characterization of a porin from the outer membrane of vibrio anguillarum
TRANSCRIPT

JOURNAL OF BACTERIOLOGY, July 1996, p. 4182–4188 Vol. 178, No. 140021-9193/96/$04.0010Copyright q 1996, American Society for Microbiology
Characterization of a Porin from the Outer Membraneof Vibrio anguillarum
MARTA SIMON,1 ANTON MATHES,2 ANICET BLANCH,1 AND HARALD ENGELHARDT2*
Department of Microbiology, University of Barcelona, 08028 Barcelona, Spain,1 and Max-Planck-Institut furBiochemie, D-82152 Martinsried, Germany2
Received 4 March 1996/Accepted 7 May 1996
The outer membranes of the 10 serovars of Vibrio anguillarum showed a common major protein with a sizeof around 40 kDa. Antibodies against the major outer membrane protein (MOMP) of V. anguillarum AO18(serovar O1) cross-reacted with the MOMPs of all the other serovars but not with the outer membrane proteinsof Escherichia coli. The MOMP of V. anguillarum serovar O1 was isolated, reconstituted to two-dimensionalcrystals, and structurally characterized by electron microscopy and image processing. The unit cell structureof the crystalline MOMP, as well as the secondary structure composition of the protein with a high amount ofb-structure, is strongly reminiscent of that of bacterial porins. The functional properties of the pores wereinvestigated by conductance measurements with the MOMP reconstituted in planar lipid membranes. The V.anguillarumMOMP is characterized by a relatively weak cation selectivity and a moderate surface charge, andit shows voltage-dependent conductance effects. The MOMP is functionally similar to OmpF from E. coli, andit can be classified as a general diffusion porin.
Vibrio anguillarum, a gram-negative bacterium, is the etio-logical agent of vibriosis, one of the most serious infectiondiseases of salmonid and marine fish species throughout theworld (2). The diagnosis and control of V. anguillarum infec-tions are of great economical importance in commercial fishfarming.Since the first descriptions of V. anguillarum infections,
many studies on the immunological characterization of thisspecies and its pathogenicity have been performed. The mo-lecular basis of these studies is closely related to the composi-tion and function of the outer membrane (OM). Most studiesof the V. anguillarum OM have been focused on the lipopoly-saccharides (LPS) which are the major agents for antigeneticspecificity (11). Several serotyping systems based on the slideagglutination method with thermostable O-antigens were de-scribed independently (29, 47). These serovars are consideredan important epizootical characteristic. The three main sero-vars O1, O2, and O3 (according to the classification of Sør-ensen and Larsen [47]) include most of the strains isolatedfrom fish infections. The remaining serovars are more com-monly isolated from the natural environment and other aquaticspecies (47). Nevertheless, there is also an increasing interestin the OM proteins of V. anguillarum which may participate inmechanisms of pathogenicity directly. So, special attention hasbeen paid the OM proteins involved in siderophore-dependentiron uptake (13, 34). Several studies analyzed and comparedthe compositions of OM proteins from different serovars (1,40). They describe the presence of usually one major OMprotein (MOMP) in the molecular size range of about 30 to 40kDa in all the strains tested. Recently, Suzuki et al. (48) sug-gested that this MOMP represents a porin because of its mo-lecular weight.In the present work, we have studied the OM protein pro-
files of different serovars of V. anguillarum. The MOMP of onestrain of V. anguillarum serovar O1 was isolated and was char-
acterized as a porin. We describe here the two-dimensional(2D) crystallization of the protein and the image processing ofelectron micrographs of the MOMP and present data on theconductance measurements addressing the pore-forming activ-ity of the OM protein.
MATERIALS AND METHODS
Bacterial strains and growth conditions. Bacterial collection strains used inthis study were the reference serovar strains of V. anguillarum ATCC 43305(serovar O1), ATCC 43306 (serovar O2), ATCC 43307 (serovar O3), ATCC43308 (serovar O4), ATCC 43309 (serovar O5), ATCC 43310 (serovar O6),ATCC 43311 (serovar O7), ATCC 43312 (serovar O8), ATCC 43313 (serovarO9), and ATCC 43314 (serovar O10) and Escherichia coli ATCC 11303. We usedstrain AO18 of V. anguillarum serovar O1 from our own strain collection (6) forthe purification of the MOMP.Bacteria were cultured in tryptic soy broth supplemented with 1.5% NaCl.
They were incubated overnight at 238C on an orbital shaker. V. anguillarumAO18 was grown in a pilot fermentor to obtain large quantities of the MOMP.To prepare the inoculum for the 50-liter fermentation, the bacteria were sub-cultured in successive volumes of 50 ml and 1 liter of tryptic soy broth–1.5%NaCl at 218C until stationary state. The 50-liter fermentation was performed inthe same medium at 238C for 19 h with agitation (200 rpm). Cells were harvestedby continuous centrifugation at 5,690 3 g for 2 h (Westfalia Separator CSA1-06-475).OM extraction. Bacterial cells from overnight cultures were collected by cen-
trifugation at 10,000 3 g for 10 min at 48C and were washed twice with 10 mMTris-HCl buffer at pH 8.0 containing 0.3% NaCl. The cell suspensions weredisintegrated by sonication, with two 100-W pulses being applied for 1 min eachpulse at 48C. The suspension was centrifuged at 16,000 3 g for 2 min to removeintact cells. The cell fragments were sedimented at 16,000 3 g for 60 min at 48C.The OM fractions were obtained with 0.75% sodium lauryl sarcosinate in 10 mMTris-HCl (pH 8.0)–0.3% NaCl as previously described (23).Electrophoresis. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) was performed by the method of Laemmli (32), with a 4% stackinggel and a 12% separating gel being used. The OM extractions were dissolved insample buffer and heated at 908C before being loaded onto the gels. Gels werestained first with Coomassie brilliant blue G-250 and second with silver nitrateaccording to the procedure of Morrisey (37). The molecular weights of theproteins were determined with the standard low-molecular-weight markers fromBio-Rad Laboratories.Antiserum and immunoblotting. In order to obtain rabbit antibodies against
the MOMP, the protein was isolated from gels after SDS-PAGE by means ofelectroelution (24). After it was stained with Coomassie brilliant blue G-250, thegel fragment containing the band of the MOMP was cut off and extracted in anelectroeluter (Bio-Rad) at 60 mA for 4 h and 48C, with an elution buffer (25 mMTris-HCl [pH 8.3], 192 mM glycine, 0.1% SDS) being used. The concentration ofthe electroeluted protein was 34.4 mg/ml. The purity of the isolated MOMP wascontrolled by SDS-PAGE. A rabbit was immunized by two subcutaneous injec-
* Corresponding author. Mailing address: Max-Planck-Institut furBiochemie, D-82152 Martinsried, Germany. Phone: 49/89/8578 2650.Fax: 49/89/8578 2641. Electronic mail address: [email protected].
4182
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
03
Febr
uary
202
2 by
191
.240
.117
.52.

tions of the purified MOMP (0.5 ml each) that were administered once a weekfor 4 weeks. Freund’s complete adjuvant and Freund’s incomplete adjuvant wereused the first and second weeks, respectively. Bleeding and serum preparationwere performed as previously described (24). The antibody titer was determinedby an enzyme-linked immunosorbent assay as outlined previously (22). Westernblotting (immunoblotting) of the extracted OMs was performed with the MiniTrans-Blot (Bio-Rad) according to the method of Towbin et al. (49) and with atransfer buffer (25 mM Tris, 192 mM glycine, 20% methanol [pH 8.1 to 8.4]).Immunoblotting was performed with the antiserum against the MOMP as de-scribed previously (41). The transferred low-molecular-weight markers werestained with amido black (24).Purification of the MOMP. The MOMP was purified from OMs of V. anguil-
larum AO18. The OM fraction prepared for 2D crystallization experiments wasobtained from the 50-liter culture essentially by the process specified above, withthe exceptions that 3 mM NaN3 was added in the 10 mM Tris-HCl buffer andsamples were stored at 48C. The cell wall preparation was incubated with ly-sozyme, bacitracin, DNase, and RNase to remove the peptidoglycan and nucleicacids. The remaining OMs were solubilized in 20 mM HEPES (N-2-hydroxyeth-ylpiperazine-N9-2-ethanesulfonic acid), pH 7.5, with 1.3% octyl polyoxyethylene(octyl-POE; BACHEM Heidelberg). The OM proteins were separated by gelfiltration with a Sephacryl S-300 HR column according to the procedure ofEngelhardt et al. (20) in 20 mM HEPES buffer, pH 7.5, plus 0.5% octyl-POE.The fractions containing the MOMP were pooled and concentrated by ultrafil-tration. The presence of LPS in the purified MOMP preparation was analyzed bySDS-PAGE and staining with silver nitrate (30). The MOMP was concentratedby means of ultrafiltration to a protein concentration of 1.9 mg/ml plus 1.1% 60.1% octyl-POE in 20 mM HEPES buffer, pH 7.5. The concentration of theMOMP was determined by the bicinchoninic acid assay (46). The concentrationof the detergent was assessed by measuring the diameters of 10-ml dropletsdeposited on a hydrophobic surface (39). For conductance measurements, aportion of the preparation was used for anion-exchange chromatography with aMonoQ column (Pharmacia) equilibrated with 10 mM methylamine buffer, pH8.5, plus 0.5% octyl-POE. The solubilized protein (2 mg) was applied to thecolumn and eluted with a NaCl gradient (0 to 2 M). The fractions containingprotein were freed from salt by dialysis and again used for chromatography, witha gradient of 0 to 400 mM being used for elution.Crystallization of the MOMP. The solubilized MOMP was two-dimensionally
crystallized in reconstituted membranes by detergent removal in controlled di-alysis. In order to optimize the crystallization conditions, several experimentswere performed in a multichamber dialysis apparatus, which allows up to sevensamples to be treated independently in a single experiment as previously de-scribed (39). In all the samples, the protein concentration was adjusted to 1mg/ml and the initial octyl-POE content was adjusted to 1%. The dialysis bufferconsisted of 20 mM HEPES, pH 7.5, plus 3 mM NaN3. The effect of additionallipid (dimyristoyl phosphatidylcholine [DMPC]) was studied at different concen-trations and at a constant temperature of 308C. The DMPC-to-protein ratio wasvaried between 0 and 1 (wt/wt). The temperature, the pH value, and the presenceof different concentrations of cations (Ca21 and Mg21) were also studied tooptimize the crystallization conditions.Electron microscopy and image processing. Purified OM preparations and
crystallized protein assemblies were examined with a Philips CM12 electronmicroscope, with a nominal magnification of 335,000 being used. The sampleswere applied on carbon-coated copper grids and were negatively stained withunbuffered uranyl acetate (2%). The electron micrographs were analyzed in anoptical diffractometer to monitor defocus and astigmatism conditions and toassess the quality of the crystalline arrays. Areas (1,024 3 1,024 pixels) of theselected micrographs were digitized with an Eikonix model 1412 camera system(Eikonix Corporation) with a pixel size of 15 mm, corresponding to 0.44 nm at thespecimen level. Image processing was done with the SEMPER 6.2 system (44).The 2D crystalline arrays were analyzed by means of correlation averaging (43).FTIR spectroscopy. Infrared (IR) spectra of the protein preparations were
recorded with a Nicolet 740 Fourier transform IR (FTIR) spectrophotometer bythe attenuated total reflection method. Between 20 to 100 mg of protein whichhad been dialyzed against water was applied to a germanium crystal and airdried. A total of 1,024 scans were averaged, and the spectra were analyzed in theamide I and II regions by means of Fourier self-deconvolution and band fitting(31) in order to assess the relative amounts of secondary structure elements. Thecarbonyl stretching vibration at around 1,720 cm21 was taken to assess the lipidcontent of the samples.Conductance measurements. Conductance measurements were performed
with a planar lipid membrane system essentially by following the techniquedescribed previously (7, 8). The lipid membranes were prepared according to themethod of Montal and Mueller (36). The current was amplified by a headstageequipped with a 200-MV and a 20-GV feedback resistor connected to an EPC-7patch clamp amplifier. The signals were monitored and stored by means of acomputer using the modular data acquisition system LabView (National Instru-ments). The special programs and procedures required for long-term measure-ments and recording of voltage-current curves, etc., were developed by us (35).Single-molecule measurements were performed at $20 mV in 100 mM KCl
buffered with 10 mM Tris-HCl, pH 8.3. Characteristically, 10 ng of proteindissolved in 1 M KCl plus 1% Genapol was applied to one side of the chamberhaving two compartments of 1 ml each. Voltage-to-current curves were obtained
by applying a slow (25-min) triangle voltage stimulus; the voltage was continu-ously changed from 0 to 2125 mV, increased to 1125 mV, and finally decreasedto 0 again. The selectivity of the porin was assessed according to the methoddescribed previously (5, 9), with black lipid-type membranes made of the samelipid mentioned above being used. Salt gradients from 0.01 to 1 M KCl wereapplied in a stepwise manner, and the voltages developed across the membranewere determined after equilibrium conditions were reached. The relative per-meability was calculated by means of the Goldman equation (5, 9).
RESULTS
OM protein profiles. The OMs of the 10 serovars of V.anguillarum showed considerable heterogeneity by SDS-PAGE(Fig. 1). Each strain exhibited a particular protein pattern,especially with the minor components. However, all the pro-tein profiles of V. anguillarum strains had a common charac-teristic, i.e., the presence of one MOMP with an apparentmolecular mass in the range from 35 to 42 kDa. The proteinprofiles of the serovar O1 strains studied (ATCC 43305 andAO18) were quite similar, showing an MOMP with an appar-ent size of 40 kDa as a common constituent. The antiserumagainst the MOMP of V. anguillarum AO18 (serovar O1) rec-ognized the MOMPs of the type collection strains of the 10serovars of V. anguillarum. No cross-reactions were observedwith proteins extracted from the OM of E. coli ATCC 11303(Fig. 2).Purification and crystallization of the MOMP. The OM
proteins were hardly completely separated by gel filtration.The MOMP was eluted in the main peak. Other minor pro-teins were still present in a very low proportion, which, how-ever, did not prevent the crystallization studies. Apparently, notrace of free LPS was detected in the MOMP fractions, asjudged by silver staining and SDS-PAGE.The reconstitution of the solubilized MOMP by removing
the detergent in a controlled manner in the dialysis apparatusyielded vesicles and membranous sheets with regularly arrayedproteins. The samples with DMPC adjusted to a protein-to-lipid ratio of up to 1 (wt/wt) contained membranes and vesicleswhich were not crystalline. This result denotes that the lipidcontent was too high to support the crystallization of the pro-tein.Differences in the sizes and shapes of the reconstituted
membranes were determined by the temperature chosen forthe experiments. Long cylindrical forms (0.2 to 0.5 by 3 to 10
FIG. 1. SDS-PAGE of the extracted OMs from the V. anguillarum serovars.A, molecular weight markers (from top to bottom in kilodaltons: 97.4, 66.2, 45.0,31.0, 21.5, and 14.4); B, ATCC 43305 serovar O1; C, ATCC 43306 serovar O2;D, ATCC 43307 serovar O3; E, ATCC 43308 serovar O4; F, ATCC 43309serovar O5; G, ATCC 43310 serovar O6; H, ATCC 43311 serovar O7; I, ATCC43312 serovar O8; J, ATCC 43313 serovar O9; K, ATCC 43314 serovar O10.
VOL. 178, 1996 PORIN OF VIBRIO ANGUILLARUM 4183
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
03
Febr
uary
202
2 by
191
.240
.117
.52.

mm) were obtained predominantly below 308C, and smallspherical vesicles ('0.1 mm in diameter) were more abundantabove 308C. The quality of the regular arrays was also affectedby the temperature. The largest crystals (5 to 10 mm2) wereobtained between 26 and 308C. The solubilized protein precip-itated at pH 5 in the dialysis experiments, forming amorphousaggregates. Crystalline tubular reassembly products and sheetswere obtained at pH 7.5 and 9. The addition of Ca21 and Mg21
ions to the dialysis buffer, often supporting the formation ofregular arrays (27), yielded multilamellar liposomes with dif-ferent sizes (0.1 to 0.3 mm) in our experiments. This effect wasincreased with higher cation concentrations.The best results were obtained at pH 9 with the original
preparation without any addition of DMPC and cations, indi-cating that residual lipid, probably intimately associated withthe protein, was sufficient for the regular reconstitution. Mostof the membranes were double- or multilayered. For imageprocessing, single-layered membranes were selected.Structure of the MOMP in 2D crystals. No regular arrange-
ment was detected for native OMs observed with an electronmicroscope. The crystalline arrays of the MOMP in the recon-stituted membrane sheets, however, revealed a typical unit cellstructure after image processing (Fig. 3). All the images ana-lyzed showed the same type of diffraction pattern, with reflec-tions usually out to the sixth diffraction order in the opticaldiffractograms (Fig. 3). The lattice parameters are as follows:a 5 14.0 6 0.1 nm, b 5 12.3 6 0.1 nm, and g 5 908. Theorthogonal lattice type belongs to a p2-related symmetrygroup. The arrangement of the trimeric structures, which arenot equivalent in neighboring positions (Fig. 3), the existenceand the positions of symmetry and mirror axes, and the (al-most) systematically missing reflections of the type 0k0, wherek is odd numbered, are consistent with a p2212 2D space group,with the corresponding symmetry group being pmg (for a re-view, see reference 18). The crystallographic unit cell containsfour identical molecules, each of them obviously representinga trimeric structure. Neighboring molecules are oppositely in-corporated with respect to the membrane plane. In the diffrac-togram, very weak 010 reflections (and 300 reflections) whichshould actually be absent according to the plane group sug-gested are detectable. This result might be due to an inhomo-geneous stain distribution on the two surfaces of the mem-brane, a phenomenon that is frequently observed with
negatively stained preparations supported by a carbon film(25). Accordingly, neighboring trimers are not absolutely mir-ror symmetric but are distinguishable by minor staining differ-ences (Fig. 3).The crystallographic resolution is 2.0 nm, with the reflection
spot (k 5 4 and h 5 5) being taken as a measure. The reso-lution assessment by means of the radial correlation functioncriterion (43) provides a value of 1.4 nm, which is apparentlythe limit that can be expected for negatively stained prepara-tions (19).The architecture of the trimeric molecular unit and its di-
mensions (diameter, '6.5 nm) are strongly reminiscent ofthose of the pore-forming protein complexes in the OMs ofbacteria, i.e., the porins (17, 21). The heavily stain-filled areasrepresent the triplets of pores, the bright structures surround-ing the pores originate mainly from the additional stain-exclud-ing protein portions outside the membrane plane, and thegreyish areas between pore triplets apparently represent thesurface of the lipid (Fig. 3).FTIR spectroscopy of the MOMP. Attenuated total reflec-
tion IR spectroscopy of the purified MOMP revealed a spec-trum typical for porins (Fig. 4). Characteristic features are thestrong absorption at 1,631 cm21, indicative of a considerablecontent of b-sheet structure (31), and the prominent shoulderat 1,697 cm21, which originates from b-turns. Fourier self-deconvolution, band form analysis, and quantitative assess-ment of the secondary structure composition suggest a b-struc-ture content of about 60%. This value is in agreement withthose for other porin molecules (9, 12, 31). Other typical fea-tures are the prominent peak at 1,515 cm21 and the compo-nent at 1,615 cm21, both of which originate from Tyr, i.e., thecharacteristic constituent of the rings of aromatic amino acidsin the 16-stranded b-barrel (12).The region between 1,700 and 1,740 cm21 is indicative for
residual lipid, i.e., for the stretching vibration of the carbonylgroups of the fatty acids bound in phospholipids and LPS. Thevery weak signal (Fig. 4) corresponds to a low content of lipid,probably LPS. The relative strengths of the lipid and proteinbands were compared with those for appropriate mixtures oftest lipids and concanavalin A, a b-structure protein. Thiscomparison suggests that the molar ratio of porin trimers tocarbonyl groups is on the order of about 1:10. Assuming fourcarbonyl groups per LPS, the ratio was approximately 2 to 3LPS molecules per porin trimer. The lipid content was evensmaller in preparations further purified by chromatographyand might not exceed a ratio of 1:1.Conductance measurements. The solubilized MOMP of V.
anguillarum caused a stepwise increase of membrane conduc-tance after the protein was added to the salt solution bathinga planar lipid bilayer (Fig. 5A). The histogram of these con-ductance steps, which originate from insertion of the pore-forming protein complexes into the membrane, shows a majorpeak at 0.2 to 0.25 nS for a single event in 100 mM KCl (Fig.5B).The histogram as well as the current trace in Fig. 5 illustrate
that frequently more than one pore-forming unit entered themembrane. A relatively clear maximum of conductance stepswhich might be attributable to the insertion of three complexeswas obtained at 0.6 to 0.7 nS. Assuming Gaussian distributions,the histogram in Fig. 5B revealed average conductance valuesof 0.22, 0.45, and 0.61 nS, with the last value having a some-what larger standard deviation. But there were even stepswhich apparently originated from six or more MOMP com-plexes. The 0.22-nS step is very likely due to the insertion of atrimeric pore-forming complex and not a single channel only.We occasionally observed conductance steps in the range of
FIG. 2. Western blot of the extracted OMs of V. anguillarum serovars withantiserum against the MOMP of V. anguillarum AO18. A, marker proteins (fromtop to bottom in kilodaltons: 97.4, 66.2, 45.0, 31.0, and 21.5); B, ATCC 43305serovar O1; C, ATCC 43306 serovar O2; D, ATCC 43307 serovar O3; E, ATCC43308 serovar O4; F, ATCC 43309 serovar O5; G, ATCC 43310 serovar O6; H,ATCC 43311 serovar O7; I, ATCC 43312 serovar O8; J, ATCC 43313 serovarO9; K, ATCC 43314 serovar O10; L, E. coli ATCC 11303; M, V. anguillarumAO18.
4184 SIMON ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
03
Febr
uary
202
2 by
191
.240
.117
.52.

FIG. 3. (A) Electron micrograph of a 2D crystal of the MOMP from V. anguillarum, with the crystal being reconstituted by means of the dialysis technique at pH9.0 and 268C and in the absence of any additional lipid. (B) Quasioptical diffractogram of an area, 0.44 by 0.44 mm square, that was digitized for image processing andthat came from the membrane central to the single-layered crystal shown in panel A. The lattice parameters are as follows: a 5 14.1 nm, b 5 12.4 nm, and g 5 908for this particular 2D crystal. (C) Correlation average of 984 unit cells, containing 3,936 trimeric molecules, i.e., 4 molecules per unit cell. Neighboring trimers areoriented oppositely to each other with respect to the membrane plane.
VOL. 178, 1996 PORIN OF VIBRIO ANGUILLARUM 4185
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
03
Febr
uary
202
2 by
191
.240
.117
.52.
0.05 to 0.1 nS, which is approximately one-third of the majorvalue. The distribution of conductance steps obtained with aporin preparation containing residual LPS was indistinguish-able from that shown in Fig. 5.The average conductance in 10 mM KCl was 0.055 nS. Sim-
ilar to that in the histogram in Fig. 5, a prominent peak oftriple events was obtained. The ratios of conductance over thebulk KCl conductivity at 10 and 100 mM KCl are 3.9 3 1028
and 1.7 3 1028 cm, respectively. The ratio should be constantif the salt concentrations in the porin channel and the electro-lyte solutions are always the same. The relatively higher valueat low salt concentrations may denote that ions are accumu-lated near the pore entrance (9).Selectivity of the MOMP. The ion selectivity of the V. an-
guillarumMOMP was assessed by means of KCl concentrationgradients across the porin-doped black lipid membrane. Themembrane potentials developed at zero-current conditionswere taken to calculate the ratio of cation permeability toanion permeability (5, 9). The MOMP behaved consistentlycation selectively in all the salt solutions tested (values wereaveraged from at least two experiments). Because of the largerdiameter of the hydrated cation with LiCl, the apparent selec-tivity was lower than that found with KCl (cation/anion per-meability ratio of 2.3). Because of the larger anion of potas-sium acetate, the selectivity value was higher than that foundwith KCl (cation/anion permeability ratio of 15). A ratio of >4for KCl identifies the MOMP of V. anguillarum as a moder-ately cation-selective porin.Voltage-dependent behavior of the MOMP. The current-to-
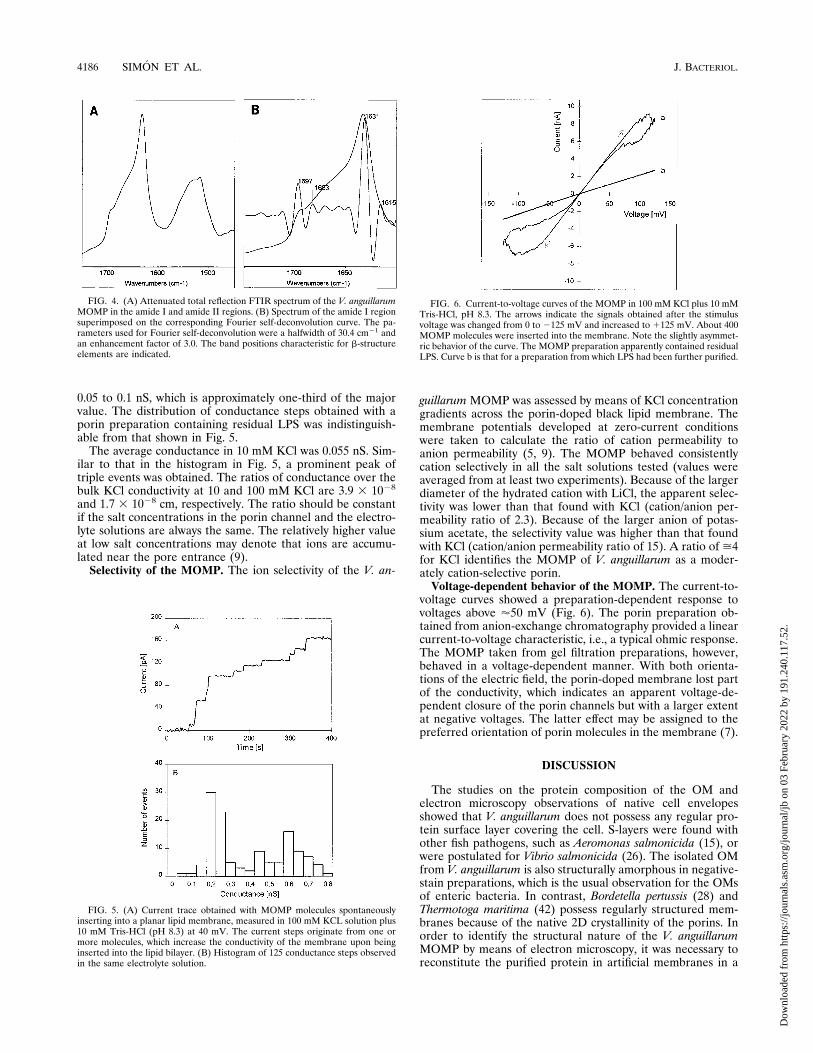
voltage curves showed a preparation-dependent response tovoltages above '50 mV (Fig. 6). The porin preparation ob-tained from anion-exchange chromatography provided a linearcurrent-to-voltage characteristic, i.e., a typical ohmic response.The MOMP taken from gel filtration preparations, however,behaved in a voltage-dependent manner. With both orienta-tions of the electric field, the porin-doped membrane lost partof the conductivity, which indicates an apparent voltage-de-pendent closure of the porin channels but with a larger extentat negative voltages. The latter effect may be assigned to thepreferred orientation of porin molecules in the membrane (7).
DISCUSSION
The studies on the protein composition of the OM andelectron microscopy observations of native cell envelopesshowed that V. anguillarum does not possess any regular pro-tein surface layer covering the cell. S-layers were found withother fish pathogens, such as Aeromonas salmonicida (15), orwere postulated for Vibrio salmonicida (26). The isolated OMfrom V. anguillarum is also structurally amorphous in negative-stain preparations, which is the usual observation for the OMsof enteric bacteria. In contrast, Bordetella pertussis (28) andThermotoga maritima (42) possess regularly structured mem-branes because of the native 2D crystallinity of the porins. Inorder to identify the structural nature of the V. anguillarumMOMP by means of electron microscopy, it was necessary toreconstitute the purified protein in artificial membranes in a
FIG. 4. (A) Attenuated total reflection FTIR spectrum of the V. anguillarumMOMP in the amide I and amide II regions. (B) Spectrum of the amide I regionsuperimposed on the corresponding Fourier self-deconvolution curve. The pa-rameters used for Fourier self-deconvolution were a halfwidth of 30.4 cm21 andan enhancement factor of 3.0. The band positions characteristic for b-structureelements are indicated.
FIG. 5. (A) Current trace obtained with MOMP molecules spontaneouslyinserting into a planar lipid membrane, measured in 100 mM KCL solution plus10 mM Tris-HCl (pH 8.3) at 40 mV. The current steps originate from one ormore molecules, which increase the conductivity of the membrane upon beinginserted into the lipid bilayer. (B) Histogram of 125 conductance steps observedin the same electrolyte solution.
FIG. 6. Current-to-voltage curves of the MOMP in 100 mM KCl plus 10 mMTris-HCl, pH 8.3. The arrows indicate the signals obtained after the stimulusvoltage was changed from 0 to 2125 mV and increased to 1125 mV. About 400MOMP molecules were inserted into the membrane. Note the slightly asymmet-ric behavior of the curve. The MOMP preparation apparently contained residualLPS. Curve b is that for a preparation from which LPS had been further purified.
4186 SIMON ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
03
Febr
uary
202
2 by
191
.240
.117
.52.

regular manner. The various dialysis experiments performedrevealed that the best 2D crystallizations were obtained whenno lipid was added to the particular protein preparation underscrutiny. This result was surprising, considering the experi-ences with the 2D crystallization of other porins (17, 21) andother membrane-bound (27) or membrane-anchored (39) sur-face proteins. But the V. anguillarum MOMP appeared tocluster even in presence of detergent, as the conductance stepanalysis suggested, which may reflect the protein’s inherentpropensity to form regular aggregates. The content of residualLPS was quite low and might not have been higher than 1 LPSper porin monomer, which is in accordance with the fact thatno traces of LPS were detected in silver-stained PAGE gels.We assume that it was this low LPS content which led to thestrong tendency of the preparation to form clusters and regular2D arrays. Yamada and Mizushima (50) showed that at least 1LPS molecule per porin trimer was required for the formationof 2D crystalline arrays. The formation of crystals with p2212symmetry and alternating orientations of the porin trimers isreminiscent of three-dimensional crystals of other porins, forwhich alternating orientations of neighboring porin trimersalso occurred along a certain crystal plane (12). The unit cellstructure of the V. anguillarumMOMP is very similar to that ofother bacterial porins reconstituted in 2D crystals (17, 27). Thesame is true for the secondary structure composition, with theb-structure content being about 60%, which is a commonstructural principle for bacterial porins (12).The functional properties identify the V. anguillarum
MOMP as a general diffusion porin exhibiting a relatively weakcation selectivity and apparently possessing a moderate surfacecharge. The latter can be concluded from the relatively higherconductance value in low salt concentrations compared withthat in higher KCl concentrations. Similar but sometimesstronger phenomena were observed with other porins, e.g.,with the Acidovorax delafieldii porin Omp34 (9). The selectivityfor cations in KCl solution is about 4, i.e., cations diffuse fourtimes more frequently across the membrane than do chlorideions. This value is close to the value for the cation selectivity ofOmpF from E. coli, i.e., 3.8 (5). The average conductivity in 0.1M KCl, too, is approximately the same for the V. anguillarumand E. coliOmpF porins, i.e., 0.22 and 0.25 nS, respectively (4).Suzuki et al. (48) discussed how the antigenetic 36-kDa OMprotein of Listonella anguillara (probably identical to that of V.anguillarum [16]) is an OmpC-type porin because of its size andextraction properties. The porin investigated here, however,has more functional similarities with OmpF of E. coli than withOmpC, which is a strongly cation-selective porin (3).The V. anguillarum porin behaves in a voltage-dependent
manner, like the other porins tested under appropriate condi-tions (33). The asymmetry and hysteresis effects are typicalproperties of the voltage current curves (7). The most inter-esting observation is that the voltage-dependent effects weremissing after the use of a porin preparation which had beenpurified by means of anion-exchange chromatography in addi-tion. It is difficult to determine exactly what parameter wasresponsible for the loss of the ability to switch between appar-ently open and closed states. But it is likely that the LPScontent of the preparation matters. FTIR measurementsshowed that the LPS-to-protein ratio was significantly reducedin the preparations after chromatography. That LPS may alterthe voltage-dependent behavior of E. coli porins has alreadybeen observed (10). LPS is probably an important parameter inaddition to others, for voltage sensing with porins, as discussedby Lakey and Pattus (33) and Brunen and Engelhardt (7). It is,however, unclear whether other conditions, such as divalentcations bound to the porin molecules or ligands, were of im-
portance. Previous experiments with A. delafieldii porinOmp34 did not reveal significant effects of Ca21 or Mg21 onthe voltage-dependent properties (7). Whether the voltage-dependent effects have any significance for the physiologicalrole of the V. anguillarum porin cannot be concluded from ourexperiments. Sen et al. (45) demonstrated that the Donnanpotential across the OM had no effect on the E. coli porins inintact cells. But recently, de la Vega and Delcour (14) foundevidence that polyanions may regulate voltage-dependent be-havior in vitro and in vivo.Immunoblotting showed that the 10 serovars of V. anguilla-
rum do have common immunogenic characteristics. This ob-servation is in agreement with previous reports of commonantigens in the OMs of the serovars (48). The presence ofsimilar epitopes in the MOMP of the different serovars couldexplain this result, though the MOMPs of each serovar showeddifferent mobilities by SDS-PAGE.The identification of the MOMP of V. anguillarum as a porin
allows a better understanding of the role of this protein in themechanisms of pathogenicity related to resistance to b-lactamantibiotics (38) and its role as a common surface antigen (11).Further studies on the antigenicity of the MOMP are necessaryin order to evaluate its potential use for diagnosis and vaccineformation.
ACKNOWLEDGMENTS
We thank Jan Lubieniecki for his valuable help with the fermenta-tion of cells and the purification of the protein and Wilk von Gustedtfor his advice on the 2D crystallization experiments.Part of this work was supported by a grant from the Deutsche
Forschungsgemeinschaft, Sonderforschungsbereich 266/D4. M. Simonhad a fellowship from CIRIT, Generalitat de Catalunya.
REFERENCES1. Aoki, T., T. Kitao, T. Itabashi, Y. Wada, and M. Sakai. 1981. Proteins andlipopolysaccharides in the membrane of Vibrio anguillarum. Dev. Biol.Stand. 49:225–232.
2. Austin, B., and D. A. Austin. 1993. Bacterial fish pathogens: diseases infarmed and wild fish, 2nd ed. Ellis Horwood, Chester, England.
3. Benz, R., and K. Bauer. 1988. Permeation of hydrophilic molecules throughthe outer membrane of gram-negative bacteria. Eur. J. Biochem. 176:1–19.
4. Benz, R., K. Janko, W. Boos, and P. Lauger. 1978. Formation of large,ion-permeable membrane channels by th matrix protein (porin) of Esche-richia coli. Biochim. Biophys. Acta 511:305–319.
5. Benz, R., K. Janko, and P. Lauger. 1979. Ionic selectivity of pores formed bythe matrix protein (porin) of Escherichia coli. Biochim. Biophys. Acta 551:238–247.
6. Blanch, A. R., and J. Jofre. 1992. Isolamento di Vibrio anguillarum sierotipoO1 in un focolaio di vibriosi di muggine dorato (Mugil auratus) nel delta delfiume Ebre (Catalogna, Spagna). Boll. Soc. Ital. Patol. Itt. 9:17–23.
7. Brunen, M., and H. Engelhardt. 1993. Asymmetry of orientation and voltagegating of the Acidovorax delafieldii porin Omp34 in lipid bilayers. Eur. J. Bio-chem. 212:129–135.
8. Brunen, M., and H. Engelhardt. 1995. Significance of positively chargedamino acids for the function of the Acidovorax delafieldii porin Omp34.FEMS Microbiol. Lett. 126:127–132.
9. Brunen, M., H. Engelhardt, A. Schmid, and R. Benz. 1991. The major outermembrane protein of Acidovorax delafieldii is an anion-selective porin. J.Bacteriol. 173:4182–4187.
10. Buehler, L. K., S. Kusumoto, H. Zhang, and J. P. Rosenbusch. 1991. Plas-ticity of Escherichia coli porin channels. Dependence of their conductance onstrain and lipid environment. J. Biol. Chem. 266:24446–24450.
11. Chart, H., and T. J. Trust. 1984. Characterization of the surface antigens ofthe marine fish pathogens Vibrio anguillarum and Vibrio ordalii. Can. J.Microbiol. 30:703–710.
12. Cowan, S. W., T. Schirmer, G. Rummel, M. Steiert, R. Ghosh, R. A. Pauptit,J. N. Jansonius, and J. P. Rosenbusch. 1992. Crystal structures explainfunctional properties of two E. coli porins. Nature (London) 358:727–733.
13. Crosa, J. H., and L. Hodges. 1981. Outer membrane proteins induced underconditions of iron limitation in the marine fish pathogen Vibrio anguillarum775. Infect. Immun. 31:223–227.
14. de la Vega, A. L., and A. H. Delcour. 1995. Cadaverine induces closing of E.coli porins. EMBO J. 14:6058–6065.
15. Dooley, J. S. G., H. Engelhardt, W. Baumeister, W. W. Kay, and T. J. Trust.
VOL. 178, 1996 PORIN OF VIBRIO ANGUILLARUM 4187
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
03
Febr
uary
202
2 by
191
.240
.117
.52.

1989. Three-dimensional structure of an open form of the surface layer fromthe fish pathogen Aeromonas salmonicida. J. Bacteriol. 171:190–197.
16. Dorsch, M., D. Lane, and E. Stackebrandt. 1992. Towards a phylogeny of thegenus Vibrio based on 16S rRNA sequences. Int. J. Syst. Bacteriol. 42:58–61.
17. Dorset, D. L., A. Engel, M. Haner, A. Massalski, and J. B. Rosenbusch. 1983.Two-dimensional crystal packing of matrix porin. J. Mol. Biol. 165:701–710.
18. Engelhardt, H. 1988. Correlation averaging and 3-D reconstruction of 2-Dcrystalline membranes and macromolecules, p. 357–413. In F. Mayer (ed.),Methods in microbiology, vol. 20. Academic Press, London.
19. Engelhardt, H. 1991. Electron microscopy of microbial cell wall proteins.Surface topography, three-dimensional reconstruction, and strategies fortwo-dimensional crystallization, p. 11–25. In J. P. Latge and D. Boucias (ed.),Fungal cell wall and immune response. NATO ASI Series H53. Springer-Verlag, Berlin.
20. Engelhardt, H., S. Gerbl-Riegen, D. Krezmar, S. Schneider-Voss, A. Engel,and W. Baumeister. 1990. Structural properties of the outer membrane andthe regular surface protein of Comamonas acidovorans. J. Struct. Biol. 105:92–102.
21. Engelhardt, H., T. Scheybani, W. von Gustedt, and W. Baumeister. 1994.Formation of two-dimensional crystals of membrane-anchored and water-soluble proteins, p. 201–208. In H. Bayley, D. Kaplan, and M. Navia (ed.),Biomolecular materials by design, vol. 330. Materials Research Society,Pittsburgh.
22. Fibla, J. 1990. Analisis immunoquımico de la alcohol-deshidrogenasa deDrosophila. Doctoral thesis. University of Barcelona, Barcelona, Spain.
23. Filip, C., G. Fletcher, J. L. Wulff, and F. Earhart. 1973. Solubilization of thecytoplasmic membrane of Escherichia coli by the ionic detergent sodiumlauryl sarcosinate. J. Bacteriol. 115:717–722.
24. Harlow, E., and D. Lane. 1988. Antibodies. A laboratory manual. ColdSpring Harbor Laboratory, Cold Spring Harbor, N.Y.
25. Harris, J. R., H. Engelhardt, S. Volker, and A. Holzenburg. 1993. Electronmicroscopy of human erythrocyte catalase: new two-dimensional crystalforms. J. Struct. Biol. 111:22–33.
26. Hjelmeland, K., K. Stensvag, T. Jorgensen, and S. Espelid. 1988. Isolationand characterization of a surface layer antigen from Vibrio salmonicida. J.Fish Dis. 6:175–181.
27. Jap, B. K., M. Zulauf, T. Scheybani, A. Hefti, W. Baumeister, U. Aebi, andA. Engel. 1992. 2D crystallization: from art to science. Ultramicroscopy46:45–84.
28. Kessel, M., M. J. Brennan, B. L. Trus, M. E. Bisher, and A. C. Steven. 1988.Naturally crystalline porin in the outer membrane of Bordetella pertussis. J.Mol. Biol. 203:275–278.
29. Kitao, T., T. Aoki, M. Fukudome, K. Kawano, Y. Wada, and Y. Mizuno. 1983.Serotyping of Vibrio anguillarum isolated from diseased freshwater fish inJapan. J. Fish Dis. 6:175–181.
30. Kittelberger, R., and F. Hilbink. 1993. Sensitive silver-staining detection ofbacterial lipopolysaccharides in polyacrylamide gels. J. Biochem. Biophys.Methods 26:81–86.
31. Kleffel, B., R. M. Garavito, W. Baumeister, and J. P. Rosenbusch. 1985.Secondary structure of a channel forming protein: porin from E. coli outermembranes. EMBO J. 4:1589–1592.
32. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature (London) 227:680–685.
33. Lakey, J. H., and F. Pattus. 1989. The voltage-dependent activity of Esche-richia coli porins in different planar bilayer reconstitutions. Eur. J. Biochem.186:303–308.
34. Lemos, M. L., P. Salinas, A. E. Toranzo, J. L. Barja, and J. H. Crosa. 1988.Chromosome-mediated iron uptake system in pathogenic strains of Vibrioanguillarum. J. Bacteriol. 170:1920–1925.
35. Mathes, A. 1994. Aufbau eines hochempfindlichen Meßsystems zur Unter-suchung spannungsabhangiger porenbildender Proteine in planaren Lipid-membranen. Diploma thesis. Technische Universitat, Munich.
36. Montal, M., and P. Mueller. 1972. Formation of bimolecular membranesfrom lipid monolayers and a study of their electrical properties. Proc. Natl.Acad. Sci. USA 69:3561–3566.
37. Morrisey, J. H. 1981. Silver stain for proteins in polyacrylamide gels: amodified procedure with enhanced uniform sensitivity. Anal. Biochem. 117:307–313.
38. Nikaido, H., and M. Vaara. 1985. Molecular basis of bacterial outer mem-brane permeability. Microbiol. Rev. 49:1–32.
39. Paul, A., H. Engelhardt, U. Jakubowski, and W. Baumeister. 1992. Two-dimensional crystallization of a bacterial surface protein on lipid vesiclesunder controlled conditions. Biophys. J. 61:172–188.
40. Pazos, F., Y. Santos, B. Magarinos, I. Bandın, S. Nunez, and A. E. Toranzo.1993. Phenotypic characteristics and virulence of Vibrio anguillarum-relatedorganisms. Appl. Environ. Microbiol. 59:2969–2976.
41. Pyle, S. W., and W. B. Schill. 1985. Rapid serological analysis of bacteriallipopolysaccharides by electrotransfer to nitrocellulose. J. Immunol. Meth-ods 85:371–382.
42. Rachel, R., A. M. Engel, R. Huber, K.-O. Stetter, and W. Baumeister. 1990.A porin-type protein is the main constituent of the cell envelope of theancestral eubacterium Thermotoga maritima. FEBS Lett. 262:64–68.
43. Saxton, W. O., and W. Baumeister. 1982. The correlation averaging of aregularly arranged bacterial cell envelope protein. J. Microsc. (Oxford) 127:127–138.
44. Saxton, W. O., T. J. Pitt, and M. Horner. 1979. Digital image processing: theSemper system. Ultramicroscopy 4:343–354.
45. Sen, K., J. Hellman, and H. Nikaido. 1989. Porin channels in intact cells ofEscherichia coli are not affected by Donnan potentials across the outermembrane. J. Biol. Chem. 263:1182–1187.
46. Smith, P. K., R. I. Krohn, G. T. Hermanson, A. K. Mallia, F. H. Gartner,M. D. Provenzano, E. K. Fujimoto, N. M. Goeke, B. J. Olson, and D. C.Klenk. 1985. Measurement of protein using bicinchoninic acid. Anal. Bio-chem. 150:76–85.
47. Sørensen, U. B. S., and J. L. Larsen. 1986. Serotyping of Vibrio anguillarum.Appl. Environ. Microbiol. 51:593–597.
48. Suzuki, S., K. Kuroe, and R. Kusuda. 1994. Characteristics of porin-likemajor outer membrane proteins of Listonella anguillara serotypes J-O-1, -2,and -3. Biochem. Mol. Biol. Int. 32:605–613.
49. Towbin, H., T. Staechelin, and J. Gordon. 1979. Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets: procedure andsome applications. Proc. Natl. Acad. Sci. USA 76:4350–4354.
50. Yamada, H., and S. Mizushima. 1980. Interaction between major outermembrane protein (O-8) and lipopolysaccharide in Escherichia coli K-12.Eur. J. Biochem. 103:209–218.
4188 SIMON ET AL. J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
03
Febr
uary
202
2 by
191
.240
.117
.52.