estudio de la fauna de los castros de lastra … · id.a.1.-mamiferos la tabla 2 recoge la...
TRANSCRIPT

KOBIE (Serie Paleoantropología). Bilbao Bizkaiko Foru Aldundia-Diputación Foral de Bizkaia N.º xxvm, pp. 51-88, año 2009. ISSN 0214-7971
ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, ALAVA)
Faunal's study fro1n "Los Castros de Lastra" (Caranca,Alava)
RESUMEN
Pedro Castaños* Jone Castaños**
(Recibido 15/XI/08) (Aceptado 15/XII/08)
En este trabajo se estudian los restos de fauna del yacimiento de Los Castros de Lastra (Caranca,AJava) cuya cronología se extiende desde finales del Bronce hasta la Edad Media. Se observa un claro predominio de las especies domésticas entre las que destaca el ganado ovicaprino seguido del bovino y del cerdo. Se compara la estructura faunística de esta muestra con la de otros asentamientos coetáneos de la Península Ibérica. Se analiza la representación anatómica de los restos, la distribución de edades y sexos así como la osteometría de cada cabaña doméstica para establecer los modelos de aprovechamiento y explotación de los distintos componentes ganaderos. En el ovicaprino se observa un aprovechamiento de carácter mixto con el sacrificio de animales jóvenes para su consumo y la conservación de otros hasta la edad adulta. En el bovino la mayoría de los individuos son adultos. Y en el cerdo la frecuencia de jóvenes es más alta.
Se incluye un estudio métrico de las distintas especies comparando sus dimensiones con las de otras muestras próximas en el espacio y en el tiempo. También se estudia el proceso de descuartizado del ganado ovicaprino.
Palabras clave: Arqueozoología, Bronce Final, Edad del Hierro, Edad Media, País Vasco, España.
* Museo Arqueológico, Etnográfico e Histórico Vasco. Bilbao; [email protected] ** Departamento de Estratigrafía y Paleontología Facultad de Ciencia y Tecnología, Universidad del País Vasco/ Euskal Herríko Unibertsitatea ,
Apartado 644, E-48080 Bilbao; [email protected]

52 PEDRO CASTAÑOS Y JONE CASTAÑOS
ABSTRACT
Bone remains coming from the site of "Los Castros de Lastra" (Caranca, Ala va) with a chronology frorn late Bronze until Middle Age, are studied on the present work. Predorninance of domestic species is verified; to point out sheep/goat and followed by bovine cattle and pig. The faunal structure of this sample is compared to other contemporary sites on the Iberian Peninsula. The anatornic representation of bones, age and sex distribution as well as the rneasures of each domestic flock is also analysed to establish profit use and exploit patters of different livestock cornponents. In the precise case of sheep/goat a rnix use is noticed where young individuals are sacrificed far consurnption while others are kept until adult age. Regarding the bovine cattle, rnost of the anirnals are adults and respect to the pig, the frequency of young individuals is higher.
A metric study of the different species comparing their dirnensions with other samples next in space and time is also included, furthermore of the butchering process of sheep/goat.
Key words: Archeozoology, Late Bronze, /ron Age, Middle Age, Bask Country, Spain.
LABURPENA
Lan honetan Lastrako Gotorlekuko (Karanka, Araba) aztarnategiko fauna hondakinak aztertzen <lira. Horien kronologia Brontze Aroaren amaieratik Erdi Arara doa. Etxeko espezieak nabarmenki nagusi direla ikusten da; horien artean, azpirnarragarriak dira ardi eta ahuntz taldeak, atzetik behi eta txerri taldeak egonez. Lagin honetako faunaren egitura alderatu egiten da Iberiar Penintsulako garai berdineko beste kokaleku batzuetan topaturikoarekin. Hondakinen irudikapen anatornikoa, adinen eta sexuen banaketa nahiz etxeko abere bakoitzaren osteornetria aztertzen dira, abeltzaintzako osagai ezberdinen aprobetxamendu eta ustiapen ereduak ezartzeko. Ardí eta ahuntzen kasuan, izaera rnistoko aprobetxarnendua ikus daiteke: kontsumorako animalia gazteak sakrifikatzen ziren eta beste batzuk heldu bihurtu arte rnantentzen ziren. Behien kasuan, ale gehienak helduak <lira. Eta txerriaren kasuan, berriz, gazteen rnaiztasuna altuagoa da.
Espezie ezberdinen azterketa rnetrikoa erantsi da, beren dirnentsioak espazioan eta denboran hurbilekoak diren beste lagin batzuek dituztenekin alderatuz. Ardí eta ahuntzak laurdentzeko prozesua aztertzen da.
Hitz gakoak: Arkeozoologia, Brontze Aroko amaiera, Burdin Aroa, Erdi Aroa, Euskal Herria, Espainia.
KOBIB (Paleoantropología n.º 28), año 2009

ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, ALAVA) 53
1.- INTRODUCCION
El presente trabajo tiene por objeto el análisis arqueozoológico de la fauna recuperada en Los Castros de Lastra (Caranca, Alava). Se trata de un yacimiento con distintos sectores y varios horizontes de ocupación que van desde el Bronce Final hasta el Medievo aunque la mayor parte del material corresponde a la Primera y Segunda Edad del Hierro. La excavación fue dirigida por Dña. F. Sáez de Urturi quien nos encomendó los materiales para su estudio.
II.- MATERIAL Y METODOS
I.a.- MATERIAL
Equus caballus 55 10
Equus asinus
Bos taurus 2833 83 11 237 Ovis/Capra 6193 73 48 315 Sus domesticus 1798 25 10 139 Canis familiaris 4 31 Pelis catus 15
Cervus elaphus 288 7 Capreolus capreolus 1 Sus ferus 2 Oryctolagus cunicu-
3 lus
Lepus capensis 7 Talpa europaea 3 Sciurus vulgaris 1 Gallus gall us 33
Coturnix coturnix 1 Alectoris rufa 2
Columbasp. 1
Corvus corone 1
Turdus sp. 1
Rapaz indet.
Unio sp. 16 2 1 Cardiumsp. 5
Venus sp. 2
Restos humanos 78
Totales 11343 229 71 692
% Número restos 72,14 1,45 0,45 4,41
El estudio completo de la muestra recuperada en Caranca ha proporcionado un total de 15.719 restos identificados anatómicamente y taxonómicamente. Hay un conjunto de fragmentos óseos que no han sido susceptibles de clasificación y otro que aunque identificado procedía de niveles con mezcla de elementos del Hierro y medievales. Este último se ha incluido en la muestra de la Edad Media.
Esta rica muestra se reparte de forma desigual en cuatro grupos zoológicos de distinta significación arqueológica. El conjunto dominante corresponde a 14 especies de Mamíferos cuyos restos suponen el 98,9% del. total de los recuperados. Hay también un pequeño grupo de restos de Aves (0,25%) y de Moluscos (0,31%). Y mezclados con la fauna se recuperaron también algunos restos humanos (O ,5%) (Tabla 1).
7 10 82 l 1
426 24 21 346 3981 1269 43 20 580 8541 326 21 11 214 2544
35
15 1 36 332
l 2 3 5
3
7 3 1
33
1
2 1
1
1 1 1
10 13 42
5
2
2039 89 53 1203 15719 12,97 0,56 0,33 7,65
Tabla 1.- Distribución del número de restos según las distintas especies y sectores excavados.
KOBIE (Paleoantropología n.º 28), año 2009

54 PEDRO CASTAÑOS Y JONE CASTAÑOS
La cantidad de fauna recuperada en cada sector de Caranca es desigual no sólo en cantidad sino también en precisión cronológica. Casi las dos terceras partes (72,16%) de todos los fragmentos identificados proceden del Sector II que además presenta la mayor variedad taxonómica ya que salvo el asno y una rapaz ha proporcionado evidencias de las restantes especies presentes en la muestra. ·
A mucha distancia en tamaño queda el conjunto recuperado en el sector VB (12,9%).El elenco de especies se reduce a las tres cabañas domésticas más comunes, una rapaz y algunas valvas de almejón de río sin que aparezca un solo resto de Mamíferos salvajes. Los sectores VIII y VA reducen su frecuencia de restos a 7 ,6% y 4,4% respectivamente quedando para las demás zonas muestras casi residuales que en el caso del sector III sólo representa el 1,45% y en las otras tres no se alcanza ni el 1 %.
Hay otra circunstancia que enfatiza la importancia de la muestra del sector II además de su riqueza cuantitativa ya de por sí destacable. La mayor parte de los fragmentos recuperados en esta zona pertenecen a horizontes cronológicos y culturales bien diferenciados desde el Bronce Final al Medievo. Sólo hay un conjunto a caballo entre la Primera y Segunda Edad del Hierro. Sin embargo, en los demás sectores
Long. máx. periférica
hay siempre una parte del material medieval mezclado con elementos del Hierro y el resto no se puede asignar con seguridad a qué fase del Hierro pertenecen. En adelante estos materiales se incluirán en los de la época Celtibérica y los anteriores en la medieval.
11.b.- METODOLOGIA
Los procedimientos empleados en la identificación de los restos así como los criterios para la estimación de la edad, sexo y otros extremos arqueozoológicos so~ los comunes en este tipo de trabajos y por muy repetidos en múltiples publicaciones omitimos aquí. :ran sólo se ofrecerán referencias bibliográficas al aphcar los factores de estimación de la altura en la c:uz y de las cohortes de edad en las distintas especies.
Todas las medidas han sido obtenidas siguiendo la m~~odología Y,ª ~lásica de A.v.d. DRIESCH (1976) utilizada en pract1camente todos los análisis de faunas ibéricas. Se han expresado todas ellas en mm. con un error estimado de 0,5 mm. en todas las que superan los 20 mm. (excepto en los metapodios de Lagomorfos) y de 0,05 mm. en las demás. Las abreviaturas utilizadas son las siguientes:
Long. olécranon
Long. serie d. inf. Número mín. individuos
KOBIE (Paleoantropologfa n.º 28), año 2009

ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, ALAVA) 55
A lo largo del trabajo se citarán con cierta frecuencia una serie de yacimientos de la Edad del Hierro situados todos ellos en la mitad septentrional de la Península en las cuencas del Ebro y del Duero cuya relación y referencia bibliográfica se adelantan con objeto de evitar posteriores repeticiones.
- Castro del Castillo de Henayo (Alava): Primera Edad del Hierro (Altuna, 1980).
Castro de Berbeia (Alava): Primera Edad del Hierro (Altuna, 1980).
- Castro de la Peñas de Oro (Alava): Primera Edad del Hierro (Altuna, 1980).
- Sacaojos (León): Primera Edad del Hierro (Driesch & Boessneck, 1980)
Cerro de Santa Ana (Rioja): Primera Edad del Hierro (de Miguel & Morales, 1983).
- Castillar de Mendavia (Navarra): Primera Edad del Hierro (Mariezkurrena, 1986).
- Castro de Ubiema (Burgos): Segunda Edad del Hierro (Castaños, 1989).
- Poblado de La Hoya (Alava): Primera y Segunda Edad del Hierro (Altuna & Mariezkurrena, 1990).
- Soto de Medinilla (Valladolid): Primera y Segunda Edad del Hierro (Liesau, 1993).
Castro de Barahones (Palencia): Primera Edad del Hierro (Líesau, 1993).
- La Mota (Valladolid): Transición Hierro I-II (Morales & Liesau, 1995)
- Cerro del Castillo (Valladolid): Primera y Segunda Edad del Hierro (Morales &Liesau, 1195)
- Era Alta (Valladolid): Primera y Segunda Edad del Hierro (Morales & Liesau, 1995)
KOBIE (Paleoantropología n.º 28), año 2009
- Las Quintanas-Valoría (Valladolid): Segunda Edad del Hierro (Morales & Liesau, 1995)
- Las Quintanas- Padilla (Valladolid): Segunda Edad del Hierro (Morales & Liesau, 1995)
- Castejón de Bargota (Navarra): Segunda Edad del Hierro (Castaños, inédito).
ID.- RESULTADOS
ID.a.- GENERALIDADES
ID.a.1.- MAMIFEROS
La Tabla 2 recoge la distribución de los 15.552 restos de Mamíferos según las distintas especies y en cada uno de los horizontes culturales. El conjunto más escaso corresponde a la ocupación del Bronce Final. A la baja cantidad de restos para este período se añade su escasa variedad taxonómica ya que a la tríada habitual de cabañas domésticas (bovino, ovicaprino y cerdo) sólo se añade la presencia del caballo y del ciervo.
Las muestras de la ocupación medieval y de la Primera Edad del Hierro se acercan en cada caso a los dos millares de restos repartidos casi entre las mismas especies. Sin embargo, el nivel Celtibérico es sin duda el más abundante tanto en restos como en número de especies. Además conviene tener presente la presencia de un importante conjunto de huesos mezclados entre los dos subperíodos del Hierro. En definitiva los fragmentos procedentes de las dos ocupaciones del Hierro suponen el 82,6% de toda la muestra identificada. Por todo lo cual se puede considerar la fauna de Caranca como perteneciente fundamentalmente a la Edad del Hierro de forma que las ulteriores consideraciones y comparaciones que a lo largo del trabajo se expongan se van a circunscribir a este horizonte cultural.

SG PEDRO CASTJ\llos y JONE CASrAJ1<os
~Wi~==~~~~~~~~~~;~~~ :l1~~it&~~~~t r~~t{ t.™!•!(tf 0~~~~m~~1 ~~~-i 1-~~:~ift~~ Equus caballus 9 43 14 11 5 82
Equui:- asinus 1 1
Bosm~ 390 1803 892 633 263 3981
Ovis aiCupr.a h. 1155 3967 1813 957 .151 8303
Ovis ruies 20 83 30 17 5 155
C:J.pr;~ hill:\IS 17 73 31 15 7 83
Sus ill)1h~~t.ictL'> 312 1274 571 285 102 2544
Canis f:uniliari!:I 34 1 35
f'c,is caws 15 15
Cervus elapbus 18 132 85 68 29 332
Cápn:olu.'> c:.pn::u,. 2 2
Sus fcrus 1 4 5
01)'Ctol~u.°' C. 1 2 J
Lepus capen.~is 6 1 7
Tulpll curopaea 1 2 3
Sciuru~ \'Ulgaris 1 1
Totales 1942 7415 3442 1991 762 15552
%NR do1uésticos 98,76 99,16 97.36 96,38 96,2
%NR sal vajcs 1,24 0,83 2,64 3.61 3.8
Tabla 2.· Distl'ibucióo de. los restos de ~lamifcr~ por especie~ y hmi7.0ntcs cuhuralc~.
El primer rasgo a desJ.acar en esta muestra es el claro predominjo de las especies domésticas respecto de la fauna salvaje. Tal como se indica al final de cada nivel de ocupación la proporción de los restos pertenecí.entes a animales domésticos supera el 96% en todos los períodos. Parece por tanto razonable afumar desde el inicio de es1e análisis que Ja subsistencia de origen anima l de Caranca es bá<icameme pecuaria desde el Bronce Final basta eJ Medievo. T..a caza supone un apone nuuicional muy pobre e incluso dentro de su escasez se observa que disminuye pro!,'TeSivamentc desde Ja ocupadón más anLigua a los tiempos históricos.
Dentro de Ja fauna doméstica el ganado ovicaprino es el mejor representado en número de res1os aumentando su frecuencia a lo largo de la ocupación de forma gradual pero continua. En la cabaña porcina
la frecuencia apenas varía a lo largo de la ocupación y ocupa siempre en el tercer Jugar (Fig. !).
70 60 lil Equus 50 •Bos 40
c O/C 30 20 a Sus
10 •Ung.s. o
1 2 3 4 5
Fig. 1.- llisto;,'í'.una de frecuencia.~ de los re.o;lns de Ong,uladc~ en fo.., distint0$ niveles ( l. f\.tcdic•;o 2. Hierro JI 3. IUeue> 1-14. Hieito 1 5. Bronct l'mul).

ESTUDIO DE LA f ,,\UNA DE"~ C;\!:il~OS DE l.A:STW. A .. (<.:AEtANCA, ,'\IÁ\"'r'A) 57
El bovino sigue un proceso inverso reduciendo su presencia también poco a poc.:o pern de forma continua. Ocupa el segundo puesto cuando se tiene en cuenta el número de restos pero pasa a primer lugar cuando el cálculo se hace a partir del peso <le los huesos. La evolución diacrónica de la biomasa representada por cada especie de Ungulados sigue las mismas pautas que las descritas al considerar el nllmero de restos (Fig. 2).
70.-~---~~~~~~-. .... ....,
60
10 o,,... ...... ....,.~-..-......,.-.ii..,.-....-1 1 2 3 4 5
a Equus
• Bos aO/C
a Sus
•Ung.s.
YJg. 2.· H.is1ogi·ruu:l d~ fro?c\1en.:ias del peso de Ungulado!!: en los distintos ni\'elci> (1. ~1ellic·•o 2. Hierro II 3. Ilicrro 1-14. Ilieno I 5. Bronce Fin3J).
Gallus gallus 10 7
AlecLoris /Perdix 2
Cotumix c.
Columba livin/oenas 1
Corvus corone/frugilegus
Turdus sp.
Accipitridac intl.
Totales 14 7
Tabl:i 3.· Djsr.ribucióu de Los rcs1os de avif:nms rlc Caranc11.
Este conjunto representa un mínimo de siete especies de la.~ que seis son salvajes. La gallina es la única rorma doméslica aunque Ja ~jor 1·epresentada tanto en mímero de restos como en individuos. El res1o son en
KOBIE (PJJro.-incropoJogia u." 2Sl, ~ 2009
El caballo y el asno muestran frecuencias casi residuales y además el asno no aparece hasta la Se&'llnda Edad del li.ierrn. La relación de especies domésticas se completa con el perro que está presente en dos horiz.ontes del Hierro con desigual frecuencia. J.lama la atenci6n la ausencia de restos de este casi inevitable compañero del hombre en el B ronce y en el nivel de transición dentro del Hierro así como su escasa frecuencia en el Hierro T. El gato e~iá presente en el n ivel medieval pero fülta como era de esperar en las ocupaciones protohistóricas .
Entre los Ungulados objeto de caza el ciervo es la especie más frecuente y está presente a lo laJ!:o de todos los momentos de ocupación. El corzo y el jabalí son mucho más raros en cantidad de restos y además su presencia parece restringida al horizonte Celtibérico. La liehre y el conejo también presentan frecuencias residuales y en la misma línea están el topo y ta ardilla.
ffi.a.2.- AVES
El gru¡x) de las Aves ha dejado solamente 44 füt&•mentos repartidos por todos los ho1izontes de ocupación (Thbla 3).
10 6 1 34
2
1 1
1
4 4
1
15 6 2 44
general ~p~-...ic::s ubiquisbls habituales en el entorno de Caranca desde finales del Wünn. La Tabla 4 recoge la distribución ana16mica de los restos de cada espc<--ie sin tener en cuenta sus ambucioncs cronológicas.

58 PEDRO CASTAÑOS Y JONE CASTAÑOS
Coracoides 8
Clavícula 1
Escápula 2
Húmero 4
Radio 3
fina 3
Carpo-Me 2
Pelvis 1
Fémur
Tibio-Tarso 7
Ts-Mt 2
Totales 34 2
Tabla 4.- Distribución anatómica de los restos de avifauna.
ill.a.3.- MOLUSCOS
Hay un total de 23 restos de Moluscos repartidos por los distintos niveles de ocupación (Tabla 5). Pertenecen todos a Moluscos Bivalvos tanto marinos como continentales aunque con mejor representación cuantitativa de éstos últimos.
Unío sp. 4 4
Cerastoderma sp. 2
Venus sp.
Totales 4 7
Tabla 5.- Distribución de los restos de Moluscos.
El almejón de río (Unio) es la forma más abundante estando presente en todos los horizontes de ocupación. Es un Molusco dulceacuícola relativamente frecuente en yacimientos del interior con corrientes fluviales próximas como es el caso de Caranca. Tanto el berberecho (Cerastodemza) como
1
8
2
3 9
3
4
3
2
8
3
4 44
4 2 2 16
2 5
2
7 3 2 23
la chirla (Venus) son marinas y su presencia en el yacimiento sólo puede ser consecuencia de su importación desde la costa canta'brica probablemente donde son habituales. Su interés para los habitantes de Caranca pudiera estar relacionado con técnicas de decoración cerámica.
KOBIB (Paleoantropología n.º 28), año 2009

ESTUDIO DE LA FAUNA DE 1.0S CASTROS DE LASTRA» (CARANCA, ALAVA) 59
ID.a.4.- RESTOS HUMANOS
Los 78 restos humanos por su naturaleza no son objeto de este estudio. A falta de un análisis antropológico se ofrecen a modo de inventario la distribución anatómica de los mismos en los distintos horizontes culturales (Tabla 6).
Maxilar
Diente a. superior 4
Diente a. inferior 2
Clavícula 2 1
Vértebra 1 3
Costilla
Radio 2
Ulna
Metacarpo 4
Pelvis 3
Fémur
Rótula
Tibia 1
Tarso
Metatarso 5 4
Metapodio ind.
Falange 1 8 3 9 2
Falange 2 1 2 4
Falange 3 3
Totales 24 26 25 3
Tabla 6.- Distribución de los restos humanos.
KOBIB (Paleoantropologfa n.0 28), año 2009

60 PEDRO CASTAÑOS Y JONE CASTAÑOS
ID.B.- DESCRIPTIVA ESPECIFICA
ID.b.1.- CABALLO
Equus caballus
El caballo con 82 restos es el Ungulado doméstico más escaso después del asno. La Tabla 7 recoge la distribución anatómica de sus fragmentos en los distintos horizontes culturales.
Maxilar
Diente a. Sup. 4 6
Mandíbula 3
Diente a. Inf. 3 14
Costillas 4
Escápula
Radío 2
Ulna
Carpo 3
Metacarpo
Tibia
Astrágalo
Resto tarso 4
Metapodio ín. 2
Falange 1
Falange 2
Falange 3
Sesamoideo
Totales 9 43
Tabla 7 .- Distribución anatómica de los restos de caballo.
La muestra más significativa procede del nivel Celtibérico. Predominan las piezas dentarias aisladas y algunas falanges. La estimación de la edad no es
1
7 1 18
3
3 20
4
4
1
3
1
1
2
5
2 4
3 5
2 3
2 3 6
1
14 l1 5 82
muy precisa debido a que los elementos de la denti-ción no están en sus correspondientes series.
KOBIB (Paleoantropología n.º 28), año 2009

ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, ALAVA) 61
1M 25,.5 LM 30,.5
AM 23,5 AM 17,8
Lprotoc. 9,4 Doble lazo
++ Postfléxido
Ap 62 74
Asp 58,5 66,5
AD 30,5
Ad 61 54,5
Asd 49,5
Ad 66,5 AM 66,5
Ed 40,5 ASart. 45
Tabla 8.· Medidas aísladas de caballo (las no señaladas son del Hierro).
Los 9 restos del nivel medieval representan como mínimo a dos individuos: un adulto y un juvenil. En la ocupación Celtibérica hay al menos cuatro distintos: un subadulto y tres adultos cada uno de ellos con un grado de desgaste mayor y por tanto una edad diferente. En cada uno de los horizontes restantes hay un solo individuo adulto salvo en el Hierro 1-II en que se trata de un subadulto.
Las medidas son pocas (Tabla 8) y se recogen todas ya que la endémica escasez del caballo en yacimientos protohistóricos hace que cualquier dato métrico sea interesante de cara a poder algún día tipificar esta especie.
ID.B.2.- GANADO VACUNO
Bos taurus
ID.b.2.1.- Representación anatómica
Los 3981 restos de ganado vacuno se reparten de forma desigual entre los distintos horizontes cronológicos. Casi la mitad ( 45 ,3%) del material procede del nivel Celtibérico seguido a distancia del de transición
KOBIE (Paleoantropología n.º 28), año 2009
27 26,3 23 24,3 32 33
18,6 18,6 16,8 16,l 14,4 13,4
19 17,4 13,9 13,2 12,9 14,6
10,5 9,1 7 ;2 8 9,5 10,7
++ +
49 Ap 52,5 52
46 Asp 48,5 44
AD 27,5 AD 43,5 42,5
Ad 40 Ad 49,5 46
p a p
(22,4%) y del correspondiente a la Primera Edad del Hierro (15,9%). Tanto la muestra medieval (9,8%) como la procedente del Bronce Final son las más escasas.
La Tabla 9 recoge la distribución anatómica de los fragmentos en cada nivel de ocupación. La porción correspondiente al esqueleto de las extremidades mantiene una proporción relativamente constante mientras que las variaciones diacrónicas más significativas se circunscriben a los elementos de la cabeza y tronco. A partir del nivel de transición se aprecia un aumento de la frecuencia relativa de los fragmentos craneales y una correlativa disminución de elementos del esqueleto axial. Este fenómeno pudiera ser consecuencia de una mayor fragmentación de los maxilares que aumenta el número de piezas dentarias aisladas.
En todos los niveles el esqueleto del tronco está escasamente representado si se compara con el de la cabeza por la elevada proporción de molares desprendidos de los arcos maxilares. Además se aprecia una disminución de las falanges en sentido distal. El dato apoya la hipótesis de prácticas de curtido que exige el desollamiento de la res y la eliminación frecuente de las terceras falanges y a veces también las segundas con la piel del animal.

62 PEDRO CASTAÑOS Y JONE CASTAÑOS
Clav.Cuerna 19 9 5 34
Cráneo 14 46 31 15 5 111
Maxilar 2 11 48,1
11 44,l
7 29,5
3 28,2
34
D.a. sup. 60 337 124 56 28 605
Mandfbula 27 82 77 46 19 251
D.a. inf. 80 46,92
372 139 58 19 668
Hiodes 3 4
Vértebra 22 13,08 116 53 74 29 294
Costilla 29 125 13,3 63 13,0 110 29,0 37 25,2 364
Escápula 9 43 38 14 4 108
Húmero 19 102 49 30 14 214
Radio 9 56 38 13 11 127
Ulna 2 21 8 32
Carpo 4 24 17 4 8 57
Metacarpo 3 15 14 4 4 40
Pelvis 11 36 15 25 8 95
Fémur 12 40,0 74 38,5 30 42,8 22 41,4 12 46,5 150
Rótula 3 5
Tibia 27 130 59 47 18 281
Calcáneo 3 7 16 13 3 42
Astrágalo 3 16 5 8 6 38
Resto tarso 3 5 3 13
Metatarso 7 34 19 3 8 71
Metap.Ind. 24 55 29 20 4 132
Falange 1 6 33 19 20 8 86
Falange 2 7 27 19 16 6 75
Falange 3 6 12 4 13 3 38
Sesamoideo 2 3 2 4 12
TOTALES 390 1803 892 633 263 3981
%NR 9,8% 45,3% 22,4% 15,9% 6,6%
Tabla 9.- Distribución anatómica de los restos de bovino doméstico.
KOBIE (Paleanntropología n.º 28), aílo 2009

ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, Al.AYA) 63
Si se compara la distribución relativa de las tres regiones del cuerpo entre distintas muestras del Hierro 1yHierro11 del Norte peninsular se observan patrones diferenciados. En el caso de la Primera Edad del Hierro (Tabla 10) la proporción de elementos apendiculares muestra la mayor constancia ya que oscila entre el 37,5% y 55,4% en las nueve muestras consideradas. La Hoya, Sacaojos y Caranca ofrecen frecuencias similares entre la cabeza y el tronco mientras que en los tres restantes se aprecia un predominio de los elementos craneales especialmente en el
Tronco 17,0 25,0 10,7 27f:J
Extrem. 42,5 37,5 44,6 40,88
NTR 688 634 289 702
Soto de Medinilla y Peñas de Oro. La variación es algo mayor en las muestras Celtibéricas (Tabla 11) ya que aquí también hay diferencias en la representación porcentual de las extremidades entre el 64,43% de Bargota y el 38,5% de Caranca. Esta parte del cuerpo sigue ofreciendo valores próximos al 50% en las tres muestras. Además se observan un dominio de elementos respecto del tronco en La Hoya, Caranca y B argota que se equilibran en Soto de Medinilla y se invierten en Ubierna.
24,36 16,58 12,7 9,14 29,06
48,92 51,75 52,1 55,4 41,39
5827 796 848 908 633
Tabla 10 .- Frecuencias porcentuales de las regiones anatómicas del bovino en el Norte de la Península para la Primera Edad del Hierro.
Cabeza 32,45 21,98 27,7 48,14 26,03
Tronco 15,78 31,27 22,05 13,36 9,53
Extremidades 51,76 46,74 55,25 38,5 64,43
NRT 2129 905 1551 1503 703
Tabla 11.- Frecuencias porcentuales de las regiones anatómicas del bovino en el Norte de la Península para la Segunda Edad del Hierro.
ID.b.2.2.~ Distribución de edades y sexos
Utilizando como criterio básico el grado de erupción y desgaste dentarios se ha estimado la edad de 129 individuos repartidos de forma desigual en los distintos horizontes cronológicos (Tabla 12).
Ml 0-6 meses Infantil 3 3 4 12
Ml+M2- 6-18 meses 2 5 3 11
M2+M3- 18-27m. Juvenil 9 6 16
M3+/- 27-30 m. 2 3 5
M3+ Subadulto 3 14 5 3 2 27
M3++ 3 13 5 5 2 28
M3+++ Adultos 3 16 7 3 30
Totales 13 60 31 15 10 129
Tabla 12.- Distribución de edad del bovino doméstico.
KOBIE (Paleoantropologfa n.º 28), año 2009

64 PEDRO CASTAÑOS Y JONE CASTAÑOS
Agrupando dichos datos en cohortes de edad, se aprecia en general un predominio de animales adultos y subadultos frente a los juveniles e infantiles. Tan solo se desvía de este patrón el conjunto correspondiente al Bronce Final que por el reducido tamaño es más sensible a los factores aleatorios inherentes a las pequeñas muestras. Con estos datos puede proponerse al menos para las ocupaciones de la Edad del Hierro, un patrón de aprovechamiento pecuario de carácter marcadamente secundario que indica una conservación de gran parte de los individuos de esta cabaña ganadera hasta la edad adulta con el fin de obtener productos derivados como la leche, trabajo y reproducción antes del simple aprovechamiento cárnico.
ID.b.2.3.- Tamaño y alzada
El conjunto de valores obtenido no es muy amplio si se tiene en cuenta el tamaño de la muestra debido al alto grado de :fragmentación de la misma. No obstante, ante la escasez de conjuntos métricos coetáneos del Norte peninsular ofrecemos el resumen estadístico de los valores más frecuentes (Tabla 14).
Mandíbula
LM3 14 32,5-37 34,39 1,41 4,12
AM3 14 1,4-16 13,42 0,9 6,7
Escápula:
LMP IO 61-75 66,5 5,03 7,5
LS IO 51,5-62,5 56,1 4,13 7,3
AS 11 42-52,5 46,8 3,56 7,6
LmC 8 47-58,5 52,5 5,46 10,4
Metacarpo:
Ap 7 49,5-62 57,5 5,09 8,8
Tibia:
Ap 2 86-86,5 86,25
Ad 7 57,5-63,5 60,35 1,79 2,9
Astrágalo:
LMl 13 59,5-67,5 63,8 2,23 3,4
LMm 14 55-63 58,71 2,16 3,6
El 14 30-38,5 35,28 1,99 5,6
Ad 14 37-49 42,25 3,47 8,2
Se han agrupado los de todo el Hierro ya que no parece haber diferencias significativas entre ellos y con el fin de no perder los datos del nivel de transición de cara a cotejarlos con los de otros asentamientos. Comparados los parámetros estadísticos de Caranca con los de Sacaojos, P. Vasco, Ubierna y Bargota se observa que la mayoría de las medias de este yacimiento alavés entran dentro del dominio de variación de las de los restantes yacimientos. La única excepción es el astrágalo que presenta una media algo superior a la observada en el resto. Nos encontramos por tanto ante un conjunto de ganado vacuno cuyo tamaño entra dentro de los límites que son habituales en la época y área geográfica en que está encuadrada.
Por lo que a la alzada se refiere tan sólo se conserva entera una tibia (LM 346) a partir de la cual y utilizando los factores de Fock (1966) y Matolcsi (1970) se ha podido estimar la altura en la cruz del ejemplar en 119,37 cms. Este valor queda también dentro del intervalo de variación de las restantes muestras citadas. En general la alzada del bovino doméstico va disminuyendo desde el Neolítico al Hierro ofreciendo en esta última época los ejemplares más pequeños. Con la Romanización la talla vuelve a aumentar presentando los valores más altos hasta los tiempos modernos.
Húmero:
Ad 2 60,5-73,5 67
AT 2 56,5-71 63,75
Radio:
Ap 5 73,5-88,5 81, 6,77 8,3
ASp 6 67,5-82 73 6,22 8,5
Ad 2 68-76,5 72,25
Fémur:
EC 3 44,5-48 46,33
Calcáneo:
LM 4 115-132 124,37
AM 4 39-43 40,75
Metatarso:
Ap 5 42,5-47 44,1
Ad 4 49,5-55 53,37
Ed 4 28,5-31 29,87
KOBIE (Paleoantropología n.º 28), año 2009

ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, ALAVA) 65
Fa!.1 ant.: Fa!J post.:
LMpe lO 49-57 52,65 2,66 5,07 LMpe 10 48,5-63-5 56,15 4,1 7;3
Ap lO 27,5-32 29,0 1,69 5,8 Ap 10 24-32 27;2 2,5 9;2
AD 10 22-26 24,15 1,33 5,5 AD lO 20-25,5 22,4 1,67 7,4
Ad 10 25,5-32 27,45 2,12 7,7 Ad 10 22,5-31 25,75 2,51 9,7
Fa!.2 ant.: Fa!.2 post.:
LM 8 32-42,5 36,5 3,68 10,0 LM 16 32,5-42,5 36;37 2,5 6,8
Ap 8 26,5-35 30,06 3,46 11,5 Ap 16 24,5-31,5 27,5 1,87 6,8
AD 8 18,8-27 23,47 3,06 13,0 AD 16 18;2-26 21;21 1,91 9,0
Ad 8 21,5-30,5 26,18 3,49 13;3 Ad 16 19,7-26 22,6 1,77 7,8
Falange 3: Centrotar.:
LSD 10 61-80 68,55 5,63 8;2 AM 3 51-59,5 55;33
Ldo 10 48,5-59 51,85 3,07 5,9
Tabla 14.- Resumen estadístico de las medidas del bovino doméstico del Hierro de Caranca.
GANADO OVICAPRINO
Ovis aries/Capra hircus
ill.b.3.1.- Representación anatómica.
También los 8541 restos de ovicaprino se reparten de forma desigual entre los distintos horizontes culturales siendo las muestras más ricas las que proceden del nivel Celtibérico y del Hierro I-Il. En esta cabaña el conjunto medieval es algo más abundante que el del Hierro I (Tabla 15) quedando el correspondiente al Bronce Final muy por debajo de los demás.
Clavija c. 3 20 8
Cráneo 20 12
Maxilar 6 24 21
Diente a.sup. 173 780 294
Mandíbula 54 33;2
212 41,5 88
Diente a. inf. 160 650 239
Hioides 6 4
Vértebras 34 82
Costillas 209 20;3 714 20,2 417
KOBIE (Paleoantropología n.º 28), año 2009
En el ganado menor la proporción de los fragmentos pertenecientes a las extremidades es de nuevo la que menos variabilidad presenta a lo largo de casi todas las etapas de ocupación del yacimiento salvo para el conjunto medieval. Sin embargo la frecuencia de esta región anatómica es más baja que en el bovino. En el esqueleto del tronco también se registra una disminución progresiva las ocupaciones más antiguas a las del Medievo. Las variaciones de los elementos craneales dependen una vez más del número de piezas dentarias que se han separado de las arcadas maxilares. Y se confinna también aquí la reducción de las falanges más distales (segundas y sobre todo terceras) como consecuencia del la separación de las pieles de cara a su aprovechamiento.
3 3 37
11 44
13 4 68
109 37 1393
35,5 56 29,6 26 32,2 436
100 46 1195
11
37 12 286
26,6 348 38,9 97 30,0 1785

66 PEDRO CASTAÑOS Y JONE CASTAÑOS
Escápula 18 65 20
Húmero 52 130 59
Radio 40 153 59
Ulna 5 19 7
Carpo 6 22 16
Metacarpo 6 45 21
Pelvis 10 49 9
Fémur 76 178 64
Rótula 7 5
Tibia 70 239 114
Calcáneo 6 31 14
Astrágalo 14 36 12
Resto tarso 3
Metatarso 13 46,l 50 38,2 26
Metap. ind. 199 420 225
Falange 1 18 71 30
Falange 2 12 36 14
Falange 3 5 16 7
Sesamoideo
Totales 1192 4123 1874
Tabla 15.- Distribución anatómica de los restos de ovicaprino.
La proporción relativa de las distintas regiones anatómicas del ovicaprino en Caranca se asemeja mucho a la que existe en La Hoya al menos durante el Hierro I con cierto equilibrio entre los elementos de las tres regiones del cuerpo. Hay cinco muestras de las nueve cotejadas en las que el esqueleto de las extremidades absorbe más de la mitad de los frag-
Cabeza 23,05 23,83
Tronco 18,35 36,23
Extremidades 58,6 39,93
TotalNR 7861 621
12 4 119
22 15 278
29 19 300
7 39
8 52
6 2 80
9 78
45 17 380
13
42 28 493
2 2 55
4 3 69
15
37,8 9 31,4 5 37,7 103
93 28 965
10 6 135
6 2 70
4 3 35
2
989 363 8541
mentas y la cabeza está mejor representada que el tronco. Y a estas dos situaciones se añade una tercera caracterizada por el predominio de fragmentos craneales (Berbeia y Peñas de Oro) como consecuencia de cierta abundancia de fragmentos de maxilar y de piezas dentarias aisladas (Tabla 16).
28,4 27,7 29,6
16,3 12,1 38,9
55,1 60,2 31,4
232 1282 989
Thbla 16.- Porcentajes de regiones anatómicas en ovicaprino de yacimientos del Hierro I.
KOBIE (Paleoantropología n.º 28), año 2009

ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, ALAVA) 67
Cabeza 34,65 41,03 38,5 31,21
Tronco 18,8 19,81 23,4 16,56
Extremidades 46,54 39,15 38,0 52,22
Total NR 1362 212 5997 157
Tabla 17.- Porcentajes de regiones anatómicas en ovicaprino de yacimientos del Hierro Il.
Sin embargo, durante la Segunda Edad del Hierro la distribución de Caranca se parece a la de Ubiema y ambas se caracterizan por un alto grado de fragmentación de las arcadas dentarias. En las tres restantes el esqueleto de las extremidades es el mejor representado (Tabla 17).
ID.b.3.2.- Presencia relativa de la oveja y de la cabra.
NRoveja 20 83
NR cabra 17 73
NR ovejalcabra 1,17 1,13
Tabla 18.- Relación entre los restos de oveja y los de cabra en Caranca.
La dificultad intrínseca para la diferenciación entre los huesos de cabra y los de oveja aumenta cuando el material está fragmentado como es el caso de Caranca. Por esta razón la mayor parte de los fragmentos de ovicaprino han quedado sin especificación. No obstante la presencia de un reducido conjunto de restos susceptibles de ser asignados a cada una de las especies permite apreciar aunque sea de forma limitada la relación entre la oveja y la cabra en los distintos horizontes (Tabla 18).
30 17 5
31 15 7
0,96 1,13 0,71
La relación entre ambas cabañas está muy equilibrada en Caranca salvo en la muestra procedente del Bronce Final que debido a su reducido tamaño puede ser más sensible a los efectos de factores aleatorios. Una representación tan pareja entre la oveja y la cabra como la que se da en Caranca no es precisamente el caso más frecuente en otras muestras de la Edad del Hierro.
Tabla 19 .- Relación entre los restos de oveja y los de cabra en yacimientos del Hierro.
Así en el Hierro 1 de siete muestras analizadas (Tabla 19) hay tres en las que la relación está próxima al equilibrio (Mendavia, Soto de Medinilla I-11yCaranca1), hay un caso en el que la oveja es más escasa que la cabra (Santa Ana 1) y en el resto se aprecia un claro predominio de la oveja. Sin embargo durante la época Celtibérica el único caso en que se mantiene el equilibrio entre las dos cabañas se da en Caranca mientras que en las otras seis se generaliza el predominio del ganado lanar.
KOBIE (Paleoantropología n.0 28), año 2009

68 PEDRO CASTAÑOS Y JONE CASTAÑOS
ill.b.3.3.- Distribución de edad y de sexo.
D4Ml- 0-3 meses 2
MI+/- 3 meses Infantil 2 3
Ml+M2- 3-6 mes. 3 5
M2+!- 6 meses 3
M2+M3- 9-15 mes. 8 Juvenil
M3+/- 15-24 m. 7 31
M3+ Subadulto 8 39
M3++ 5 39
M3+++ Adulto 6 29
Totales 32 159
Tabla 20.- Distribución de edad del ovicaprino de Caranca.
Teniendo en cuenta que los maxilares son las regiones mejor representadas de la muestra de ovicaprino, las estimaciones de la edad se han realizado a partir del estado de emergencia de las piezas dentarias y del desgaste de los últimos molares. Al tratarse de una región del esqueleto que no permite atribuciones específicas la distribución de edades se realiza conjuntamente para ambas cabañas (Tabla 20).
A la vista de los datos se puede afirmar que no se observan diferencias significativas de la estructura de edad en los distintos horizontes cronológicos salvo en el más antiguo. En el Medievo y en toda la Edad del Hierro el predominio de los animales mayores de dos años (subadultos y adultos) es notable y constante.Al
ill.b.3.4.- Tamaño y alzada.
OVEJA
4
2 7
3 2 14
6
3 2 15
3 3 45
16 8 5 76
14 10 2 70
9 2 47
52 24 17 284
igual que en el ganado mayor con estos datos es razonable postular para el ovicaprino un patrón de aprovechamiento pecuario también de carácter secundario. Son menos los animales inmaduros que se sacrifican en vistas a su exclusivo consumo cárnico. La mayoría se conservan hasta la madurez con el fin de aprovechar antes de su muerte productos secundarios como la leche, la reproducción o la lana en el caso de la oveja. Sin embargo, al igual que ocurría con el bovino la muestra correspondiente al Bronce presenta una distribución más "equilibrada" entre juveniles y adultos. Estas diferencias en el ovicaprino pudieran estar relacionadas con fenómenos de trashumancia aunque la escasez de datos en Caranca no permite conclusiones definitivas.
DMB
DmB
50,5
31,5
Ad
AT
25,5
25
28 25,5 28 28,5 26,5 Ap
ASp
30,5
28,5 26,5 24,5 26,5 28,5 26
m
KOBIB (Paleoantropologfa n.º 28), año 2009

ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, ALAVA)
LMP
LS
AS
LmC
Ad
LM
Ap
AD
Ad
Ed
Húme.AT
Metac.Ap
31,5
25,5
20,5
18,2
23
116,5
20,5
11,7
21,5
14,7
6
6
26,5 LMI
215 LMm
16,3 El
16,6 Ad
21,5 24
118,5 102
21,5 18
13 8,8
23,5
14,4
24,5-28,5
18-21,5
24
19
26 27,5 22
24 26 21,5
14,6 15,9 11,9
16 17,5 14,5
24,5 22,5 23,5
20,5 18,7
26,16 1,4 5,3
19,7 1,33 6,7
Tabla 22.- Medidas aisladas y resumen estadístico de la oveja de Caranca.
27,5 26 LM
26 24 AM
14,9 14,l
17,5 . 16,4
25 22 23,5 22,5
LM
Ap
AD
Ad
Ed
Tibia Ad 11 21,5-25 23,2
Astr. LMl 5 22-27,5 25,8
69
52,5
16,8
120,5
17,7 18,8
8,8
21
14,5
1,08 4,6
2,2 8,7
La Tabla 22 recoge las escasas medidas de oveja entre las que destacan los extremos distales de húmero y tibia, el proximal de metacarpo y los astrágalos de los que se ofrecen los correspondientes resúmenes estadísticos. Los valores medios de las medidas más frecuentes indican que la oveja de Caranca está en cuanto a dimensiones dentro del dominio de variación de sus congéneres de la Edad del Hierro de la Península que presenta en esta cabaña doméstica una disminución osteológica a partir del Bronce (Tabla 23). Los datos de esta última tabla comparativa están recogidos a partir de la mayoría de los yacimientos publicados en la Península para cada horizonte cronológico incluyendo con el mismo criterio la altura en la cruz.
Neolítico 66 24,6-32 28,16 Neolítico 24 19-27 21,88
Calcolítico 279 24,5-33,5 28,7 Calcolítico 165 19,5-30 22,7
Bronce 199 22,5-32,5 26,78 Bronce 133 17-25,5 21,22
Hierro 80 22,7-30,5 26,38 Hierro 75 16,6-24,5 20,34
Astrág.LMI Alt. Cruz
Neolítico 113 23,5-31,5 28,03
Calcolítico 369 24,5-33,5 28,3 Calcolítico 94 60-74 65,34
Bronce 197 23-31,7 26,79 Bronce 53 52,6-69 59,7
Hierro 23 23-39 27,64 Hierro 22 51,7-65,8 60,8
Tabla 23.- Variación de algunas medidas y de la altura en la cruz en ovejas de la Península.
KOBIE (Paleoantropologfa n.º 28), año 2009

70 PEDRO CASTAÑOS Y JONE CASTAÑOS
Por lo que se refiere a la alzada de la oveja de Caranca, se ha podido estimar la altura en la cruz a partir de media docena de metapodios utilizando los factores de Teichert (Driesch & Boessneck, 1973) que se recogen en la Tabla 21. Se incluyen en la misma las dos estimaciones de cabra a partir de los factores de Schramm (Driesch & Boessneck, 1973).
Metacarpo Medievo
Metacarpo Medievo
Metacarpo Hierro II
Metacarpo Hierro II
Metacarupo Hierro II
Metatarso Hierro II
Radio Hierro I
Metacarpo Hierro 1
Las cuatro estimaciones del Hierro para la oveja están dentro del dominio de variación de la oveja peninsular del Hierro (Tabla 24) e incluso su media queda ligeramente por debajo de las de sus coetáneas. Se constata con Caranca la disminución de la altura del ganado lanar después del Calcolítico en los yacimientos peninsulares.
115,5 4,89 56,4
108,5 4,89 53,05
116,5 4,89 56,9
118,5 4,89 579
102 4,89 49,8
120,5 4,54 54,7
152,5 3,98 60,7
106 5,75 60,9
Tabla 24.- Estimaciones de la altura en la cruz de la oveja y cabra de Caranca.
CABRA
Al igual que para la oveja se recogen las medidas de cabra y los resúmenes estadísticos de la tibia y del astrágalo en la Tabla 25. Como puede comprobarse los datos osteométricos para la cabra son más pobres y ello no es exclusivo de esta muestra sino que ocurre en la mayor parte de las faunas protohistóricas publicadas en la Península Ibérica. Por esta razón las comparaciones son más escasas y además menos fiables.
Clavija: Radio:
DMB 34 31 LM 152,5
DmB 21,5 18,3 Ap 29,5
Asp 28
Húmero: AD 17,8
Ad 38 26,5 Ad 29,5
AT Tibia:
Ad 23,5 25
Lo primero que puede constatarse es que las medidas de la cabra de Caranca están dentro del área de dispersión de las de sus congéneres del Hierro peninsular (Tabla 26). Lo mismo cabe decir de las dos únicas estimaciones de altura en la cruz obtenidas (Tabla 24). En esta especie no se aprecia una disminución osteométrica tan clara como en la oveja aunque la escasez de datos pueda tener algo que ver con este hecho.
Escápula: Metacarpo:
LMP 31,5 LM 106
LS 25 Ap 23,5
AS 19,2 AD 15,8
LmC 19 Ad 26
24,5 Ed 15,7
26,5 24 22,5 23 22,5
KOBIB (Puleoantropología n.º 28), año 2009
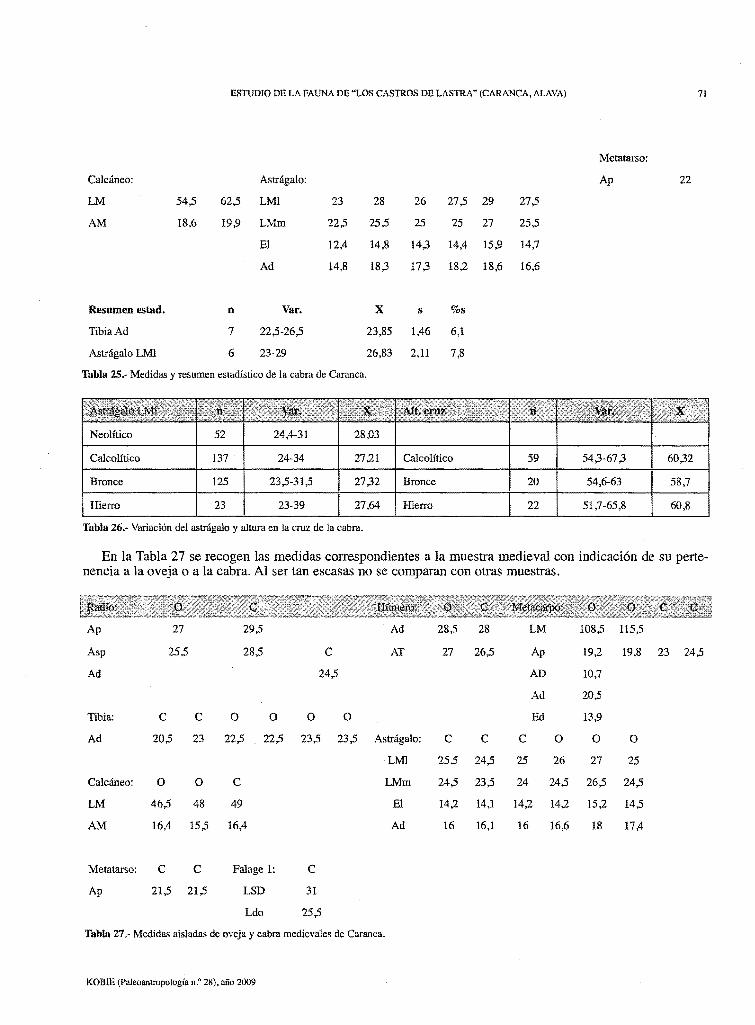
ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, ALAVA) 71
Metatarso:
Calcáneo: Astrágalo: Ap 22
LM 54,5 62,5 LMI 23 28 26 27,5 29 27,5
AM 18,6 19,9 LMm 22,5 25,5 25 25 27 25,5
El 12,4 14,8 14,3 14,4 15,9 14,7
Ad 14,8 18,3 17,3 18,2 18,6 16,6
Resumen estad. n Var. X s %s
TibiaAd 7 22,5-26,5 23,85 1,46 6,1
Astrágalo LMI 6 23-29 26,83 2,11 7,8
Tabla 25.- Medidas y resumen estadístico de Ja cabra de Caranca.
Neolítico 52 24,4-31 28,03
Calcolítico 137 24-34 27,21 Calco lítico 59 54,3-67,3 60,32
Bronce 125 23,5-31,5 27,32 Bronce 20 54,6-63 58,7
Hierro 23 23-39 27,64 Hierro 22 51,7-65,8 60,8
Tabla 26.- Variación del astrágalo y altura en la cruz de la cabra.
En la Tabla 27 se recogen las medidas correspondientes a la muestra medieval con indicación de su pertenencia a la oveja o a la cabra. Al ser tan escasas no se comparan con otras muestras.
Ap 27 29,5 Ad 28,5 28 LM 108,5 115,5
Asp 25,5 28,5 e AT 27 26,5 Ap 19,2 19,8 23 24,5
Ad 24,5 AD 10,7
Ad 20,5
Tibia: e e o o o o Ed 13,9
Ad 20,5 23 22,5 22,5 23,5 23,5 Astrágalo: e e e o o o
LMI 25,5 24,5 25 26 27 25
Calcáneo: o o e LMm 24,5 23,5 24 24,5 26,5 24,5
LM 46,5 48 49 El 14,2 14,1 14,2 14,2 15,2 14,5
AM 16,4 15,5 16,4 Ad 16 16,l 16 16,6 18 17,4
Metatarso: e e Falage 1: e Ap 21,5 21,5 LSD 31
Ldo 25,5
Tabla 27 .- Medidas aisladas de oveja y cabra medievales de Caranca.
KOBIE (Paleoantrnpología n.º 28), año 2009
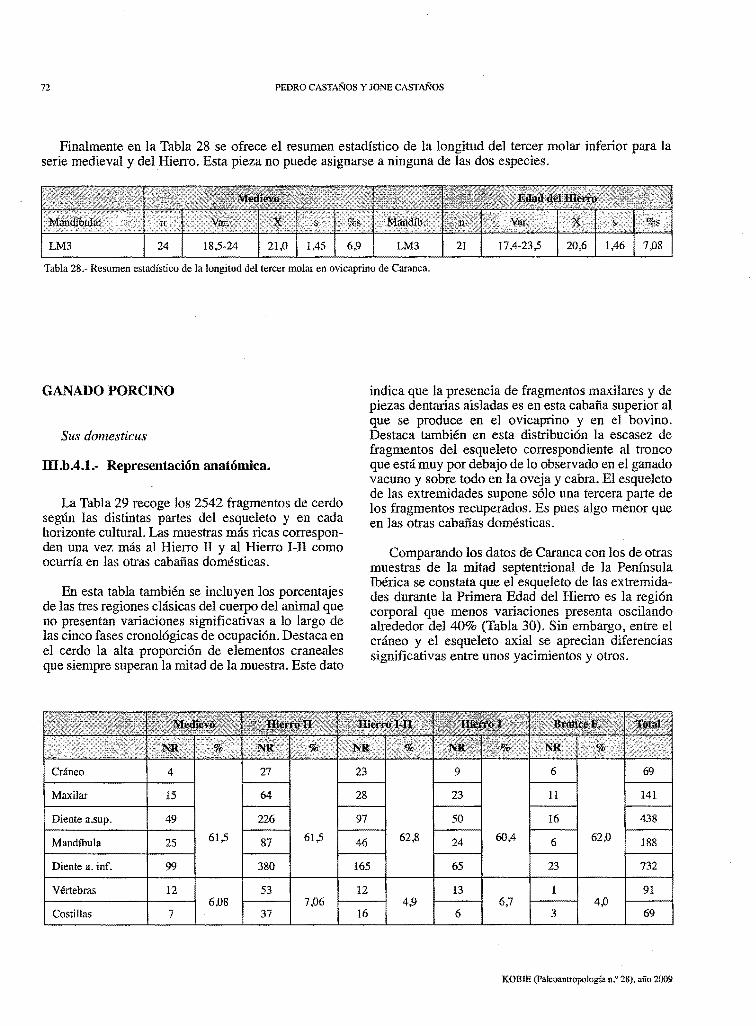
72 PEDRO CASTAÑOS Y JONE CASTAÑOS
Finalmente en la Tabla 28 se ofrece el resumen estadístico de la longitud del tercer molar inferior para la serie medieval y del Hierro. Esta pieza no puede asignarse a ninguna de las dos especies.
Tabla 28.- Resumen estadístico de la longitud del tercer molar en ovicaprino de Caranca.
GANADO PORCINO
Sus domesticus
ID.b.4.1.- Representación anatómica.
La Tabla 29 recoge los 2542 fragmentos de cerdo según las distintas partes del esqueleto y en cada horizonte cultural. Las muestras más ricas corresponden una vez más al Hierro II y al Hierro 1-II como ocurría en las otras cabañas domésticas.
En esta tabla también se incluyen los porcentajes de las tres regiones clásicas del cuerpo del animal que no presentan variaciones significativas a lo largo de las cinco fases cronológicas de ocupación. Destaca en el cerdo la alta proporción de elementos craneales que siempre superan la mitad de la muestra. Este dato
Cráneo 4 27
Maxilar 15 64
Diente a.sup. 49 226
Manch'bula 25 61,5 87 61,5
Diente a. inf. 99 380
Vértebras 12 53 6,08 7,06
Costillas 7 37
23
28
97
46
indica que la presencia de fragmentos maxilares y de piezas dentarias aisladas es en esta cabaña superior al que se produce en el ovicaprino y en el bovino. Destaca también en esta distribución la escasez de fragmentos del esqueleto correspondiente al tronco que está muy por debajo de lo observado en el ganado vacuno y sobre todo en la oveja y cabra. El esqueleto de las extremidades supone sólo una tercera parte de los fragmentos recuperados. Es pues algo menor que en las otras cabañas domésticas.
Comparando los datos de Caranca con los de otras muestras de la mitad septentrional de la Península Ibérica se constata que el esqueleto de las extremidades durante la Primera Edad del Hierro es la región corporal que menos variaciones presenta oscilando alrededor del 40% (Tabla 30). Sin embargo, entre el cráneo y el esqueleto axial se aprecian diferencias significativas entre unos yacimientos y otros.
9 6 69
23 11 141
50 16 438
62,8 24 60,4 6 62,0 188
165 65 23 732
12 13 1 91 4,9 6,7 4,0
16 6 3 69
KOBIE (Paleoantropología n.• 28), año 2009

ESTUDIO DE LA FAUNA DE "LOS CAS1ROS DE LAS1RA" (CARANCA, ALAVA) 73
Escápula 6 30
Húmero 10 39
Radio 5 32
illna 2 19
Carpo 6 22
Metacarpo 8 18
Pelvis 4 10
Fémur 10
Rótula
Tibia 13 34
Calcáneo 3 9
Astrágalo 2 12
Resto tarso 2 32,3 9 31,4
Metatarso 9
Metapodio ind. 7 50
Falange 1 21 61
Falange 2 8 39
Falange 3 8 18
Totales 312 1274
Tabla 29.- Distribución anatómica de los restos de cerdo.
El modelo más frecuente y que se repite en ocho de los nueve conjuntos considerados es aquel que da a la cabeza una proporción relativa mayor que al tronco e incluso en dos ocasiones (Mendavia y Caranca) mucho mayor. Parece claro que en estas dos
Cabeza 38,6 41,3 32,9 36,9
Tronco 22,03 20,6 21,2 22,3
Extrem. 39,3 37,9 45,8 40,7
Tot. NR 608 232 325 376
14 9 6 65
17 8 5 79
9 2 19 48
13 6 41
16 2 2
7 4 3 40
6 21
2 17 14
4
23 5 4 79
2 16
8 4 27
4 32,2 32,8 36,0 16
6 4 2 22
225 93 28 965
27 18 5 132
16 7 71
11 5 43
571 283 102 2542
localidades la fragmentación de las arcadas dentarias es más intensa. Sin embargo Santa Ana es el único caso en el que la presencia de fragmentos de vértebras y costillas supera a los del cráneo.
32,5 22,5 47,08 46,5 60,0
26,1 30,4 6,8 10,7 6,6
41,3 47.05 46,1 42,7 33,3
1562 102 206 456 285
Tabla 30.- Frecuencias anatómicas del cerdo en el Norte de la Península para la Primera Edad del Hierro.
KOBIE (Paleoantropología n.º 28), año 2009

74 PEDRO CASTANOS Y JONE CASTANOS
En la Epoca Celtibérica las tres muestras consideradas ofrecen una distribución bastante semejante con escasa representación de elementos del tronco y equilibrio entre los de la cabeza y extremidades que en el caso de Caranca se desvía a favor del cráneo (Tabla 31).
Tronco 10,9 9,7 7,06
Extremidad 40,9 43,2 31,4
Total NR 1032 606 1274
Tabla 31.- Frecuencias anatómicas del cerdo en el Norte de Ja Península para la Segunda Edad del Hierro.
ill.h.4.2.- Distribución de edad y sexo.
La Tabla 32 recoge la distribución por edades de la cabaña porcina en los distintos horizontes cronoló-
D4Ml- 0-6 meses 3 Infantil
Ml+/- 6 meses 2
Ml+M2- 6-12 mes. 2
M2+/- 12meses Juvenil
M2+M3- 12-20 mes. 3
M3+/- 20-24 mes. 6
M3+ Subadulto 9
M3++ 5 Adulto
M3+++ 3
Totales 34
Machos 16
Hembras 14
Tabla 32.- Distribución de edad y sexo del cerdo.
ill.b.4.3.- Tamaño
La cabaña porcina suele proporcionar pocos datos osteométricos más que por lo reducido de sus conjun-tos óseos debido al predominio de animales inmadu-
5
2
6
3
2
gicos así como la estimación del sexo cuando ésta era posible.
En el cerdo se observa un modelo de aprovechamiento que difiere de las otras cabañas. Mientras en el bovino y ovicaprino el predominio de subadultos y adultos era constate, la explotación de las piaras está más equilibrada entre los inmaduros y los adultos. En general se sacrifican más animales entre el año y los dos años de edad de forma que la cohorte de juveniles muestra porcentajes similares a los que corresponden a la cohorte de adultos. Estas pautas de selección parecen adaptarse a las propias características biológicas de la especie. La mejor relación entre el peso y la edad de sacrificio en el ganado porcino actualmente se estima en los doce meses en ejemplares estabulados. Esta optimización se retrasa casi un año en aquellos ejemplares criados en semilibertad. Más allá de los dos años el único producto secundario que suele obtenerse del cerdo es la reproducción. Bien diferente a lo que ocurre en la oveja y en el bovino doméstico en los que la lana, leche o fuerza de tracción de cara a labores agrícolas son aportaciones sustantivas en la economía agropecuaria.
3 12
3 6
3 1 7
4
4 JO
6 2 3 18
10 3 5 28
10 3 22
4 2 3 14
22 44 12 9 121
11 16 7 50
11 30 6 1 62
ros en sus muestras. En Caranca las medidas son escasas sobre todo en las ocupaciones medieval y del Bronce Final por lo que sólo ofrecemos estos valores de fonna aislada (Tabla 34).
KOBIE (Paleoantropología n.• 28), año 2009
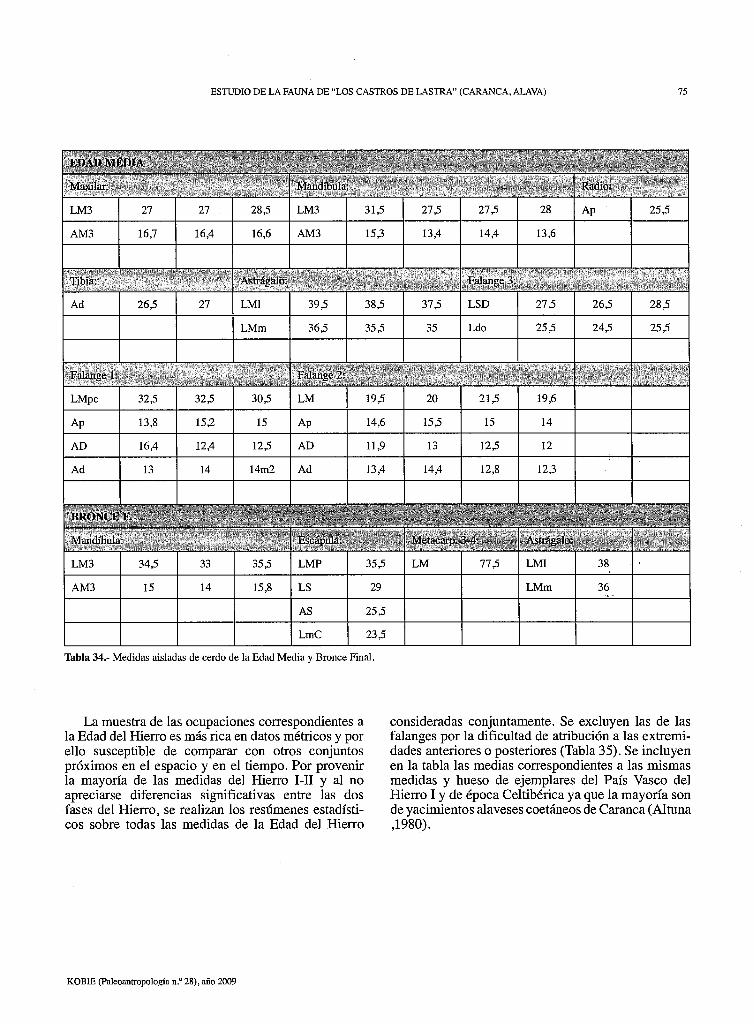
ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, ALAVA) 75
LM3 27 27 28,5 LM3 31,5
AM3 16,7 16,4 16,6 AM3 15,3
Ad 26,5 27 LMl 39,5 38,5
LMrn 36,5 35,5
LMpe 32,5 32,5 30,5 LM 19,5
Ap 13,8 15,2 15 Ap 14,6
AD 16,4 12,4 12,5 AD 11,9
Ad 13 14 14m2 Ad 13,4
LM3 34,5 33 35,5 LMP 35,5
AM3 15 14 15,8 LS 29
AS 25,5
LmC 23,5
Tabla 34.- Medidas aisladas de cerdo de la Edad Media y Bronce Final.
La muestra de las ocupaciones correspondientes a la Edad del Hierro es más rica en datos métricos y por ello susceptible de comparar con otros conjuntos próximos en el espacio y en el tiempo. Por provenir la mayoría de las medidas del Hierro 1-II y al no apreciarse diferencias significativas entre las dos fases del Hierro, se realizan los resúmenes estadísticos sobre todas las medidas de la Edad del Hierro
KOBIE (Paleoantropologfa n.º 28), año 2009
27,5 27,5 28 Ap 25,5
13,4 14,4 13,6
37,5 LSD 27,5 26,5 28,5
35 Ldo 25,5 24,5 25,5
20 21,5 19,6
15,5 15 14
13 12,5 12
14,4 12,8 12,3
LM 77,5 LMl 38
LMrn 36
consideradas conjuntamente. Se excluyen las de las falanges por la dificultad de atribución a las extremidades anteriores o posteriores (Tabla 35). Se incluyen en la tabla las medias correspondientes a las mismas medidas y hueso de ejemplares del País Vasco del Hierro 1 y de época Celtibérica ya que la mayoría son de yacimientos alaveses coetáneos de Caranca (Al tuna ,1980).

76 PEDRO CASTAÑOS Y JONE CASTAÑOS
Mandíbula:
LM3
AM3
Escápula:
LMP
LS AS
LmC
Húmero:
Ad
AT
Radio:
Ap
Pelvis:
LA
AA
Tibia:
Ad
Calcáneo:
LM
AM
Astrágalo:
LMl
LMm
22
21
4
4
4
3
14
14
14
4
4
7
2
2
14
14
25,5-33,5
13,7-16,6
30-35,5
27-31,5
21,5-26,5
19,5-23,5
30-40
29-34
23,5-27,5
32-34,5
31-33
25,5-28
65,5-72
18,3-20
33-42,5
31,5-39
30,54
14,73
33,25
28,25
24,0
21,83
36,64
31,85
26,42
33,12
32,12
26,92
68,75
19,15
37,96
34,96
1,93
0,72
2,68
1,72
1,49
0,83
2,66
2,38
6,3
4,9
7;3
5,4
5,6
3,1
7,01
6,8
3
5
8
12
12
11
8
8
11
4
2
6
17
31,2
14,4
34,6
29,2
23,6
23,l
37,4
30,7
28,3
30,6
28,2
29,7
38,9
21
20
23
24
23
29
18
18
16
7
6
12
10
29,8
14,8
33,5
28,3
23
22,2
35,9
29,8
26,9
31,8
29
26,6
37,7
Tabla 35.- Resumen estadístico y comparación de medidas de cerdo de la Edad del Hierro.
Hay una gran semejanza métrica entre el cerdo de la muestra estudiada y los ejemplares del resto de Alava especialmente de los que corresponden a niveles celtibéricos que a su vez son ligeramente más pequeños que los de la Primera Edad del Hierro. También puede extenderse esta apreciación a los ejemplares navarros de Mendavia, a los riojanos de Santa Ana y a los burgaleses del Castro de Ubiema entre los conjuntos más próximos geográficamente.
ID.b.5.- PERRO
Canis familiaris
De los 35 restos de perro presentes todos menos uno proceden de la zona IIl y corresponden a la ocupación Celtibérica. Representan un mínimo de tres individuos distintos: dos adultos y un cachorro de unos seis meses que conserva aún el último molar de leche. Uno de los ejemplares adultos presenta una importante exostosis en el extremo distal del fémur y en el proximal de la tibia que por su desarrollo tuvo que dificultar sensiblemente el juego de dicha articulación (Tabla 36).
KOBIE (Paleoantropología n.º 28), año 2009

ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, Al.AYA) 77
Maxilar 2 Calcáneo
D.a. sup. 2 Astrágalo
D.a. inf. Metatarso 4
Vértebra 1 Falange 1 4
Costilla 12 Totales 34
Escápula 1 Tabla 36.- Distribución de Jos restos de perro.
Húmero 1
Metacarpo La Tabla 37 recoge las escasas medidas obtenidas
Fémur 2 así como la estimación de la altura en la cruz de uno
Tibia de los animales utilizando los factores de Koudelka (1885) y Harcourt (1974).
LMP 29 LMl 21 LM 171 Ad 21,5 LM 66,5
LS 26 AMI 8,3 Ap 36 Ap 9
AS 18 AD 13,3 AD 7
LmC 23,5 Ad 34 Ad 8,8
Koud. Harcourt Alt. cruz
Fémur 171 3,01 3,4 51,47 58,14
Tabla 37 .- Medidas aisladas y altura en la cruz del perro.
Se trata de un perro de tamaño mediano a grande cuya presencia en el entorno geográfico de Caranca está registrada desde la Primera Edad del Hierro Maxilar 1 (Altuna, 1980) y que convive con otra variedad
Mandíbula canina de menor tamaño. Vértebra 2
ill.b.6.- GATO Costilla 6
Felis catus Escápula 1
El gato está representado mediante 15 restos que Húmero·
parecen corresponder a un mismo individuo de la Pelvis 2 ocupación medieval del Sector II (Tabla 38). La pre-
Tibia 1 sencia de dentición de leche y la falta de fusión epifi-sacia en los dos extremos del húmero y de la tibia indican que se trata de una cría. Carece por tanto de
Total 15
interés osteométrico. Tabla 38.- Restos de gato.
KOBIE (Paleoantropología n.º 28), año 2009

78 PEDRO CASTAÑOS Y JONE CASTAÑOS
ffi.b.7.- CIERVO
Cervus elaphus
El ciervo con 332 restos es el Ungulado salvaje mejor representado en la muestra de Caranca. Está presente en todos los horizontes cronológicos aunque la mayor parte de los restos corresponden a los niveles del Hierro (Tabla 39).
Cuerna 16
Cráneo
Maxilar
Diente a. sup. 16
Mandíbula 3
Diente a. inf. 6 17
Vértebras 4
Costillas 9
Escápula 2
Húmero 9
Radio 3
Ulna 1 3
Carpo 2
Metacarpo 2 6
Pelvis
Fémur 5
Rótula
Tibia 2 14
Calcáneo 2
Astrágalo 3
Resto tarso 4
Metatarso 1 5
Metapodio ind. 5
Falange 1 2 5
Falange 2
Falange 3
Totales 18 132
Tabla 39.- Distribución de Jos restos de ciervo.
9
1
1
12
2
7
4
3
3
4
6
1
4
2
8
2
2
6
3
2
85
7 33
3
4
5 2 36
5 11
7 3 40
2 7
6 2 21
2 2 10
2 14
8
2 14
2
9 24 80
2
9
2
5 4 33
6
1 6
4 10
2 15
7 16
10
2
2
68 29 332
KOBIE (Paleoanlropología n.º 28), año 2009

ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, ALAVA) 79
La distribución anatómica de los fragmentos muestra una escasa proporción de elementos del tronco (vértebras y costillas), elevada frecuencia de restos craneales debido a un gran número de piezas dentarias aisladas y porciones de todos los huesos de las extremidades aunque en diferente número. Parece por tanto razonable suponer que los venados cazados eran transportados al asentamiento ya despiezados y tras eliminar probablemente las vfsceras y trocear el
Ml+M2-
Ml+M2+/-
M2+M3-
M3+
M3++
M3+++ 1
Totales 2
Tabla 40.· Distribución de edad del ciervo.
Ad 45,5
Ed 29,5
LMP 59 Ad 58 57 48,5
LS 49 AT 54 52 47
AS 45,5
LmC 40,5
KOBIE (Paleoantropologfa n.º 28), año 2009
2
tronco del animal.
Las estimaciones de edad en este conjunto son escasas pero indican que la captura de venados se centraba preferentemente en lo animales adultos (Tabla 40). En la muestra del Bronce Final están representados tres adultos y un juvenil sin poder concretar más su edad.
2
3
2 2
3
11 3
Ad 18 20,5
51,5 Ap 52,5 59,5
49,5 Asp 51,5 53
Ad 47,5

80 PEDRO CASTAÑOS Y JONE CAS'D\ÑOS
EC 38 36,5 LM 53 Ad
AM 45,5
LMm 55 56 55,5 51,.5 Ad
El 31,.5 34 33,5 30,.5 Ed
Ad 36 37 37 36
LMpe 56 55,5 51 LM 37,5
Ap 22 21 19,4 Ap 20
AD 179 17,2 16,l AD 14,8
LMP 61 LM 269.
LS 47 Ap 46
AS 46,.5 AD 24,5
LmC 41,.5 Ad 46
Ed 30
Tabla 41.- Medidas aisladas de ciervo.
Las medidas obtenidas en esta muestra no son abundantes por lo que evitan cualquier comentario comparativo respecto de otros conjuntos próximos a Caranca desde el punto de vista espaciotemporal (Tabla 41). También es cierto que los conjuntos ricos en ciervo en estas épocas protohistóricas brillan por su ausencia. Sí puede decirse que las medidas aisladas obtenidas entran dentro de dominio de variación de los ciervos holocénicos peninsulares más pequeños que sus congéneres würmienses
No todo el material del ciervo llegó al yacimiento como consecuencia de las actividades cinegéticas. Hay un conjunto de media docena de fragmentos de
42,5 46 LDS 45,.5
Ldo 42,.5
42
27,5
AM 42,5 46,5 44,5 49 41,.5
LM 117,5
AM 37
cornamenta cuyo acarreo al lugar de habitación tuvo otra fmalidad. Se trata de una percha de desmogue o muda de la que se conserva la roseta basal acompañada de cinco fragmentos de candil claramente seccionados y separados de la misma. Uno al menos conserva íntegro el plano de sección realizada con un instrumento cortante. Estos candiles del asta de ciervo se utilizaban para diversos instrumentos artesanales especialmente mangos. El vecino yacimiento del Castro de Ubiema presenta un conjunto de candiles trabajados y retocados que aún conservan la huella del ánima del instrumento cortante (hoz, cuchillo o espada) en el que se utilizaron como asidero.
KOBIE (Paleoantropología n.º 28), año 2009

ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA,ALAVA) 81
ill.b.8.- CORZO
Capreolus capreolus
El corzo es el Ungulado salvaje con más baja cantidad de evidencias óseas en la muestra. Hay un fragmento de hemimandíbula izquierda que conserva el primer molar en la Zona III y un molar superior aislado en el Sector II correspondiente a la ocupación de la Segunda Edad del Hierro. Proceden en ambos casos de individuos adultos. Además hay una cuerna de desmogue en el mismo sector pero en la muestra del Hierro I-II.
ill.b.9.- JABALÍ
Susferus
Hay cinco restos atribuibles al jabalí por sus dimensiones. Todos ellos proceden del Sector II. Un fragmento distal de tibia (Ad 35,5) corresponde a la Segunda Edad del Hierro y los demás al conjunto del Hierro 1-II. Se trata de un fragmento de cráneo, otro de mandíbula y dos molares aislados que parecen corresponder a una hembra adulta. Conviene recordar que la dificultad de discriminación entre la forma doméstica y su agriotipo salvaje es alta y por tanto puede haber restos de jabalí incluidos entre los que se atribuyen al cerdo.
ill.b.10.- ZORRO
Vulpes vulpes
El zorro es el único Carnívoro salvaje presente en Caranca lo que indica el carácter urbanizado del asentamiento. Está presente con tres restos: un fragmento proximal de ulna izquierda y otro de metatarsiano 2 izquierdo en el nivel medieval además de un tercero también de ulna pero del nivel correspondiente al Hierro I-II.
ill.b.11.- CONEJO
Oryctolagus cuniculus
Este pequeño Lagomorfo está representado por otros tres restos: una vértebra en el Hierro II y sendos fragmentos de mandíbula y tibia en el Hierro l. Su escasa frecuencia parece reflejar o bien la rareza de esta especie en el entorno del yacimiento o el desinterés del grupo en la caza del mismo.
KOBJE (Paleoantropología n.° 28), año 2009
ill.b.12.- LIBBRE
Lepus capensis
La liebre aunque presente con siete restos también es una especie casi residual en el conjunto de Caranca. Hay media docena de restos en la fase medieval: maxilar, mandíbula (SDI 15,2), un fragmento de ulna y otro de pelvis (LA 10,2; AA 9,9). Representan un mínimo de tres individuos adultos. A ellos se añade un fragmento de metatarsiano del Hierro l.
ill.b.13.- ARDILLA
Sciurus vulgaris
Hay un cráneo fragmentado de esta especie en el Sector II y que corresponde al Hierro l. Su presencia indica la presencia de masas forestales en el propio asentamiento o sus alrededores.
ill.b.14.- TOPO
Talpa europaea
El elenco de Mamíferos de Caranca se completa con tres restos de topo: dos húmeros correspondientes al Hierro I y Hierro 1-II y un fragmento de fémur del Hierro I.
ill.b.15.- GALLINA
Gallus gallus
Los 34 restos de gallina se distribuyen de forma desigual a lo largo de las distintas fases de ocupación de Caranca. Extraña sobremanera la presencia de un húmero izquierdo completo en el conjunto atribuido al Bronce Final. Hace más de una década que tras la revisión de todos los hallazgos de gallina procedentes de yacimientos protohistóricos se constataba que la aparición de esta especie en la Península Ibérica no fue anterior a la Primera Edad del Hierro (Hemández, 1992). Este hueso de Caranca integrado entre la fauna del Bronce Final parece más bien el resultado de alguna intrusión posterior o de la mezcla parcial de restos entre niveles.
Más improbable resultaría atribuir la presencia de seis fragmentos del Hierro I y de 10 del Hierro I-TI a similares alteraciones. Por tanto la muestra de Caranca confirma la presencia del gallo doméstico en el norte peninsular a partir de la Primera Edad del Hierro.

82 PEDRO CASTAÑOS Y JONE CASTAÑOS
Ap 15,7 13,6 18,9
AD 5,9 4,5 6,6
Ad 12,5 10,5 14,3
Tabla 42.- Medidas aisladas de gallina (M=Medieval; H=Hierro; Br=Bronce)
La Tabla 42 recoge las medidas obtenidas que aunque escasas tienen su interés debido a la poca información osteométrica que se tiene hasta el presente de esta especie.
ID.b.16.- PERDIZ
Alectoris rufa/Perdix perdix
Hay sendos fragmentos proximales de húmero y fémur que resultan imposible atribuir con seguridad a la perdiz común (Alectoris rufa) o a la pardilla (Perdix perdix). En ambos casos se trata de taxones adaptados a espacios más o menos abiertos o a lindes boscosas.
ID.b.17.- CODORNIZ
Coturnix coturnix
El único resto de codorniz es un fragmento distal de Carpo-Metacarpo correspondiente a la ocupación del Bronce Final. Se trata en esta caso de una especie con amplia distribución y que ocupa habitualmente terrenos abiertos evitando el matorral denso y el bosque.
ID.b.18.- PALOMA
Columba livia/oenas
Hay un Tarso-Metatarso completo de una paloma en el conjunto medieval. Sus dimensiones son más pequeñas que las de la paloma torcaz (e .palumbus) pero con este simple resto resulta difícil determinar con garantías si se trata de la paloma bravía (C. livia) o de la zurita (C. oenas). Si se acepta que la primera suele presentar un marcado carácter mediterráneo habría que suponer que pudiera tratarse de la paloma zurita que suele además ocupar zonas próximas a masas forestales que la bravía evita.
ID.b.19.- CORNEJA O GRAJA
Corvus corone/frugilegus
Hay cuatro restos en el conjunto del Hierro I-II que corresponden a un córvido de mediano tamaño. Tres están fragmentados y sólo se conserva un húmero completo. Sus dimensiones permiten atribuirle tanto a la corneja (C. corone) como a la graja (Cfrugilegus). Se trata en ambos casos de especies ubiquistas con amplía distribución.
Húmero: LM 62,SAD 5 ,5
Ap 17 Ad 13,9 14,4
ID.b.20.- MIRLO/ZORZAL
Turdus sp.
En la ocupación medieval hay un húmero completo de Turdus. Se trata del Passeriforme de menor talla aparecido en Caranca. La variedad específica de este género dificulta su atribución concreta a partir de un solo resto.
Húmero: LM 29,8 AD 3
Ap 9,2 Ad 7,2
ID.b.21.- RAPAZ GRANDE
La muestra de fauna aviaria se completa con un fragmento distal de ulna correspondiente al Hierro sin más precisión y que pudiera pertenecer a una rapaz de la envergadura de un buitre.
KOBIE (Paleoantropología n." 28), año 2009

EST'ODlO llB LA F"Llt-:A DF. '"'1.-0S CASTROS Of. f__.\S'l'KA~ (CAltANCA, Al...AVA} 83
IV.- DISCUSIÓN
IV .a •• Fragmentación del material.
El grado de fragmemación de Jos huesos, estimado indirectamente por Ja relación del peso de Ja fracción identificada respeclo úe la que queda sin identificar, está dentro de Jos valores de Ja mayoría de las muestras de Ja Edad del Hierro de Ja mitad septentrional de Ja Península (Tabla 43) .
SsCJtojos 90.8 9.2
Sta. Ano 1 97.S 2.5
Sta. An.'1! 00.S 9.5
~·fenJa\'ia 86,3 13,7
So10 Medill. I 71.7 28,3
Soto Mcdin. U 81,4 18,6
Ca~tillü 67,1 32,9
Er.i Alta 76,8 2~,2
A pesar de que Caranca ofrece una de las porciooes más altas <lc material sin identificar conviene también señalar que se trata de uno de Jos yacimieutos con más riqueza faunfstica de su enlomo . Sólo las ex.cepcionales muestras óscas procedentes de Sacaojos y La Hoya (con 16.227 y 16.068 restos identificados respectivamente) aportan conjuntos comparables cuantitativamente al formado por los 12.848 restos de los niveles correspondicmes a Ja Edad de Hierro en Caranca.
Soto Mc<lin. lll 73,3 26,7
ca~ti1lu 6 1,4 38,6
Era /\Jt:.1 21.8 78,2
V&lori~ 82,9 17;1
Padilla 69 31
Ubien<1 96.5 3"¡1
Cara:nca 70,0 30,0
ThbJa 43.- Pon:.-eot<ijes Ud pi::so del cuattrial idenliílrudu y i.iD identificar del. Hierro.
IV.b.- Presencia relativa de las distintas cabañas.
Las figuras siguientes (Fig. 3 a 5) representan los porcentajes de restos y pesos de Jos Ungulados en distintas muestras de la Primcra y Segunda. Edad del Hierro en yacimientos de las cuencas del Duero y Ebro con el fin de contextualizar Jos datos de Ja fauna de Caranca.
Iln Jos asentamientos del Hierro T de la zona sep· tcnlrional <le la Península hay un e>'Pectro faunístico <¡ue se repite en la mayor parte de Jos casos. Cuando se calculllJl las frecuencias a partir del número de restos el ganado ovicaprino ocupa el primer puesto seguido del bovino y del cerdo. Sin embargo si las estimaciones se realizan a partir del peso de los huesos los úos primcros puestos se invierten e incluso el cerdo unas veces iguala y otras incluso supera a la oveja y cabra.
1 1.~, ~ 1 ' 1 ' 11'' ~ · '. l r:.1· 1, 1·1 1 1~ 1 : 1, . l ~' ·11'· 1 ·.•;,. 1 · 1 · ' 1 ~ 1, ¡ 11
·.11d_~Jd1 ¡1_
100%
80%
40%
20%
0% 1 2 3 4 5 s 7 a e 10 11 12
• Ung.s. e Sus CO/C • Bos • Equus
Fig. 3.· Histogrii.nu etc frecuencia ele restos de Uogulttdos en yOéimlentos llel llierru 1 (l. H~nayo 2. La Hoya D2·1 l. P. De Orn 4. Sta. Ana I 5. S~caojos 6. Me:.l)d~\'ia 7. S..f\.tedinilla J.[1 8. C.'\stillo H.I 9. Era AJta HJ 10. C.W-dnca 11. Bcrbeia 12. Boruhonc,<;.)

PEDRO CASTJ\NOS Y JONE CASTANOS
120
100
80
60 40
20
o
• Uog.s. a Sus
aO/C
•Bos •Equus
Fig. 4.- Histograma.o; de frocucncilL" de pcsns de Un;;ulados en algunas yaciwientos del Hie.111> 1.(1. Bel'bei3 2. Sta.Ana l 3. S::ic::aojos 4. ~1.cnda\'ia 5. l..11 Hnyilo B2-1 6. S..1Vtedinil111 1-11 7 . Darabe>ncs 8. Ouaoca D.
Esta situación se repite en diez de las doce muestras de esta época entre las cuales está la de Caranca. Hay dos casos en los que el bovino es la cabaña domiuante con cualquiera de los parámeu·os con los que se haga el c:ilculo (Berbeia y Barahones). Y en el caso de Peñas de Oro la cabaoa porcina Loma el protagonismo seguida de cerca por el bovino. Se trata del único caso con esta distribución.
La situación en el Hierro Il es más favorable al ganado mayor ya que éste es el mejor representado en seis de los nueve conjuntos considerados eu número de re.$los (Fig. 5). Tan sólo La Mota. Castillo y Caranca se desvían ligeramente de este modelo ya que el ovicaprino presenta la mayor frecuencia cuando se trata del número de restos pero se asimila al resto si considcr.llllos la biomasa. Escc dominio en peso del ganado vacuno en todos los asentrunientos de las cuencas del Ebro y Duero contrasta con ouas
lnfuntil 8.!16 14.z
Juvenil 19,3 21,4
SubaduJto 45,1 28.5
Adulto 22.~ 35,7
TotaJ l\1MI 62 14
muestras más meridionales en las que el ovicaprino ocupa e l primer puesto.
100%
60% • Ung.s.
60% o Sus oOIC
40% • Bos 20% Equus
0% 2 3 4 5 6 7 8 9
Fi¡¡:. 5..--Hi..,.tn~'l"llmas de frocui:nciu." de n:stn" de UnguJados en el Hierro Il. (l. Ubierna 2. ).., Hoya A 1-3 n 3. S. Modinill• W 4.Ela Alm H.11 5. Villoría 6. Podilla 7. La Mu"' 1-Il 8. Casúllo H.Il 9. Coronca 11).
IV.C.· APKOVECBAl.\UENTO PECUARIO.
IV.e. l.- Ganado vacuno
Si se comparan la escrucntra de edad del bovino doméstico de Caranca con la de ou-as muestras· (Tabla 13) se aprecia que en general el citado predominío de subadultos y adultos es conSUll)lc tanto en el Hierro J como durante los niveles Celtibéricos al menos eo la zona más septentríonal de la Península. Lamentablemente el nivel de fragmentación de la mue~tra apenas permite estimaciones acerca del sexo de los animales. Para el Hierro l en Ja muestrJ de Ja cuenca del Duero se incluyen los datos de Soto de Medinilla, Era Alta y CastiUo mientras que e11 Ja muestra del Hierro 11 están sumados Jos efe<..1ivos de Ubierna, Castillo, Era Alta , Valoria y Padilla.
9.7 6.6 9.64
171!7 20 19,4
9,7 20 2.'1,8
63.4 53,3 43,7
41 15 132

ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA,ALAVA) 85
Infantil 7,4 2,7 6,6 5,5
Juvenil 13,9 22,2 23,03 19,7
Subadulto 24,06 19,4 24,1 22,5
Adulto 54,5 55,5 45,05 51,8
Total NMI 187 36 91 314
Tabla 13.- Porcentajes de cohortes de edad del bovino en yacimientos del Hierro.
IV.c.2.- Ovicaprino
En el contexto de la Primera Edad del Hierro de las cuencas del Ebro y del Duero hay algunas variaciones significativas entre las distribuciones de edad del ganado ovicaprino de unos yacimientos y de otros (Tabla 21). Así Sacaojos ofrece una distribución
Infantiles 23,1 14,7
Juveniles 24,5 20,5
Sub adultos 33,1 8,8
Adultos 19,0 55,8
Total NMI 220 34
Infantiles 12,2 6,2
Juveniles 22,6 2,64
Subadultos 23,2 24,5
Adultos 41,6 42,7
Total NMI 526 159
cercana a una curva normal cuyo valor modal coincide con las cohortes intermedias (juveniles y subadultos). Sin embargo Caranca presenta una mayor desviación hacia los adultos. Mendavia es la muestra que más se parece a las muestras valHsoletanas que ofrecen en conjunto una distribución bimodal con máximos relativos en los juveniles y sobre todo en los adultos.
15,l 4,1 14,2
27,5 12,5 22,0
12,0 33,3 21,8
45,3 50,0 42,5
86 24 364
9,2
24,5
23,8
42,l
685
Tabla 21.- Porcentajes de cohortes de edad del ovicaprino en yacimientos del Hierro.
KOBIE (Paleoantropología n.º 28), año 2009

86 PEDRO CASTAÑOS Y JONE CASTAÑOS
El cuadro es más homogéneo aunque también más reducido en el Hierro II donde la frecuencia relativa de cada cohorte de edad es bastante parecida en Caranca y en las cinco muestras del valle del Duero. Además no parece que se den diferencias significativ~s entre los datos globales del Hierro I · y los del Hierro II en esta región septentrional de la Península. A lo largo de todo el período se mantiene un predominio de los subadultos y adultos aunque con frecuencias no tan marcadas como en el ganado vacuno.
Como suele ser habitual las estimaciones del sexo son tan escasas que cualquier comentario al respecto carece de significación.
20,8
Juvenil 33,3 71,4 31,2
Subadultos 33,3 6,2
Adultos 20,5 14,2 41,6
TotalNMI 39 14 48
IV.c.3.- Cabaña porcina
El patrón de aprovechamiento pecuario en el cerdo de Caranca coincide con el de otros asentamíentos coetáneos de la mitad septentrional de la Península (Tabla 33). Durante la Primera Edad del Hierro las frecuencias relativas de las distintas cohortes de edad de esta muestra alavesa se asemejan sobr~ !º?º a l~ ?e Sacaojos mientras que en época Celtibenca comc1den con las de los yacimíentos del valle del Duero.
La estimación del sexo muestra un equilibrio entre machos y hembras en la mayor parte de los períodos de ocupación. Solamente en la muestra del Hierro I-II las hembras duplican a los machos.
8,3 21,3 18,1
25,0 27,4 46,l 28,7
25,0 24,3 30,7 24,2
41,6 26,4 23,07 28,7
12 197 13 66
Tabla 33.· Porcentajes de cohortes de edad del cerdo en la Edad del Hierro del norte peninsular.
AGRADECIMIENTOS
Agradecemos a Paquita Sáez de Urturi los datos sobre el yacimíento y su confianza al encomendarnos el material de fauna para su estudio.
BIBLIOGRAFIA
AltunaJ.
1980 Historia de la domesticación animal en el País Vasco desde sus orígenes hasta la Romanización. Munibe, 32. San Sebastián.
Altuna J. & Marlezkurrena, K.
1990 Nourishment of animal origin of the inhabitants of the north of the Iberian Península during th first millennium before Crist Festschrift jür Hans R. Stampfli, Helbing & Lichtenhahn.
Castaños, P.
1989 Estudio de los restos óseos del Castro de Ubierna (Burgos), Kobie (Serie Paleoantropología) XVIII, 88-97. Bilbao.
Driesch v.d. A.
1976 Das vennessen von tierknochen aus vor und frühgeschinchtlichen siedlungen. Müních.
Driesch v.d. A. & Boessneck J.
1980 Tierknochenfunde aus Sacaojos bei La Bañeza (León), Studien uber frühe Tierknochenfunde von der lberischen Halbinsel, 7, 122-155. München.
Harcourt, R.A.
1974 Th_e ?og in Prehistoric and Early Historie Bntam. Joumal of Archaeol. Science J 151-175 ' '
KOBIE (Paleoantropo1ogía n.º 28). año 2009

ESTUDIO DE LA FAUNA DE "LOS CASTROS DE LASTRA" (CARANCA, ALAVA) 87
Fock, J.
1966 Metrische Untersuchungen an Metapodieneinigereuropaischer Ronderrasen. Dissertation Tieriirztl, Fakultiit der Universitiit München.
Hemandez, F.
1992 Sorne comments on the introduction of domestic fowl in Iberia, Archaeofauna I, 45-53.
Koudelka, F.
1885 Das Verhfiltnis der ossa longa zur SkeletthOhe bei den Saugethieren. Verhand.d. Naturforsh. Ver.Brünn,24, 127-153.
Liessau von Letow-Vorbeck C.
1994 Contribución al estudio arqueofaunístico durante la Edad del Hierro en la Submeseta Norte de la Península Ibérica, Publicaciones de la U .A. M. Madrid.
Mariezkurrena, K.
1986 La cabaña ganadera del Castillar de Mendavia (Navarra). Munibe (Antropología y Arqueología), 38, 119-169. San Sebastián.
Matolcsi, J. 1970 Historesche Erforschung der Kopergrosse
des Rindes auf Grund von ungarischen Knochenmaterial .Zeitschriftfür Tierzüchtung und Züchtungsbiologie, 87, 2,89-137. Hamburg und Berlín.
KOBIE (Paleoantropología n.º 28), año 2009
Miguel F. & Morales A.
1983 Informe sobre los restos faunísticos recuperados en el yacimiento del cerro de Santa Ana (Entrena, Logroño). Berceo I, 49-165.
Morales, A. & Liesau Von Lettow-Vorbeck, C.
1995 Análisis comparado de las faunas arqueológicas en el valle Medio del Duero (prov. Valladolid) durante la Edad del Hierro. In. : Delibes G. Et alii: Arqueología y Medio Ambiente, Junta de Castilla y León.
Schramm,Z.
1967 Long bones and eight in withers of goat. Koscedlugie a wysokosc w klebie u kozy. Poznan. Rocznide wyzsezj szkoly roiniczej w poznaniu, 36, 89-105.
Teichert, M.
1975 Osteometrische untersuchm;igen zur berchung der widerristhohe bei schafen. In. A.T. Clason Archaezoological Studies. Archaeozoological Conference 1974 Groningen, 51-59.
