efeitos do captopril e do enalapril sobre a resposta imune
TRANSCRIPT

UNIVERSIDADE FEDERAL DE MATO GROSSO
FACULDADE DE MEDICINA
COORDENAÇÃO DE PROGRAMAS DE PÓS-GRADUAÇÃO EM CIÊNCIAS DA
SAÚDE
Efeitos do captopril e do enalapril sobre a resposta imune humoral e celular
LIZIANE CRISTINA DE ALMEIDA
Cuiabá, MT.
2016.

Efeitos do captopril e do enalapril sobre a resposta imune humoral e celular
LIZIANE CRISTINA DE ALMEIDA
Orientadora: Deijanira Alves de Albuquerque
Tese apresentada ao Programa de Pós-
Graduação em Ciências da Saúde, para
obtenção do título de doutor em Ciências da
Saúde, área de concentração Doenças
Infecciosas e Parasitárias.
Cuiabá, MT.
2016


DEDICATÓRIA
Com profunda gratidão e amor, dedico este trabalho aos meus queridos pais, Herbert
e Vânia.

AGRADECIMENTOS
É um desafio muito grande utilizar duas páginas para agradecer tantas pessoas especiais
que marcaram essa minha trajetória.
Em primeiro lugar, quero agradecer a Deus, pela saúde e sabedoria para vencer meus
desafios e a oportunidade de concretizar mais uma etapa de minha caminhada
acadêmica.
Sem palavras para agradecer aos meus pais, Herbert e Vânia, ao meu irmão, Paulo
Henrique, pelo apoio incondicional e por sempre me incentivarem a realizar os meus
sonhos.
Ao meu marido, Flávio Arruda, pela compreensão, paciência, amor e apoio em todas
as minhas decisões.
Aos meus filhos, João Guilherme e Arthur, que trazem tanta luz e alegria para minha
vida.
A minha orientadora, Deijanira Alves de Albuquerque, pela contribuição para minha
carreira acadêmica e pela amizade, carinho e respeito dedicados a mim em todo este
período de convivência.
Obrigada a cada pessoa que ajudou em cada etapa dos procedimentos laboratoriais bem
como na fase final do trabalho, em especial à Carina de Souza Menezes, minha aluna
de iniciação científica, que cuidou diariamente com alegria e disposição dos animais.
Vocês foram muito queridos e essenciais.
Não posso deixar de agradecer minha amiga e grande incentivadora, Professora Maria
Gregório de Souza, que marcou importante presença na minha vida
acadêmica.
Ao Centro universitário Cândido Rondon (Unirondon), pelo incentivo a realização
deste trabalho.

Agradeço imensamente aos meus amigos do Unirondon pelo apoio, amizade e por
transformarem muitos momentos de angústia em momentos de entusiasmo e diversão.
Eu serei eternamente grata ao meu chefe, Professor Alceu Vidotti, pelo apoio,
compreensão e incentivo e também pela constante demonstração de sabedoria e
humildade.
Agradeço a todos que, de alguma forma, participaram e contribuíram com este
trabalho.
Agradeço também ao Departamento de Ciências Básicas em Saúde, Faculdade de
Medicina, UFMT, em cujas dependências foi realizado esse
trabalho.
E, por fim, ao Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq
# 471234/2011-7) e à Fundação de Amparo à Pesquisa do Estado de Mato Grosso
(FAPEMAT # 751305/2011) pelo apoio
financeiro.

RESUMO
Os inibidores da enzima conversora de angiotensina (ECA) atuam não só no tratamento
de hipertensão e nas doenças cardiovasculares, mas têm também propriedades
imunomoduladoras. O objetivo deste estudo foi determinar o efeito do captopril e
enalapril sobre a resposta imune humoral e celular em camundongos C57BL/6. No
presente trabalho, nós investigamos o efeito de enalapril sobre a produção de IgG1 e
IgG2c específicos para ovalbumina em camundongos imunizados com ovalbumina
EndoFit na presença de Alhydrogel, como adjuvante. Além disso, foi investigado o
efeito de captopril e enalapril sobre a frequência, número absoluto, e a ativação dos
linfócitos T e as células B e células natural killers (NK) in vivo utilizando citometria
de fluxo. Os resultados mostraram que o enalapril aumenta significativamente anti-
OVA IgG2c no soro em resposta secundária, sem afetar a síntese de IgG1. Além disso,
que o captopril, diferentemente do enalapril, aumenta a frequência, o número absoluto
e a ativação da subpopulação de linfócitos T CD4+. Assim, estes dados expandem o
nosso entendimento sobre as propriedades do captopril e enalapril sobre a resposta
imune.
Palavras chave: inibidores da ECA, captopril, enalapril, linfócitos, anticorpos IgG,
ovalbumina.

ABSTRACT
The angiotensin-converting-enzyme (ACE) inhibitors act not only in the treatment of
hypertension and in cardiovascular diseases but also have immunomodulatory
properties. The objective of this study was to determine the effect of captopril and
enalapril on humoral and cellular immune responses in mice C57BL6. In the present
work, we have investigated the effect of enalapril on ovalbumin-specific IgG1 and
IgG2c production in mice immunized with EndoFit ovalbumin in the presence of
Alhydrogel, as adjuvant. Additionally, we investigated the effect of captopril and
enalapril on the frequency, absolute number, and activation of T lymphocytes and B
and natural killer (NK) cells in vivo using flow cytometry. The results showed that
enalapril significantly increases anti-OVA serum IgG2c in the secondary response
without affecting IgG1 synthesis. Moreover, captopril, unlike enalapril, increased the
frequency, absolute number, and activation of the CD4+ lymphocyte subpopulation.
Thus, these data expand our understanding on the properties of enalapril on the immune
response.
Keywords: ACE inhibitors, captopril, enalapril, lymphocytes, IgG antibodies,
ovalbumin.

LISTA DE FIGURAS
Figura 1: Sistema Renina-Angiotensina-Aldosterona. O angiotensinogênio serve de
substrato para a renina dando origem à angiotensina I, que por sua vez, é convertida em
angiotensina II pela ação da ECA. Angiotensina II atua em receptores específicos.
Figura 2: Estrutura química do captopril (A) e do enalapril (B).
Figura 3: Tamanho vs. granularidade de células e gate na região dos linfócitos.
Figura 4: Ilustração das etapas utilizadas para a análise dos dados por citometria de
fluxo. Histograma de frequência da intensidade de expressão da molécula de superfície
CD4, demonstrando a população de linfócitos CD4+ na região P1. Distribuição pontual
de FL3/anti-CD4 versus FL4/anti-CD69 nos linfócitos esplênicos de animais do grupo
controle (painel A), grupo captopril (painel B) e grupo enalapril (painel C).
Figura 5: (A) Histograma de número de células vs. intensidade de fluorescência.
Células não marcadas lidas nos canais FL2, FL3 e FL4 representadas no primeiro pico,
e células marcadas e também lidas nos canais FL2, FL3 e FL4, representadas no segundo
pico. (B) Dot plots a partir dos quais se faz o cálculo da intensidade de
fluorescência dos fluorocromos referentes às células marcadas ou não.
Figura 6: Quantificação de IgG1 sérica anti-OVA.
Figura 7: Quantificação de IgG2c sérica anti-OVA.
Figura 8: Frequência de linfócitos T expressando CD4 em animais não tratados
(controle) ou tratados com captopril ou enalapril durante três meses. O asterisco
representa diferença estatística significativa.
Figura 9: Frequência dos linfócitos TCD8+ (painel A), dos linfócitos B CD19+ (painel
B) e dos linfócitos NK CD49b+ (painel C) em animais não tratados (controle) ou
tratados com captopril ou enalapril.
Figura 10: Número de linfócitos T expressando CD4 em animais não tratados (controle)
ou tratados com captopril ou enalapril. O asterisco representa diferença estatística
significativa.

Figura 11: Número absoluto dos linfócitos T CD8+ (painel A), dos linfócitos B CD19+
(painel B) e dos linfócitos NK CD49b+ (painel C) em animais não tratados (controle)
ou tratados com captopril ou enalapril.
Figura 12: Frequência de linfócitos T CD4 ativados em animais não tratados (controle)
ou tratados com captopril ou enalapril. O asterisco representa diferença estatística
significativa.
Figura 13: Frequência dos linfócitos T CD8+ (painel A), dos linfócitos B CD19+
(painel B) e dos linfócitos NK CD49b+ (painel C) ativados em animais não tratados
(controle) ou tratados com captopril ou enalapril.

LISTA DE TABELAS
Tabela 1: Anticorpos monoclonais conjugados com fluorocromos específicos para o
CD4, CD8, CD19, CD49b, CD69 ou MHC de classe II.
Tabela 2: Anticorpos monoclonais distribuídos em tubos separados para serem
analisados por citometria de fluxo.

LISTA DE SIGLAS E ABREVIATURAS
Ac - Anticorpo
Ang I - Angiotensina I
Ang II - Angiotensina II
ANOVA - Análise de variância APC - Aloficocianina
AT1-R - Receptor tipo 1da angiotensina II
AT2-R - Receptor tipo 2 da angiotensina II
C57BL/6 - Linhagem de camundongos C57 black 6
CD - Cluster of differentation ou grupo de diferenciação
CEMIB - Centro Multidisciplinar para Investigação Biológica
CMRV - Conselho Regional de Medicina Veterinária
CNPQ - Conselho Nacional de Desenvolvimento Científico e Tecnológico
CTL - Linfócito T citotóxico
DNA - Ácido desoxirribonucleico
ECA - Enzima conversora de angiotensina
ELISA - Enzyme Linked Immunosorbent Assay
FAPEMAT - Fundação de Amparo à Pesquisa do Estado de Mato Grosso
FC - Fraçãocristalizável FL - fluorescence laser
FSC - Forward Scatter
IFN-γ - Interferon gama
Ig - Imunoglobulina
IL - Interleucina
LPS - Lipopolissacarídeo
MFI - Média de intensidade de fluorescência
MHC- Complexo de histocompatibilidade principal
OVA - Ovalbumina
PAMP - Padrão molecular associado aos patógenos
PBS - Solução salina tamponada com fosfato
PE - Picoeritrina
PerCP- eFluor710 - Proteína peridina-clorofila eFluor 710nm
PerCP-Cy5.5 - Proteína peridina-clorofila cianina 5.5
RPMI - Roswell Park Memorial Institute
SAS - Statistical Analysis System

SRAA - Sistema renina angiotensina aldosterona
SSC - Side Scatter
Th - Linfócito T helper
TLR - Receptor do tipo toll
TNF - Tumor necrosis factor alfa ou fator de necrose tumoral alfa
UFMT - Universidade Federal do Mato Grosso
UNICAMP - Universidade Estadual de Campinas

SUMÁRIO
1. INTRODUÇÃO...................................................................................................... 16
1.1 Sistema renina-angiotensina-aldosterona ............................................................ 17
1.2 Aspectos gerais da resposta imunológica............................................................. 18
1.3 Bloqueio do sistema renina angiotensina-aldosterona: inibição da enzima de
conversões da angiotensina........................................................................................... 20
1.4 Efeitos dos inibidores da ECA no sistema imunológico...........................................21
2. OBJETIVOS............................................................................................................ 23
2.1 Objetivo geral........................................................................................................ 24
2.1 Objetivos específicos............................................................................................. 24
3. MATERIAL E MÉTODOS..................................................................................... 25
3.1 Animais................................................................................................................. 26
3.2 ELISA.................................................................................................................... 26
3.2.1 Medicamento e Tratamento................................................................................ 26
3.2.2 Antígeno e adjuvante.......................................................................................... 27
3.2.3 Imunização......................................................................................................... 27
3.2.4 Coleta de sangue................................................................................................ 27
3.2.5 Ensaio para determinação de IgG1 e IgG2c anti-OVA...................................... 27
3.2.6 Análise estatística............................................................................................... 28
3.3 CITOMETRIA DE FLUXO.................................................................................. 28
3.3.1 Medicamentos e tratamento................................................................................ 28
3.3.2 Obtenção de células esplênicas.......................................................................... 29
3.3.3 Imunofenotipagem celular.................................................................................. 29
3.3.4 Análise de imunofenotipagem............................................................................ 31
3.3.5 Análise estatística.............................................................................................. 34

4. RESULTADOS...................................................................................................... 35
4.1 Avaliação da produção de IgG1 e IgG2c anti-OVA............................................. 36
4.2 Avaliação da frequência de linfócitos T CD4+ in vivo......................................... 38
4.3 Avaliação da frequência das populações de linfócitos T CD8+, B e NK in 39
vivo..............................................................................................................................
4.4 Captopril aumenta o número absoluto de linfócitos T CD4+ in vivo.................... 40
4.5 Avaliação do número absoluto das populações de linfócitos T CD8+, B e NK 41
in vivo..........................................................................................................................
4.6 Avaliação da frequência de linfócitos TCD4 ativados in vivo.............................. 42
4.7 Avaliação da frequência das populações de linfócitos T CD8+, B e NK ativados in
vivo................................................................................................................................. 43
5. DISCUSSÃO........................................................................................................... 46
6. CONCLUSÕES....................................................................................................... 49
7. BIBLIOGRAFIA..................................................................................................... 51
8. ANEXOS................................................................................................................. 56

16
1. INTRODUÇÃO

17
1.1 Sistema renina angiotensina-aldosterona
O sistema renina-angiotensina-aldosterona (SRAA) é um dos mais importantes
sistemas hormonais envolvidos na regulação da pressão arterial e no controle do
equilíbrio hidroeletrolítico [1].
Os principais elementos da cascata incluem o angiotensinogênio, a renina, a
angiotensina I (Ang I), a enzima conversora de angiotensina (ECA) e a angiotensina II
(Ang II). A cascata começa com a renina, agindo sobre o angiotensinogênio para
produzir a Ang I, a qual por sua vez, é convertida em Ang II pela ação da ECA. Ang II
é o principal peptídeo efetor do SRAA, promovendo uma variedade de ações
fisiológicas e fisiopatológicas. A maioria dos efeitos da Ang II deve-se à sua ligação
aos receptores tipo 1 (AT1-R) e tipo 2 (AT2-R) amplamente distribuídos pelo
organismo [2]. (Figura 1).
Figura 1: Sistema renina-angiotensina-aldosterona. O angiotensinogênio serve de
substrato para a renina dando origem à angiotensina I, que por sua vez, é convertida em
angiotensina II pela ação da ECA. Angiotensina II atua em receptores específicos.

18
A Ang II apresenta importante efeito vasoconstritor pela síntese da aldosterona,
retenção de sódio e água, desempenha importante papel no processo inflamatório e
na transcrição de fatores de crescimento responsáveis pela proliferação celular [3].
Desta forma, a alta concentração de Ang II desencadeia respostas inflamatórias,
principalmente porque é possível encontrar a ECA em muitos tecidos e células
do corpo, incluindo pulmão, intestino, rim, células endoteliais, monócitos, linfócitos e
também na forma circulante no plasma [4,5].
1.2 Aspectos gerais da resposta imunológica
O sistema imunológico é constituído por um conjunto de órgãos, células e
moléculas, e tem por finalidade manter a homeostase do organismo, combatendo as
agressões em geral. A imunidade inata celular é exercida por células que reconhecem
padrões moleculares associados aos patógenos (PAMP) usando receptores do tipo toll
(TLR) e constitui a primeira linha defesa do organismo frente a danos teciduais,
infeciosos ou traumáticos, envolvendo diferentes etapas. Embora a imunidade inata
constitua a primeira linha de defesa do hospedeiro, ela atua em conjunto com a
imunidade adaptativa e as principais células efetoras desse eixo da imunidade são: os
macrófagos, os neutrófilos e as células natural killer (NK). Em contraposição à
resposta inata, as células de imunidade adaptativa, representadas somente pelos
linfócitos B e T, reconhecem antígenos de forma específica e, sendo ativadas, elas
diferenciam em células efetoras e células de memória imunológica para o antígeno que
as ativou [6,7].
Considerando somente o receptor de reconhecimento específico para antígenos
dos linfócitos T, nos vertebrados superiores, existem duas subpopulações
fenotipicamente distintas de linfócitos T: os linfócitos T alfa:beta (T α:β) e os linfócitos
T gama:delta (T γ:δ). No homem, até 99% dos linfócitos T, são do fenótipo α:β. Além
disso, os linfócitos T α:β maduros dividem-se em duas subpopulações: linfócitos T α:β
que expressam o correceptor CD4+ (T α:β CD4+) e linfócitos T α:β que expressam o
correceptor CD8+ (T α:β CD8+). Os linfócitos T α:β CD4+, ao serem ativados pelo
antígeno, diferenciam em linfócitos T α:β CD4+ efetores, que secretam citocinas, e
linfócitos T α:β CD4+ de memória imunológica para o antígeno [6-8].
Os linfócitos T α:β CD4+ efetores, secretores de citocinas, são denominados
linfócitos T auxiliares ou T helper (Th), uma vez que eles colaboram com múltiplos

19
eventos imunológicos através das citocinas que produzem. Os linfócitos Th,
subdividem-se me linfócitos T auxiliares tipo 1 (Th1) e linfócitos T auxiliares tipo 2
(Th2). Essa divisão dos linfócitos Th em T1 e Th2 decorre do padrão diferente de
citocinas que eles produzem. Os Th1 produzem principalmente interleucina-2 (IL-2) e
interferon-gama (IFN-γ), enquanto os Th2 produzem preferencialmente IL-4, IL-5,
IL-6 e IL-13. Como as citocinas tem efeitos autócrinos, elas promovem inclusive a
expressão de novas moléculas na superfície celular, como por exemplo, a molécula
CD69, que é uma molécula que caracteriza a ativação de diversas subpopulações de
células [7-9].
No que tange aos linfócitos T α:β CD8+, ao serem ativados pelo antígeno,
diferenciam em linfócitos T α:β CD8+ efetores e T α:β CD8+ de memória imunológica
para o antígeno. Os linfócitos T α:β CD8+ efetores são denominados linfócitos T
citotóxicos (CTL), uma vez que eles eliminam células hospedeiras indesejadas, como
por exemplo, células hospedeiras infectadas por patógenos intracelulares [6,8].
Como mencionado anteriormente, a outra subpopulação de células da imunidade
adaptativa ou específica são os linfócitos B, que são as únicas células produtoras de
anticorpos (Ac) ou imunoglobulinas (Ig). Os Ac são glicoproteínas secretadas por
linfócitos B, inclusive em resposta a um estímulo antigênico. Existem cinco isotipos ou
classes de anticorpos (IgA, IgD, IgE, IgG e IgM), além do que algumas classes de Ac
se subdividem em subclasses, como por exemplo a IgG do camundongo que apresenta
as subclasses: IgG1, IgG2a, IgGb, IgG2c e IgG3. Os Ac são os únicos elementos da
imunidade humoral específica e medeiam a imunidade contra antígenos extracelulares.
Os linfócitos B também atuam como células apresentadoras de antígeno, nos quais os
peptídeos gerados pelo processamento do antígeno são expressos na membrana dos
linfócitos B ligados às moléculas do complexo maior de histocompatibilidade (MHC)
classe II, para apresentação aos linfócitos Th [6-8].
A ativação de linfócitos B por antígenos proteicos requer a ativação simultânea
de linfócitos T α:β CD4+ (Th) específicos para o mesmo antígeno e a colaboração entre
B e Th. No que tange à colaboração dos linfócitos Th com os linfócitos B, os Th, via
citocinas e moléculas coestimuladores, promovem a expansão clonal dos linfócitos T,
a diferenciação dos linfócitos B em linfócitos de memória imunológica para o antígeno
e a troca do isotipo e o aumento de afinidade dos Ac produzidos em resposta ao
antígeno.[8].

20
Os camundongos da linhagem C57 black 6 (C57Bl/6), usados no presente
trabalho, em resposta a antígenos proteicos, como por exemplo a albumina de ovo de
galinha (ovalbumina; OVA) em presença de adjuvantes à base de sais de alumínio,
produzem IgM, IgG1 e IgE. A IgM é o Ac preferencialmente produzido após
primeiro contato do animal com a ovalbumina, o que caracteriza a resposta humoral
primaria. No entanto, a produção preferencial da IgM é gradativamente substituída pela
secreção preferencial de IgG1 e IgE, quando a imunização do animal é feita com
ovalbumina em presença de adjuvantes à base de sais de alumínio [10].
1.3 Bloqueio do sistema renina angiotensina-aldosterona: inibição da
enzima de conversão da angiotensina.
O SRAA pode ser bloqueado em diferentes pontos da cascata e por elementos
endógenos e exógenos. No que tange ao bloqueio do SRAA por elementos exógenos,
existem, atualmente, agentes farmacológicos, tais como os inibidores da renina,
inibidores da ECA, antagonistas do receptor de Ang II do tipo 1 (AT1-R) e do tipo II
(AT2-R) e outras classes de drogas [11,12]. Como o nome sugere, os inibidores da ECA
inibem a conversão da Ang I em Ang II, que é o principal peptídeo bioativo do SRAA
[1, 11]. Os inibidores da ECA são amplamente prescritos para tratar a hipertensão
primária, além de serem também usados para o tratamento de doenças relacionadas com
o sistema cardiovascular e insuficiência renal, devido aos efeitos imunoreguladores
desses fármacos [13,14]. Entre os inibidores da ECA largamente prescritos para tratar
a hipertensão primária estão o captopril e enalapril. O captopril é um medicamento que
contém o grupo sulfidrila em sua molécula, enquanto o enalapril contém o grupo
carbonila em sua estrutura (Figura 2). Além das diferenças nas estruturas químicas
desses dois inibidores da ECA, o enalapril é uma pró-droga, o maleato de enalapril, que
no organismo é convertida à droga biologicamente ativa, o enalaprilato [15]. Além
disso, a farmacocinética destes dois inibidores da ECA é diferente, o captopril tem
início de ação rápida com curta duração de ação, enquanto enalapril tem início mais
lento e, relativamente, longa duração de ação [16].

21
A) B)
Figura 2: Estrutura química do captopril (A) e do enalapril (B)
1.4 Efeitos dos inibidores da ECA no sistema imunológico
Os inibidores da ECA regulam uma variedade de funções imunes, incluindo
ações não-hemodinâmicas como a síntese de citocinas, produção de anticorpos e a
resposta celular. Nesse sentido, foi mostrado que o captopril reduz a produção do fator
de necrose tumoral do tipo alfa (TNF-alfa) na artrite reumatoide experimental [17].
Além disso, uma ampla quantidade de estudos clínicos e experimentais tem mostrado
que os inibidores da ECA, captopril, enalapril e lisinopril tem propriedades
pleiotrópicas, não-hemodinâmicas sobre a resposta de células T através da indução da
síntese de citocinas [18]. Nesse contexto, foi mostrado anteriormente que captopril
inibe a produção da IL-4 e da IL-10 sem alterar a síntese da IL-2, da IL-5 e do IFN-γ
[19]. Corroborando esses dados, foi mostrado recentemente que captopril reduziu a
produção de TNF-α, IL-1α, IL-10, IL-12 e IL-18 por células dendríticas estimuladas
com lipopolissacarídeo (LPS) [20]. Em outro estudo, foi mostrado que enalapril
aumenta o número de células T CD4+CD103+CD25-negativas no baço de
camundongos BALB/c junto com o aumento da produção de IL-10 [21]. Além disso,
foi recentemente mostrado que enalapril induz a expansão de células e regularização
de macrófagos para o perfil 1 (M1) em rins de camundongos diabéticos [22].
Recentemente, foi relatado que os inibidores de ECA também são utilizados para o
tratamento de doenças neurodegenerativas, tais como doença de Alzheimer e doença
de Parkinson [23]. De longe, a maioria dos estudos sobre as propriedades imuno-
mediadas dos inibidores da ECA tem enfatizado seus efeitos sobre a produção de
citocinas. Pouca atenção, no entanto, tem sido voltada para possíveis efeitos dos

22
inibidores da ECA sobre a síntese de anticorpos e sobre as subpopulações de linfócitos
T CD4+, T CD8+, B e NK in vivo. Diante do exposto e, sobretudo para alargar a
nossa compreensão sobre os efeitos dos inibidores da ECA sobre a produção de
anticorpos, procurou-se verificar se o inibidor de ECA, enalapril, seria capaz de
interferir na resposta humoral específica em camundongos C57BL/6. O enalapril foi
escolhido como o inibidor da ECA porque ele regula a produção de citocinas e, tanto
quanto sabemos, não há dados na literatura sobre o efeito deste inibidor da ECA na
resposta humoral a antígenos estranhos em modelos pré-clínicos. No presente trabalho,
investigou-se o efeito de enalapril sobre a resposta humoral de camundongos C57BL/6
imunizados com ovalbumina EndoFit (OVA) na presença de Alhydrogel, como
adjuvante. Do mesmo modo, com o intuito de ampliar o conhecimento sobre o efeito
do captopril e do enalapril sobre a resposta imune celular, no presente trabalho avaliou-
se a expressão de marcadores fenotípicos e de ativação de populações de linfócitos
esplênicos de camundongo C57BL/6 após o tratamento crônico com captopril ou
enalapril.

23
2. OBJETIVOS

24
2.1 Objetivo geral
O objetivo geral deste trabalho foi investigar o efeito do captopril e do
enalapril sobre a resposta imunológica humoral e celular in vivo.
2.2 Objetivos específicos
- Avaliar, por ELISA, a produção de IgG1 e IgG2c em camundongos
imunizados com ovalbumina EndoFit (OVA) na presença de Alhydrogel,
como adjuvante.
- Avaliar, por citometria de fluxo, a frequência de linfócitos T (CD4+ e
CD8+), de linfócitos B (CD19+) e de NK (CD49b+) após o tratamento
crônico com captopril ou enalapril;
- Avaliar, por citometria de fluxo, o número absoluto de linfócitos T
(CD4+ e CD8+), de linfócitos B (CD19+) e de NK (CD49b+) in vivo após
o tratamento de longa duração com captopril ou enalapril;
- Verificar se o tratamento crônico com captopril ou enalapril promove
ativação de linfócitos T (CD4+ e CD8+), de linfócitos B (CD19+) e de NK
(CD49b+) in vivo.

25
3. MATERIAL E MÉTODOS

26
3.1 Animais
Os camundongos, isogênicos da linhagem C57BL/6, usados nesse experimento
estavam com oito semanas de idade no início do experimento e foram obtidos do Centro
Multidisciplinar para Investigação Biológica na Área da Ciência em Animais de
Laboratório (CEMIB) da Universidade Estadual de Campinas (UNICAMP), Campinas,
SP. Durante o experimento, os animais foram mantidos em gaiolas de polipropileno
com acesso livre a água e ração autoclavadas, com ciclos alternados de claro/escuro
de
12 horas e temperatura a 25°C. A manipulação dos animais foi feita de acordo com as
regras do uso de animais em pesquisa e com os princípios da ética da experimentação
animal (#23108.039341/12-4 ANEXO 1). Para a avaliação da resposta imune humoral,
os camundongos foram divididos aleatoriamente em dois grupos com quatro animais
por grupo, como segue:
• Grupo 1: camundongos imunizados com ovalbumina tratados com enalapril.
• Grupo 2: camundongos imunizados com ovalbumina não tratados.
Da mesma forma, para a avaliação da imune celular, os camundongos foram
divididos aleatoriamente em três grupos com seis a nove animais por grupo, como segue:
• Grupo 1: camundongos não tratados, que serviram de controle.
• Grupo 2: camundongos tratados cronicamente com captopril.
• Grupo 3: camundongos tratados cronicamente com enalapril.
3.2 ELISA
3.2.1 Medicamento e Tratamento
Os animais imunizados com OVA foram tratados com enalapril ou foram
deixados sem tratamento durante quatro semanas. O enalapril (Hexal Laboratoty, São
Paulo, SP, Brasil), obtido comercialmente, foi dissolvido em água potável autoclavada
e oferecido ad libitum, na concentração de 0,02 mg/mL para o enalapril e substituído
a cada 24 h. A dosagem diária foi de 5 mg/kg peso corporal para maleato de enalapril,
assumindo um consumo diário de 5 ml por animal. A dose do enalapril foi selecionada
para se ter um nível normal de pressão sistólica e está dentro da média terapêutica usada
em camundongos [16,24

27
3.2.2 Antígeno e adjuvante
O antígeno utilizado para a imunização dos camundongos foi a ovalbumina (OVA;
EndoFit) e como adjuvante foi usado gel de hidróxido de alumínio (Alhydrogel a 2%;
Aluminium hydroxide gel) ambos da Invivogen (San Diego, CA, USA).
3.2.3 Imunização
Os camundongos foram imunizados por via subcutânea com ovalbumina (OVA, 10 µg)
na presença de Alhydrogel (1 mg). No 21º dia após a imunização inicial, os animais que
haviam sido imunizados receberam uma dose de reforço nas mesmas condições da
imunização primária.
3.2.4 Coleta de sangue
Amostras de sangue foram coletadas por punção venosa do plexo retro-orbital
de cada animal individual com o auxílio de pipetas Pasteur, no dia zero, ou seja,
imediatamente antes da primeira imunização e nos dias 7, 14, 21 e 28. O sangue
coletado foi colocado em tubos cônicos e deixado em repouso por 2 h à temperatura
ambiente para a retração do coágulo. Em seguida, as amostras de sangue foram
centrifugadas a 2.000g por 20 min e os soros obtidos foram acondicionados a -20°C
para análise posterior da presença de anticorpos específicos para OVA.
3.2.5 Ensaio para determinação de IgG1 e IgG2c anti-OVA
Os soros obtidos dos animais imunizados com OVA e tratados ou não com
enalapril, foram analisados pelo Enzyme Linked Immunosorbent Assay (ELISA), a
fim de se pesquisar a presença de IgG1 e IgG2c anti-OVA. Resumidamente: As reações
foram realizadas em microplacas de poliestireno de 96 poços de fundo plano
(MAxsorp). As placas foram sensibilizadas com OVA a 10 µg/mLe incubadas durante
a noite a 4°C. Após a sensibilização, descartou-se o excesso de antígeno, lavou-se cada
placa 3 vezes com salina tamponada com fosfato (PBS), pH 7.2, contendo Tween-20
(Sigma) a 0,05% (PBS-Tween) e as ligações inespecíficas foram bloqueadas com

28
albumina sérica bovina (BSA, Sigma) a 1% em PBS durante 1h, à temperatura
ambiente. Em seguida, o excesso de solução bloqueadora foi descartado, os soros
diluídos a 1/100 foram adicionados a cada poço e as placas foram incubadas
novamente durante a noite a 4°C. Removeu-se o excesso dos soros, lavando as placas
5 vezes com PBS-Tween, adicionou-se 50 µL por poço de anticorpo monoclonal anti-
IgG1 ou anti-IgG2c (Zymed®) marcados com peroxidase, na diluição de 1/4000 e as
placas foram incubas por 1 h à temperatura ambiente. O excesso de anticorpo-enzima
foi descartado lavando a placa 6 vezes com PBS-Tween e a reação antígeno-anticorpo
foi revelada adicionando água oxigenada (H2O2) com o cromógeno ortho-
phenyldiamine (OPD; Sigma Aldrich ® USA). As placas foram incubadas por 20 min
à temperatura ambiente no escuro e a reação enzima-substrato foi interrompida
adicionando 50 µL de H2SO4 2N. As reações foram determinadas em leitor de
microplacas de ELISA (Thermo Plate-Reader, NM, modelo X) a 492 nm e os
resultados foram expressos como a média da densidade ótica (DO) ± DP. A
concentração de IgG2c foi determinada por extrapolação a partir da curva padrão.
O nível mínimo de detecção para IgG2c é 0,39ng/ mL.
3.2.6 Análise estatística
A análise estatística foi feita usando o software Graph-Pad Prism. Para comparar os dados
entre os grupos foi utilizada a análise de variância de uma via (ANOVA), seguida do
teste Tukey. As diferenças foram consideradas significantes com p<0,05.
3.3 CITOMETRIA DE FLUXO
3.3.1 Medicamentos e tratamento
O captopril (Captotec®) e o enalapril (Enaprotec®) foram obtidos comercialmente
(Hexal Laboratoty, São Paulo, SP, Brasil). O captopril e o enalapril foram dissolvidos em
água potável autoclavada, na concentração de 0,2 mg/mL para o captopril e 0,02 mg/mL
para o enalapril. Os medicamentos dissolvidos em água potável foram oferecidos ad
libitum e a dosagem diária foi de 30 mg/kg de peso corporal para o captopril e de 5
mg/kg para enalapril, assumindo uma ingestão diária de 5 mL de água por animal. A
dose do captopril foi previamente estabelecida para manter a pressão arterial normal

29
[19]. Já a dose do ena lapr i l e s t á dentro das margens das doses normalmente
usadas em experimentos com camundongos [16]. Além disso, a administração desses
medicamentos por via oral e oferecidos ad libitum, está descrita na literatura pertinente
[19,21]. Os medicamentos dissolvidos em água potável autoclavada foram repostos a
cada 24 horas. O tratamento foi realizado durante três meses ininterruptos para a avalição
da imunidade celular.
3.3.2 Obtenção de células esplênicas
Três meses após o início do experimento, os animais foram sacrificados,
de forma assistida por uma médica veterinária (CRMV-MT 1698). Imediatamente após
o sacrifício assistido, o baço de cada camundongo foi removido em condições
assépticas e macerado individualmente em meio incompleto para cultura de células
(RPMI 1640, Sigma Chemical Co. St. Louis, EUA) para a obtenção das células
esplênicas. As células esplênicas assim obtidas foram colocadas em tubos cônicos
esterilizados e foram lavadas duas vezes por centrifugação com staining buffer (solução
salina contendo soro bovino fetal e 0.09% de azida sódica) (eBioscience, San Diego,
Califórnia, EUA). Após as lavagens, o sobrenadante foi descartado e as células foram
ressuspensas em 1 mL de salina tamponada com fosfato (PBS). Em seguida, uma
alíquota de células foi diluída em solução de Turk a 1:40 e foram contadas em câmara
de Neubauer, usando microscópio de luz com objetiva de 40x. Simultaneamente foi
feita a verificação da viabilidade celular pela exclusão com azul de Trypan.
3.3.3 Imunofenotipagem celular
A marcação das subpopulações de linfócitos com anticorpos monoclonais
conjugados com fluorcromos foi feita de acordo com os protocolos do fabricante
(eBioscience). Resumidamente: Suspenssão de células contendo 2x106células/ml
foram preparadas em aliquotas de 100 ml de staining buffer por tubo e incubadas
por 20 minutos com anticorpo monoclonal anti-CD16/CD32 para bloquear as ligações
inespecíficas para a fração cristalizável (Fc) do anticorpo. Após a incubação, as células
foram marcadas com anticorpos monoclonais específicos para as moléculas de
superfície CD4, CD8, CD19 e CD49b. Além desses marcadores fenotípicos, foram

30
também utilizados os anticorpos monoclonais específicos para moléculas de ativação
como o CD69 e as moléculas do complexo de histocompatibilidade pricinpal tipo
II (MHC II). Todos os anticorpos monoclonais conjugados com fluocromos usados
nesse trabalho foram obtidos da eBioscience, (San Diego, Califórnia, EUA) e estão
mostrados na tabela 1.
Tabela 1 : Anticorpos monoclonais conjugados com fluorocromos específicos para
o CD4, CD8, CD19 e CD49b, CD69 ou MHC de classe II.
Anticorpos monoclonais Fluorocromos
Anti-CD4 PerCP-Cy5.5
Anti-CD8 PerCP-Cy5.5
Anti-CD19 PE
Anti-CD49b PE
Anti-CD69 APC
Anti MHC classe II PerCP-eFluor710
PerCP-Cy5.5, proteína peridina-clorofila cianina 5.5; PE ficoeritrina; APC,
aloficocianina; PerCP- eFluor710, proteína peridina-clorofila eFluor 719nm
Como os anticorpos anti-CD4 e anti-CD8 eram ambos conjugados com a
proteína peridina-clorofila cianina 5.5 (PerCP-Cy5.5) e os anticorpos anti-CD19 e anti-
CD49b eram marcados com ficoeritriina (PE), as marcações das suspensões celulares
foram feitas em 4 tubos separados, como mostrado na tabela 2.
Tabela 2: Anticorpos monoclonais distribuídos em tubos separados para serem
analisados por citometria de fluxo
Tubos Células e anticorpos
1 Células esplênicas
Anti-CD4 PerCP-Cy5.5
Anti-CD69 - APC
2 Células esplênicas
Anti-CD8 PerCP-Cy5.5
Anti-CD69 - APC
3 Células esplênicas

31
Anti-CD19 - PE
Anti-MHC classe II - PerCP-eFluor710
4 Células esplênicas
Anti-CD49b - PE
Anti-CD69 - APC
Após a marcação, as células foram incubadas a 4°C por 30 minutos, no escuro.
Em seguida, as suspensões celulares foram lavadas com PBS, para a remoção do excesso
de anticorpos, foram fixadas com formaldeído a 4%e analisadas no citômetro de fluxo
(BD Accuri™ C6, Becton Dickinson, San José, CA, EUA). As fluorescências emitidas
pelos fluorocromos conjugados aos anticorpos monoclonais foram lidas nos seguintes
canais de fluorescência (FL): Em FL2–PE, FL3-PerCP-Cy5.5 e PerCP-e Fluor710 e FL4
- APC.
3.3.4 Análise de imunofenotipagem
A partir de cada tubo foram adquiridos 20.000 eventos em uma gate de
linfócitos baseada na dispersão de luz frontal (forward scatter, FSC) e lateral (side
scatter, SSC). O forward scatter (FSC) relaciona se com o tamanho e o side scatter
(SSC) com a granularidade da célula ou partícula analisada. Na sequência, a população
celular de interesse, que se constituiu de linfócitos, foi selecionada dentro da região
P1(Figura 3).
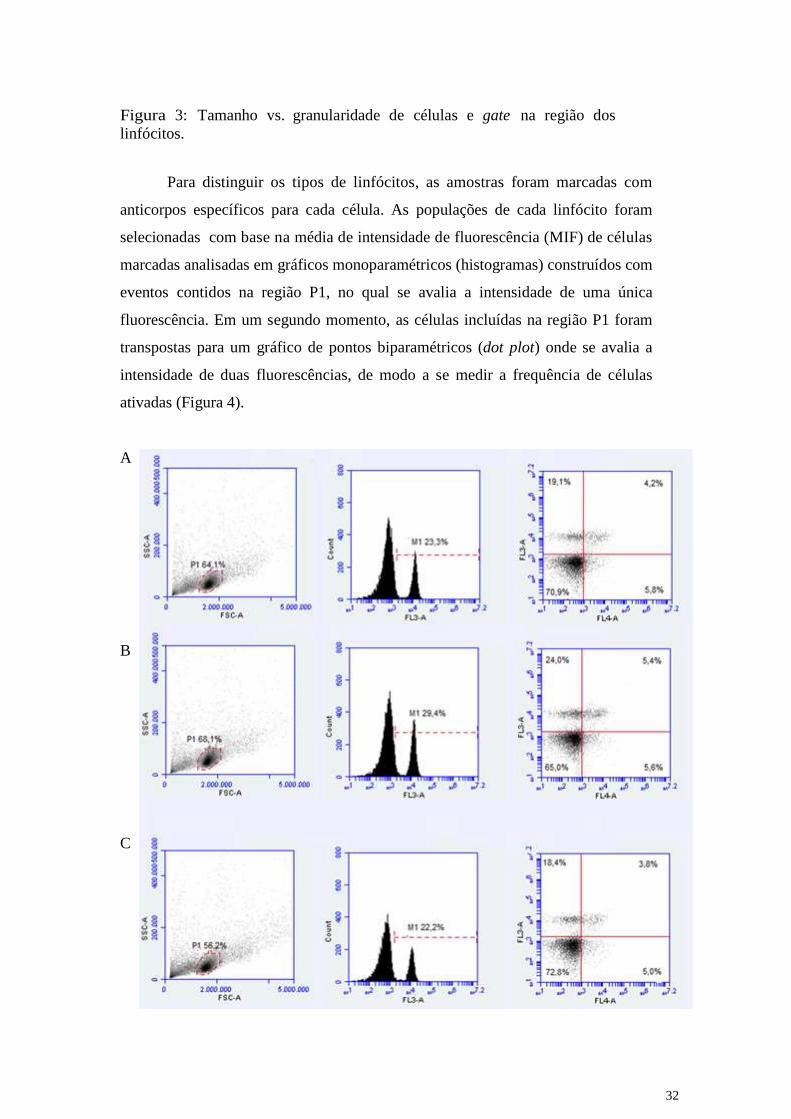
32
Figura 3: Tamanho vs. granularidade de células e gate na região dos
linfócitos.
Para distinguir os tipos de linfócitos, as amostras foram marcadas com
anticorpos específicos para cada célula. As populações de cada linfócito foram
selecionadas com base na média de intensidade de fluorescência (MIF) de células
marcadas analisadas em gráficos monoparamétricos (histogramas) construídos com
eventos contidos na região P1, no qual se avalia a intensidade de uma única
fluorescência. Em um segundo momento, as células incluídas na região P1 foram
transpostas para um gráfico de pontos biparamétricos (dot plot) onde se avalia a
intensidade de duas fluorescências, de modo a se medir a frequência de células
ativadas (Figura 4).
A
B
C
33
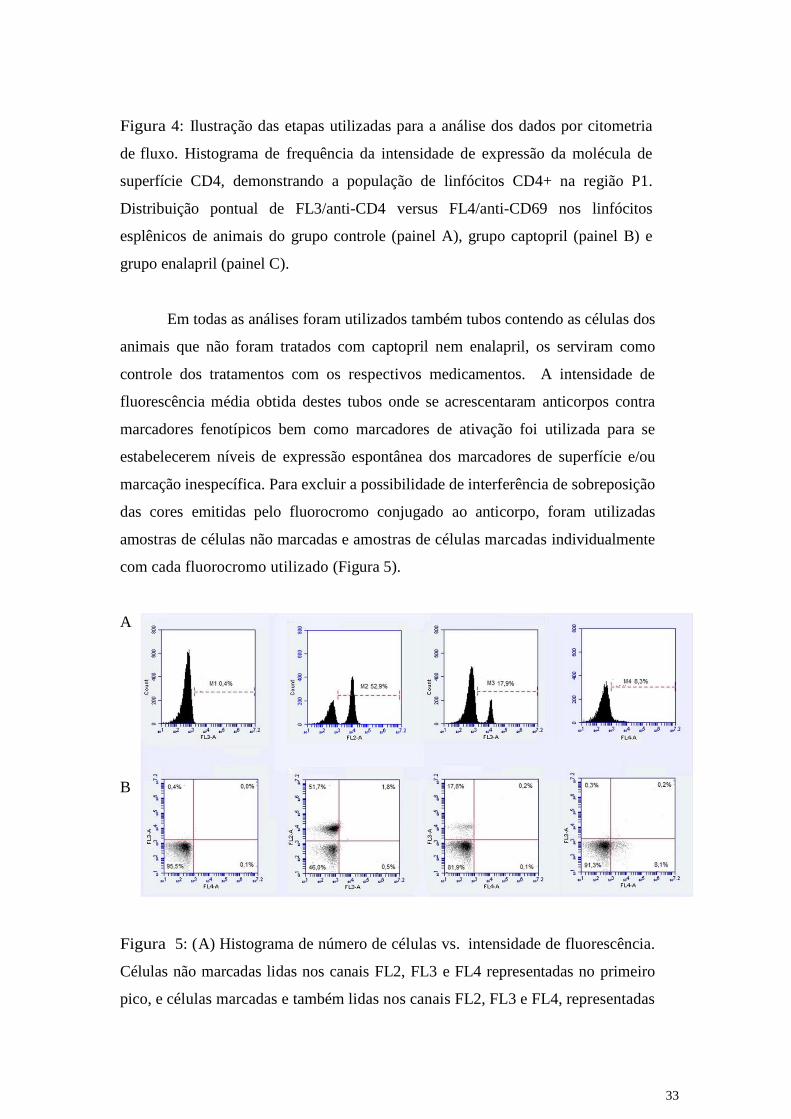
Figura 4: Ilustração das etapas utilizadas para a análise dos dados por citometria
de fluxo. Histograma de frequência da intensidade de expressão da molécula de
superfície CD4, demonstrando a população de linfócitos CD4+ na região P1.
Distribuição pontual de FL3/anti-CD4 versus FL4/anti-CD69 nos linfócitos
esplênicos de animais do grupo controle (painel A), grupo captopril (painel B) e
grupo enalapril (painel C).
Em todas as análises foram utilizados também tubos contendo as células dos
animais que não foram tratados com captopril nem enalapril, os serviram como
controle dos tratamentos com os respectivos medicamentos. A intensidade de
fluorescência média obtida destes tubos onde se acrescentaram anticorpos contra
marcadores fenotípicos bem como marcadores de ativação foi utilizada para se
estabelecerem níveis de expressão espontânea dos marcadores de superfície e/ou
marcação inespecífica. Para excluir a possibilidade de interferência de sobreposição
das cores emitidas pelo fluorocromo conjugado ao anticorpo, foram utilizadas
amostras de células não marcadas e amostras de células marcadas individualmente
com cada fluorocromo utilizado (Figura 5).
A
B
Figura 5: (A) Histograma de número de células vs. intensidade de fluorescência.
Células não marcadas lidas nos canais FL2, FL3 e FL4 representadas no primeiro
pico, e células marcadas e também lidas nos canais FL2, FL3 e FL4, representadas

34
no segundo pico. (B) Dot plots a partir dos quais se faz o cálculo da intensidade
de fluorescência dos fluorocromos referentes às células marcadas ou não.
3.3.5 Análise estatística
Os dados obtidos foram submetidos à análise de variância por One-way analysis
of variance (ANOVA) seguida pelo teste de Bonferroni, utilizado para comparação
entre os grupos. Foram consideradas diferenças significativas quando o valor de P foi
menor ou igual a 0,05 (P ≤ 0,05). As análises estatísticas foram realizadas usando o
software Statistical Analysis System (SAS, Institute Inc., 2002-2008, Cary, Carolina
do Norte, EUA) versão 9.3. A análise estatística foi realizada sob a supervisão e
orientação da Professora Édila Cristina de Souza do Departamento de Estatística da
Universidade Federal de Mato Grosso.

35
4. RESULTADOS

36
4.1 Avaliação da produção de IgG1 e IgG2c anti-OVA
Para a análise da produção de anticorpos específicos para ovalbumina, os
camundongos imunizados com OVA mais Alhydrogel (Al) foram tratados com
enalapril durante quatro semanas. Para fins comparativos, animais foram imunizados
com OVA mais Al e não foram tratados com enalapril. Com relação aos isotipos, os
dados apresentados na figura 6 mostram que a cinética da produção IgG1 anti-OVA
apresentou um perfil clássico, ou seja, os níveis séricos de IgG1 aumentam com
o tempo e após o reforço, demonstrando que esses resultados não foram afetados pelo
tratamento com enalapril. Deve notar-se que os animais imunizados com ovalbumina
EndoFit mais Alhydrogel e tratados ou não com o enalapril não exibiram IgG1
específica para OVA, nos dias 7 e 14 após a imunização, considerando-se que a
absorbância obtida a partir dos soros analisados foi de aproximadamente 0,1 (Figura
6). A ausência de IgG1 anti-OVA no soro na resposta primária inicial pode ser
explicada pela falta de lipopolissacarídeo (LPS) na ovalbumina EndoFit, visto que a
ligação do LPS aos TLR4 em linfócitos B parece ser necessária para a completa
resposta de anticorpos contra antígenos T-dependentes [25].
Figura 6: Quantificação de IgG1 sérica anti-OVA. Dois grupos de animais (3-4 / grupo)
foram imunizados com OVA mais Al e receberam uma dose de reforço no dia 21. Os
animais foram tratados oralmente com o enalapril ou foram deixados sem tratamento. Os
anticorpos IgG1 específicos para OVA foram determinados por ELISA usando soro
37
diluído a 1:400. Os resultados são expressos em ng/mL e representam dois experimentos
independentes.
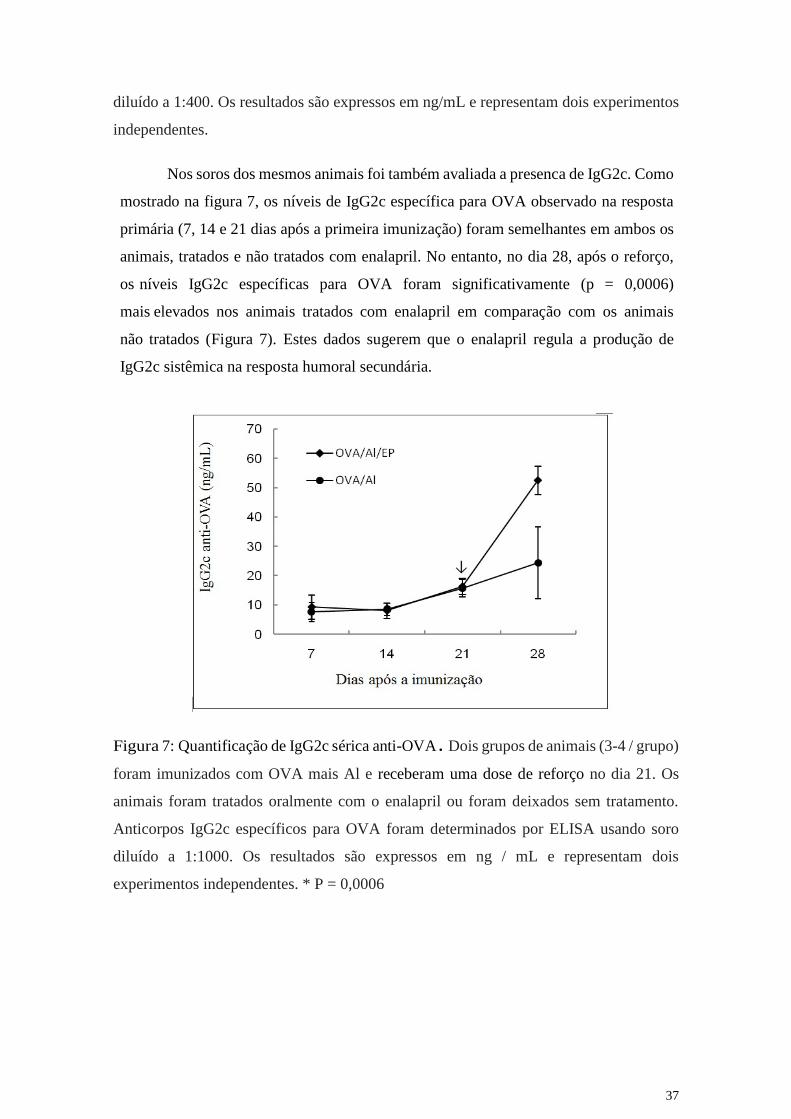
Nos soros dos mesmos animais foi também avaliada a presenca de IgG2c. Como
mostrado na figura 7, os níveis de IgG2c específica para OVA observado na resposta
primária (7, 14 e 21 dias após a primeira imunização) foram semelhantes em ambos os
animais, tratados e não tratados com enalapril. No entanto, no dia 28, após o reforço,
os níveis IgG2c específicas para OVA foram significativamente (p = 0,0006)
mais elevados nos animais tratados com enalapril em comparação com os animais
não tratados (Figura 7). Estes dados sugerem que o enalapril regula a produção de
IgG2c sistêmica na resposta humoral secundária.
Figura 7: Quantificação de IgG2c sérica anti-OVA. Dois grupos de animais (3-4 / grupo)
foram imunizados com OVA mais Al e receberam uma dose de reforço no dia 21. Os
animais foram tratados oralmente com o enalapril ou foram deixados sem tratamento.
Anticorpos IgG2c específicos para OVA foram determinados por ELISA usando soro
diluído a 1:1000. Os resultados são expressos em ng / mL e representam dois
experimentos independentes. * P = 0,0006
38
4.2 Avaliação da frequência de linfócitos T CD4+ in vivo
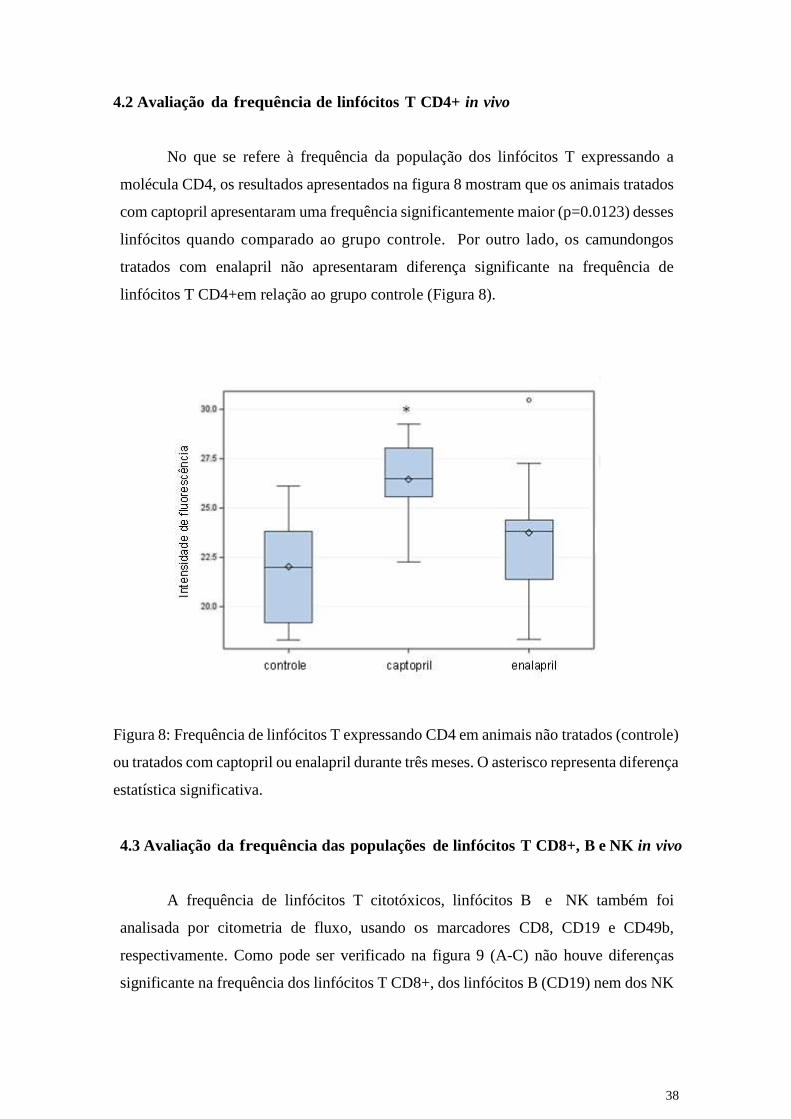
No que se refere à frequência da população dos linfócitos T expressando a
molécula CD4, os resultados apresentados na figura 8 mostram que os animais tratados
com captopril apresentaram uma frequência significantemente maior (p=0.0123) desses
linfócitos quando comparado ao grupo controle. Por outro lado, os camundongos
tratados com enalapril não apresentaram diferença significante na frequência de
linfócitos T CD4+em relação ao grupo controle (Figura 8).
Figura 8: Frequência de linfócitos T expressando CD4 em animais não tratados (controle)
ou tratados com captopril ou enalapril durante três meses. O asterisco representa diferença
estatística significativa.
4.3 Avaliação da frequência das populações de linfócitos T CD8+, B e NK in vivo
A frequência de linfócitos T citotóxicos, linfócitos B e NK também foi
analisada por citometria de fluxo, usando os marcadores CD8, CD19 e CD49b,
respectivamente. Como pode ser verificado na figura 9 (A-C) não houve diferenças
significante na frequência dos linfócitos T CD8+, dos linfócitos B (CD19) nem dos NK
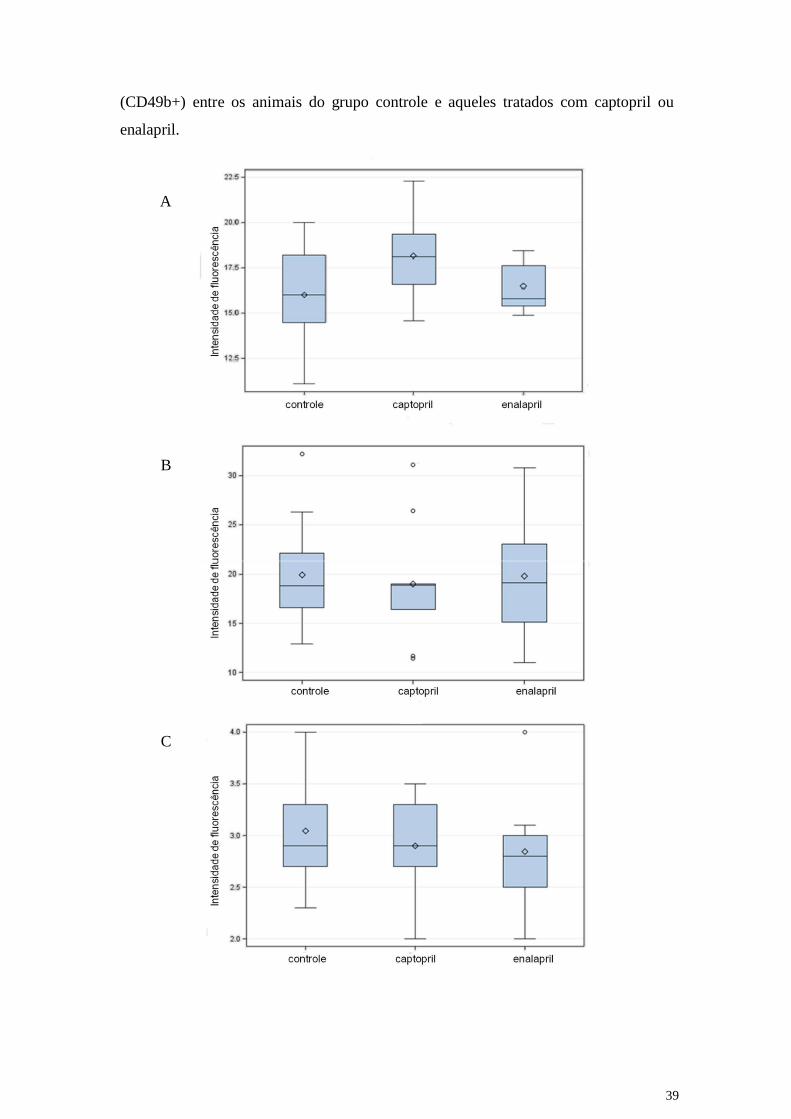
39
(CD49b+) entre os animais do grupo controle e aqueles tratados com captopril ou
enalapril.
A
B
C
40
Figura 9: Frequência dos linfócitos TCD8+ (painel A), dos linfócitos B CD19+ (painel
B) e dos NK CD49b+ (painel C) em animais não tratados (controle) ou tratados com
captopril ou enalapril.
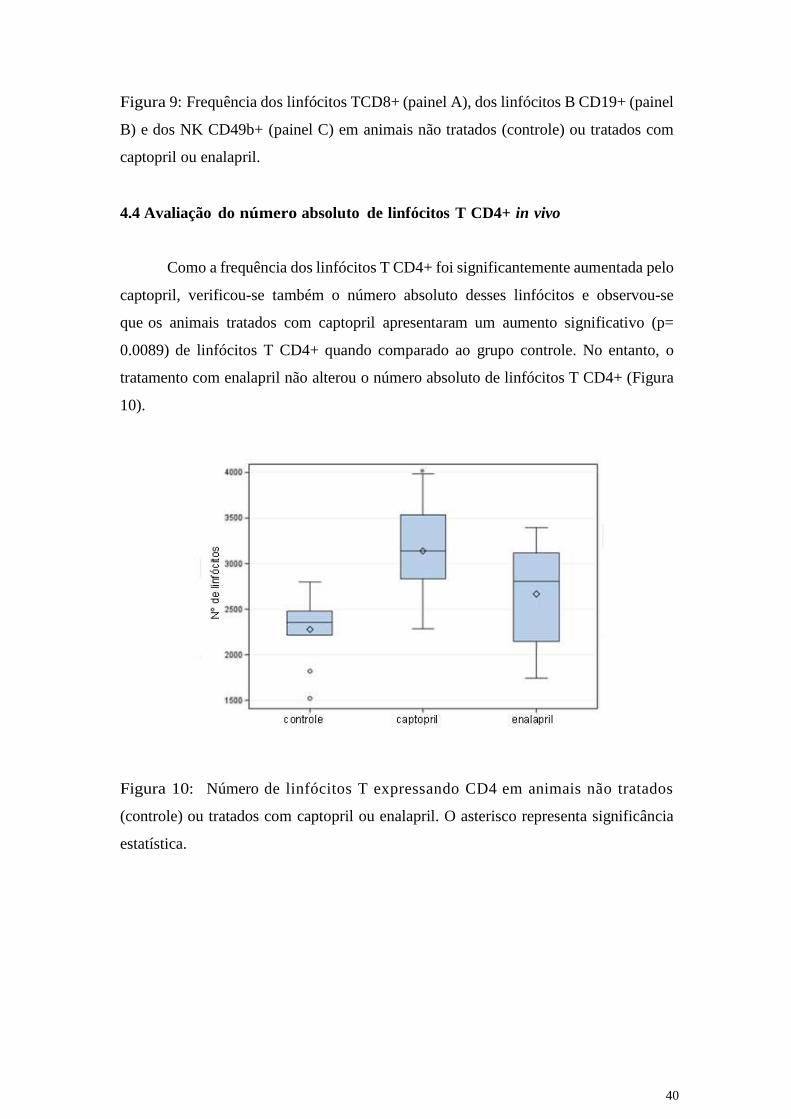
4.4 Avaliação do número absoluto de linfócitos T CD4+ in vivo
Como a frequência dos linfócitos T CD4+ foi significantemente aumentada pelo
captopril, verificou-se também o número absoluto desses linfócitos e observou-se
que os animais tratados com captopril apresentaram um aumento significativo (p=
0.0089) de linfócitos T CD4+ quando comparado ao grupo controle. No entanto, o
tratamento com enalapril não alterou o número absoluto de linfócitos T CD4+ (Figura
10).
Figura 10: Número de linfócitos T expressando CD4 em animais não tratados
(controle) ou tratados com captopril ou enalapril. O asterisco representa significância
estatística.
41
4.5 Avaliação do número absoluto das populações de linfócitos T CD8+, B e
NK in vivo
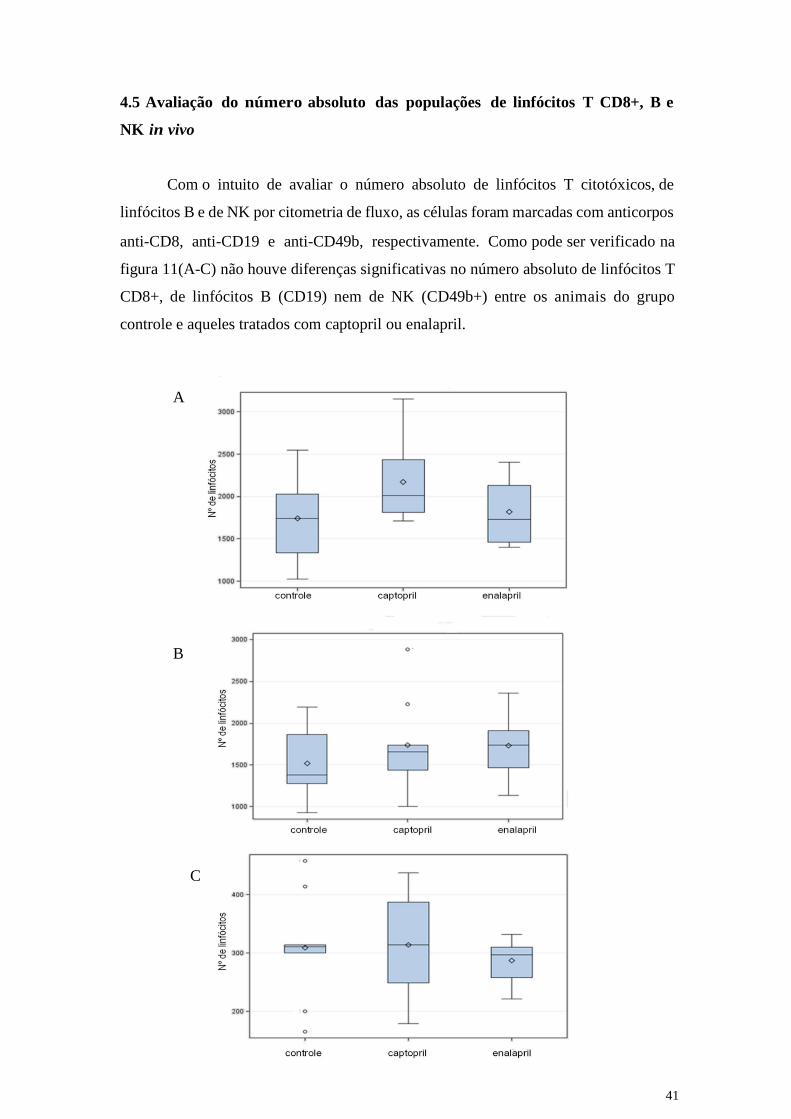
Com o intuito de avaliar o número absoluto de linfócitos T citotóxicos, de
linfócitos B e de NK por citometria de fluxo, as células foram marcadas com anticorpos
anti-CD8, anti-CD19 e anti-CD49b, respectivamente. Como pode ser verificado na
figura 11(A-C) não houve diferenças significativas no número absoluto de linfócitos T
CD8+, de linfócitos B (CD19) nem de NK (CD49b+) entre os animais do grupo
controle e aqueles tratados com captopril ou enalapril.
A
B
C
42
Figura 11: Número absoluto dos linfócitos T CD8+ (painel A), dos linfócitos B CD19+
(painel B) e dos NK CD49b+ (painel C) em animais não tratados (controle) ou tratados
com captopril ou enalapril.
4.6 Avaliação da frequência de linfócitos TCD4 ativada in
vivo
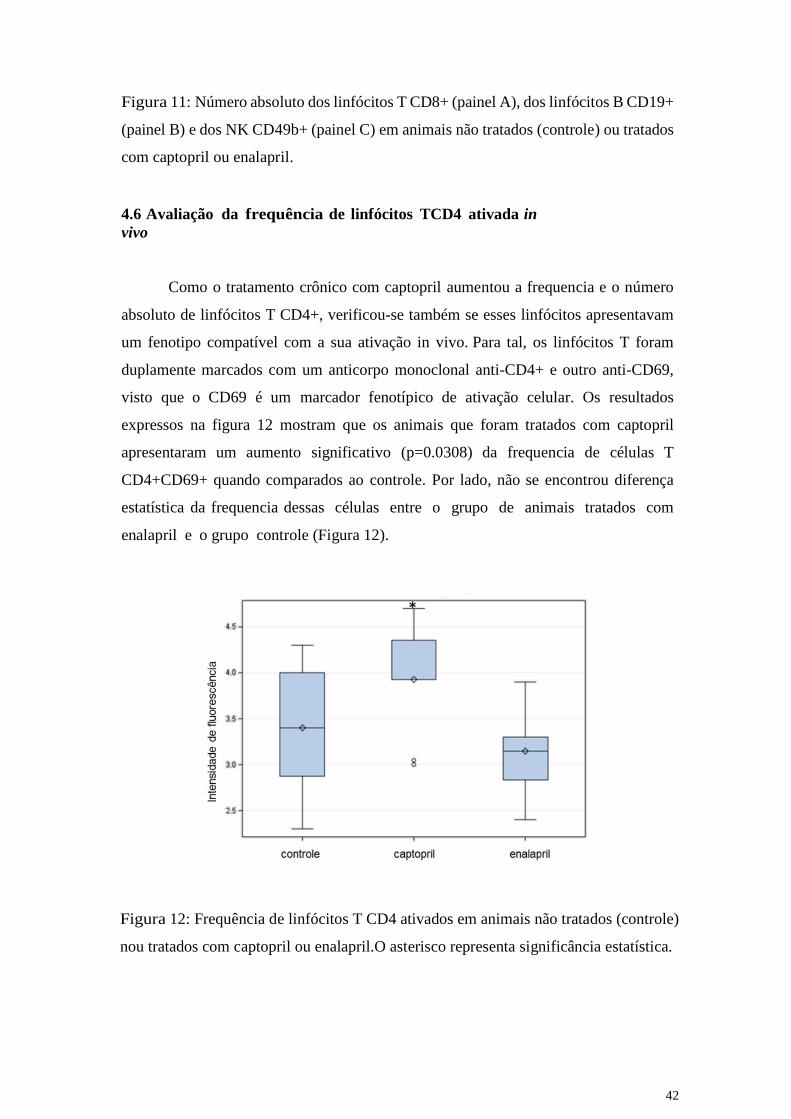
Como o tratamento crônico com captopril aumentou a frequencia e o número
absoluto de linfócitos T CD4+, verificou-se também se esses linfócitos apresentavam
um fenotipo compatível com a sua ativação in vivo. Para tal, os linfócitos T foram
duplamente marcados com um anticorpo monoclonal anti-CD4+ e outro anti-CD69,
visto que o CD69 é um marcador fenotípico de ativação celular. Os resultados
expressos na figura 12 mostram que os animais que foram tratados com captopril
apresentaram um aumento significativo (p=0.0308) da frequencia de células T
CD4+CD69+ quando comparados ao controle. Por lado, não se encontrou diferença
estatística da frequencia dessas células entre o grupo de animais tratados com
enalapril e o grupo controle (Figura 12).
Figura 12: Frequência de linfócitos T CD4 ativados em animais não tratados (controle)
nou tratados com captopril ou enalapril.O asterisco representa significância estatística.

43
4.7 Avaliação da frequência das populações de linfócitos T CD8+, B e NK
ativados in vivo
Ao contrário do que foi observado para os linfócitos TCD4+, não foram
encontradas mudanças significativas na frequência de linfócitos TCD8+, B e NK
ativados nos grupos dos animais tratados com captopril ou enalapril quando comparado
com os animais controle (Figura 13).
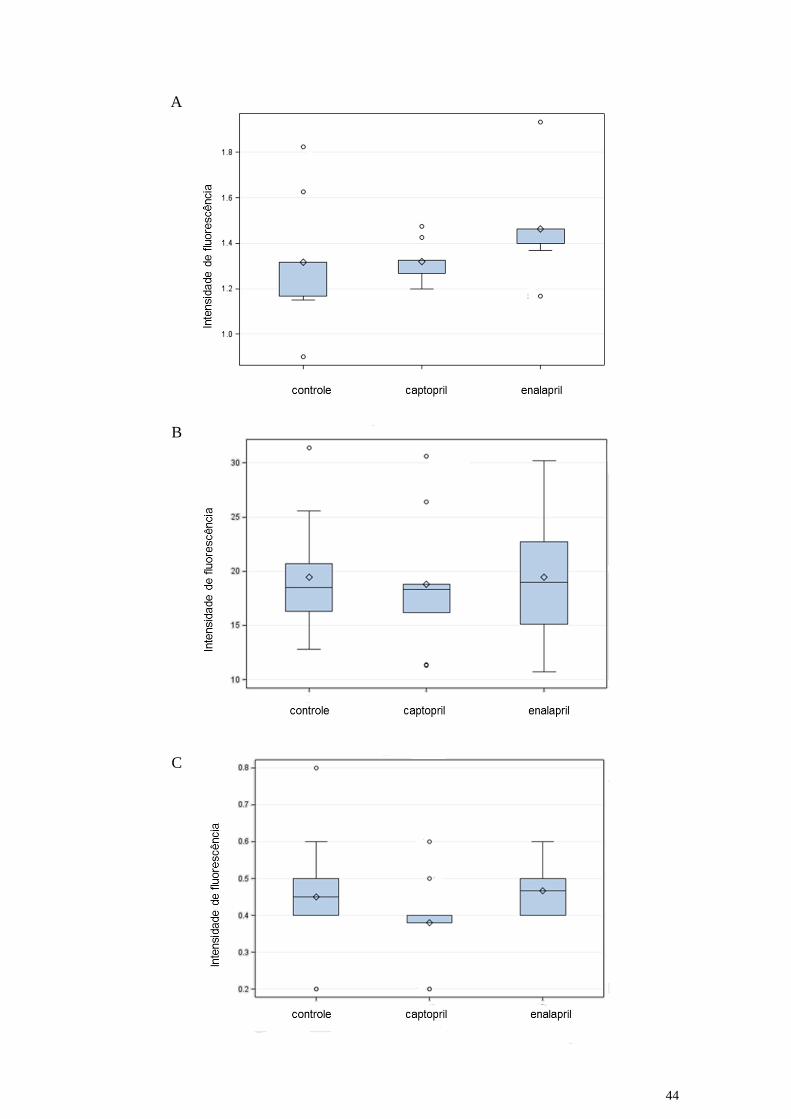
44
A
B
C

45
Figura 13: Frequência dos linfócitos T CD8+ (painel A), dos linfócitos B CD19+
(painel B) e dos NK CD49b+ (painel C) ativados em animais não tratados (controle) ou
tratados com captopril ou enalapril.

46
5. DISCUSSÃO

47
Nos experimentos apresentados, no que concerne à reposta imune humoral, os
dados mostram que a produção de IgG1 anti-OVA não foi alterada pelo tratamento com
enalapril. Esses resultados, mostrando que enalapril não altera a produção de IgG1 anti-
OVA no modelo murino, estão de acordo com os dados da literatura mostrando que o
tratamento de camundongos com captopril também não afeta a produção de IgG1 anti-
OVA em animais sensibilizados com esse antígeno mais o adjuvante incompleto de
Freund [26]. Contudo, no presente experimento, observou-se um aumento
significante dos níveis séricos de IgG2c anti-OVA na resposta secundária (7 dias após
o reforço) (Figura 7). Até onde se sabe, os resultados sobre o aumento da produção de
IgG2c promovido por enalapril são novos e suportam um possível papel dos inibidores
da ECA em resposta humoral contra antígenos estranhos em camundongos. Como a
mudança de isotipo para IgG2c é, em grande parte, dependente de sinais de IFN-γ
[27,28], pode-se especular que o aumento da produção IgG2c por enalapril seja devido
ao aumento de IFN-γ na circulação. Essa é uma hipótese plausível, que deve ser
avaliada em experimentos futuros.
A hipótese racional para se estudar o efeito de inibidores da ECA sobre resposta
humoral no modelo pré-clínico baseia-se no fato do enalapril e captopril estarem
incluídos na lista de medicamentos implicados no lúpus induzido por drogas [29]. Como
os inibidores da ECA são amplamente utilizados para o tratamento de pacientes
hipertensos e a hipertensão primária é uma condição durável, pode-se postular que esses
medicamentos usados por longo tempo por pacientes hipertensos deslocariam a resposta
Th do paciente para o padrão Th1, que poderia favorecer o desenvolvimento de doenças
de auto-imunidade regidas pela resposta Th1.
Concluindo, os resultados sugerem que a inibição sistêmica da angiotensina
II por enalapril estimula a produção IgG2c no modelo pré-clínico. Esses resultados têm
implicações importantes para a compreensão da regulação da resposta humoral por
inibidores da ECA, principalmente a abertura de uma nova perspectiva de pesquisa
sobre o papel destes medicamentos na resposta humoral a antígenos T–dependentes.
No que diz respeito à resposta imune celular, nesse estudo foi avaliado o efeito
do captopril e do enalapril sobre as subpopulações de linfócitos T CD4+ e CD8+, B
e NK em camundongos C57BL/6. Os resultados mostram que o captopril aumenta a
frequência, o número absoluto e promove a ativação de linfócitos T CD4+. No entanto,
sob as mesmas condições experimentais, o captopril não alterou a frequência, nem o

48
número absoluto e nem promoveu a ativação de linfócitos T CD8+, B e NK. Os dados
mostrando que o captopril aumenta a frequência, o número absoluto e promove
ativação de linfócitos T CD4+ corroboram com dados da literatura mostrando que
enalapril, outro inibidor da ECA, aumenta o número de células T CD4+CD103+CD25-
negativas no baço de camundongos BALB/c [21]. Estudos adicionais são necessários
para avaliar se o perfil genético da linhagem de camundongos usados e o tempo de
tratamento com enalapr i l ou captopril interferem com o efeito imunoregulador desses
medicamentos in vivo. Embora o captopril e o enalapril sejam largamente estudados
por seus efeitos benéficos na diabetes mellitus tipo 2 [30], na doença de Chagas [31],
na aterosclerose [32] e em alguns tipos de câncer [33], já foi também mostrado que
pacientes tratados com captopril desenvolvem IgM anti-DNA de fita dupla [34] O
captopril está, inclusive, elencado como uma droga indutora de lúpus [35]. Embora
nesse estudo, o modelo experimental seja saudável, pode postular que a ativação de
linfócitos T CD4+ pode ser responsável por síndromes imune-mediatas causadas pelo
captopril, visto que a indução de síndromes similar ao lúpus eritematoso sistêmico
requer a ativação de linfócitos T. Embora o captopril tenha ativado linfócitos T CD4+,
ressalte-se que aparentemente, o captopril não altera as subpopulações de linfócitos T
CD8+, linfócitos B nem NK, sugerindo assim um efeito preferencial sobre linfócitos T
CD4+. Estudos posteriores são requeridos para esclarecer o mecanismo pelo qual o
captopril age somente sobre linfócitos T CD4+. De modo contrário ao que foi visto para
o captopril, o enalapril, que é também um inibidor da ECA, não promoveu alterações
nas populações de linfócitos T, B nem NK. Uma hipótese plausível para os efeitos
diferentes desses dois inibidores da ECA sobre os linfócitos é a diferença estrutural
desses medicamentos, uma vez que o grupo sulfidrila que está presente no captopril está
ausente no enalapril.
Em conjunto, os dados mostram que o captopril e enalapril podem influenciar
na reposta imunológica seja por um efeito na produção de anticorpos ou na expressão
de moléculas envolvidas na ativação celular.

49
6. CONCLUSÕES

50
A partir dos resultados conclui-se que:
Enalapril aumenta significantemente a produção de IgG2c
anti-OVA.
Captopril aumenta significantemente a frequência e o número absoluto de
linfócitos T CD4+ in vivo. Entretanto, as subpopulações linfócitos T CD8+, B
e NK não foram afetadas pelo tratamento com captopril.
A ativação celular, caracterizada pela expressão da molécula CD69, foi
observada de forma significante apenas nos linfócitos T CD4+ dos camundongos
submetidos ao tratamento com captopril.
O tratamento com enalapril não alterou os parâmetros estudados nas
subpopulações de linfócitos.

51
7. BIBLIOGRAFIA

52
1.Fyhrquist F, Saijonmaa O. Renin-angiotensin system revisited. J Intern Med. 2008;
264: 224–36.
2. Regulski M, Regulska K, Stanisz BJ, Murias M, Gieremek P, Wzgarda A, Niznik B.
Chemistry and pharmacology of Angiotensin-converting enzyme inhibitors. Curr
PharmDes. 2015; 21(13):1764-75.
3. Ferrario CM. Role of angiotensin II in cardiovascular disease therapeutic implications
of more than a century of research. J Renin Angiotensin Aldosterone Syst. 2006
Mar;7(1):3-14.
4. Costerousse O, Allegrini J, Lopez M, Alhenc-Gelas F. Angiotensin I-converting
enzyme in human circulating mononuclear cells: genetic polymorphism of expression in
T-lymphocytes. Biochem. J. (1993) 290, 33-40.
5. Sparks MA, Crowley SD, Gurley SD, Mirotsou M, Coffman TM. Classical Renin-
Angiotensin System in Kidney Physiology. Compr Physiol. 2014 July; 4(3): 1201–
1228.
6 . Abbas AK, Lichtman AH, Pillai S. Cellular and Molecular Immunology, 6ª ed,
Editora Saunders 2007.
7. Cruvinel WM, Mesquita Júnior D, Araújo JA, Cate lan TT, Souza AW, Silva NE,
Andrade LE. Sistema Imunitário – Parte I Fundamentos da imunidade inata com ênfase
nos mecanismos moleculares e celulares da resposta inflamatória Rev Bras Reumatol
2010;50(4):434-61.
8. Mesquita Júnior D, Araújo JA, Catelan TT, Souza AW, Cruvinel WM, Andrade LE.
Sistema imunitário - parte II: fundamentos da resposta imunológica mediada por
linfócitos T e B. Rev Bras Reumatol. 2010;50(5):552-80.
9. Ziegler SF, Ramsdell F, Alderson MR. The activation antigen CD69. Stem Cells.
1994 Sep;12(5):456-65.

53
10. Prouvost-Danon A, Binaghi R, Rochas S, Boussac-Aron Y. Immunochemical
identification of mouse IgE. Immunology 1972; 23: 481-491.
11. Robles NR, Cerezo I, Hernandez-Gallego R. Renin-angiotensin system blocking
drugs. J Cardiovasc Pharmacol Ther. 2014 Jan;19(1):14-33.
12. Te Riet L, van Esch JH, Roks AJ, Van den Meiracker AH, Danser
AH.HypertensionRenin–Angiotensin–Aldosterone System Alterations. Circ Res. 2015;
116(6):960-75.
13. Atlas SA. The Renin-Angiotensin Aldosterone System: Pathophysiological Role
andPharmacologic Inhibition. J Manag Care Pharm. 2007; 13(8 Suppl B):9-20.
14. Li EC, Heran BS, Wright JM. Angiotensin converting enzyme (ACE) inhibitors
versus angiotensin receptor blockers for primary hypertension. Cochrane Database Syst
Rev. 2014; 22;8: CD009096.
15. Davies RO, Gomez HJ, Irvin JD, Walker JF. An overview of the clinical
pharmacology of enalapril. Br J Clin Pharmacol. 1984; 18 (Suppl 2):215S-29S.
16. Raia JJ Jr, Barone JA, Byerly WG, Lacy CR. Angiotensin-converting enzyme
inhibitors: a comparative review. DICP. 1990; 24(5):506-25.
17. Liu HM, Wang KJ. Therapeutic effect of Captopril on rheumatoid arthritis in rats.
Asian Pac J Trop Med. 2014; 7(12):996-9.
18. Gullestad L, Aukrust P, Ueland T et al. Effect of high- versus low-dose angiotensin
converting enzyme inhibition on cytokine levels in chronic heart failure. J Am Coll
Cardiol 1999; 34:2061–67.
19. De Albuquerque DA, Saxena V, Adams DE, Boivin GP, Brunner HI, Witte DP, Singh
RR. An ACE inhibitor reduces Th2 cytokines and TGF-beta1 and TGF-beta2 isoforms in
murine lupus nephritis. Kidney Int. 2004; 65(3):846-59.

54
20. Lapteva N, Ide K, Nieda M et al. Activation and suppression of renin-angiotensin
system in human dendritic cells. Biochem Biophys Res Commun 2002; 296:194-200.
21. Albuquerque D, Nihei J, Cardillo F, Singh R. The ACE inhibitors enalapril and
captopril modulate cytokine responses in Balb/c and C57Bl/6 normal mice and
increaseCD4(+) CD103(+) CD25(negative) splenic T cell numbers. Cell Immunol. 2010;
260(2):92-7.
22. Cucak H, Nielsen Fink L, Hojgaard Pedersen M, Rosendahl A. Enalapril treatment
increases T cell number and promotes polarization towards M1-like macrophages
locally in diabetic nephropathy. Int Immunopharmacol. 2015; 25(1):30-42.
23. Kaur P, Muthuraman A, Kaur M. The Implications of Angiotensin-Converting
Enzymes and Their Modulators in Neurodegenerative Disorders: Current and Future
Perspectives. ACS Chem Neurosci 2015; 15:508-521.
24. De Lema PG, De Wit CD, Nieto CE et al. Angiotensin inhibition reduces glomerular
damage and renal chemokine expression in MRL/lpr mice. J Pharmacol Exp Ther 2003;
307:275–281.
25. Pasare C., Medzhitov R. Control of B-cell responses by Toll-like receptors. Nature
2005; 438: 364–8.
26. Godsel LM, Leon JS, Wang K et al. Captopril prevents experimental autoimmune
myocarditis. J Immunol 2003; 171:346–52.
27. Bossie A, Vitetta ES. IFN-γ enhances secretion of IgG2a from IgG2a-committed LPS-
stimulated murine B cells: implications for the role of IFN-γ in class switching. Cell
Immunol 1991; 135:95–104.
28. Snapper CM, Peschel C, Paul WE. IFN-γ stimulates IgG2a secretion by murine B
cells stimulated with bacterial lipopolysaccharide. J Immunol 1988; 140:2121–27.

55
29. Carter JD, Valeriano-Marcet J, Kanik KS et al. Antinuclear antibody-negative, drug-
induced lupus caused by lisinopril. South Med J 2001; 94:1122-3.
30. Mahmood IH, Abed MN, Merkhan MM. Effects of blocking of angiotensin system
on the prevalence of metabolic syndrome in type 2 diabetic patients. Pak J Med Sci
2013; 29(1):140-3.
31. Coelho dos Santos JS, Menezes CA, Villani FN, Magalhães LM, Scharfstein J,
Gollob KJ, Dutra WO. Captopril increases the intensity of monocyte infection by
Trypanosoma cruzi and induces human T helper type 17 cells. Clin Exp Immunol. 2010;
162(3):528-36.
32. Montecucco F, Pende A, Mach F. The renin-angiotensin system modulates
inflammatory processes in atherosclerosis: evidence from basic research and clinical
studies. Mediators Inflamm. 2009; 2009:752406.
33. Wysocki PJ1, Kwiatkowska EP, Kazimierczak U, Suchorska W, Kowalczyk DW,
Mackiewicz Captopril, an angiotensin-converting enzyme inhibitor, promotes growth of
immunogenic tumors in mice. Clin Cancer Res. 2006; 12(13):4095-102.
34. Kallenberg CGM, Hoorntje SJ, A. J. Smit AJ et al. Antinuclear and antinative DNA
antibodies during captopril treatment. Acta Med Scand 1982; 21: 297-300.
35. Carter JD, Valeriano-Marcet J, Kanik KS, Vasey FB Antinuclear antibody- negative,
drug-induced lupus caused by lisinopril. South Med J. 2001 Nov;94(11):1122-3.

56
8. ANEXOS

57
ANEXO 1

58
ANEXO 2
Parte dos resultados deste t rabalho compôs um artigo aceito para
publicação no periódico Brazilian Journal of Medical and Biological Research, sob
o título: Enhancing of anti-OVA IgG2c production in vivo by enalapril
Liziane Cristina de Almeida, Lívia Saab Muraro and Deijanira Albuquerque

59
Enhancing of anti-OVA IgG2c production in vivo by enalapril
Liziane Cristina de Almeida1, Deijanira Albuquerque2 and Lívia Saab Muraro3
1Departamento de Biomedicina, Centro Universitário Cândido Rondon, Cuiabá, MT,
Brasil
2Departamento de Ciências Básicas, Faculdade de Medicina, Universidade Federal de
Mato-Grosso, Cuiabá, MT, Brasil
3Departamento de Ciências Veterinárias, Faculdade de Medicina Veterinária,
Universidade Federal do Mato Grosso, Cuiabá, MT, Brasil
Correspondence: Profª Deijanira Alves de Albuquerque
Department of Basic Health Sciences, Faculty of Medicine, Federal University of Mato
Grosso. Avenida Fernando Corrêa, 2.367 Boa Esperança, 78060-900, Cuiabá, MT, Brazil.
Tel.: +556536158847. E-mail address: [email protected]
Senior author: Deijanira Albuquerque
Short title: Enalapril enhances OVA-specific IgG2c
Key-words: ACE inhibitors; Enalapril humoral response; IgG2c antibodies.
Abbreviations used in this paper: ACE, angiotensin-converting enzyme; OVA, EndoFit
ovalbumin; Al, Alhydrogel; EP, enalapril.

60
Abstract
Angiotensin-converting enzyme (ACE) inhibitors have non-hemodynamic, pleiotropic
effects on the immune response. The effects of ACE inhibitors on the production of
cytokines and T-cell functions are well established. However, little is known on the
effects of these medicines on humoral response to foreign antigens. In this study we
investigated the effect of enalapril-long-term treatment on ovalbumin-specific IgG1 and
IgG2c production in mice determined by ELISA. The results show that enalapril
significantly increases anti-OVA serum IgG2c in the secondary response without
affecting IgG1 synthesis. These data expand our understanding on the properties of
enalapril on the immune response, including antibody production.
Introduction
The antibody response to proteins depends on simultaneous activation of Ag-specific
cognate B and T cells. Additionally, the antibody isotope, like IgG2a/IgG2c and IgG1
produced by B cells in response to T-dependent immunogens is droven by cytokines
produced by Th1 and Th2 lymphocytes, respectively (1-3). T-cell polarization into either
Th1 or Th2 profile is influenced by several endogens signals, including cytokines
produced by Ag-presenting cells during the onset of T-cell response. It is also well
established that exogens agents such as adjuvants and some medicines are involved in
shaping the following immune response and thus have a major impact on the profile of
the subsequent T-cell response. In this regard, a large body of clinical and experimental
studies has established that angiotensin-converting (ACE) inhibitors, such as enalapril,
captopril, and lisinopril have pleiotropic, non-hemodynamic properties on T-cell response
by inducing cytokine synthesis (4,5). Accordingly, we have demonstrated that captopril,
an ACE inhibitor with thiol group, inhibits the production of IL-10 and IL-4 without
affecting IL-5, IFN-γ, and IL-2 synthesis in lupus mice (6). In agreement with our
findings, it was recently reported that captopril reduced the production of TNF-α, IL-1α,
IL-10, IL-12, and IL-18 by LPS-stimulated dendritic cells (DC) (7). In a previous study,
we showed that enalapril, an ACE inhibitor without thiol group significantly increases
the number of CD4+CD103+CD25-negative T cells in spleen of normal Balb/c mice
together with the increasing production of IL-10 (8). Moreover, it was recently shown
that enalapril induced an expansion of T cells and re-polarization of macrophages towards

61
a M1-like state in kidneys of diabetic mice (9). So far, most of the studies on immune-
mediated properties of ACE inhibitors have emphasized their effects on cytokine
production and T cell activation (4-9). Little attention however, has been paid to possible
immunemodulatory roles of ACE inhibitors on antibody synthesis. In regard, data from
two clinical studies showed that patients treated with captopril or lisinopril developed,
respectively IgM anti-double-stranded DNA and IgG anti-(H 2A-H 2B)-DNA antibodies
(10,11). However, using the same pharmacological approach, we showed that captopril
does not affect IgG anti-dsDNA antibodies in lupus-prone BWF1 mice (6). Reinforcing
our data, it has been shown that captopril does not alter the production of myosin-specific
antibodies in antigen-immunized mice (12). Based on ours and other author’s findings
(10-12), it could be hypothesized that, at least regarding to captopril effects on
autoantibody production, data from clinical and experimental studies are contradictories.
To extend our overall comprehension on the effects of ACE inhibitors on antibody
production, we sought to analyze whether the widely used ACE inhibitor enalapril would
interfere with anti-ovalbumin (OVA) humoral response in mice. Enalapril was chosen as
the model ACE inhibitor because it regulates cytokine production and, as far as we know,
there is no data on literature on the effect of this ACE inhibitor on humoral response to
foreign antigens in pre-clinical models. In the present work, we have investigated the
effect of enalapril on the humoral response of C57BL/6 mice immunized with EndoFit
ovalbumin (OVA) in the presence of Alhydrogel, as adjuvant. Our results show that
enalapril significantly enhances anti-OVA serum IgG2c without any apparent effect on
OVA-specific IgG1.
Materials and methods
Animals
The 8-week-old C57BL/6 mice used in this study were purchased from CEMIB,
UNICAMP, Campinas, SP. The animals were kept in micro-isolators and all experiments
were performed according to the institutional ethical guidelines on the use of animals in
research (# 23108.039341/12-4).
Antigen and adjuvant
EndoFit ovalbumin (OVA) and Alhydrogel (Al) were used as Ag and adjuvant,
respectively (InvivoGen. San Diego, CA, USA).

62
Immunization
Mice were subcutaneously immunized with OVA (10 µg) in the presence of Alhydrogel
(1 mg) in a total volume of 0.1 mL per animal. 21 days after priming, all animals received
a booster under the same condition of the first immunization. Blood samples were
collected from each individual mouse on days 0, 7, 14, 21, and 28 after primary
immunization and serum was kept at -20oC until used.
Treatment
OVA-immunized mice were treated with enalapril or were left without treatment.
Enalapril (Hexal, Brazil) was dissolved in drinking water at a concentration of 0.02
mg/mL and replaced every 24 h. The daily dosage for enalapril was 5 mg/kg body weight,
assuming a daily fluid intake of 5 mL per mouse. This dose of enalapril is within the usual
therapeutic ranges used in mice (8, 13-15).
ELISA assay
OVA-specific IgG1 was assayed by standard ELISA procedures. In short, polystyrene
plates (Maxsorp) were coated overnight at 4°C with OVA (10 μg/mL), washed with PBS
containing 0.05% Tween-20, blocked with 1%-BSA for 1 h at room temperature and
incubated with serial dilutions of each mouse antiserum starting at 1/100. After overnight
incubation at 4°C, the plates were washed, incubated for 1 h at 37°C with rabbit anti-
mouse IgG1 conjugated to peroxidase (Zymed), washed and developed by addition of
H2O2 and orthophenylenediamine (OPD). The reaction was stopped by addition of
H2SO4, the optical density (OD) at 492 nm was read using a microplate reader (Thermo
Plate-Reader, NM, X model) and the results are expressed as the mean (OD) ± SD. Anti-
OVA IgG2c was assayed following manufacturer’s instructions (eBioscience, USA).
IgG2c concentration was determined by extrapolation from the standard curve. Minimum
level of detection for IgG2c is 0.39 ng/ml.
Measurement of serum IFN-γ
Serum IFN-γ levels were determined by ELISA, using monoclonal antibody pairs and
recombinant cytokines (eBioscience, USA), as previously described (6).
Statistical analysis

63
The results were analyzed using one-way analysis of variance (ANOVA) followed by the
Tukey test. P≤0.05 were considered statistically significant.
Results
Evaluation of the effect of enalapril on ovalbumin-specific IgG1 antibodies.
To investigate whether enalapril would regulate antibody response against ovalbumin,
mice injected with EndoFit ovalbumin (OVA) plus Alhydrogel (Al) were treated with
enalapril for four weeks. For comparative purpose animals were immunized with OVA/Al
and were left without enalapril-treatment. Data presented in figure 1 show that kinetics
of OVA-specific IgG1 response exhibited a classic pattern, with anti-OVA serum IgG1
levels increasing with the time and after the booster. Those results were not affected by
enalapril-treatment. In a similar study, it was shown that captopril, an ACE inhibitor with
thiol group, does not affect OVA-induced IgG responses in mice immunized with this
antigen in complete Freund’s adjuvant (12). Taken together, these data add novel
information on the lack of effects of ACE inhibitors on anti-OVA IgG1 response in the
mouse model. It should be noted that mice immunized with EndoFit ovalbumin plus
Alhydrogel and treated or not with enalapril did not exhibit OVA-specific IgG1, on days
7 and 14 after priming (data not shown). The lack of OVA-specific serum IgG1 at early
primary response may be explained by the lack of LPS in EndoFit ovalbumin, as TLR
ligation on B cells may be necessary for T-dependent antibody responses (16).
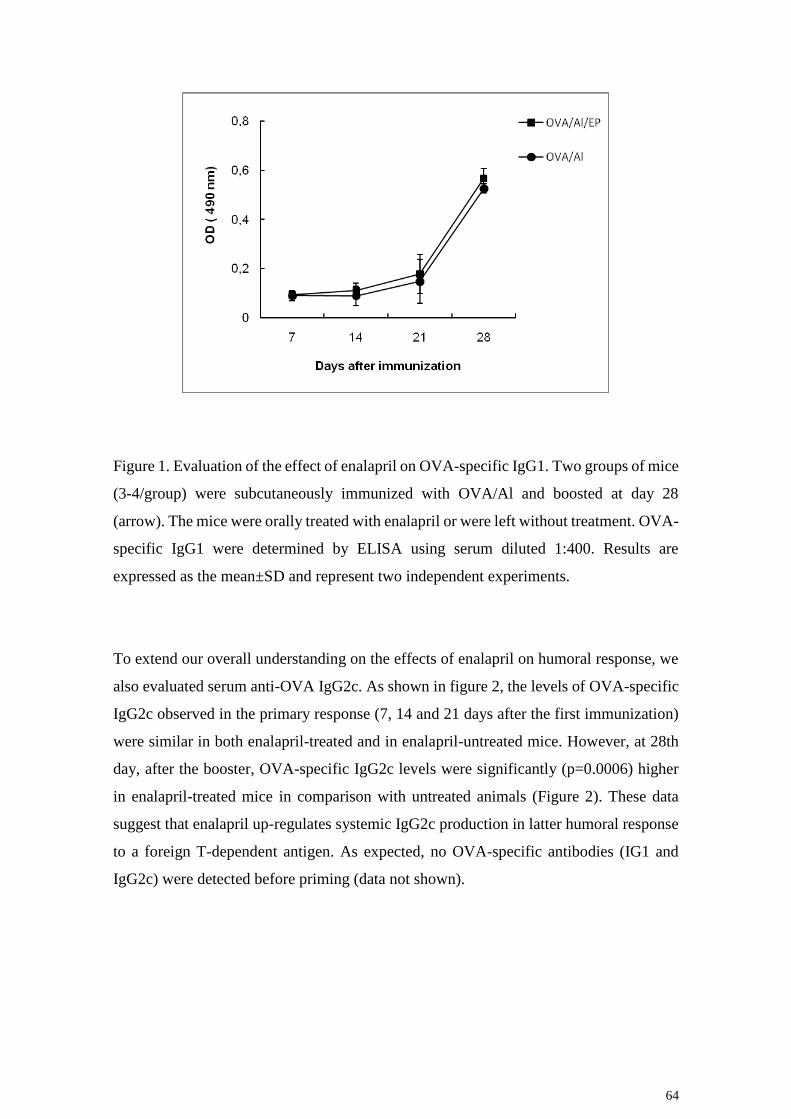
64
Figure 1. Evaluation of the effect of enalapril on OVA-specific IgG1. Two groups of mice
(3-4/group) were subcutaneously immunized with OVA/Al and boosted at day 28
(arrow). The mice were orally treated with enalapril or were left without treatment. OVA-
specific IgG1 were determined by ELISA using serum diluted 1:400. Results are
expressed as the mean±SD and represent two independent experiments.
To extend our overall understanding on the effects of enalapril on humoral response, we
also evaluated serum anti-OVA IgG2c. As shown in figure 2, the levels of OVA-specific
IgG2c observed in the primary response (7, 14 and 21 days after the first immunization)
were similar in both enalapril-treated and in enalapril-untreated mice. However, at 28th
day, after the booster, OVA-specific IgG2c levels were significantly (p=0.0006) higher
in enalapril-treated mice in comparison with untreated animals (Figure 2). These data
suggest that enalapril up-regulates systemic IgG2c production in latter humoral response
to a foreign T-dependent antigen. As expected, no OVA-specific antibodies (IG1 and
IgG2c) were detected before priming (data not shown).
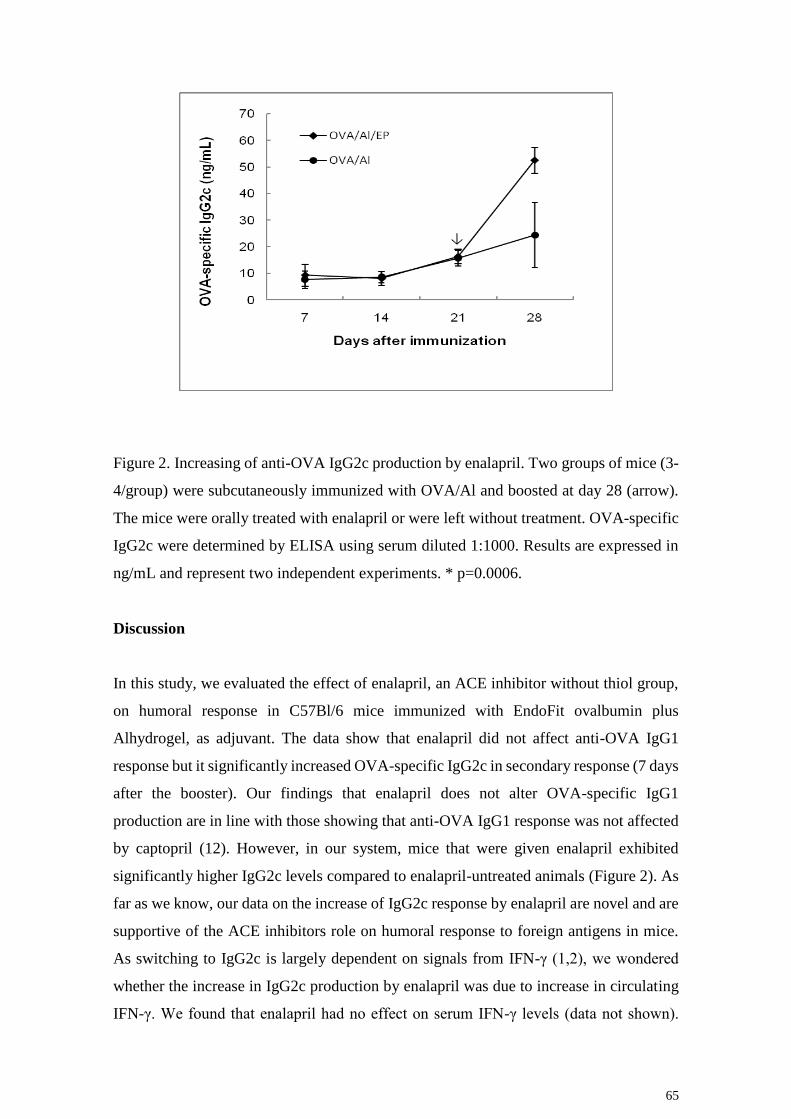
65
Figure 2. Increasing of anti-OVA IgG2c production by enalapril. Two groups of mice (3-
4/group) were subcutaneously immunized with OVA/Al and boosted at day 28 (arrow).
The mice were orally treated with enalapril or were left without treatment. OVA-specific
IgG2c were determined by ELISA using serum diluted 1:1000. Results are expressed in
ng/mL and represent two independent experiments. * p=0.0006.
Discussion
In this study, we evaluated the effect of enalapril, an ACE inhibitor without thiol group,
on humoral response in C57Bl/6 mice immunized with EndoFit ovalbumin plus
Alhydrogel, as adjuvant. The data show that enalapril did not affect anti-OVA IgG1
response but it significantly increased OVA-specific IgG2c in secondary response (7 days
after the booster). Our findings that enalapril does not alter OVA-specific IgG1
production are in line with those showing that anti-OVA IgG1 response was not affected
by captopril (12). However, in our system, mice that were given enalapril exhibited
significantly higher IgG2c levels compared to enalapril-untreated animals (Figure 2). As
far as we know, our data on the increase of IgG2c response by enalapril are novel and are
supportive of the ACE inhibitors role on humoral response to foreign antigens in mice.
As switching to IgG2c is largely dependent on signals from IFN-γ (1,2), we wondered
whether the increase in IgG2c production by enalapril was due to increase in circulating
IFN-γ. We found that enalapril had no effect on serum IFN-γ levels (data not shown).

66
Although, circulating IFN-γ levels were not affected in enalapril-treated mice it could not
be ruled out a possible inhibition of Th2 response by enalapril, as we only measured IFN-
γ by ELISA with minimum level of detection for IFN-γ being 15 pg/ml. Supporting our
hypothesis that enalapril may inhibit local Th2 response, it was recently shown that
enalapril re-polarized macrophages towards a M1-like state in kidneys of diabetic mice
(9). Whether the enhancement of IgG2c production that we verified in our experimental
system is related to inhibition of the local Th2 response by enalapril, it needs to be further
evaluated. The rational for studying the effect of ACE inhibitors on humoral response
relies on the fact that enalapril and captopril are included in the list of medicines
implicated in drug-induced lupus (11). As ACE inhibitors are widely used to treat
hypertensive patients and primary hypertension is a durable condition, one would be
concerned on the effects of these medicines in shifting patient’s Th response to the Th1
profile which might favor the development of autoimmunity diseases governed by Th1
response. Concluding, our results suggest that systemic inhibition of angiotensin II by
enalapril up-regulates IgG2c production without increasing serum IFN-γ levels. Our
results have important implications to our understanding of the regulatory effects of ACE
inhibitors on humoral response mainly opening a new perspective of research on the role
of these medicines on humoral response to foreign T-dependent antigens.
Acknowledgments
The work was supported by grants from the National Council National Council for
Scientific and Technological Development (CNPq) and by fellowship from CNPq (D.
Albuquerque). We thank Prof. Dr. Maria da Guia Silva Lima for critical reading the
manuscript.
Disclosures
Authors have no potential conflicts of interest

67
References
1. Bossie A, Vitetta ES. IFN-γ enhances secretion of IgG2a from IgG2a-committed LPS-
stimulated murine B cells: implications for the role of IFN-γ in class switching. Cell
Immunol 1991; 135:95–104.
2. Snapper CM, Peschel C, Paul WE. IFN-γ stimulates IgG2a secretion by murine B cells
stimulated with bacterial lipopolysaccharide. J Immunol 1988; 140:2121–27.
3. Snapper CM, Finkelman FD, Paul WP. Differential regulation of IgG1 and IgE
synthesis by interleukin 4. J Exp Med 1988; 167:183-197.
4. Gullestad L, Aukrust P, Ueland T et al. Effect of high- versus low-dose angiotensin
converting enzyme inhibition on cytokine levels in chronic heart failure. J Am Coll
Cardiol 1999; 34:2061–67.
5. Platten M, Youssef S, Hur EM et al. Blocking angiotensin-converting enzyme induces
potent regulatory T cells and modulates TH1- and TH17-mediated autoimmunity. Proc
Natl Acad Sci USA 2009; 106:14948–53.
6. De Albuquerque DA, Saxena V, Adams DE et al. An ACE inhibitor reduces Th2
cytokines and TGF-β1 and TGF-β2 isoforms in murine lupus nephritis. Kidney Int 2004;
65:846–59.
7. Lapteva N, Ide K, Nieda M et al. Activation and suppression of renin-angiotensin
system in human dendritic cells. Biochem Biophys Res Commun 2002; 296:194-200.
8. Albuquerque D, Nihei J, Cardillo F et al. The ACE inhibitors enalapril and captopril
modulate cytokine responses in Balb/c and C57Bl/6 normal mice and increase
CD4+CD103+CD25negative splenic T cell numbers. Cell Immunol 2010; 260:92–97.
9. Cucak H, Fink LN, Pedersen MH et al. Enalapril treatment increases T cell number and
promotes polarization towards M1-like macrophages locally in diabetic nephropathy. Int
Immunopharmacol 2015; 25:30–42.

68
10. Kallenberg CGM, Hoorntje SJ, A. J. Smit AJ et al. Antinuclear and antinative DNA
antibodies during captopril treatment. Acta Med Scand 1982; 21: 297-300.
11. Carter JD, Valeriano-Marcet J, Kanik KS et al. Antinuclear antibody-negative, drug-
induced lupus caused by lisinopril. South Med J 2001; 94:1122-3.
12. Godsel LM, Leon JS, Wang K et al. Captopril prevents experimental autoimmune
myocarditis. J Immunol 2003; 171:346–52.
13. Tarkowski A, Carlsten H, Herlitz H et al. Differential effects of captopril and
enalapril, two angiotensin converting enzyme inhibitors, on immune reactivity in
experimental lupus disease. Agents Actions 1990; 31:96–101.
14. Herlitz H, Svalander C, Tarkowski A et al. Effect of captopril on murine systemic
lupus erythematosus disease. J Hypertens 1988; Suppl. 6:
S684–S686.
15. De Lema PG, De Wit CD, Nieto CE et al. Angiotensin inhibition reduces glomerular
damage and renal chemokine expression in MRL/lpr mice.
J Pharmacol Exp Ther 2003; 307:275–281.
16. Pasare C., Medzhitov R. Control of B-cell responses by Toll-like receptors. Nature
2005; 438: 364–8.
69
ANEXO 3
Parte dos resultados deste trabalho está inserida em um artigo submetido ao periódico
Brazilian Journal of Medical and Biological Research, sob o título: Captopril enhances
CD4+ and CD69+ spleen cells in vivo.
Liziane Cristina de Almeida, Carina Souza de Menezes, Suize Silva Oliveira, Édila
Cristina de Souza, Deijanira Alves de Albuquerque.

70
Captopril enhances CD4+ and CD69+ spleen cells in vivo
Liziane Cristina de Almeida1, Carina Souza de Menezes1, Suize Silva Oliveira2, Édila
Cristina de Souza3, Deijanira Alves de Albuquerque4
1Department of Biomedicine. University Center Cândido Rondon, Cuiabá, Mato Grosso,
Brazil.
2Department of Biomedicine. University Center of Várzea Grande, Várzea Grande, Mato
Grosso, Brazil
3Department of Statistics. Federal University of Mato Grosso. Cuiabá, Mato Grosso,
Brazil.
4Department of Basic Health Sciences, Faculty of Medicine, Federal University of Mato
Grosso, Cuiabá, Mato Grosso, Brazil.
Keywords: captopril, enalapril, angiotensin converting enzyme, lymphocytes.
Correspondence: [email protected]
Postal address: Department of Basic Health Sciences, Faculty of Medicine, Federal
University of Mato Grosso. Avenida Fernando Corrêa, 2367. Boa Esperança, 78060-900,
Cuiabá, MT.
Tel.: +556536158847.
E-mail address: [email protected] (D. Albuquerque)

71
Abstract
Angiotensin-converting enzyme (ACE) inhibitors treatment of hypertension and they
possess immunomodulatory properties. The objective of this study was to determine the
effect of captopril and enalapril on the frequency, absolute number, and activation of T
lymphocytes and B and natural killer (NK) cells in vivo using flow cytometry. Female
C57BL6 mice were treated with captopril (30 mg/kg body weight), enalapril (5mg/Kg
body weight) or were orally administered normal drinking water daily for three months.
After treatment, the spleen cells of the mice were labeled with monoclonal antibodies
specific for surface molecules CD4, CD8, CD19, and CD49b. In addition to these
phenotypic markers, monoclonal antibodies specific for activation molecules such as
CD69 and molecules of the major histocompatibility complex type II (MHC II) were also
used. The results showed that captopril increased the frequency and absolute number and
promoted CD4+ activation. However, treatment with enalapril showed no significant
changes in the studied parameters. Thus, we can conclude that captopril, unlike enalapril,
increased the frequency, absolute number, and activation of the CD4+ lymphocyte
subpopulation.
Introduction
Angiotensin-converting enzyme (ACE) inhibitors, such as captopril and enalapril, are
widely prescribed for primary hypertension and for the treatment of cardiovascular
system-related diseases and renal insufficiency; they mainly exert an inhibitory effect on
the renin–angiotensin–aldosterone system (RAAS) (1). ACE inhibition is the main
mechanism by which peptides can modulate RAS function and exert antihypertensive
effects. Angiotensin II (Ang II), the key effector of the RAAS, plays a central role in the
regulation of blood pressure and the fluid system. Various studies indicate that Ang II and
its receptores possess nonhemodynamic activities, which induce inflammatory responses
(2). Additionally, RAAS components are present in immunocompetent cells and may
influence the regulation of immunological functions (3). Captopril and enalapril are
involved in maintaining the balance between vasoconstrictive, hypertrophic, and
salt/water-retentive Ang II and vasodilatory and natriuretic bradykinin by the inhibition
of Ang II biosynthesis and bradykinin degradation (4). Captopril, the first commercially

72
available orally active ACE inhibitor, is a sulfhydryl-containing compound. In contrast,
enalapril, a nonsulfhydryl ACE inhibitor, is a prodrug that releases the active enzyme
inhibitor enalaprilat following hydrolysis. The pharmacokinetic profiles of these three
ACE inhibitors differ. Captopril has a rapid onset with relatively short duration of action,
whereas enalapril has a slower onset and relatively long duration of action (5). In addition
to their ability to reduce blood pressure, captopril and enalapril exert several other
biological activities, such immunomodulatory functions. Beneficial effects of prolonged
enalapril therapy in a type II diabetic rat model are well documented (6). Captopril
ameliorates myocarditis associated with acute Trypanosoma cruzi infection in an
experimental model of acute Chagas heart disease (7). Captopril also exhibits therapeutics
effects in patients with rheumatoid arthritis through a reduction in the serum levels of
tumor necrosis factor-alpha (TNF- α) in rats with rheumatic arthritis (8). Treatment with
various ACE inhibitors reduced atherosclerotic lesions in different animal models (9).
The beneficial effects of treatment with captopril on lupus nephritis have also been
observed in murine models of lupus (10). Further, ACE inhibitors are also used for the
treatment of neurodegenerative disorders, such as Alzheimer's disease and Parkinson's
disease (11). Additionally, captopril and enalapril might affect pro-inflammatory
cytokine and chemokine production (12, 13). Additionally, enalapril increased the subset
population of macrophages and locally increased the number of CD4+ T cells in diabetic
patients with diabetic nephropathy (14). Captopril can reduce the permeability of blood
vessels and eliminate various free radicals, thereby reducing tissue edema and
inflammatory exudation (15). Moreover, captopril interferes with polymorphonuclear
neutrophils-induced cytotoxicity in endothelial cells by modulating pro-inflammatory
surface receptors (16). Furthermore, enalapril treatment potently increases the number of
CD4+ T cells in the spleen along with increasing interleukin-2 (IL-2) and interleukin-10
(IL-10) production (17). There has been considerable interest in the use of ACE inhibitors
in inflammatory disease and for several immune-related diseases. A majority of studies
on the immunomodulatory properties of ACE inhibitors have emphasized their effect on
cytokine production in immune cells. Little attention, however, has been paid to
immunologic roles of ACE inhibitors on splenic lymphocyte populations in non-
manipulated animals. Hence, our objective was to compare captopril- and enalapril-
related phenotypic changes in splenic lymphocytes of C57BL/6 mice.

73
Materials and Methods
Animals
Eight-week-old C57BL/6 mice used in this study were purchased from CEMIB,
Campinas, SP. The animals were kept in micro-isolators, and all experiments were
performed according to the institutional ethical guidelines on the use of animals in
research (# 23108.039341/12-4).
Treatment
The mice were treated with captopril, enalapril or were administered normal drinking
water for three months. Enalapril and captopril (Hexal Laboratory, Brazil) were dissolved
in the drinking water at a concentration of 0.02 and 0.2 mg/mL, respectively. The daily
dosage was 5 mg/kg body weight for enalapril and 30 mg/kg for captopril, assuming a
daily fluid intake of 5 mL. These doses of the two antihypertensive medicines were within
the usual therapeutic ranges used in mice (10,17-18).
Staining of spleen cells for flow cytometry
Staining was performed according to the manufacturer's protocols. Briefly, cells were first
blocked for unspecific binding with anti-CD16/CD32, followed by surface staining with
anti-mouse CD4 peridinin chlorophyll protein-cyanine5.5 (PerCP-Cyanini5.5), anti-
mouse CD8 PerCP-Cyanini5.5, anti-mouse CD19 phycoerythrin (PE), anti-mouse CD49b
PE, anti-mouse CD69 allophycocyanin (APC), and anti-mouse MHC II peridinin
chlorophyll protein-eFluor (PerCP-efluor) (all antibodies were obtained eBioscience, San
Diego, California, USA). In this study, two acquisitions were performed for every sample;
one for total cells, in order to determine the absolute frequencies, and the other
deliberately gating positive events for further analysis. A total of 20,000 events were
recorded per sample. Acquisitions and data analysis were performed on BD Accuri™ C6
flow cytometer (Becton Dickinson, San Jose, CA, USA).
Statistical analysis
The results were analyzed by one-way analysis of variance (ANOVA) followed by the
Bonferroni test. P ≤ 0.05 was considered statistically significant.
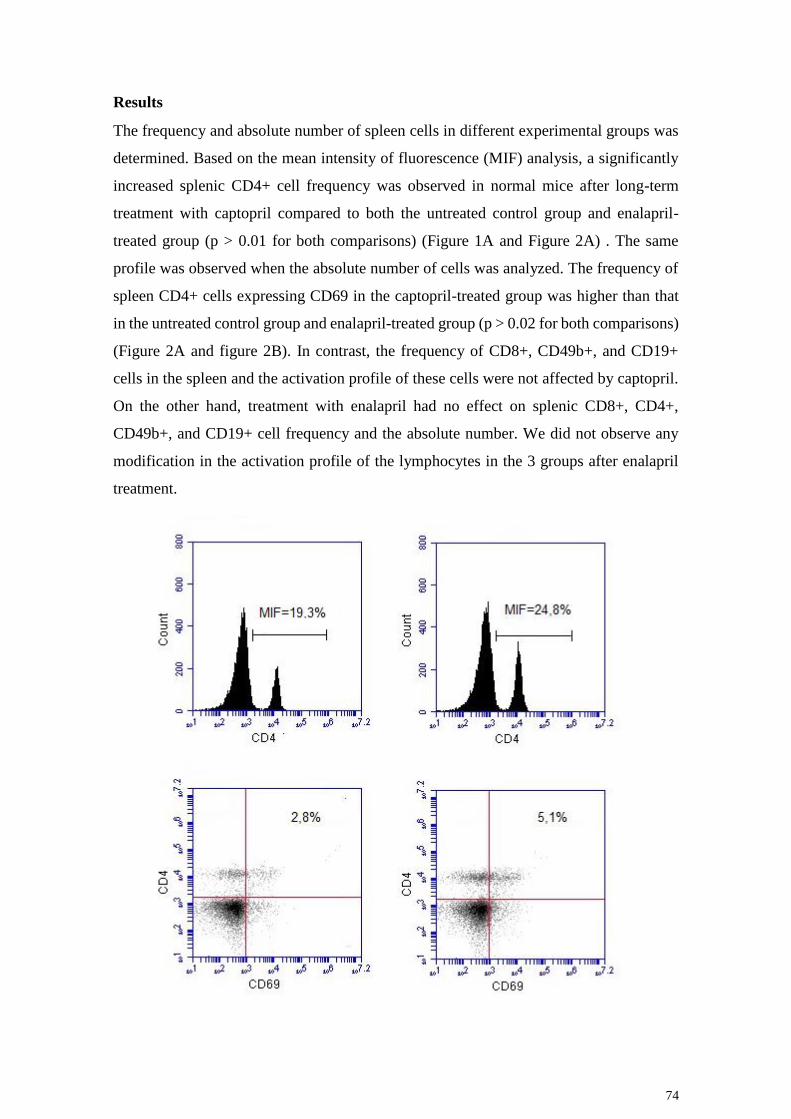
74
Results
The frequency and absolute number of spleen cells in different experimental groups was
determined. Based on the mean intensity of fluorescence (MIF) analysis, a significantly
increased splenic CD4+ cell frequency was observed in normal mice after long-term
treatment with captopril compared to both the untreated control group and enalapril-
treated group (p > 0.01 for both comparisons) (Figure 1A and Figure 2A) . The same
profile was observed when the absolute number of cells was analyzed. The frequency of
spleen CD4+ cells expressing CD69 in the captopril-treated group was higher than that
in the untreated control group and enalapril-treated group (p > 0.02 for both comparisons)
(Figure 2A and figure 2B). In contrast, the frequency of CD8+, CD49b+, and CD19+
cells in the spleen and the activation profile of these cells were not affected by captopril.
On the other hand, treatment with enalapril had no effect on splenic CD8+, CD4+,
CD49b+, and CD19+ cell frequency and the absolute number. We did not observe any
modification in the activation profile of the lymphocytes in the 3 groups after enalapril
treatment.
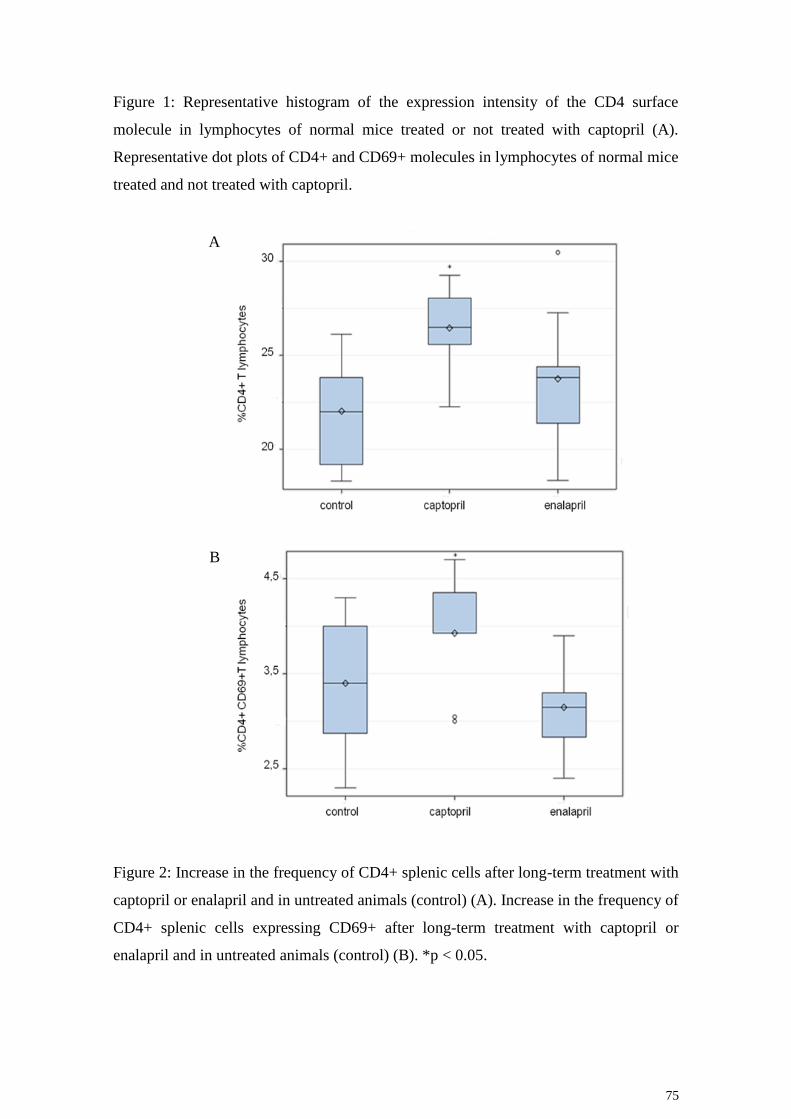
75
Figure 1: Representative histogram of the expression intensity of the CD4 surface
molecule in lymphocytes of normal mice treated or not treated with captopril (A).
Representative dot plots of CD4+ and CD69+ molecules in lymphocytes of normal mice
treated and not treated with captopril.
A
B
Figure 2: Increase in the frequency of CD4+ splenic cells after long-term treatment with
captopril or enalapril and in untreated animals (control) (A). Increase in the frequency of
CD4+ splenic cells expressing CD69+ after long-term treatment with captopril or
enalapril and in untreated animals (control) (B). *p < 0.05.

76
Discussion
In this work, we investigated the effects of captopril and enalapril on subpopulations of
lymphocytes CD4+ and CD8+ and B and NK in normal C57BL/6 mice. Our results
showed that captopril significantly increased the frequency and absolute number of CD4+
lymphocytes in vivo. However, under the same experimental conditions, CD8+
lymphocyte subsets and B and NK were not affected by treatment with captopril.
Moreover, the findings of our study revealed that captopril induces the expression of
early-activating antigen CD69 only in CD4+ lymphocytes. These results support the
concept that CD69 negatively regulates the immune response and inflammatory
phenomenon (19). In contrast, the results showed that treatment with enalapril did not
change the parameters studied in the lymphocyte subpopulations. The study in normal
BALB/c mice showed that enalapril increased the number of CD4+, CD103+, and CD25-
cells (17). Previous studies showed that various biological activities of captopril were
associated with its influence on the immune system in experimental animal models of
various diseases (8-10). Our experimental study reported the effect of captopril in
lymphocytes in a non-manipulated animal. Further studies are required to clarify the
mechanism by which the captopril acts only on CD4+ lymphocytes. The effects observed
in our study may be related to the presence of sulfhydryl group in its structure and are
independent of its effect on the renin angiotensin system. Finally, the results of flow
cytometry indicated that captopril might interfere with the immune response to their
beneficial on immunologic cells, by enhancing CD4+T lymphocytes, thereby suggesting
that a novel therapeutic agent can be used for the inflammatory process that is not
necessarily related to the blood pressure.
Acknowledgments
This work was supported by the Research Foundation of Mato Grosso State (FAPEMAT)
(#751305/2011) and National Counsel of Technological and Scientific Development
(CNPq) findings (#471234/2011-7).
Conflict of interest
The authors declare no conflicts of interests.

77
References
1. Li EC, Heran BS, Wright JM. Angiotensin converting enzyme (ACE) inhibitors versus
angiotensin receptor blockers for primary hypertension. Cochrane Database Syst Rev
2014; 8: CD009096.
2. Cheng ZJ, Vapaatalo H, Mervaala E. Angiotensin II and vascular inflammation. Med.
Sci. Monit 2005; 11: RA194–RA205.
3. Hoch NE, Guzik TJ, Chen W., Deans T, Maalouf SA, Gratze P, Weyand C, Harrison
DG. Regulation of T-cell function by endogenously produced angiotensin II. Am J
Physiol Regul Integr Comp Physiol 2009; 296: R208–R216.
4. Regulski M, Regulska K, Stanisz BJ, Murias M, Gieremek P, Wzgarda A, Niznik B.
Chemistry and pharmacology of Angiotensin-converting enzyme inhibitors. Curr Pharm
Des 2015; 21:1764-1775.
5. Raia JJ Jr, Barone JA, Byerly WG, Lacy CR. Angiotensin-converting enzyme
inhibitors: a comparative review. DICP 1990; 24:506–525.
6. Kang YS, Ko GJ, Lee MH, Song HK, Han SY, Han KH, Kim KH, Han JY, Cha DR.
Effect of eplerenone, enalapril and their combination treatment on diabetic nephropathy
in type II diabetic rats. Nephrol Dial Transplant 2009; 24:73–84.
7. Leon JS, Wang K, Engman DM. Captopril Ameliorates Myocarditis in Acute
Experimental Chagas Disease. Circulation 2003; 107: 2264-2269.
8. Liu HM, Wang KJ. Therapeutic effect of Captopril on rheumatoid arthritis in rats.
Asian Pac J Trop Med 2014; 7:996-999.
9. Montecucco F, Pende A, Mach F. The renin-angiotensin system modulates
inflammatory processes in atherosclerosis: evidence from basic research and clinical
studies. Mediators Inflamm 2009; 2009:752406.

78
10. Albuquerque DA, Saxena V, Adams DE, Boivin GP, Brunner HI, Witte DP, Singh
RR. An ACE inhibitor reduces Th2 cytokines and TGF-beta1 and TGF-beta2 isoforms in
murine lupus nephritis. Kidney Int 2004; 65:846-859.
11. Kaur P, Muthuraman A, Kaur M. The Implications of Angiotensin-Converting
Enzymes and Their Modulators in Neurodegenerative Disorders: Current and Future
Perspectives. ACS Chem Neurosci 2015; 15:508-521.
12. Alla JA, Langer A, Elzahwy SS, Arman-Kalcek G, Streichert T, Quitterer U.
Angiotensin-converting enzyme inhibition down-regulates the pro-atherogenic
chemokine receptor 9 (CCR9) -chemokine ligand 25 (CCL25) axis. Biol Chem 2010;
23:23496-23505.
13. Andrzejczak D, Górska D, Czarnecka E. Influence of enalapril, quinapril and losartan
on lipopolysaccharide (LPS) -induced serum concentrations of TNF-alpha, IL-1 beta, IL-
6 in spontaneously hypertensive rats (SHR). Pharmacol Rep 2007; 59:437-446.
14. Cucak H, Nielsen FL, Hojgaard Pedersen M, Rosendahl A. Enalapril treatment
increases T cell number and promotes polarization towards M1-like macrophages locally
in diabetic nephropathy. Int Immunopharmacol 2015; 25:30-42.
15. Liu HM, Guo YN. Effect of captopril on serum TNF- alfa level in acute lung injury
rats induced by HCL. Asian Pac J Trop Med. 2014; 7:905-908.
16. Andersson P, Bratt J, Heimbürger M, Cederholm T, Palmblad J. Inhibition of
neutrophil-dependent cytotoxicity for human endothelial cells by ACE inhibitors. Scand
J Immunol 2014; 80:339-345.
17. Albuquerque D, Nihei J, Cardillo F, Singh R. The ACE inhibitors enalapril and
captopril modulate cytokine responses in Balb/c and C57Bl/6 normal mice and increase
CD4(+)CD103(+)CD25(negative) splenic T cell numbers. Cell Immunol 2010; 260: 92-
97.

79
18. Lema GP, Wit C, Cohen CD, Nieto E, Molina A, Banas B, Luckow B, Vicente AB,
Mampaso F, Schlöndorff D. Angiotensin inhibition reduces glomerular damage and renal
chemokine expression in MRL/lpr mice.J. Pharmacol. Exp. Ther 2003; 307:275–281.
19. González-Amaro R, Cortés JR, Sánchez-Madrid F, Martín P. Is CD69 an effective
brake to control inflammatory diseases? Trends Mol Med 2013; 19:625–632.