antigen-dependent b cell development prof. sylvie fournier
TRANSCRIPT

Antigen-dependent B cell development
Prof. Sylvie Fournier

“Traditional” view of B lymphocyte development
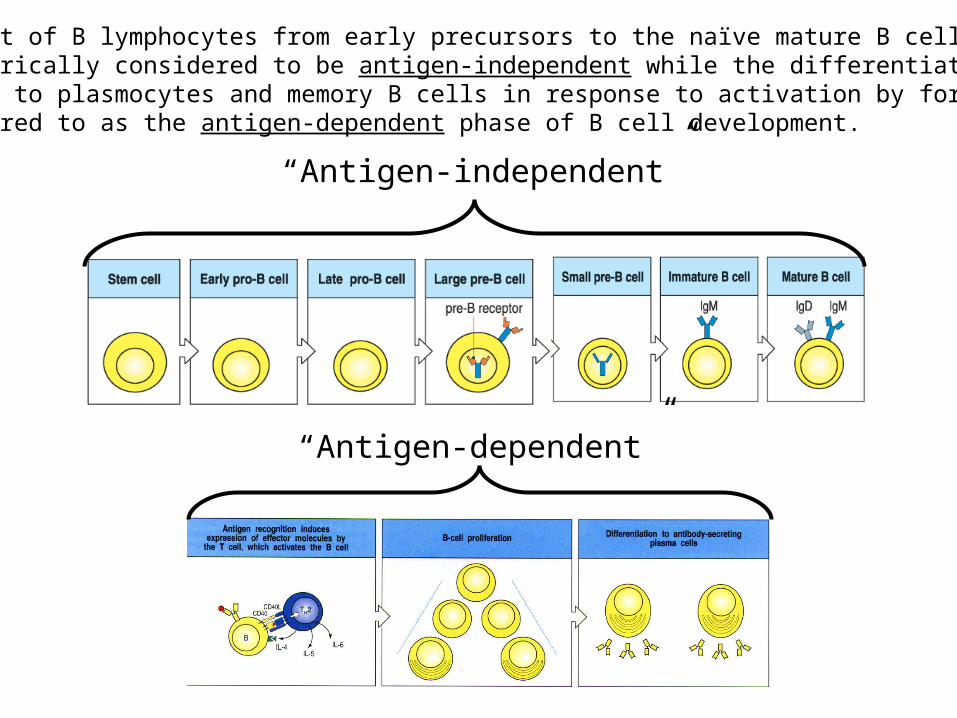
“Antigen-independent”
The development of B lymphocytes from early precursors to the naïve mature B cell stagehas been historically considered to be antigen-independent while the differentiation of naïve mature B cells to plasmocytes and memory B cells in response to activation by foreign Ags has been referred to as the antigen-dependent phase of B cell development.
“Antigen-dependent”
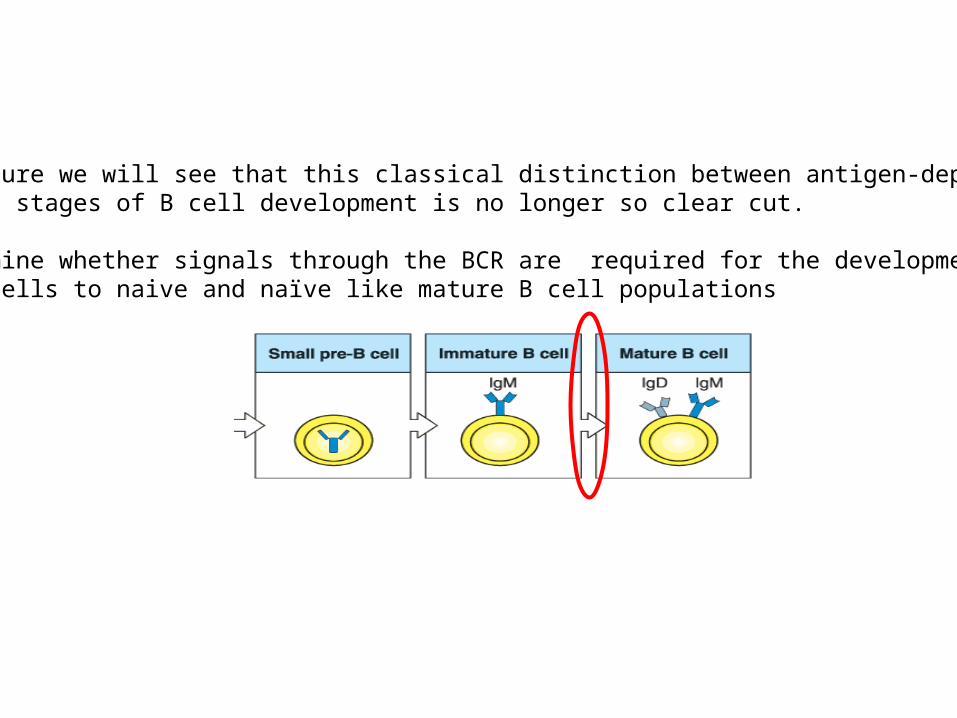
In this lecture we will see that this classical distinction between antigen-dependent and -independent stages of B cell development is no longer so clear cut.
We will examine whether signals through the BCR are required for the development of immature B cells to naive and naïve like mature B cell populations
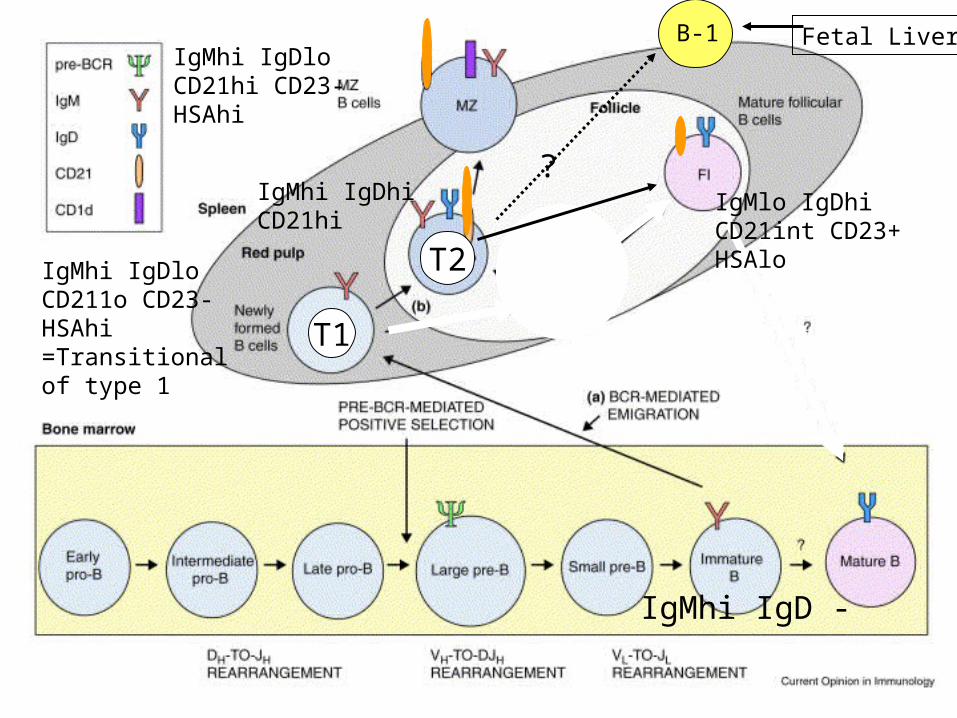
IgMhi IgD -
IgMhi IgDloCD211o CD23-HSAhi=Transitionalof type 1
IgMhi IgDloCD21hi CD23-HSAhi
IgMhi IgDhiCD21hi
T1
T2
IgMlo IgDhiCD21int CD23+HSAlo
B-1
?
Fetal Liver

T1 cells are recent emigrants from the bone marrow. Their phenotype is closely related to thatof immature B cells in the bone marrow: T1: IgMhigh , IgDlow/neg, CD21low/neg, CD23 low/neg.
T1 cells that successfully enter splenic B-cell follicles become T2 cells. T2 cells have a mixedphenotype between that of T1 cells and mature follicular (FO) B cells:T2: IgMhigh , IgDhigh, CD21high, CD23 +.FO: IgMlow , IgDhigh, CD21int, CD23 +.
T2 B cells are considered to be precursors for FO and MZ B cells, and some B-1 cells.MZ: IgMhigh , IgDlow, CD21high, CD23low/neg.
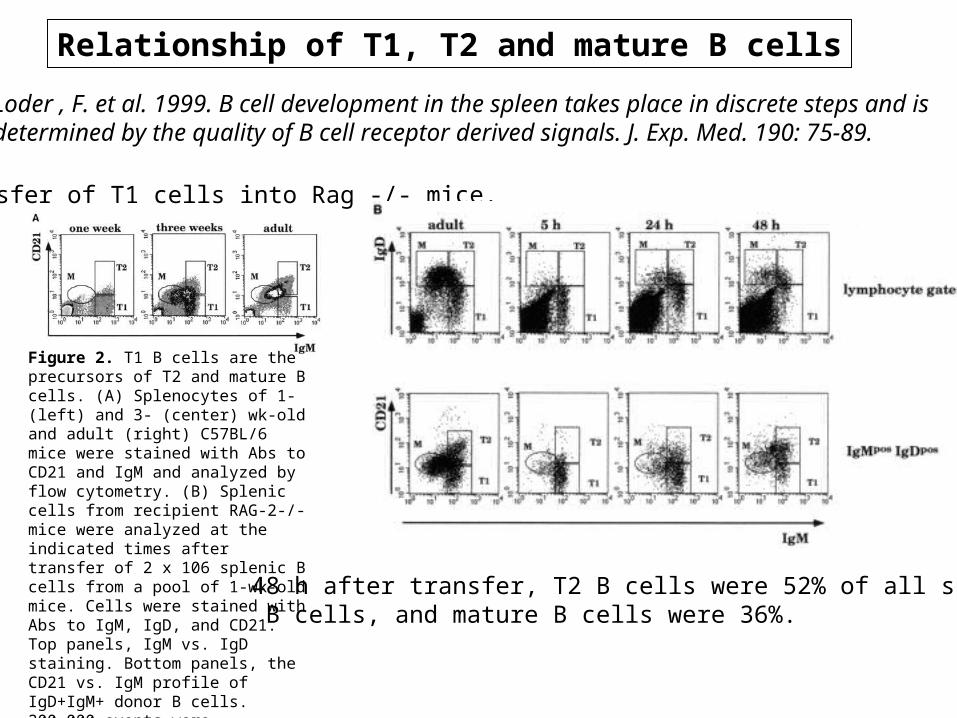
Relationship of T1, T2 and mature B cells
Loder , F. et al. 1999. B cell development in the spleen takes place in discrete steps and is determined by the quality of B cell receptor derived signals. J. Exp. Med. 190: 75-89.
Transfer of T1 cells into Rag -/- mice.
48 h after transfer, T2 B cells were 52% of all splenic B cells, and mature B cells were 36%.
Figure 2. T1 B cells are the precursors of T2 and mature B cells. (A) Splenocytes of 1- (left) and 3- (center) wk-old and adult (right) C57BL/6 mice were stained with Abs to CD21 and IgM and analyzed by flow cytometry. (B) Splenic cells from recipient RAG-2-/- mice were analyzed at the indicated times after transfer of 2 x 106 splenic B cells from a pool of 1-wk-old mice. Cells were stained with Abs to IgM, IgD, and CD21. Top panels, IgM vs. IgD staining. Bottom panels, the CD21 vs. IgM profile of IgD+IgM+ donor B cells. 200,000 events were collected. Data is shown as dot plots to highlight the few transferred cells that home to the spleen. In the dot plots corresponding to the control (adult) spleen, only 5% of the collected events are shown.
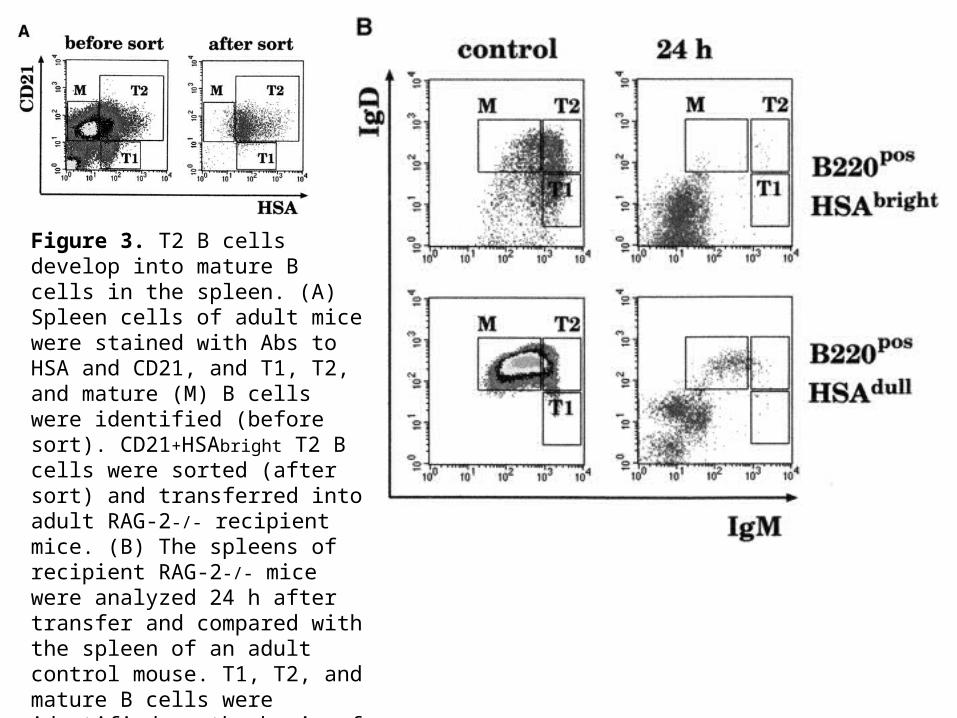
Figure 3. T2 B cells develop into mature B cells in the spleen. (A) Spleen cells of adult mice were stained with Abs to HSA and CD21, and T1, T2, and mature (M) B cells were identified (before sort). CD21+HSAbright T2 B cells were sorted (after sort) and transferred into adult RAG-2-/- recipient mice. (B) The spleens of recipient RAG-2-/- mice were analyzed 24 h after transfer and compared with the spleen of an adult control mouse. T1, T2, and mature B cells were identified on the basis of the expression of HSA, B220, IgM, and IgD. The plots show the IgD and IgM staining of cells that were positive for B220 and either bright or dull for HSA.

What supports massive cell loss at or soon after the immature B-cellStage in the bone marrow?
• The high rate of B cell production: Immature B cells: 15-20 x 106/day
(Osmond, D. G. 1986. Population dynamics of bone marrow B lymphocytes. Immunol. Rev. 93:103.)
• Half-life of mature follicular B cells: 4.5 months
(Hao, Z. and Rajewsky, K. 2001. Homeostasis of peripheral B cells in the absence of B cell influx from the bone marrow. J. Exp. Med. 194: 1151-1163.)
One way the relatively long lifespan of follicular B cells can be reconciled with the highrate of production is to assume that a large proportion of recently generated B cells are lost
Emigration and loss of recently generated B cells

At the immature stage, about 90% of the immature B cells produced are lost. A large part of thisloss can probably be explained by the deletion of autoreactive B cells in the bone marrow.
However, signaling through the BCR appears to play a role in the emigration of immatureB cells from the bone marrow to the spleen.
• Mice with an engineered mutation in the cytoplasmic tail of Ig: B cell development up to the immature B cell stage in the bone marrow is minimally affected, but emigration to the peripheryappears to be markedly reduced. (Torres, R. M., et al. 1996. Aberrant B cell development and immune response in mice with a compromised BCR complex. Science 272:1804).
• BCR Tg mice with a deletion of Syk: B cells at the immature stage are detected in the bonemarrow but very few are present in the spleen. (Turner, M. et al. (1997). Syk tyrosine kinase is requiredfor the positive selection of immature B cells into the recirculating B cell pool. J. Exp. Med. 186:2013-2021.)
• CD45-/- and Xid (Btk defective) mice: Marked increase of immature B cells in the spleen thatmay be consistent with an accelerated emergence of immature B cells from the bone marrow.
According to these findings, it is therefore assumed that emigration from the bone marrow probably represents a checkpoint in B cell development, and that this process is regulatedby signals received through the BCR.

Now if there is a BCR-driven checkpoint exits at this stage of B cell development in the bonemarrow, the question is whether this BCR signal is constitutive or generated by self-antigens.The most commonly held view is that it is probably a constitutive signal. If the BCR-generated signal is constitutive it may be ligand independent (“tonic” signaling; the mere processof receptor assembly), or it may depend on some extracellular ligand that recognizes anonvariable portion of every BCR.Perhaps not not every possible immunoglobulin heavy-light chain is capable of associating wellenough to generate appropriate levels of properly assembled receptor to initiate a tonic signal.It is possible that what is being selected at the immature B cell stage in the bone marrow is anappropriate fit between pairs of heavy and light chains
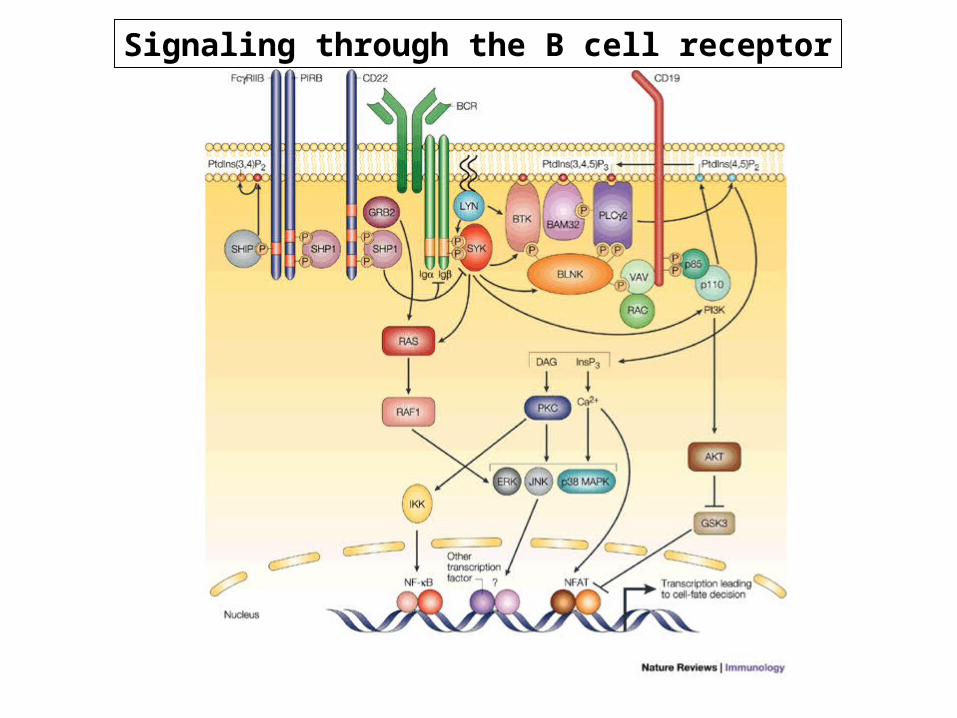
Signaling through the B cell receptor

Signals required for the T1 to T2 transition
Only a small percentage of T1 cells proceed to the T2 stage. Most T1 undergo clonal deletionor anergy. These represent B cells that cross-react with self-antigens expressed only in peripheral tissues. However, basal BCR signals are required for the transition from T1 to T2 B cell stage.
T1 T2
Syk -/-Ig ∆c/∆c Ig ∆c/∆c
Baff -/-A/WySnJ (mutation in BaffR)
T1
Syk -/-Ig ∆c/∆c Ig ∆c/∆c
Baff -/-A/WySnJ (mutation in BaffR)
T2 FO
Btk -/-CD45 -/-
PLC2 -/-Dominant negative of MEK
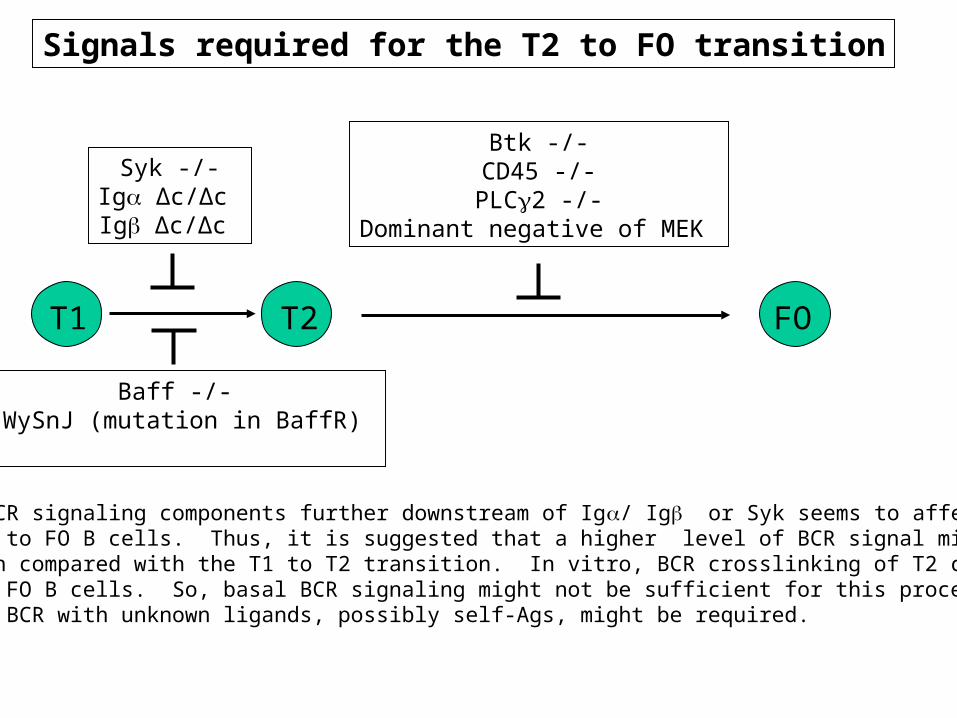
Signals required for the T2 to FO transition
As disruption of BCR signaling components further downstream of Ig/ Ig or Syk seems to affect thetransition from T2 to FO B cells. Thus, it is suggested that a higher level of BCR signal might be required for this transition compared with the T1 to T2 transition. In vitro, BCR crosslinking of T2 cells induces theirdifferentiation to FO B cells. So, basal BCR signaling might not be sufficient for this process, andinteraction of the BCR with unknown ligands, possibly self-Ags, might be required.
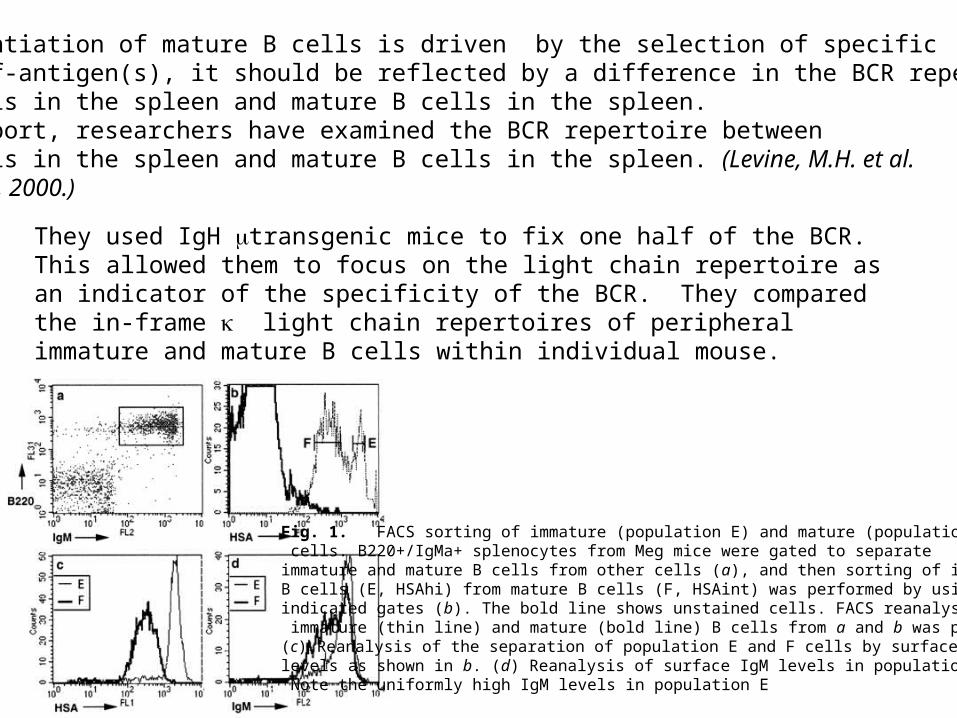
If the differentiation of mature B cells is driven by the selection of specificB cells by self-antigen(s), it should be reflected by a difference in the BCR repertoire betweenimmature B cells in the spleen and mature B cells in the spleen.In a recent report, researchers have examined the BCR repertoire betweenimmature B cells in the spleen and mature B cells in the spleen. (Levine, M.H. et al.PNAS 97:2743-2748, 2000.)
Fig. 1. FACS sorting of immature (population E) and mature (population F) B cells. B220+/IgMa+ splenocytes from Meg mice were gated to separate immature and mature B cells from other cells (a), and then sorting of immature B cells (E, HSAhi) from mature B cells (F, HSAint) was performed by using the indicated gates (b). The bold line shows unstained cells. FACS reanalysis of immature (thin line) and mature (bold line) B cells from a and b was performed. (c) Reanalysis of the separation of population E and F cells by surface HSA levels as shown in b. (d) Reanalysis of surface IgM levels in population E and F. Note the uniformly high IgM levels in population E
They used IgH transgenic mice to fix one half of the BCR. This allowed them to focus on the light chain repertoire as an indicator of the specificity of the BCR. They compared the in-frame light chain repertoires of peripheral immature and mature B cells within individual mouse.

In all three mice of this line, the immature B cell light chain sequences were diverse (Fig. 2 a-c). However, in the mature B cells of all three mice, the frequency of a single member of the V24/25 family, sequence 80 as denoted by Strohal et al. (15) was significantly enhanced (Fig. 2 a-c). The frequency of the sequence 80 light chain was 3- to 7-fold higher in the mature B cell population compared with the immature B cell population within the same mouse. The magnitude of difference in all mice analyzed was similar, and in all cases the sequence 80 light chain was the most prevalent light chain isolated from the mature B cells.

Fig. 2. In-frame light chain repertoire is more restricted in mature naive B cells than in immature B cells from three Meg (VH186.2 transgenic ¥ JH/) mice. light chain joins to J2 from immature (population E,white bars) and mature (population F, black bars) B cell populations were amplified, cloned, and sequenced. Each panel compares the in-frame light chain repertoire of immature and mature splenic B cells from an individual mouse. The horizontal axis is labeled with V families, and chain numbers as assigned by Strohal et al. (15). Chains not identified by Strohal are labeled N1-N9. The vertical axis depicts standardized light chain expression frequency as a percentage of total chains sequenced.(a) V24/25 family member number 80 (marked with an asterisk) was found to be at a three-fold greater percentage in mature (black bars) than immature (white bars) B cells within the first mouse (E, n = 37; F, n = 42). This V24/25 family member also represents the most common light chain in the mature B cell population. (b) In the second mouse, the same V24/25 family member number 80 was found to be enhanced at the E to F transition,representing 12% of total in-frame V chains in the mature B cell population and only 2% in the immature population (E, n = 48; F, n = 59). (c) In a third mouse analyzed, the same V24/25 family member number 80 once again appeared at a much greater frequency in the mature B cell population (12%) than the immature B cell population (2.5%) (E, n = 40; F, n = 50). (d) Tabulation of all V-J2 sequences from the Meg line for immature (E) and mature (F) cells is shown. The horizontal axis is labeled with as above whereas the vertical axis depicts total number of sequences with the immature numbers normalized to be equivalent to the mature numbers (n = 151 for both populations). V24/25 sequence 80 (marked with an asterisk) represents just under 5 sequences of 151 in immature B cells (3.2%) and represents 19 of 151 sequences (12.6%) in mature B cells. This enhancement is highly significant (P < 0.003)

The fundamental significance of these data is that they demonstrate that the loss of the majority of B cells at the peripheral immature to mature follicular transition is selective rather than stochastic.
Although not proved directly by these studies, the authors favor the interpretation that certain cells are positively selected.
If positive selection is occurring at this transition, the question is raised: Why must B cells be positively selected, and what is the nature of the selecting ligand?
It is possible that environmental antigens and nonpathogenic flora may be "preselecting" a naive recirculating B cell repertoire that is predisposed to recognize pathogenic antigens it may later encounter.
Regardless of the purpose of receptor-specific selection of immature B cells into the mature compartment, the present work adds another dimension to the mounting evidence that receptor-ligandinteractions signal for the ongoing survival of lymphocytes in the periphery.

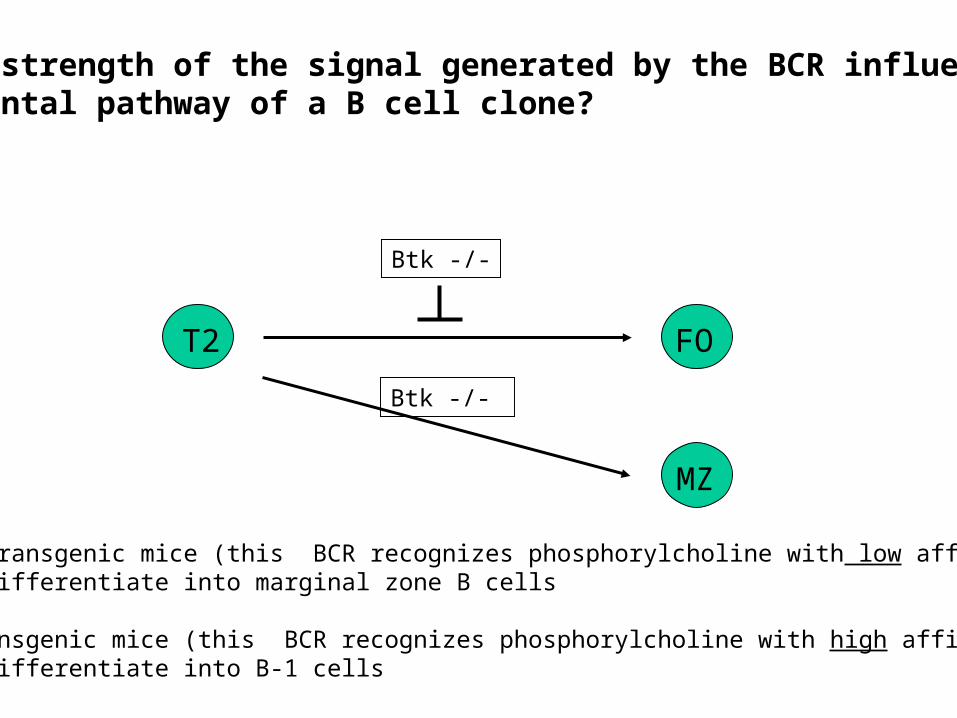
Does the specificity of the BCR dictate the developmental pathwayof a B cell clone?
• VH81X transgenic mice (this BCR recognizes phosphorylcholine with low affinity): B cells differentiate into marginal zone B cells
• Anti-HEL transgenic mice :B cells differentiate into follicular B cells
• A restricted BCR repertoire that is weakly self-reactive is selected preferentially in MZ and B1 B cells.
• Microbial products, recognized specifically through TLRs, are sufficient to drive the differentiation of T2 cells to MZ B cells in vitro.
• In mice that lack the normal commensal intestinal flora, the generation of MZ and B1 cells is impaired.
T2 FO
MZ
Btk -/-
Btk -/-
Does the strength of the signal generated by the BCR influence thedevelopmental pathway of a B cell clone?
• VH81X transgenic mice (this BCR recognizes phosphorylcholine with low affinity): B cells differentiate into marginal zone B cells
• T15 transgenic mice (this BCR recognizes phosphorylcholine with high affinity):B cells differentiate into B-1 cells

We will examine more closely data that supports the notion that the strength of theBCR signal that B cells receive during the late phases of their differentiation drives their differentiation into different mature B cell subsets.
Cariappa A. et al.Immunity, 14:603-615, 2001.The follicular versus marginal zone B lymphocyte fate decision is regulated byAiolos, Btk and CD21.

Aiolos -/-: BCR signaling is increased.
Xid (Btk is not functional):BCR signaling is decreased.
T1: IgMhigh IgDlowT2: IgMhigh IgDhighFO: IgMlow IgDhigh
The ratio of mature B cells tonewly formed B cells is increased inAiolos -/- mice but decreased in Xidmice.
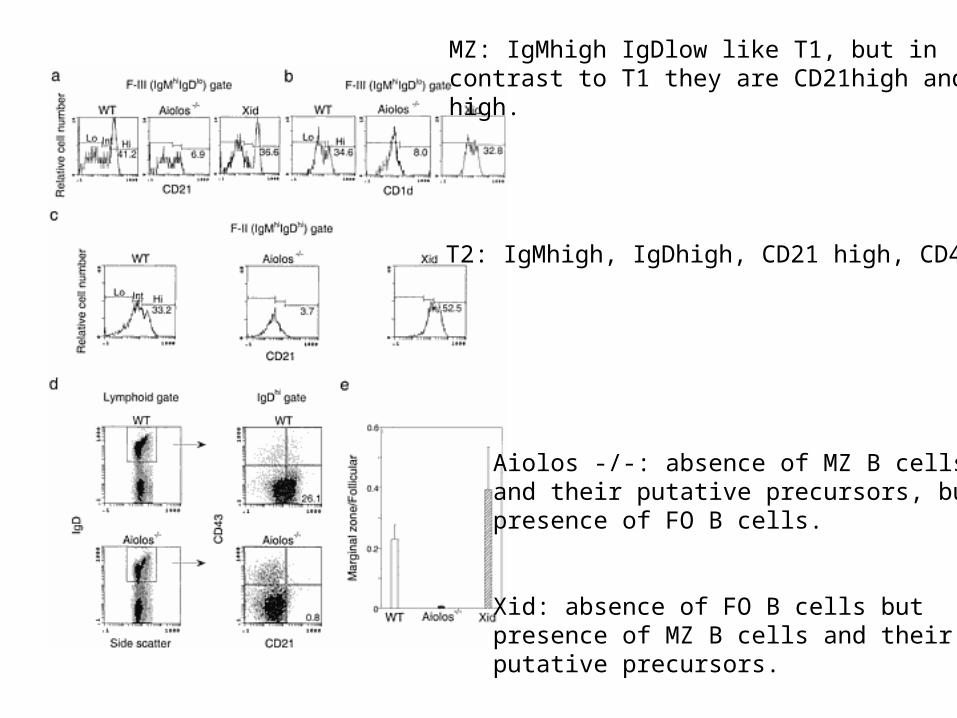
Aiolos -/-: absence of MZ B cellsand their putative precursors, butpresence of FO B cells.
Xid: absence of FO B cells butpresence of MZ B cells and theirputative precursors.
MZ: IgMhigh IgDlow like T1, but in contrast to T1 they are CD21high and CD1high.
T2: IgMhigh, IgDhigh, CD21 high, CD43-
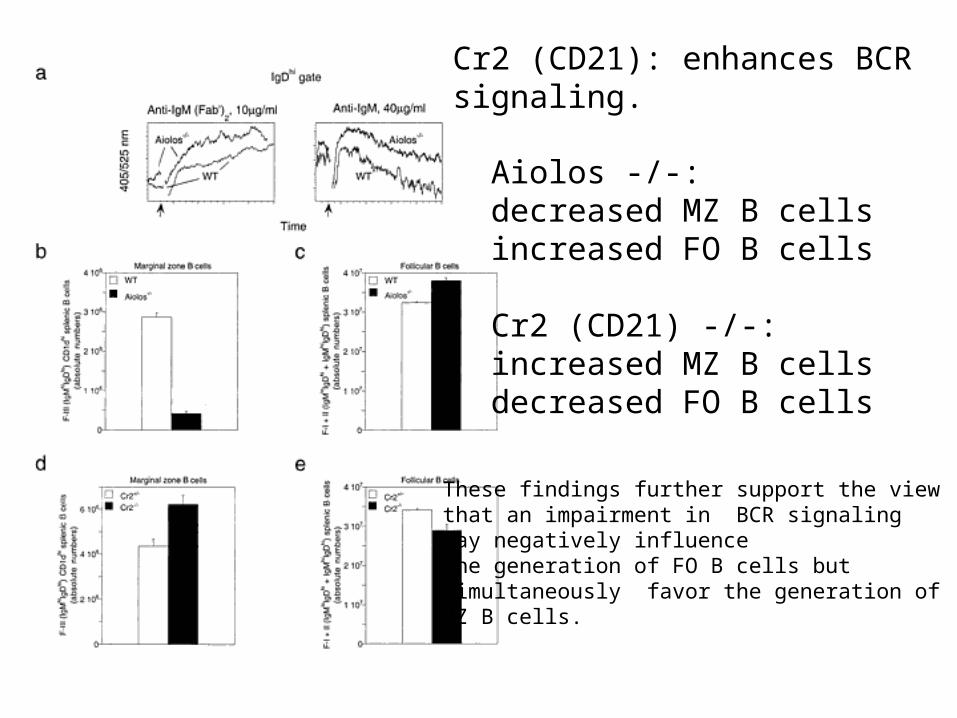
Aiolos -/-:decreased MZ B cellsincreased FO B cells
Cr2 (CD21) -/-:increased MZ B cellsdecreased FO B cells
Cr2 (CD21): enhances BCRsignaling.
These findings further support the viewthat an impairment in BCR signalingmay negatively influencethe generation of FO B cells butsimultaneously favor the generation ofMZ B cells.

It is known that B cells bearing high-affinity receptors for PC (bearing the T15 idiotype for instance) develop into B-1 cells whereas antigen-receptors with low avidity for PC acquire a MZ B-cell phenotype.
Although specific BCRs can clearly favor B-1 differentiation, reducing the level of surface expression of a “B-1 committed BCR” can induce a cell to acquire a follicular or B-2 fate.
B-1-cell numbers are enhanced in mice that lack the immunoreceptor tyrosine-based inhibition motif(ITIM)-containing negative regulator of B-cell signaling, CD72, which is known to recruit SHP-1, as well as in viable motheaten mice, which harbor a defect in SHP-1.
These data argue that very strong BCR signals favor B-1-cell generation.

An emerging theme for peripheral B-cell development is that the strength of signal delivered via self-antigen/BCR interactions contributes in a significant way to the differentiative fate of an emerging B-cell clone. This model is based on the study of B cell development in mice that lack various componentsof the BCR signaling machinery.
• Disruption of the proximal and pivotal kinase Syk leads to the absence of the three subsets of mature B cells(MZ, FO, B-1).
• In the absence of Btk alone (“Weak BCR signals” only) or of its major substrate, PLC2, follicular B cells and B-1 B cells are lost. However, MZ B-cell development is by large normal. • In the absence of PKC alone, which is downstream of PLC2 in the signaling cascade of the BCR,(`Weak' and `Intermediate BCR signals' only), follicular B cells are not lost and MZ B cells presumably remain unaffected, but B-1 cells are markedly deficient.
Weak BCR signals are therefore permissive for MZ B-cell development, but not for follicular and B-1 cells, intermediate signals are required for follicular B-cell development, and strong BCR signals are required for the positive selection of B-1 cells.
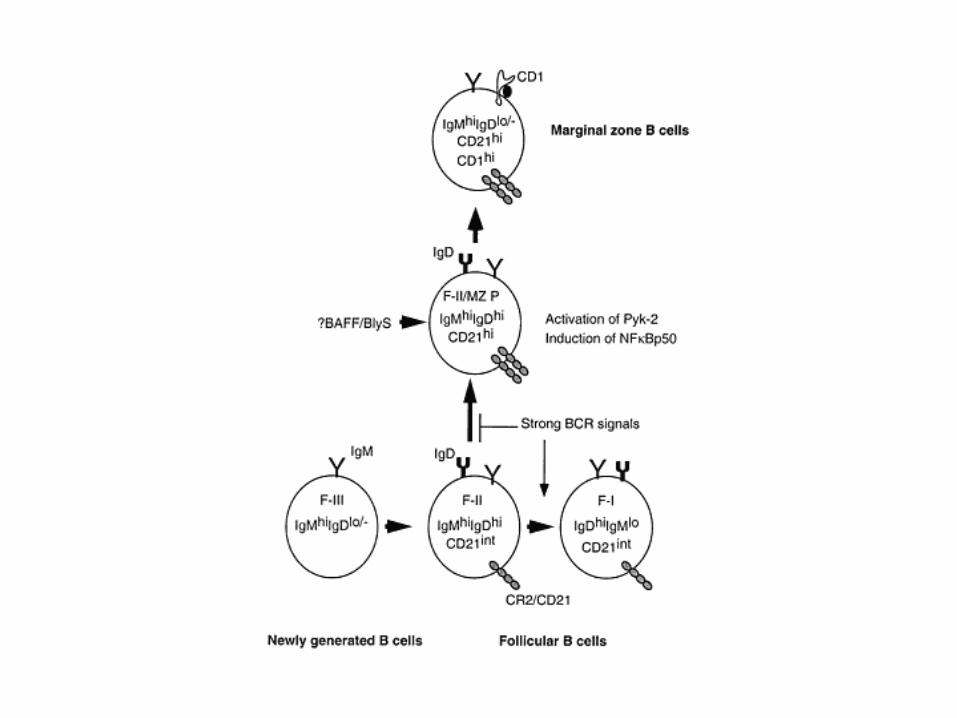
The BCR signal-strength model for lineage commitment in peripheral B cells
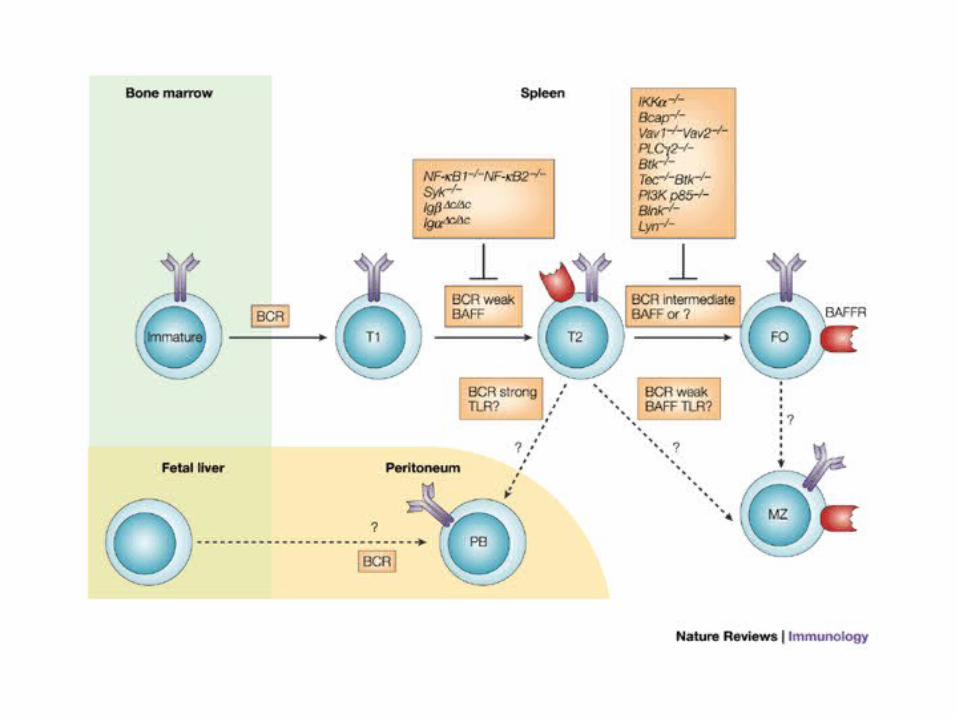

Figure 3 | Development of transitional and mature B cells.
Basal (weak) B-cell receptor (BCR) signals are required for the transition from transitional type 1 (T1) to T2 B-cell stages. Disruption of proximal BCR signalling components — the cytoplasmic tails of immunoglobulin-associated protein- (Ig) or Ig, and SYK — blocks this transition. BAFF (B-cell-activating factor of the tumour-necrosis-factor family) is also required for the maintenance of T2 cells. For the transition from T2 to follicular (FO) B-cell stages, intermediate levels of BCR signals are required. Disruption of some BCR signalling components leads to a block at this stage. Again, BAFF might be required for the maintenance of follicular B cells. A weak BCR signal and a stronger BCR signal are required for the generation of marginal-zone (MZ) B cells and peritoneal B cells, respectively (see also Table 1). BAFF might be required for the maintenance of MZ B cells, but not peritoneal B cells. The Toll-like receptor (TLR) signals that are provided by blood-borne and commensal pathogens might regulate the survival of MZ B cells also. BAFFR; BAFF receptor; BCAP, B-cell adaptor for PI3K; BLNK, B-cell linker; IKK, inhibitor of NF-B (IB) kinase; NF-B, nuclear factor-B; PB cell, peritoneal B cell; PI3K, phosphatidylinositol 3-kinase; PLC, phospholipase C.