kinetic characterization of in vitro lead transport across the rat small intestine: mechanism of...
TRANSCRIPT

TOXICOLOGY AND APPLIED PHARMACOLOGY 61, 39-47 ( 198 1)
Kinetic Characterization of in vitro Lead Transport across the Rat Small Intestine
Mechanism of Intestinal Lead Transport
BRUCE J. AUNGST AND HO-LEUNG FUNGI
Department of Pharmaceutics, School of Pharmacy, State University of New York at Buffalo, Amherst, New York 14260
Received February 18. 1981: accepted June 4, 1981
Mechanism of Intestinal Lead Transport. Kinetic Characterization of in Vitro Lead Trans- port across the Rat Small Intestine. AUNGST, B. J., AND FUNG, H.-L. (198 1). Toxicol. Appl. Pharmacol. 61, 39-47. Lead transport through the everted rat small intestine was used as an in vitro model to examine the kinetics and mechanism of gastrointestinal lead absorption. Mucosal-to-serosal lead flux increased nonlinearly with increasing mucosal lead concentrations (0.5-48.3 PM), and an apparent capacity-limited lead flux was observed with intestines of both adult and adolescent rats. Uptake of lead by the adult intestinal tissue, measured at the end of the flux experiments, was also nonlinearly related to mucosal lead concentration. Both intestinal uptake and mucosal-to-serosal flux were reduced by anoxia and by inhibition of glycolysis using fluoride. The data were consistent with a model of lead intestinal transport which included both a carrier-mediated component and passive diffusion. Kinetic parameters for these transport processes were obtained. The relative contribution of passive diffusion to total lead llux increased with increasing lead concentration, but was quantitatively minor (~20%) at all lead concentrations tested. The apparent capacity-limited component of lead flux was not due artifactually to reduced lead solubility in the mucosal solution at high metal concentrations, nor was it due apparently to reduced viability of the intestinal tissue, since lead did not alter 3-Gmethylglucose transport over the range of lead concentration used. When the same lead solution was placed on both sides of an everted intestine, net serosal-to- mucosal lead flux was observed at low lead concentrations. It is therefore likely that intestinal lead transport may be bidirectional, similar to calcium transport.
The identification of impaired intelligence in children with abnormally high concentra- tions of lead in dentine (Needleman et al., 1979) emphasizes the need for limiting ex- posure to environmental lead. Major sources of the body lead burden at normal levels of exposure are food (Mahaffey, 1977; Rabi- nowitz et al., 1977), including specific foods with high concentrations of lead (Settle and
’ To whom correspondence should be addressed: De- partment of Pharmaceutics, School of Pharmacy, 517 Hochstetter Hall, State University of New York at Buf- falo, Amherst, N.Y. 14260.
Patterson, 1980), and water (Thomas et al., 1979). In children, ingestion of dust and soil also contributes significantly to elevated blood lead levels (Roels et al., 1980). Thus, lead toxicity could be prevented, in principle, by inhibiting its absorption through the gas- trointestinal tract. A thorough understand- ing of the mechanism of lead absorption is essential if efforts toward prevention of lead exposure via this approach are to be ratio- nally formulated.
The mechanism of gastrointestinal ab- sorption of lead has not been clearly de- scribed. It has been postulated that lead
39 0041-008X/81/130039-09$02.00/0 Copyright 8 1981 by Academic Press, Inc. All rights of reproduction in any form reserved.

40 AUNGST AND FUNG
shares intestinal absorption sites with cal- cium and iron, because of the apparent changes in lead absorption when dietary cal- cium (Six and Goyer, 1970; Meredith et al., 1977) and iron (Six and Goyer, 1972) are manipulated. Indeed, competitive inhibition of lead binding to mucosal proteins by cal- cium (Barton et al., 1978a) and iron (Barton et al., 1978b) has been demonstrated. The absorption of orally administered lead, like that of calcium and iron, is highest in new- borns (Kostial et al., 1971) and decreases with age (Forbes and Reina, 1972). Thus, it appears likely that absorption of lead through the gastrointestinal tract is regu- lated, and perhaps limited, by a requirement of binding to transport protein(s).
The role of passive diffusion in lead ab- sorption has not been delineated. Meredith et al. (1977) proposed that more than one mechanism may be responsible for lead ab- sorption because calcium, even at very high concentrations in the gut, had a limited com- petitive effect on lead absorption. Based on the differences in absorption kinetics be- tween normal and iron-deficient mice, Flan- agan et al. (1979) also suggested multiple mechanisms for lead absorption. No quan- titative data exist, however, which assess the relative contribution of passive diffusion vs active transport in lead absorption. In this report, we examined this aspect of lead ab- sorption by studying the effect of lead con- centration on its in vitro flux through everted intestines from both adult and adolescent rats. In vitro kinetic parameters for passive and active lead transport were determined.
METHODS
Animals. Male Sprague-Dawley rats (Holtzman Breeders, Madison, Wise.) were used for all experi- ments. One group consisted of adult animals weighing 200-300 g, and another group was 27- to 33-day-old rats which weighed 40-90 g. All rats were housed prior to the experiments in stainless-steel cages in a room with a 12-hr dark, 12-hr light illumination cycle. The diet was Charles River RMH 1000 (Agway). The experi- ments were performed following an overnight fast.
Preparation of isolated intestine. Rates of lead and 3-Qmethylglucose transfer across the rat small intes- tines were determined in vitro using the everted intestine preparation of Crane and Wilson (1958). Rats were decapitated and a single 12-cm-length segment of in- testine was removed from each rat approximately 10 cm from the pylorus. This was rinsed with saline, everted,
and tied at one end, with the other end attached to glass tubing for removal of samples. The intestine was then put into 90 ml of an isotonic Tris buffer (pH 7.4) con- taining 0.2% glucose and a known concentration of lead and/or 3-O-methylglucose. The buffer was kept at 37°C and continually gassed with 95% 0,/5% CO*. A Tris buffer was used instead of the usual buffers containing carbonate or phosphate in order to avoid lead precipi- tation.
3-0-Methylglucoseflux. As an index of the biochem- ical viability of the intestine preparations, flux of 3-O [ “C]methyl-d-glucose (Amersham) was determined in the absence of lead and in the presence of 0.5 and 48.3 &M lead. Intestines from adult rats were prepared as described and placed in Tris buffer containing 0.2% glucose and 0.5 pCi 3-O-[‘4C]methylglucose (specific activity, 58.9 mCi/mmol). Tris buffer placed inside the intestine sacs was free of lead, glucose, and 3-Ometh- ylglucose. The solution inside the intestine sac was re- placed five times during 40 min of incubation. Total radioactivity in the samples removed from the sac was then determined, and the counting efficiency was ob- tained with the addition of [“C]toiuene (Packard) in- ternal standard. 3-0-methylglucose flux was measured in intestines from five rats at each lead concentration.
Lead flux and intestinal lead uptake. Lead flux was measured in intestines from both adult and adolescent rats. The range of lead concentrations in the mucosal bathing medium was from 0.5 to 48.3 NM. One milliliter of a lead-free Tris buffer was placed inside the intestinal sac from the adult rats, while 0.7 ml was put into the sacs from the adolescent rats. The solution inside the sac was removed entirely four or five times during 40 min of incubation and was replaced with lead-free buffer each time. At the lowest mucosal lead concentration used (0.5 PM), only three samples, at 15-min intervals, were removed. The samples were assayed for lead as described later. Intestines from five to nine rats were used at each lead concentration. To examine the effects of anoxia, experiments also were performed in which the mucosal buffer solution was gassed with N2 instead of the Or/CO2 mixture. The effect of metabolic inhi- bition on lead flux was studied using an isotonic Tris buffer containing 0.02 M NaF and 0.2% glucose.
Following the 40-min incubation, each adult rat in- testine was removed from the apparatus and blotted to remove excess buffer. The segment of intestinal tissue was then weighed, minced, and frozen until analysis for lead concentration as described later.
In those experiments in which the buffer within the
LEAD TRANSPORT ACROSS SMALL INTESTINE 41
intestinal sac was removed at regular intervals and re- placed with lead-free buffer, the mucosal-to-serosal lead concentration gradient was maintained, creating a “sink” condition. Lead transport by the rat intestine at a nonsink condition also was investigated. Everted in- testines were prepared as described, but instead of in- serting a lead-free buffer inside the sac, the same lead- containing buffer was placed on both sides of the in- testine. For these experiments a single sample was taken from both the mucosal and serosal buffers after 40 min of incubation, and the lead concentration ratio was cal- culated. Two initial lead concentrations were used for these experiments (2.4 and 48.3 PM), with six animals in each group.
Sample analyses. All glassware was washed with 10% nitric acid prior to use to prevent lead contamination. Lead concentrations in the Tris buffer samples were determined by atomic absorption spectrophotometry (AAS) after diluting the samples with 2 or more vol of 0.1 N nitric acid. Five milliliters of an organic base tissue solubilizer (TS- I, Research Products International) was added to minced intestines. This mixture was then son- icated and the resulting solution, which was free of par- ticulate matter, was diluted to at least 10 ml with toluene and analyzed by AAS. Intestinal lead uptake was cal- culated by subtracting individual endogenous intestinal lead concentrations from those obtained following in- cubation in the lead-containing buffer.
For all analyses, a Perkin-Elmer Model 603 atomic absorption spectrophotometer equipped with an HGA- 2200 graphite furnace and deuterium arc background correction was used. Tbe analytical wavelength was 283.3 nm. The optimum dry, char, and atomization pa- rameters were determined for both the buffer and sol- ubilized intestine matrices. The buffer samples were dried for 70 set at 135”C, charred for 15 set at 400°C. and atomized for 10 set at 2lOO“C. The optimum pro- gram for the solubilixed intestine samples was to dry at 60°C for 60 set, char 30 set at 4OO”C, and atomize 10 set at 2100°C. The graphite tube was purged with NZ, with flow interrupted for 7 set at the start of atomixa- tion. Calibration curves were made daily using blank
buffer or intestine samples spiked with a certified atomic absorption lead standard (4.8 mM lead in dilute nitric acid, Fisher). This standard was also used to prepare the mucosal buffer solutions prior to the flux experi- ments.
RESULTS
Chronic administration of lead to rats im- paired intestinal transport mechanisms for glucose, sodium, glycine, lysine, and phe- nylalanine noncompetitively (Wapnir ef al., 1977). To determine whether short-term
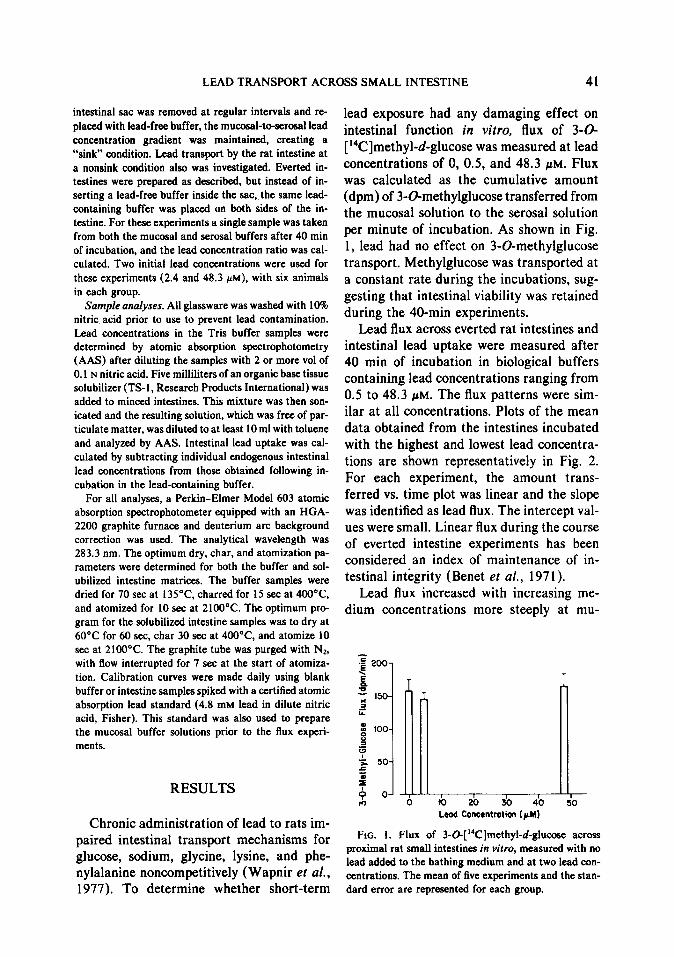
lead exposure had any damaging effect on intestinal function in vitro, flux of 3-G [ “C]methyl-d-glucose was measured at lead concentrations of 0, 0.5, and 48.3 pM. Flux was calculated as the cumulative amount (dpm) of 3-O-methylglucose transferred from the mucosal solution to the serosal solution per minute of incubation. As shown in Fig. 1, lead had no effect on 3Gmethylglucose transport. Methylglucose was transported at a constant rate during the incubations, sug- gesting that intestinal viability was retained during the 40-min experiments.
Lead flux across everted rat intestines and intestinal lead uptake were measured after 40 min of incubation in biological buffers containing lead concentrations ranging from 0.5 to 48.3 pM. The flux patterns were sim- ilar at all concentrations. Plots of the mean data obtained from the intestines incubated with the highest and lowest lead concentra- tions are shown representatively in Fig. 2. For each experiment, the amount trans- ferred vs. time plot was linear and the slope was identified as lead flux. The intercept val- ues were small. Linear flux during the course of everted intestine experiments has been considered an index of maintenance of in- testinal integrity (Benet et al., 197 1).
Lead flux increased with increasing me- dium concentrations more steeply at mu-
I I 1 I 0
10 20 30 40 Lead Concrntrotion (pM)
I 50
FIG. 1. Flux of 3-0-[‘4C]methytd-glucose across proximal rat small intestines in vitro, measured with no lead added to the bathing medium and at two lead con- centrations. The mean of five experiments and the stan- dard error are represented for each group.

42 AUNGST AND FUNG
A is the area of membrane exposed to lead, and h is the thickness of the membrane (Bates and Gibaldi, 1971). If A and h are considered constant and C,,, b C,, Eq. (1) can be simplified to
=KC,,,, (2) passive
0 15 30 45 Time (min)
where K is a constant with units of liters per minute. Flux due to a capacity-limited pro- cess can be represented by
FIG. 2. Results obtained from everted intestine ex-
periments using buffer lead concentrations of 0.5 (0) and 48.3 PM (0), as the mean + SE of six and seven
intestines, respectively. Lead flux was calculated as the regression slope of the amount transferred vs time plot
for each individual intestine.
V max *Gl = K,,, -I- C,,,
(3) capacity-limited
with V,,,,, and K,,, being Michaelis-Menten parameters for the transport system.
cosal lead concentrations below 2.4 PM than those above it. In fact, lead flux at concen- trations above 4.8 PM was only marginally dependent on mucosal lead concentration (Fig. 3). In all the experiments, the lead con- centrations measured in buffer from the mucosal side following 40 min of incubation and after centrifugation were within 90% of the initial concentrations. Thus, the plateau- ing of flux at higher lead concentrations was not due artifactually to precipitation of in- soluble lead salts from the medium bathing the mucosa.
The total flux contributed jointly by both passive diffusion and capacity-limited pro- cesses can then be described using
V max. Cm
tota, = Km + Cm +KC,,,. (4)
The mean data reported on Fig. 3 were fit to Eq. (4) with the nonlinear least-squares computer program NONLIN (Metzler, 1969). The estimates (mean + SD) obtained for the transport kinetic parameters were: K,,,, 0.59 f 0.13 PM; V,,,,,, 1.44 X lo-5 + 7.7 X lo-’ pmol/min; and K, 7 t- 2
The flux data were consistent with a lead transport model in which both capacity-lim- ited and passive processes were involved. The rate of appearance of lead on the serosal side of the membrane due to passive diffusion can be expressed by
2.5-
8 x 2.0-
.f I T i
dx, c-1
= MCln - Cd tA z 0.5
dt hX 1000 ’ (1) passive
4 F I I I I I
0 lo 20 30 40 50 where X and C are amount and concentra- Maeowl Lead Concentration (FM)
tion (nM), respectively, with the subscripts s and m representing serosal and mucosal
FIG. 3. Lead flux across adult rat small intestines
levels, respectively; k is a diffusion constant (mean f SE) measured at various buffer lead concen-
with units of square centimeters per minute, trations. The line was drawn from the parameters ob- tained by computer fitting the data to Eq. (4).

LEAD TRANSPORT ACROSS SMALL INTESTINE 43
TABLE 1
EXPERIMENTAL AND PREDICTED FLUXIS USING EQ. (4). WITH K,,, = 0.59 CM, V,, = 1.44 X 10-s cmol/min, K = 7 x lo-* liters/min AND THE PREDICTED RELATIVE CONTRIBUTIONS OF CARRIER-MEDIATED TRANSPORT
AND PASSIVE DIFFUSION
G (PM)
Measured Prexlicted flux bmol/min) flux (pmol/ AIT
min) Total (7) Passive (P) Active (A) XlOO?G
0.5 5.98 x IO+ 1.2 1.03 x lo-5 2.4 1.19 x lo-s 4.8 1.32 X lo-’
12.1 1.39 x lo-+ 24.1 1.66 x lo-5 36.2 1.60 X lo-’ 48.3 1.80 x lo+
6.50 x lo-+ 9.76 X lo+ 1.17 x 10-S 1.32 X lo-’ 1.46 X lo+ 1.58 X lo+ 1.68 x lo-’ 1.77 x 10-s
3.47 x 10-s 8.74 x 10-s 1.74 x lo-’ 3.49 x lo-’ 8.72 X lo-’ 1.74 x 10-6 2.62 x lO-6 3.49 x 10-6
6.46 X lo+ 99.4 9.68 X 10d 99.2 1.16 X 1O-5 99.1 1.28 X lO-5 97.0 1.37 x lo+ 93.8 1.40 x 10-J 88.6 1.41 x lo+ 83.9 1.42 X 10-s 80.2
X lo-* liters/min. Table 1 gives a compar- ison of the predicted flux, based on these parameters, the measured flux, and the pre- dicted contributions of capacity-limited transport and diffusion to the total flux. Lead transport was effected almost exclu- sively through the active route at low lead concentrations, with increasing relative con- tribution from passive diffusion as the lead concentration was increased. However, even at the highest lead concentration studied, passive diffusion contributed less than 20% to the total flux of lead through the intestine.
Intestinal lead concentrations also were measured in the adult rats at the end of the flux experiments. The tissue concentration is expected to reflect both the buffer/tissue partition coefficient and the extent of tissue binding that might have occurred. Uptake of lead by the intestines increased with in- creasing buffer concentrations, but as was seen with flux, the increases in uptake were not proportional to the increases in lead con- centration (Fig. 4). Because of the irregular shape of the uptake/concentration curve, these data were not suitable for computer fitting to obtain estimates for the binding parameters and partition coefficient. Within any group there was no apparent relation- ship between the intestinal lead content and the measured lead flux.
Because lead absorption in vivo is age de- pendent, the kinetics of intestinal lead trans- port in vitro in young rats were compared to those in adult rats. Due to technical prob- lems associated with the handling of intes- tines from very young rats, the youngest an- imals used for the in vitro work were 27 days old. Forbes and Reina (1972) have shown that in 27-day-old rats lead absorption in vivo was approximately double that of adult rats, with greater interanimal variability than adult rats; absorption also decreased rapidly after 27 days. There were similarities in lead transport between the adult and ad- olescent rats in vitro in that the flux of lead
Mucosal Led comantmtion (pM)
FIG. 4. Intestinal lead accumulation (mean 2 SE) determined after 40 min of incubation in buffers of var- ious lead concentrations. These are the same prepara- tions from which flux was determined (see Fig. 3).

44 AUNGST AND FUNG
TABLE 2
LEAD FLUX ACROSS EVERTED INTE~TINE~ 0~
TAINED FROM AD~LF.SCENT (27- to 33-Day Old) AND
ADULT (200-300 g) RATS AT VARIOUS MUCOSAL ME- DIUM LEAD CONCENTRATIONS (C,)
Adolescent Adult
Cm Flux (xl06 Flux (Xl06 (PM) N ~mol/min) N rmol/min)
0.5 7 9.5 + 2.2 6 6.0 f 0.4 1.2 7 10.3 f 2.3 2.4 6 11.9 f 2.7 4.8 6 11.8 f 1.5 9 13.2 f 2.0
12.1 6 13.9 + 1.1 24.1 4 14.6 + 1.4 9 16.6 + 2.5 36.2 6 16.0 f. 2.5 48.3 5 12.3 f 4.1 7 18.0 f 3.0
Note. The number of animals (IV) and mean + SE are given for each group.
across the adolescent intestines was also ap- parently saturable (Table 2). A direct com- parison of the values of flux in adult vs ad- olescent rat intestine is difficult, however, because different weights of intestines were used, and there were probably differences between these two groups in membrane sur- face area as well. A rough estimate indicated that the surface area of the adolescent in- testine used was approximately 60-70s of that of the adult intestine. If this correction factor is applied to the measured flux values (Table 2), it can be seen that the flux per unit surface area across the adolescent in- testine is, in most cases, greater than that observed for the adult intestine at the same mucosal lead concentration.
One characteristic common to active transport systems is the requirement for metabolic energy. The dependence of intes- tinal uptake and transport of lead on a met- abolic energy supply were investigated using adult rat intestines. Lead flux and intestinal lead concentrations at 40 min were mea- sured as before, using a mucosal buffer lead concentration at 4.8 PM. Anoxia was pro- duced by removing the oxygen supply and
replacing it with nitrogen. Flux was reduced by about 40% under anoxic conditions (Ta- ble 3). The intestinal uptake of lead was like- wise reduced. Similar results were observed in the presence of fluoride, an inhibitor of glycolysis; flux was reduced by about 50% and the mean intestinal concentration of lead appeared to also decrease, albeit only modestly. The dependence of intestinal up- take and flux of lead on metabolic energy suggest then that the saturable component of the flux might be an active transport system.
We also attempted to demonstrate, using adult rat intestines, active lead transport against a concentration gradient by measur- ing serosal-to-mucosal concentration ratios after 40 min of uninterrupted incubation. Lead concentrations initially were equal on both sides of the intestinal membrane. When the lead concentration was 48.3 PM, the 40- min serosal-to-mucosal lead concentration ratio (s/m) was 0.97 5 0.04 (mean + SE, n = 6). However, at a lead concentration of 2.4 PM the final s/m ratio was 0.72 + 0.07 (n = 6) which was significantly different from that at 48.3 PM using an unpaired Stu-
TABLE 3
DEPENDENCE OF INTESTINAL LEAD FLUX AND
UPTAKE ON A METABOLIC ENERGY SUPPLY~
Flux (Xl@ pmollmin)
Intestinal lead uptake
(X 10’ pmol/g)
Control Anoxia by
removal of 02 supply
In presence of 0.02 M NaF
13.2 + 2.0 9.5 + 0.9
8.4 + 0.4 4.6 + 0.4’
6.1 + l.ob 7.5 + 0.8
’ The mucosal lead concentration in each group was 4.8 PM and there were nine animals in the control group and seven in each other group. Data are expressed as mean f SE.
‘p < 0.05 significantly different from the control group using unpaired Student’s f test.

LEAD TRANSPORT ACROSS SMALL INTESTINE 45
dent’s t test. Thus, it appeared that net trans- port of lead in the serosal-to-mucosal direc- tion was also present and this process was also concentration dependent. The intestinal transport mechanism for lead may therefore be bidirectional in nature.
DISCUSSION
In order to understand the mechanism of absorption of lead through the rat gastroin- testinal tract more completely, we have eval- uated the concentration dependency of in- testinal transport and accumulation of lead in vitro. Under circumstances in which in vivo absorption is influenced by many vari- ables (e.g., presence or absence of food, gas- tric emptying rate, GI motility), the everted intestine can be a valuable tool for exam- ining absorptive mechanisms. Our methods differed from the previous in vitro work us- ing everted intestines (Blair et al., 1979) in that the buffer on the serosal side of the in- testine was continually replenished with lead-free buffer in our experiments, creating a “sink” condition. The maintenance of sink conditions was essential for measurement of a true lead flux because serosal-to-mucosal back-transport was identified.
From our measurements of mucosal-to- serosal intestinal lead flux at various lead concentrations, we conclude that flux pro- ceeds by a capacity-limited process in par- allel with passive diffusion. Lead transport was reduced under conditions of anoxia or inhibition of the metabolic energy supply. Using our estimates of transport parameters obtained using adult rat intestines, the rel- ative contributions of active vs passive trans- port can be assessed. At a buffer lead con- centration on the mucosal side of 0.5 PM, the capacity-limited process contributed approx- imately 200 times more to the mucosal-to- serosal lead flux than did diffusion. At a lead concentration of 48.3 pM, diffusion still ac- counted for less than 20% of the flux ac- cording to our model.
The substantial contribution of capacity- limited intestinal lead binding and transport is consistent with evidence obtained by oth- ers implying that the mechanism of intes- tinal lead absorption is similar to, and pos- sibly the same as those which transport the essential metals, calcium and iron. The ac- tivities of the calcium and iron transport sys- tems are regulated by the requirement for maintenance of relatively constant serum concentrations of these minerals. When the diet is deficient in these metals, absorption efficiency improves. Dietary calcium and iron deficiencies have also been associated with elevated tissue lead concentrations, pre- sumably due to improved efficiency of lead absorption.
Another similarity in the way the rat in- testine handles calcium and lead is its ap- parent ability to transport both metals in both directions. In situ experiments by Ghis- han et al. (1980) have demonstrated the ability of the immature rat intestine to transport calcium, at low mucosal calcium concentrations, in a serosal-to-mucosal di- rection. In this study, we identified net ser- osal-to-mucosal lead transport at low lead concentration using adult rat intestines.
The age dependency in lead absorption could be due to greater activity of the trans- port systems (possibly calcium and iron) or greater diffusion, or both. It has also been proposed that in suckling mice intestinal pi- nocytosis contributed significantly to lead absorption (Keller and Doherty, 1980). Al- though we could not estimate kinetic param- eters with our flux data from adolescent (27- to 33-day-old) rats, it would appear that ca- pacity-limited absorption is operative.
The in vitro data we have presented sug- gest that in vivo lead absorption might also be concentration dependent. However, con- flicting evidence exists in the literature re- garding this point. In the in situ work by Barton and Conrad (1978) and Conrad and Barton (1978), the percentage of lead doses absorbed was dependent on the magnitude

46 AUNGST AND FUNG
of the dose. However, Barltrop and Meek (1975) reported that tissue lead concentra- tions increased in proportion to increases in the magnitude of chronic oral doses. Others have reported dose-dependent lead absorp- tion in starved (Garber and Wei, 1974), or iron-deficient (Hamilton, 1978) animals, but not in fed or iron-replete animals. Rigorous bioavailability experiments have not ap- peared in the literature. In a follow-up study which examined blood and tissue lead con- centrations in adult and suckling rats given a broad range of oral and intravenous lead doses, we showed that lead absorption in vivo was indeed dose dependent (Aungst et al., 198 1). If lead absorption in man is simi- larly determined largely by capacity-limited transport, the primary objective in reducing absorption should be to limit the activity of the transport system(s).
ACKNOWLEDGMENT
This work was supported in part by NIH Grant ES01317.
REFERENCES
AUNGST, B. J., DOLCE, J. A., AND FUNG, H.-L. (1981). The effect of dose on the disposition of lead in rats after intravenous and oral administration. Toxicol.
Appl. Pharmacol. 61,48-57. BARLTROP, D., AND MEEK, F. (1975). Absorption of
different lead compounds. Postgrad. Med. J. 51,805- 809.
BARTON, J. C., AND CONRAD, M. E. (1978). Effects of ethanol on absorption and retention of lead. Proc. Sot. Exp. Biol. Med. 159, 213-218.
BARTON, J. C., CONRAD, M. E., HARRISON, L., AND NUBY, S. (1978a). Effects of calcium on the absorp- tion and retention of lead. J. Lnb. Clin. Med. 91,366- 376.
BARTON, J. C., CONRAD, M. E., NUBY, S., AND HAR- RISON, L. (1978b). Effects of iron on the absorption and retention of lead. J. Lab. Clin. Med. 92,536-547.
BATES, T. R., AND GIBALDI, M. (1971). Gastrointes- tinal absorption of drugs. In Current Concepts in the Pharmaceutical Sciences: Biopharmaceutics (J. Swarbrick, ed.). Lea & Febiger, Philadelphia.
BENET, L. Z., ORR, J. M., TURNER, R. H., AND WEBB, H. S. (197 1). Effect of buffer constituents and time on drug transfer across in vitro rat intestine. J. Pharm. Sci. 60, 234-238.
BLAIR, J. A., COLEMAN, 1. P. L., AND HILBURN, M. E. (1979). The transport of the lead cation across the intestinal membrane. J. Physiol. 286, 343-350.
CONRAD, M. E., AND BARTON, J. C. (1978). Factors affecting the absorption and excretion of lead in the rat. Castroenrerology 74, 731-740.
CRANE, R. K., AND WILSON, T. H. (1958). In vitro method for the study of the rate of intestinal absorp- tion of sugars. .I. Appt. Physiol. 12, 145-146.
FLANAGAN, P. R., HAMILTON, D. L., HAIST, J., AND VALBERG, L. S. (1979). Interrelationships between iron and lead absorption in iron-deficient mice. Gas- froenterology 77, 1074- 108 1.
FORBES, G. B., AND REINA, J. C. (1972). Effects of age on gastrointestinal absorption (Fe, Sr, Pb) in the rat. J. Nutr. 102, 647-652.
GARBER, B. T., AND WEI, E. (1974). Influence of di- etary factors on gastrointestinal absorption of lead. Toxicol. Appl. Pharmacol. 27, 685-691.
GHISHAN, F. K.. JENKINS, J. T., AND YOUNOSZAI, M. K. (1980). Maturation of calcium transport in the rat small and large intestine. 1. Nutr. 1%0,1622-1628.
HAMILTON, D. L. (1978). Interrelationships of lead and iron retention in iron-deficient mice. Toxicol. Appl. Pharmacol. 46,65 l-666.
KELLER, C. A., AND DOHERTY, R. A. (1980). Corre- lation between lead retention and intestinal pinocy- tosis in the suckling mouse. Amer. J. Physiol. 239, G114-G122.
KOSTIAL, K., SIMONOVIC, I., AND PISONIC, M. (1971). Lead absorption from the intestine in newborn rats. Nature (London) 233, 564.
MAHAFFEY, K. R. (1977). Quantities of lead producing health effects in humans: Sources and Bioavailability. Environ. Health Perspect. 19, 285-295.
MEREDITH, P. A., MOORE, M. R., AND GOLDBERG, A. (1977). The effect of calcium on lead absorption in rats. Biochem. J. 166, 531-537.
METZLER, C. M. (1969). NONLIN: A Computer Pro gram for Parameter Estimation in Nonlinear Situ- ations, Technical Report 7297/69/7292/005. The Upjohn Co., Kalamazoo, Mich.
NEEDLEMAN, H. L., GUNNOE, C., LEVITON, A., REED, R., PERESIE, H., MAHER, C.. AND BARRETT, P. (1979). Deficits in psychologic and classroom perfor- mance of children with elevated dentine lead levels. N. Engl. J. Med. 300, 689-695.
RABINOWITZ, M. B., WETHERILL, G. W., AND KOPPLE. J. D. (1977). Magnitude of lead intake from respi- ration by normal man. J. Lab. Clin. Med. 90, 238- 248.

LEAD TRANSPORT ACROSS SMALL INTESTINE 47
ROELS, H. A., BUCHET, J.-P., LAUWERYS. R. R., BRUAUX, P., CLAEYS-THOREAU, F., LAFONTAINE, A., AND VERDUYN, G. (1980). Exposure to lead by the oral and pulmonary routes of children living in the vicinity of a primary lead smelter. Environ. Res.
22, 8 l-94. SEITLE, D. M., AND PATTERSON, C. C. (1980). Lead
in albacore: Guide to lead pollution in Americans. Science 207, 1167- 1176.
SIX, K. M., AND GOYER, R. A. (1970). Experimental enhancement of lead toxicity by low dietary calcium. J. Lab. Clin. Med. 76, 933-942.
SIX, K. M., AND GOYER, R. A. (1972). The influence of iron deficiency on tissue content and toxicity of ingested lead in the rat. J. Lob. Clin. Med. 79, 128-
136. THOMAS, H. F., ELWOOD, P. C., WELSBY. E., AND ST.
LEGER, A. S. (1979). Relationship of blood lead in women and children to domestic water lead. Nature (London) 282, 712-713.
WAPNIR, R. A., EXENI, R. A., MCVICAR, M., AND LIFSHITZ, F. (1977). Experimental lead poisoning and intestinal transport of glucose, amino acids, and so- dium. Pediar. Res. 11, 153-157.