morphologic development and clay redistribution in lysimeter soils under chaparral and pine
TRANSCRIPT

Morphologic Development and Clay Redistribution in Lysimeter Soilsunder Chaparral and Pine
R. C. Graham* and H. B. Wood
ABSTRACTLysimeter soils under 41-yr-old stands of chaparral species and
a pine (Pinus coulteri B. Don) were studied to determine the bioticinfluence on soil morphologic development. The large (5.3 by 5.3 by2.1 m) unconfined lysimeters at the San Dimas Experimental Forestin southern California were filled in 1937 with homogenized finesandy loam soil material derived from diorite. After a 9-yr stabili-zation period, the lysimeters were planted with monocultures of spe-cies common to California chaparral ecosystems. The speciesrepresented, each on a separate lysimeter and its surrounding bufferzone, are scrub oak (Quercus dumosa Nutt.), ceanothus (Ceanothuscrassifolia Torn), chamise (Adenostomafasciculatum Hook. & Am.),and Coulter pine. The soils were described and sampled in triplicatein 1987. Differences in earthworm (including Aporrectodea trape-zoides Duges and Allolobophora chlorotica Savigny) activity wererelated to plant species and played a major role in differentiatingthe soils. Earthworm activity under scrub oak and ceanothus pro-duced A horizons (7 and 2 cm thick, respectively), composed largelyof worm casts, that were darker and enriched in clay compared withunderlying C horizons and archived fill material. Earthworm activitywas minimal under chamise, and relatively little morphologic de-velopment was apparent. The soil under pine, where earthwormswere absent, had a clay-depleted A horizon (1 cm thick) and Bthorizons with sufficient clay increase, illuviation argillans, and thick-ness to qualify as an argillic horizon. Mixing and sorting of soil byearthworms has acted against argillic horizon development underthe other plant species.
VEGETATION has been recognized as important insoil formation since the earliest scientific con-
siderations of soils (Buol et al., 1989). Jenny (1941)included biota as one of the five soil-forming factors,along with climate, topography, parent material, andtime, in his formal expressions of soil-forming func-tions. Jenny's work inspired much soil genesis researchutilizing the factor functions, or sequence, approach.In such studies, situations in nature are sought whereall factors are constant except the one under investi-gation so that the effect of that individual factor onsoil formation can be assessed. However, because soilproperties and vegetation (the main form of the bioticfactor) commonly covary in response to other factors(i.e., climate, topography, parent material, time), theplant factor in soil genesis has been difficult to assess(Jenny, 1961;Crocker, 1967;Yaalon, 1975). Situationsin nature where vegetation is the only variable in soilformation are exceedingly rare (Birkeland, 1984). Ar-tificially prepared experiments for plant-factor soilgenesis studies also are rare because such experimentsrequire several decades—a time frame that exceeds thescope of most research plans.
The large unconfined lysimeters at the San DimasR.C. Graham, Dep. of Soil and Environmental Sciences, Univ. ofCalifornia, Riverside, CA 92521-0424; and H.B. Wood, U.S. ForestService, Pacific Southwest Forest and Range Exp. Stn., Riverside,CA 92507. Received 18 Sept. 1990. Corresponding author.
Published in Soil Sci. Soc. Am. J. 55:1638-1646 (1991).
Experimental Forest in southern California present aunique opportunity to study soil formation in a se-quence of soils where the: biotic factor is the only var-iable, the site history is well documented, and archivedsamples of parent material are available for compar-ison. The lysimeters were filled with homogenized soilmaterial in 1937 and, after a 9-yr period to allow set-tling and stabilization, were planted with monocul-tures of native plant species (Colman and Hamilton,1947). Since water-budget studies at the installationwere abandoned in 1960, soils research has been lim-ited to periodic monitoring of soil N and base-cationnutrients (Zinke, 1969; 1977), yet the soils and estab-lished plant communities remain—an unintentionalexperiment in soil genesis. The objective of this re-search was to assess the morphologic development andredistribution of clay in the soils of the San Dimasunconfined lysimeters 41 yr after the establishment ofchaparral and pine species.
MATERIALS AND METHODSEnvironmental Setting
The San Dimas Experimental Forest is a U.S. Forest Ser-vice watershed research laboratory 56 km northeast of LosAngeles, CA, in the San Gabriel Mountains. The lysimeterinstallation is in an area known as Tanbark Flat at an ele-vation of 830 m. The surrounding natural vegetation is cha-parral, dominantly chamise, hoaryleaf ceanothus, bigberrymanzanita (Arctostaphylos glauca Lindl.), and scrub oak.The mean annual precipitation is 678 mm with annual pre-cipitation ranging from 292 to 1224 mm (Dunn et al., 1988).Most of the precipitation falls as rain between Decemberand March. The mean annual air temperature is. 14.4 °C,with summer temperatures frequently exceeding 38 °C andwinter temperatures rarely below — 4 °C (Dunn et al., 1988).
Soil temperature data collected at the lysimeters in 1952through 1959 show the mean annual soil temperature at the61-cm depth to be 13.6 °C under oak and 16.1 °C underpine (Qashu and Zinke, 1964). These data indicate mesicand thermic soil temperature regimes under oak and pine,respectively (Soil Survey Staff, 1990). Soil temperatures werenot monitored for this study, but denser canopies and thick-ened litter layers, particularly under the pine, have probablyresulted in lower soil temperatures. It is likely that in 1987all of the soils met the mesic temperature regime criteria.
Most of the San Dimas Experimental Forest is in steep(30-60% slopes), highly dissected terrain of plutonic andmetamorphic rock. The topography at Tanbark Rat is moresubdued. The nearly level lysimeter installation was devel-oped on the footslope of a broad spur ridge underlain bydiorite. Soils on the steeper slopes are mostly Lithic andTypic Xerorthents (Ryan, 1991). On less steep slopes, soilsderived from diorite and granodiorite typically have illuvialaccumulations of clay in the subsoil and underlying fracturedrock (Crawford, 1962) and are Haploxeralfs.
The LysimetersIn the late 1930s, several types of lysimeters were con-
structed within a 0.8-ha enclosure to study the influence ofchaparral species on the disposition of rainfall and soil water.
1638

GRAHAM & WOOD: MORPHOLOGIC DEVELOPMENT IN LYSIMETER SOILS 1639
Of specific interest to this study are the unconfined (earthen-walled) lysimeters, which have 5.3 by 5.3 m horizontal di-mensions and are 2.1 m deep. The arrangement of the lys-imeters was specifically designed to minimize variation ofall environmental influences except vegetation (Colman andHamilton, 1947; Patric, 1961a). In 1937, all of the lysimeterswere filled with a brown (7.5YR 5/4, dry; 7.5YR 4/4, moist)fine sandy loam derived on site from the weathering of di-orite. The excavated material from the lysimeter site wasmixed thoroughly, passed through a 19-mm-mesh screen,and stockpiled in a large, covered bin. Great care was takento achieve a homogeneous material for filling the lysimeters.Analyses at the time showed no statistically significant dif-ference in texture in 100 random samples taken from thestorage bin (Patric, 1961a). The lysimeters were filled in 7.5-cm-thick increments, with each successive layer beingchopped thoroughly with a flat-bladed spade after placement.During the filling, composite samples of each layer wereplaced in labeled glass jars and archived.
The lysimeters were overfilled by 30 cm to allow for set-tling. During the settling period (1937-1940), the soil sur-faces were maintained free of vegetation and protected fromerosion by an excelsior mulch. In the fall of 1940, the overfillwas removed to establish the final 2.1-m depth, a 5% slopeto the south was imposed on the surfaces, and an annualgrass (Bromus mollis L.) was planted. In 1946, the grass coverwas burned and replaced by chaparral species: buckwheat(Eriogonum fasciculatum var. foliolosum Nutt), chamise,hoaryleaf ceanothus, scrub oak, and Coulter pine. Thesewere planted in monocultures that covered 17 by 24 m areasthat included the unconfined lysimeters and buffer strips de-signed to eliminate edge effects. The monocultures weremaintained by periodic weeding, and by the mid-1950s allof the lysimeters supported pure stands of wildland speciessimilar in size and density to natural stands (Patric, 1961 a,b).
Water-budget research using the large lysimeters was besetwith problems from the beginning and was finally abandonedin 1960 after a wildfire burned all vegetation on the instal-lation, except the pine. The other plants resprouted, as istypical of chaparral species, and by 1972 vegetation wasagain vigorous and in virtually pure stands as originallyplanted (J.H. Patric, 1974, unpublished data). For the pine,oak, ceanothus, and chamise stands, these conditions pre-vailed with no further disturbance until the time of samplingin 1987. The buckwheat stand was excluded from this studybecause, in 1987, it was thin and had been invaded by otherspecies.
It should be noted that the lysimeter fill material may notbe a realistic analog for many types of natural sediments. Itdiffers from alluvial and eolian sediments in that, except forthe removal of coarse fragments >19 mm in diameter, it isunsorted. It may be most similar to thoroughly mixed, un-sorted debris or mud-flow colluvium.
Field and Laboratory MethodsThe soils were sampled in May and June of 1987. On each
lysimeter plot, three pedons were sampled from exposed pro-files in pits. One pedon on each lysimeter was sampled to adepth of 1 m, the other two were sampled to the 35-cm depth.Precise locations disturbed by sampling are on file with theUSDA Pacific Southwest Forest and Range Experiment Sta-tion, Riverside, CA. Pedons were described and sampled bymorphologic horizons using conventional procedures (SoilConservation Service, 1984). Below the apparent morpho-logic horizons exposed in the 1-m-deep pits, 15-cm incre-ments were sampled to 80 cm and a final 20-cm incrementto 100 cm. Bulk samples were taken from all pedons to pro-vide triplicate sampling of the horizons within the upper 35cm under each vegetation type. Clod samples for bulk den-sity and micromorphological analyses were taken from the
pedons sampled to 1 m. Archived samples from matchingfill layers were subsampled from the glass jars in which theyhad been stored.
Bulk samples were air dried and sieved to remove coarsefragments (>2 mm). The pH was measured in 1:1 soil/watersuspensions. Particle-size distribution was determined by thepipet method (Gee and Bauder, 1986). Water repellency wasmeasured using the water-drop penetration test (King, 1981).Bulk density was determined on saran-coated clods, or usingcores in the case of thin, fragile A horizons (Soil Conser-vation Service, 1984). Thin sections (26 by 46 mm) wereprepared from epoxy-impregnated clods. Clod samples werechosen from the small (<19-mm-diam.) clods present in thearchived samples and from the intact fabric of 1987 lysimetersoils. In most cases, only one thin section was prepared foreach sample because, particularly for the archived samples,clod-material quantities were very limited. Micromorphol-ogical features were described following the terminology ofBrewer (1976) and were quantified by counting 1000 pointsalong evenly spaced line transects of thin sections using apetrographic microscope. Two archived clod samples weretoo small to allow 1000 points to be counted, thus 428 pointswere counted for the fill depths of 74 to 81 cm under oakand 752 points for the 88 to 96 cm fill depth under pine.
RESULTS AND DISCUSSIONField Morphology
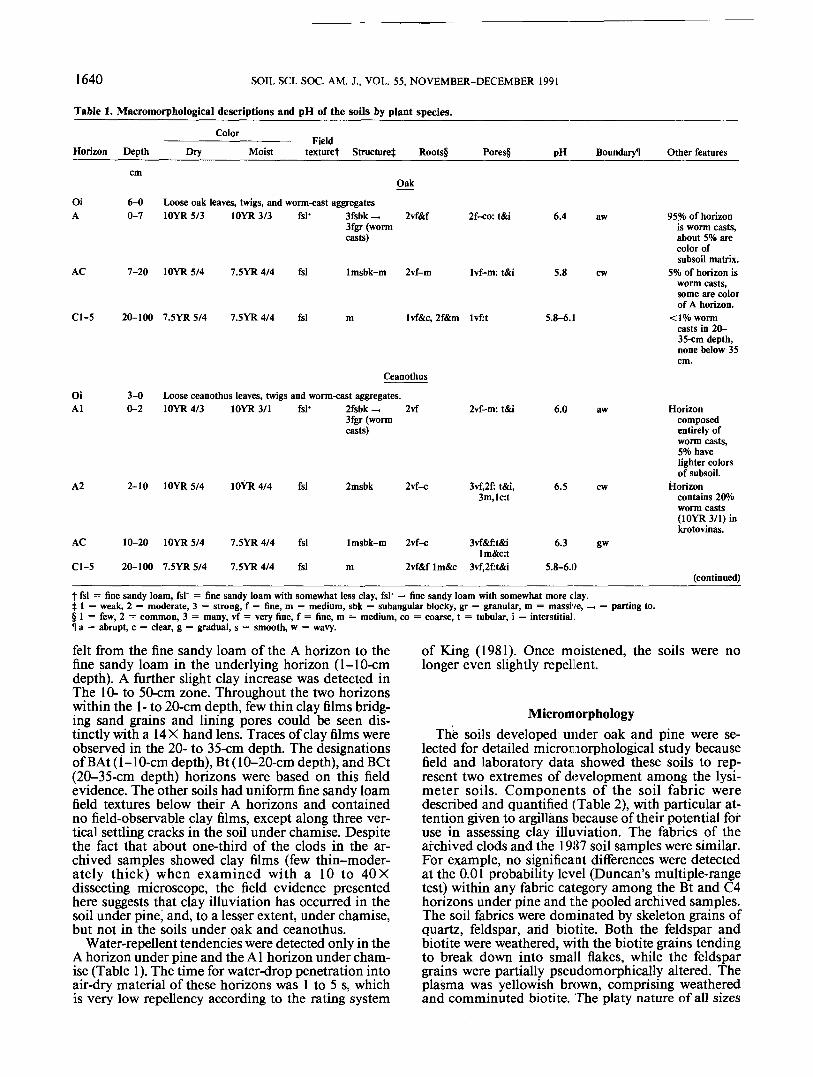
Macromorphological descriptions of the soils arepresented in Table 1. All soils had litter accumulatedas O horizons; the thickest was under pine (10 cm),and the thinnest was under ceanothus (3 cm). Underoak, ceanothus, and chamise, the litter layer consistedof a single horizon of loose, relatively fresh leaves andtwigs, designated Oi. Earthworm casts were also pre-sent in these O horizons. In contrast, the litter layerunder pine consisted of three distinct horizons: an Oilcomposed of fresh pine needles, an Oi2 with partiallydecomposed pine needles, and an Oe of mostly de-composed needles. No worm casts were found in thepine litter. Guild (1955) reported that pine needles areamong the least preferred of possible foods in earth-worms' diets.
Development of A horizons varied among the soilsand was related to the activity of earthworms. The Ahorizon under oak was 7 cm thick, dark brown (10YR3/3, moist), and consisted nearly entirely of wormcasts. The casts were aggregated, resulting in strongfine subangular blocky structure that broke down tostrong fine granular structure of individual casts.About 5% of the casts were the color of the subsoilmatrix (7. SYR 5/4, dry; 7.5 YR 4/4, moist). The tex-ture of this A horizon felt somewhat finer than thoseof the underlying horizons, although all were withinthe fine sandy loam textural class. A similar horizonunder ceanothus was only 2 cm thick. The earthworm-worked A horizon under chamise was only 1 cm thickand had only about 50% worm casts. In all of thesesoils, worm krotovinas extended into A subhorizonswith decreasing frequency to about the 20- to 35-cmdepth. Below the depth of worm activity, the soils weremassive, had the uniform color of the archived parentmaterial, and were designated C horizons.
In contrast, the soil under pine had a 1-cm-thick Ahorizon that was completely devoid of worm casts. Byhand texturing, a slight, but distinct, clay increase was
1640 SOIL SCI. SOC. AM. J., VOL. 55, NOVEMBER-DECEMBER 1991
Table 1. Macromorphological descriptions and pH of the soils by plant species.
ColorHorizon Depth Dry Moist
Fieldtexturef Structure}: Roots§ Pores§ pH BoundaryH Other features
Oak
Oi 6-0 Loose oak leaves, twigs, and worm<ast aggregatesA 0-7 10YR5/3 10YR 3/3 fsl* 3fsbk — 2vf&f
3fgr (wormcasts)
2f-co: t&i 6.4 aw
AC 7-20 10YR 5/4 7.5YR 4/4 fsl Imsbk-m 2vf-m Ivf-m: t&i 5.8 cw
Cl-5 20-100 7.5YR5/4 7.5YR 4/4 fsl m Ivf&c, 2f&m lvf:t 5.8-6.1
OiAl
Ceanothus
3-0 Loose ceanothus leaves, twigs and worm-cast aggregates.0-2 10YR4/3 10YR3/1 fsl* 2fsbk _ 2vf
3fgr (wormcasts)
2vf-m: t&i 6.0
A2 2-10 10YR 5/4 10YR 4/4 fsl 2msbk 2vf-c 3vf,2f: t&i,3m,lc:t
6.5
95% of horizonis worm casts,about 5% arecolor ofsubsoil matrix.
5% of horizon isworm casts,some are colorof A horizon.
<1% wormcasts in 20-35-cm depth,none below 35cm.
Horizoncomposedentirely ofworm casts,5% havelighter colorsof subsoil.
Horizoncontains 20%worm casts(10YR 3/1) inkrotovinas.
AC
Cl-5
10-20
20-100
10YR 5/4
7.5YR 5/4
7.5YR 4/4
7.5YR 4/4
fsl
fsl
Imsbk-m
m
2vf-c
2vf&f Im&c
3vf&f:t&ilm&c:t
3vf,2f:t&i
6.3 gw
5.8-6.D(continued)
t fsl = fine sandy loam, fsl~ = fine sandy loam with somewhat less clay, fsl* = fine sandy loam with somewhat more clay,j 1 = weak, 2 — moderate, 3 = strong, f = fine, m = medium, sbk = subangular blocky, gr = granular, m = massive, —§ 1 = few, 2 = common, 3 = many, vf = very fine, f = fine, m = medium, co = coarse, t = tubular, i = interstitial.11 a = abrupt, c = clear, g = gradual, s — smooth, w = wavy.
parting to.
felt from the fine sandy loam of the A horizon to thefine sandy loam in the underlying horizon (1-10-cmdepth). A further slight clay increase was detected inThe 10- to 50-cm zone. Throughout the two horizonswithin the 1- to 20-cm depth, few thin clay films bridg-ing sand grains and lining pores could be seen dis-tinctly with a 14 X hand lens. Traces of clay films wereobserved in the 20- to 35-cm depth. The designationsof BAt (i-10-cm depth), Bt (10-20-cm depth), and BCt(20-35-cm depth) horizons were based on this fieldevidence. The other soils had uniform fine sandy loamfield textures below their A horizons and containedno field-observable clay films, except along three ver-tical settling cracks in the soil under chamise. Despitethe fact that about one-third of the clods in the ar-chived samples showed clay films (few thin-moder-ately thick) when examined with a 10 to 40 Xdissecting microscope, the field evidence presentedhere suggests that clay illuviation has occurred in thesoil under pine, and, to a lesser extent, under chamise,but not in the soils under oak and ceanothus.
Water-repellent tendencies were detected only in theA horizon under pine and the Al horizon under cham-ise (Table 1). The time for water-drop penetration intoair-dry material of these horizons was 1 to 5 s, whichis very low repellency according to the rating system
of King (1981). Once moistened, the soils were nolonger even slightly repellent.
Microm orphologyThe soils developed under oak and pine were se-
lected for detailed micromorphological study becausefield and laboratory data showed these soils to rep-resent two extremes of development among the lysi-meter soils. Components of the soil fabric weredescribed and quantified (Table 2), with particular at-tention given to argillans because of their potential foruse in assessing clay illuviation. The fabrics of thearchived clods and the 1987 soil samples were similar.For example, no significant differences were detectedat the 0.01 probability level (Duncan's multiple-rangetest) within any fabric category among the Bt and C4horizons under pine and the pooled archived samples.The soil fabrics were dominated by skeleton grains ofquartz, feldspar, and biotite. Both the feldspar andbiotite were weathered, with the biotite grains tendingto break down into small flakes, while the feldspargrains were partially pseudomorphically altered. Theplasma was yellowish brown, comprising weatheredand comminuted biotite. The platy nature of all sizes

GRAHAM & WOOD: MORPHOLOGIC DEVELOPMENT IN LYSIMETER SOILS 1641
Table 1. (continued).
Color
Horizon Depth Dry MoistField
texturef Structure}: Roots§ PH BoundaryH Other features
Chamise
OiAl
A2
AC
Cl
5-0 Loose chamise leaves, twigs, and worm-cast aggregates. Fungal mycelia occupy <5% of horizon, up to 50% in pockets.0-1 10YR 4/4 10YR 3/3
1-7 10YR5/4
7-20 7.5YR 5/4
20-35 7.5YR 5/4
10YR 4/4
10YR 4/4
7.5YR 4/4
fsl-
fsl
fsl
fsl
3msbk _ Ivf3fgr (wormcasts)
2msbk
Imsbk
m
2vf&f
3vf&f, 2m &co:t&i
3vf,2f-co:t&i
2vf&f, 1m 3vf,2f, 1m &co: t&i
2vf-m 3vf,2f,lm:t&i
6.4
6.4
6.0
cw
gw
50% worm casts(10YR 3/2).Slightly waterrepellent.
10% worm casts(10YR 3/2) inkrotovinas.
<2% wormcasts
Very few wormkrotovinas,same color asmatrix. Fewthin clay filmsbridging grainsand on facesalong verticalsettling cracks.
C2-5
011012OeA
BAt
Bt
BCt
ClC2-4
35-100 7.5YR 5/4 7.5YR 4/4 fsl
10-6 Fresh pine needles, twigs, some grass fragments.6-4 Partially decomposed pine needles.4-0 Mostly decomposed pine needles.0-1 10YR4/4 10YR4/3 fsl- 2msbk
1-10 10YR 5/4
10-20 7.5YR 5/4
20-35 7.5YR 5/4
35-50 7.5YR 5/450-100 7.5YR5/4
7.5YR 4/4
7.5YR 4/4
7.5YR 4/4
7.5YR 4/47.5YR 4/4
fsl
fsl*
fsl*
fsl*fsl
2msbk
Imsbk
mm
2vf-mPine
2vf&f3vf,2f3vf,2f
2vf&f,Im&co
Ivf-co
Ivf-m
Ivf-coIvf-co
3vf,2f:t&i
3vf:t&i
3vf:t
2vf:t
2vf:t
2vf:t2vf:t
6.0-6.2
5.5
6.0
6.0
5.9
6.06.1-6.2
asasawaw
gw
gw
gw
Slightly waterrepellent.
Few thin clayfilms bridgingsand grainsand liningpores.
Few thin clayfilms bridgingsand grainsand liningpores.
Trace of clayfilms.
t fsl = fine sandy loam, fsl~ = fine sandy loam with somewhat less clay, fsl* = fine sandy loam with somewhat more clay.j 1 = weak, 2 = moderate, 3 = strong, f = fine, m = medium, sbk = subangular blocky, gr = granular, m = massive, —> = parting to.§ 1 = few, 2 = common, 3 = many, vf = very fine, f = fine, m = medium, co = coarse, t = tubular, i = interstitial.H a = abrupt, c = clear, g = gradual, s = smooth, w = wavy.
of weathered biotite grains enhanced their tendency tobecome oriented, particularly in the vicinity of larger,less flexible skeleton grains. All samples had inskel-sepic plasmic fabrics.
Simple packing voids were the dominant void typein all samples reported in Table 2. Root channels werealso present, and they were more common in the ar-chived clods than in the 1987 lysimeter soils. Organicroot material was present in some channels in the 1987samples (Table 2).
Both free and embedded types of grain argillans werepresent in the 1987 samples, whereas only the free typewas present in the archived samples. The free-grainargillans had moderate and strong continuous orien-tation, while the embedded-grain argillans (Fig. la andIb) had weak to moderate striated orientation, pos-sibly the result of pressure stresses in the denselypacked lysimeter soil material (Brewer, 1976).
Because grain argillans may result from processes
other than illuviation and because they are readily pre-served in disturbed materials, they were not usefulindicators of illuvial clay accumulation in the 1987soils relative to the archived fill material. On the otherhand, the channel argillans were associated with con-ducting voids and are clearly illuviation features,based on the diagnostic criteria of Brewer (1976).Channel argillans with moderate to strong continuousorientation were present in both the clods from thearchived samples and the 1987 soil fabrics (Table 2,Fig. lc and Id). Therefore, it is likely that the 1987soils contain illuviation features inherited from the fillmaterial. In fact, a few apparent pedorelicts of originalclods with strongly oriented argillans were observedin the thin sections of the 1987 soil fabrics. Unfor-tunately, no independent criteria could be identifiedto allow inherited argillans to be distinguished fromthose that may have formed since the lysimeters werefilled.

1642 SOIL SCI. SOC. AM. J., VOL. 55, NOVEMBER-DECEMBER 1991
Mean values for channel-argillan contents of ar-chived clods and 1987 Bt and C4 horizons under pineare compared in Table 2. Although not significantlydifferent at the 0.01 probability level, as previouslymentioned, the means for the archived (4.4%) and Bt(4.0%) samples are significantly greater than the meanfor the C4 (0.8%) samples if a higher probability ofType I error is accepted (P = 0.1). An important con-sideration when comparing the micromorphology of
the archived clods to the 1987 soil fabrics is that, inaddition to the <19-mm-diameter clods, much of thefill material consisted of unaggregated fines and ag-gregates too small to contain voids with channel ar-gillans intact. Thus, the values reported for thearchived clods in Table 2 overestimate the proportionof channel argillans in the fill material as a whole. TheAC and C4 horizons under oak and the BCt and C4horizons under pine all contained <1% channel ar-
Table 2. Quantitative micromorphology of the archived fill-material clods and the 1987 lysimeter soils under oak and pine.fArgillanst
Lysimeter Horizon Depth Plasma Grains Voids Grain Channel Organic
Archived samplesOakOakOakPinePinePineOak + Pine (n = 6)\x ± SD
0-29-16
74-810-22-9
88-96
25.716.510.021.013.517.2
17.3 ± 5.5a
54.146.153.358.648.664.8
54.3 ± 6.8a1987 samples
18.228.525.512.927.96.5
19.9 ± 9.0a
1.53.16.86.32.84.4
4.2 ± 2.1a
0.55.84.41.17.27.2
4.4 ± 3.0a
000000
Oa
OakOakPinePinePinePine
ACC4BAtBt (n = 3);BCtC4 (a = 3)
X ± SD
KS ± SD
7-2065-80
1-1010-2020-3580-100
16.713.716.1
13.3 ± 2.3a11.1
15.6 ± 4.6a
64.266.156.0
59.7 ± 2.6ab62.2
64.3 ± 0.8b
12.313.817.9
15.3 ± O.Sa21.3
12.8 ± 3.8a
5.45.05.6
7.0 ± Mb4.4
6.6 ± 1.3ab
0.50.53.1
4.0 ± O.Sa0.9
0.8 ± 0.3b
0.90.91.3
0.6 + 0.6b0.1Oa
f Within columns, means followed by the same letter are not significantly different at the 0.1 probability level, as determined by Duncan's multiple-range test.No significant differences were found at the 0.01 level.
J Microlaminated typic clay coatings, according to Bullock et al. (1985).
Fig. 1. Thin-section photomicrographs of the BAt horizon (1-10 cm depth) under pine showing an embedded grain argillan on a quartz grain,in (a) plane light and (b) cross-polarized light; and channel illuviation argillans (indicated by arrows) 0:1 a pore wall surface, in (c) planelight and (d) cross-polarized light.

GRAHAM & WOOD: MORPHOLOGIC DEVELOPMENT IN LYSIMETER SOILS 1643
gillans and, in this respect, probably typify fill materialin which illuvial clay has not accumulated. With thisin mind, the Bt horizon developed under pine appar-ently contains more channel argillans than the originalfill material, as well as more than the underlying C4horizon. These results tend to support field evidenceof illuvial clay accumulation in the Bt horizon underpine.
Particle-Size DistributionsThe texture of the original fill material was fine
sandy loam with little variation in particle-size distri-
bution among subsamples. The mean clay content of24 samples of archived parent material (from six filllayers each of the oak, ceanothus, chamise, and pinelysimeters) is 11.0% with a spread of only 1.8 per-centage points between the maximum and the mini-mum values (Table 3). The clay values of the Chorizons sampled in 1987 (Table 4) are within, orslightly higher than, the range of values found for thearchived samples (Table 3). No significant (P = 0.01)differences in clay content were detected among the1987 soil horizons to the 35-cm depth and the archivedsamples, except in the uppermost horizon of the soilsunder oak, ceanothus, and pine (Table 5). The A ho-
Table 3. Mean particle-size distribution of archived parent-material samples from the oak, ceanothus, chamise, and pine lysimeters (n = 24).
XSD
Range
Gravel% (w/w) <whole so
91.66-12
Very coarse Coarsejfil
3.80.8
7.30.5
2.5-5.5 6.3-8.1t Very coarse = 2-1 mm, coarse =JSand =
Table 4.
Horizon
Oi
AACClC2C3C4C5
SandtMedium
i
9.90.5
9.2-10.7
Total*
Fine Very fine
% (w/w) of <2-mm fraction -
18.71.2
17.2-23.5
1-0.5 mm, medium = 0.5-0.25 mm, fine = 0.25-0.12-0.05 mm, silt - 0.05-0.002 mm, clay= <0.002 mm.
Particle-size distributions and bulk densities of the pedons
Depth
cm
(wormcasts)0-77-20
20-3535-5050-6565-8080-100
Gravel
% (w/w) ofwhole soil
0
61010117
109
Very coarse
0.1
2.55.13.63.85.04.74.3
Coarse
3.9
8.08.27.37.18.17.57.9
Sandf
Medium
t
11.0
11.410.510.210.29.89.5
10.1
18.11.1
14.1-19.4
mm, very fine
Sand Silt
57.81.5
31.31.7
52.0-60.1 29.2-38.1
= 0.1-0.05 mm.
Clay
11.00.5
9.8-11.6
sampled to the 100 cm depth, by plant species.
Fine Very fine
Vo (w/w) of <2-mm fraction -
Oak17.8
17.217.419.617.817.918.317.4
15.4
15.217.216.918.316.917.617.7
Sand
48.2
54.358.457.657.257.757.657.4
Total*
Silt
36.0
32.130.431.131.430.931.632.4
Clay
15.7
13.611.211.411.611.410.710.2
— Bulkdensity (SD)
Mg nr3
0.92 (0.60)1.70 (0.06)1.71 (0.01)1.71 (0.01)1.70 (0.03)1.72 (0.04)1.71 (0.03)
CeanothusAlA2ACClC2C3C4C5
0-22-10
10-2020-3535-5050-6565-8080-100
58
108
12111011
3.23.23.64.23.43.73.84.1
8.77.48.68.16.87.37.37.6
10.510.410.69.8
10.29.59.29.8
18.119.218.817.519.518.218.817.7
15.617.616.818.217.318.818.418.3
56.157.858.457.857.257.557.557.5
29.130.729.930.231.231.031.931.7
14.811.511.711.911.711.510.610.8
0.92 (0.13)1.67 (0.06)1.67 (0.03)1.72 (0.02)1.69 (0.03)1.61 (0.08)1.71 (0.01)1.74 (0.02)
ChamiseAlA2ACClC2C3C4C5
ABAtBtBCtClC2C3C4
0-11-77-20
20-3535-5050-6565-8080-100
0-11-10
10-2020-3535-5050-6565-8080-100
489
107777
99
1077687
2.83.65.03.43.13.33.43.9
3.94.64.43.93.84.54.73.7
8.47.57.57.06.66.57.77.3
7.57.78.28.07.37.87.17.8
11.310.29.9
10.59.49.89.99.3
10.910.310.510.29.79.8
10.39.9
18.318.519.318.619.418.719.417.6
Pine19.918.218.017.918.818.017.418.7
16.617.916.918.017.718.217.318.8
18.617.316.516.617.117.418.117.6
57.457.758.657.556.256.557.756.9
60.858.157.656.656.757.557.657.7
31.331.529.831.031.931.631.132.8
30.530.830.431.531.131.231.731.8
11.410.911.611.512.011.811.110.1
8.811.011.911.912.111.410.710.5
1.38 (0.09)1.65 (0.03)1.79 (0.02)1.69 (0.02)1.74 (0.01)1.73 (0.02)1.73 (0.03)1.75 (0.01)
1.08 (0.26)1.75 (0.01)1.76 (0.02)1.72 (0.03)1.74 (0.03)1.74 (0.01)1.72 (0.02)1.80 (0.05)
t Very coarse = 2-1 mm, coarse = 1-0.5 mm, medium = 0.5-0.25 mm, fine = 0.25-0.1 mm, very fine = 0.1-0.05 mm.t Sand = 2-0.05 mm, silt = 0.05-0.002 mm, clay <0.002 mm.

1644 SOIL SCI. SOC. AM. J., VOL. 55, NOVEMBER-DECEMBER 1991
Table 5. Mean clay contents and standard deviations for the 1987soil horizons to the 35-cm depth and for the pooled archived sam-ples (from Table 3).f_____________________
ClayVegetation
Oak
Ceanothus
Chamise
Pine
Archived
Horizon
AACClAlA2ACClAlA2ACClABAtBtBCt
Depthcm0-77-20
20-350-22-10
10-2020-350-11-77-20
20-350-11-10
10-2020-35
X
— —— — <X13.9a11. 7b11.7b13.7a11. Ob11. 4b11. 4b10.3b10.8b11. 2b11. 4b8.5c
11. 4b11. 8b11. 7b11. Ob
SD> ——————
0.30.40.41.30.50.30.51.20.90.50.10.60.50.20.40.5
t All 1987 soil horizons sampled in triplicate (i.e., three separate pedons).Means followed by the same letter are not significantly different at the 0.01probability level, as determined by Duncan's multiple-range test.
rizon under oak and the Al horizon under ceanothuscontained more clay than the underlying horizons andarchived parent materials, whereas the A horizon un-der pine contained less clay.
Graphical comparisons of 1987 clay distributions tothe clay contents of selected original fill layers are pre-sented in Fig. 2. For each lysimeter soil, clay distri-butions are plotted for one pedon sampled to the 100-cm depth, two pedons sampled to the 35-cm depth,and archived samples from six fill layer depths. Nodiscernible textural changes occurred in the C horizonsin the 50 yr since the lysimeters were filled. In contrast,the A horizons under oak, ceanothus, and pine hadclay contents distinctly different from those of the orig-inal fill material.
The A horizons of the soils under oak and ceanothushave higher clay contents (means of 13.9 and 13.7%,respectively; Fig. 2) than their archived parent mate-rials (11-1 and 10.6%, respectively). These increasescoincide with the dominance of worm casts, as notedin Table 1. Worm casts picked from the Oi horizon
8 10 12 14 16.6 8 10 12 14
under oak contained 15.7% clay, as well as more siltand less sand than the C horizons (Table 4) or theoriginal fill material (Table 3). Earthworms preferen-tially ingest clay- and silt-size particles (Lee, 1985).This sorting mechanism may have been responsiblefor concentrating clay in A horizon soil material, be-cause earthworms have brought up clay-enriched ma-terial from below. Furthermore, the binding action oforganic constituents causes dried worm casts to havegreater aggregate stability (Lee, 1985) and to be lessdispersible (Shipitalo and Protz, 1988) than uningestedsoil material. Eluviation of clay is inhibited in A ho-rizons composed of stable worm-cast aggregates (Rus-sell, 1973).
In contrast to the extensively worm-worked soils,the A horizon under pine;, which contained no wormcasts, has been depleted of clay relative to the originalfill material (Fig. 2). Macro- and micromorphologicalobservations suggest an accumulation of illuviatedclay in the 1- to 20-cm depth. Field textures (Table 1)and particle-size distributions (Fig. 2) also indicate aslight clay enrichment in this zone. The absence ofearthworm activity has favored clay translocationfrom the thin (1-cm-thick) A horizon into the BAt andBt horizons. In addition, organic acids may have pro-moted clay dispersion, as demonstrated by Durgin andChaney (1984).
Unlike the other soils, the soil under chamise doesnot exhibit a consistent trend of clay redistributionamong the pedons analyzed (Fig. 2). The variation inclay-distribution patterns may result from the minorand irregularly distributed earthworm activity, whichcould be sufficient to counteract illuviation effects insome pedons but not in cithers.
An alternative hypothesis to explain the depletionof clay in the 0- to 1-cm depth under pine is thatsurface runoff may have preferentially eroded clay-sizeparticles. The lysimeter installation was designed toexclude runoff from the surrounding landscape, soonly rain falling directly on the lysimeters could havecontributed to surface erosion. The pedons sampledfor our study were located within the runoff plot oneach lysimeter. These plots for monitoring runoff mea-sured 2.3 by 4.5 m and had sheet metal (galvanized
Clay, %16 6 8 10 12 14 16 6 8 10 12 14 16
20
o 40
r»01 60Q
80
100
6
OAK CEANOTHUS CHAMISE PINEFig. 2. Clay distribution (weight basis) with depth under the different plant species. For each species, plots are for one pedon sampled to the
100-cm depth, two pedons sampled to the 35-cm depth, and archived parent material from several fill layers. Open circles represent archivedsamples, solid symbols represent 1987 samples.

GRAHAM & WOOD: MORPHOLOGIC DEVELOPMENT IN LYSIMETER SOILS 1645
iron) borders that extended 15 cm above the soil sur-face. Sediment yield from the plots was not measured,but runoff and site-history information provide an in-dication of the possible role of erosion. Photographstaken in 1946 show that all species were densely plant-ed, and Patric (1961b) recorded an original plantingdensity of 10.7 pine seedlings nr2. During the first yearafter planting (1946-1947), runoff on the vegetated lys-imeters ranged from 30 to 48% (pine and oak lysi-meters, respectively) of the total rainfall (703 mm),compared with 69% on bare soil (J.H. Patric, 1974,unpublished data). For the entire period of monitoring(1946-1959), the disposition of total rainfall (7608mm) as runoff was 28% under oak and pine, 26% underceanothus, 16% under chamise, and 66% on bare soil.Photographs of the lysimeter under pine taken duringthe 1950s show accumulated litter on the surface. Pa-tric (196la) noted that, by 1952, complete vegetationcanopies had developed and erosion seldom occurred.Photographs taken immediately after the fire in 1960show that the Coulter pine stand was unburned, thescrub oak was partially burned, and the other specieswere so severely burned that only charred stems stoodabove a bare surface. Although erosional loss of claycannot be ruled put, site history and runoff data in-dicate that the soil under pine has been at least as wellprotected from erosion as the soils under the otherspecies.
Bulk DensityThe lysimeter soils have high bulk densities (X =
1.72 Mg nr3) except in their surface horizons (Table4). Lowest bulk densities are in the soils under oakand ceanothus, where there has been much earthwormactivity producing relatively large and abundant pores(Table 1). The zone of reduced bulk density extendsdeepest under oak, corresponding with its thicker,worm-worked A horizon. Bulk densities of soils underpine (no earthworm activity) and chamise (minorearthworm activity) were decreased only to a depth of1 cm and not to the same degree as under oak andceanothus. Earthworm activity is an important processby which soil bulk density has been reduced.
Implications for Soil GenesisIn the relatively short time since the establishment
of the vegetation sequence, distinct differences in soilmorphology have developed under the four plant spe-cies. Differences in earthworm activity correspondingwith plant species have been largely responsible fordifferentiating the soils. Where present, earthwormspromoted the thickening of dark A horizons and in-creased the clay content of A horizons relative to un-derlying horizons. The effect of earthworms has beengreatest under scrub oak, where a dark, 7-cm-thick Ahorizon has developed (Table 1). Although the soil un-der oak is still an Entisol (coarse-loamy, mixed, mesicTypic Xerorthent), the evolutionary trend is toward thedevelopment of a Mollisol. The same interpretation istrue for the soil under ceanothus, although A horizonthickening has been slower under this species. The de-velopment of argillic horizons in the soils under oakand ceanothus is unlikely because the earthworms pref-
erentially move clay-size material to the surface anddeposit it within stable worm casts, thus counteractingilluviation and inhibiting eluviatipn.
Earthworm activity has been minimal under cham-ise and the A horizon is only 1 cm thick (Table 1).Two of the pedons analyzed have been depleted ofclay near the surface (Fig. 2), but morphologic evi-dence of illuvial accumulation of clay in the subsoilis minimal. Because neither earthworm activity norilluviation dominate, both mpllic epipedon and argil-lic horizon development are impeded. Further devel-opment of subsoil structure may result in a cambichorizon before either mollic or argillic horizons form.Although the soil under chamise is still an Entisol(coarse-loamy, mixed, mesic Typic Xerorthent), its de-velopmental progression is toward an Inceptisol.
The absence of earthworm activity in the soil underpine has allowed clay translocation and its accumu-lation in the subsoil. In each of the three pedons sam-pled, the illuvial horizon contains at least 3%(absolute) more clay than the eluvial horizon (Fig. 2).This clay increase results from eluvial depletion in theA horizon and illuvial accumulation and weatheringin the subsoil. Because the soil under pine contains <15% clay, the subsoil increase in clay satisfies the clay-distribution criterion for an argillic horizon (Soil Sur-vey Staff, 1990). The illuvial horizon further qualifiesas an argillic horizon by being sufficiently thick (>7.5cm thick) and by having >1% oriented clay in cross-section. The soil under pine has an argillic horizon,albeit minimal, and is an Alfisol (coarse-loamy, mixed,mesic Typic Haploxeralf).
Most research has indicated that argillic horizon for-mation is a slow process (Birkeland, 1984). One of thefastest rates of argillic horizon formation has been re-ported by Holliday (1985), who found possible argillichorizons in soils developed in 450-yr-old eolian sed-iments at an archeological site on the southern HighPlains of Texas. More typically, Parsons and Herriman(1976) found an argillic horizon developed within2350 yr in a soil on a low Holocene terrace in theRogue River Valley, Oregon. Our study illustrates thestrong influence of pedoturbation, in this case the re-sult of faunal activity that is plant-species dependent,on the direction and rate of soil development. In theabsence of pedoturbation, it is possible for argillic ho-rizons to form within a matter of decades.
ACKNOWLEDGMENTSThis research was supported in part by the U.S. Forest
Service Pacific Southwest Forest and Range Experiment Sta-tion under Cooperative Agreement PSW-88-0003CA. Theauthors thank Charles Colver, San Dimas Experimental For-est Manager, for access to the lysimeters. Appreciation isalso extended to Bob Tissell for sampling assistance; KenHoltzclaw, Leslee Roistacher, and Jerry Ervin for laboratoryassistance; Wendy Weitkamp, Eileen Kladivko, and SamJames for earthworm identifications; and Paul Zinke formaking the archived samples available.

1646 SOIL SCI. SOC. AM. J., VOL. 55, NOVEMBER-DECEMBER 1991