intracellular cargo transport: molecular motors playing tug-of-war
TRANSCRIPT

Intracellular cargo transport:Molecular motors playing
tug-of-war
Melanie J.I. Müller,Stefan Klumpp, Reinhard Lipowsky
Department of Theory & Bio-Systems
Max Planck Institute for Colloids and Interfaces

Outline
1) Intracellular cargo transport & molecular motors
3) Why such weird motion?
2) Motors playing tug-of-wara) fair playb) unfair

1) Intracellular cargo transport& molecular motors
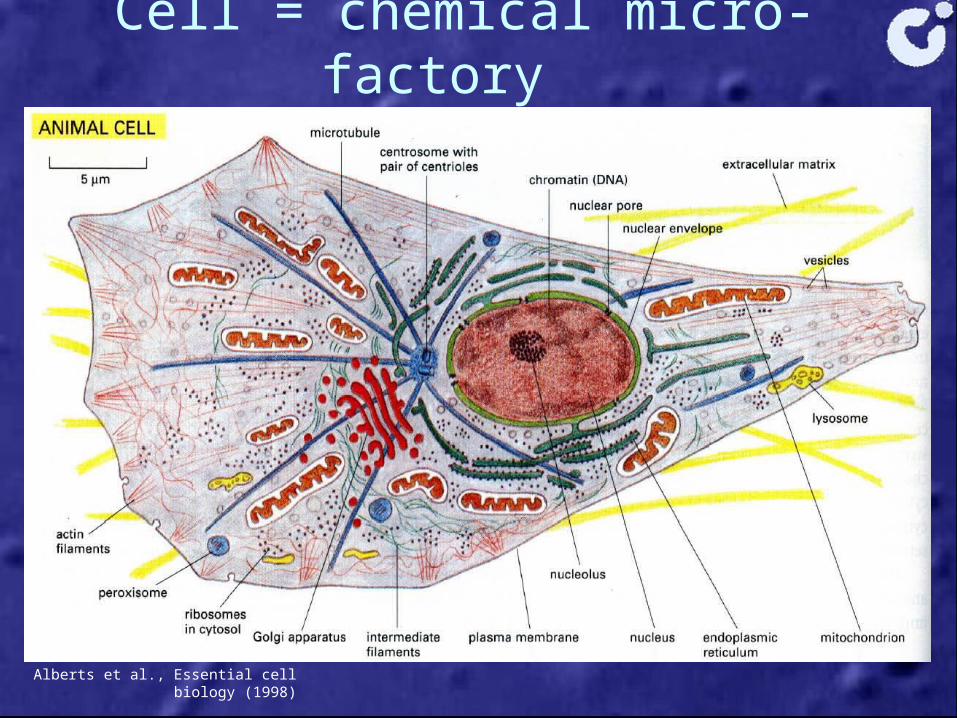
Cell = chemical micro-factory
Alberts et al., Essential cell biology (1998)

Molecular motors• Molecular motors
= nanotrucks - Roads: filaments - Fuel: ATP - Cargo: vesicles, organelles…
Travis, Science 261:1112 (1993) www.herculesvanlines.com (2008)
www.inetnebr.com/stuart/ja (2008)
• Nanoscale → Stochastic (Brownian) motion→ Unbinding from filament ('fly') after ~ 1 μm
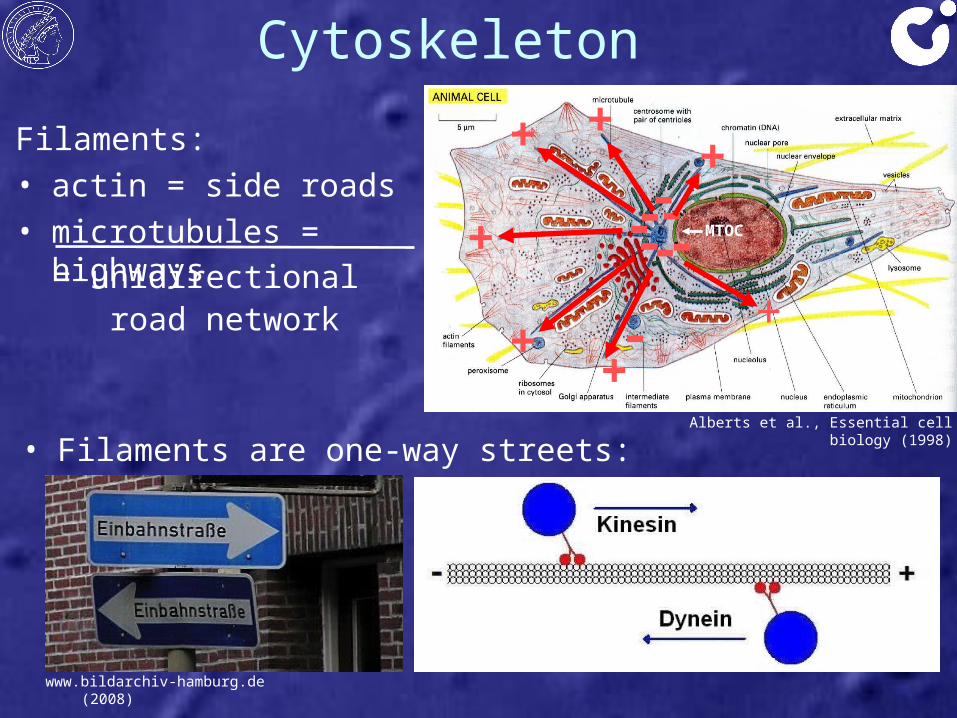
Cytoskeleton
MTOC+
+ +
+ +
+
+----
-
-- -
Alberts et al., Essential cell biology (1998)
• Filaments are one-way streets:
Filaments:• actin = side roads• microtubules = highways
= unidirectional road network
www.bildarchiv-hamburg.de (2008)
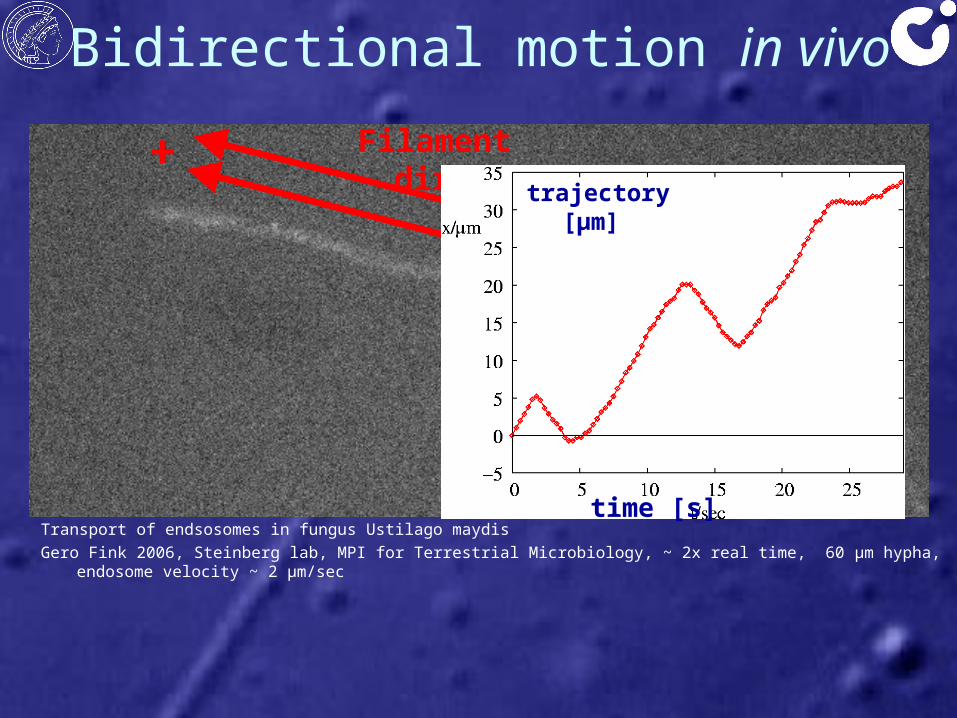
Bidirectional motion in vivo
Transport of endsosomes in fungus Ustilago maydisGero Fink 2006, Steinberg lab, MPI for Terrestrial Microbiology, ~ 2x real time, 60 μm hypha, endosome velocity ~ 2 μm/sec
+
_
Filament direction
time [s]
trajectory [μm]

Bidirectional motion• Bidirectional motion on
unidirectional filaments→ plus and minus motors on one cargo
trajectory [μm]
time [s]
Gero Fink, MPI for Terrestristrial Mikcrobiology (2006)
Ashkin et al., Nature 348: 346 (1990)
0.1 μm
• Teams of 1-10 motors

Why?
• Why bidirectional motion? → later
• How does it work?Why no blockade?
trajectory [μm]
time [s]
~ 2 μm/sas for one species alone

Coordination
• Hypothetical coordination complex
Coordination complex
• mechanical interaction• Tug-of-war model:
- model for single motor- mechanical interaction
or tug-of-war?
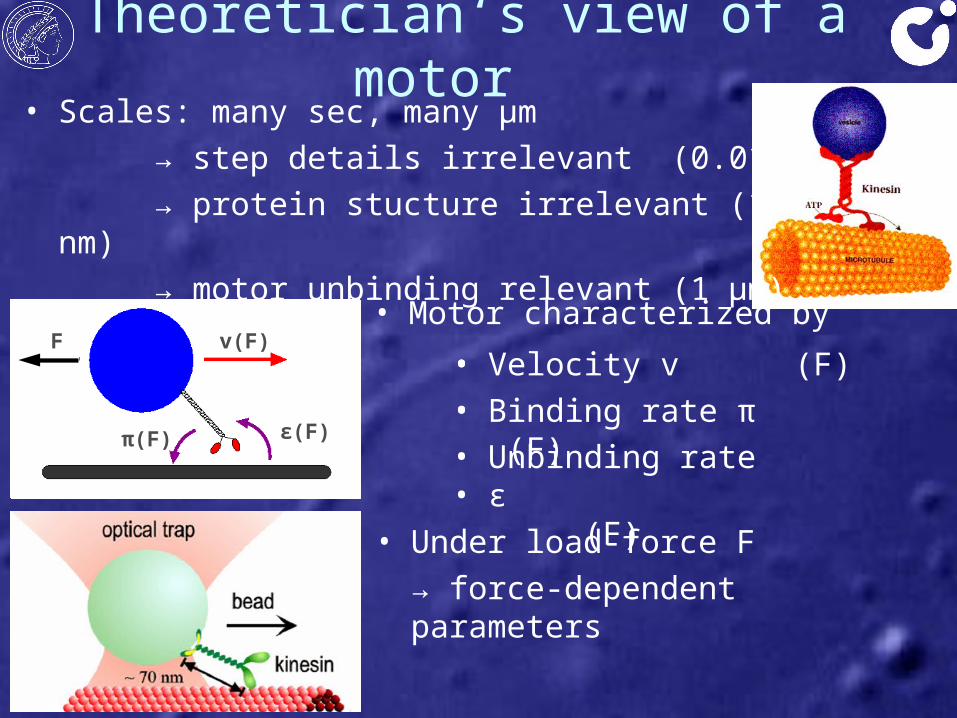
v
π ε
• Velocity v• Binding rate π• Unbinding rate ε
Theoretician‘s view of a motor
• Motor characterized byv(F)F
π(F) ε(F)
• Under load-force F → force-dependent parameters
• (F)• (F)• (F)
• Scales: many sec, many μm → step details irrelevant (0.01 s)→ protein stucture irrelevant (100 nm)→ motor unbinding relevant (1 μm)
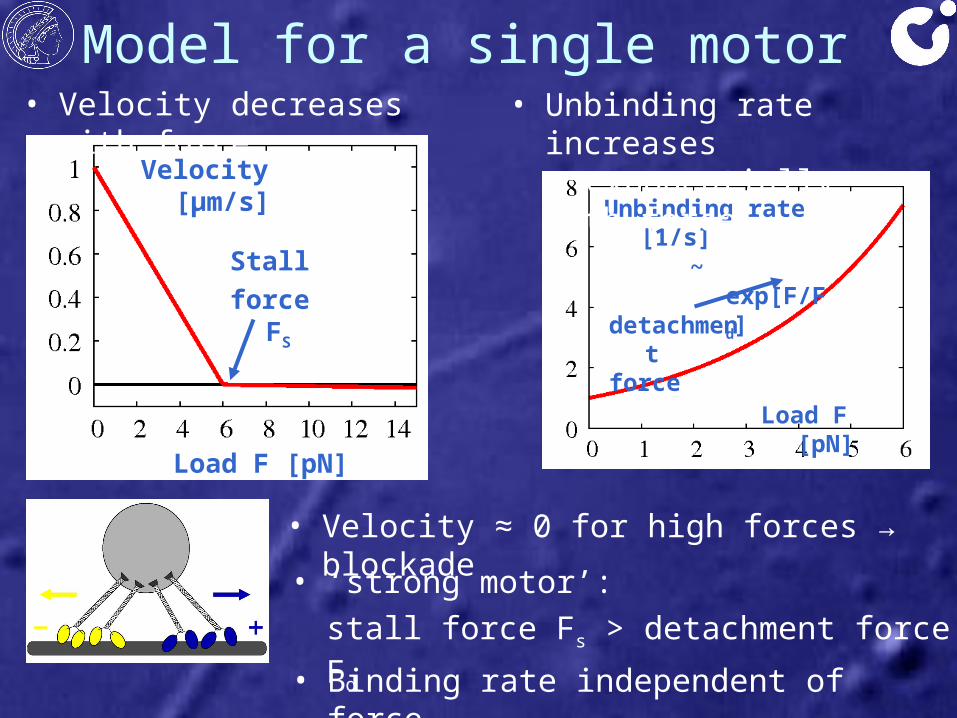
• Velocity decreases with force
Model for a single motor
Velocity [μm/s]
Load F [pN]
Stall force FS
• Velocity ≈ 0 for high forces → blockade
Load F [pN]
Unbinding rate [1/s]
~ exp[F/Fd]
detachment force
• Unbinding rate increases exponentially with force
• Binding rate independent of force
• ‘strong motor’: stall force Fs > detachment force Fd

2) Molecular motorsplaying
tug-of-war

Tug-of-war model
v(F)F
π ε(F)
Single motors with rates from single molecule experiments
• Opposing motors → load force• Motors of one team share force
• Forces determined by:- Balance of motor forces (+ cargo friction + external force)- All motors move with one velocity

• Master equation• Observation time sec - min → stationary state• Analysis: numerical calculations, simulations,
analytical approximations
Tug-of-war model

Types of motion
Motors block each other → no motion
Minus motors win → motion to minus
end
Plus motors win → motion to plus end
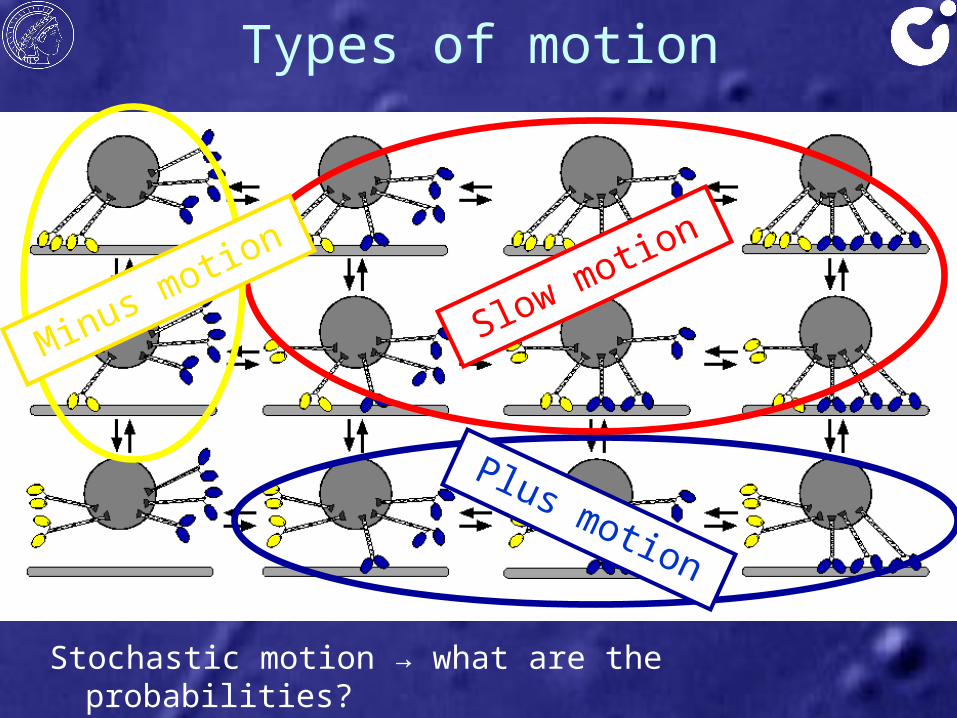
Types of motion
Stochastic motion → what are the probabilities?
Plus motion
Slow motion
Minus motion
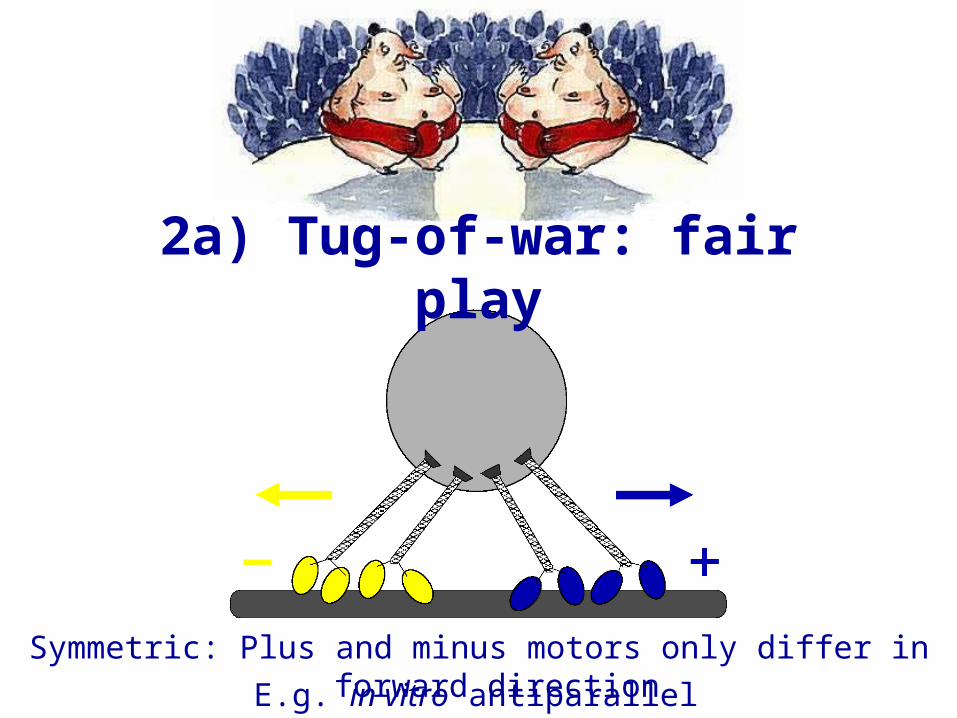
Symmetric: Plus and minus motors only differ in forward directionE.g. in vitro antiparallel microtubules
2a) Tug-of-war: fair play

Weak motors• Weak motors := exert less force than they can sustain stall force Fs < detachment force Fd
• Motors don’t feel each other→ random binding and unbinding
x x

Weak motors• highest probability for same number of bound motors
(0,0)
• blockade!
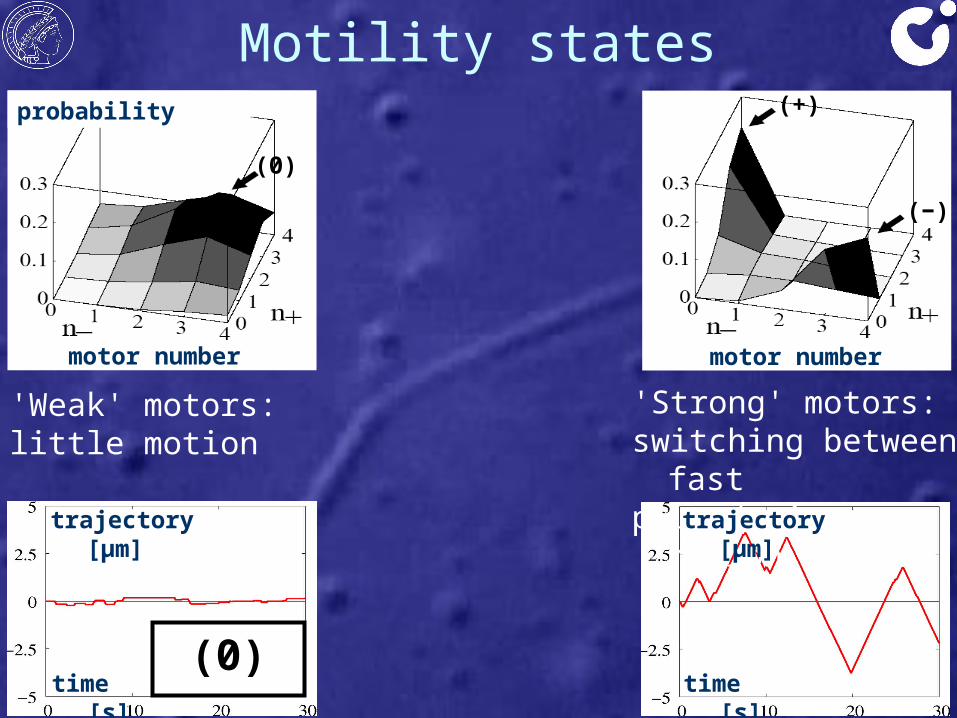
'Strong' motors: switching between fastplus / minus motion
'Weak' motors:little motion
motor number
trajectory [μm]
time [s]
(−)
(+)
(0)
motor number
probability
(0)
Motility states
trajectory [μm]
time [s]
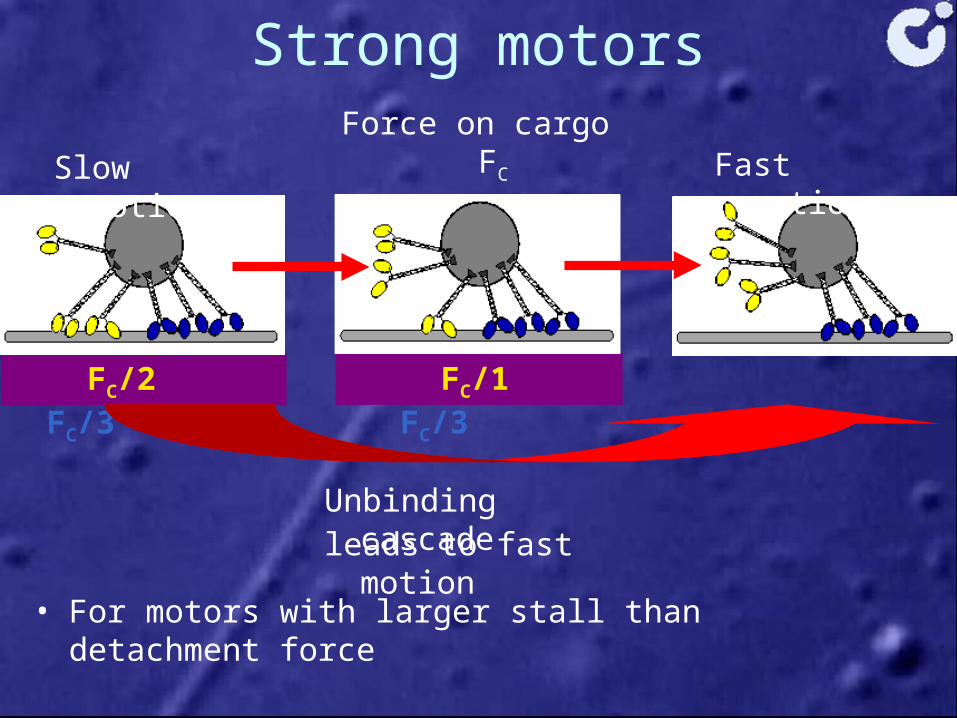
Strong motors
• For motors with larger stall than detachment force
Force on cargo FC
FC/2 FC/3 FC/1 FC/3
Slow motion Fast motion
Unbinding cascade leads to fast motion
Strong motors• Unbinding cascade → only one team remains bound
(0,0)
• Unbinding cascade
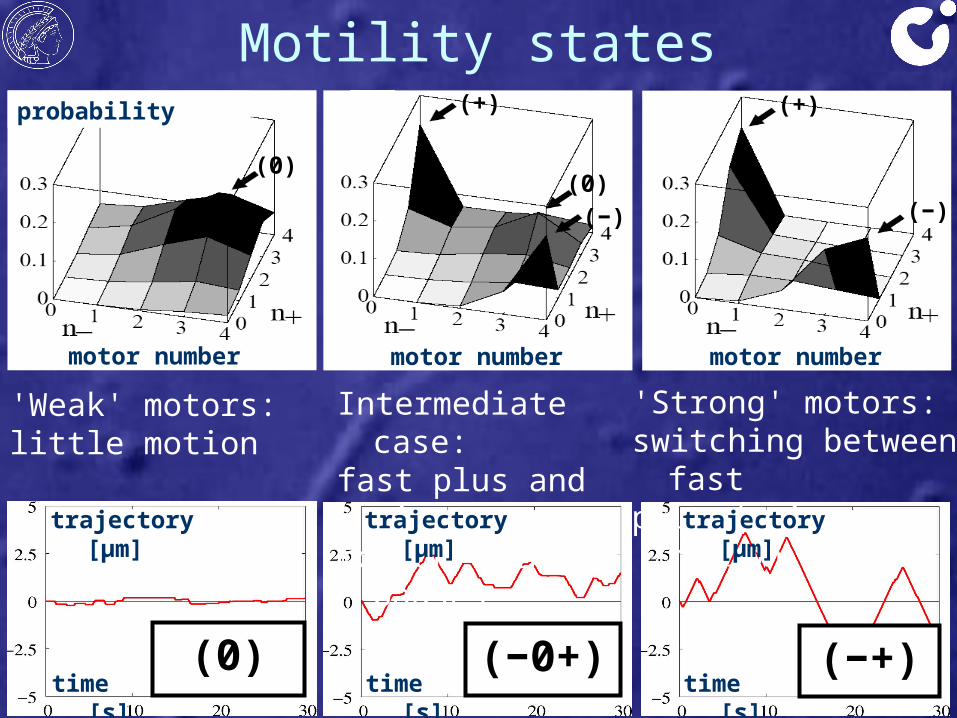
'Strong' motors: switching between fastplus / minus motion
Intermediate case:fast plus and minusmotion with pauses
'Weak' motors:little motion
motor number
trajectory [μm]
time [s]
(−)
(+)
(0)
(−)
motor numbermotor number
probability
(0)
(+)
(0) (−+)(−0+)
Motility states
trajectory [μm]
time [s]
trajectory [μm]
time [s]
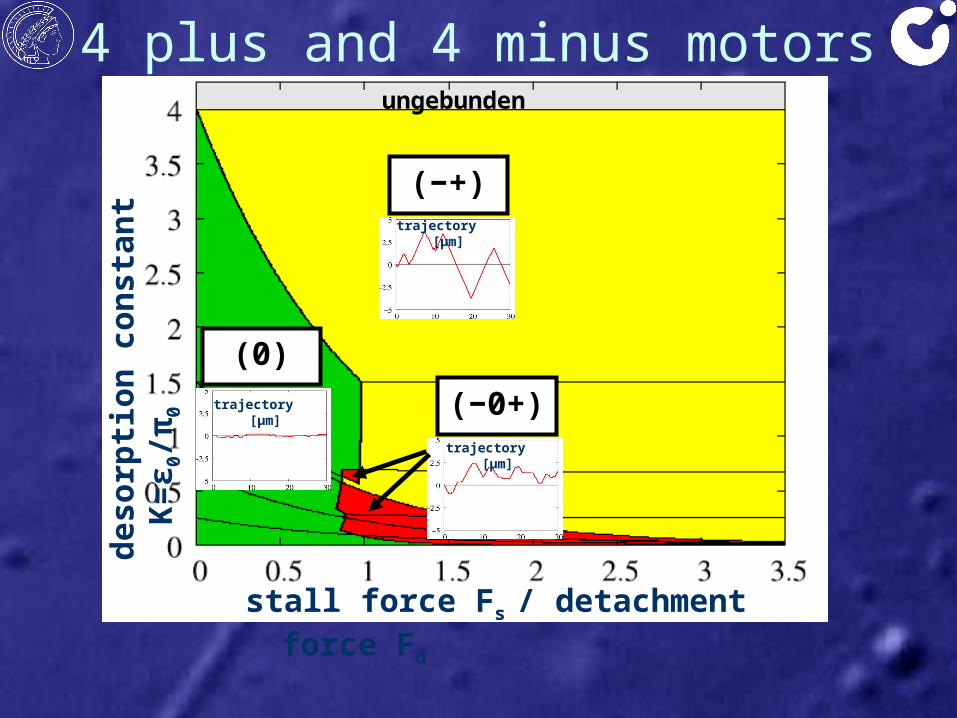
4 plus and 4 minus motors
zz
deso
rptio
n co
nsta
nt K
=ε0/π
0
stall force Fs / detachment force Fd
ungebunden
(0)
(−+)
(−0+)trajectory [μm]
trajectory [μm]
trajectory [μm]

Weak motors‘Weak' motors:stall force Fs < detachment force Fd
→ motors don’t feel each other→ analytical solution
deso
rptio
n co
nsta
nt K
=ε0/π
0
(0)
(−+)
stall force Fs / detachment force Fd
→ slow motion (0)
(−0+)

Strong motors
Unbindingcascade
→ fast bidirectional motion (−+)
deso
rptio
n co
nsta
nt K
=ε0/π
0
(0)
(−+)
stall force Fs / detachment force Fd
(−0+)
‘Strong' motors:stall force Fs > detachment force Fd
Strong motors• Unbinding cascade → 1D random walk → exact solution
(0,0)
• Unbinding cascade
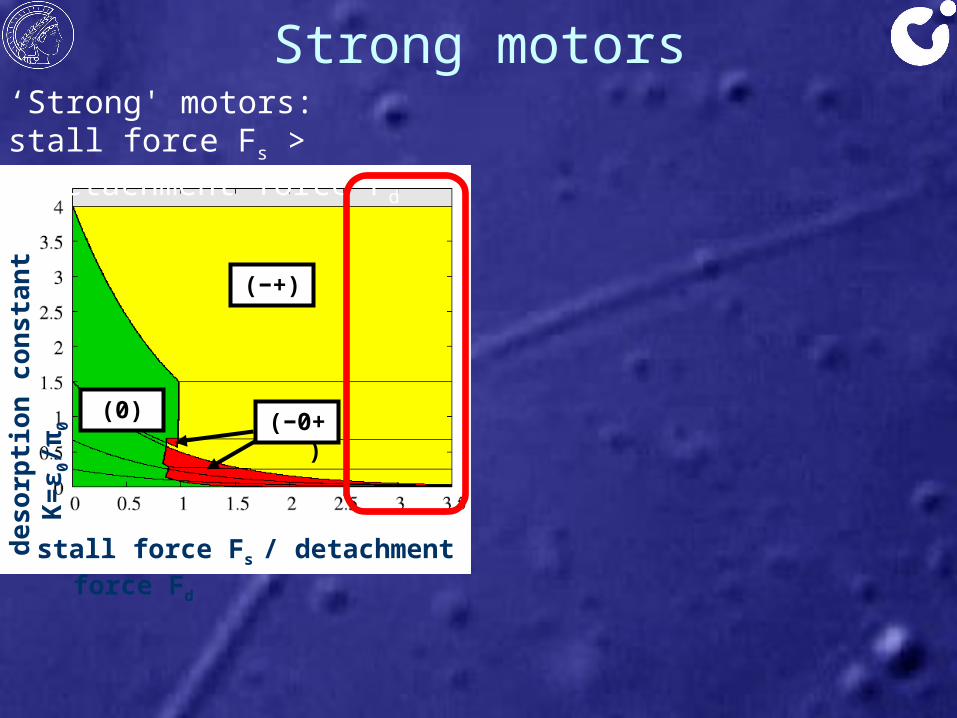
Strong motorsde
sorp
tion
cons
tant
K=ε
0/π0
(0)
(−+)
stall force Fs / detachment force Fd
(−0+)
‘Strong' motors:stall force Fs > detachment force Fd
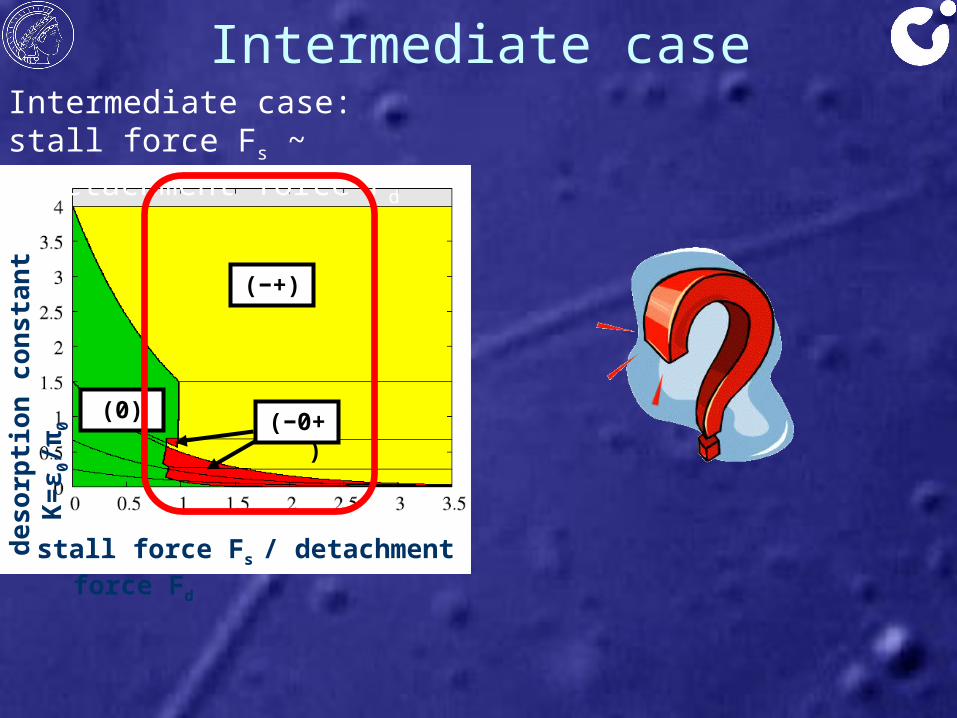
Intermediate casede
sorp
tion
cons
tant
K=ε
0/π0
(0)
(−+)
stall force Fs / detachment force Fd
(−0+)
Intermediate case:stall force Fs ~ detachment force Fd
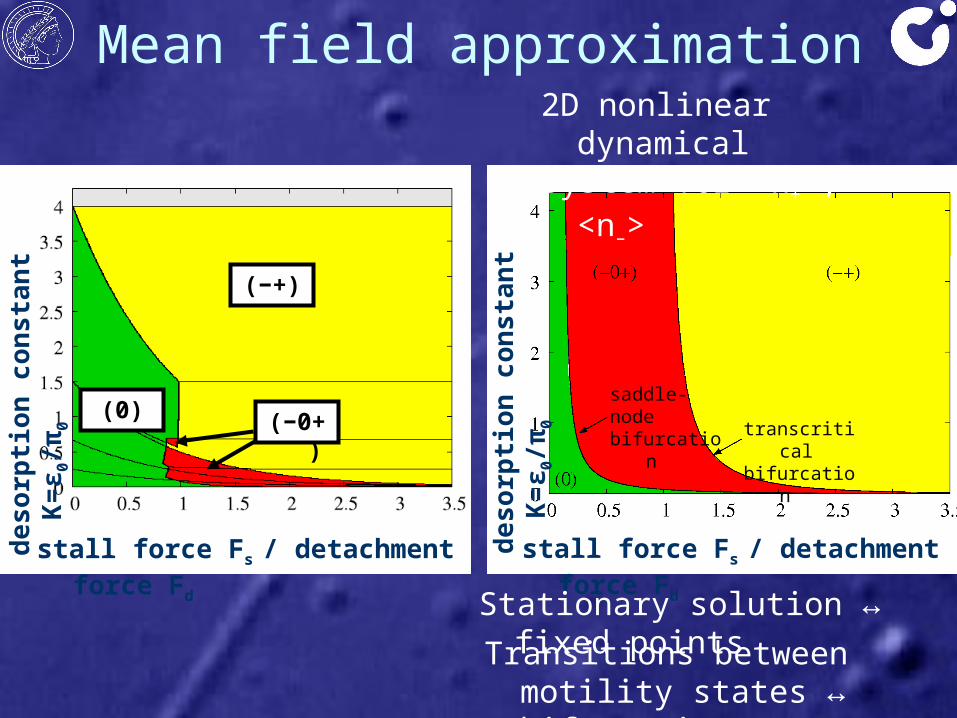
Mean field approximationde
sorp
tion
cons
tant
K=ε
0/π0
(0)
(−+)
stall force Fs / detachment force Fd
(−0+)
deso
rptio
n co
nsta
nt K
=ε0/π
0
stall force Fs / detachment force Fd
Stationary solution ↔ fixed pointsTransitions between motility
states ↔ bifurcations
saddle-nodebifurcation transcritical
bifurcation
2D nonlinear dynamical system for <n+>, <n–>
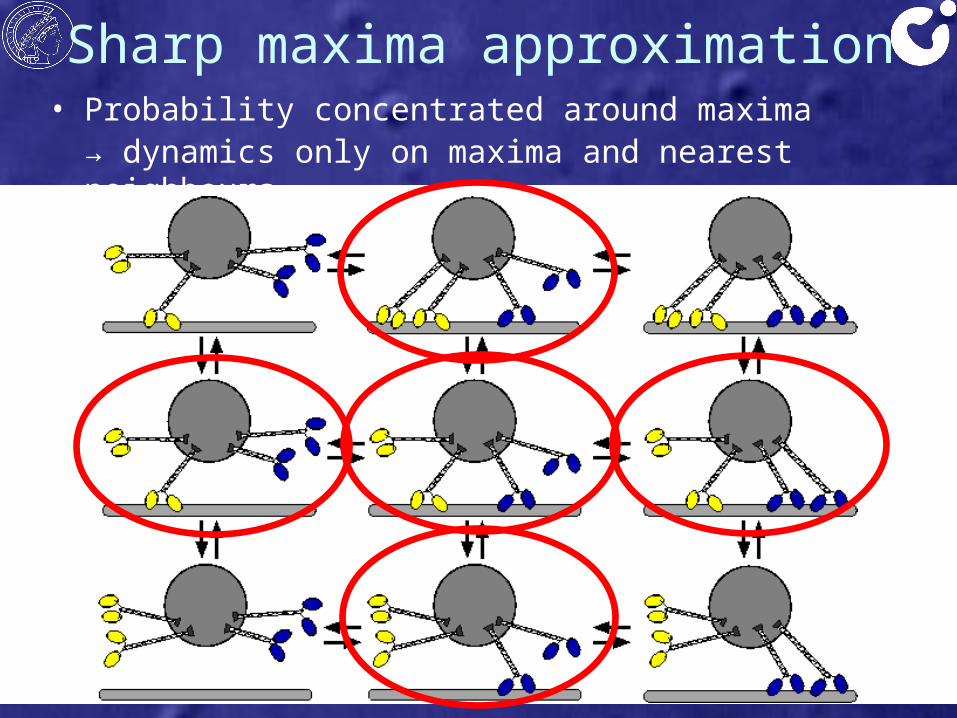
Sharp maxima approximation• Probability concentrated around maxima
→ dynamics only on maxima and nearest neighbours
(0,0)
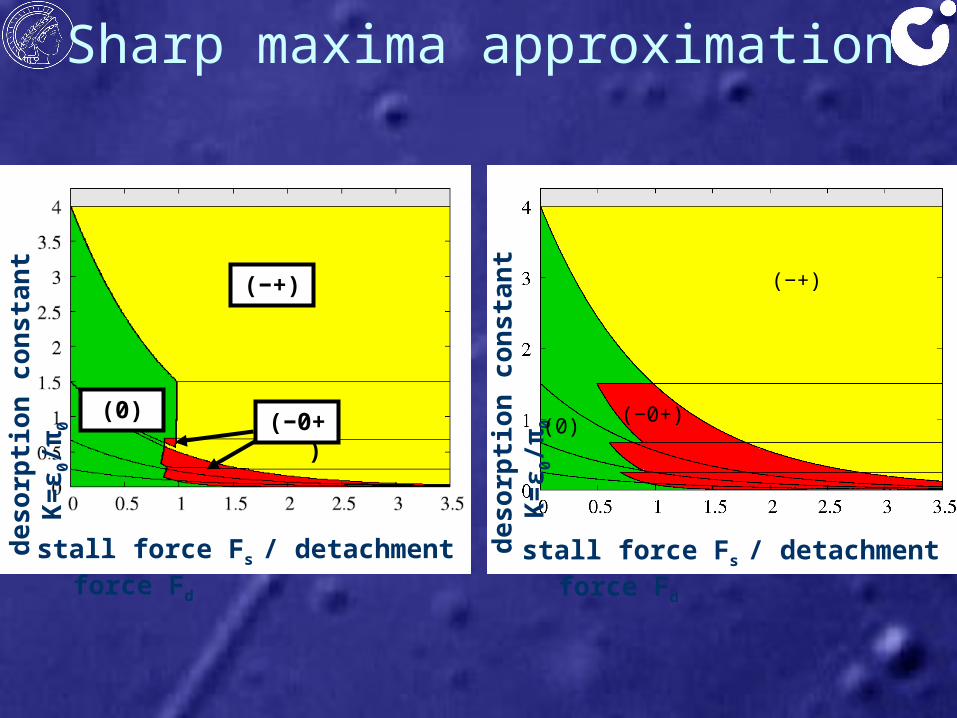
Sharp maxima approximationde
sorp
tion
cons
tant
K=ε
0/π0
(0)
(−+)
stall force Fs / detachment force Fd
(−0+)
deso
rptio
n co
nsta
nt K
=ε0/π
0
(−0+)
(−+)
(0)
stall force Fs / detachment force Fd

Tug-of-war animation
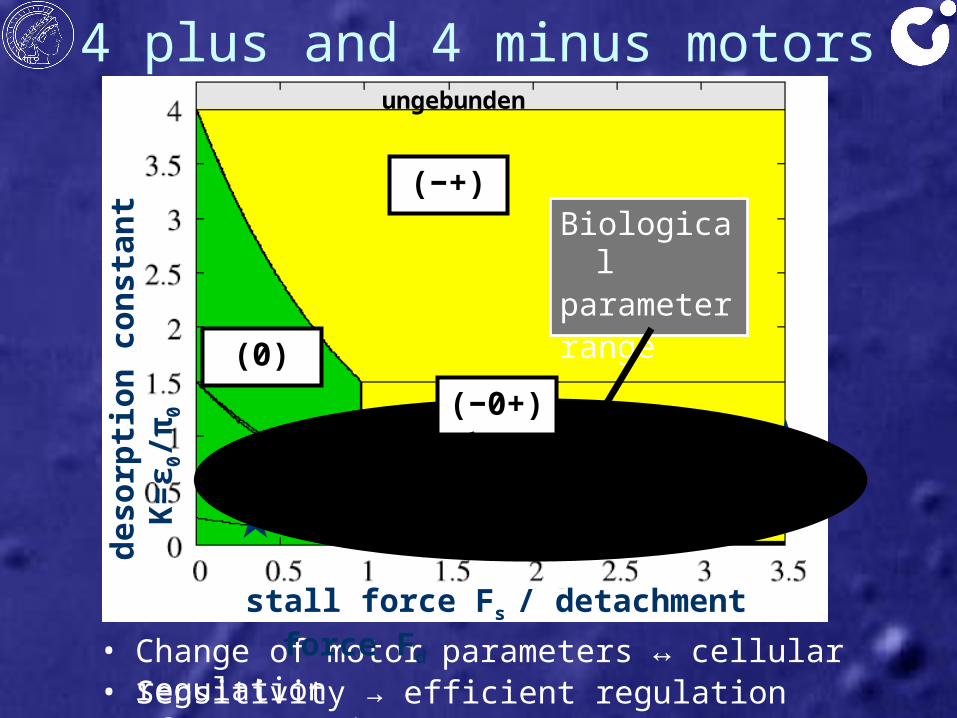
4 plus and 4 minus motors
• Change of motor parameters ↔ cellular regulation
zz
deso
rptio
n co
nsta
nt K
=ε0/π
0
(0)
stall force Fs / detachment force Fd
ungebunden
Kin1cDyn cDynKin2 Kin3
Kin5
• Sensitivity → efficient regulation of cago motion
Biological parameterrange
(−0+)
(−+)
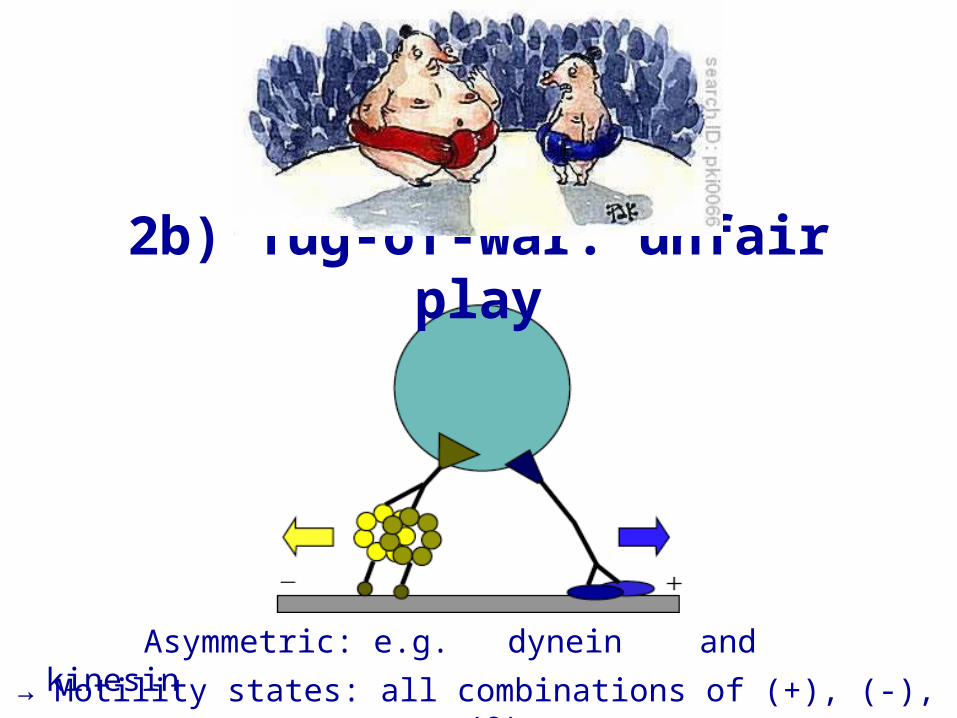
Asymmetric: e.g. dynein and kinesin→ Motility states: all combinations of (+), (-), (0)
2b) Tug-of-war: unfair play
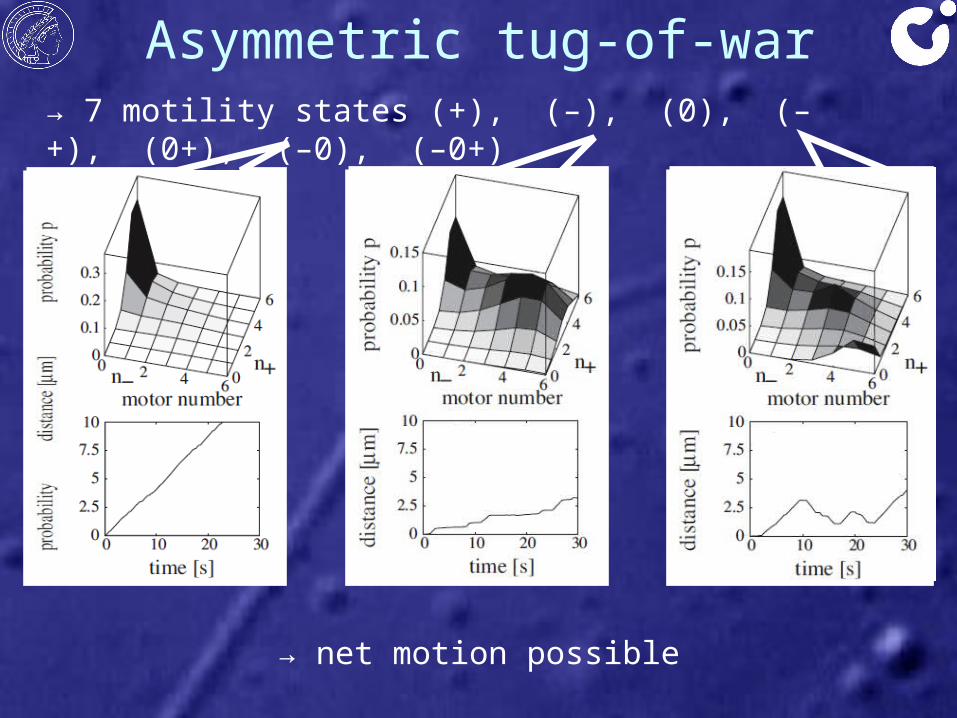
Asymmetric tug-of-war→ 7 motility states (+), (–), (0), (–+), (0+), (–0), (–0+)
→ net motion possible

Comparison to experiment
• Slow motion (blockade)
Experiment:Fast motion in each direction
• Why people didn’t believe in a tug-of-war before:
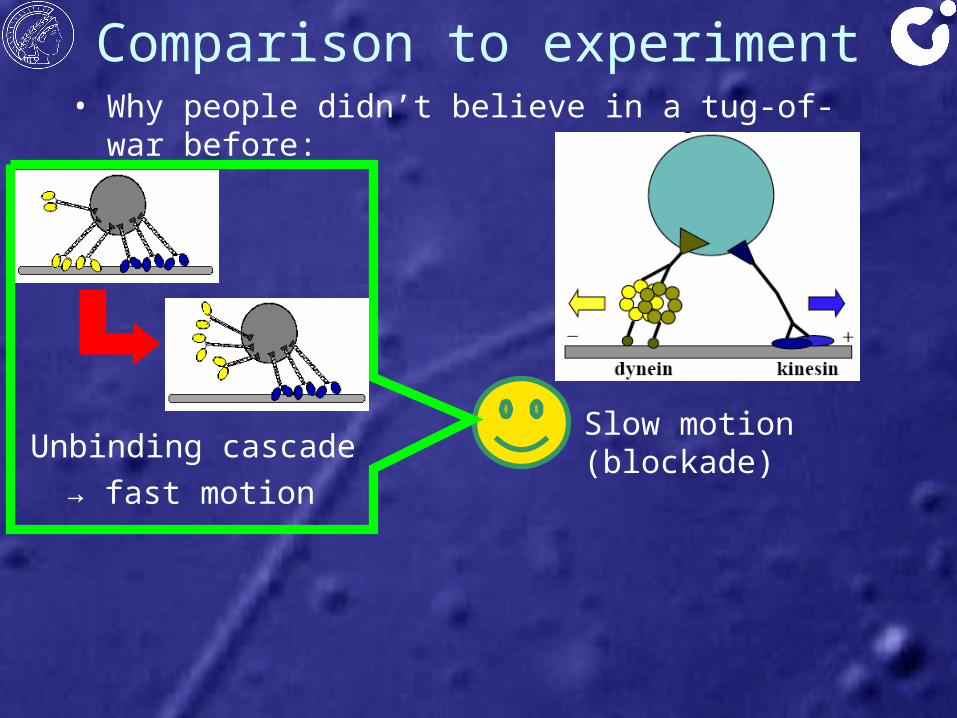
• Slow motion (blockade)Unbinding cascade
→ fast motion
Comparison to experiment• Why people didn’t believe in a tug-of-war before:
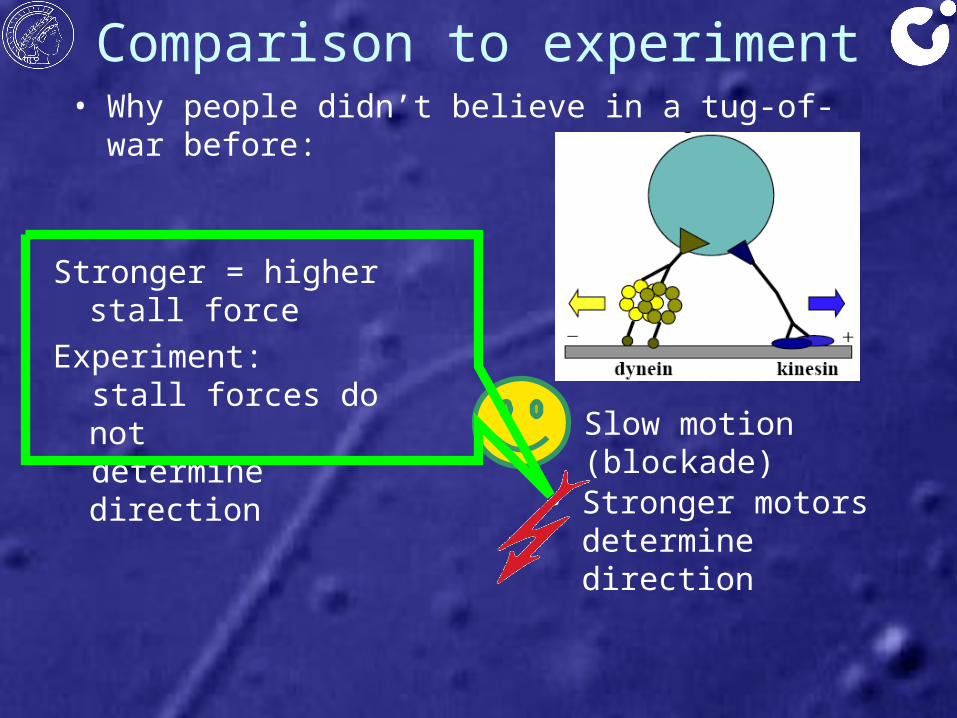
• Slow motion (blockade)
• Stronger motors determine direction
Stronger = higher stall forceExperiment: stall forces do not determine direction
Comparison to experiment• Why people didn’t believe in a tug-of-war before:

• Slow motion (blockade)
• Stronger motors determine direction
→ 'Stronger' can mean- generate larger force- bind stronger to filament- resist pulling force better
→ direction not only determined by (stall) forces
Comparison to experiment• Why people didn’t believe in a tug-of-war before:
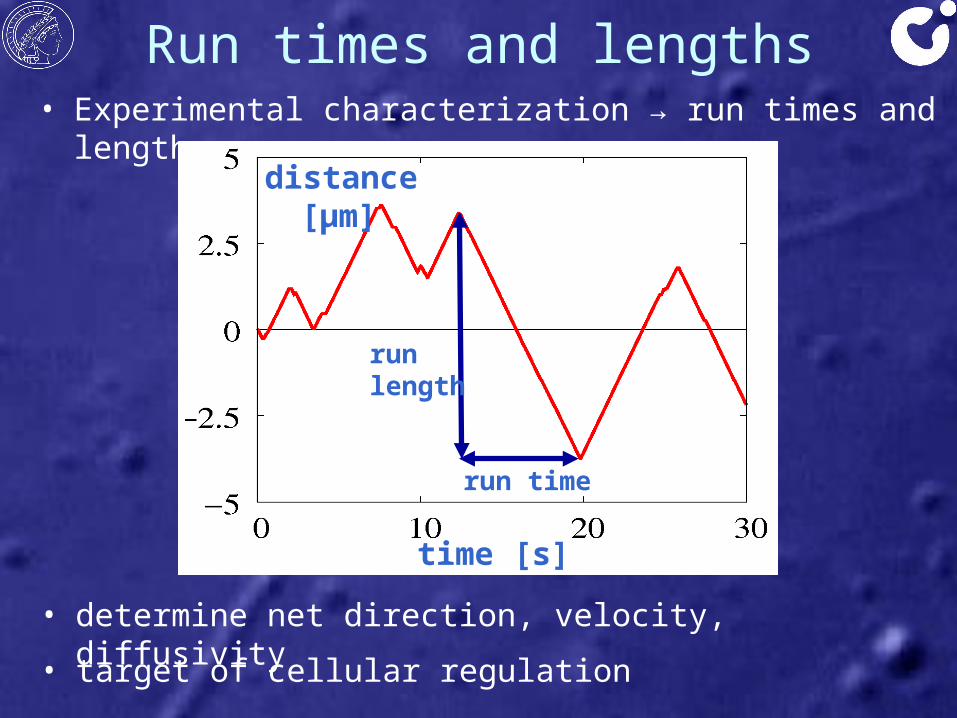
Run times and lengths• Experimental characterization → run times and lengths
distance [μm]
time [s]
runlength
run time
• determine net direction, velocity, diffusivity
• target of cellular regulation

• Slow motion (blockade)
• Stronger motors determine direction
• Impairing one direction enhances the other
→ impairing one direction can have various effects
plus minuscellular regulation ↓ ─dynein mutations ↓ ↓kinesin mutation ↓ ↑
Comparison to experiment• Why people didn’t believe in a tug-of-war before:

Regulation and mutation
• Dynein mutation = changing dynein properties
• Examples:- increase dynein's unbinding rate
→ minus runs, plus runs
• Cellular regulation = changing motor properties
• Change one parameter → impair / enhance• Change several parameters
→ various effects of changing many properties
shorter
longer
longer
shorter- increase dynein's resistance to force
→ minus runs, plus runs

• Slow motion (blockade)
• Stronger motors determine direction
• Impairing one direction enhances the other
Impairing one direction(regulation / mutation)can have various effectson the other direction
Comparison to experiment• Why people didn’t believe in a tug-of-war before:
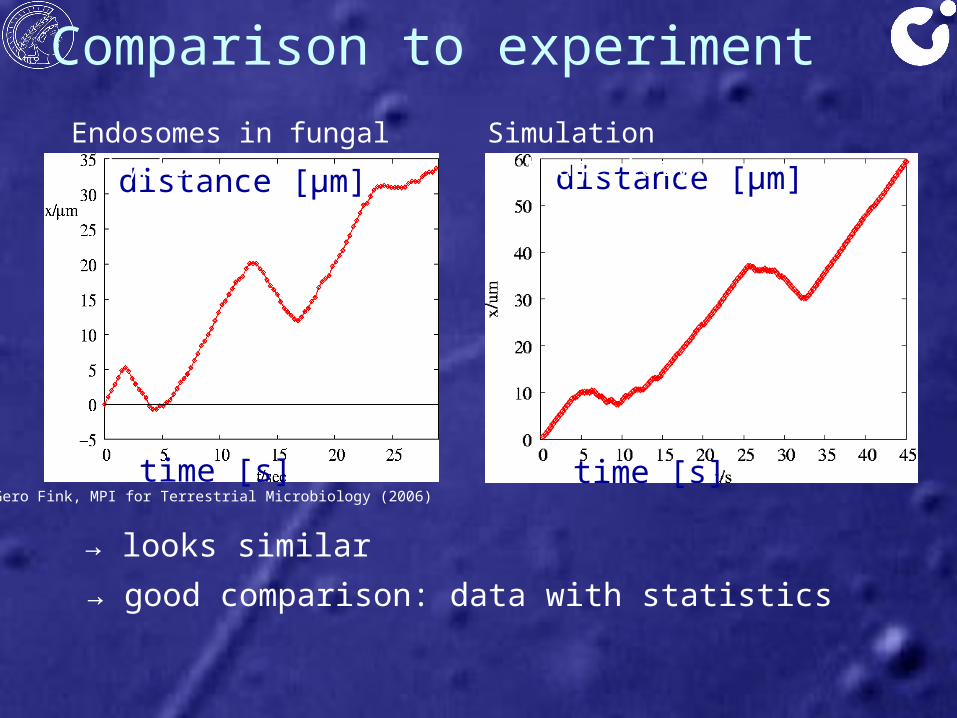
Comparison to experiment
Gero Fink, MPI for Terrestrial Microbiology (2006)time [s]
distance [μm]Endosomes in fungal hypha:
time [s]
distance [μm]Simulation trajectory:
→ looks similar→ good comparison: data with statistics

Comparison to experiment• Bidirectional transport
of lipid-droplets in Drosophila embryos
trajectory [nm]
time [s]
Gross et al., J. Cell Biol. 148:945 (2000)quest.nasa.gov/projects/flies/LifeCycle.html
• Data from Gross lab (UC Irvine):
- Statistics on run lengths, velocities, stall forces
- effect of cellular regulation (2 embryonic phases)
- effect of 3 dynein mutations
→ Tug-of-war reproduces experimental data within 10 %
→ no coordination complex necessary

3) Why

Why bidirectional motion?
Why instead of ?
• Search for target• Error correction• Avoid obstacles• Cargos without destination• Easy and fast regulation
• Bidirectional transport of cargo and motors
Why instead of ?

SummaryBidirectional transport as tug-of-war of molecular motors
• simple model, but complex and cooperative motility
• fast bidirectional motion ‘despite’ tug-of-war
• complex parameter-dependence→ efficient regulation of motility
• consistent with in vivo data

Thank you
Yan Chai
Stefan Klumpp
Janina BeegChristian
KornSteffen
Liepelt
Thank youfor your attention!
Reinhard Lipowsky