notch signalling pathway
TRANSCRIPT
S
e
e
o
n
l
i
n
e
v
e
r
s
i
o
n
f
o
r
l
e
g
e
n
d
a
n
d
r
e
f
e
r
e
n
c
e
s
.
1 2 4
6
C
e
l
l
1
2
8
,
M
a
r
c
h
2
3
,
2
0
0
7
©
2
0
0
7
E
l
s
e
v
i
e
r
I
n
c
.
D
O
I
1
0
.
1
0
1
6
/
j
.
c
e
l
l
.
2
0
0
7
.
0
3
.
0
1
1
SnapShot: Notch Signaling PathwayMa. Xenia G. Ilagan and Raphael Kopan
Department of Molecular Biology and Pharmacology, Washington University School of Medicine, St. Louis, MO 63110, USA

SnapShot:Notch Signaling PathwayMa. Xenia G. Ilagan and Raphael Kopan
Department of Molecular Biology and Pharmacology, Washington University School of Medicine,
St. Louis, MO 63110, USA
1246.e1 Cell 128, March 23, 2007 ©2007 Elsevier Inc. DOI 10.1016/j.cell.2007.03.011
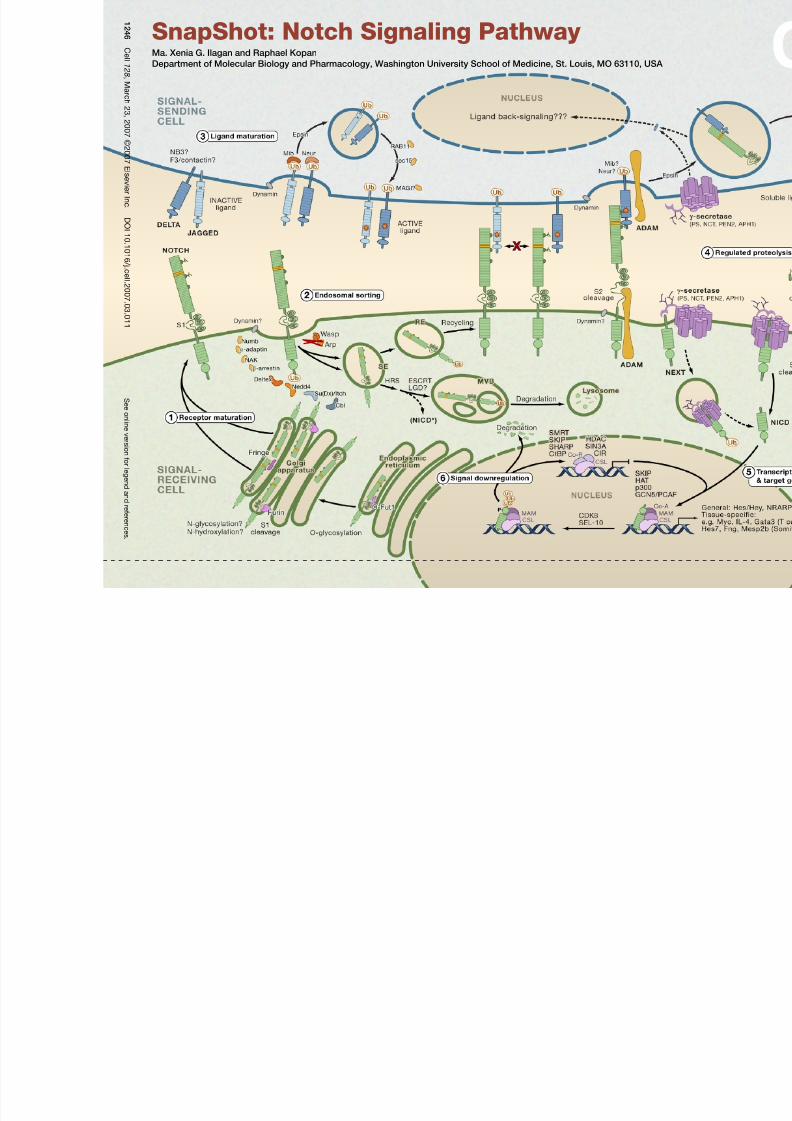
The Notch signaling pathway is a short-range communication transducer that is involved in regulating many cellular processes (proliferation, stem cell and stem cell
niche maintenance, cell fate specification, differentiation, and cell death) during development and renewal of adult tissues. Notch signaling is mediated by proteolysis
and does not appear to involve any secondary messengers. However, depending on cellular context, the amplitude and timing of Notch activity can be regulated by
posttranslational modifications to ligands and receptors and their trafficking.
(1) Receptor MaturationNotch receptors are large single pass type I transmembrane proteins. Upon translation, the Notch protein is fucosylated by the chaperone O-fut, a modification es-
sential for the production of a functional receptor. In cells expressing Fringe, the fucose is extended by the glycosyltransferase activity of Fringe, altering the ability of
specific ligands to activate Notch (see below). The mature receptor undergoes proteolytic cleavage by protein convertases (PC5; Furin) at site 1 (S1) and then is targeted
to the cell surface as a heterodimer held together by noncovalent interactions.
(2) Endosomal SortingSeveral mechanisms control the steady-state levels of the Notch receptor at the cell surface and therefore regulate its availability for binding of ligand. For example,
Numb, in cooperation with the AP2 component α-adaptin and AP2- or Numb-associated kinase (NAK), can promote Notch endocytosis and degradation. Several E3
ubiquitin ligases—Deltex, Nedd4, Su(Dx)/Itch, Cbl—target Notch, shifting receptor trafficking toward degradation or recycling. Other proteins prevent inappropriate
receptor activation in the absence of ligand binding. Mutations in certain ESCRT complex proteins lead to accumulation of Notch in endosomal vesicles, which surpris-
ingly permits ectopic activation of Notch via γ-secretase-dependent proteolysis (NICD*). Another protein, Lethal Giant Discs (LGD), is also required to maintain Notch
in the OFF state. Therefore, ESCRT and LGD complexes are normally involved in Notch downregulation, indicating that endosomal sorting is a key way to restrict
activation of Notch to the cell surface and that defects in endosomal sorting may contribute to pathogenesis. SE, sorting endosome; RE, recycling endosome; MVB,
multivesicular body.
(3) Ligand MaturationNotch ligands are also type I transmembrane proteins characterized by an N-terminal DSL domain. The two major classes of ligands are Delta and Jagged (Serrate in
Drosophila ), the latter containing a cysteine-rich domain. In addition to Delta and Jagged, the neural adhesion molecule F3/contactin, the related NB-3 protein, the EGF
repeat protein DNER, and a diffusible protein in C. elegans have been identified as potential Notch ligands. Endocytic trafficking of the DSL ligands is crucial for en-
hancing their signaling activity: Ligands are ubiquitinated by the E3 ubiquitin ligases, Neur and Mib, triggering Epsin-mediated endocytosis; an undefined modification
produces an active ligand that recycles to the cell surface in a Rab11-dependent process. Current models explaining the nature of ligand modification include ligand
clustering, posttranslational modifications, and/or recycling into specific membrane domains.
(4) Regulated ProteolysisThe Notch receptor is activated by binding to a ligand presented by a neighboring cell. Productive receptor-ligand interactions depend on the glycosylation state, for
example, a Fringe-modified receptor may favor binding of Delta. Ligand endocytosis is thought to generate sufficient force to produce partial or complete domain
dissociation, thereby exposing Notch to cleavage at site S2 by ADAM metalloproteases (perhaps following heterodimer dissociation at S1). The Notch extracellular
domain is transendocytosed into the signal-sending cell, whereas the membrane-anchored NEXT ( N otch ex tracellular t runcation) fragment is recognized by the inactive
aminopeptidase domain of nicastrin (NCT), which transfers NEXT to the active site of γ-secretase, an enzymatic complex composed of presenilin (PS), NCT, PEN2, and
APH1. γ-secretase then cleaves the Notch transmembrane domain sequentially starting near the cytosolic surface (sites S3 and S4) to release the Notch intracellular
domain (NICD) and Nβ peptides, respectively.γ-secretase cleavage can occur at the cell surface or in endosomal compartments, perhaps following monoubiquitination.
The apical polarity protein Crumbs appears to play a role in restricting γ-secretase activity thereby limiting the extent of Notch activation. Like many type I proteins,
Notch ligands are also subject to extracellular cleavage by ADAM proteases followed by transmembrane domain cleavage by γ-secretase. Ligand processing may be
important to reduce its ability to antagonize Notch signaling in cis and for its downregulation and membrane clearance. Alternatively, it could generate biologically active
fragments, e.g., soluble ligands (that may act as antagonists of Notch signaling) and/or ligand intracellular domain fragments.
(5) Transcriptional Switch and Target Gene ExpressionIn the absence of NICD, the DNA-binding protein CSL associates with ubiquitous corepressor (Co-R) proteins and histone deacetylases (HDACs) to repress transcrip-
tion of target genes. When NICD enters the nucleus, its binding to CSL may trigger an allosteric change that facilitates displacement of transcriptional repressors. The
NICD/CSL interface is then recognized by Mastermind (MAM), and this tri-protein complex recruits coactivators (Co-A)—such as histone acetylases (HATs), chromatin-
remodeling factors, and a mediator complex— to assemble an active transcription complex on target promoters.
(6) Signal DownregulationDuring transcriptional activation, NICD is phosphorylated on its PEST domain by kinases such as CDK8 and targeted for proteasomal degradation by E3 ubiquitin
ligases such as Sel10/Fbw7. This terminates the Notch signal and resets the cell for the next round of signaling.
REFERENCES
Bray, S.J. (2006). Notch signalling: a simple pathway becomes complex. Nat. Rev. Mol. Cell Biol. 7 , 678–689.
Ehebauer, M., Hayward, P., and Martinez-Arias, A. (2006). Notch signaling pathway. Sci. STKE 2006, cm7.
Haines, N., and Irvine, K.D. (2003). Glycosylation regulates Notch signalling. Nat. Rev. Mol. Cell Biol. 4, 786–797.
Kadesch, T. (2004). Notch signaling: the demise of elegant simplicity. Curr. Opin. Genet. Dev.14, 506–512.
Kovall, R.A. (2007). Structures of CSL, Notch and Mastermind proteins: piecing together an active transcription complex. Curr. Opin. Struct. Biol. 17 , 117–127.
Le Borgne, R. (2006). Regulation of Notch signalling by endocytosis and endosomal sorting. Curr. Opin. Cell Biol. 18, 213–222.
Lubman, O.Y., Korolev, S.V., and Kopan, R. (2004). Anchoring notch genetics and biochemistry; structural analysis of the ankyrin domain sheds light on existing data.
Mol. Cell 13, 619–626.
Mumm, J.S., and Kopan, R. (2000). Notch signaling: from the outside in. Dev. Biol. 228, 151–165.
Schweisguth, F. (2004). Regulation of notch signaling activity. Curr. Biol. 14, R129–R138.
Wilkin, M.B., and Baron, M. (2005). Endocytic regulation of Notch activation and down-regulation (review). Mol. Membr. Biol. 22, 279–289.
ACKNOWLEDGMENTS
M.X.G.I., R.K., and Washington University may receive income based on a license of Notch-related technology by the University to Merck. Merck did not support this work.
