identification and typing of vibrio anguillarum: a
TRANSCRIPT

General rights Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights.
Users may download and print one copy of any publication from the public portal for the purpose of private study or research.
You may not further distribute the material or use it for any profit-making activity or commercial gain
You may freely distribute the URL identifying the publication in the public portal If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim.
Downloaded from orbit.dtu.dk on: Feb 07, 2022
Identification and typing of Vibrio anguillarum: A comparison of different methods.
Austin, B.; Alsina, M.; Austin, D. A.; Blanch, A. R.; Grimont, F.; Grimont, P. A. D.; Jofre, J.; Koblavi, S.;Larsen, J. L.; Pedersen, KarlTotal number of authors:13
Published in:Systematic and Applied Microbiology
Link to article, DOI:10.1016/S0723-2020(11)80400-5
Publication date:1995
Document VersionPublisher's PDF, also known as Version of record
Link back to DTU Orbit
Citation (APA):Austin, B., Alsina, M., Austin, D. A., Blanch, A. R., Grimont, F., Grimont, P. A. D., Jofre, J., Koblavi, S., Larsen, J.L., Pedersen, K., Tiainen, T., Verdonck, L., & Swings, J. (1995). Identification and typing of Vibrio anguillarum: Acomparison of different methods. Systematic and Applied Microbiology, 18(2), 285-302.https://doi.org/10.1016/S0723-2020(11)80400-5

System. App!. !--.1icrobio!' 18,285-302 (1995) © Gustav Fischer Verlag, Stuttgart· Jena . New York
Identification and Typing of Vibrio anguillarum: A Comparison of Different Methods
B. AUSTIN" M. ALSINAb, D. A. AUSTIN', A. R. BLANCHh, F. GRIMONT", P. A. D. GRIMONTc, J. JOFREb ,
S. KOBLAVIc, J. L. LARSENd
, K. PEDERSENd, T. TIAINENd
, L. VERDONCKc, and J. SWINGS"
From the Department of Biological Sciences, Heriot-Watt University, Riccarton, Edinburgh EHI4 4AS, Scotland'; Departament de Microbiologia, Universitat de Barcelona. Av. Diagonal 645, 08028 Barcelona, Spain"; Centre National de Typage Moleculaire Enterique, Unite des Enterobacteries, Institut Pasteur, 28 rue du Docteur Roux. F-75724 Paris Cedex \5, France\ Department of Veterinary Microbiolo~, Laboratory of Fish Diseases, Royal Veterinary and Agricultural University, Bulowsvej 13, DK-1870 Frederiksberg C, Denmark ; and Laborarorium voor Microbiologie, Faculteit Wetenschappen, Universiteit Gem, K. L. Ledeganckstraat 35, B-9000 Gent, Belgium<
Received February 26, 1995
Summary
The majority (91 %) of 260 isolates initially identified as Vibrio anguillarum, that were obtained from a wide range of hosts, habitats and geographical locations, were recovered in a single cluster based on the ribotype and were pathogenic to Adamic salmon. A significant proportion of isolates (78% of the total) were allocated to 15 serogroups (01 - 010 and five previously undescribed groups referred ro as VaNTl, VaNT2, VaNT4, NaNT5 and VaNT7). A minority of isolates (6%) reacted with more than one antiserum or were self-agglutinating, and the remainder did not react with any of the antisera tested. Good correlation was noted between serogroups and lipopolysaccharide profiles, particularly with respect ro isolates belonging to serogroups 01, 02 and 04 - 010. Plasmids were recognized in some serogroups, especially 01, which contained the 67 kb plasmid associated with virulence. However, the) 9 profiles based on outer membrane protein patterns did not correspond to the results obtained with the other typing methods. Generally, the isolates were heterogeneous in their biochemical characteristics; 117 profiles were obtained with the API 20E system, and 9 and 32 clusters recognised from the results of BIOLOG fingerprinting and Biotype-l00 biotyping methods, respectively. Three dominant clusters were defined from fatty acid methyl esters profiles.
Key words: Vibrio - Fish isolates - Fish pathogens - Identification - Typing methods
Introduction
Vibriosis, caused by Vibrio anguillarum, is a significant limiting factor in mariculture, worldwide (Austin and Austin, 1993). Although formally reclassified into a newly proposed genus Listonella, as Listonella a1tguillara (MacDcmell et aI., 1986), this classification was and is not widely accepted. Consequently, the pathogen is still commonly referred to as V. anguillarum_ There is considerable heterogeneity between descriptions of the species (e_g_ West et aI., 1983; Kaper et aI., 1983; Larsen and Olsen, 1991), especially with regard to the recognition of a plethora of biotypes (Austin and Austin, 1993) and serotypes (Sorensen and Larsen, 1986). Initially, two biotypes, A and B (Nybelin, 1935) were recognised, followed by bioty-
pe C (Smith, 1961) and then biotype D and E (see Austin and Austin, 1993). Heterogeneity was recognised, insofar as two to four subgroups were defined after numerous taxonomy studies, including those of Kusuda et al. (1979), West et al. (1983) and Kaper et al. (1983). Furthermore, 10 serotypes/serogroups were defined (Muroga et aI., 1984; Kitao et aI., 1983; Sorensen and Larsen, 1986), some of which, e.g. serogroup 02, traversed species boundaries, i.e. V. anguillarum and V. ordalii (Larsen and Olsen, 1991). In part, this heterogeneity is a reflection of the ad hoc way in which vibriosis is often diagnosed, i.e. purely from pathological signs on diseased animals. Thus, the aetiological agent is frequently allocated to V. anguil-

286 B. Austin et al.
larum on no sound bacteriological basis (Allstin and AIIstin, 1993). When employed, identification techniques for pure cultures have relied heavily on serology, and biochemistry (Austin and Allstin, 1993). Attempts have been made to use rapid methods, such as the API 20E system (e.g. Kent, 1982; Maugeri et aI., 1983; Grisez et aI., 1991). However, the biochemical characteristics of V. anguillarum need clarification, because there is some dispute about the profile of the taxon (Larsen and Olsen, 1991; Wiik et aI., 1991). Although a diverse range of techniques have been used for identification and typing of V. angl/illarum (Austin and Austill, 1993), the comparative merits of the methods have not been assessed. Consequently, this study has sought to compare the efficacy of different methods for the identification and typing of cultures received as V. anguillarum.
Materials and Methods
Examination of cllitt/res. Two hundred and sixty cultures provisionally idenrified as V. angllillanmz. obtained from 17 species of fish, Artemia, rotifers, sediment and water in 16 countries, were examined (Table I). These were maintained in 20-25% (wi v) glycerol at -70, -80 or -40°c, and as plate cultures at room temperature on marine 2216E agar (Difco, Detroit, Michigan, USA) and tryptone soya agar (TSA; Oxoid, Basingsroke, England) supplemenred with 1 % or 1.5% (w/v) sodium chloride.
Ribotyping. Bacteria were grown in a shaking incubator for 18 h at 37°C in trypro casein soy (TCS) broth (Diagnostic Pasteur, Sanofi, France) supplemented with 2% (w/v) sodium chloride (TCSN). DNA was extracted from pellcts with the automate AutoGen 540 (AutoGen Instrumenrs, USA), and, after dialysis, cleaved for >3 h at 37°C with Bgn (Boehringer, Mannheim, Germany) (Grimont et aI., 1989). Restriction fragments were separated e1ectrophoretically through 0.8°;', (w/v) agarose (Appligene, IIIkirch, France) in Tris-borate buffer (89 mM Tris, 89 mM boric acid, 2 mM EDTA buffer, pH 8.3) for 16 h at 1.5 VI cm, before transfer to nylon membranes (Hybond-N; Amersham International, Amersham, England) using a VacuGene System (Pharmacia). The DNA was fixed (15 min at 80°C) before hybridization with an acetylaminofluorene (AAF)-labelled ribosomal probe (Eurogenrec, Seraing, Belgium), as described by Grimont et al. (1989). After blocking, the membranes were incubated at room temperature for 2-3 h with mouse anti-acetylaminofluorene monoclonal antibodies, and washed three times for 10 min in buffer (0.1 M maleic acid, 0.15 M NaCI, 0.3% Tween 20; pH 7.5) to eliminate excess antibody. Antimouse IgG alkaline phosphatase-conjugated anribody (1 Ilg/ml) was added for 1 h, then, the systems were washed three times for 10 min. Revelation was in 0.1 M Tris HCI, 0.1 M NaCI, 50 mM MgCI 2 ; pH 9.5, after addition of nitroblue tetrazolium (Sigma) and 5 -bromo-4-chloro-3-indolyl phosphate (Sigma) as substrate. Purplish-blue bands appeared within a time span of a few minutes to one hour. The data were examined using the Dice (SAB) coefficient with clustering by single linkage (Sneath and Sokal, 1973; Priest and Austin, 1993).
Serotyping. The preparation of absorbed pol}'donal antisera and O-antigen have been described previously (Maniatis et aI., 1982; Larsen et aI., 1994). A loopful of undiluted antiserum was mixed with an equal volume of O-antigen on a glass microscope slide, and agglutination reactions were recorded after visual inspection.
Determination of lipopolysaccharide (LPS) profiles. LPS was extracted after enzymic digestion of lysed cells with proteinase K (Hitchcock and Brown, 1983), polyacrylamide electrophoresis using Mini-Protean" Ready Gels (Bio-Rad, J-krcules, California, USA) without 5DS, and with LPS silver stain (I\.tttelberger and Hilbink,1993).
Determination of outer membrane protein (OMP) profiles. OMP was extracted from young cells (overnight broth cultures) which were disrupted by alternate freezing and thawing in TrisHCI (10 mM) buffer prepared in 0.3% (w/v) sodium chloride with the sarkosyl technique (tambert. 1986), polyacrylamide electrophoresis using Mini-Protean" Ready Gels (Bio-Rad) without 5DS, and later with Coomassie Brillianr Blue stain. The gels were stained with protein-silver stain (Merril et aI., 1981), as necessary.
Gas chr01l1atogr,lphic analysis of cellular fatty acid methyl esters (FAMEs). A subset of 147 V. anguillartlm cultures was examined by this method. Quanritative analysis of cellular fatty acid compositions was performed using the gas-liquid chromatographic procedure, as described by De Boer and Sasser (1986). Cultures were grown for 24 h at 28°C on TSA supplemented with 1.5% (w/v) sodium chloride. Approximately 70 mg of cells were added to 1 ml of 3.75 M NaOH in 50% (v/v) aqueous methanol, and heated for 30 min in a boiling water bath for saponification. Methylation was achieved by adding 2 ml of 6 N hydrochloric acid in 50% (v/v) aqueous methanol and heating for 10 min at 80°C. After cooling to room temperature, fatty acid methyl esters (FAMEs) were extracted with a 1: I mixture of hexane and methyl-iso butylether. The FAMEs were analysed with a Hewlett-Packard model 5898A gas chromatograph and identified using the Microbial Identification System software package (MIS version no. 3.7, obtained from Microbial ID Inc., Newark, Delaware, USA). Clustering was by numerical analysis using the Euclidean distance coefficient and the unweighted pairgroup method of averages (UPGMA; Sneath and Sakal, 1973; Priest and Austin, 1993). Clusters were delineated at Euclidean distance 69.
Plasmid profiling. Overnight cultures in Luria-Berrani broth base (LB; Gibco, Paisley, Scotland) or veal infusion broth (Difco) supplemented with 0.5% (w/v) sodium chloride were used for the determination of plasmid profiles. The plasmids were extracted from the bacteria by the method of Kado and Liu (1981). Plasmids were separated by gel electrophoresis in 0.6-1.0% agarose gels (SeaKem GTG; FMC, Bio Products, USA) in T AE buffer, at pH 8.0. The size of the plasmids was determined by the length of migration, according to the method of Rochelle et al. (1985). All cultures were examined at least four rimes.
Examination of ClIltllres by the API 20E rapid idmtifiwtion s)'stem. Inocula were prepared from 24-28 h cultures incubated at 20 DC on marine 2216E agar (Difco). Cells were harvested in phosphate buffered saline (PBS) supplemenred with 2% (w/v) sodium chloride, and the suspensions adjusted with fresh PBS to achieve a standard opacity equivalent to McFarland No.3. The API 20E rapid identification sysrems (BioMerieux, Marcyl'Etoile, France) were inoculated using the modification for marine bacteria described by Kent (1982) and Grisez er al. (1991) with incubation at 20°C for 24 and 48 h.
BIOLOG fingerprints. Strains were grown for 24 h at 25 DC on brain heart infusion agar (Difco) supplemented with 1.5% (wI v) sodium chloride. Inocula were prepared in 1.5% (w/v) sodium chloride, and the cell density standardized (0 berween 0.261 and 0.3 OD using a spectrophotometer at 590 nm. Each well in the BIOLOG GN microplate (Biolog Inc., Hayward, CA., USA) was inoculated with 150 fd of cell suspension, and the microplates were incubated at 25°C for 24 h. Changes in colour were measured using a Multiscan Multisoft filter photometer (Labsystems, Helsinki, Finland) at 590 nm. The BIOLOG profiles were com-

pared by numerical analysis (PhenoCompar, L. Vallterin) with similarities calculated by the Pearson product moment correlation coefficient between strains_ The strains were grouped by UPGMA. Phenotypic clusters were delineated at 80% r. Differentiating characters between pairs of clusters were calculated using the BIONUM software package (L. Vallterin et al.), and were defined as those tests yielding >90% and < 10% of positive responses, respectively. In order to extract differentiating characters, optical density values were recalculated between maximum and minimum values, whereby the cut-off factor to obtain positives and negatives was set at 30%. The internal homogeneity within each delineated cluster was calculated using the same software package.
Biotyping. Carbon source utilization tests were carried out using Biotype-IOO strips (BioMerieux; La Balme-Ies Grones, France) that contained 99 pure carbon sources. Volumes (60 ml) of Biotype medium No.1 were supplemented with 2% (w/v) of sodium chloride and 100 mM MgClb and inoculated with 2 ml amounts of a 100- Klett unit suspension of bacterial cells in 2 % (w/v) sodium chloride. The strips were completely filled so as ro obtain a slightly convex meniscus, and incubated at 30°C for 2 and 4 days. The cupules were examined visually for the presence of growth, and the data were examined by the Jaccard (S)) coefficient with UPGMA clustering (Sneath and Sokal, 1973; Priest and Austin, 1993), using the Recognizer program (Institut Pasteur Taxolab, Paris, France).
Fish pathogenicity experiments. Bacterial cultures were grown overnight at 22°C in tryptone soya broth (Oxoid) supplemented with 1 % (w/v) sodium chloride, centrifuged at 5,000 x 'g' for 10 min and resuspended in 10 ml volumes of 0.9% (w/v) saline to approximately ION cellslml. Ten fold dilutions were prepared to 104 cells/ml. A salmonid fish model (Atlantic salmon [Salma salar]) was used to assess pathogenicity, as follows:
Groups of 10 fish wre infected by intraperitoneal injection with 0.1 ml volumes of the washed bacterial suspensions to achieve doses of 104 _106 cells/fish. The infected animals were maintained for up to 14 days in covered polypropylene tanks supplied with dechlorinated, aerated static freshwater (the water was changed daily) at a temperature of -15°C. Dead and moribund fish were removed, and subjected to standard bacteriological and pathological examination (Austin and Allstin, 1989). Any survivors at the end of the experiment were sacrificed and examined, as above. The disease signs were recorded, and attempts made to recover and confirm the identity of the pathogen by the methods described by Allstin and Austin (1993).
Test reproducibility. The test results for 20 duplicate cultures were compared for all methods, and an estimate of individual test variance (S,2) was calculated (formula 15; Sneath and Johnson. 1972). The average test variance was calculated to obtain a pooled variance (52), which was used to estimate the average probability (p) of an erroneous test result (formula 4; Sneath and Johnsorl, 1972).
Results
1. Test reproducibility
Inclusion of duplicate isolates in the study enabled experimental test error to be estimated. The average probability (p) of an erroneous result was 5_6%, calculated from the pooled variance (52 = 0.0558) of all the methods scored for the duplicate cultures. Values of 5,z ranged from zero for some of the test regimes, e.g. ribotyping, through 0.03 for OMP and pathogenicity, 0.05 for serotyping, 0.15 for LPS profiles, 0.44 for the API 20E system, and 0_9 for BIOLOG.
Identification and Typing of Vibrio angllillarum 287
2. Ribotypillg
The majority (236/260 = 91 %) of the cultures were ascribed to a single ribotype cluster defined at or above the 80% similarity level (Table 1; Fig. 1). The cultures in this cluster were characterised by three groups of bands. Thus, there were 2-7,4-9 and 2 bands between 10 kb and 20 kb, 4 kb and 8 kb, and 2 kb, respectively. The remaining 24 cultures were recovered in 19 separate clusters. Of these, clusters 2, 3, 4, 6, 8, 9, 10, 11, 12, and 15-20 comprised single isolates; clusters 5, 13 and 14 each contained two isolates, and cluster 7 comprised 6 isolates (Table 1; Fig. 1).
% SIMILARITY 40 !
100 rlU~rn ~O Of mm ij~ oour~ IDENTITY
mmrt
2
3 4 5 6
7
8 9
\0 11
12 \3 14
IS 16
17 18 11)
20
236
1 2
6
2 2
v. anguillarum
VIB 225
VIB66
VIB 76 VIB 189. VIB 190 VlB 239
VIB 192. VIB 193. VIB 195. VIB 210. VIB 21I, VIB 223 VIB 224 VlB 212
VIB 194 VlB 215
VIB 23 VlB 186, VIB 213 VlB 144. VIB 145
VlB 187 VlB 218 VIB 191 VIB 214
VIB 185 VlB IR4
Fig. 1. Simplified dendrogram based on the examination of ribotyping data by the SAS coefficient and single linkage clustering.

Tab
le 1
. B
acte
rial
cul
ture
s tv
0
0
00
Lab
orat
ory
Col
lect
ion'
/ So
urce
and
/or
coun
try
Rib
otyp
e S
ero·
LP
S O
MP
FA
:VIE
Pl
asm
id
BIO
LO
G
API
20E
B
ioty
pe
Pat
ho-
refe
renc
e no
. st
rain
no.
of
ori
gin
clus
ter
grou
p pr
ofil
e pr
ofil
e pr
ofil
e c
on
terl
( pr
ofil
e pr
ofil
e pr
ofil
e ge
nici
ty
?'
VIB
1
AT
CC
433
05
Ollc
orh)
'IIc/
ms
lIIy
kiss
, 0
1
2 2
67 k
b 30
4752
556
+
>- c D
enm
ark
~
VIB
2
AT
CC
433
06
Gad
us m
orim
a, D
enm
ark
02
a
2 1
2 0
3247
5275
7 +
5'
n
VIB
3
AT
CC
433
07
O. m
ykis
s, D
enm
ark
03
3
3 2
0 32
4752
757
25
+
~ '"
VIB
4
AT
CC
433
08
G.l
llor
hua,
Den
mar
k 0
4
4 2
2 0
3247
5275
7 15
+
:-
VIB
5
AT
CC
433
09
G. m
orhu
a, D
enm
ark
05
5
3 2
0 30
4712
557
30
+
VIB
6
AT
CC
43
3\0
G
.lll
orhu
a, D
enm
ark
06
6
3 2
0 30
4752
757
+
VIB
7
AT
CC
433
11
Ang
uill
a an
guill
a, D
enm
ark
07
7
1 2
0 32
4752
757
15
+
VIB
8
AT
CC
433
12
G. m
orl"
,a,
Den
mar
k 0
8
8 1
2 0
3247
5255
7 16
+
V
IB 9
A
TC
C 4
3313
G
. mor
hua,
Den
mar
k 0
9
9 3
2 0
1247
1255
7 +
V
IB 1
0 A
TC
C 4
3314
C
. mor
hua,
Den
mar
k 10
10
.3
2 0
3247
5275
7 27
+
(w
eak)
V
IB 1
1 A
VL
90-9
-22
Dic
entr
arch
us la
bm.>
:, 0
1
1 3
67 k
b 30
4773
757
23
+
Gre
ece
V1B
12
AV
L 89
-2-5
4 D
.lab
rax,
Gre
ece
02
3
2 2
2 0
3047
5164
7 24
+
V
IB 1
3 A
VL
68-2
8-C
D
.lab
rax,
Gre
ece
02
b
2 1
0 32
4753
757
8 +
V
IB 1
4 H
WU
44
O. m
ykis
s, I
taly
0
1
3 67
kb
3247
7375
7 23
+
V
IB 1
5 H
WU
48
D.la
hra.
>:,
Gre
ece
01
3
67 k
b 32
4752
457
I +
V
IB 1
6 H
WU
49
D.l
ahra
x, G
reec
e 0
1
2 6
7k
b
3047
7245
7 4
+
VIB
17
HW
U 5
0 D
. lah
rax,
Gre
ece
01
3
67 k
b 30
4753
757
2 +
V
IB 1
8 A
VL
27.
2 O
. myk
iss,
Den
mar
k 0
1
2 2
50
,67
kb
3046
7375
7 +
V
IB 1
9 A
VL
2&.2
O
.III
),ki
ss,
Den
mar
k 0
1
1 4
50
,67
kb
I 30
4773
757
+
VIB
20
AV
L 2
8.3
O. m
),ki
ss,
Den
mar
k 1
01
I
2 5
0,6
7 k
b I
3247
7375
7 32
+
V
IB 2
1 A
VL
28.
4 O
. m),
kiss
, D
enm
ark
I 0
1
1 2
50
,67
kb
I 72
4753
757
+
VIB
22
AV
L 20
/90
D.l
abra
x, G
reec
e I
05
24
10
4
6,7
6 k
b 8
5346
2255
7 +
V
IB 2
3 A
VL
33/
91
Spar
us t
lura
ta,
Gre
ece
12
05
24
10
76
kb
8 53
4622
557
+
VIB
24
AV
L 1
8/91
Sp
. aur
ata,
Gre
ece
24
10
1 5
0,7
0 k
b 8
5346
2255
7 +
V
IB 2
5 H
WU
63
Sa
llllo
sal
ar,
Chi
le
01
1
2 3
67 k
b 1
3047
7275
7 +
V
IB 2
6 H
WU
64
fish
02
b
2 0
1 32
4712
757
9 +
V
IB 2
8 H
WU
V0
2
Sa.
sala
r, N
orw
ay
01
2
2 67
kb
3047
5275
7 +
V
IB 2
9 H
WU
VA
73
D.l
ahra
x, G
reec
e cr
oss-
2 2
0 32
4712
657
+
reac
tion
s be
twee
n 0
2a&
05
V
IB 3
0 H
WU
VA
75
Sp. a
ura
ta,
Gre
ece
(ros
s-2
3 0
3247
1265
7 7
+
reac
tion
s be
twee
n 0
2a&
05
V
IB 3
1 N
CM
B 8
28
Sa. s
afar
, U
.K.
02
a
2 2
2 0
3247
5245
7 +
V
IB 3
2 N
CM
B 5
72
O. m
ykis
s. J
apan
0
1
2 67
kb
3047
5275
7 +
V
IB 3
4 U
B 0
1/91
~Iugilidac,
Ital
y 0
1
1 3
67 k
b 32
4752
457
2 +
" V
IB 3
5 U
B 2
8/89
D
.lab
rax,
Ira
ly
01
1
2 67
kb
3247
5245
7 2
+
VIB
36
UB
76/
91
Mug
ilid
ae,
Ital
y 0
1
1 2
2 67
kb
3247
5245
7 +
(w
eak)
V
IB 3
7 U
B 1
09/9
0 Sp
. aI/
rata
, It
aly
04
4
19
0 32
4752
757
25
+
VIB
39
UB
178/
90
D.l
ahra
x, I
tal)
, 0
1
1 1
2 67
kb
3247
5244
7 +
V
IB 4
0 U
B 19
1/90
D
.lab
rax,
Ita
l),
01
1
78,
84 k
b 32
4752
557
+
VIB
41
UB
2071
90
Sp. a
urat
a, I
taly
08
8 0
3247
1245
7 +

Tab
le I
. C
on
tin
ued
Lab
orat
ory
Col
lect
ion'
1 S
ourc
e an
dlor
cou
ntry
R
ibot
)'pe
Sero
-LP
S O
MP
F
AM
E
Pla
smid
B
IOL
OG
A
PI2
0E
B
iot)
'pe
Pat
ho-
refe
renc
e no
. sr
rain
no.
of
ori
gin
clus
rer
grou
p pr
ofil
e pr
ofil
e pr
ofil
e co
nten
t pr
ofil
e pr
ofil
e pr
ofil
e ge
nici
t)'
VlB
42
UB
217
/90
D./
abra
x, I
taly
2
2 0
3247
5275
7 +
V
IB 4
4 U
B 2
6119
1 D
./ab
rax,
Ita
ly
0)
1 1
35
,60
kb
3247
5275
7 +
VIB
45
UB
295
/91
D./
abra
x, I
taly
0
4
4 12
0
3247
5257
7 +
" V
IB 4
6 U
B 3
27/9
0 D
./ab
rax,
Ita
ly
01
I
2 67
kb
3247
5245
7 1
,3
+"
VlB
47
UB
342
191
D./
abra
x, I
taly
0
1
1 2
67 b
32
4752
457
1 +
b V
lB 4
8 U
B 3
9319
1 D
./ab
rax,
Ira
l)'
01
1
2 67
, 84
kb
3247
5265
7 23
+
" V
lB4
9
UB
397
/90
D.l
abra
x, I
taly
4
2
3 0
3247
5276
7 +"
V
lB 5
0 U
B 4
17/9
0 D
./ab
rax,
Ita
ly
02
a
2 2
0 32
4752
446
+
VIB
5!
UB
498
/90
fi.h,
Ita
ly
02
a
42
3 0
3247
5275
7 15
+
V
IB 5
2
UB
505
/91
D./
abra
x, I
taly
46
1
2 0
3247
5257
7 16
+
V
IB 5
3 U
B 2
58/9
1 D
./ab
rax,
Ita
ly
02
a
2 2
0 32
4732
657
+ V
IB 5
4 U
B 5
74/9
1 Sp
. aur
ata,
Ita
ly
07
7
2 0
3247
5255
7 +
V
IS 5
5 U
B 5
78/9
0 D
. /a
brax
, It
aly
02
a
2 2
0 32
4732
657
6 +
V
lB 5
6 U
B 6
01
19
) D
./ab
rax,
Ita
ly
01
2
67
,84
kb
-32
4752
657
+
VlB
57
UB
86
1/8
9
D./
abra
x, I
raly
0
1
1 2
67 k
b 32
4752
757
+
VIB
58
UB
904
/89
D./
abra
x, I
taly
13
6
0 32
4752
757
15
+ V
IB 5
9 U
S 90
9/89
D
.lab
rax,
Ita
ly
01
1
1 0
3247
5245
7 +
(w
eak)
V
IB 6
0 U
B 9
10/9
0 Sp
. aur
ata,
Ita
ly
03
42
13
0
3247
5277
7 31
+
V
IB 6
1 U
B 9
60/9
0 D
.lab
rax,
Ita
ly
03
42
2
2 0
3247
5265
7 27
+
" V
IB 6
2 U
B 1
0321
90
D./
abra
x, I
taly
0
3
42
2 0
3047
5275
7 V
IB 6
3 U
B 1
075/
90
D./
abra
x, I
taly
0
3
42
2 0
7347
5275
7 24
+
V
IB 6
4 U
B A
023
Scop
htha
/mus
max
im/I
S,
01
2
2 26
kb
3047
5275
7 23
+
Spai
n V
IB 6
5 U
B A
024
Sc. m
axim
us,
Spa
in
1 0
1
1 2
67 k
b
3247
5275
7 +
c;:
V
IB 6
6 U
B A
044
Sc. m
axim
/IS,
Spa
in
3 8
11
0 30
4652
556
+
" V
IB 6
7 U
B A
055
Sc. m
axim
us.
Spa
in
01
1
2 67
kb
3247
7265
7 23
+
" a
VlB
68
UB
A05
6 Sc
. max
imu5
, S
pain
0
1
1 2
67 k
b 30
4652
457
23
+
n'
~
VIB
69
UB
A07
S Sp
. allr
ata,
Spa
in
02a
2 6
2 0
3247
5275
7 24
+
o·
VIB
70
UB
A08
5 ro
tife
r, S
pain
5
2 0
3047
5275
7 15
+
" ::J
'" V
IB 7
1 L
MG
441
1 S
cotl
and
02
a
14
3 2
0 4
3247
5245
6 " c...
VlB
72
L
MG
443
7T
G. m
orhu
a, N
orw
ay
02
a
2 2
2 0
1 32
4752
757
+
-I
VIB
73
LM
G 1
1684
1
02
a
2 2
2 0
3247
5264
7 .....
+
""
VlB
75
LM
G 1
0939
U
.S.A
. 1
01
I
2 2
67
kb
4
3207
5275
6 +
5 C/O
VIB
76
LM
G 1
2098
Ja
pan
4 0
2a
2 1
2 0
4 30
4752
547
+
~
VIB
77
LM
G 1
2099
P/
ecog
loss
/4S
a/tiv
e/is
. Ja
pan
I 0
2a,
b
2 6
2 0
3247
5255
7 19
+
~
VIS
78
LM
G 1
2100
P.
a/ti
ve/is
, Jap
an
1 14
9
2 5
0 k
b
3047
1245
7 +
<3
-
VIS
79
LM
GI2
1O
! 0
1
1 2
2 77
kb
3047
5265
7 +
a<
VIS
80
LM
G 1
2102
0
2a
2 9
2 0
3047
3265
7 +
t:.
VIB
81
LM
G 1
3185
D
./ab
rax,
Gre
ece
cros
s 43
1
2 4
6,6
8,
3247
7255
7 16
+
~ ::
reac
tion
s 83
kb
~ V
IS 8
2 L
MG
131
86
D.l
abra
x. G
reec
e V
aNT
2 43
1
2 6
8,8
3 k
b 32
4772
577
10
+ ...
VIB
83
LM
G 1
3187
D
./ab
rax,
Fra
nce
01
1
2 2
0 30
4752
457
1 +
(w
eak)
~
VIS
84
LM
G 1
3188
D
./ab
rax,
Ita
ly
01
2
2 67
kb
3047
7245
7 +
VlB
85
RV
AU
87-
9-11
7 O
. myk
iss,
Fin
land
0
1
2 67
kb
3247
7265
7 +
b
VIB
86
RV
AU
169
2 O
. myk
iss,
Ita
ly
01
2
67 k
b 30
4752
757
+
1'-'
00
\0

Tah
le I
. C
on
tin
ued
, .... 'C
0
Lab
orat
ory
Col
lect
ion"
1 S
ourc
e an
d/or
cou
ntry
R
ibot
ype
Ser
o·
LPS
OM
P
FA!v
IE
Pla
smid
B
IOL
OG
A
PI2
0E
B
ioty
pe
Pat
ho·
refe
renc
e no
. st
rain
no.
o
f or
igin
cl
uste
r gr
oup
prof
ile
prof
ile
prof
ile
cont
ent
prof
ile
prof
ile
prof
ile
geni
city
?"
V
IB 8
7 N
CM
B 1
873
Sa. s
al"r
, U
.K.
cros
s 2
5.3,
11.
5,
30
47
72
65
7
+ (w
eak)
;.
.
reac
tion
s 5
3k
b
~ be
twee
n :5' "
01
&
M '"
VaN
Tl
-V
IB 8
8 R
VA
U 5
1182
12
O. m
ykis
s, G
erm
any
01
1
2 67
, 10
5 kb
3
04
77
26
57
V
IB 8
9 R
VA
U 8
2061
7·11
6 O
. myk
iss,
Den
mar
k 0
1
1 2
67
kb
3247
5275
7 +
VIB
90
RV
AU
820
901·
2/2
O. m
ykis
s, S
wed
en
01
I
2 67
kb
3247
5274
7 +
h V
IB 9
1 R
VA
U 8
3042
2·1/
1 O
. myk
iss,
Den
mar
k 0
1
I 2
2 67
kb
3247
7275
7 +
VIB
92
RV
AU
810
530·
111
O. m
ykis
s, D
enm
ark
01
1
2 67
kb
1 32
4772
757
+ V
IB 9
3 R
VA
U 8
5061
O-1
/6a
O. m
ykis
s, D
enm
ark
01
I
2 67
kb
I 32
4772
757
VIB
94
NC
MB
829
O
. myk
iss,
U.K
. 0
2a
2 4
2 0
4 32
4752
456
8 +
VIB
95
RV
AU
222
8/80
sa
lmon
, S
wed
en
02
a
2 2
0 32
4750
747
+ V
IB 9
6 R
VA
U 2
887
O. m
yki5
s, I
taly
0
2a
2 2
2 64
, 32
4732
657
+ -2
00
kh
VIB
97
RV
AU
850
617·
1/1
Eso
x lu
cius
, F
inla
nd
02
a
2 I
0 32
4752
757
18
+ V
IB 9
8 R
VA
U 8
6090
8·3
Sc. m
axim
us,
Nor
way
0
2a
2 2
0 30
4752
657
21
+ (w
eak)
V
IB 9
9 R
VA
U 8
9-2-
62
Sc. m
axim
lls,
Nor
way
0
2a
2 7
2 0
3047
5375
7 +
VIB
100
R
VA
U 9
1-7-
175
Sc. m
axim
lls,
Den
mar
k 0
2a
2 2
0 32
4712
757
7 +
(wea
k)
VIB
101
R
VA
U 8
2072
3-21
8 Sc
. max
imus
, S
wed
en
02
b
2 I
2 2
0 k
b 30
4752
757
7 +
(wea
k)
VIB
102
R
VA
U 1
076/
2 G
. mor
hlla
, D
enm
ark
02
b
2 2
0 30
4752
457
14
+ V
IB 1
03
RV
AU
108
9/2
G. m
orhu
a, D
enm
ark
02
b
2 1
>2
00
kb
3247
5245
6 14
+
VIB
104
R
VA
U 1
4741
1 G
. mor
hua,
Den
mar
k 0
2b
2
7 2
>2
00
kb
3247
5275
7 +
VIB
105
R
VA
U 6
828c
Sc
. max
illlu
s, D
enm
ark
02
b
2 7
0 32
4752
757
14
+ V
IB 1
06
RV
AU
25/
82/2
O
.lIIy
kiss
, G
erm
an),
0
2b
2
2 0
3247
5275
7 +
VIB
108
R
VA
U 8
6061
9·3/
2 O
. lII
ykis
s, D
enm
ark
02
b
2 2
2 >
20
0 k
b 32
4772
757
+ V
IB 1
09
RV
AU
860
828-
1I3a
O
. myk
iss,
Den
mar
k 0
2b
2
2 0
3247
7275
7 +b
V
IB 1
10
RV
AU
90·
2-6
Pol
lach
il/s
(lire
llS,
Nor
way
0
2b
2
2 2
>2
00
kb
3247
3245
6 +
(wea
k)
VIB
III
R
VA
U 1
3211
1 G
. m
orhu
a, D
enm
ark
self
agg
l-45
2
2 0
3247
5265
7 +b
ur
inat
ing
VIB
112
R
VA
U 8
2080
6·11
2 O
. myk
iss,
Den
mar
k cr
oss
45
3 0
3247
5275
7 28
re
acti
ons
VIB
113
R
VA
U 8
2080
6-1/
3 O
.lIIy
kiss
, D
enm
ark
03
3
.3 2
0 3
24
75
27
57
28
V
IB 1
14
RV
AU
171
2/1
G. m
orhl
/a,
Den
mar
k 0
4
4 2
2 0
3247
5275
7 2
0
VIS
115
R
VA
U 7
4/82
/1
O. m
ykis
s, G
erm
any
04
4
8 0
3247
5275
7 V
IB 1
17
RV
AU
89·
9·18
1 Sa
. sal
ar,
Nor
way
0
4
4 8
2 0
32
47
52
75
7
VIB
118
R
VA
U 1
629
G. m
orhl
/a,
Den
mar
k 0
5
5 I
2 0
30
47
12
55
7
VIB
119
R
VA
U 8
5081
6-11
17b
O. m
ykis
5, D
enm
ark
05
5
1 2
0 32
4712
557
VIB
120
R
VA
U 9
0-2-
9 D
.lab
rax,
Nor
way
15
2
0 3
04
75
27
57
15
+
V
IB 1
22
RV
AU
682
8g
Sc. m
axill
lus,
Den
mar
k 0
7
7 3
0 3
24
75
27
57
+h
V
IB 1
23
RV
AU
170
113b
A
ngui
lla s
p.,
Den
mar
k cr
oss
2 14
0
3247
5245
6 8
+ re
acti
ons
VIB
124
R
VA
U 1
7021
1 A
ngui
lla s
p.,
Den
mar
k 0
7
7 3
0 3
04
75
27
57
15
+b
V
IB 1
25
RV
AU
88-
12-7
16
Sa.
sala
r, N
orw
ay
08
8
2 0
3247
5257
7 16
+
VIB
126
R
VA
U 8
4060
6·21
5 O
. myk
iss,
Den
mar
k V
aNT
I 45
2
53
,56
kb
3247
5275
7 23
+b
V
IB 1
27
RVAU
373
7/1
O. m
ykiss
, Ita
ly
VaN
T4
47
28
2 0
3247
3245
7 +

,~
Tab
le I
. C
onti
nued
0 ~
< ~
Lab
orat
or),
C
olle
ctio
n'l
Sour
ce a
nd/o
r co
untr
),
Rib
otyp
e Se
ro-
LPS
OM
P
FAM
E
Plas
mid
B
IOL
OG
A
PI 2
0E
Bio
t)'p
e Pa
th,,-
;,-re
fere
nce
no_
stra
in n
o.
of o
rigi
n du
ster
gr
oup
prof
ile
prof
ile
prof
ile
cont
ent
prof
ile
prof
ile
prof
ile
geni
cit)
' .., "2
- ~ V
IB 1
28
RV
AU
850
610-
lIS
b O
. myk
iss,
Den
mar
k V
aNT
2 0
3 2
11.5
kb
3247
5275
7 +
" g.
VIB
129
R
VA
U 8
5061
7-11
3 G
. mor
hua,
Fin
land
V
aNT
5 43
2
0 72
4752
557
+
~ V
IB 1
30
RV
AU
91-
7-15
2 Sc
. max
im us
, D
enm
ark
2 2
2 0
3246
1055
6
~ V
IB 1
31
RV
AU
88-
6-73
12
O.
myk
iss,
Den
mar
k 0
2a
2 2
2 5
,5.5
, 32
4752
757
+
11 k
b "" .~
VIB
132
R
VA
U 9
1-7-
143
0_ m
),ki
ss,
Den
mar
k 0
3
0 2
2 0
3247
5275
7 V
IB 1
34
RV
AU
91-
8-17
8 Sc
. max
im us
, N
orw
ay
01
2
53
,67
kb
3247
5275
7 23
+
V
IB 1
35
RV
AU
91-
11-2
44
Sc. m
axim
us,
Nor
way
0
1
I 2
2 3
3,6
7 k
b 32
4772
757
21
+"
VIS
136
R
VA
U 9
2-8-
158
All
guil
la s
p.,
Den
mar
k 0
3
3 2
0 32
4752
757
17
+
VIB
137
R
VA
U 9
2-8-
161
Allg
uilla
sp.
, D
enm
ark
02
a
2 2
0 30
4652
457
+
VlB
138
R
VA
U 9
2-8-
163
Allg
uilla
sp.
, D
enm
ark
1 0
2a
2 2
2 0
1046
5245
6 8
+
VIB
139
R
VA
U 9
2-8-
\68
Ang
uill
a sp
., D
enm
ark
I 0
3
3 2
2 0
3247
5275
7 +
V
IB 1
40
RV
AU
910
614-
111
G. m
orhu
a, D
enm
ark
I 0
2a
2 2
0 32
4712
657
5 +
V
IB 1
41
RV
AU
840
523-
2f4b
O
. myk
iss,
Den
mar
k I
VaN
T2
45
3 0
3247
7275
7 13
V
IB 1
42
R V
AU
840
808-
3/4a
O
. m),
kiss
, D
enm
ark
I V
aNT
2 2
3 2
0 7
7246
1255
7 V
IB 1
43
RV
AU
VI
312
wat
er,
Den
mar
k I
01
I
2 0
I 32
4752
577
VIB
144
R
VA
U V
i 4/
2 w
ater
. D
enm
ark
14
48
2 0
7 32
4612
547
VIB
145
R
VA
U V
I 5/
2 w
ater
, D
enm
ark
14
48
2 2
0 7
7046
1255
7 V
IB 1
46
RV
AU
V2
112
wat
er,
Den
mar
k 0
2a
21
2 0
7346
1265
7 +
V
IB 1
47
RV
AU
V2
2/2
wat
er,
Den
mar
k 0
9
0 2
7347
5245
7 V
IB 1
48
RV
AU
V2
4/3
wat
er,
Den
mar
k 0
3
45
I 2
0 1
3247
5275
7 +
(w
eak)
V
IB 1
49
RV
AU
V2
5/3
wat
er,
Den
mar
k V
aNT
2 45
3
2 0
3247
7275
7 12
V
IB 1
50
RV
AU
VI
1/4
wat
er.
Den
mar
k 0
2 2
0 32
4752
777
17
VIB
151
R
VA
U V
I 2/
4 w
ater
, D
enm
ark
0 2
2 0
3247
1275
7 c;:
V
IB 1
52
RV
AU
V2
114
wat
er,
Den
mar
k 0
3
3 2
0 32
4752
757
17
+
" g V
IB 1
53
RV
AU
Sel
112
se
dim
ent,
Den
mar
k 0
3 0
3247
5275
7 21
S1
V
IB 1
54
RV
AU
Sci
3/2
se
dim
ent,
Den
mar
k 0
5
I 0
3247
5275
7 22
,.,
+
~
VIB
ISS
R
VA
U S
e1 5
13
sedi
men
t, D
enm
ark
I 3
2 0
3247
5275
7 +
o·
sedi
men
t, D
enm
ark
05
2
2 2
0 32
4712
557
::>
VIB
156
R
VA
U S
e2 1
/2
., V
IB 1
57
RV
AU
Se2
2/2
se
dim
ent,
Den
mar
k 0
5
2 2
0 32
4712
557
30
~
VIB
158
R
VA
U S
e22/
4 se
dim
ent,
Den
mar
k 4
2 2
0 32
4772
757
...;
VIB
159
R
VA
U S
ci 3
/4
sedi
men
t, D
enm
ark
08
69
2
2 0
3257
0254
7 29
. .,
+
"0
VIB
160
R
VA
U S
el l
iS
sedi
men
t, D
enm
ark
02
a
2 2
2 0
3247
5255
7 5'
Q<
O
VIB
161
R
VA
U S
el 5
15
sedi
men
t, D
enm
ark
cros
s 44
0
3247
1255
7 29
0 .....
re
acti
ons
~ V
IB 1
62
RV
AU
Se2
315
sedi
men
t, D
enm
ark
04
4
18
0 32
4752
752
20
~.
VIS
163
R
VA
U P
T24
P.
a/ti
ve/is
, Ja
pan
01
1
2 0
3047
5275
7 +
(w
eak)
0
VIB
164
R
VA
U P
T49
3 P.
a/ti
velis
, Ja
pan
cros
s 45
35
2
0 32
4752
756
25
+
'" ;:s
reac
tion
s Q
\:)
;:
VIB
165
R
VA
U P
T21
3 P.
a/ti
velis
, Ja
pan
01
2
2 0
1 30
4752
757
~ V
IB 1
66
RV
AU
PB
I5
P. a
/tive
lis, J
apan
0
5
2 2.
6 kb
I
i1 V
IS 1
67
RV
AU
PB
28
P. a
/tive
/is,
Japa
n 45
1
2 0
4 30
0612
457
::! V
IB 1
68
RV
AU
ET
I P.
a/ti
ve/is
, Ja
pan
07
(w
eak)
0
I 30
4752
557
19
VIB
169
R
VA
U 7
75
salm
on.
U.S
.A.
01
2
2 67
kb
3207
7265
7 +
V
IB 1
70
RV
AU
53-
507
salm
on,
U.S
.A.
01
2
2 67
kb
3247
7275
7 17
+
1'
-'
RV
AU
RG
75-
834
salm
on,
U.S
.A.
01
2
2 6
7,3
6 k
b 32
4752
757
'Ci
VIB
171
+

Tab
le I
. C
on
tin
ued
I'
" -0
t"
Llb
ora
tory
C
olle
ctio
n'/
Sou
rce
and/
or c
ount
ry
Rib
otyp
e S
ero-
LPS
OM
P
fAM
E
Pla
smid
B
IOL
OG
A
PI2
0E
B
ioty
pe
Pat
ho-
refe
renc
e no
. st
rain
no.
of
ori
gin
clus
ter
grou
p pr
ofil
e pr
ofil
e pr
ofil
e co
nten
t pr
ofil
e pr
ofil
e pr
ofil
e gc
nici
t)'
?'
VIB
17
5 R
VA
U V
AI2
O
. myk
iss,
Can
ada
01
2
67 k
b 3
24
77
27
57
+
(w
eak)
;..
V
IB 1
76
RV
AU
VA
41
O. m
~'ki
ss,
Can
ada
01
2
2 6
7 k
b 3
24
77
27
57
+
" ~
VIB
17
8 R
VA
U V
A43
O
. Iny
kiss
, C
anad
a 0
1
2 67
kb
30
47
72
75
7
+
5'
VIB
18
0 R
VA
U V
A61
O
. ne
rka,
Can
ada
01
2
67 k
b 3
24
77
27
57
+
~
VIB
181
R
VA
U V
A70
SI
I. sa
/ar,
Can
ada
01
2
67 k
b 3
24
77
27
57
.,
+
:-V
IB 1
82
RV
AU
VA
72
O. m
ykis
s. C
anad
a 0
1
I 2
67 k
b 1
32
47
72
75
7
+
VIB
18
4 L
MG
132
11
Art
emia
, S
pain
29
19
5
2 0
3 3
04
41
37
57
+
V
IB 1
85
l.M
G 1
3212
A
rtem
ia.
Spdi
n 19
0
4
16
16
2 32
kb
8 4
14
71
25
56
+
V
IB 1
86
LM
G 1
3213
A
rtem
ia,
Spa
in
13
19
5 2
4.4
kb
.3
72
44
13
75
6
+
VIB
187
L
MG
132
14
D./
abra
x, S
pain
15
w
eak
17
3 2
2.2
.3.4
, 5
00
04
11
45
6
+
cros
s 4
.2,5
.8
reac
tion
s 6.
8, 5
3 kb
V
IB 1
88
LM
G 1
3215
D
./llb
rax,
Spa
in
cros
s 17
2
2 3.
3,
5 10
0412
456
+ reaction~
12
3 k
h V
IB 1
89
LM
G 1
3216
ro
tife
rs,
Gre
ece
5 17
2
2 88
kb
.3 3
24
45
37
57
+
V
IB 1
90
LM
G 1
3217
ro
tile
rs,
Gre
ece
5 17
2
2 88
kb
3 12
4413
757
+
VIB
19
1 L
MG
132
18
roti
fers
, G
reec
e 17
cr
oss
18
2 2
50
kb
3 3
24
41
27
57
+
re
acti
ons
VIB
19
2 L
MG
132
19
roti
fers
, G
reec
e 7
19
3 2
-19
0 k
b 6
30
46
10
55
6
+
VIB
193
L
MG
132
20
roti
fers
, G
reec
e 7
21
3 2
-20
0 k
b 6
32
46
10
55
6
+
VIB
194
L
MG
132
21
rori
fers
, G
reec
e 10
20
2
2 0
6 3
24
61
05
56
+
V
IB 1
95
LM
G 1
3222
A
rlem
ia,
Spa
in
7 V
aNT
1 21
2
2 0
6 3
24
61
05
56
+
V
IB 1
96
LM
G 1
3223
ro
tife
rs,
Gre
ece
0 2
2 °
1
32
47
72
55
7
+
VIB
197
L
MG
132
24
roti
fers
, G
reec
e V
aNT
7 I
2 16
kb
I
32
47
72
55
7
+
VIB
198
L
MG
132
25
roti
fers
, G
reec
e V
aNT
7 1
2 16
kb
32
47
72
55
7
+
VIB
199
L
MG
132
26
wat
er,
Gre
ece
VaN
T7
2
16
,49
, 3
24
77
25
57
11
+
68
kh
VIB
200
L
MG
132
27
Sp. a
ural
a. G
reec
e V
aW
7
2 16
kb
3
24
77
25
57
10
+
V
IB 2
01
LM
G 1
3228
Sp
. aum
ta,
Gre
ece
VaN
T7
2 16
kb
32
47
72
55
7
10
+
VIB
202
L
MG
132
29
D./
abra
x, G
reec
e V
aNT
7 2
16
,49
, 3
24
77
25
57
11
+
6
8,8
0 k
b V
lB 2
03
LM
G 1
3230
Sp
. aur
ata,
Spa
in
06
6
2 3
.2,5
.3,
32
47
52
75
7
16
+
50,
70 k
b V
IB 2
04
LM
G 1
3231
D
.labr
ax.
Spa
in
6 6
3 0
32
47
52
65
7
22
+
VIB
205
L
MG
132
32
wat
er,
Spa
in
06
(w
eak)
6
6 0
32
47
52
75
7
22
+
VIB
206
L
MG
132
33
roti
fers
, S
pain
0
6
6 6
2 0
32
47
52
65
7
22
+
VlB
207
L
MG
132
34
roti
lers
, G
reec
e 19
2
2 36
kb
6 3
24
61
05
56
+
V
IB 2
08
LM
G 1
3235
D
./abr
ax,
Gre
ece
20
2 2
36
kb
6 3
04
61
25
56
+
V
IB 2
09
LM
G 1
3236
D
. /a
brax
, G
reec
e 1
19
2 2
0 6
32
46
10
55
6
+
VIB
21
0
LM
G 1
3237
w
ater
, G
reec
e 7
wea
k 18
2
2 0
6 3
24
61
05
56
+
(w
eak)
cr
oss
reac
tion
s V
IB 2
11
LM
G 1
3238
w
ater
, G
reec
e 7
05
(w
eak)
20
2 2
120
kb
6 3
24
61
25
56
+
V
IB 2
12
LM
G 1
3239
D
./ab
rax,
Gre
ece
9 19
9
2 3
7 k
b 6
32
46
10
55
6
VIB
213
L
MG
132
40
wat
er,
Spa
in
13
19
17
2 4.
4 k
b
3 7
24
41
37
56
+"
V
IB 2
14
LM
G 1
3241
w
ater
, Gre
ece
18
04
26
15
2
49 k
b 5
15
61
25
57
+
"

Tab
le l
. C
onti
nued
Lab
orat
ory
Col
iect
ion'
l So
urce
and
/or
coun
try
Ribotyp~
Scro
-L
l'S
OM
I'
FAM
E
Plas
mid
B
IOL
OG
A
PI2
0E
Bio
type
I'a
tho-
refe
renc
e no
. st
rain
no.
of
ori
gin
clus
ter
grou
p pr
ofil
e pr
ofil
e pr
ofil
e co
nten
t pr
ofil
e pr
ofil
e pr
ofil
e ge
nici
ty
VIB
215
L
MG
132
42
D.la
br<
lx,
Spai
n 11
V
aNT
5 27
2
2 39
kb
6 30
4614
556
+ V
IB 2
16
LM
G 1
3243
D
.lab
rax,
Spa
in
VaN
T4
49
2 2
0 32
4752
757
15
+
VIB
217
L
MG
132
44
D.la
brax
, Sp
ain
VaN
Tl
49
2 2
0 1
3247
5275
7 +
V
IB 2
18
LM
G 1
3245
D
.lab
rax,
Spa
in
16
28
3 2
0 3
3006
5655
6 +
VIB
219
L
MG
132
46
roti
fers
, Sp
ain
06
2
6 2
0 1
3247
5275
7 +
V
IB 2
20
LM
G 1
3247
w
ater
, G
reec
e 1
1 2
0 1
3247
7255
7 11
+
VIB
221
L
MG
132
48
wat
er,
Gre
ece
1 2
2 0
1 32
4752
557
+ V
IB 2
22
LM
G 1
3249
D
.lab
rax.
Spa
in
06
6
1 2
44 k
b 32
4752
557
+
VIB
223
L
MG
132
50
Sp. a
llra/
a, G
reec
e 7
21
5 2
ISS
kb
6 32
4610
556
+
VIS
224
L
MG
132
51
D.l
abra
x, G
reec
e 8
05
21
11
2
0 6
3246
1255
6 +
VIB
225
L
MG
132
52
wat
er,
Gre
ece
2 21
7
2 14
7 kb
6
3246
1055
6 +
VIB
226
C
IP G
83.0
1 Sc
. max
imus
, Fr
ance
1
01
J
2 2
67 k
b 32
4752
657
+ V
IB 2
27
CII
'P03
.82
$(. m
axim
us,
Fran
ce
1 0
1
2 2
67 k
b 30
4752
677
+b
VIB
228
C
I1'1
9.22
Sp
. allr
a/a,
Fra
nce
01
1
67 k
b 32
4752
657
+ V
IB 2
29
clr 4
08F
O
. myk
iss.
Fra
nce
01
1
2 67
kb
3247
5275
7 +
V
IB 2
30
CI1
'1'0
9.80
O
. myk
iss,
Fra
nce
01
2
67
,78
kb
-32
4752
757
+ V
IB 2
31
err
07.
84
O. m
ykis
s, I
taly
01
I
67 k
b 32
4752
757
+ V
IS 2
32
ell' 0
4.88
O
. myk
iss,
Ira
ly
01
6
7k
b
3247
5265
7 +
VIB
233
ell
' 03.
88
O. m
ykis
s, I
taly
0
1
67 k
b 32
4752
757
+ V
IB 2
34
eiP
VA
408
trou
t, F
ranc
e 0
1
67 k
b 32
4752
757
+
VlB
235
el
l'V
62
D
.lab
rax,
Fra
nce
01
2
67 k
b 30
4752
457
+ V
IB 2
36
CIP
102
83
Sc. ,
nax
imll
s, F
ranc
e 1
01
2
2 67
kb
3247
7265
7 21
+
VIB
237
C
II'I
04
82
D
.lab
rax,
Fra
nce
I 0
1
2 3
6,6
7 k
b 30
4772
457
+b
VIB
238
el
P 1
0466
Sc
.max
;mIl
S, F
ranc
e 1
01
2
2 67
kb
3047
5265
7 21
+
0:
V
IB 2
39
CIP
106
21
sea
perc
h, G
reec
e 6
21
2 2
0 6
3246
1455
7 rc
+
~ V
IB 2
40
elP
/07
12
D.l
abra
x, M
orro
co
1 18
2
0 5
2246
5265
7 +
(w
eak)
n'
V
IB 2
41
ell
' 11
331
sea
perc
h, F
ranc
e 1
cros
s 2
1 2
0 1
3047
5247
7 +
~
reac
tion
s 0 ::l
V
IS 2
42
CIl
'1l1
64
Sc
. max
im liS
, Sp
ain
02
a 2
1 2
0 32
4752
757
20
+
co V
IB 2
43
NC
MB
187
5 O
. ner
ka,
U.S
.A.
01
&
2 5.
3, 1
1.5,
32
4772
757
26
+ ::
l 0
-
VaN
Tl
67 k
b -l
'<
V
IB 2
44
NC
MB
571
O
. myk
iss,
Jap
an
01
2
2 67
kb
3047
3275
7 +
'<
l
VIB
245
L
ee,
VL
3355
un
nam
ed f
ish,
U.K
. 0
1
2 2
67,
78 k
b 32
4772
657
+ S ""
VlB
246
H
WU
BA
136
Sa. t
rutta
, Ir
aly
01
2
2 67
kb
3247
5275
7 +
2.,
VlB
247
R
VA
U 1
989
salm
onid
fis
h, F
inla
nd
01
3
67 k
b 32
4772
557
30
+ <
V
IB 2
48
RV
AU
199
0 O
. myk
iss.
Fin
land
0
1
3 2
67 k
b 32
4752
757
23
+ 5- ...
VIB
249
R
VA
U 1
990
O. m
ykis
s, F
inla
nd
01
3
67 k
b 32
4752
757
+
<5
VIB
250
R
VA
U 1
990
O. m
ykis
s, F
inla
nd
01
3
67 k
b 32
4752
457
30
+ .. ~
VIB
251
C
arso
n 85
1352
6 O
. myk
iss,
Tas
man
ia
01
2
67 k
b 32
4752
757
15
+b
E.
VIB
252
C
arso
n 86
1383
1-2
O. m
yk;s
s, T
asm
ania
0
1
28
67 k
b 30
4752
757
+b
if
VIB
253
C
arso
n 85
/395
4-4
O. m
ykis
s, T
asm
ania
0
1&
2
67 k
b 32
4752
757
+b
... 1:
VaN
Tl
;:s
VIB
254
C
arso
n 89
/362
0 O
. myk
iss,
Tas
man
ia
01
2
67 k
b 9
3247
5275
7 18
+
VIB
255
C
arso
n 89
1287
3 O
. 11l
ykis
s, T
asm
ania
O
J 67
kb
2 30
4752
757
26
+ V
IB 2
56
Car
son
89/4
046
O. m
ykis
s. T
asm
ania
0
1
67
kb
1
3247
5275
7 15
+
N '"
VIB
257
C
arso
n 89
1066
O
. myk
iss,
Tas
man
ia
01
67
kb
I 32
4752
757
+
w
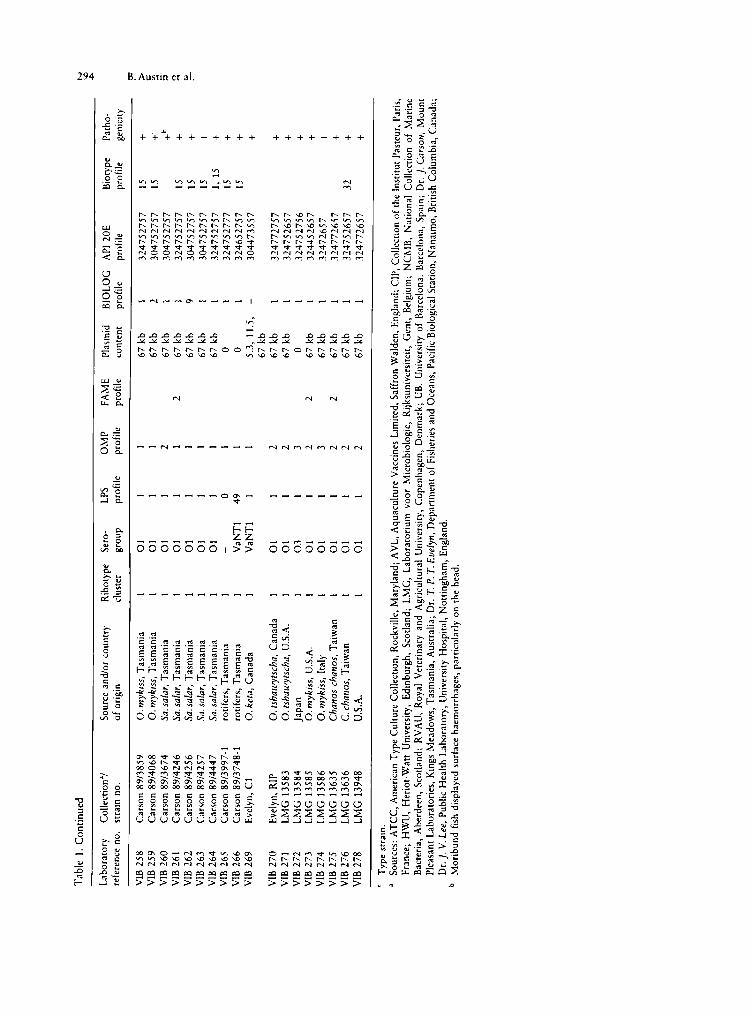
Tab
le 1
. C
on
tin
ued
t"
-c
.,. L
abor
ator
y C
olle
ctio
n'/
Sour
ce a
nd/o
r co
untr
y R
ibot
ype
Sero
-LP
S O
MP
FA~IE
Plas
mid
B
IOL
OG
A
PI2
0E
Bio
type
P
atho
-re
fere
nce
no.
stra
in n
o.
of
orig
in
dust
er
grou
p pr
ofil
e pr
ofil
e pr
ofil
e co
nten
t pr
ofil
e pr
ofil
e pr
ofil
e ge
nici
ty
?"
VIB
258
C
arso
n 89
/385
9 O
. myk
iss,
Tas
man
ia
01
1
67 k
b 1
3247
5275
7 15
+
;>
t:
V
IB 2
59
Car
son
89/4
068
O. m
ykis
s, T
asm
ania
O
J 1
67 k
b 2
3047
5275
7 15
+
'" ..
VIB
260
C
arso
n 89
/367
4 Sa
. saf
ar.
Tas
man
ia
01
1
2 67
kb
1 30
4752
757
+b
:; V
IB 2
61
Car
son
89/4
246
Sa. s
afar
, T
asm
ania
0
1
1 2
67 k
b 1
3247
5275
7 15
+
g
VIB
262
C
arso
n 89
/425
6 Sa
. saf
ar, T
asm
ania
0
1
1 67
kb
9 30
4752
757
'" 15
+
-
VIB
263
C
arso
n 89
/425
7 Sa
. saf
ar,
Tas
man
ia
01
1
67 k
b 30
4752
757
15
+
VIB
264
C
arso
n 89
/444
7 S,1
. saf
ar,
Tas
man
ia
01
1
67 k
b 32
4752
757
I, 1
5 +
V
IB 2
65
Car
son
89/3
997-
1 ro
tife
rs,
Tas
man
ia
0 0
3247
5277
7 15
+
V
IB 2
66
Car
son
89/3
748-
1 ro
tife
rs,
Tas
man
ia
VaN
TI
49
0 32
4652
757
15
+
VIB
269
E
vely
n, C
l O
. ket
a, C
anad
a V
aNT
I 1
5.3,
11.
5,
3044
7355
7 +
67
kb
VIB
270
E
vely
n, R
IP
O. t
shaw
ytsc
ha,
Can
ada
01
2
67 k
b 32
4772
757
+
VIB
271
L
MG
135
83
O. t
shaw
ytse
ha,
U.S
.A.
01
2
67 k
b 32
4752
657
+
VIB
272
L
MG
135
84
Japa
n 0
3
3 0
3247
5275
6 +
V
IB 2
73
LM
G 1
3585
O
. myk
iss,
U.S
.A.
01
2
2 67
kb
3244
5265
7 +
V
IB 2
74
LM
G 1
3586
O
. myk
iss,
Ita
ly
01
3
67 k
b 32
4726
57
+
VIB
275
L
MG
136
35
Cha
n os
chan
as,
Tai
wan
0
1
2 2
67 k
b 32
4772
657
+
VIB
276
L
MG
136
36
C. c
hano
s, T
aiw
an
01
2
67 k
b 32
4752
657
32
+
VIB
278
L
MG
139
48
U.S
.A.
Ot
2 67
kb
3247
7265
7 +
!
Typ
e st
rain
. 3
Sour
ces:
AT
CC
, A
mer
ican
Typ
e C
ultu
re C
olle
ctio
n, R
ockv
ille
, M
aryl
and;
AV
L,
Aqu
acul
ture
Vac
cine
s L
imit
ed,
Saf
fron
Wal
den,
Eng
land
; C
IP,
Col
lect
ion
of
the
Inst
itut
Pas
teur
, Pa
ri"
Fra
nce;
HW
U,
Her
iot-
Wat
t U
nive
rsit
y, E
dinb
urgh
, S
cotl
and;
LM
G,
Lab
orat
oriu
m v
oor
Mic
robi
olog
ie,
Rij
ksun
iver
site
it,
Gen
t, B
elgi
um;
NC
MB
, N
atio
nal
Col
lect
ion
of
Mar
ine
Bac
teri
a, A
berd
een,
Sco
tlan
d; R
VA
U,
Roy
al V
eter
inar
y an
d A
gric
ultu
ral
Uni
vers
ity,
Cop
enha
gen,
Den
mar
k; U
B,
Uni
vers
ity o
f B
arce
lona
. B
arce
lona
, S
pain
; D
r. j
. C
arso
t/, M
ount
P
leas
ant
Lab
orat
orie
s, K
ings
Mea
dow
s, T
asm
ania
, A
ustr
alia
; D
r. T
. P. T
. Eve
/yn,
Dep
artm
ent
of
Fish
erie
s an
d O
cean
s, P
acif
ic B
iolo
gica
l S
tati
on,
Nan
aim
o, B
ritis
h C
olum
bia,
Can
ada;
D
r. J.
V. L
ee,
Publ
ic H
ealt
h L
abor
atot
y, U
nive
rsity
Hos
pita
l, N
otti
ngha
m,
Eng
land
. b
Mor
ibun
d fis
h di
spla
yed
surf
ace
haem
orrh
ages
, pa
rric
ular
ly o
n th
e he
ad.

3. Serology
The majoriry (184/260 = 71 %) of the cultures, received as V. anglli//arum, were recovered in serogroups .01 -010, of which serogroup 01 with 95 isolates dommated (Table 1). Twenty isolates were considered to represent five previously undescribed serogroups, and are referred to
as VaNTl VaNT2, VaNT4, VaNT5 and VaNT7. A further con~plication concerned 16 cultures, of which 15 reacted with antisera representing two serogroups, and one was self-agglutinating (Table 1). The remaining 40 cultures did not react with any of the antisera used or cross reacted with more than two different antisera, and were regarded as untyped. It is noteworthy that the majority (24/40 = 60%) of the untyped isolates were not denved from fish but from anemia, rotifers, sediment and water.
Apart from serogroups 01 and 02a and 02b, which accommodated 134 (= 51 %) of the isolates from many fish species and geographical locations, smaller numbers of cultures were associated with serogroups 03 - 010 and VaNTl - VaNT7. Again, serogroups 04 and 05 with 9 and 11 isolates respectively, contained cultures obtained from a wide range of countries and habitats. In cont~ast, the 12 isolates recovered in serogroup 03 were obtamed only from fish in Denmark, Italy and Japan, and water from Denmark (Table 1). Isolates labelled as serogroups 06 and 08, with 6 and 4 isolates respectively, were obtained from many habitats in Europe. In particular, the isolates allocated to serogroup 06 were derived from Denmark and Spain. The cultures regarded as serogroup 08 were obtained from Denmark and Italy. Five isolates, recovered from fish (ayu [Plecoglossus altivelis], eel [Anguilla anguilla], sea bream [Sparus aurata] and turbot [Scophthalmus maximus] in Denmark, Italy and Japan, belonged to serogroup 07. Three isolates from cod (Gadus morhua) obtained from Denmark were equated with serogroup 09 (2 isolates) and 010 (1 isolate) (Table 1).
All of the cultures, typed in serogroups 01, 02b, 03, 06,07,08,09,010, VaNT2 VaNT5 and VaNt:7, were recovered in cluster 1 as defined by ribotyping. It IS relevant to note that VIB 76, which was regarded as representing serogroup 02a, was recovered as ribotype cluster 4; two isolates (VIB 185 and VIB 214) from serogroup 04 comprised riborype clusters 19 and 18, respectively; three cultures (VIB 23, VIB 211 and VIB 224) of serogroup 05 were defined as ribotype clusters 12, 7 and 8, respectively; and VIB 195 (serogroup VaNTI) and VIB 215 (serogroup VaNT5) were recovered in ribotype group 7 and 11, respectively. The remaining cultures ~ere unrypeable or demonstrated serological cross reactions between several serogroups. Yet, these were mostly found within ribotype cluster 1.
4. Lipopolysaccharide (LPS) profiles
There was good agreement between the composition of serogroups 01, 02, 04, 06, 07, 08, 09 and 010 and LPS profiles. Thus, all of the isolates in serogroup 01 were homogeneous in terms of their LPS profile, being equated with LPS profile 1. Interestingly, 12 isolates in LPS group 1 were recovered in serogroup 03, 07, VaNT1 and
Identification and Typing of Vibrio anguillarum 295
VaNT7 and as untypable. Serogroup 02, which was subdivided into 02a and 02b by serology, was regarded as a single LPS grouping, namely LPS profile. 2. In additio~, some of the isolates (3130 = 10%), whICh clustered 111
serogroup 02a, were equated with <;>ther LPS groups, namely LPS profile 14, 21 and 42. Five Isolates, which were ~quated with LPS profile 2, were recovered in serogroups OS, 06, VaNT2 and as untypable. It should be emphasized that these apparently ab.berant strams were recovered in cluster 1, as defined by nbotypmg (Table 1). Cultures ryped as serogroup 04 were recovered principally in LPS profile 4. Again, a minority (2/9 = 22%) were labelled as LPS profile 16 and 26. Of these, VIB 185 (serogroup 04) was recovered as a distinct single member cluster by ribotyping. Seven isolates, recovered in serogroup 03 (VIB 60-63, VIS 132, VIB 148 and VIB 272), were considered to represent LPS profiles 1, 42 and 45 (Table 1). Four cultures (VIB 22, VIB 23, VIS 211 and VIB 224), regarded as representatives of serogroup 05.' were ~quated with LPS profiles 20, 21 and 24. Indeed, It IS qu~stlonable whether these isolates are bona fide representatives of V. anguillarum.
Overall most cultures with LPS profile 1 and 2 corresponded t~ serogroups 01 and 02a and b, respectively. Nevertheless, LPS profile 1 was also recorded for cultures in serogroup 03, 07, VaNTI and VaNT7. Similarly, LPS profile 2 was noted among cultures in serogroups OS, O~ and VaNT2. Also, the distribution of the other LPS profiles traversed serogroup boundaries. Thus, it would appear that typing systems based on LPS profiles do not coincide exactly with serogroups. However, some serogroups, e.? 01 and 02, demonstrated a higher level of homology 111
terms of the LPS profile than others.
5. OMP profiles There was negligible agreement between the composi
tion of the serogroups and groupings resulting from examination of the OMP profiles. It is notable that serogroup VaNT7, in which the component organisms ~ere equated exclusively with OMP group 1, was charactenzed by the presence of a 34 kDa protein. Yet, overall, the 260 cultures were recovered in 19 groups based on OMP profiles. Cultures from serogroup 01 were equated with 4 groups based on the OMP profiles (Table 1). f:Iere, the dominant OMP profiles were 1, 2 and 3, which were characterized by the presence of dominant proteins of 33-35, 36-38 and 39-41 kDa, respectively. Again, the cultures recovered in serogroup 02 (a and b), 03, 04, OS, 06 07 and 08 were heterogeneous in terms of their OMP pro'fiIes. With serogroups 09 and 010
7 there wereinsuffi
cient cultures to make meaningful detailed compansons. It should be emphasized that representatives of the dominant OMP groups appeared in a wide range of serogroups. For example, OMP profile 1 was found in cultures from serogroups 01, 02a, 02b, 03, 05,06, 07, 08, VaNTl, VaNT2 and VaNT7 (Table 1). It was apparent that most of the cultures, recovered in ribotype clusters 2-20, represented unique OMP profiles. For example, OMP profile 15 was only recorded from VIB 214, which represented ribotype cluster 10 (Table 1).

296 B. Austin et al.
6. FAME profiles
The major fatty acids were identified as C12: 0, C12: 0 30H, C14: 0, C16: 0 iso, C16: I w7c and C16 : 0 (Table 2). From the numerical analysis of the data, all isolates, except for four, were recovered in cluster 2 as delineated at Euclidean distance .6.9. The farry acid content of the isolates was very variable (Table 2). Within cluster 2, one group of isolates, which belonged to BIOLOG cluster 6, were distinct from the other cultures insofar as they exhibited a significantly higher C12: 01C12: 0 30H ratio. However, no correlation could be found between phenotypic or serological diversity and the FAME profiles.
Cluster 1, which comprised two cultures, namely VIB 22 (serogroup 05) and VIB 24 (untyped by serology), were also phenotypically distinct and were recovered in BIOLOG cluster 8. Another two isolates, i.e. VIE 25 and VIB 204, comprised cluster 3, and were aberrant from other isolates insofar as they lacked C16: 0 iso, and possessed a higher C16 : 0 content. Nevertheless, these may be regarded as authentic V. anguillarum cultures when examined by other methods.
7. Plasmid profiles
Approximately half (137/260 = 53%) of the isolates carried plasmids. Isolates belonging to serogroups 03, 07,08,09 and 010 and VaNT4 were empty of plasm ids (Table 1). However, the majority (86/95 = 90%) of isolates in serogroup 01 contained the -67 kb virulence plasmid, equivalent to pJMl (Crosa et aI., 1980), either singly in most cases, or in association with one or two other plasm ids of 26-105 kb. The 67 kb plasmid was also found in VIE 243 and VIB 253, which demonstrated cross reactions between serogroups 01 and VaNTI, and VIB 269, which was labelled as VaNT1. Although most (32139 = 82%) of the isolates in serogroup 02 (a and b) were empty of plasm ids, 7 cultures contained one or two plasmids of 5->200 kb (Table 1). Again, the minority (2/9 = 22 %) of isolates recovered as serogroup 04 possessed single plasm ids of either 32 or 49 kb. From serogroup OS, 5 isolates (45%) contained one (4 isolates) or two (1 iso-
Table 2. Major fatty acids of Vibrio anguilla rum
Fatty acid
C12:0 C12:0 30H C14:0 C16:0 iso C16:1OJ7c C16:0 l:F3 LF7
Ratios:
C12:0/14:0 CI2:0/12:0 30H CI6:0/16:1 w7c
3.1-6.4 1.7-3.4 4.1-7.9 2.4-3.7
30.4-39 18.4-25.4 2.2-4.5 9.6--21.5
0.79 1.88 0.63
Cluster:
2
1.4-7.1 0.1-4.8 3.4-10.1 0.1-10.5
34.1-44.4 17.5-28.6
1.3-5.3 9.3-22.0
0.72 1.62 0.62
3
7.4-8.2 4.5-6.0 8.4-11.8 0
41.2-43.2 41.5-43.2 5.16-6.4 5.6-7.3
0.77 1.42 0.46
late) plasmids of 2.6-120 kb. Two cultures (33%) from serogroup 06 contained one (44 kb) or four plasmids (3.2-70 kb). Isolates from the new serogroups were most-
9
8
7
6
5 4
:3
2
100 90 80 70 60 so 40
Fig. 2. Simplified dendrogram based on the examination of BIOLOG data by Pearson product correlation and UPGMA clustering.

ly devoid of plasmids. However, two (40%) of VaNTl isolates comprised two (53 and 56 kb) and three (5.3, 11.5 and 67 kb) plasm ids, respectively. Again, 2 (40%) of the isolates from VaNT2 comprised single (11.5 kb) or two (68 and 83 kb) plasmids. A single (50%) isolate from VaNT5 contained a 39 kb plasmid. All the cultures from VaNT7 contained a 16 kb plasmid. In addition, VIB 199 and VIB 202 both contained a 49 and a 68 kb plasmid. The majority of the serologically untyped cultures were empty of plasm ids. Expressed in terms of ribotyping, it was apparent that cultures from ribotypes 3, 4, 6, 8, 10, II, 12, 14, 16 and 20 were empty of plasm ids. However, a wide range of plasmid profiles was recorded from some of the isolates (Table 1).
8. BTOLOG fingerprints
Nine clusters were delineated at 78% r among the 260 isolates, using the Pearson correlation coefficient (Fig.2). All isolates tested in the BIOLOG GN-microplates were positive for oxidation of dextrin, glycogen, N-aceryl-Dglucosamine, D-fructose, D-glucose and maltose. Conversely, all isolates were negative for adonitol, D-arabitol,
Table 3. Tests differemiating the clus-Test: ters as defined after numerical analysis
using the Pearson correlation coeffi-cient and UPGlvlA clustering of BIOLOG fingerprints Negative control
Cyclodextrin N-acetyl-D-galactosamine L-arabinose Gentobiose m-inositol a-D-Iactose D-mannose j)-methylglucoside D-sorbitol mono-methyl-succinate Acetic acid D-gluconic acid Succinic acid Bromosuccinic Alaninamide acid L-alanine L-alanylglycine L-aspartic acid Glycyl-L-aspartic acid Glycal-L-glutamic acid L-histidine L-proline L-threonine Uridine Thymidine Glycerol D,L-a-glycerolphosphate Glucose-I-phosphate Glucose-6-phosphate
Identification and Typing of Vibrio anguillarum 297
erythrirol, L-fucose, lactulose, D-melibiose, D-raffinose, turanose, xylitol, formic acid, galactonic acid lactone, Dgalacruronic acid, D-glucosaminic acid, y-hydroxybutyric acid, p-hydroxy-phenylacetic acid, itaconic acid, a-ketovalerie acid, malonic acid, D-saccharic acid, sebacic acid, L-leucine, L-phenylalanine, L-pyroglutamic acid, D,L-carnitine, y-amino-butyric acid, urocanic acid, phenyl-ethylamine, putrescine, 2-amino-ethanol and 2,3-butanediol. The differentiating characters (which were set at a discriminating level of 89%) between each cluster has been given in Table 3. The clusters did not possess any unique single features permitting differentiation from all other groups. Nevertheless, the number of characters needed to be used to distinguish between each of the delineated clusters has been presented in Fig. 3. From this diagram, it is apparent that dusters I, 2, 8 and 9 could not be distinguished from each other. Clusters 2 and 9 each comprised two isolates deemed to be from serogroup 01. These clusters were similar to the majoriry of V. anguillarum isolates studied by other methods.
In common with the results of riboryping, the majority of the isolates, particularly those regarded as belonging in serogroup 01 (= 90%) and including the type strain, were
Cluster:
2 3 4 5 6 7 8 9
.B
54 + 50 20 + + + 20 33 60
65 + 83 20 33 + 33 50 + 33 + 80 + 47 + 20 + 25 17 20 + 20 + + 83 + + + + + + + + 60 + 80 + + + 50 80 33 60 + 85 50 17 + 80 + 76 50 17 67 60 33 + + + + 50 80 67 20 + + + + 17 67 87 + + + 85 50 17 20 + 80 + 73 + 60 + 65 + 20 + + + + + 83 + 40 + + + + + + + + + + + + 68 + 67 + + + 50 20 50 + 33 60 53 + 33 + + + 67 + + + + 30 50 + + 67 + 85 50 33 + + + + 63 67 + + 67 + + + 33 + + + + + 84 + 20 + 60 + + + 67 40 80 + + + + + 33 60 80 + + + . '+' and '-' correspond [Q >89% and <11% of positive responses. respectively. The
numbers correspond to the percentage of positive responses.

298 B. Austin et al.
Cluster: 1 2 3 4 5 6 7 8 9
1 0 2 0 0 3 2 9 0 4 4 8 1 0 5 4 9 5 6 0 6 2 8 8 9 2 0 7 2 7 7 7 9 5 0 8 0 3 6 9 4 1 1 0 9 Q Q B 12 1a 1~ 9 :2 Q
Fig. ]. Number of tests differentiating the 9 clusters defined by examination of the BIOLOG fingerprints.
recovered in a single large cluster 1, which accomodated 205 (= 79%) cultures. This group had an intragroup homogeneity of 80%. Yet, the membership of BIOLOG cluster 1 was not identical to that of the ribotype cluster 1. Thus, 21 of the isolates from ribotype cluster 1 were recovered in BIOLOG clusters 2, 4, 5, 6, 7,8 and 9 (Table 1). Of the other isolates in serogroup 01, these were equated with BIOLOG fingerprint clusters 2, 4 and 9, whereas VIB 18 and VIB 28 were unclustered. Interestingly, BIOLOG cluster 1 contained representatives of all serogroups.
Serologically untyped isolates, which were mostly recovered from water and live food, i.e. Artemia and rotifers, were grouped in cluster 3. These isolates corresponded to ribotype clusters 5, 13, 16, 17 and 20. The four isolates in BIOLOG cluster 2 were equated with serogroups 02 and 09. Cluster 4 contained 5 isolates of untyped V. anguillarum and representatives of serogroups 01 and 02a from Japan, U.K. and U.S.A. One of these isolates corresponded to ribotype cluster 4.
The three isolates of cluster 5 were obtained from sea bass, and were untyped by serology. One of these isolates belonged to ribotype cluster 15, whereas the other represented ribotype cluster l.
Most isolates in cluster 6 were isolated from Greece. Yet only a few of the isolates could be typed by serology, and were equated with serogroups OS, VaNTI and VaNT5. These isolates were dispersed among ribotype clusters 2, 6, 7, 8, 9, 10 and 11. Interestingly, the isolates in cluster 7 were recovered from water and rainbow trout (Oncorhynchlls mykiss; VIB 142) in Denmark. Although most of these isolates were untyped by serology, VIB 142 was regarded as a member of serogroup VaNT2. Two of the isolates of cluster 7 constituted ribotype cluster 14.
A geographical pattern was also apparent with the origin of the four isolates in cluster 8, being obtained from sea bass (Dicmtrarchus labrax) in Greece and Artemia in Spain. These isolates were either untyped or represented serogroups 04 and 05. Two of these isolates constituted ribotype clusters 12 and 19.
Overall, the isolates regarded as belonging to serogroup 02 were biochemically diverse, being distributed among BIOLOG clusters 1 and 4. Similarly, the isolates equated with serogroups 04, OS, 09, VaNT2 and VaNT5 were
recovered in many clusters based on BIOLOG profiles. Isolates, which were untyped by serology, were found in clusters 1,3,5,6,7 and 8.
9. API 20E profiles
Overall, the cultures of V. anguillarum were fermentative, motile, produced arginine dihydrolase, I~-galactosidase, indole and oxidase, but not lysine or ornithine decarboxylase, degraded gelatin but no·t urea, reduced nitrate to nitrite but not to nitrogen, gave a positive response to the Voges Proskauer reaction, and produced acid from arabinose, mannitol and saccharose, but not from amygdalin, inositol or melibiose. Yet marked heterogeneity was recorded in the individual API 20E profiles for V. anguilfarum. Thus, 117 different profiles were recorded, of which '324772757', '324752757', '324772557', '304752757' and '324772757' accounted for 112 (= 43%) of the isolates (Table 1). Generally, there was little relationship between these profiles and the composition of the serogroups. For example, the cultures of serogroup 01 were represented by 25 API 20E profiles, of which '324752757' was dominant. Yet, this profile was the most common, being recorded for 55 isolates representing 11 serogroups. Indeed, this API 20E profile predominated among the isolates of serogroups 03, 04 and 010 (Table 1). Thus, the API 20£ profiles permitted certain differentiation among the isolates of V. anguillarum. Yet, this differentiation could not be equated with the other typing methods used in this study. Nevertheless, it was apparent that the most extreme API 20£ profiles belonged to cultures which were equated with ribotypes other than 1. For example, the profile '000411456' was recorded from isolates of ribotype 15 (Table 1).
10. Biotyping
The minority (106/260 = 41 %) of the cultures were recovered in 32 clusters, which were defined at or above the 80% similarity level using the data for Biotype-100 (Table 1). The Biotype clusters were defined as groups of strains (or isolated strains) with identical (dissimilarity = 0) nutritional patterns. The remaining 154 isolates comprised single members clusters, each with a separate Biotype profile. However, these isolates did not use many substrates, possibly reflecting the lack of sufficient growth factors in the media. The differentiating characteristics between the clusters have been included in Table 4. It was apparent that the majority of isolates utilized a wide range of carbon compounds in four days, including N-acetyl-Dglucosamine, L-aspartate (1 exception), D-fructose, Dgluconate (1 exception), D-glucose, L-glutamate, L-malate (1 exception), D-mannitol, D-mannose, 1-0-methyl-/3-Dglucoside, L-proline and sucrose. Conyersely, the isolates were uniformly negative in their ability to utilize transaconitate, adonitol, 4-aminobutyrate, 5-aminovalerate, Darabitol, L-arabitol, benzoate, betaine, caprate, caprylate, m-coumarate, dulcitol, i-erythritol, aesculin, ethanolamine, L-fucose, D-galacturonate, gentobiose, gentisate, D-glucuronate, glutarate, histamine, L-histidine, 3-hydroxybenzoate, 4-hydroxybenzoate, 3-hydroxybutyrate,

Idenriiication and Typing of Vibrio angllillamm 299
Table 4. Tests diiferenriating the c1usrers as defined by use of biotyping
Carbon conraining compound·':
Cluster:
1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32
4
+ + + + + + + + + + + + + + + +
+
+ + + + + + + +
14
+
+
+
+ +
16
+ + + +
+ + + + + + + + + + + + + + + + + +
20
+
+
+ + + + +
+ + + + + + +
+ + + + + + + + + + + + +
21
+ + +
+
+ +
+
+ +
+ + + + + + + + +
+
+
+ + + +
37
+ + + +
+ + + + + + + + + + + + + + + + + + + + +
+ +
49
+ +
+ +
+ +
76
+ + + + + + + + + + + + + + + + + +
+ + + +
+ + + + + +
93
+ + + + + + +
+ + + +
+ + + +
+ + +
+
+ + +
+
94
+ + + + + + + + + + + + +
+ + + +
+ + +
+
+ + +
95
+ + + + + + + + + + + + +
+ + + +
+ + + + + + + + + + + +
98
+ +
99
+ +
+ +
+
a 4, 14, 16, 20, 21, 33, 37,49, 76, 93, 94, 95, 98 and 99 correspond to D-trehalose, 1-0-methyl-l3-galactopyranoside, D(+) cellobiose, D( -) ribose, L( +) arabinose, myo-inositol, D-sorbitol, D( +) malate, DL-Iactate, D-alanine, L-alanine, L-serine, L-serine, Ltyrosine and a-ketoglutarate, respectively.
hydroxyquinoline-B-glucuronide, itaconate, 2-ketogluconate, 5-ketogluconate, lactose, lactulose, D-Iyxose, malonate, maltitol, D-melezitose, D-melibiose, 1-0-methyl-agalactoside, 3-0-methyl-D-glucose, 1-0-methyl-a-Dglucoside, mucate, palatinose, phenylacetate, 3-phenylpropionate, propionate, protocatechuate, putrescine, quinate, D-raffinose, L-rhamnose, D-saccharate, L-sorbose, D-tagatose, D-tartrate, L-tartrate, meso-tartrate, tricarballylate, trigonelline, tryptamine, tryptophan, Dturanose, xylitol and D-xylose. Differential results, In
terms of the % positive response to the utilization of the carbon containing compounds, was recorded for cisaconitate (91 %), D-alanine (72%), L-alanine (75%), Larabinose (60%), D-cellobiose (77%), citrate (94%), fumarate (91%), D-galactose (98%), D-glucosamine (89%), DL-glycerate (91 %), glycerol (97%), myo-inositol (9%), 2-ketoglutarate (8%), DL-Iactate (83%), D-malate (14%), maltose (95%), maltotriose (96%), 1-0-methyl-f3-
galactoside (6%), D-ribose (86%), L-serine (85%), D-sorbitol (91 %), succinate (95%), D-trehalose (72%) and Lryrosine (2%).
11. Pathogenicity
The majority (226/260 = 87%) of the isolates were pathogenic to salmonids at a dose of 105 - 106 cells/fish, as determined by the development of overt clinical signs of disease and evenrual mortalities. Overall, high proportions of the isolates from serogroup 01 (96%), 02a (93%), 02b (100%), 03 (75%), 06 (100%), 07 (80%), 08 (100%), 10 (100%), VaNTl (100%), VaNT4 (100%), YaNT5 (100%) and VaNT7 (100%) and unrypable (83 %) were pathogenic. In contrast, less isolates from serogroup 04 (55%), 05 (54%),09 (40%) and YaNT2 (40%) demonstrated pathogeniciry. Of the non-pathogenic culrures, only 4 (12%) contained plasmids. Thus,

300 B. Austin et al.
VIB 88, VIB 93, VIB 166 and VIB 212 contained 67 kb and 105 kb, 67 kb, 2.6 kb, and 37 kb plasmids, respectively. It should be emphasised that other cultures, which did not contain plasm ids, were pathogenic. By comparing pathogenicity with the ribotype, it was apparent that 100% of the isolates from ribotype 2-8 and 10-20 were pathogenic. In contrast, pathogenicity levels of 98% and 0% was recorded for the isolates from ribotype 1 and 9, respectively. Generally, the pathology was consistent with septicaemia, namely the development of gastro-enteritis, ascites, haemorrhaging in the musculature, and liquefaction of the internal organs, especially the kidney. Some cultures (total = 26), representing a diverse range of serogroups and origin, resulted in the development of extensive surface haemorrhaging, particularly on the head and in the mouth (Table 1).
Discussion
Overall, this study sought to clarify the identificationl typing of isolates, considered to be bona fide represent,l[ives of V. anguillarum. The strengths of the investigation centred on the large collection of bacterial strains, which were obtained from a diverse range of geographical locations, hosts and environmental samples. However, the influence of the different methods of analyses of the data obtained in this study is largely unknown (Priest and Austin, 1993). In contrast to previous phenetic based studies, e.g. Kaper et al. (1983), the bulk of these isolates were recovered in a single homogeneous group, as defined by ribotyping. Undoubtedly, this group (cluster 1) constituted authentic isolates of V. angui/larum. It is accepted that the other clusters as defined by ribotyping, i.e. clusters 2-20, may not, in fact, be V. anguillarum. The taxonomic position of these small number of isolates must await further investigation. Yet, it was apparent that the various methods did not give congruent results. For example, some of the other clusters defined by ribotyping could be equated with existing serogroups of V. anguillamm. In short, the various methods enabled recognition of core members of the taxon, but there was a problem with boundary definition. The greatest discrepancies occurred with the non-typed isolates (by serology), some of which must be considered to be on the periphery of the species. In contrast, the highest congruence in results occurred with the bulk of the isolates, which were considered to represent serogroup 01. This is arguably the best understood of all the subgroups of V. allguilfarum (Pedersen and Larsen, 1993; Austin and Austin, 1993). Thus, the majority of these isolates belonged to LPS group 1, possessed the 67 kb plasmid, which was found to be associated with virulence (Crosa et aI., 1980), were recovered in the same BIOLOG cluster, i.e. 1, and were highly pathogenic to fish. However, a small number of isolates from serogroup 01 possessed the virulence plasmid but were not pathogenic. Moreover, some isolates were pathogenic and did not possess the 67 kb plasmid. Thus, in contrast to previous studies, it is apparent that there is not an absolute correlation between serogrouping and LPS profiles (Aoki
et aI., 1981) and plasmid profiles and virulence (Crosa et aI., 1980).
The validity of separating serogroup 02 into 02a and 02b must be questioned in view of some of the data from this study, notably the results of the LPS analysis which could not distinguish between the sub-groups. This has also been reported by Mutharia et al. (1993).
The LPS of Gram-negative bacteria corresponds to the O-antigen, and inevitably forms the basis of serogrouping. Therefore, a close relationship between serogroup and LPS profile may usually be anticipated. However in the present study, this association was most evident with certain serogroups, i.e. 01, 02, 04, and 06-010. With the other serogroups, the correlation to LPS profile was lower. Indeed, in serogroups 03, VaNTl, VaNT4 and VaNT5, the isolates belonged to separate LPS profiles. An interpretation of these results is that certain LPS antigens have immunological cross-reactivity. Alternatively, the bacteria may contain common heat-stable antigens, other than LPS. This possibility needs further investigation. Clearly, serogrouping is not always adequate for predicting the nature and composition of the LPS of V. anguilla rum, insofar as some isolates revealed identical typing patterns with the O-antisera but possessed different LPS profiles. Overall, the pool of serologically non-typable isolates were the most diverse in terms of LPS profiles. This indicates that there are additional serogroups still to be defined. Interestingly, only 78% of the isolates, examined in this study, could be typed by serology, which is a lower proportion than previously reported (Larsen et aI., 1994). A possible explanation may reflect the comparatively large number of environmental isolates examined here in compari~on to previolls studie~.
The discriminatory power of plasmid profiling was generally low. Therefore, it is conceded that plasmid profiles are of minor value for typing V. anguillarum. An increased discriminatory value of plasmid patterns, particularly for representatives of serogroup 01, may be achieved by restriction enzyme analysis of isolated plasmids (Olsell and Larsen, 1990). Certainly, the association of the 67 kb plasmid with serogroup 01 is in agreement with previous work (Wolf and Crosa, 1986). The presence of this plasmid in the previously undescribed serogroup, coined VaNTl, and the frequent cross-reaction between serogroups 01 and VaNTl, raises the possibility that the new group is merely a variant/sub-group of serogroup 01. This needs further investigation. The presence of a 67 kb plasmid encoding an iron sequestering mechanism, consisting of a siderophore and an outer membrane protein, OM2, has long been considered to explain the virulence of isolates of serogroup 01 (Crosa, 1980), with curing of the plasmid leading to a marked increase in LDso values (Crosa et aI., 1980). Yet, it should not be overlooked that in the present study nine isolates, equated with serogroup 01, were either non- or weakly pathogenic. Three isolates contained the 67 kb plasmid. The reason for the lack of virulence by these few cultures is unclear. Conceivably, the iron sequestering mechanism may not have been expressed.
The pathogenicity of V. anguilla rum for fish, especially salmonids, is well established (Austin and Austill, 1993).

The association of some serogroups, i.e. 01, 02 and 03, with epizootics has been reported (S(Jrensen and Larse'l, 1986; Larsen et aI., 1994). Indeed, the data from this study have highlighted the virulence of representatives of these serogroups. An inference that other serogroupS are mostly non-pathogenic and comprise environmental isolates is partially supported by this study, insofar as only approximately half of the cultures from serogroups 04, OS, 09 and YaNT2 caused mortalities in Atlantic salmon. Of course, it may be argued that the apparently nonpathogenic isolates could well cause disease in other fish species under less favourable environmental conditions (= stress) (Austin and Austin. 1993).
Overall, the number of profiles obtained with the API 20E rapid identification system greatly exceeded previous studies (Kent, 1982; Maugeri et aI., 1983; Grisez et aI., 1991), and, together with the results of biotyping demonstrated the biochemical heterogeneity of V. anguillarum. Yet, this was not so clear with BIOLOG, which revealed a smaller number of profiles. It should be emphasised that caution needs to be exercised when using systems designed for other uses. Thus, the API 20E system was originally developed for medically important isolates of the family Enterobacteriaceae, which grew at 37°C within 48 h. Yet for non-human isolates, notably V. anguillarum as used in this study, lower incubation temperatures often need to be adopted. Thus, the comparison of data to schemes designed for use with medically important bacteria may well give rise to erroneous identifications. However, a good identification of V. anguillarum has been achieved by using the results of the API 20E rapid identification system with specially formulated keys for vibrios (Alsina and Blanch, 1994a, b).
It will remain for further work to clarify the precise taxonomic position of V. anguilla rum and its relationship to the genus Listonella. Yet, the validity of the genus Listonella has been thrown into doubt when V. damsela was reclassified to Listonella (MacDonell and Colwell, 1985) and thence to Photo bacterium (Smith et aI., 1991). V. anguillarum has been considered to be more closely related to V. pelagius than to organisms with the specific epithet of damsela in terms of ribotyping (Olsen and Larsen, 1993). The current view from this study is that strains with the specific epithet of anguillarum would be better suited in the genus Vibrio, as V. anguillarum.
As a result of this study, the characteristics of typical isolates of V. anguillarum may be amended from the description in Baumann et al. (1984), to:
Gram-negative, motile fermentative, rod-shaped cells, which produced arginine dihydrolase, catalase, f)-galactosidase, HlS, indole and oxidase but not lysine or ornithine decarboxylase or tryptophan deaminase, reduced nitrate to nitrite but not to nitrogen, were positive for the Yoges-Proskauer reaction, degraded gelatin but not urea, utilized citrate, and produced acid from amygdalin, arabinose, glucose, sorbitol and sucrose but not from inositol, mannose, melibiose or rhamnose. Dextrin, D-fructose, glycogen, N-acetyl-D-glucosamine, D-fructose, maltose and sucrose were utilized, but not so adonitol, Darabitol, erythritol. L-fucose, lactulose, D-mannose, D-
Identification and Typing of Vibrio anguillarum 301
melibiose, D-raffinose, turanose, xylitol, formic acid, galactonic acid lactone, D-galacturonic acid, Dglucosamininic acid, y·hydroxybutyric acid, p-hydroxyphenylacetic acid, itaconic acid, a-keto-valeric acid, malonic acid, D-saccharic acid, sebacic acid, L-leucine, Lphenylalanine, L-pyroglutamic acid, D,L-carnitine, yamino-butyric acid, urocanic acid, phenylethylamine, putrescine, 2-amino-ethanol or 2,3-butanediol. Growth occurred on MacConkey agar. Cultures comprised a single dominant ribotype, but may be typed in any of a diverse range of serogroups, of which representatives of serogroup 01 may contain a 67 kb plasmid associated with viru· lence. Two dominant groups have been defined with LPS profiles. OMP were typically of 33-40 KDa in size. Pathogenicity was exhibited to fish.
Acknowledgement. Financial support was provided by CEC Contract No: AIR-Cf92-0341. We are grateful to colleagues for the gift of cultures.
References
Alsina, M., Blanch, A. R.: A set of keys for biochemical identifi· cation of environmental Vibrio species. J. Appl. Bacteriol. 76, 79-85 (1994a)
Alsina, M., Bla"ch, A. R.: Improvement and update of a set of keys for biochemical identification of Vibrio species. J. Appl. Bacterial. 77, 719-721 (1994b)
Aoki, T., Kitao, T., [tabashi, T., Wada, Y., Sakai, M.: Proteins and lipopolysaccharides in the membrane of Vibrio anguillarum. Devel. BioI. Stand. 49, 225-232 (1981)
Austin, B., Austin D. A.: Methods for the Microbiological Examination of Fish and Shellfish. Chichester, England, Ellis Horwood (1989)
Austin, B., Austin, D. A.: Bacterial Fish Pathogens Disease of Farmed and Wild Fish, 2nd edn. Chichester, England, Ellis Horwood (1993)
Baumann, P., Furniss, A. L., Lee, J. V.: Genus 1. Vibrio Pacini 1854, 411AL, pp. 518-538. In: Bergey's Manual of Systematic Bacteriology, Vol. 1 (N. R. Krieg, J. G. Holt, eds.). Baltimore, Williams & Wilkins (1984)
Crosa, J. H.: A plasmid associated with virulence in the marine fish pathogen, Vibrio anguillarum specifies an iron sequester· ing system. Nature (London) 284, 566-568 (1980)
Crosa,J. H., Hodges, L. L., Schiewe, M. H.: Curing of a plasmid is correlated with attenuation of virulence in the marine fish pathogen Vibrio anguillarum. Infect. Immun. 27, 897-902 (1980)
DeBoer, S. H., Sasser, M.: Differentiation of Erwinia carotovora ssp. carotovora and E. carotovora ssp. atroseptica on the basis of cellular fatty acid composition. Can. J. Microbiol. 32, 796-800 (1986)
Grimont, F., Chevrier, D., Grimont, P. A. D., Lefevre, M., Gllesdon, J. L.: Acetyiaminofluorene-iabelled ribosomal RNA for use in molecular epidemiology and taxonomy. Res. Microbiol. 140,447-454 (1989)
Grisez, L., Ceusters, R., Ollevier, F.: The use of AP[ 20E for the identification of Vibrio anguilla rum and V. ordalii. J. Fish Dis. 14,359-365 (1991)
Hitchcock, P. J., Brown, T. M.: Morphological heterogeneity among Salmonella lipopolysaccharide chemotypes in silverstained polyacrylamide gel. J. Bacteriol. 154, 269-277 (1983)

302 B. Austin et al.
Kado, C. T., Lill, S.-T.: Rapid procedure for detection and isolation of large and small plasmids. J. Bacterial. 145, 1365-1373 (1981)
Kaper, J. B., Lockman, H., Remmers, E. F., Kristensen, K., Colwell, R. R.: Numerical taxonomy of vibrios isolated from estuarine environment. Int. J. System. Bacterial. 33, 229-255 (1983)
Kent, M. L.: Characteristics and identification of Pasteurella and Vibrio species pathogenic to fishes using API-20E (Analytab Products) multitube test strips. Can. J. Fish. A'Iuat. Sci. 39, 1725-1729 (1982)
Kitao, T., Aoki, T., Fllkodome, M., Kawano, K., Wada, Y., MiZllno, Y.: Serotyping of Vibrio anguillarum isolated from diseased freshwater fish in Japan. J. Fish Dis. 6, 175-181 ( 1983)
Kittclberger. R .• Hilbink. F.: Sensitive silver-staining detection of bacterial lipopolysaccharides in polyacrylamide gels. J. Biochern. Biophys. Ivlethods 26,81-86 (1993)
Kllsuda, R .. Sako, H., Kawai. K.: Classification of vibrios isolated from diseased fishes - 1. On the morphological, biological and biochemical properties. Fish Pathol. /3, 123-137 (1979)
Lambert. P. A.: Isolation and purification of outer membrane proteins from Gram-negative bacteria, pp. 110-125. In; Bacterial Cell Surface Techniques (I. Hancock. I. Poxton. eds.). Chichester, England, John Wiley & Sons (1986)
Larsen, J. L., Olsen. J. E.: Occurrence of plasm ids in Danish isolates of Vibrio angllillarum serovars Oland 02 and association of plasm ids with phenotypic characteristics. Appl. Environ. Microbiol. S7, 2158-2163 (1991)
Larsen. J. L.. Pedersen, K .• D'l/sgaard, 1.: Vibrio anguilla rum serovars associated with vibriosis in fish. J. Fish Dis. 17, 259-267 (1994)
Maniatis, T., Fritsch. E. F., Sambrook, J.: Molecular cloning; a laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1982)
MacDonell, M. T., Colwell, R. R.: Phylogeny of the Vibrionaceae and recommendations for two new genera, Listonella and Shewanella. System. Appl. Microbial. 6, 171-182 (1985)
MacDonell, M. T., Swartz, D. G., Ortiz-Conde, B. A., Last, G. A., Colwell, R. R.: Ribosomal RNA phylogenies for the vibrioenteric group of eubacteria. Microbiol. Sci. 3, 172-179 (1986)
Maugeri, T. L., Crisafi, E., Genovese, L., Scoglio, M. E. R.: Identification of Vibrio angllillarum with the API-20E system. Microbiologica I, 73-79 (1983)
Merrill. C. R., Goldman, D., Sedman. S. A .• Ebert, M. H.: UItrasensitive stain for proteins in polyacrylamide gcls shows regions in cerebrospinal fluid proteins. Science 21,1437-1438 (1981)
Muroga. K., Yama11Oi, H., Hiranaka, Y., Yamamoto, S .. Tatani. M .. Io, Y .• Takahashi, S., Hanada. H.: Detection of Vibrio
anguilla rum from wild fingerlings of ayu Plecoglosslis altive/is. Bull. Jap. Soc. Sci. Fish. 50, 591-596 (1984)
Mutharia, L. W., Raymond, B. T.. Dekie~'it. T. R .• Stevenson, R. M. W.: Antibody specificities of polyclonal rabbit and rainbow trout antisera against Vibrio ordalii and serotype 0; 2 strains of Vibrio angllillarum. Can. J. ~1icrobiol. 39, 492-499 (1993)
Nybelin, 0.: Untersuchungen iiber den bei Fischen krankheitserregenden Spaltpilz Vibrio anguillarttm. Med. Stat. Undersok. Fors. Anst. Sotvat. Stockholm 8, 1-62 (1935)
Olsen, j. E., Larsen, j. L.: Restriction fragment length polymorphism of the Vibrio anguilla rum serovar 01 virulence plasmid. Appl. Environ. Microbiol. S6, 3130-31>2 (1990)
Olsen, j. E., Larse11, j. L.: Ribotypes and plasmid contents of Vibrio anguillaru»l strains in relation to serovar. App!. Environ. Microbiol. 59, 3863-3870 (1993)
Pedersen, K., Larsen, J. L.: rRNA gene restriction patterns of Vibrio anguilla rum serogroup 01. Dis. Aquat. Org. 16, 121-126 (1993)
Priest, F. G., Austin, B.: Modern Bacterial Taxonomy, 2nd. edn. London, Chapman & Hall (1993)
Rochelle, P. A., Fry, J. c., Day, M. j., Bale, M. J.: An accurate method for estimating sizes of small and large plasmids and DNA fragments by gel electrophoresis. J. Gen. Microbiol. 132, 53-59 (1985)
Smith, I. W.: A disease of finnock due to Vibrio anguillarum. J. Gen. Microbiol. 74, 247-252 (1961)
Smith, S. K., Sutton, D. C, Fuerst, J. A., Reichelt. j. L.: Evaluation of the genus Listonella and reassignment of Listonella damsela (Love et al.) MacDonell and Colwell to the genus Photobacterium as Photobacterium damsela comb. nov. with an emended description. Int. J. Syst. Bacteriol. 41, 529-534 (1991)
Sneath, P. H. A .• Johnson, R.: The influence on numerical taxonomic similarities of errors in microbiological tests. J. Gen. Microbiol. 72, 377-392 (1972)
Sneath, P. H. A., Sokal, R. R.: Numerical Taxonomy: The Principles and Practice of Numerical Classification. San Francisco, W. H. Freeman (1973)
Sorensen. U. B .• Larsen. j. L.: Serotyping of Vibrio anguillarum. Appl. Environ. Microbiol. SI, 593-597 (1986)
West, P. A., Lee, j. V., Bryant, T. N.: A numerical taxonomic study of species of Vibrio isolated from the aquatic environment and birds in Kent, England. J. Appl. Bacteriol. SS, 263-282 (1983)
Wiik, R., Hoff. K., Andersen. K., Da,/e. F. L.: Relationship between plasm ids and phenotypes of presumptive strains of Vibrio anguilla rum isolated from different fish species. Appl. Environ. Microbiol. 55, 826-831 (1989)
Wolf, M. K., Crosa, j. H.: Evidence for the role of a siderophore in promoting Vibrio anguillamm infections. J. Gen. Microbiol. 132,2949-2952 (1986)
Professor B. Austin (Department of Biological Sciences, Heriot-Watt University, Riccarron, Edinburgh EH14 4AS, Scotland UK)