinity of avrPphE in the chromosome and theplasmid (ie in the two transposons) we analysed byPCR the two transposons in the genome of PsmM6integron Figure 4B shows that the unique regions Aand C are no longer located on the same transposonbut are instead separated by the entire plasmidsequence (Fig 4C) This indicated that the integrationsite of the plasmid is located in the region betweenregion A and region C ie within the homology providedby region B (Fig 4)
We used this sequence data to investigate the rateof excision occurring in Psm M6integron grown indifferent conditions by PCR The rate of excision wasestimated at around 10-4 (data not shown) No differencein rate of excision could be detected between bacteria
Fig 3 pFKN can integrate into the Psm M6 chromosomeA Total DNA from Psm M6 (lanes F) and Psm M6integron (lanes I) was digested with EcoRI HindIII EcoRV SacI PvuII SmaI BglII and NsiI and probed with various pFKN fragments Here the DNA blot was probed with the avrRpm1 ORF The two DNA samples exhibit a similar pattern except for a polymorphism (eg EcoRI BglII and SmaI) near the site of pFKN integra-tion into the chromosomeB The integration of the plasmid alters the overall fitness of Psm M6integron The growth of Psm M6integron in KB (empty symbols) and MM (filled symbols) was compared with the growth of Psm M6 and Psm M6C Bacteria were inoculated at 104 cfu ml-1 and cultures were titrated at 0 6 12 29 32 54 and 60 h after the inoculation Psm M6pFKN is indicated by squares Psm M6C is indicated by triangles and rhombi indicate Psm M6integron Data points represent the mean plusmn standard error of four samples (some error bars are not visible because too small) Psm M6pFKN and Psm M6C grew equally well in both minimal medium and KB Psm M6integron grows slower than Psm M6pFKN and Psm M6C in both media
pFKN a virulence plasmid in P syringae pv maculicola 1553
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
grown in KB MM and on resistant or susceptibleplants
The genomic context of avrPphE is different in various P syringae strains
We investigated the regions surrounding the avrPphEtransposon on the chromosome of Psm M6 (Fig 5) byTAIL-PCR The left border shows 97 nt identity to the 5centend of the insertion sequence ISPsy3 (AB063176) fromP syringae pv eriobotryae This element is interrupted bythe avrPphE transposon The translation product of theinterrupted ORF found on this border is homologous tothe IS801 transposase (79 identity and 86 homology)
and IS801 has been associated with other type III effec-tors in various P syringae pathovars (Kim et al 1998)On the right border the 200 bp sequence following theterminal repeat of the avrPphE transposon encodes aprotein homologous (81 nucleotide identity) to the 3centend of an unrelated putative transposase gene fromPseudomonas aeruginosa The next 1062 bp sequencedownstream shows high nucleotide homology tosequence identified in P syringae pv pisi (Arnold et al2001) This DNA fragment may be a transposon remnantand carries the avrPpiA1 allele of avrRpm1 from P syrin-gae pv pisi among other genes In the chromosome ofPsm M6 this sequence shows homology to the P s pisigenes ORFD (94 identity) encoding a putative trans-
Fig 4 The pFKN integration site is a second copy of the avrPphE transposonA The region in the Psm M6 chromosome that hybridizes to the avrPphE and TnpA probes was amplified and sequenced The sequence of the chromosomal region is 100 identical to the transposon in the plasmid but contains two additional regions (regions A and C) TnpA encodes a transposase and res a resolvase Based on homology the chromosomal avrPphE transposon could be complete and functional The avrPphE transposon carried on the plas-mid lacks the 5cent end of TnpA and res (region A) and the absence of region C results in the truncation and a frameshift (marked FS) of the gene encoding the transposase The primers used for PCR amplification (Fig 4B) are indi-cated by arrows and their names are in boldB To investigate where regions A and C were distributed after integration of the plasmid into the chromosome PCRs were performed on the DNA of Psm M6integron and Psm M6 using Larp3 and K110 and K110 and K113 (see Fig 4A) The product amplified from Psm M6integron is run in lane I The product ampli-fied from Psm M6 is run in lane F The product amplified from Psm M6C is run in lane C The difference in size observed between the PCR products from Psm M6integron and Psm M6 using Larp3 and K110 indicates that regions A and C are not carried on the same putative transposon in the chromosome of Psm M6integron (in contrast to the chromosome of Psm M6) The PCR products from Psm M6integron and Psm M6 using K113 and K110 show that neither region A nor region C has been lost in the chromosome of Psm M6integron The identical size of the PCR prod-ucts from Psm M6integron and Psm M6 using K113 and Larp2 indicates that region C is on the right border of pFKN when integrated into the chromosome The region A is on the left border as it is carried by the other putative transposonC After integration of pFKN into the chromo-some TnpA on the left border contains region A but not region C TnpA on the right border contains region C The primers used for the PCR amplification are indicated by arrows and their names are in bold (in Fig 4A)
1554 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
posase and ORF3 encoding a putative type III effector(100 identity) Thus this region in Psm M6 is a remnantof the putative transposon found in P s pisi
We detected the avrPphE transposon in other strains ofP syringae pv maculicola and in the sequence of the
genome of P s tomato DC3000 In Psm M2 Psm M8Psm M12 Psm m90 and Psm m10831 the borders arethe same as in Psm M6 (data not shown) On the otherhand they differ in Psm M5 and Psm M9 and these alldiffer from Pst DC3000 In Psm M5 (Fig 5) and Psm M9
Fig 5 Sequences surrounding the avrPphE transposon define four different genetic contexts The figure is a graphical representation of the four genomic environments in which the avrPphE transposon was detected The regions represented were characterized by sequence comparison using the BLASTX algorithm It is not known whether the avrPphE transposon is located in the chromosome or on a plasmid in the genome of Psm M5 and Pst DC3000
pFKN a virulence plasmid in P syringae pv maculicola 1555
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(not shown) the left border consists of the 5cent end of agene sharing 99 identity to the replicase of pPS10 aplasmid found in P syringae pv savastanoi (Nieto et al1992) This replicase is not related to that of pFKN (aPT23A-like plasmid see above) The right bordersequence is a remnant of a gene encoding a putative ABCtransporter (23 identity and 43 homology to a putativesugar uptake ABC transporter ATP-binding protein fromSinorhizobium meliloti)
In the genome of Pst DC3000 we looked at 30 kbsurrounding the transposon (Fig 5) The proteinsencoded on this 30 kb region share homology to eitherDNA mobility functions (transposases plasmid partition-ing proteins replicases) or type III effectors (identical toavrPpiB Cournoyer et al 1995) The region also containsother sequences often found associated with plasmidsand genes involved in pathogenicity such as rulA and rulB(Sesma et al 2000) Sequences directly neighbouringthe transposon on the left side are homologous to a geneencoding the transposase of the IS100 elements fromYersinia pestis Ralstonia solanacearum and Salmonellaenterica On the right-hand side of the avrPphE transpo-son the first homology is to a fragment of a putative ABCtransporter identical to the one identified in Psm M5 Thissuggests that this part of the two genomes is morerecently diverged than either is from Psm M6
No common sequence was identified in the four differ-ent environments surrounding the avrPphE transposonthat would help to identify the integration target of theavrPphE transposon although the proximal right bordersin Psm M5 and Pst DC3000 are nearly identical Thissuggests that the avrPphE transposon and this border-ing sequence were acquired together by these twostrains and that subsequent events at both the distalright flank and on the left border are responsible for thedifferences in genomic context In contrast the insertionsites of the avrPphE transposon in Psm M5 and PsmM6 are completely different In sum the features ofthese integration sites are consistent with the model thatPAIs evolve by repeated insertion and deletion intopre-existing PAIs (Hensel et al 1999 Tauschek et al2002)
Discussion
We sequenced pFKN an ordf40 kb plasmid that contributesto the virulence of Psm M6 The plasmid carries two typeIII effectors on remnants of transposons contiguous toORF8 itself neighbouring an element closely related toIS801 avrRpm1 encodes a known virulence factor andavrPphE a type III effector The expression of ORF8 isinduced in planta and therefore ORF8 may play a role inthe interaction with the host plant The portion of theplasmid carrying avrRpm1 avrPphE and ORF8 may thus
be considered as a PAI Interestingly the G+C content ofthe ORFs carried on this region are very heterogeneousand the three genes are flanked by sequences related tomobile elements and therefore are likely to have differentorigins This is consistent with the idea that the formationof PAIs requires a multistep process (Hensel et al 1999Alfano et al 2000 Tauschek et al 2002) It is also con-sistent with the findings of Kim et al (1998) showing thatmost of the known type III effectors are associated withthis type of sequence pFKN (and therefore the PAI that itcarries) sometimes integrates into and can excise fromthe chromosome as demonstrated by the discovery of thevariant of the strain Psm M6 We identified the region ofthe chromosome into which the integration took placeThis region is a transposon carrying a second copy ofavrPphE
Alleles of the avrPphE gene have been detected inmany P syringae pathovars (Mansfield et al 1994Alfano et al 2000 Fouts et al 2002 Guttman et al2002) suggesting that it provides an important virulencefunction on a variety of host plants Our findings are thefirst to associate avrPphE with a transposon In PsyB728a and in Pph 1302A for example avrPphE residesdirectly next to the hrp locus in a region called thelsquoexchangeable effector locusrsquo or EEL (Alfano et al 2000)and is not associated with a transposase In fact in PsyB728a avrPphE is predicted to be the third gene in anoperon controlled by the hrpK promoter (Alfano et al2000) This striking finding illustrates the dynamism ofPAIs
We determined by complementation assay that the par-tial loss of virulence in the strain cured of pFKN resultsfrom the absence of avrRpm1 The virulence function ofavrRpm1 has already been demonstrated in the strainPsm M2 (Ritter and Dangl 1995) Recently AvrRpm1 hasbeen demonstrated to interact with RIN4 a proteininvolved in the regulation of some of the plant defenceresponses (Mackey et al 2002) Mackey and coworkersproposed that the role of AvrRpm1 is to reduce the levelof plant defence response triggered by the attack of thepathogen This hypothesis might be consistent with thefact that only quantitative differences in growth wereobserved between Psm M6C and Psm M6 No differencein growth was observed between Psm M6C and PsmM6CKO (lacking avrPphE) Similar results were observedwith the avrPphE allele of P s phaseolicola (Stevenset al 1998) This observation could be explained byeither a functional redundancy with another gene in thegenome of the Psm M6 or the fact that the assay isinappropriate to assess the virulence function of this typeIII effector
The sequence and the integration mechanism of pFKNinto the Psm M6 chromosome provide insights on theevolutionary dynamics of the genomes of P syringae
1556 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
pathovars First the analysis of the G+C content of frag-ments of pFKN suggests that they may have differentorigins This hypothesis is further supported by the pres-ence of some defined fragments from pFKN in thegenomes of Pst DC3000 and Psy B728a The fragment 3(see Table 2) encodes one putative transcriptional regu-lator proteins predicted to be located in the bacterialinner or outer membrane by PSORT (inorganic pyrophos-phatase sugar transporter putative protein) and a puta-tive integraserecombinase homologous to ripX fromBacillus subtilis The presence of this latter gene sug-gests that fragment 3 may be the remnant of abacteriophage
The integration and excision mechanism is very proba-bly a homologous recombination in the 15 kb sequenceregion B of TnpA carried on the avrPphE transposon(Fig 6) It is not clear whether the integrationexcisionprocess relies on general recombination or on a site-specific recombination dependent on an integraserecombinase As indicated by the sequence of the twotransposons before and after integration of pFKN nonucleotide sequences were altered at the site ofexchange and not a single nucleotide was lost or gained(this excludes a mechanism of transpositional site-specificrecombination) General recombination requires a largeDNA region whereas the site-specific recombinationrequires as few as 240 bp for the lambda phage for exam-ple (Thompson and Landy 1989) The rate of generalrecombination is unknown for P syringae but has beenestimated at 10-3 in Pseudomonas tolaasii (and whenrecA is inactivated at 10-4 Sinha et al 2000) The rateof excision of pFKN has been estimated between 10-3 and10-4 and would therefore be consistent with a mechanismof general recombination Additionally the region B is
15 kb long enough to ensure general recombinationOlder reports describe the integration of pMMC7501 intothe chromosome of P s pv phaseolicola (Curiale andMills 1982) An imprecise excision of pMMC7501 led tothe formation of eight derivative plasmids all sharing thesame origin of replication (Szabo and Mills 1984a) Thisexcision occurred through recombination events betweensets of repeats in the chromosome exhibiting sequencehomology with IS elements (Szabo and Mills 1984bPoplawsky and Mills 1987)
The integration of a virulence gene into the chromo-some may be a first step towards its stabilization in thegenome Indeed lack of selection can lead to the stochas-tic loss of the plasmid and many steps in the pathogenlife cycle probably do not require the presence of virulencefunctions The loss of a plasmid carrying the type III effec-tor avrPpiB was observed in P syringae pv pisi (Bavageet al 1991) We propose that the integration of pFKNcould lead to the stabilization of avrRpm1 in the genomeof Psm M6 Interestingly a DNA segment of at least 42 kbcarrying avrPphB in P syringae pv phaseolicola canexcise from the chromosome but it is not maintained inthe strain as a plasmid This region may represent anancient plasmid integration event followed by successivegenomic rearrangements deleterious for plasmid mainte-nance (Jackson et al 2000) The fact that we can stillobserve the excision of pFKN from the chromosome ofPsm M6integron shows that the newly generated geneticunit is not rapidly stabilized in the genome and that furtherevents might be necessary to prevent excision Further-more the integration of pFKN into the chromosomeslightly impairs the fitness of Psm M6integron The pres-ence of the functional origin of replication of pFKN mayinterfere with the replication of the chromosome Integra-
Fig 6 The integration of pFKN may have occurred via homologous recombination between the regions B common to the two TnpA genes The TnpA genes are constituted of four DNA sequence regions (1) before the integration the chromosomal copy of TnpA contains all four regions (A B C and D) The TnpA on the plasmid is missing the 5cent end of the gene (region A) and a fragment towards the 3cent end (region C) which results in a frameshift (FS) After the homologous recombination between two identical regions (region B) in the coding region of the TnpA genes (2) the region A is found in the TnpA of the left border and region C in the TnpA of the right border (3)
pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

1550 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
lence function of avrRpm1 We compared the growth ofPsm M6C and Psm M6CKO when each carried avrRpm1on a plasmid No difference in growth was observed(Fig 2B) indicating that avrPphE does not affectavrRpm1 virulence function
Identification of additional genes putatively involved in pathogenicity based on their expression pattern
The expression of known P syringae virulence factors isspecifically induced in planta Minimal media (MM) plus asugar source seems to mimic the plant apoplast leadingto the expression of the type III pilus genes and effectors
(Huynh et al 1989 Innes et al 1993) We compared theexpression of the pFKN ORFs in MM and in rich medium(KB) using reverse transcription RT-PCR methodsWhen the expression of an ORF was observed in bothmedia we assumed that the gene was constitutivelyexpressed Expression of a gene in MM but not in KB mayindicate a role for this gene during interaction with thehost Most of the pFKN genes are constitutivelyexpressed However the expression of both avrRpm1(Ritter and Dangl 1995) and avrPphE was detected inMM but not in KB Additionally ORF26 is expressed inMM but not in KB ORFs 14 and 24 were not expressedin either of the two conditions The expression of ORF8
Table 2 pFKN ORFs sorted by descending G+C content
a Some pFKN ORFs are contiguous on the plasmid and are collinear in this tableindicating that they possess a similar G+C content When this is the case an emptyrow separates them from the next ORF in the tableb The DNA fragments from the Pst DC3000 genome that contain sequenceshomologous to ORFs from pFKN were colour-coded (1 green 2 red 3 blue4 pink) The table indicates that fragment 4 has a relatively high G+C contentwhereas fragment 3 has a relatively low content
pFKN a virulence plasmid in P syringae pv maculicola 1551
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
and ORF11 could not be assessed using this techniquedespite many attempts
The six genes (avrRpm1 avrPphE ORF8 ORF11ORF14 and ORF24) for which no constitutive expressionwas observed by RT-PCR were selected for further studyWe constructed transcriptional fusions between the 5cent endof the genes of interest and a gene encoding green fluo-rescent protein (GFP) and chose ORF10 as a constitutivecontrol The fluorescence of PsmM6C carrying the tran-scriptional fusions was assessed in KB MM and in plantaby exposing the bacteria grown in these conditions to bluelight (488 nm) and observing them by fluorescencemicroscopy A strain carrying promoterless GFP did notexhibit any fluorescence in the conditions tested whereas
all cells carrying the ORF10ndashGFP fusion did Only theavrPphE avrRpm1 and ORF8 GFP fusions were active inplants (Table 1)
pFKN can integrate into the Psm M6 chromosome
One colony of Psm M6 derived during routine mainte-nance exhibited unusual features Although this straintriggered recognition (measured by HR) on RPM1 plantsand disease symptoms on rpm1 plants no plasmid couldbe isolated from it AvrRpm1 was still present in totalDNA as assayed by DNA blot (Fig 3A) We thereforeconcluded that avrRpm1 was now located on the PsmM6 chromosome To determine whether additional por-tions of pFKN were also present on the variance chromo-some total DNA from the variant Psm M6 was digestedwith EcoRI HindIII EcoRV SacI PvuII SmaI BglII andNsiI and probed with the DNA from different parts of thepFKN plasmid The DNA of Psm M6 containing pFKNwas used as a positive control This analysis revealed apolymorphism between the avrRpm1 gene in this strain(eg an 80 kb BglII fragment) compared with pFKN (egan 88 kb BglII fragment Fig 3A) All restriction frag-ments from pFKN were detected in the DNA of PsmM6integron Therefore pFKN has integrated into thechromosome For the purpose of clarity the strain bear-ing avrRpm1 in the chromosome will be referred to asPsm M6integron
We have on occasion isolated a replicon from indi-vidual colonies of Psm M6integron (see next section)The plasmid purified from these bacteria always exhib-ited a restriction profile identical to that of pFKNTherefore pFKN can excise from the chromosome andPsm M6integron can revert to Psm M6 Interestinglythe overall fitness of Psm M6 is affected by the integra-tion status of pFKN into the chromosome PsmM6integron grows in media (KB and MM) more slowlythan either the original Psm M6 strain or a revertantfrom Psm M6integron that is cured of the plasmid(Fig 3B) Because Psm M6 (with a pFKN replicon) andthe plasmid-cured derivative of Psm M6integron bothgrew equivalently we can conclude that it is the inte-gration of pFKN per se that results in this growthimpairment This is not uncommon as several plasmidreplicons are known to slow growth of the organismwhen integrated (see Discussion)
The pFKN integration site in Psm M6integron is the avrPphE transposon
We noticed from the DNA blot that the genomic environ-ment of avrRpm1 is different in Psm M6 and PsmM6integron This is also the case for both copies of avrP-phE (data not shown) suggesting that the pFKN integra-tion site is near avrRpm1 and avrPphE and that the
Fig 2 avrRpm1 encodes the determining virulence factor on pFKNA The growth of Psm M6C on rpm1-3 plants was compared with the growth of the wild-type Psm M6 strain and Psm M6C strains comple-mented with different fragments from pFKN The pFKN subclone carrying avrRpm1 restored full virulence to Psm M6C (the other subclones did not restore any virulence and are not shown) Leaves were hand inoculated with 105 cfu ml-1 and a titration of bacteria in the leaves was performed 3 days after inoculation Data points rep-resent the mean plusmn standard error of four samplesB The avrPphE gene was deleted from the chromosome of Psm M6C to generate Psm M6CKO To assess the virulence function of avrPphE or its influence on the virulence function of avrRpm1 the growth of Psm M6CKO on rpm1-3 plants was compared with the growth of Psm M6C Psm M6C + avrRpm1 and Psm M6CKO + avrRpm1 Data points represent the mean plusmn standard error of four samples
1552 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
chromosomal location into which the plasmid integrates isnear the chromosomal copy of avrPphE We first definedthat the chromosomal avrPphE is carried on the sametransposon found in the sequence of pFKN using PCRamplification on total DNA of Psm M6C with primersspecific to sequence on the avrPphE transposon(Table 5) The PCR products were sequenced and com-pared with the corresponding sequenced regions ofpFKN As shown in Fig 4A the chromosomal copy of thetransposon contains two sequences not present on pFKNOne 1244 bp sequence (region A) is located betweenavrPphE and the coding region of TnpA It carries an ORFwith a sequence that is nearly identical to various genesencoding resolvases of Tn3 transposons Additionally aportion of 661 bp of this region encodes the 5cent portion ofTnpA Another region of 271 bp unique to the chromo-somal copy is located within the coding region of thetransposase (region C) and is 3cent from a region shared byboth pFKN and chromosomal copies of TnpA (region B)The addition of regions A and C to the chromosomal copyresults in a complete TnpA ORF correcting the frameshiftat the 3cent end of TnpA carried on pFKN
TnpA avrPphE and the putative resolvase fragmentare located between nearly identical inverted repeats of66 bp on both the plasmid and the chromosome Theirorganization is reminiscent of Tn3-like transposons(Sherrat 1989) Moreover this TnpA is 81 identical tothose of the Tn21 subfamily of Tn3 elements (Diveret al 1983) As we suspected that the integration site isin the vicinity of avrPphE in the chromosome and theplasmid (ie in the two transposons) we analysed byPCR the two transposons in the genome of PsmM6integron Figure 4B shows that the unique regions Aand C are no longer located on the same transposonbut are instead separated by the entire plasmidsequence (Fig 4C) This indicated that the integrationsite of the plasmid is located in the region betweenregion A and region C ie within the homology providedby region B (Fig 4)
We used this sequence data to investigate the rateof excision occurring in Psm M6integron grown indifferent conditions by PCR The rate of excision wasestimated at around 10-4 (data not shown) No differencein rate of excision could be detected between bacteria
Fig 3 pFKN can integrate into the Psm M6 chromosomeA Total DNA from Psm M6 (lanes F) and Psm M6integron (lanes I) was digested with EcoRI HindIII EcoRV SacI PvuII SmaI BglII and NsiI and probed with various pFKN fragments Here the DNA blot was probed with the avrRpm1 ORF The two DNA samples exhibit a similar pattern except for a polymorphism (eg EcoRI BglII and SmaI) near the site of pFKN integra-tion into the chromosomeB The integration of the plasmid alters the overall fitness of Psm M6integron The growth of Psm M6integron in KB (empty symbols) and MM (filled symbols) was compared with the growth of Psm M6 and Psm M6C Bacteria were inoculated at 104 cfu ml-1 and cultures were titrated at 0 6 12 29 32 54 and 60 h after the inoculation Psm M6pFKN is indicated by squares Psm M6C is indicated by triangles and rhombi indicate Psm M6integron Data points represent the mean plusmn standard error of four samples (some error bars are not visible because too small) Psm M6pFKN and Psm M6C grew equally well in both minimal medium and KB Psm M6integron grows slower than Psm M6pFKN and Psm M6C in both media
pFKN a virulence plasmid in P syringae pv maculicola 1553
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
grown in KB MM and on resistant or susceptibleplants
The genomic context of avrPphE is different in various P syringae strains
We investigated the regions surrounding the avrPphEtransposon on the chromosome of Psm M6 (Fig 5) byTAIL-PCR The left border shows 97 nt identity to the 5centend of the insertion sequence ISPsy3 (AB063176) fromP syringae pv eriobotryae This element is interrupted bythe avrPphE transposon The translation product of theinterrupted ORF found on this border is homologous tothe IS801 transposase (79 identity and 86 homology)
and IS801 has been associated with other type III effec-tors in various P syringae pathovars (Kim et al 1998)On the right border the 200 bp sequence following theterminal repeat of the avrPphE transposon encodes aprotein homologous (81 nucleotide identity) to the 3centend of an unrelated putative transposase gene fromPseudomonas aeruginosa The next 1062 bp sequencedownstream shows high nucleotide homology tosequence identified in P syringae pv pisi (Arnold et al2001) This DNA fragment may be a transposon remnantand carries the avrPpiA1 allele of avrRpm1 from P syrin-gae pv pisi among other genes In the chromosome ofPsm M6 this sequence shows homology to the P s pisigenes ORFD (94 identity) encoding a putative trans-
Fig 4 The pFKN integration site is a second copy of the avrPphE transposonA The region in the Psm M6 chromosome that hybridizes to the avrPphE and TnpA probes was amplified and sequenced The sequence of the chromosomal region is 100 identical to the transposon in the plasmid but contains two additional regions (regions A and C) TnpA encodes a transposase and res a resolvase Based on homology the chromosomal avrPphE transposon could be complete and functional The avrPphE transposon carried on the plas-mid lacks the 5cent end of TnpA and res (region A) and the absence of region C results in the truncation and a frameshift (marked FS) of the gene encoding the transposase The primers used for PCR amplification (Fig 4B) are indi-cated by arrows and their names are in boldB To investigate where regions A and C were distributed after integration of the plasmid into the chromosome PCRs were performed on the DNA of Psm M6integron and Psm M6 using Larp3 and K110 and K110 and K113 (see Fig 4A) The product amplified from Psm M6integron is run in lane I The product ampli-fied from Psm M6 is run in lane F The product amplified from Psm M6C is run in lane C The difference in size observed between the PCR products from Psm M6integron and Psm M6 using Larp3 and K110 indicates that regions A and C are not carried on the same putative transposon in the chromosome of Psm M6integron (in contrast to the chromosome of Psm M6) The PCR products from Psm M6integron and Psm M6 using K113 and K110 show that neither region A nor region C has been lost in the chromosome of Psm M6integron The identical size of the PCR prod-ucts from Psm M6integron and Psm M6 using K113 and Larp2 indicates that region C is on the right border of pFKN when integrated into the chromosome The region A is on the left border as it is carried by the other putative transposonC After integration of pFKN into the chromo-some TnpA on the left border contains region A but not region C TnpA on the right border contains region C The primers used for the PCR amplification are indicated by arrows and their names are in bold (in Fig 4A)
1554 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
posase and ORF3 encoding a putative type III effector(100 identity) Thus this region in Psm M6 is a remnantof the putative transposon found in P s pisi
We detected the avrPphE transposon in other strains ofP syringae pv maculicola and in the sequence of the
genome of P s tomato DC3000 In Psm M2 Psm M8Psm M12 Psm m90 and Psm m10831 the borders arethe same as in Psm M6 (data not shown) On the otherhand they differ in Psm M5 and Psm M9 and these alldiffer from Pst DC3000 In Psm M5 (Fig 5) and Psm M9
Fig 5 Sequences surrounding the avrPphE transposon define four different genetic contexts The figure is a graphical representation of the four genomic environments in which the avrPphE transposon was detected The regions represented were characterized by sequence comparison using the BLASTX algorithm It is not known whether the avrPphE transposon is located in the chromosome or on a plasmid in the genome of Psm M5 and Pst DC3000
pFKN a virulence plasmid in P syringae pv maculicola 1555
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(not shown) the left border consists of the 5cent end of agene sharing 99 identity to the replicase of pPS10 aplasmid found in P syringae pv savastanoi (Nieto et al1992) This replicase is not related to that of pFKN (aPT23A-like plasmid see above) The right bordersequence is a remnant of a gene encoding a putative ABCtransporter (23 identity and 43 homology to a putativesugar uptake ABC transporter ATP-binding protein fromSinorhizobium meliloti)
In the genome of Pst DC3000 we looked at 30 kbsurrounding the transposon (Fig 5) The proteinsencoded on this 30 kb region share homology to eitherDNA mobility functions (transposases plasmid partition-ing proteins replicases) or type III effectors (identical toavrPpiB Cournoyer et al 1995) The region also containsother sequences often found associated with plasmidsand genes involved in pathogenicity such as rulA and rulB(Sesma et al 2000) Sequences directly neighbouringthe transposon on the left side are homologous to a geneencoding the transposase of the IS100 elements fromYersinia pestis Ralstonia solanacearum and Salmonellaenterica On the right-hand side of the avrPphE transpo-son the first homology is to a fragment of a putative ABCtransporter identical to the one identified in Psm M5 Thissuggests that this part of the two genomes is morerecently diverged than either is from Psm M6
No common sequence was identified in the four differ-ent environments surrounding the avrPphE transposonthat would help to identify the integration target of theavrPphE transposon although the proximal right bordersin Psm M5 and Pst DC3000 are nearly identical Thissuggests that the avrPphE transposon and this border-ing sequence were acquired together by these twostrains and that subsequent events at both the distalright flank and on the left border are responsible for thedifferences in genomic context In contrast the insertionsites of the avrPphE transposon in Psm M5 and PsmM6 are completely different In sum the features ofthese integration sites are consistent with the model thatPAIs evolve by repeated insertion and deletion intopre-existing PAIs (Hensel et al 1999 Tauschek et al2002)
Discussion
We sequenced pFKN an ordf40 kb plasmid that contributesto the virulence of Psm M6 The plasmid carries two typeIII effectors on remnants of transposons contiguous toORF8 itself neighbouring an element closely related toIS801 avrRpm1 encodes a known virulence factor andavrPphE a type III effector The expression of ORF8 isinduced in planta and therefore ORF8 may play a role inthe interaction with the host plant The portion of theplasmid carrying avrRpm1 avrPphE and ORF8 may thus
be considered as a PAI Interestingly the G+C content ofthe ORFs carried on this region are very heterogeneousand the three genes are flanked by sequences related tomobile elements and therefore are likely to have differentorigins This is consistent with the idea that the formationof PAIs requires a multistep process (Hensel et al 1999Alfano et al 2000 Tauschek et al 2002) It is also con-sistent with the findings of Kim et al (1998) showing thatmost of the known type III effectors are associated withthis type of sequence pFKN (and therefore the PAI that itcarries) sometimes integrates into and can excise fromthe chromosome as demonstrated by the discovery of thevariant of the strain Psm M6 We identified the region ofthe chromosome into which the integration took placeThis region is a transposon carrying a second copy ofavrPphE
Alleles of the avrPphE gene have been detected inmany P syringae pathovars (Mansfield et al 1994Alfano et al 2000 Fouts et al 2002 Guttman et al2002) suggesting that it provides an important virulencefunction on a variety of host plants Our findings are thefirst to associate avrPphE with a transposon In PsyB728a and in Pph 1302A for example avrPphE residesdirectly next to the hrp locus in a region called thelsquoexchangeable effector locusrsquo or EEL (Alfano et al 2000)and is not associated with a transposase In fact in PsyB728a avrPphE is predicted to be the third gene in anoperon controlled by the hrpK promoter (Alfano et al2000) This striking finding illustrates the dynamism ofPAIs
We determined by complementation assay that the par-tial loss of virulence in the strain cured of pFKN resultsfrom the absence of avrRpm1 The virulence function ofavrRpm1 has already been demonstrated in the strainPsm M2 (Ritter and Dangl 1995) Recently AvrRpm1 hasbeen demonstrated to interact with RIN4 a proteininvolved in the regulation of some of the plant defenceresponses (Mackey et al 2002) Mackey and coworkersproposed that the role of AvrRpm1 is to reduce the levelof plant defence response triggered by the attack of thepathogen This hypothesis might be consistent with thefact that only quantitative differences in growth wereobserved between Psm M6C and Psm M6 No differencein growth was observed between Psm M6C and PsmM6CKO (lacking avrPphE) Similar results were observedwith the avrPphE allele of P s phaseolicola (Stevenset al 1998) This observation could be explained byeither a functional redundancy with another gene in thegenome of the Psm M6 or the fact that the assay isinappropriate to assess the virulence function of this typeIII effector
The sequence and the integration mechanism of pFKNinto the Psm M6 chromosome provide insights on theevolutionary dynamics of the genomes of P syringae
1556 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
pathovars First the analysis of the G+C content of frag-ments of pFKN suggests that they may have differentorigins This hypothesis is further supported by the pres-ence of some defined fragments from pFKN in thegenomes of Pst DC3000 and Psy B728a The fragment 3(see Table 2) encodes one putative transcriptional regu-lator proteins predicted to be located in the bacterialinner or outer membrane by PSORT (inorganic pyrophos-phatase sugar transporter putative protein) and a puta-tive integraserecombinase homologous to ripX fromBacillus subtilis The presence of this latter gene sug-gests that fragment 3 may be the remnant of abacteriophage
The integration and excision mechanism is very proba-bly a homologous recombination in the 15 kb sequenceregion B of TnpA carried on the avrPphE transposon(Fig 6) It is not clear whether the integrationexcisionprocess relies on general recombination or on a site-specific recombination dependent on an integraserecombinase As indicated by the sequence of the twotransposons before and after integration of pFKN nonucleotide sequences were altered at the site ofexchange and not a single nucleotide was lost or gained(this excludes a mechanism of transpositional site-specificrecombination) General recombination requires a largeDNA region whereas the site-specific recombinationrequires as few as 240 bp for the lambda phage for exam-ple (Thompson and Landy 1989) The rate of generalrecombination is unknown for P syringae but has beenestimated at 10-3 in Pseudomonas tolaasii (and whenrecA is inactivated at 10-4 Sinha et al 2000) The rateof excision of pFKN has been estimated between 10-3 and10-4 and would therefore be consistent with a mechanismof general recombination Additionally the region B is
15 kb long enough to ensure general recombinationOlder reports describe the integration of pMMC7501 intothe chromosome of P s pv phaseolicola (Curiale andMills 1982) An imprecise excision of pMMC7501 led tothe formation of eight derivative plasmids all sharing thesame origin of replication (Szabo and Mills 1984a) Thisexcision occurred through recombination events betweensets of repeats in the chromosome exhibiting sequencehomology with IS elements (Szabo and Mills 1984bPoplawsky and Mills 1987)
The integration of a virulence gene into the chromo-some may be a first step towards its stabilization in thegenome Indeed lack of selection can lead to the stochas-tic loss of the plasmid and many steps in the pathogenlife cycle probably do not require the presence of virulencefunctions The loss of a plasmid carrying the type III effec-tor avrPpiB was observed in P syringae pv pisi (Bavageet al 1991) We propose that the integration of pFKNcould lead to the stabilization of avrRpm1 in the genomeof Psm M6 Interestingly a DNA segment of at least 42 kbcarrying avrPphB in P syringae pv phaseolicola canexcise from the chromosome but it is not maintained inthe strain as a plasmid This region may represent anancient plasmid integration event followed by successivegenomic rearrangements deleterious for plasmid mainte-nance (Jackson et al 2000) The fact that we can stillobserve the excision of pFKN from the chromosome ofPsm M6integron shows that the newly generated geneticunit is not rapidly stabilized in the genome and that furtherevents might be necessary to prevent excision Further-more the integration of pFKN into the chromosomeslightly impairs the fitness of Psm M6integron The pres-ence of the functional origin of replication of pFKN mayinterfere with the replication of the chromosome Integra-
Fig 6 The integration of pFKN may have occurred via homologous recombination between the regions B common to the two TnpA genes The TnpA genes are constituted of four DNA sequence regions (1) before the integration the chromosomal copy of TnpA contains all four regions (A B C and D) The TnpA on the plasmid is missing the 5cent end of the gene (region A) and a fragment towards the 3cent end (region C) which results in a frameshift (FS) After the homologous recombination between two identical regions (region B) in the coding region of the TnpA genes (2) the region A is found in the TnpA of the left border and region C in the TnpA of the right border (3)
pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

pFKN a virulence plasmid in P syringae pv maculicola 1551
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
and ORF11 could not be assessed using this techniquedespite many attempts
The six genes (avrRpm1 avrPphE ORF8 ORF11ORF14 and ORF24) for which no constitutive expressionwas observed by RT-PCR were selected for further studyWe constructed transcriptional fusions between the 5cent endof the genes of interest and a gene encoding green fluo-rescent protein (GFP) and chose ORF10 as a constitutivecontrol The fluorescence of PsmM6C carrying the tran-scriptional fusions was assessed in KB MM and in plantaby exposing the bacteria grown in these conditions to bluelight (488 nm) and observing them by fluorescencemicroscopy A strain carrying promoterless GFP did notexhibit any fluorescence in the conditions tested whereas
all cells carrying the ORF10ndashGFP fusion did Only theavrPphE avrRpm1 and ORF8 GFP fusions were active inplants (Table 1)
pFKN can integrate into the Psm M6 chromosome
One colony of Psm M6 derived during routine mainte-nance exhibited unusual features Although this straintriggered recognition (measured by HR) on RPM1 plantsand disease symptoms on rpm1 plants no plasmid couldbe isolated from it AvrRpm1 was still present in totalDNA as assayed by DNA blot (Fig 3A) We thereforeconcluded that avrRpm1 was now located on the PsmM6 chromosome To determine whether additional por-tions of pFKN were also present on the variance chromo-some total DNA from the variant Psm M6 was digestedwith EcoRI HindIII EcoRV SacI PvuII SmaI BglII andNsiI and probed with the DNA from different parts of thepFKN plasmid The DNA of Psm M6 containing pFKNwas used as a positive control This analysis revealed apolymorphism between the avrRpm1 gene in this strain(eg an 80 kb BglII fragment) compared with pFKN (egan 88 kb BglII fragment Fig 3A) All restriction frag-ments from pFKN were detected in the DNA of PsmM6integron Therefore pFKN has integrated into thechromosome For the purpose of clarity the strain bear-ing avrRpm1 in the chromosome will be referred to asPsm M6integron
We have on occasion isolated a replicon from indi-vidual colonies of Psm M6integron (see next section)The plasmid purified from these bacteria always exhib-ited a restriction profile identical to that of pFKNTherefore pFKN can excise from the chromosome andPsm M6integron can revert to Psm M6 Interestinglythe overall fitness of Psm M6 is affected by the integra-tion status of pFKN into the chromosome PsmM6integron grows in media (KB and MM) more slowlythan either the original Psm M6 strain or a revertantfrom Psm M6integron that is cured of the plasmid(Fig 3B) Because Psm M6 (with a pFKN replicon) andthe plasmid-cured derivative of Psm M6integron bothgrew equivalently we can conclude that it is the inte-gration of pFKN per se that results in this growthimpairment This is not uncommon as several plasmidreplicons are known to slow growth of the organismwhen integrated (see Discussion)
The pFKN integration site in Psm M6integron is the avrPphE transposon
We noticed from the DNA blot that the genomic environ-ment of avrRpm1 is different in Psm M6 and PsmM6integron This is also the case for both copies of avrP-phE (data not shown) suggesting that the pFKN integra-tion site is near avrRpm1 and avrPphE and that the
Fig 2 avrRpm1 encodes the determining virulence factor on pFKNA The growth of Psm M6C on rpm1-3 plants was compared with the growth of the wild-type Psm M6 strain and Psm M6C strains comple-mented with different fragments from pFKN The pFKN subclone carrying avrRpm1 restored full virulence to Psm M6C (the other subclones did not restore any virulence and are not shown) Leaves were hand inoculated with 105 cfu ml-1 and a titration of bacteria in the leaves was performed 3 days after inoculation Data points rep-resent the mean plusmn standard error of four samplesB The avrPphE gene was deleted from the chromosome of Psm M6C to generate Psm M6CKO To assess the virulence function of avrPphE or its influence on the virulence function of avrRpm1 the growth of Psm M6CKO on rpm1-3 plants was compared with the growth of Psm M6C Psm M6C + avrRpm1 and Psm M6CKO + avrRpm1 Data points represent the mean plusmn standard error of four samples
1552 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
chromosomal location into which the plasmid integrates isnear the chromosomal copy of avrPphE We first definedthat the chromosomal avrPphE is carried on the sametransposon found in the sequence of pFKN using PCRamplification on total DNA of Psm M6C with primersspecific to sequence on the avrPphE transposon(Table 5) The PCR products were sequenced and com-pared with the corresponding sequenced regions ofpFKN As shown in Fig 4A the chromosomal copy of thetransposon contains two sequences not present on pFKNOne 1244 bp sequence (region A) is located betweenavrPphE and the coding region of TnpA It carries an ORFwith a sequence that is nearly identical to various genesencoding resolvases of Tn3 transposons Additionally aportion of 661 bp of this region encodes the 5cent portion ofTnpA Another region of 271 bp unique to the chromo-somal copy is located within the coding region of thetransposase (region C) and is 3cent from a region shared byboth pFKN and chromosomal copies of TnpA (region B)The addition of regions A and C to the chromosomal copyresults in a complete TnpA ORF correcting the frameshiftat the 3cent end of TnpA carried on pFKN
TnpA avrPphE and the putative resolvase fragmentare located between nearly identical inverted repeats of66 bp on both the plasmid and the chromosome Theirorganization is reminiscent of Tn3-like transposons(Sherrat 1989) Moreover this TnpA is 81 identical tothose of the Tn21 subfamily of Tn3 elements (Diveret al 1983) As we suspected that the integration site isin the vicinity of avrPphE in the chromosome and theplasmid (ie in the two transposons) we analysed byPCR the two transposons in the genome of PsmM6integron Figure 4B shows that the unique regions Aand C are no longer located on the same transposonbut are instead separated by the entire plasmidsequence (Fig 4C) This indicated that the integrationsite of the plasmid is located in the region betweenregion A and region C ie within the homology providedby region B (Fig 4)
We used this sequence data to investigate the rateof excision occurring in Psm M6integron grown indifferent conditions by PCR The rate of excision wasestimated at around 10-4 (data not shown) No differencein rate of excision could be detected between bacteria
Fig 3 pFKN can integrate into the Psm M6 chromosomeA Total DNA from Psm M6 (lanes F) and Psm M6integron (lanes I) was digested with EcoRI HindIII EcoRV SacI PvuII SmaI BglII and NsiI and probed with various pFKN fragments Here the DNA blot was probed with the avrRpm1 ORF The two DNA samples exhibit a similar pattern except for a polymorphism (eg EcoRI BglII and SmaI) near the site of pFKN integra-tion into the chromosomeB The integration of the plasmid alters the overall fitness of Psm M6integron The growth of Psm M6integron in KB (empty symbols) and MM (filled symbols) was compared with the growth of Psm M6 and Psm M6C Bacteria were inoculated at 104 cfu ml-1 and cultures were titrated at 0 6 12 29 32 54 and 60 h after the inoculation Psm M6pFKN is indicated by squares Psm M6C is indicated by triangles and rhombi indicate Psm M6integron Data points represent the mean plusmn standard error of four samples (some error bars are not visible because too small) Psm M6pFKN and Psm M6C grew equally well in both minimal medium and KB Psm M6integron grows slower than Psm M6pFKN and Psm M6C in both media
pFKN a virulence plasmid in P syringae pv maculicola 1553
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
grown in KB MM and on resistant or susceptibleplants
The genomic context of avrPphE is different in various P syringae strains
We investigated the regions surrounding the avrPphEtransposon on the chromosome of Psm M6 (Fig 5) byTAIL-PCR The left border shows 97 nt identity to the 5centend of the insertion sequence ISPsy3 (AB063176) fromP syringae pv eriobotryae This element is interrupted bythe avrPphE transposon The translation product of theinterrupted ORF found on this border is homologous tothe IS801 transposase (79 identity and 86 homology)
and IS801 has been associated with other type III effec-tors in various P syringae pathovars (Kim et al 1998)On the right border the 200 bp sequence following theterminal repeat of the avrPphE transposon encodes aprotein homologous (81 nucleotide identity) to the 3centend of an unrelated putative transposase gene fromPseudomonas aeruginosa The next 1062 bp sequencedownstream shows high nucleotide homology tosequence identified in P syringae pv pisi (Arnold et al2001) This DNA fragment may be a transposon remnantand carries the avrPpiA1 allele of avrRpm1 from P syrin-gae pv pisi among other genes In the chromosome ofPsm M6 this sequence shows homology to the P s pisigenes ORFD (94 identity) encoding a putative trans-
Fig 4 The pFKN integration site is a second copy of the avrPphE transposonA The region in the Psm M6 chromosome that hybridizes to the avrPphE and TnpA probes was amplified and sequenced The sequence of the chromosomal region is 100 identical to the transposon in the plasmid but contains two additional regions (regions A and C) TnpA encodes a transposase and res a resolvase Based on homology the chromosomal avrPphE transposon could be complete and functional The avrPphE transposon carried on the plas-mid lacks the 5cent end of TnpA and res (region A) and the absence of region C results in the truncation and a frameshift (marked FS) of the gene encoding the transposase The primers used for PCR amplification (Fig 4B) are indi-cated by arrows and their names are in boldB To investigate where regions A and C were distributed after integration of the plasmid into the chromosome PCRs were performed on the DNA of Psm M6integron and Psm M6 using Larp3 and K110 and K110 and K113 (see Fig 4A) The product amplified from Psm M6integron is run in lane I The product ampli-fied from Psm M6 is run in lane F The product amplified from Psm M6C is run in lane C The difference in size observed between the PCR products from Psm M6integron and Psm M6 using Larp3 and K110 indicates that regions A and C are not carried on the same putative transposon in the chromosome of Psm M6integron (in contrast to the chromosome of Psm M6) The PCR products from Psm M6integron and Psm M6 using K113 and K110 show that neither region A nor region C has been lost in the chromosome of Psm M6integron The identical size of the PCR prod-ucts from Psm M6integron and Psm M6 using K113 and Larp2 indicates that region C is on the right border of pFKN when integrated into the chromosome The region A is on the left border as it is carried by the other putative transposonC After integration of pFKN into the chromo-some TnpA on the left border contains region A but not region C TnpA on the right border contains region C The primers used for the PCR amplification are indicated by arrows and their names are in bold (in Fig 4A)
1554 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
posase and ORF3 encoding a putative type III effector(100 identity) Thus this region in Psm M6 is a remnantof the putative transposon found in P s pisi
We detected the avrPphE transposon in other strains ofP syringae pv maculicola and in the sequence of the
genome of P s tomato DC3000 In Psm M2 Psm M8Psm M12 Psm m90 and Psm m10831 the borders arethe same as in Psm M6 (data not shown) On the otherhand they differ in Psm M5 and Psm M9 and these alldiffer from Pst DC3000 In Psm M5 (Fig 5) and Psm M9
Fig 5 Sequences surrounding the avrPphE transposon define four different genetic contexts The figure is a graphical representation of the four genomic environments in which the avrPphE transposon was detected The regions represented were characterized by sequence comparison using the BLASTX algorithm It is not known whether the avrPphE transposon is located in the chromosome or on a plasmid in the genome of Psm M5 and Pst DC3000
pFKN a virulence plasmid in P syringae pv maculicola 1555
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(not shown) the left border consists of the 5cent end of agene sharing 99 identity to the replicase of pPS10 aplasmid found in P syringae pv savastanoi (Nieto et al1992) This replicase is not related to that of pFKN (aPT23A-like plasmid see above) The right bordersequence is a remnant of a gene encoding a putative ABCtransporter (23 identity and 43 homology to a putativesugar uptake ABC transporter ATP-binding protein fromSinorhizobium meliloti)
In the genome of Pst DC3000 we looked at 30 kbsurrounding the transposon (Fig 5) The proteinsencoded on this 30 kb region share homology to eitherDNA mobility functions (transposases plasmid partition-ing proteins replicases) or type III effectors (identical toavrPpiB Cournoyer et al 1995) The region also containsother sequences often found associated with plasmidsand genes involved in pathogenicity such as rulA and rulB(Sesma et al 2000) Sequences directly neighbouringthe transposon on the left side are homologous to a geneencoding the transposase of the IS100 elements fromYersinia pestis Ralstonia solanacearum and Salmonellaenterica On the right-hand side of the avrPphE transpo-son the first homology is to a fragment of a putative ABCtransporter identical to the one identified in Psm M5 Thissuggests that this part of the two genomes is morerecently diverged than either is from Psm M6
No common sequence was identified in the four differ-ent environments surrounding the avrPphE transposonthat would help to identify the integration target of theavrPphE transposon although the proximal right bordersin Psm M5 and Pst DC3000 are nearly identical Thissuggests that the avrPphE transposon and this border-ing sequence were acquired together by these twostrains and that subsequent events at both the distalright flank and on the left border are responsible for thedifferences in genomic context In contrast the insertionsites of the avrPphE transposon in Psm M5 and PsmM6 are completely different In sum the features ofthese integration sites are consistent with the model thatPAIs evolve by repeated insertion and deletion intopre-existing PAIs (Hensel et al 1999 Tauschek et al2002)
Discussion
We sequenced pFKN an ordf40 kb plasmid that contributesto the virulence of Psm M6 The plasmid carries two typeIII effectors on remnants of transposons contiguous toORF8 itself neighbouring an element closely related toIS801 avrRpm1 encodes a known virulence factor andavrPphE a type III effector The expression of ORF8 isinduced in planta and therefore ORF8 may play a role inthe interaction with the host plant The portion of theplasmid carrying avrRpm1 avrPphE and ORF8 may thus
be considered as a PAI Interestingly the G+C content ofthe ORFs carried on this region are very heterogeneousand the three genes are flanked by sequences related tomobile elements and therefore are likely to have differentorigins This is consistent with the idea that the formationof PAIs requires a multistep process (Hensel et al 1999Alfano et al 2000 Tauschek et al 2002) It is also con-sistent with the findings of Kim et al (1998) showing thatmost of the known type III effectors are associated withthis type of sequence pFKN (and therefore the PAI that itcarries) sometimes integrates into and can excise fromthe chromosome as demonstrated by the discovery of thevariant of the strain Psm M6 We identified the region ofthe chromosome into which the integration took placeThis region is a transposon carrying a second copy ofavrPphE
Alleles of the avrPphE gene have been detected inmany P syringae pathovars (Mansfield et al 1994Alfano et al 2000 Fouts et al 2002 Guttman et al2002) suggesting that it provides an important virulencefunction on a variety of host plants Our findings are thefirst to associate avrPphE with a transposon In PsyB728a and in Pph 1302A for example avrPphE residesdirectly next to the hrp locus in a region called thelsquoexchangeable effector locusrsquo or EEL (Alfano et al 2000)and is not associated with a transposase In fact in PsyB728a avrPphE is predicted to be the third gene in anoperon controlled by the hrpK promoter (Alfano et al2000) This striking finding illustrates the dynamism ofPAIs
We determined by complementation assay that the par-tial loss of virulence in the strain cured of pFKN resultsfrom the absence of avrRpm1 The virulence function ofavrRpm1 has already been demonstrated in the strainPsm M2 (Ritter and Dangl 1995) Recently AvrRpm1 hasbeen demonstrated to interact with RIN4 a proteininvolved in the regulation of some of the plant defenceresponses (Mackey et al 2002) Mackey and coworkersproposed that the role of AvrRpm1 is to reduce the levelof plant defence response triggered by the attack of thepathogen This hypothesis might be consistent with thefact that only quantitative differences in growth wereobserved between Psm M6C and Psm M6 No differencein growth was observed between Psm M6C and PsmM6CKO (lacking avrPphE) Similar results were observedwith the avrPphE allele of P s phaseolicola (Stevenset al 1998) This observation could be explained byeither a functional redundancy with another gene in thegenome of the Psm M6 or the fact that the assay isinappropriate to assess the virulence function of this typeIII effector
The sequence and the integration mechanism of pFKNinto the Psm M6 chromosome provide insights on theevolutionary dynamics of the genomes of P syringae
1556 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
pathovars First the analysis of the G+C content of frag-ments of pFKN suggests that they may have differentorigins This hypothesis is further supported by the pres-ence of some defined fragments from pFKN in thegenomes of Pst DC3000 and Psy B728a The fragment 3(see Table 2) encodes one putative transcriptional regu-lator proteins predicted to be located in the bacterialinner or outer membrane by PSORT (inorganic pyrophos-phatase sugar transporter putative protein) and a puta-tive integraserecombinase homologous to ripX fromBacillus subtilis The presence of this latter gene sug-gests that fragment 3 may be the remnant of abacteriophage
The integration and excision mechanism is very proba-bly a homologous recombination in the 15 kb sequenceregion B of TnpA carried on the avrPphE transposon(Fig 6) It is not clear whether the integrationexcisionprocess relies on general recombination or on a site-specific recombination dependent on an integraserecombinase As indicated by the sequence of the twotransposons before and after integration of pFKN nonucleotide sequences were altered at the site ofexchange and not a single nucleotide was lost or gained(this excludes a mechanism of transpositional site-specificrecombination) General recombination requires a largeDNA region whereas the site-specific recombinationrequires as few as 240 bp for the lambda phage for exam-ple (Thompson and Landy 1989) The rate of generalrecombination is unknown for P syringae but has beenestimated at 10-3 in Pseudomonas tolaasii (and whenrecA is inactivated at 10-4 Sinha et al 2000) The rateof excision of pFKN has been estimated between 10-3 and10-4 and would therefore be consistent with a mechanismof general recombination Additionally the region B is
15 kb long enough to ensure general recombinationOlder reports describe the integration of pMMC7501 intothe chromosome of P s pv phaseolicola (Curiale andMills 1982) An imprecise excision of pMMC7501 led tothe formation of eight derivative plasmids all sharing thesame origin of replication (Szabo and Mills 1984a) Thisexcision occurred through recombination events betweensets of repeats in the chromosome exhibiting sequencehomology with IS elements (Szabo and Mills 1984bPoplawsky and Mills 1987)
The integration of a virulence gene into the chromo-some may be a first step towards its stabilization in thegenome Indeed lack of selection can lead to the stochas-tic loss of the plasmid and many steps in the pathogenlife cycle probably do not require the presence of virulencefunctions The loss of a plasmid carrying the type III effec-tor avrPpiB was observed in P syringae pv pisi (Bavageet al 1991) We propose that the integration of pFKNcould lead to the stabilization of avrRpm1 in the genomeof Psm M6 Interestingly a DNA segment of at least 42 kbcarrying avrPphB in P syringae pv phaseolicola canexcise from the chromosome but it is not maintained inthe strain as a plasmid This region may represent anancient plasmid integration event followed by successivegenomic rearrangements deleterious for plasmid mainte-nance (Jackson et al 2000) The fact that we can stillobserve the excision of pFKN from the chromosome ofPsm M6integron shows that the newly generated geneticunit is not rapidly stabilized in the genome and that furtherevents might be necessary to prevent excision Further-more the integration of pFKN into the chromosomeslightly impairs the fitness of Psm M6integron The pres-ence of the functional origin of replication of pFKN mayinterfere with the replication of the chromosome Integra-
Fig 6 The integration of pFKN may have occurred via homologous recombination between the regions B common to the two TnpA genes The TnpA genes are constituted of four DNA sequence regions (1) before the integration the chromosomal copy of TnpA contains all four regions (A B C and D) The TnpA on the plasmid is missing the 5cent end of the gene (region A) and a fragment towards the 3cent end (region C) which results in a frameshift (FS) After the homologous recombination between two identical regions (region B) in the coding region of the TnpA genes (2) the region A is found in the TnpA of the left border and region C in the TnpA of the right border (3)
pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

1552 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
chromosomal location into which the plasmid integrates isnear the chromosomal copy of avrPphE We first definedthat the chromosomal avrPphE is carried on the sametransposon found in the sequence of pFKN using PCRamplification on total DNA of Psm M6C with primersspecific to sequence on the avrPphE transposon(Table 5) The PCR products were sequenced and com-pared with the corresponding sequenced regions ofpFKN As shown in Fig 4A the chromosomal copy of thetransposon contains two sequences not present on pFKNOne 1244 bp sequence (region A) is located betweenavrPphE and the coding region of TnpA It carries an ORFwith a sequence that is nearly identical to various genesencoding resolvases of Tn3 transposons Additionally aportion of 661 bp of this region encodes the 5cent portion ofTnpA Another region of 271 bp unique to the chromo-somal copy is located within the coding region of thetransposase (region C) and is 3cent from a region shared byboth pFKN and chromosomal copies of TnpA (region B)The addition of regions A and C to the chromosomal copyresults in a complete TnpA ORF correcting the frameshiftat the 3cent end of TnpA carried on pFKN
TnpA avrPphE and the putative resolvase fragmentare located between nearly identical inverted repeats of66 bp on both the plasmid and the chromosome Theirorganization is reminiscent of Tn3-like transposons(Sherrat 1989) Moreover this TnpA is 81 identical tothose of the Tn21 subfamily of Tn3 elements (Diveret al 1983) As we suspected that the integration site isin the vicinity of avrPphE in the chromosome and theplasmid (ie in the two transposons) we analysed byPCR the two transposons in the genome of PsmM6integron Figure 4B shows that the unique regions Aand C are no longer located on the same transposonbut are instead separated by the entire plasmidsequence (Fig 4C) This indicated that the integrationsite of the plasmid is located in the region betweenregion A and region C ie within the homology providedby region B (Fig 4)
We used this sequence data to investigate the rateof excision occurring in Psm M6integron grown indifferent conditions by PCR The rate of excision wasestimated at around 10-4 (data not shown) No differencein rate of excision could be detected between bacteria
Fig 3 pFKN can integrate into the Psm M6 chromosomeA Total DNA from Psm M6 (lanes F) and Psm M6integron (lanes I) was digested with EcoRI HindIII EcoRV SacI PvuII SmaI BglII and NsiI and probed with various pFKN fragments Here the DNA blot was probed with the avrRpm1 ORF The two DNA samples exhibit a similar pattern except for a polymorphism (eg EcoRI BglII and SmaI) near the site of pFKN integra-tion into the chromosomeB The integration of the plasmid alters the overall fitness of Psm M6integron The growth of Psm M6integron in KB (empty symbols) and MM (filled symbols) was compared with the growth of Psm M6 and Psm M6C Bacteria were inoculated at 104 cfu ml-1 and cultures were titrated at 0 6 12 29 32 54 and 60 h after the inoculation Psm M6pFKN is indicated by squares Psm M6C is indicated by triangles and rhombi indicate Psm M6integron Data points represent the mean plusmn standard error of four samples (some error bars are not visible because too small) Psm M6pFKN and Psm M6C grew equally well in both minimal medium and KB Psm M6integron grows slower than Psm M6pFKN and Psm M6C in both media
pFKN a virulence plasmid in P syringae pv maculicola 1553
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
grown in KB MM and on resistant or susceptibleplants
The genomic context of avrPphE is different in various P syringae strains
We investigated the regions surrounding the avrPphEtransposon on the chromosome of Psm M6 (Fig 5) byTAIL-PCR The left border shows 97 nt identity to the 5centend of the insertion sequence ISPsy3 (AB063176) fromP syringae pv eriobotryae This element is interrupted bythe avrPphE transposon The translation product of theinterrupted ORF found on this border is homologous tothe IS801 transposase (79 identity and 86 homology)
and IS801 has been associated with other type III effec-tors in various P syringae pathovars (Kim et al 1998)On the right border the 200 bp sequence following theterminal repeat of the avrPphE transposon encodes aprotein homologous (81 nucleotide identity) to the 3centend of an unrelated putative transposase gene fromPseudomonas aeruginosa The next 1062 bp sequencedownstream shows high nucleotide homology tosequence identified in P syringae pv pisi (Arnold et al2001) This DNA fragment may be a transposon remnantand carries the avrPpiA1 allele of avrRpm1 from P syrin-gae pv pisi among other genes In the chromosome ofPsm M6 this sequence shows homology to the P s pisigenes ORFD (94 identity) encoding a putative trans-
Fig 4 The pFKN integration site is a second copy of the avrPphE transposonA The region in the Psm M6 chromosome that hybridizes to the avrPphE and TnpA probes was amplified and sequenced The sequence of the chromosomal region is 100 identical to the transposon in the plasmid but contains two additional regions (regions A and C) TnpA encodes a transposase and res a resolvase Based on homology the chromosomal avrPphE transposon could be complete and functional The avrPphE transposon carried on the plas-mid lacks the 5cent end of TnpA and res (region A) and the absence of region C results in the truncation and a frameshift (marked FS) of the gene encoding the transposase The primers used for PCR amplification (Fig 4B) are indi-cated by arrows and their names are in boldB To investigate where regions A and C were distributed after integration of the plasmid into the chromosome PCRs were performed on the DNA of Psm M6integron and Psm M6 using Larp3 and K110 and K110 and K113 (see Fig 4A) The product amplified from Psm M6integron is run in lane I The product ampli-fied from Psm M6 is run in lane F The product amplified from Psm M6C is run in lane C The difference in size observed between the PCR products from Psm M6integron and Psm M6 using Larp3 and K110 indicates that regions A and C are not carried on the same putative transposon in the chromosome of Psm M6integron (in contrast to the chromosome of Psm M6) The PCR products from Psm M6integron and Psm M6 using K113 and K110 show that neither region A nor region C has been lost in the chromosome of Psm M6integron The identical size of the PCR prod-ucts from Psm M6integron and Psm M6 using K113 and Larp2 indicates that region C is on the right border of pFKN when integrated into the chromosome The region A is on the left border as it is carried by the other putative transposonC After integration of pFKN into the chromo-some TnpA on the left border contains region A but not region C TnpA on the right border contains region C The primers used for the PCR amplification are indicated by arrows and their names are in bold (in Fig 4A)
1554 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
posase and ORF3 encoding a putative type III effector(100 identity) Thus this region in Psm M6 is a remnantof the putative transposon found in P s pisi
We detected the avrPphE transposon in other strains ofP syringae pv maculicola and in the sequence of the
genome of P s tomato DC3000 In Psm M2 Psm M8Psm M12 Psm m90 and Psm m10831 the borders arethe same as in Psm M6 (data not shown) On the otherhand they differ in Psm M5 and Psm M9 and these alldiffer from Pst DC3000 In Psm M5 (Fig 5) and Psm M9
Fig 5 Sequences surrounding the avrPphE transposon define four different genetic contexts The figure is a graphical representation of the four genomic environments in which the avrPphE transposon was detected The regions represented were characterized by sequence comparison using the BLASTX algorithm It is not known whether the avrPphE transposon is located in the chromosome or on a plasmid in the genome of Psm M5 and Pst DC3000
pFKN a virulence plasmid in P syringae pv maculicola 1555
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(not shown) the left border consists of the 5cent end of agene sharing 99 identity to the replicase of pPS10 aplasmid found in P syringae pv savastanoi (Nieto et al1992) This replicase is not related to that of pFKN (aPT23A-like plasmid see above) The right bordersequence is a remnant of a gene encoding a putative ABCtransporter (23 identity and 43 homology to a putativesugar uptake ABC transporter ATP-binding protein fromSinorhizobium meliloti)
In the genome of Pst DC3000 we looked at 30 kbsurrounding the transposon (Fig 5) The proteinsencoded on this 30 kb region share homology to eitherDNA mobility functions (transposases plasmid partition-ing proteins replicases) or type III effectors (identical toavrPpiB Cournoyer et al 1995) The region also containsother sequences often found associated with plasmidsand genes involved in pathogenicity such as rulA and rulB(Sesma et al 2000) Sequences directly neighbouringthe transposon on the left side are homologous to a geneencoding the transposase of the IS100 elements fromYersinia pestis Ralstonia solanacearum and Salmonellaenterica On the right-hand side of the avrPphE transpo-son the first homology is to a fragment of a putative ABCtransporter identical to the one identified in Psm M5 Thissuggests that this part of the two genomes is morerecently diverged than either is from Psm M6
No common sequence was identified in the four differ-ent environments surrounding the avrPphE transposonthat would help to identify the integration target of theavrPphE transposon although the proximal right bordersin Psm M5 and Pst DC3000 are nearly identical Thissuggests that the avrPphE transposon and this border-ing sequence were acquired together by these twostrains and that subsequent events at both the distalright flank and on the left border are responsible for thedifferences in genomic context In contrast the insertionsites of the avrPphE transposon in Psm M5 and PsmM6 are completely different In sum the features ofthese integration sites are consistent with the model thatPAIs evolve by repeated insertion and deletion intopre-existing PAIs (Hensel et al 1999 Tauschek et al2002)
Discussion
We sequenced pFKN an ordf40 kb plasmid that contributesto the virulence of Psm M6 The plasmid carries two typeIII effectors on remnants of transposons contiguous toORF8 itself neighbouring an element closely related toIS801 avrRpm1 encodes a known virulence factor andavrPphE a type III effector The expression of ORF8 isinduced in planta and therefore ORF8 may play a role inthe interaction with the host plant The portion of theplasmid carrying avrRpm1 avrPphE and ORF8 may thus
be considered as a PAI Interestingly the G+C content ofthe ORFs carried on this region are very heterogeneousand the three genes are flanked by sequences related tomobile elements and therefore are likely to have differentorigins This is consistent with the idea that the formationof PAIs requires a multistep process (Hensel et al 1999Alfano et al 2000 Tauschek et al 2002) It is also con-sistent with the findings of Kim et al (1998) showing thatmost of the known type III effectors are associated withthis type of sequence pFKN (and therefore the PAI that itcarries) sometimes integrates into and can excise fromthe chromosome as demonstrated by the discovery of thevariant of the strain Psm M6 We identified the region ofthe chromosome into which the integration took placeThis region is a transposon carrying a second copy ofavrPphE
Alleles of the avrPphE gene have been detected inmany P syringae pathovars (Mansfield et al 1994Alfano et al 2000 Fouts et al 2002 Guttman et al2002) suggesting that it provides an important virulencefunction on a variety of host plants Our findings are thefirst to associate avrPphE with a transposon In PsyB728a and in Pph 1302A for example avrPphE residesdirectly next to the hrp locus in a region called thelsquoexchangeable effector locusrsquo or EEL (Alfano et al 2000)and is not associated with a transposase In fact in PsyB728a avrPphE is predicted to be the third gene in anoperon controlled by the hrpK promoter (Alfano et al2000) This striking finding illustrates the dynamism ofPAIs
We determined by complementation assay that the par-tial loss of virulence in the strain cured of pFKN resultsfrom the absence of avrRpm1 The virulence function ofavrRpm1 has already been demonstrated in the strainPsm M2 (Ritter and Dangl 1995) Recently AvrRpm1 hasbeen demonstrated to interact with RIN4 a proteininvolved in the regulation of some of the plant defenceresponses (Mackey et al 2002) Mackey and coworkersproposed that the role of AvrRpm1 is to reduce the levelof plant defence response triggered by the attack of thepathogen This hypothesis might be consistent with thefact that only quantitative differences in growth wereobserved between Psm M6C and Psm M6 No differencein growth was observed between Psm M6C and PsmM6CKO (lacking avrPphE) Similar results were observedwith the avrPphE allele of P s phaseolicola (Stevenset al 1998) This observation could be explained byeither a functional redundancy with another gene in thegenome of the Psm M6 or the fact that the assay isinappropriate to assess the virulence function of this typeIII effector
The sequence and the integration mechanism of pFKNinto the Psm M6 chromosome provide insights on theevolutionary dynamics of the genomes of P syringae
1556 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
pathovars First the analysis of the G+C content of frag-ments of pFKN suggests that they may have differentorigins This hypothesis is further supported by the pres-ence of some defined fragments from pFKN in thegenomes of Pst DC3000 and Psy B728a The fragment 3(see Table 2) encodes one putative transcriptional regu-lator proteins predicted to be located in the bacterialinner or outer membrane by PSORT (inorganic pyrophos-phatase sugar transporter putative protein) and a puta-tive integraserecombinase homologous to ripX fromBacillus subtilis The presence of this latter gene sug-gests that fragment 3 may be the remnant of abacteriophage
The integration and excision mechanism is very proba-bly a homologous recombination in the 15 kb sequenceregion B of TnpA carried on the avrPphE transposon(Fig 6) It is not clear whether the integrationexcisionprocess relies on general recombination or on a site-specific recombination dependent on an integraserecombinase As indicated by the sequence of the twotransposons before and after integration of pFKN nonucleotide sequences were altered at the site ofexchange and not a single nucleotide was lost or gained(this excludes a mechanism of transpositional site-specificrecombination) General recombination requires a largeDNA region whereas the site-specific recombinationrequires as few as 240 bp for the lambda phage for exam-ple (Thompson and Landy 1989) The rate of generalrecombination is unknown for P syringae but has beenestimated at 10-3 in Pseudomonas tolaasii (and whenrecA is inactivated at 10-4 Sinha et al 2000) The rateof excision of pFKN has been estimated between 10-3 and10-4 and would therefore be consistent with a mechanismof general recombination Additionally the region B is
15 kb long enough to ensure general recombinationOlder reports describe the integration of pMMC7501 intothe chromosome of P s pv phaseolicola (Curiale andMills 1982) An imprecise excision of pMMC7501 led tothe formation of eight derivative plasmids all sharing thesame origin of replication (Szabo and Mills 1984a) Thisexcision occurred through recombination events betweensets of repeats in the chromosome exhibiting sequencehomology with IS elements (Szabo and Mills 1984bPoplawsky and Mills 1987)
The integration of a virulence gene into the chromo-some may be a first step towards its stabilization in thegenome Indeed lack of selection can lead to the stochas-tic loss of the plasmid and many steps in the pathogenlife cycle probably do not require the presence of virulencefunctions The loss of a plasmid carrying the type III effec-tor avrPpiB was observed in P syringae pv pisi (Bavageet al 1991) We propose that the integration of pFKNcould lead to the stabilization of avrRpm1 in the genomeof Psm M6 Interestingly a DNA segment of at least 42 kbcarrying avrPphB in P syringae pv phaseolicola canexcise from the chromosome but it is not maintained inthe strain as a plasmid This region may represent anancient plasmid integration event followed by successivegenomic rearrangements deleterious for plasmid mainte-nance (Jackson et al 2000) The fact that we can stillobserve the excision of pFKN from the chromosome ofPsm M6integron shows that the newly generated geneticunit is not rapidly stabilized in the genome and that furtherevents might be necessary to prevent excision Further-more the integration of pFKN into the chromosomeslightly impairs the fitness of Psm M6integron The pres-ence of the functional origin of replication of pFKN mayinterfere with the replication of the chromosome Integra-
Fig 6 The integration of pFKN may have occurred via homologous recombination between the regions B common to the two TnpA genes The TnpA genes are constituted of four DNA sequence regions (1) before the integration the chromosomal copy of TnpA contains all four regions (A B C and D) The TnpA on the plasmid is missing the 5cent end of the gene (region A) and a fragment towards the 3cent end (region C) which results in a frameshift (FS) After the homologous recombination between two identical regions (region B) in the coding region of the TnpA genes (2) the region A is found in the TnpA of the left border and region C in the TnpA of the right border (3)
pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

pFKN a virulence plasmid in P syringae pv maculicola 1553
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
grown in KB MM and on resistant or susceptibleplants
The genomic context of avrPphE is different in various P syringae strains
We investigated the regions surrounding the avrPphEtransposon on the chromosome of Psm M6 (Fig 5) byTAIL-PCR The left border shows 97 nt identity to the 5centend of the insertion sequence ISPsy3 (AB063176) fromP syringae pv eriobotryae This element is interrupted bythe avrPphE transposon The translation product of theinterrupted ORF found on this border is homologous tothe IS801 transposase (79 identity and 86 homology)
and IS801 has been associated with other type III effec-tors in various P syringae pathovars (Kim et al 1998)On the right border the 200 bp sequence following theterminal repeat of the avrPphE transposon encodes aprotein homologous (81 nucleotide identity) to the 3centend of an unrelated putative transposase gene fromPseudomonas aeruginosa The next 1062 bp sequencedownstream shows high nucleotide homology tosequence identified in P syringae pv pisi (Arnold et al2001) This DNA fragment may be a transposon remnantand carries the avrPpiA1 allele of avrRpm1 from P syrin-gae pv pisi among other genes In the chromosome ofPsm M6 this sequence shows homology to the P s pisigenes ORFD (94 identity) encoding a putative trans-
Fig 4 The pFKN integration site is a second copy of the avrPphE transposonA The region in the Psm M6 chromosome that hybridizes to the avrPphE and TnpA probes was amplified and sequenced The sequence of the chromosomal region is 100 identical to the transposon in the plasmid but contains two additional regions (regions A and C) TnpA encodes a transposase and res a resolvase Based on homology the chromosomal avrPphE transposon could be complete and functional The avrPphE transposon carried on the plas-mid lacks the 5cent end of TnpA and res (region A) and the absence of region C results in the truncation and a frameshift (marked FS) of the gene encoding the transposase The primers used for PCR amplification (Fig 4B) are indi-cated by arrows and their names are in boldB To investigate where regions A and C were distributed after integration of the plasmid into the chromosome PCRs were performed on the DNA of Psm M6integron and Psm M6 using Larp3 and K110 and K110 and K113 (see Fig 4A) The product amplified from Psm M6integron is run in lane I The product ampli-fied from Psm M6 is run in lane F The product amplified from Psm M6C is run in lane C The difference in size observed between the PCR products from Psm M6integron and Psm M6 using Larp3 and K110 indicates that regions A and C are not carried on the same putative transposon in the chromosome of Psm M6integron (in contrast to the chromosome of Psm M6) The PCR products from Psm M6integron and Psm M6 using K113 and K110 show that neither region A nor region C has been lost in the chromosome of Psm M6integron The identical size of the PCR prod-ucts from Psm M6integron and Psm M6 using K113 and Larp2 indicates that region C is on the right border of pFKN when integrated into the chromosome The region A is on the left border as it is carried by the other putative transposonC After integration of pFKN into the chromo-some TnpA on the left border contains region A but not region C TnpA on the right border contains region C The primers used for the PCR amplification are indicated by arrows and their names are in bold (in Fig 4A)
1554 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
posase and ORF3 encoding a putative type III effector(100 identity) Thus this region in Psm M6 is a remnantof the putative transposon found in P s pisi
We detected the avrPphE transposon in other strains ofP syringae pv maculicola and in the sequence of the
genome of P s tomato DC3000 In Psm M2 Psm M8Psm M12 Psm m90 and Psm m10831 the borders arethe same as in Psm M6 (data not shown) On the otherhand they differ in Psm M5 and Psm M9 and these alldiffer from Pst DC3000 In Psm M5 (Fig 5) and Psm M9
Fig 5 Sequences surrounding the avrPphE transposon define four different genetic contexts The figure is a graphical representation of the four genomic environments in which the avrPphE transposon was detected The regions represented were characterized by sequence comparison using the BLASTX algorithm It is not known whether the avrPphE transposon is located in the chromosome or on a plasmid in the genome of Psm M5 and Pst DC3000
pFKN a virulence plasmid in P syringae pv maculicola 1555
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(not shown) the left border consists of the 5cent end of agene sharing 99 identity to the replicase of pPS10 aplasmid found in P syringae pv savastanoi (Nieto et al1992) This replicase is not related to that of pFKN (aPT23A-like plasmid see above) The right bordersequence is a remnant of a gene encoding a putative ABCtransporter (23 identity and 43 homology to a putativesugar uptake ABC transporter ATP-binding protein fromSinorhizobium meliloti)
In the genome of Pst DC3000 we looked at 30 kbsurrounding the transposon (Fig 5) The proteinsencoded on this 30 kb region share homology to eitherDNA mobility functions (transposases plasmid partition-ing proteins replicases) or type III effectors (identical toavrPpiB Cournoyer et al 1995) The region also containsother sequences often found associated with plasmidsand genes involved in pathogenicity such as rulA and rulB(Sesma et al 2000) Sequences directly neighbouringthe transposon on the left side are homologous to a geneencoding the transposase of the IS100 elements fromYersinia pestis Ralstonia solanacearum and Salmonellaenterica On the right-hand side of the avrPphE transpo-son the first homology is to a fragment of a putative ABCtransporter identical to the one identified in Psm M5 Thissuggests that this part of the two genomes is morerecently diverged than either is from Psm M6
No common sequence was identified in the four differ-ent environments surrounding the avrPphE transposonthat would help to identify the integration target of theavrPphE transposon although the proximal right bordersin Psm M5 and Pst DC3000 are nearly identical Thissuggests that the avrPphE transposon and this border-ing sequence were acquired together by these twostrains and that subsequent events at both the distalright flank and on the left border are responsible for thedifferences in genomic context In contrast the insertionsites of the avrPphE transposon in Psm M5 and PsmM6 are completely different In sum the features ofthese integration sites are consistent with the model thatPAIs evolve by repeated insertion and deletion intopre-existing PAIs (Hensel et al 1999 Tauschek et al2002)
Discussion
We sequenced pFKN an ordf40 kb plasmid that contributesto the virulence of Psm M6 The plasmid carries two typeIII effectors on remnants of transposons contiguous toORF8 itself neighbouring an element closely related toIS801 avrRpm1 encodes a known virulence factor andavrPphE a type III effector The expression of ORF8 isinduced in planta and therefore ORF8 may play a role inthe interaction with the host plant The portion of theplasmid carrying avrRpm1 avrPphE and ORF8 may thus
be considered as a PAI Interestingly the G+C content ofthe ORFs carried on this region are very heterogeneousand the three genes are flanked by sequences related tomobile elements and therefore are likely to have differentorigins This is consistent with the idea that the formationof PAIs requires a multistep process (Hensel et al 1999Alfano et al 2000 Tauschek et al 2002) It is also con-sistent with the findings of Kim et al (1998) showing thatmost of the known type III effectors are associated withthis type of sequence pFKN (and therefore the PAI that itcarries) sometimes integrates into and can excise fromthe chromosome as demonstrated by the discovery of thevariant of the strain Psm M6 We identified the region ofthe chromosome into which the integration took placeThis region is a transposon carrying a second copy ofavrPphE
Alleles of the avrPphE gene have been detected inmany P syringae pathovars (Mansfield et al 1994Alfano et al 2000 Fouts et al 2002 Guttman et al2002) suggesting that it provides an important virulencefunction on a variety of host plants Our findings are thefirst to associate avrPphE with a transposon In PsyB728a and in Pph 1302A for example avrPphE residesdirectly next to the hrp locus in a region called thelsquoexchangeable effector locusrsquo or EEL (Alfano et al 2000)and is not associated with a transposase In fact in PsyB728a avrPphE is predicted to be the third gene in anoperon controlled by the hrpK promoter (Alfano et al2000) This striking finding illustrates the dynamism ofPAIs
We determined by complementation assay that the par-tial loss of virulence in the strain cured of pFKN resultsfrom the absence of avrRpm1 The virulence function ofavrRpm1 has already been demonstrated in the strainPsm M2 (Ritter and Dangl 1995) Recently AvrRpm1 hasbeen demonstrated to interact with RIN4 a proteininvolved in the regulation of some of the plant defenceresponses (Mackey et al 2002) Mackey and coworkersproposed that the role of AvrRpm1 is to reduce the levelof plant defence response triggered by the attack of thepathogen This hypothesis might be consistent with thefact that only quantitative differences in growth wereobserved between Psm M6C and Psm M6 No differencein growth was observed between Psm M6C and PsmM6CKO (lacking avrPphE) Similar results were observedwith the avrPphE allele of P s phaseolicola (Stevenset al 1998) This observation could be explained byeither a functional redundancy with another gene in thegenome of the Psm M6 or the fact that the assay isinappropriate to assess the virulence function of this typeIII effector
The sequence and the integration mechanism of pFKNinto the Psm M6 chromosome provide insights on theevolutionary dynamics of the genomes of P syringae
1556 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
pathovars First the analysis of the G+C content of frag-ments of pFKN suggests that they may have differentorigins This hypothesis is further supported by the pres-ence of some defined fragments from pFKN in thegenomes of Pst DC3000 and Psy B728a The fragment 3(see Table 2) encodes one putative transcriptional regu-lator proteins predicted to be located in the bacterialinner or outer membrane by PSORT (inorganic pyrophos-phatase sugar transporter putative protein) and a puta-tive integraserecombinase homologous to ripX fromBacillus subtilis The presence of this latter gene sug-gests that fragment 3 may be the remnant of abacteriophage
The integration and excision mechanism is very proba-bly a homologous recombination in the 15 kb sequenceregion B of TnpA carried on the avrPphE transposon(Fig 6) It is not clear whether the integrationexcisionprocess relies on general recombination or on a site-specific recombination dependent on an integraserecombinase As indicated by the sequence of the twotransposons before and after integration of pFKN nonucleotide sequences were altered at the site ofexchange and not a single nucleotide was lost or gained(this excludes a mechanism of transpositional site-specificrecombination) General recombination requires a largeDNA region whereas the site-specific recombinationrequires as few as 240 bp for the lambda phage for exam-ple (Thompson and Landy 1989) The rate of generalrecombination is unknown for P syringae but has beenestimated at 10-3 in Pseudomonas tolaasii (and whenrecA is inactivated at 10-4 Sinha et al 2000) The rateof excision of pFKN has been estimated between 10-3 and10-4 and would therefore be consistent with a mechanismof general recombination Additionally the region B is
15 kb long enough to ensure general recombinationOlder reports describe the integration of pMMC7501 intothe chromosome of P s pv phaseolicola (Curiale andMills 1982) An imprecise excision of pMMC7501 led tothe formation of eight derivative plasmids all sharing thesame origin of replication (Szabo and Mills 1984a) Thisexcision occurred through recombination events betweensets of repeats in the chromosome exhibiting sequencehomology with IS elements (Szabo and Mills 1984bPoplawsky and Mills 1987)
The integration of a virulence gene into the chromo-some may be a first step towards its stabilization in thegenome Indeed lack of selection can lead to the stochas-tic loss of the plasmid and many steps in the pathogenlife cycle probably do not require the presence of virulencefunctions The loss of a plasmid carrying the type III effec-tor avrPpiB was observed in P syringae pv pisi (Bavageet al 1991) We propose that the integration of pFKNcould lead to the stabilization of avrRpm1 in the genomeof Psm M6 Interestingly a DNA segment of at least 42 kbcarrying avrPphB in P syringae pv phaseolicola canexcise from the chromosome but it is not maintained inthe strain as a plasmid This region may represent anancient plasmid integration event followed by successivegenomic rearrangements deleterious for plasmid mainte-nance (Jackson et al 2000) The fact that we can stillobserve the excision of pFKN from the chromosome ofPsm M6integron shows that the newly generated geneticunit is not rapidly stabilized in the genome and that furtherevents might be necessary to prevent excision Further-more the integration of pFKN into the chromosomeslightly impairs the fitness of Psm M6integron The pres-ence of the functional origin of replication of pFKN mayinterfere with the replication of the chromosome Integra-
Fig 6 The integration of pFKN may have occurred via homologous recombination between the regions B common to the two TnpA genes The TnpA genes are constituted of four DNA sequence regions (1) before the integration the chromosomal copy of TnpA contains all four regions (A B C and D) The TnpA on the plasmid is missing the 5cent end of the gene (region A) and a fragment towards the 3cent end (region C) which results in a frameshift (FS) After the homologous recombination between two identical regions (region B) in the coding region of the TnpA genes (2) the region A is found in the TnpA of the left border and region C in the TnpA of the right border (3)
pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

1554 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
posase and ORF3 encoding a putative type III effector(100 identity) Thus this region in Psm M6 is a remnantof the putative transposon found in P s pisi
We detected the avrPphE transposon in other strains ofP syringae pv maculicola and in the sequence of the
genome of P s tomato DC3000 In Psm M2 Psm M8Psm M12 Psm m90 and Psm m10831 the borders arethe same as in Psm M6 (data not shown) On the otherhand they differ in Psm M5 and Psm M9 and these alldiffer from Pst DC3000 In Psm M5 (Fig 5) and Psm M9
Fig 5 Sequences surrounding the avrPphE transposon define four different genetic contexts The figure is a graphical representation of the four genomic environments in which the avrPphE transposon was detected The regions represented were characterized by sequence comparison using the BLASTX algorithm It is not known whether the avrPphE transposon is located in the chromosome or on a plasmid in the genome of Psm M5 and Pst DC3000
pFKN a virulence plasmid in P syringae pv maculicola 1555
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(not shown) the left border consists of the 5cent end of agene sharing 99 identity to the replicase of pPS10 aplasmid found in P syringae pv savastanoi (Nieto et al1992) This replicase is not related to that of pFKN (aPT23A-like plasmid see above) The right bordersequence is a remnant of a gene encoding a putative ABCtransporter (23 identity and 43 homology to a putativesugar uptake ABC transporter ATP-binding protein fromSinorhizobium meliloti)
In the genome of Pst DC3000 we looked at 30 kbsurrounding the transposon (Fig 5) The proteinsencoded on this 30 kb region share homology to eitherDNA mobility functions (transposases plasmid partition-ing proteins replicases) or type III effectors (identical toavrPpiB Cournoyer et al 1995) The region also containsother sequences often found associated with plasmidsand genes involved in pathogenicity such as rulA and rulB(Sesma et al 2000) Sequences directly neighbouringthe transposon on the left side are homologous to a geneencoding the transposase of the IS100 elements fromYersinia pestis Ralstonia solanacearum and Salmonellaenterica On the right-hand side of the avrPphE transpo-son the first homology is to a fragment of a putative ABCtransporter identical to the one identified in Psm M5 Thissuggests that this part of the two genomes is morerecently diverged than either is from Psm M6
No common sequence was identified in the four differ-ent environments surrounding the avrPphE transposonthat would help to identify the integration target of theavrPphE transposon although the proximal right bordersin Psm M5 and Pst DC3000 are nearly identical Thissuggests that the avrPphE transposon and this border-ing sequence were acquired together by these twostrains and that subsequent events at both the distalright flank and on the left border are responsible for thedifferences in genomic context In contrast the insertionsites of the avrPphE transposon in Psm M5 and PsmM6 are completely different In sum the features ofthese integration sites are consistent with the model thatPAIs evolve by repeated insertion and deletion intopre-existing PAIs (Hensel et al 1999 Tauschek et al2002)
Discussion
We sequenced pFKN an ordf40 kb plasmid that contributesto the virulence of Psm M6 The plasmid carries two typeIII effectors on remnants of transposons contiguous toORF8 itself neighbouring an element closely related toIS801 avrRpm1 encodes a known virulence factor andavrPphE a type III effector The expression of ORF8 isinduced in planta and therefore ORF8 may play a role inthe interaction with the host plant The portion of theplasmid carrying avrRpm1 avrPphE and ORF8 may thus
be considered as a PAI Interestingly the G+C content ofthe ORFs carried on this region are very heterogeneousand the three genes are flanked by sequences related tomobile elements and therefore are likely to have differentorigins This is consistent with the idea that the formationof PAIs requires a multistep process (Hensel et al 1999Alfano et al 2000 Tauschek et al 2002) It is also con-sistent with the findings of Kim et al (1998) showing thatmost of the known type III effectors are associated withthis type of sequence pFKN (and therefore the PAI that itcarries) sometimes integrates into and can excise fromthe chromosome as demonstrated by the discovery of thevariant of the strain Psm M6 We identified the region ofthe chromosome into which the integration took placeThis region is a transposon carrying a second copy ofavrPphE
Alleles of the avrPphE gene have been detected inmany P syringae pathovars (Mansfield et al 1994Alfano et al 2000 Fouts et al 2002 Guttman et al2002) suggesting that it provides an important virulencefunction on a variety of host plants Our findings are thefirst to associate avrPphE with a transposon In PsyB728a and in Pph 1302A for example avrPphE residesdirectly next to the hrp locus in a region called thelsquoexchangeable effector locusrsquo or EEL (Alfano et al 2000)and is not associated with a transposase In fact in PsyB728a avrPphE is predicted to be the third gene in anoperon controlled by the hrpK promoter (Alfano et al2000) This striking finding illustrates the dynamism ofPAIs
We determined by complementation assay that the par-tial loss of virulence in the strain cured of pFKN resultsfrom the absence of avrRpm1 The virulence function ofavrRpm1 has already been demonstrated in the strainPsm M2 (Ritter and Dangl 1995) Recently AvrRpm1 hasbeen demonstrated to interact with RIN4 a proteininvolved in the regulation of some of the plant defenceresponses (Mackey et al 2002) Mackey and coworkersproposed that the role of AvrRpm1 is to reduce the levelof plant defence response triggered by the attack of thepathogen This hypothesis might be consistent with thefact that only quantitative differences in growth wereobserved between Psm M6C and Psm M6 No differencein growth was observed between Psm M6C and PsmM6CKO (lacking avrPphE) Similar results were observedwith the avrPphE allele of P s phaseolicola (Stevenset al 1998) This observation could be explained byeither a functional redundancy with another gene in thegenome of the Psm M6 or the fact that the assay isinappropriate to assess the virulence function of this typeIII effector
The sequence and the integration mechanism of pFKNinto the Psm M6 chromosome provide insights on theevolutionary dynamics of the genomes of P syringae
1556 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
pathovars First the analysis of the G+C content of frag-ments of pFKN suggests that they may have differentorigins This hypothesis is further supported by the pres-ence of some defined fragments from pFKN in thegenomes of Pst DC3000 and Psy B728a The fragment 3(see Table 2) encodes one putative transcriptional regu-lator proteins predicted to be located in the bacterialinner or outer membrane by PSORT (inorganic pyrophos-phatase sugar transporter putative protein) and a puta-tive integraserecombinase homologous to ripX fromBacillus subtilis The presence of this latter gene sug-gests that fragment 3 may be the remnant of abacteriophage
The integration and excision mechanism is very proba-bly a homologous recombination in the 15 kb sequenceregion B of TnpA carried on the avrPphE transposon(Fig 6) It is not clear whether the integrationexcisionprocess relies on general recombination or on a site-specific recombination dependent on an integraserecombinase As indicated by the sequence of the twotransposons before and after integration of pFKN nonucleotide sequences were altered at the site ofexchange and not a single nucleotide was lost or gained(this excludes a mechanism of transpositional site-specificrecombination) General recombination requires a largeDNA region whereas the site-specific recombinationrequires as few as 240 bp for the lambda phage for exam-ple (Thompson and Landy 1989) The rate of generalrecombination is unknown for P syringae but has beenestimated at 10-3 in Pseudomonas tolaasii (and whenrecA is inactivated at 10-4 Sinha et al 2000) The rateof excision of pFKN has been estimated between 10-3 and10-4 and would therefore be consistent with a mechanismof general recombination Additionally the region B is
15 kb long enough to ensure general recombinationOlder reports describe the integration of pMMC7501 intothe chromosome of P s pv phaseolicola (Curiale andMills 1982) An imprecise excision of pMMC7501 led tothe formation of eight derivative plasmids all sharing thesame origin of replication (Szabo and Mills 1984a) Thisexcision occurred through recombination events betweensets of repeats in the chromosome exhibiting sequencehomology with IS elements (Szabo and Mills 1984bPoplawsky and Mills 1987)
The integration of a virulence gene into the chromo-some may be a first step towards its stabilization in thegenome Indeed lack of selection can lead to the stochas-tic loss of the plasmid and many steps in the pathogenlife cycle probably do not require the presence of virulencefunctions The loss of a plasmid carrying the type III effec-tor avrPpiB was observed in P syringae pv pisi (Bavageet al 1991) We propose that the integration of pFKNcould lead to the stabilization of avrRpm1 in the genomeof Psm M6 Interestingly a DNA segment of at least 42 kbcarrying avrPphB in P syringae pv phaseolicola canexcise from the chromosome but it is not maintained inthe strain as a plasmid This region may represent anancient plasmid integration event followed by successivegenomic rearrangements deleterious for plasmid mainte-nance (Jackson et al 2000) The fact that we can stillobserve the excision of pFKN from the chromosome ofPsm M6integron shows that the newly generated geneticunit is not rapidly stabilized in the genome and that furtherevents might be necessary to prevent excision Further-more the integration of pFKN into the chromosomeslightly impairs the fitness of Psm M6integron The pres-ence of the functional origin of replication of pFKN mayinterfere with the replication of the chromosome Integra-
Fig 6 The integration of pFKN may have occurred via homologous recombination between the regions B common to the two TnpA genes The TnpA genes are constituted of four DNA sequence regions (1) before the integration the chromosomal copy of TnpA contains all four regions (A B C and D) The TnpA on the plasmid is missing the 5cent end of the gene (region A) and a fragment towards the 3cent end (region C) which results in a frameshift (FS) After the homologous recombination between two identical regions (region B) in the coding region of the TnpA genes (2) the region A is found in the TnpA of the left border and region C in the TnpA of the right border (3)
pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

pFKN a virulence plasmid in P syringae pv maculicola 1555
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(not shown) the left border consists of the 5cent end of agene sharing 99 identity to the replicase of pPS10 aplasmid found in P syringae pv savastanoi (Nieto et al1992) This replicase is not related to that of pFKN (aPT23A-like plasmid see above) The right bordersequence is a remnant of a gene encoding a putative ABCtransporter (23 identity and 43 homology to a putativesugar uptake ABC transporter ATP-binding protein fromSinorhizobium meliloti)
In the genome of Pst DC3000 we looked at 30 kbsurrounding the transposon (Fig 5) The proteinsencoded on this 30 kb region share homology to eitherDNA mobility functions (transposases plasmid partition-ing proteins replicases) or type III effectors (identical toavrPpiB Cournoyer et al 1995) The region also containsother sequences often found associated with plasmidsand genes involved in pathogenicity such as rulA and rulB(Sesma et al 2000) Sequences directly neighbouringthe transposon on the left side are homologous to a geneencoding the transposase of the IS100 elements fromYersinia pestis Ralstonia solanacearum and Salmonellaenterica On the right-hand side of the avrPphE transpo-son the first homology is to a fragment of a putative ABCtransporter identical to the one identified in Psm M5 Thissuggests that this part of the two genomes is morerecently diverged than either is from Psm M6
No common sequence was identified in the four differ-ent environments surrounding the avrPphE transposonthat would help to identify the integration target of theavrPphE transposon although the proximal right bordersin Psm M5 and Pst DC3000 are nearly identical Thissuggests that the avrPphE transposon and this border-ing sequence were acquired together by these twostrains and that subsequent events at both the distalright flank and on the left border are responsible for thedifferences in genomic context In contrast the insertionsites of the avrPphE transposon in Psm M5 and PsmM6 are completely different In sum the features ofthese integration sites are consistent with the model thatPAIs evolve by repeated insertion and deletion intopre-existing PAIs (Hensel et al 1999 Tauschek et al2002)
Discussion
We sequenced pFKN an ordf40 kb plasmid that contributesto the virulence of Psm M6 The plasmid carries two typeIII effectors on remnants of transposons contiguous toORF8 itself neighbouring an element closely related toIS801 avrRpm1 encodes a known virulence factor andavrPphE a type III effector The expression of ORF8 isinduced in planta and therefore ORF8 may play a role inthe interaction with the host plant The portion of theplasmid carrying avrRpm1 avrPphE and ORF8 may thus
be considered as a PAI Interestingly the G+C content ofthe ORFs carried on this region are very heterogeneousand the three genes are flanked by sequences related tomobile elements and therefore are likely to have differentorigins This is consistent with the idea that the formationof PAIs requires a multistep process (Hensel et al 1999Alfano et al 2000 Tauschek et al 2002) It is also con-sistent with the findings of Kim et al (1998) showing thatmost of the known type III effectors are associated withthis type of sequence pFKN (and therefore the PAI that itcarries) sometimes integrates into and can excise fromthe chromosome as demonstrated by the discovery of thevariant of the strain Psm M6 We identified the region ofthe chromosome into which the integration took placeThis region is a transposon carrying a second copy ofavrPphE
Alleles of the avrPphE gene have been detected inmany P syringae pathovars (Mansfield et al 1994Alfano et al 2000 Fouts et al 2002 Guttman et al2002) suggesting that it provides an important virulencefunction on a variety of host plants Our findings are thefirst to associate avrPphE with a transposon In PsyB728a and in Pph 1302A for example avrPphE residesdirectly next to the hrp locus in a region called thelsquoexchangeable effector locusrsquo or EEL (Alfano et al 2000)and is not associated with a transposase In fact in PsyB728a avrPphE is predicted to be the third gene in anoperon controlled by the hrpK promoter (Alfano et al2000) This striking finding illustrates the dynamism ofPAIs
We determined by complementation assay that the par-tial loss of virulence in the strain cured of pFKN resultsfrom the absence of avrRpm1 The virulence function ofavrRpm1 has already been demonstrated in the strainPsm M2 (Ritter and Dangl 1995) Recently AvrRpm1 hasbeen demonstrated to interact with RIN4 a proteininvolved in the regulation of some of the plant defenceresponses (Mackey et al 2002) Mackey and coworkersproposed that the role of AvrRpm1 is to reduce the levelof plant defence response triggered by the attack of thepathogen This hypothesis might be consistent with thefact that only quantitative differences in growth wereobserved between Psm M6C and Psm M6 No differencein growth was observed between Psm M6C and PsmM6CKO (lacking avrPphE) Similar results were observedwith the avrPphE allele of P s phaseolicola (Stevenset al 1998) This observation could be explained byeither a functional redundancy with another gene in thegenome of the Psm M6 or the fact that the assay isinappropriate to assess the virulence function of this typeIII effector
The sequence and the integration mechanism of pFKNinto the Psm M6 chromosome provide insights on theevolutionary dynamics of the genomes of P syringae
1556 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
pathovars First the analysis of the G+C content of frag-ments of pFKN suggests that they may have differentorigins This hypothesis is further supported by the pres-ence of some defined fragments from pFKN in thegenomes of Pst DC3000 and Psy B728a The fragment 3(see Table 2) encodes one putative transcriptional regu-lator proteins predicted to be located in the bacterialinner or outer membrane by PSORT (inorganic pyrophos-phatase sugar transporter putative protein) and a puta-tive integraserecombinase homologous to ripX fromBacillus subtilis The presence of this latter gene sug-gests that fragment 3 may be the remnant of abacteriophage
The integration and excision mechanism is very proba-bly a homologous recombination in the 15 kb sequenceregion B of TnpA carried on the avrPphE transposon(Fig 6) It is not clear whether the integrationexcisionprocess relies on general recombination or on a site-specific recombination dependent on an integraserecombinase As indicated by the sequence of the twotransposons before and after integration of pFKN nonucleotide sequences were altered at the site ofexchange and not a single nucleotide was lost or gained(this excludes a mechanism of transpositional site-specificrecombination) General recombination requires a largeDNA region whereas the site-specific recombinationrequires as few as 240 bp for the lambda phage for exam-ple (Thompson and Landy 1989) The rate of generalrecombination is unknown for P syringae but has beenestimated at 10-3 in Pseudomonas tolaasii (and whenrecA is inactivated at 10-4 Sinha et al 2000) The rateof excision of pFKN has been estimated between 10-3 and10-4 and would therefore be consistent with a mechanismof general recombination Additionally the region B is
15 kb long enough to ensure general recombinationOlder reports describe the integration of pMMC7501 intothe chromosome of P s pv phaseolicola (Curiale andMills 1982) An imprecise excision of pMMC7501 led tothe formation of eight derivative plasmids all sharing thesame origin of replication (Szabo and Mills 1984a) Thisexcision occurred through recombination events betweensets of repeats in the chromosome exhibiting sequencehomology with IS elements (Szabo and Mills 1984bPoplawsky and Mills 1987)
The integration of a virulence gene into the chromo-some may be a first step towards its stabilization in thegenome Indeed lack of selection can lead to the stochas-tic loss of the plasmid and many steps in the pathogenlife cycle probably do not require the presence of virulencefunctions The loss of a plasmid carrying the type III effec-tor avrPpiB was observed in P syringae pv pisi (Bavageet al 1991) We propose that the integration of pFKNcould lead to the stabilization of avrRpm1 in the genomeof Psm M6 Interestingly a DNA segment of at least 42 kbcarrying avrPphB in P syringae pv phaseolicola canexcise from the chromosome but it is not maintained inthe strain as a plasmid This region may represent anancient plasmid integration event followed by successivegenomic rearrangements deleterious for plasmid mainte-nance (Jackson et al 2000) The fact that we can stillobserve the excision of pFKN from the chromosome ofPsm M6integron shows that the newly generated geneticunit is not rapidly stabilized in the genome and that furtherevents might be necessary to prevent excision Further-more the integration of pFKN into the chromosomeslightly impairs the fitness of Psm M6integron The pres-ence of the functional origin of replication of pFKN mayinterfere with the replication of the chromosome Integra-
Fig 6 The integration of pFKN may have occurred via homologous recombination between the regions B common to the two TnpA genes The TnpA genes are constituted of four DNA sequence regions (1) before the integration the chromosomal copy of TnpA contains all four regions (A B C and D) The TnpA on the plasmid is missing the 5cent end of the gene (region A) and a fragment towards the 3cent end (region C) which results in a frameshift (FS) After the homologous recombination between two identical regions (region B) in the coding region of the TnpA genes (2) the region A is found in the TnpA of the left border and region C in the TnpA of the right border (3)
pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

1556 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
pathovars First the analysis of the G+C content of frag-ments of pFKN suggests that they may have differentorigins This hypothesis is further supported by the pres-ence of some defined fragments from pFKN in thegenomes of Pst DC3000 and Psy B728a The fragment 3(see Table 2) encodes one putative transcriptional regu-lator proteins predicted to be located in the bacterialinner or outer membrane by PSORT (inorganic pyrophos-phatase sugar transporter putative protein) and a puta-tive integraserecombinase homologous to ripX fromBacillus subtilis The presence of this latter gene sug-gests that fragment 3 may be the remnant of abacteriophage
The integration and excision mechanism is very proba-bly a homologous recombination in the 15 kb sequenceregion B of TnpA carried on the avrPphE transposon(Fig 6) It is not clear whether the integrationexcisionprocess relies on general recombination or on a site-specific recombination dependent on an integraserecombinase As indicated by the sequence of the twotransposons before and after integration of pFKN nonucleotide sequences were altered at the site ofexchange and not a single nucleotide was lost or gained(this excludes a mechanism of transpositional site-specificrecombination) General recombination requires a largeDNA region whereas the site-specific recombinationrequires as few as 240 bp for the lambda phage for exam-ple (Thompson and Landy 1989) The rate of generalrecombination is unknown for P syringae but has beenestimated at 10-3 in Pseudomonas tolaasii (and whenrecA is inactivated at 10-4 Sinha et al 2000) The rateof excision of pFKN has been estimated between 10-3 and10-4 and would therefore be consistent with a mechanismof general recombination Additionally the region B is
15 kb long enough to ensure general recombinationOlder reports describe the integration of pMMC7501 intothe chromosome of P s pv phaseolicola (Curiale andMills 1982) An imprecise excision of pMMC7501 led tothe formation of eight derivative plasmids all sharing thesame origin of replication (Szabo and Mills 1984a) Thisexcision occurred through recombination events betweensets of repeats in the chromosome exhibiting sequencehomology with IS elements (Szabo and Mills 1984bPoplawsky and Mills 1987)
The integration of a virulence gene into the chromo-some may be a first step towards its stabilization in thegenome Indeed lack of selection can lead to the stochas-tic loss of the plasmid and many steps in the pathogenlife cycle probably do not require the presence of virulencefunctions The loss of a plasmid carrying the type III effec-tor avrPpiB was observed in P syringae pv pisi (Bavageet al 1991) We propose that the integration of pFKNcould lead to the stabilization of avrRpm1 in the genomeof Psm M6 Interestingly a DNA segment of at least 42 kbcarrying avrPphB in P syringae pv phaseolicola canexcise from the chromosome but it is not maintained inthe strain as a plasmid This region may represent anancient plasmid integration event followed by successivegenomic rearrangements deleterious for plasmid mainte-nance (Jackson et al 2000) The fact that we can stillobserve the excision of pFKN from the chromosome ofPsm M6integron shows that the newly generated geneticunit is not rapidly stabilized in the genome and that furtherevents might be necessary to prevent excision Further-more the integration of pFKN into the chromosomeslightly impairs the fitness of Psm M6integron The pres-ence of the functional origin of replication of pFKN mayinterfere with the replication of the chromosome Integra-
Fig 6 The integration of pFKN may have occurred via homologous recombination between the regions B common to the two TnpA genes The TnpA genes are constituted of four DNA sequence regions (1) before the integration the chromosomal copy of TnpA contains all four regions (A B C and D) The TnpA on the plasmid is missing the 5cent end of the gene (region A) and a fragment towards the 3cent end (region C) which results in a frameshift (FS) After the homologous recombination between two identical regions (region B) in the coding region of the TnpA genes (2) the region A is found in the TnpA of the left border and region C in the TnpA of the right border (3)
pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

pFKN a virulence plasmid in P syringae pv maculicola 1557
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
tion of multicopy plasmids is commonly lethal to E coliunless the replication origin of the integrated element issuppressed (Yamaguchi and Tomizawa 1980 Froehlichet al 1983)
pFKN is not the only genetic unit potentially stabilizedby integration The chromosomal avrPphE transposonthat acts as the insertion site for pFKN may be activebefore integration The chromosomal TnpA has a full ORFand we observed transcription in both KB and MM usingRT-PCR (data not shown) The pFKN integration eventdisrupts the chromosomal avrPphE transposon thus pre-venting autonomous transposition As transposition canbe detrimental the stabilization of the avrPphE transpo-son could be advantageous for the bacterium
Strikingly pFKN integrates into a region of the Psm M6chromosome already encoding two type III effectors (avr-PphE and ORF3 identified in Ppi) The avrPphE transpo-son has been detected at four different loci (chromosomaland on pFKN in Psm M6 one in Psm M5 and one in PstDC3000) In all cases genes related to DNA mobilitywere identified in the vicinity In at least three out of fourcases (Psm M6 chromosome pFKN and Pst DC3000)additional genes encoding type III effectors were alsofound in the vicinity of the avrPphE transposon (Fig 5)The sequence available for PsmM5 is the shortest andwe detected no homology to virulence genes in the flank-ing sequence However part of the sequence on the rightborder is 100 identical to the immediate right borderidentified in Pst DC3000 It is likely that the largergenomic context of the avrPphE transposon in bothstrains has a common ancestor (or was transferred fromone strain to the other) Finally the transposon in PstDC3000 and Psm M5 is 100 identical to the transposonin Psm M6 This suggests that the acquisition of the trans-poson was rather recent As a comparison avrRpm1 itsallele avrPpiA1 and their surrounding sequences are 97identical in nucleotide sequence The rather recent acqui-sition of avrPphE and its association with other genesrelated to pathogenicity and DNA mobility may suggestthat the avrPphE transposon is part of a PAI in formationThe integration of pFKN into this site would be the nextstep in the process The proposed mechanism of integra-tion of pFKN suggests that this plasmid could integrate inthe chromosome of any strain that carries the avrPphEtransposon (provided that the strain acquires the plas-mid) Transfer of genes related to pathogenicity or adap-tation to various conditions may be facilitated or evenexplained by the integration of plasmids in the bacterialchromosome followed by the imprecise excision of plas-mids Excised plasmids would contain all or part of theintegrated plasmid and regions from the chromosomeThe transfer of these plasmids could explain how differentstrains share nearly identical regions in their respectivegenomes
Experimental procedures
Bacterial strains and culture conditions
The bacterial strains used in this study are shown in Table 3and the recombinant plasmids in Table 4 The Pseudomonasstrains were grown in Kingrsquos B media (KB) (King et al 1954)shaken at 28infinC or grown on KB agar plates at 28infinC The Ecoli strains were grown in 2yen YT broth or on LuriandashBertani(LB) broth agar plates at 37infinC (Maniatis et al 1989) ForPseudomonas strains antibiotics were used at the followingconcentrations (mg l-1) rifampicin 25ndash50 kanamycin 50carbenicillin 100 spectinomycin 10 Plates contained cyclo-heximide (50 mg l-1) For E coli strains antibiotics were usedas follows (mg l-1) ampicillin 100 nalidixic acid 10 kana-mycin 100 spectinomycin 100
DNA manipulations
Plasmid DNA isolations were performed by alkaline lysisextraction as described by Maniatis et al (1989) GenomicDNA isolations were performed as described by Syn andSwarup (2000) All molecular manipulations were done viastandard procedures Enzymes were purchased from NewEngland BioLabs or Boehringer Mannheim and used accord-ing to the manufacturerrsquos specifications For Southern analy-sis ordf2 mg of DNA was loaded per lane on agarose gel DNAwas transferred to Hybond N+ membranes (Amersham Phar-macia Biotech) and hybridization was performed in hybrid-ization solution [15yen SSC 7 SDS 10 polyethylene glycol(PEG) molecular weight 8000 100 mg ml-1 sonicated anddenatured herring sperm DNA 250 mg ml-1 heparin] at 65infinCThe DNA probes were labelled with a-ATP using reagentsprovided in the Prime-It II random primer labelling kit(Stratagene)
Growth curves and inoculations
To assess the growth of bacteria in planta bacteria wereinoculated at a density of 105 cfu ml-1 in 10 mM MgCl2 on fourleaves per plant and four leaf discs from individual plantswere collected at day 0 (to assess the original inoculum) day1 day 3 and day 5 The discs were ground in 1 ml of 10 mMMgCl2 and serial dilutions were plated on KB plates to titratethe bacteria Four technical repetitions were performed perexperiment (Ritter and Dangl 1995)
Shotgun cloning and DNA sequencing
Purified pFKN DNA (225 mg) was partially digested withTsp5091 Fragments ranging in size from 2 to 8 kb wereisolated from an agarose gel with a QIAquick gel isolation kit(Qiagen) The extracted fragments were ligated into EcoRI-digested pUC18 (Pharmacia) The ligation was transformedinto electrocompetent E coli strain DH5a (Electro CellManipulatorR 600 from BTX 50 mF 129 W 25 kV) Thismethod has been described by Ansorge et al (1997) Theinserts of 150 randomly picked clones were sequenced usingABI dye terminator chemistry by the UNC-CH AutomatedDNA sequencing facility with M13F (TTCAGGGAGCCTGCG
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143
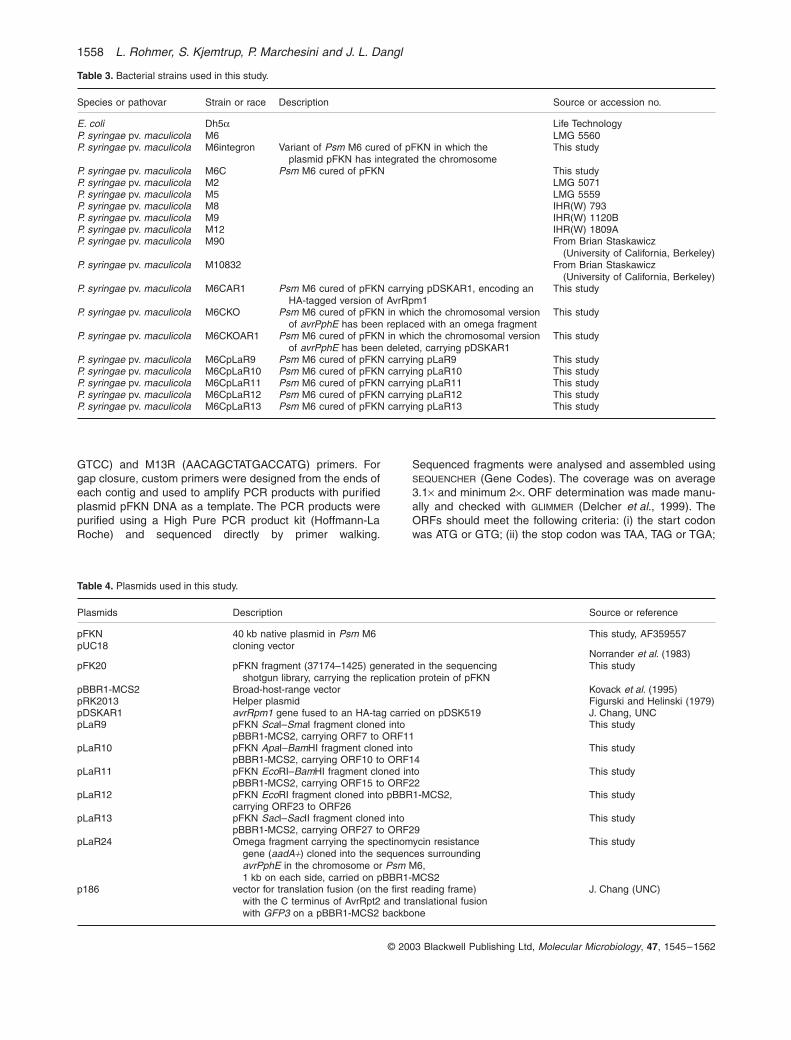
1558 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
GTCC) and M13R (AACAGCTATGACCATG) primers Forgap closure custom primers were designed from the ends ofeach contig and used to amplify PCR products with purifiedplasmid pFKN DNA as a template The PCR products werepurified using a High Pure PCR product kit (Hoffmann-LaRoche) and sequenced directly by primer walking
Sequenced fragments were analysed and assembled usingSEQUENCHER (Gene Codes) The coverage was on average31yen and minimum 2yen ORF determination was made manu-ally and checked with GLIMMER (Delcher et al 1999) TheORFs should meet the following criteria (i) the start codonwas ATG or GTG (ii) the stop codon was TAA TAG or TGA
Table 3 Bacterial strains used in this study
Species or pathovar Strain or race Description Source or accession no
E coli Dh5a Life TechnologyP syringae pv maculicola M6 LMG 5560P syringae pv maculicola M6integron Variant of Psm M6 cured of pFKN in which the
plasmid pFKN has integrated the chromosomeThis study
P syringae pv maculicola M6C Psm M6 cured of pFKN This studyP syringae pv maculicola M2 LMG 5071P syringae pv maculicola M5 LMG 5559P syringae pv maculicola M8 IHR(W) 793P syringae pv maculicola M9 IHR(W) 1120BP syringae pv maculicola M12 IHR(W) 1809AP syringae pv maculicola M90 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M10832 From Brian Staskawicz
(University of California Berkeley)P syringae pv maculicola M6CAR1 Psm M6 cured of pFKN carrying pDSKAR1 encoding an
HA-tagged version of AvrRpm1This study
P syringae pv maculicola M6CKO Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been replaced with an omega fragment
This study
P syringae pv maculicola M6CKOAR1 Psm M6 cured of pFKN in which the chromosomal versionof avrPphE has been deleted carrying pDSKAR1
This study
P syringae pv maculicola M6CpLaR9 Psm M6 cured of pFKN carrying pLaR9 This studyP syringae pv maculicola M6CpLaR10 Psm M6 cured of pFKN carrying pLaR10 This studyP syringae pv maculicola M6CpLaR11 Psm M6 cured of pFKN carrying pLaR11 This studyP syringae pv maculicola M6CpLaR12 Psm M6 cured of pFKN carrying pLaR12 This studyP syringae pv maculicola M6CpLaR13 Psm M6 cured of pFKN carrying pLaR13 This study
Table 4 Plasmids used in this study
Plasmids Description Source or reference
pFKN 40 kb native plasmid in Psm M6 This study AF359557pUC18 cloning vector
Norrander et al (1983)pFK20 pFKN fragment (37174ndash1425) generated in the sequencing
shotgun library carrying the replication protein of pFKNThis study
pBBR1-MCS2 Broad-host-range vector Kovack et al (1995)pRK2013 Helper plasmid Figurski and Helinski (1979)pDSKAR1 avrRpm1 gene fused to an HA-tag carried on pDSK519 J Chang UNCpLaR9 pFKN ScaIndashSmaI fragment cloned into This study
pBBR1-MCS2 carrying ORF7 to ORF11pLaR10 pFKN ApaIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF10 to ORF14pLaR11 pFKN EcoRIndashBamHI fragment cloned into This study
pBBR1-MCS2 carrying ORF15 to ORF22pLaR12 pFKN EcoRI fragment cloned into pBBR1-MCS2 This study
carrying ORF23 to ORF26pLaR13 pFKN SacIndashSacII fragment cloned into This study
pBBR1-MCS2 carrying ORF27 to ORF29pLaR24 Omega fragment carrying the spectinomycin resistance
gene (aadA+) cloned into the sequences surroundingavrPphE in the chromosome or Psm M61 kb on each side carried on pBBR1-MCS2
This study
p186 vector for translation fusion (on the first reading frame)with the C terminus of AvrRpt2 and translational fusionwith GFP3 on a pBBR1-MCS2 backbone
J Chang (UNC)
pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

pFKN a virulence plasmid in P syringae pv maculicola 1559
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(iii) the size of the ORF was over 300 bp We may not havepredicted the correct initiation codon Where possible wechose an initiation codon that is preceded by an upstreamribosome binding site (RBS) sequence (optimally 5ndash13 bpbefore the initiation codon) The algorithms BLASTN andBLASTX were used to compare pFKN DNA sequences withthose in the GenBank database (httpwwwncbinlmnihgovblast) The predicted proteins were also analysed forfunctionally important motifs using PROSITE and for func-tional and structural domains using PFAM (httpwwwsangeracukSoftwarePfamsearchshtml) PRODOM (httpproteintoulouseinrafrprodomdocprodomhtml) SMART
(httpsmartembl-heidelbergde) and BLOCKS (httpblocksfhcrcorgblocksblocks_searchhtml) The sequencedata have been submitted to the DDJEMBLGenBank data-bases under the accession number AF359557 Thesequence of pFKN was compared with the genomesequence of P syringae pv tomato DC3000 available athttptigrblasttigrorgufmg and P syringae pv syringaeB728a available at httpwwwjgidoegovJGI_microbialhtmlindexhtml
Plasmid curing and complementation
The subclone pFK20 from the shotgun library carries thereplicase gene (ORF1) of pFKN that is also an incompatibilitydeterminant for plasmids belonging to the PT23A plasmidfamily (Gibbon et al 1999) pFK20 was electroporated intoPsm M6 to cure the strain of pFKN Single transformantswere grown in three consecutive liquid cultures without selec-tion for pFK20 (carbenicillin) The bacteria retrieved aftergrowth no longer contained pFK20 indicating that the frag-ment carried by pFK20 was not sufficient for plasmid main-tenance No pFKN sequences could be detected by DNAblot
For complementation tests fragments of pFKN werecloned into pBBR1-MCS2 (pLaR9 pLaR10 pLaR11pLaR12 and pLaR13 see Table 4) and transferred to thecured strain by triparental mating using the helper plasmidpRK2013
We deleted the avrPphE gene from the cured strain bymarker exchange as described by Hendrickson et al (2000)In brief avrPphE was replaced by an omega fragment carry-ing a gene conferring resistance to spectinomycin and strep-tomycin (Prentki and Krisch 1984) Sequences flanking thegene on both sides (1 kb each) were cloned into pBBR1-MCS2 carrying sacB (a lethal gene when the bacteria isgrown on sucrose Zagorec and Steinmetz 1990) Betweenthe two flanking regions was inserted the omega fragmentcarrying a gene conferring resistance to spectinomycin andstreptomycin (Prentki and Krisch 1984) The resulting plas-mid (pLaR24) was transformed by triparental mating into PsmM6C using the helper plasmid pRK2013 Deletion mutantswere then selected on spectinomycin (for integration of theomega fragment) and sucrose (for loss of pLaR24)
RT-PCR analysis
RNA was extracted from exponentially growing Psm M6 asdescribed by Innes et al (1993) The bacteria were subjected
to growth in the test medium for 3 h The RNA was thenextracted using TrizolR LS reagent (Gibco BRL) RNA sam-ples were treated with 10 U of RNase-free DNase I(Hoffmann-La Roche) for 1 h at 37infinC according to the man-ufacturerrsquos instructions The enzyme was then inactivated for5 min at 75infinC Reverse transcription reaction was performedusing the RETROscriptTM kit (Ambion) according to the pro-tocol provided by the manufacturer PCRs included primersspecific for each ORF to be tested and control primers allow-ing the amplification of part of the gene encoding for theglyceraldehyde-3P-dehydrogenase The amplification withthe control primers indicates that the non-amplification of thetested ORF results from the lack of expression of the geneand not a failed reaction The primer sets used for this controlwere Larp17 (CAGCAACTTGCCGCTCAC) and Larp10(ATGACTCTCCGTATCGC) giving rise to an amplificationproduct of 1070 bp or Larp17 and Larp11 (CTGCACCGTGAACTGGG) giving rise to a product of 406 bp The follow-ing protocol was used 3 min at 95infinC 30 cycles of 45 s at94infinC 45 s at 54infinC and 1 min at 72infinC and 5 min at 72infinCThe primer sequences used for each ORF are available onrequest
Transcriptional fusion and expression assay in planta
The 5cent ends of the genes for testing were cloned into p186(J Chang personal communication) to fuse the N-termini ofthe genes for testing to the gene encoding GFP3 Psm M6Cwas transformed with the constructs by triparental matingbetween Psm M6C DH5a carrying the construct or thehelper plasmid pRK2013 Leaves of eight rpm1-3 plants wereinoculated with each transformant at a density of 5 yen 107
cfu ml-1 The bacteria were retrieved from the inoculatedleaves by shaking the leaves in 10 mM MgCl2 and 002Silwet They were resuspended in glycerol and immediatelyobserved by fluorescence microscopy (Microscope EclipseE800 Nikon) For the GFP to be activated the bacteria wereexposed to blue light (488 nm)
TAIL-PCR method
The TAIL-PCR was a modified version of the methoddescribed by Liu and Whittier (1995) Primers used are listedin Table 5 The degenerate primers (AD1 to AD8 Table 5)were provided by P Tornero (UNC Chapel Hill NC USA)On average three to five degenerate primers yielded onespecific amplification product The biggest products wereordf1 kb PCR products were isolated on a 08 agarose gel(QIAquick gel isolation kit Qiagen) and sequenced directlyThe surrounding sequences of the avrPphE transposon inthe chromosome of Psm M6 are available in GenBank(AF544992) as well as the sequences from Psm M5(AF544991)
Acknowledgements
We thank Drs Jeff Chang Petra Epple Sarah R Grant GopalSubramaniam and Ben F Holt III and Zack Nimchuk for theirpatience and advice during the course of this work We areespecially grateful to The Institute for Genome Research
1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

1560 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
(TIGR) and the DOE-Joint Genome Initiative (JGI) for makingthe genome sequences of P syringae DC3000 and B728aavailable before publication Our work is supported by DOEDivision of Energy Biosciences grant DE-FG05-95ER20187
References
Alarcon-Chaidez FJ Penaloza-Vazquez A Ullrich Mand Bender CL (1999) Characterization of plasmidsencoding the phytotoxin coronatine in Pseudomonassyringae Plasmid 42 210ndash220
Alfano JR Charkowski AO Deng WL Badel JLPetnicki-Ocwieja T van Dijk K and Collmer A (2000)The Pseudomonas syringae Hrp pathogenicity island hasa tripartite mosaic structure composed of a cluster of typeIII secretion genes bounded by exchangeable effector andconserved effector loci that contribute to parasitic fitnessand pathogenicity in plants Proc Natl Acad Sci USA 974856ndash4861
Ansorge W Voss H and Zimmermann J (1997) DNASequencing Strategies Automated and AdvancedApproaches New York Wiley
Arnold DL Jackson RW Fillingham AJ Goss SCTaylor JD Mansfield JW and Vivian A (2001) Highlyconserved sequences flank avirulence genes isolation ofnovel avirulence genes from Pseudomonas syringae pvpisi Microbiology 147 1171ndash1182
Bavage AD Vivian A Atherton GT Taylor JD andMalik AN (1991) Molecular genetics of Pseudomonassyringae pathovar pisi plasmid involvement in cultivar-specific incompatibility J Gen Microbiol 137 2231ndash2239
Boch J Joardar V Gao L Robertson TL Lim M andKunkel BN (2002) Identification of Pseudomonas syrin-gae pv tomato genes induced during infection of Arabidop-sis thaliana Mol Microbiol 44 73ndash88
Cornelis GR and Van Gijsegem F (2000) Assembly and
function of type III secretory systems Annu Rev Microbiol54 735ndash774
Cournoyer B Sharp JD Astuto A Gibbon MJ TaylorJD and Vivian A (1995) Molecular characterization ofthe Pseudomonas syringae pv pisi plasmid- borne aviru-lence gene avrPpiB which matches the R3 resistance locusin pea Mol PlantndashMicrobe Interact 8 700ndash708
Curiale MS and Mills D (1982) Integration and partialexcision of a cryptic plasmid in Pseudomonas syringae pvphaseolicola J Bacteriol 152 797ndash802
Dangl JL and Jones JD (2001) Plant pathogens andintegrated defence responses to infection Nature 411826ndash833
Dangl JL Ritter C Gibbon MJ Mur LA Wood JRGoss S et al (1992) Functional homologs of the Arabi-dopsis RPM1 disease resistance gene in bean and peaPlant Cell 4 1359ndash1369
Delcher AL Harmon D Kasif S White O and SalzbergSL (1999) Improved microbial gene identification withGLIMMER Nucleic Acids Res 27 4636ndash4641
Diver WP Grinsted J Fritzinger DC Brown NLAltenbuchner J Rogowsky P and Schmitt R (1983)DNA sequences of and complementation by the tnpRgenes of Tn21 Tn501 and Tn1721 Mol Gen Genet 191189ndash193
Figurski DH and Helinski DR (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans Proc Natl Acad SciUSA 76 1648ndash1652
Fouts DE Abramovitch RB Alfano JR Baldo AMBuell CR Cartinhour S et al (2002) Genomewide iden-tification of Pseudomonas syringae pv tomato DC3000promoters controlled by the HrpL alternative sigma factorProc Natl Acad Sci USA 99 2275ndash2280
Froehlich BJ Tatti K and Scott JR (1983) Evidence forpositive regulation of plasmid prophage P1 replicationintegrative suppression by copy mutants J Bacteriol 156205ndash211
Table 5 Primers used in this study
Primer name Sequence (5cent-3cent) Purpose
fkp1405 lt GTGAAATCGCTTAGCGTACTTT TAIL-PCR on the left border of the avrPphE transposonfkp1444 lt AGCGGCAACAGCGCGGGGAAA TAIL-PCR on the left border of the avrPphE transposonfkp1509 lt GCCATAGCCGCTGAAGCAACATT TAIL-PCR on the left border of the avrPphE transposonfkp1559 lt TGTCGTCATTTGGCCGGGTC TAIL-PCR on the left border of the avrPphE transposonfkp4637 gt TGGGCGAAATCCGAGACCGTAGCT TAIL-PCR on the right border of the avrPphE transposonfkp4802 gt TGTCGCCGCTGGGCTGGG TAIL-PCR on the right border of the avrPphE transposonfkp6431 gt CGCCTCGGTCGTGACTGCATTG TAIL-PCR on the right border of the avrPphE transposonK110 TCGGCACTTTGTAGCCAGT Sequencing of the avrPphE transposonK113 GTCTTGCCCTGGCACCAC Sequencing of the avrPphE transposonLarp2 AATAGGTTTACACCTGTTTCACG Sequencing of the avrPphE transposonLarp3 TTTCACGCCCTTGGCCTTGG Sequencing of the avrPphE transposonAD1 NTCGASTWTSGWGTT Degenerated primers for TAIL-PCRAD2 NGTCGASWGANAWGAA Degenerated primers for TAIL-PCRAD3 WGTGNAGWANCANAGA Degenerated primers for TAIL-PCRAD4 NCTAGWASTWGSTTG Degenerated primers for TAIL-PCRAD5 NTGGCGWSATNTSATA Degenerated primers for TAIL-PCRAD6 TGWGNAGSANCASAGA Degenerated primers for TAIL-PCRAD7 AGWGNAGWANCAWAGG Degenerated primers for TAIL-PCRAD8 STTGNTASTNCTNTGC Degenerated primers for TAIL-PCR
pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

pFKN a virulence plasmid in P syringae pv maculicola 1561
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Gibbon MJ Sesma A Canal A Wood JR Hidalgo EBrown J et al (1999) Replication regions from plant-pathogenic Pseudomonas syringae plasmids are similar toColE2-related replicons Microbiology 145 325ndash334
Guttman DS Vinatzer BA Sarkar SF Ranall MVKettler G and Greenberg JT (2002) A functional screenfor the type III (Hrp) secretome of the plant pathogenPseudomonas syringae Science 295 1722ndash1726
Hacker J and Kaper JB (1999) The concept of pathoge-nicity islands In Pathogenicity Islands and Other MobileVirulence Elements Hacker J and Kaper JB (eds)Washington DC American Society for Microbiology Presspp 1ndash12
Hacker J and Kaper JB (2000) Pathogenicity islands andthe evolution of microbes Annu Rev Microbiol 54 641ndash679
Hendrickson EL Guevera P Penaloza-Vazquez AShao J Bender C and Ausubel FM (2000) Virulenceof the phytopathogen Pseudomonas syringae pv maculi-cola is rpoN dependent J Bacteriol 182 3498ndash3507
Hensel M Nikolaus T and Egelseer C (1999) Molecularand functional analysis indicates a mosaic structure ofSalmonella pathogenicity island 2 Mol Microbiol 31 489ndash498
Hobbs M and Reeves PR (1994) The JUMPstartsequence a 39 bp element common to several polysac-charide gene clusters Mol Microbiol 12 855ndash856
Hou YM (1999) Transfer RNAs and pathogenicity islandsTrends Biochem Sci 24 295ndash298
Huynh TV Dahlbeck D and Staskawicz BJ (1989) Bac-terial blight of soybean regulation of a pathogen genedetermining host cultivar specificity Science 245 1374ndash1377
Innes RW Bent AF Kunkel BN Bisgrove SR andStaskawicz BJ (1993) Molecular analysis of avirulencegene avrRpt2 and identification of a putative regulatorysequence common to all known Pseudomonas syringaeavirulence genes J Bacteriol 175 4859ndash4869
Jackson RW Athanassopoulos E Tsiamis G MansfieldJW Sesma A Arnold DL et al (1999) Identificationof a pathogenicity island which contains genes for viru-lence and avirulence on a large native plasmid in the beanpathogen Pseudomonas syringae pathovar phaseolicolaProc Natl Acad Sci USA 96 10875ndash10880
Jackson RW Mansfield JW Arnold DL Sesma APaynter CD Murillo J et al (2000) Excision from tRNAgenes of a large chromosomal region carrying avrPphBassociated with race change in the bean pathogenPseudomonas syringae pv phaseolicola Mol Microbiol 38186ndash197
Jin Q and He SY (2001) Role of the Hrp pilus in type IIIprotein secretion in Pseudomonas syringae Science 2942556ndash2558
Karaolis DK Somara S Maneval DR Jr Johnson JAand Kaper JB (1999) A bacteriophage encoding a patho-genicity island a type-IV pilus and a phage receptor incholera bacteria Nature 399 375ndash379
Kim JF Charkowski AO Alfano JR Collmer A andBeer SV (1998) Sequences related to transposableelements and bacteriophages flank avirulence genes ofPseudomonas syringae Mol PlantndashMicrobe Interact 111247ndash1252
King EO Ward MK and Raney DE (1954) Two simplemedia for the demonstration of phycocynin and fluorescineJ Lab Clin Med 44 301ndash307
Kovach ME Elzer PH Hill DS Robertson GT FarrisMA Roop RM II and Peterson KM (1995) Four newderivatives of the broad-host-range cloning vectorpBBR1MCS carrying different antibiotic-resistance cas-settes Gene 166 175ndash176
Li CM Brown I Mansfield J Stevens C Boureau TRomantschuk M and Taira S (2002) The Hrp pilus ofPseudomonas syringae elongates from its tip and acts asa conduit for translocation of the effector protein HrpZEMBO J 21 1909ndash1915
Lindgren PB Peet RC and Panopoulos NJ (1986)Gene cluster of Pseudomonas syringae pv lsquophaseolicolarsquocontrols pathogenicity of bean plants and hypersensitivityof nonhost plants J Bacteriol 168 512ndash522
Lindgren PB Panopoulos NJ Staskawicz BJ andDahlbeck D (1988) Genes required for pathogenicity andhypersensitivity are conserved and interchangeableamong pathovars of Pseudomonas syringae Mol GenGenet 211 499ndash506
Liu YG and Whittier RF (1995) Thermal asymmetricinterlaced PCR automatable amplification and sequencingof insert end fragments from P1 and YAC clones for chro-mosome walking Genomics 25 674ndash681
Lu SE Scholz-Schroeder BK and Gross DC (2002)Characterization of the salA syrF and syrG regulatorygenes located at the right border of the syringomycin genecluster of Pseudomonas syringae pv syringae Mol PlantndashMicrobe Interact 15 43ndash53
Mackey D Holt BF Wiig A and Dangl JL (2002) RIN4interacts with Pseudomonas syringae type III effector mol-ecules and is required for RPM1-mediated resistance inArabidopsis Cell 108 743ndash754
Maniatis T Fritsch EF and Sambrook J (1989) Molecu-lar Cloning A Laboratory Manual Cold Spring Harbor NYCold Spring Harbor Laboratory Press
Mansfield J Jenner C Hockenhull R Bennett MA andStewart R (1994) Characterization of avrPphE a gene forcultivar-specific avirulence from Pseudomonas syringaepv phaseolicola which is physically linked to hrpY a newhrp gene identified in the halo-blight bacterium Mol PlantndashMicrobe Interact 7 726ndash739
Nagy G Dobrindt U Kupfer M Emody L Karch H andHacker J (2001) Expression of hemin receptor moleculeChuA is influenced by RfaH in uropathogenic Escherichiacoli strain 536 Infect Immun 69 1924ndash1928
Nagy G Dobrindt U Schneider G Khan AS HackerJ and Emody L (2002) Loss of regulatory protein RfaHattenuates virulence of uropathogenic Escherichia coliInfect Immun 70 4406ndash4413
Nieto C Giraldo R Fernandez-Tresguerres E and DiazR (1992) Genetic and functional analysis of the basicreplicon of pPS10 a plasmid specific for Pseudomonasisolated from Pseudomonas syringae pathovar savastanoiJ Mol Biol 223 415ndash426
Nimchuk Z Rohmer L Chang JH and Dangl JL (2001)Knowing the dancer from the dance R-gene products andtheir interactions with other proteins from host and patho-gen Curr Opin Plant Biol 4 288ndash294
1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143

1562 L Rohmer S Kjemtrup P Marchesini and J L Dangl
copy 2003 Blackwell Publishing Ltd Molecular Microbiology 47 1545ndash1562
Norrander J Kempe T and Messing J (1983) Construc-tion of improved M13 vectors using oligodeoxynucleotide-directed mutagenesis Gene 26 101ndash106
Petnicki-Ocwieja T Schneider DJ Tam VC ChanceyST Shan L Jamir Y et al (2002) Genomewide iden-tification of proteins secreted by the Hrp type III proteinsecretion system of Pseudomonas syringae pv tomatoDC3000 Proc Natl Acad Sci USA 99 7652ndash7657
Poplawsky AR and Mills D (1987) Identification and map-ping of regions that confer plasmid functions and of sitesfor excisive recombination of plasmid pMMC7105 Mol GenGenet 206 265ndash272
Prentki P and Krisch HM (1984) In vitro insertionalmutagenesis with a selectable DNA fragment Gene 29303ndash313
Rajakumar K Bulach D Davies J Ambrose LSasakawa C and Adler B (1997) Identification of a chro-mosomal Shigella flexneri multi-antibiotic resistance locuswhich shares sequence and organizational similarity withthe resistance region of the plasmid NR1 Plasmid 37159ndash168
Rakin A Noelting C Schropp P and Heesemann J(2001) Integrative module of the high-pathogenicity islandof Yersinia Mol Microbiol 39 407ndash415
Ritter C and Dangl JL (1995) The avrRpm1 gene ofPseudomonas syringae pv maculicola is required for viru-lence on Arabidopsis Mol PlantndashMicrobe Interact 8 444ndash453
Santangelo TJ and Roberts JW (2002) RfaH a bacterialtranscription antiterminator Mol Cell 9 698ndash700
Sesma A Sundin GW and Murillo J (1998) Closelyrelated plasmid replicons coexisting in the phytopathogenPseudomonas syringae show a mosaic organization of thereplication region and altered incompatibility behavior ApplEnviron Microbiol 64 3948ndash3953
Sesma A Sundin GW and Murillo J (2000) Phylog-eny of the replication regions of pPT23A-like plasmidsfrom Pseudomonas syringae Microbiology 146 2375ndash2384
Sherrat D (1989) Tn3 and related TE site-specific recom-bination and transposition In Mobile DNA Berg DE andHowe MM (eds) Washington DC American Society forMicrobiology Press pp 163ndash184
Sinha H Pain A and Johnstone K (2000) Analysis of therole of recA in phenotypic switching of Pseudomonastolaasii J Bacteriol 182 6532ndash6535
Stevens C Bennett MA Athanassopoulos E TsiamisG Taylor JD and Mansfield JW (1998) Sequencevariations in alleles of the avirulence gene avrPphER2from Pseudomonas syringae pv phaseolicola lead to lossof recognition of the AvrPphE protein within bean cells anda gain in cultivar-specific virulence Mol Microbiol 29 165ndash177
Syn CK and Swarup S (2000) A scalable protocol for theisolation of large-sized genomic DNA within an hour fromseveral bacteria Anal Biochem 278 86ndash90
Szabo LJ and Mills D (1984a) Characterization of eightexcision plasmids of Pseudomonas syringae pv phaseoli-cola Mol Gen Genet 195 90ndash95
Szabo LJ and Mills D (1984b) Integration and excision ofpMC7105 in Pseudomonas syringae pv phaseolicolainvolvement of repetitive sequences J Bacteriol 157 821ndash827
Tauschek M Strugnell RA and Robins-Browne RM(2002) Characterization and evidence of mobilization ofthe LEE pathogenicity island of rabbit-specific strains ofenteropathogenic Escherichia coli Mol Microbiol 441533ndash1550
Thompson JF and Landy A (1989) Regulation of thebacteriophage lambda site-specific recombination InMobile DNA Berg DE and Howe MM (eds) Wash-ington DC American Society for Microbiology Press pp1ndash22
Van der Hoorn RAL De Wit PJGM and JoostenMHAJ (2002) Balancing selection favors guarding resis-tance proteins Trends Plant Sci 7 67ndash71
Vivian A Murillo J and Jackson RW (2001) The roles ofplasmids in phytopathogenic bacteria mobile arsenalsMicrobiology 147 763ndash780
Xiao Y Heu S Yi J Lu Y and Hutcheson SW (1994)Identification of a putative alternate sigma factor and char-acterization of a multicomponent regulatory cascade con-trolling the expression of Pseudomonas syringae pvsyringae Pss61 hrp and hrmA genes J Bacteriol 1761025ndash1036
Yamaguchi K and Tomizawa J (1980) Establishment ofEscherichia coli cells with an integrated high copy numberplasmid Mol Gen Genet 178 525ndash533
Zagorec M and Steinmetz M (1990) Expression of levan-sucrase-beta-galactosidase hybrids inhibits secretion andis lethal in Bacillus subtilis J Gen Microbiol 136 1137ndash1143