activación del sistema plasminógeno-plasmina y el...
TRANSCRIPT

171
Manuel Nolasco, Mauricio Salcedo y Guelaguetza Vázquez-Ortiz
Laboratorio de Oncología Genómica, Unidad de Investigación Médica en Enfermedades Oncológicas,UMAE Hospital de Oncología, CMN Siglo XXI-IMSS, México.
Abstract•
Cells’ interaction with their environment is essential for the integrity and proper function of tissues, this is mainly due to the fact that tissues are not only composed by cells but also by a complex net of proteins called extracellular matrix (ECM). ECM is a dynamic structure, which is in constant remodeling (degradation and synthesis of components). This remodeling depends on several mechanisms, such as the activation of the plasminogen/plasmine system that participates in the degra-dation of some ECM components like fibrin and collagen. An important component of this system is PAI-1 protein, which regulates this system suppressing plasmine and by this, the ECM degradation is avoided. The alteration of this system is associated with several pathogenic processes like fibrosis, thrombosis and atherosclerosis, among others. Besides this, PAI-1 and uPA proteins participate in some cancer develop-ment. PAI-1 acts in several ways; it can inhibit ECM degrada-tion, and also acts in other processes like angiogenesis and cell migration. In this manuscript, we review the participation of PAI-1 in different pathologies, the aspects that allows this protein to participate in such processes, and also the way by which PAI-1 interacts with specific signaling pathways like MAPK, FAK, JAK-STAT, among others.
Key words: PAI-1, Extracellular Matrix, Cancer
Resumen•
LAS INTERACCIONES DE LAS CÉLULAS con su entorno son fundamentales para el buen funcionamiento e integridad de los teji-dos, ya que éstos no solo están conformados
por células sino que además contienen una red de componentes proteicos denominada matriz extrace-lular (MEC). La MEC es una estructura dinámica que está en constante remodelación (síntesis de nuevos componentes y degradación). Esta remodelación depende de varios mecanismos, como el sistema de activación plasminógeno/plasmina, el cual participa en la degradación de algunos componentes de la MEC, como la fibrina y algunas colágenas. Un componente importante de este sistema es la proteína PAI-1, en-cargada de regular dicho sistema, inhibiendo a la plas-mina y, consecuentemente, evitando la degradación de la MEC. Algunas alteraciones en la regulación de este sistema se asocian a diversos procesos patoló-gicos como fibrosis, trombosis y aterosclerosis, entre otras. Además, se ha encontrado que proteínas como PAI-1 y uPA participan en diferentes tipos de cáncer. PAI-1 es una proteína muy versátil, ya que además de inhibir la degradación de la MEC, interviene en otros procesos como angiogénesis y migración celular. Aquí se revisan las implicaciones de PAI-1 en diferentes pa-tologías, así como los aspectos que influyen para que esta proteína participe en ellos; así mismo se revisa la manera en que PAI-1 interactúa con varias vías de señalización como las MAPK, FAK, JAK-STAT, entre otras, y sus posibles repercusiones.
Palabras Clave: PAI-1, Matriz Extracelular, Cáncer.
Nolasco et al, Cancerología 2 (2007): 171-183
Correspondencia a:Dra. Guelaguetza Vázquez OrtizLaboratorio de Oncología GenómicaUnidad de Investigación Médica en Enfermedades OncológicasUMAE Hospital de OncologíaCentro Médico Nacional Siglo XXI-IMSSAv. Cuauhtémoc 330. Col Doctores. México D. F. C. P. 06720Tel. 56276900 Ext: 22708e-Mail: [email protected]
Activación del Sistema Plasminógeno-Plasmina
y el Papel de PAI-1en Patologías Humanas

172 Activación del Sistema Plasminógeno-Plasmina
Introducción•
Las interacciones entre las células son parte fun-damental de las entidades multicelulares, esto, con el fin de mantener el equilibrio y el buen funcionamiento del organismo. Estas interaccio-nes implican a una gran cantidad de moléculas que tienen diferentes funciones de acuerdo al tipo de tejido que conforman y a su ubicación dentro de la célula. Un tejido está conformado principalmente por células pero también por la matriz extracelular (MEC) que sirve de soporte para las células y además le da la conformación al tejido. La MEC está constituida por una gran variedad de moléculas como son: a) las coláge-nas, que son proteínas fibrilares, b) los proteo-glicanos, que son macromoléculas con un com-ponente proteico y que además están unidas a disacáridos sulfatados, c) las proteínas elásticas, como la elastina que proporciona cierta elas-ticidad y flexibilidad a la MEC, y por último d) las estructuras glicoproteicas, como son la fibronectina y laminina entre otras. Todos los componentes anteriores pueden unirse entre si para formar una red que le da conformación a la MEC. (1)También existen otras moléculas que pueden pasar a través de la MEC, sin embargo, éstas no se consideran parte de la misma; por ejemplo: algunos factores de crecimiento o pro-teínas encargadas de remodelación (las cuales se mencionarán más adelante). En este escrito se describirán algunos de los mecanismos para la regulación de la remode-lación de la matriz extracelular poniendo un énfasis especial en el sistema de activación pla-minógeno/plasmina, así como también en su mecanismo de regulación, en el cual participa el inhibidor del activador del plasminógeno (PAI-1). La proteína PAI-1 es importante ya que inhibe la degradación de la MEC mediada por plasmina, además de estar involucrada en un gran número de patologías como son diabe-tes tipo 2, trombo-embolia, hiperinsulinemia, hipertensión, hipertriglicerinemia, obesidad, fibrosis epitelial y cáncer.(2-7)
Matriz Extracelulare Interacción con Células•
La MEC es un complejo insoluble de proteínas y carbohidratos producidos por todas las células de los metazoarios, en donde a su vez las células se en-cuentran embebidas sirviéndoles de sostén y dándo-le conformación al tejido. Las proteínas más impor-tantes que la componen, incluyen a las colágenas, las fibras elásticas, y algunas proteínas de adhesión como son: la fibronectina y la laminina entre otras. Mientras que los carbohidratos presentes incluyen a los proteoglicanos y los glucosaminglicanos.
Las células pueden responder a su entorno me-diante la interacción de sus componentes con lo que se encuentra a su alrededor. La célula con-tiene receptores membranales para componentes extracelulares, de tal manera que, dependiendo del tipo de tejido, la MEC se formará de diferentes componentes y proporciones de los mismos. Por ejemplo, la MEC de un tejido de cartílago contie-ne mayoritariamente fibras elásticas como elasti-na, mientras que la MEC de membranas básales contiene principalmente colágena del tipo IV. En el Cuadro 1 se muestran algunos de los componen-tes generales representativos de la MEC. Los com-ponentes de la MEC que son fibras de colágena y elásticas, fibronectina, vitronectina entre otros, pueden interaccionar entre si, proporcionando ca-racterísticas físicas al tejido y la manera en la que estos influyen al interaccionar con la célula.(1-8)
Por ejemplo, las fibras de colágena tienen una función estructural en la MEC y en el ser humano existen 13 tipos diferentes. Las fibras elásticas están conformadas principalmente por: a) elastina (pre-
Proteínas Distribución Principal
Colágena tipo
Fibras Elásticas Fibronectina
Laminina
IIIIV
Piel, Hueso, Tendones, CórneaNotocorda EmbrionariaMembranas BasalesLigamentos Elásticos, Pulmón, PielSangre (Dímero Soluble) Membranas Básales
Cuadro 1•Componentes de la matriz extracelular y su localización
173Nolasco et al, Cancerología 2 (2007): 171-183
MMPs
Pro-MMPs
ECM
Factores deCrecimiento Activados
Factores deCrecimiento Latentes
Plasmina
Plasminogeno
Pro-uPA uPA
PAI1
uPAR
JAK
FAK
SRCSHC
ActinaPaxilina
Vinculina
Talina
Sobrevivencia
STAT
STAT
AP1
MAPK PI3K AKT
RAS
GRB2
Transcripción
uPAR
α-2-AP
α5 β1 Sitio de la Unióna Fibrina
Sitio de la Unióna Colágena
Sitio de la Unióna Integrinas
Fibronectina
senta una mayor proporción), b) proteínas microfi-brilares, c) lisil oxidasa y d) proteoglicanos.
La fibronectina, que contribuye en adhesión y mi-gración celular, tiene algunos dominios de unión a diferentes tipos de colágenas como la tipo I; además de interaccionar con receptores celulares de mem-brana permitiendo que el dinamismo entre la MEC y las mismas células se vuelva aún más complejo.
Remodelación de laMatriz Extracelular•
La MEC es una estructura dinámica lo cual impli-ca un constante recambio de las proteínas y ele-mentos que la conforman. Durante los procesos de diferenciación en el desarrollo embrionario o en procesos de migración celular, la remodela-ción se da de acuerdo a los requerimientos ce-lulares. En tejidos adultos dicha remodelación ocurre en la neovascularización así como en los cambios que sufren las glándulas mamarias y el tejido uterino durante el ciclo menstrual.(9)
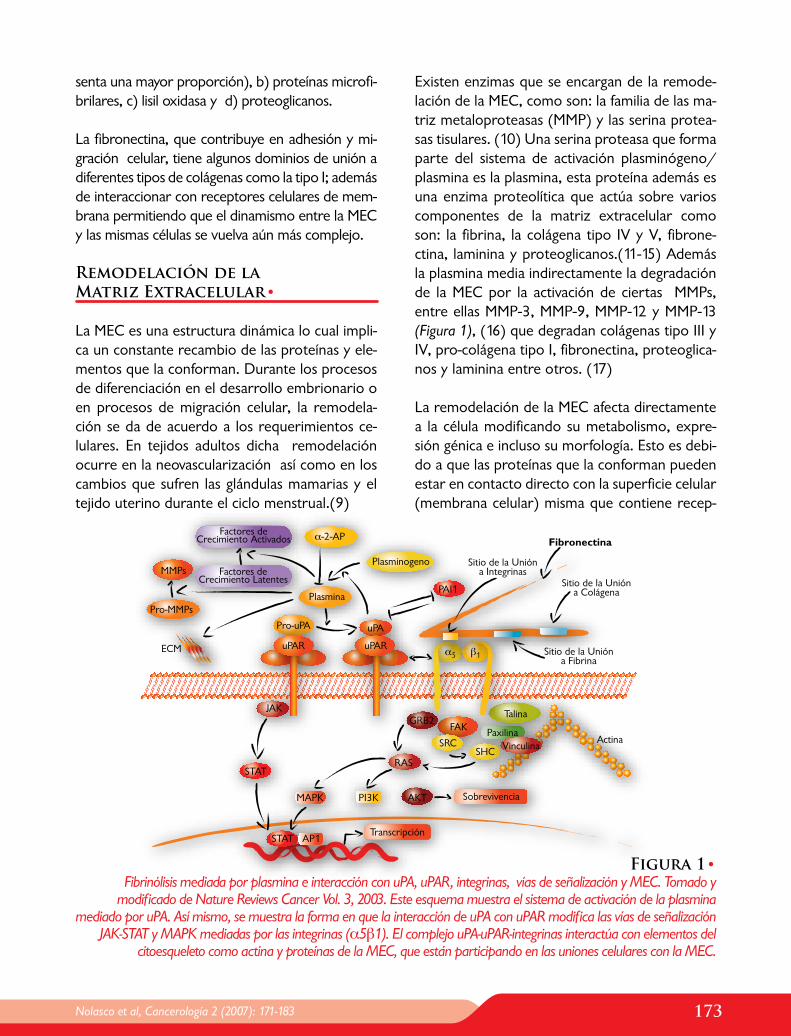
Figura 1•Fibrinólisis mediada por plasmina e interacción con uPA, uPAR, integrinas, vías de señalización y MEC. Tomado y
modificado de Nature Reviews Cancer Vol. 3, 2003. Este esquema muestra el sistema de activación de la plasmina mediado por uPA. Así mismo, se muestra la forma en que la interacción de uPA con uPAR modifica las vías de señalización
JAK-STAT y MAPK mediadas por las integrinas (α5β1). El complejo uPA-uPAR-integrinas interactúa con elementos del citoesqueleto como actina y proteínas de la MEC, que están participando en las uniones celulares con la MEC.
Existen enzimas que se encargan de la remode-lación de la MEC, como son: la familia de las ma-triz metaloproteasas (MMP) y las serina protea-sas tisulares. (10) Una serina proteasa que forma parte del sistema de activación plasminógeno/plasmina es la plasmina, esta proteína además es una enzima proteolítica que actúa sobre varios componentes de la matriz extracelular como son: la fibrina, la colágena tipo IV y V, fibrone-ctina, laminina y proteoglicanos.(11-15) Además la plasmina media indirectamente la degradación de la MEC por la activación de ciertas MMPs, entre ellas MMP-3, MMP-9, MMP-12 y MMP-13 (Figura 1), (16) que degradan colágenas tipo III y IV, pro-colágena tipo I, fibronectina, proteoglica-nos y laminina entre otros. (17)
La remodelación de la MEC afecta directamente a la célula modificando su metabolismo, expre-sión génica e incluso su morfología. Esto es debi-do a que las proteínas que la conforman pueden estar en contacto directo con la superficie celular (membrana celular) misma que contiene recep-

174 Activación del Sistema Plasminógeno-Plasmina
tores “específicos” para estas proteínas. Activan-do o desactivando vías de señalización que con-secuentemente activan o desactivan factores de trascripción, cambiando la expresión.
Sistema de ActivaciónPlasminógeno/plasmina•
La fibrinólisis o degradación de las fibras, es la lisis o remodelación de los componentes de la MEC que se genera mediante una cascada de eventos proteo-líticos. Debido a que el sistema plasminógeno/plas-mina participa en la degradación de la MEC tanto en tejidos “sólidos” como en el sistema sanguíneo, puede decirse que este sistema participa en la fibri-nólisis. Este sistema también interviene en algunos otros procesos biológicos como la regeneración ti-sular, menstruación e implantación embrionaria así como en la invasión tumoral.(18)
Activación de la Plasminaa Partir del Plasminógenopara la Remociónde Coágulos Sanguíneos•
Al existir lesiones en el endotelio del sistema vas-cular se activa el sistema de coagulación, el coágulo formado principalmente por fibrina debe ser poste-riormente degradado por la plasmina. La plasmina
Plasminógeno Plasmina A-2-AP
tPA
FX Fxa / FVa
Factor TisularFVIIa
Pro-Trombina
Trombina
Fibrina
Trombo
Fibrinógeno
tPA
a) Fibrinólisis
Agregación dePlaquetas
Producto deDegradación
b) Cascada de Coagulación
deriva de otra proteína denominada plasminógeno que de forma natural está en el plasma sanguíneo.
El plasminógeno es activado por los PAs (activa-dores del plasminógeno), entre ellos la serina proteasa uPA (activador del plasminógeno tipo urocinasa) y tPA (activador del plasminógeno tipo tisular), los cuales efectúan una proteólisis parcial de la proteína para convertirla en plasmi-na activa.(19) La degradación de fibrina durante los procesos de remoción de coágulos sanguí-neos es llevada a cabo principalmente por tPA, mientras que uPA degrada la MEC en tejidos sólidos.(6) (19) Por lo tanto, la activación del plasminógeno se lleva a cabo bajo dos esquemas, uno en donde principalmente actúa tPA y en el otro uPA (ver Figuras 1 y 2).
De manera natural, tPA en el plasma sanguíneo tie-ne una actividad enzimática débil, pero esta activi-dad aumenta cuando existe la presencia de fibrina, es decir de un coágulo.(20) Esta actividad aumenta porque tPA tiene un domino de unión especifico a fibrina (uPA no lo tiene), el cual se puede unir a residuos de lisina de la región carboxilo terminal, haciendo que la concentración de tPA aumente en el sitio donde se encuentra el coágulo. También, el plasminógeno tiene afinidad por la fibrina, y enton-ces la presencia de ambas proteínas hace que la
Figura 2•Proceso de fibrinólisis mediado por plasmina y cascada de coagulación. En el panel a) podemos observar el
esquema de la activación del plasminógeno y degradación del trombo mediado por la plasmina, este sistema está regulado por PAI-1, α-2-AP y TAFI. En donde, PAI-1 evita la generación de plasmina mediante la inhibición de
tPA, α-2-AP y se unen a la plasmina y evita que ésta degrade la fibrina. En el panel b) se muestra la activación de la cascada de coagulación que favorece a la formación del coágulo de fibrina y a la agregación plaquetaria.

175Nolasco et al, Cancerología 2 (2007): 171-183
concentración de plasmina aumente en el sitio y se facilite la remoción del coágulo.(21) Activación de la Plasminaa Partir del Plasminógenopara la Degradaciónde la MEC en Tejido Sólido•
En el segundo esquema, el plasminógeno se en-cuentra en la MEC de los tejidos “sólidos” y el activador es uPA. Este activador es inicialmente sintetizado como una proteína de una sola cade-na (scuPA o pro-uPA) que presenta una actividad enzimática baja, scuPA forma un complejo con el receptor del activador del plasminógeno tipo urocinasa uPAR que es un receptor transmembra-nal, al cual se le denomina scuPA-uPAR. Una vez que este complejo es formado algunas proteínas como plasmina, calicreina, tripsina, factor de coa-gulación XIIa y catepsina digieren parcialmente al complejo convirtiendo a scuPA en una proteína de dos cadenas polipeptídicas. Incrementándose así, la actividad proteolítica o enzimática del complejo y a esta nueva conformación se le denomina com-plejo uPA-uPAR (Figura 1).(22)
El complejo uPA-uPAR permite que el plasminó-geno sea convertido a plasmina por uPA y que la plasmina a su vez degrade componentes de la MEC como las fibras de colágena y algunos inhi-bidores de las metaloproteasas (TIMPs).(23)
Hasta este punto los dos tipos de activadores del plasminógeno tPA y uPA realizan una función muy similar en diferentes tipos de tejido, sin embargo, estructuralmente presentan algunos dominios di-ferentes que les confieren una funcionalidad dis-tinta. La proteína uPA tiene tres dominios: a) el dominio N-terminal que se denomina dominio de factor de crecimiento (GFD) y que además es el interactúa con uPAR (24;25), b) el dominio proteolítico (C-terminal) y c) el dominio Kringle, del cual aún no se conoce claramente su función. Por otro lado, tPA cuenta con dominios similares y algunos adicionales, uno de los cuales le permi-te interaccionar con fibrina.(21)
La degradación de la MEC mediada por plasmina tiene dos puntos de control, uno es la inactiva-ción de plasmina mediante la unión de las proteí-nas α-2-AP (α-2- antiplasmina) o TAIF (inhibidor de la fibrinólisis activado por trombina) que blo-quean la actividad proteolítica de esta proteína. El otro punto de control son los inhibidores: PAI-1, PAI-2 y PAI-3 (Inhibidor del activador del plas-minógeno 1, 2 y 3 respectivamente) y la protea-sa nexina.(26) En particular se ha descrito que PAI-1 mantiene la homeostasis, regulación de la fibrinólisis, angiogénesis y metástasis. (27, 28)
Vías de Señalización Mediadaspor la Interacción UPA-UPARen Tejidos Sólidos•
La formación del complejo uPA-uPAR activa a algunas vías de señalización intracelulares involu-cradas en la regulación de la migración y prolife-ración celular como: JAK-STAT y MAPK.(29-31) Estas señales contribuyen de manera natural a la regulación de varias funciones fisiológicas como: angiogénesis, implantación embrionaria, reacción inflamatoria y cicatrización cutánea.(29)
La vía de señalización JAK-STAT se activa por la unión del receptor a scuPA o pro-uPA inde-pendientemente de la activación del complejo uPA-uPAR (Figura 2). JAK1 cinasa se asocia con uPAR llevando a la fosforilación y dimerización del factor de transcripción STAT, lo que activa diferentes rutas de señalización.
Otra forma en que las vías de señalización son activadas es mediante la interacción de uPAR con algunas integrinas principalmente αVβ3, αVβ5, α3β1 y α5β1 que son receptores de vitronec-tina y fibronectina respectivamente y que están involucrados en migración celular, remodelación tisular, crecimiento celular y tumorigénesis.(32)
También el complejo uPA-uPAR interacciona con la vía de señalización de FAK (focal adhesion ki-nase) que participa en la organización del citoes-queleto, esta interacción es mediada por inte-

176 Activación del Sistema Plasminógeno-Plasmina
grinas (33-36). Además de que la interacción de uPAR con integrinas permite que se provoquen cambios en la organización del citoesqueleto, ya que las integrinas interactúan con componentes del mismo, como la actina, talina, vinculina entre otras. Además de que estas integrinas interac-túan con componentes de la MEC.
El complejo uPA-uPAR está implicado en la altera-ción de diferentes vías de señalización, las cuales, pueden llevarnos al cambio de la expresión génica y en el caso concreto de la interacción con algu-nas integrinas como la α5β1 y αVβ3. Esta inte-racción además altera a la MEC ya que cuando el complejo uPA-uPAR interacciona con los recepto-res αVβ3 y α5β1 se afecta la unión de la célula a la vitronectina y fibronectina respectivamente, causando un cambio en la arquitectura del tejido.
Inhibición de UPA por PAI-1•
PAI-1 juega un papel muy importante en varias funciones celulares como angiogénesis, homeos-tasis e invasión de células tumorales. Esta proteí-na es el Inhibidor del Activador del plasminógeno Tipo 1 y es el principal inhibidor de uPA y tPA. La interacción PAI-1-uPA provoca la inhibición de la actividad de proteasa de uPA, por lo tanto no transforma el plasminógeno en plasmina y se in-hibe la fibrinólisis o la degradación de MEC.
El hecho de que PAI-1 se una a uPA para formar un complejo uPAR-uPA-PAI-1 influye en la altera-ción de las vías de señalización en la cuales está implicado uPAR (PAI-1 también inhibe a uPA en estado libre). Al formarse este complejo, se in-ternaliza en la célula por la proteína relacionada con el receptor de lipoproteínas de baja densi-dad LRP, PAI-1, entonces, induce el decremento en la superficie celular de uPAR y por lo tanto interrumpe las vías de señalización así como la organización del citoesqueleto y MEC que son mediadas por uPAR.(36-39)
PAI-1 puede inhibir las uniones célula-célula me-diada por uPAR y vitronectina, mediante la unión
de vitronectina-PAI-1 evitando la unión de vitrone-ctina-integrina. La vitronectina también interactúa con el complejo uPA-uPAR para formar un nuevo complejo denominado uPA-uPAR-vitronectina de tal manera que PAI-1 puede evitar la formación de este complejo uniéndose tanto a vitronectina (vi-tronectina-PAI-1) como a uPA (uPAR-uPA-PAI-1) y por lo tanto se interrumpe la activación de las vías de señalización MAPK y FAK que están mediadas por uPAR-vitronectina e integrinas. Así mismo, se pueden perder contactos focales entre la célula y la MEC mediadas por las integrinas y en donde también interviene tanto vitronectina como fibro-nectina, lo cual permite inferir la importancia de PAI-1 las vías de señalización y las interacciones de la célula con la MEC. (40-43)
Como ya se mencionó anteriormente, el comple-jo uPA-uPAR interactúa con varias integrinas entre las cuales están α3β1, α5β1 que son receptores de fibronectina. Czekay y colaboradores mencio-nan que PAI-1 disocia a las células de la MEC me-diante la inactivación de integrinas y la unión de PAI-1 con uPA y no con la interacción PAI-1-vitro-nectina. También observaron que la presencia de uPA y PAI-1 induce la disociación entre la células y los componentes de la MEC.(44)
Características de PAI-1•
PAI-1 es una glicoproteína de 50 kDa que está conformada por 359 aminoácidos, pertenece a la superfamilia de las serpinas (inhibidoras de serina protesas) que la conforman aproximada-mente 40 miembros cuyas funciones principales son de inhibición.(45) El gen que codifica a esta proteína se denomina serpine1 y se localiza en el brazo largo del cromosoma 7 (7q21.3-q22). Este gen está constituido por 15 exones y puede pre-sentar cinco isoformas (www.genecards.org).
PAI-1 presenta tres conformaciones diferentes, las cuales además son ínterconvertibles entre sí: a) una conformación activa con propiedad inhibitoria (forma un complejo covalente estable con la pro-teína uPA), b) una conformación de sustrato, y c)

177Nolasco et al, Cancerología 2 (2007): 171-183
una conformación latente no reactiva. Inicialmen-te PAI-1 es sintetizada en su conformación activa pero cambia espontáneamente a su conformación latente con una vida media de 1 a 2 h. (46-48)
Regulación de PAI-1•
PAI-1 está regulado directamente por las proteínas involucradas en la remodelación de la MEC, en la Figura 3 se muestra de forma resumida las vías de señalización que están implicadas en la regulación de la expresión del gen SERPI-NE1 y por ende de la proteína PAI-1.
En patologías como el cáncer, las células cambian su morfología cambiando las interacciones con su entorno, tanto en los contactos célula-célula como en las interacciones célula-MEC. En cáncer de mama se ha visto que los cambios en la estruc-tura del citoesqueleto se dan por la interrupción de los contactos célula-célula dependientes de E-caderina y esta pérdida de contacto estimula la expresión de uPA y PAI-1.(49) Además de que la expresión de PAI-1 es regulada por la ruptura de microfilamentos y microtúbulos del citoesquele-to,(50) por lo cual el cambio morfológico de la célula permite cambios en su expresión.
Otros factores que regulan la expresión de PAI-1 es TGF- β1, (factor de crecimiento transforman-te β1) (ver Figura 3), TNF (factor de necrosis tumoral), insulina/proinsulina, y la angiotensina II; así como ácidos grasos libres y triglicéridos.
Con respecto a TGF- β1, la regulación de PAI-1 se puede dar de dos formas: 1) la activación me-diante la vía de señalización Smad (el promotor de PAI-1 tiene tres sitios de unión para los facto-res de trascripción Smad3 y Smad4), y b) por la ruta MAPK a través de p38, aunque TNF-β también puede activar la vía p38. (51, 52).
Se han descrito dos receptores para la angioten-sina II, el receptor tipo 1 y el tipo 2, mismos que tienen características de receptores acoplados a proteína G, con siete dominios trans-membra-
Perturbación delCitoesqueleto y
Contactos Celulares
Vía deSeñalización SMAD(SMAD4 y SMAD3)
Vía deSeñalización MAPK
(p38)
Promotor Serpine1
Vía deSeñalización
MAPK MedianteMER/ERK
SeñalizaciónMediada por Vía
JNK
TGF-β1
PAI-1
Angiotensina II
Figura 3•Vías de señalización involucradas en la expresión de PAI-1
nales. La unión de angiotensina II a su receptor, promueve la activación de varios factores y vías de señalización mediante la fosforilación, uno de estos factores es la proteína PKC (proteína cinasa C). Se ha observado que la angiotensina II induce
la síntesis de MEC, así como la expresión de PAI-1 promueve la acumulación de MEC mediante la in-hibición de la degradación de ésta, cabe mencio-nar que la expresión de esta última está mediada por la proteína PKC, (53;54) ya que SERPINE1 tiene sitios de reconocimiento para los factores de trascripción Sp1 y Sp3 y estos factores pueden ser activados mediante PKC(55;56)
La Proteína PAI-1en las Patologías•
La proteína PAI-1 se ha visto involucrada en un gran número de patologías, de ahí que su estudio esté cobrando gran importancia actualmente. El efecto que esta proteína puede generar es muy variable ya que puede tener diferentes funciones dependiendo de las patologías estudiadas.(57, 58)
Aterosclerosis, InfartoAl Miocardio y Trombosis Venal•
La sobre-expresión de PAI-1 se ha visto relacio-nada con las enfermedades vasculares. En mo-delos murinos, el incremento de esta proteína da como resultado la inhibición excesiva de uPA

178 Activación del Sistema Plasminógeno-Plasmina
y tPA perdiendo la eficiencia en la degradación de los trombos o coágulos generando entonces trombosis venal y un alto riesgo potencial en pa-decer un infarto al miocardio. (59)
PAI-1 y el Síndrome de Resisten-cia a la Insulina y Obesidad•
La obesidad es un padecimiento de los países desarrollados. En obesos la proteína PAI-1 tiene niveles altos en el plasma sanguíneo, ya que los adipositos sintetizan a esta proteína y esto ha co-rrelacionado con los tratamientos para bajar de peso en donde los niveles de esta proteína bajan.
Los pacientes con síndrome de resistencia a insulina se caracterizan entre otras cosas por padecer obe-sidad, un incremento en niveles de ácidos grasos libres y triglicéridos, además de hipertensión, hipe-rinsulinemia e hipofibrinólisis. En modelos murinos y en seres humanos se ha identificado que TNF y TGF-β está incrementado en tejido adiposo(59;60) y éstos en conjunto con la insulina/proinsulina, triglicéridos y ácidos grasos libres estimulan la ex-presión de PAI-1.(61-63). Por otro lado, se ha de-mostrado que la hipofibrinólisis es causada por un incremento de PAI-1 y que las dietas hipocalóricas y el ejercicio físico tienen como resultado una baja en los niveles de PAI-1 en este síndrome.(64)
PAI-1 en la Fibrosis y el Asma•
Como se ha venido mencionado, PAI-1 juega un papel fundamental en el control de la fibrinólisis mediante la inhibición de los PAs. Esta proteína es expresada por varios tipos celulares como las células endoteliales, hepatocitos y adipositos entre otros.(65-67)
Oh y colaboradores observaron que al inducirse la expresión de PAI-1 en pulmón se genera un acu-mulamiento o deposición de proteínas fibrilares generándose una fibrosis epitelial.(68) En ratones transgénicos con fibrosis pulmonar inducida se ha identificado que esta enfermedad es más agresiva en los ratones que sobre-expresan PAI-1 compara-
da con los que no la presentan.(69) Sin embargo, existen reportes contradictorios que indican que en ratones con deficiencia de PAI-1 se presenta una resistencia a desarrollar fibrosis pulmonar.(70)
Con respecto a la fibrosis renal, se ha reportado que la proteína PAI-1 está sobre-expresada y esta sobre-ex-presión también se presenta en la glomerulosis.(71)
En seres humanos con asma se ha visto que los mastocitos sobre-expresan PAI-1(72) y en mode-los murinos con asma crónica, esta proteína pro-mueve la acumulación de componentes de la MEC en las vías aéreas, además de que sus niveles están incrementados en fluido bronco-alveolar. (68) Como podemos observar PAI-1 está involucra-da en muchos mecanismos moleculares, por lo tanto la alteración del control y la expresión de esta proteína puede afectar de diferentes mane-ras la homeostasis del organismo, de ahí que sea importante estudiar y tratar de descifrar los me-canismos por los cuales esta proteína es activada así como los mecanismos y vías de señalización que pueden ser afectadas por ésta.
PAI-1 en la Angiogénesis y Cáncer•
La angiogénesis tanto en procesos normales (neovascularizacion) como en patológicos (tu-morales), requiere que las células endoteliales de los vasos y capilares migren para formar los nuevos vasos sanguíneos y para llevar a cabo esto, el sistema de activación plasminógeno/plasmina está implicado tanto en la degradación de la MEC, para que las células puedan migrar, como en la inhibición de la degradación de la misma, mediante PAI-1, para que se puedan es-tablecer los nuevos vasos sanguíneos.
La hipoxia es uno de los factores que inducen angio-génesis y bajo estas condiciones se ha reportado que las células endoteliales presentan un incremento en la expresión de uPA y PAI-1, además de otros factores angiogénicos como VEGF y bFGF que promueven la expresión de uPA, tPA, uPAR y PAI-1.(73)

179Nolasco et al, Cancerología 2 (2007): 171-183
El papel que PAI-1 juega dentro de la angiogé-nesis puede verse de dos maneras; la primera se basa en la capacidad para inhibir la fibrinólisis de la MEC y la otra, implica el rompimiento de los contactos célula-célula y célula-MEC, permi-tiendo la migración celular.
En varios estudios in vitro e in vivo, se ha visto que la sobre-expresión de PAI-1 reduce la metástasis y el crecimiento del tumor. Por ejemplo, en carcinoma de próstata la sobre-expresión de PAI-1 inhibe tanto el crecimiento como la angiogénesis en el tumor.(74) También, esta proteína inhibe la angiogénesis en un ensayo CAM (Chick Chorioallantoic Membrane), in-dicando que estos procesos son debidos a la inhibi-ción de la degradación de la MEC.(75)
Sin embargo, existen datos contradictorios en los que se observa que la presencia de PAI-1 es nece-saria para que se pueda dar la angiogénesis. En un modelo know-out murino para PAI-1 se reporta una reducción significativa del volumen de tumor así como en la vascularizacion.(76, 77) Así mismo, se ha observado que en líneas celulares derivadas de cáncer de pulmón el proceso metastático se incrementa al administrar PAI-1 endógeno.(78)
Como se vio anteriormente, existen reportes contradictorios sobre la inhibición o activación de PAI-1 sobre procesos de angiogénesis y la metástasis. Con respecto a estos resultados, McMahon y colaboradores utilizaron diferentes concentraciones de PAI-1 en un modelo in vivo y observaron que a bajas concentraciones de la proteína se promueve la angiogénesis, mientras que a concentraciones altas ésta se inhibe.(79)
Cáncer Gástrico•
En tejidos de cáncer gástrico se han encontra-do niveles altos de PAI-1 así como de uPA y ésto se ha relacionado con una baja sobrevida, en contraste, existen otros trabajos donde se indica que los niveles de estas proteínas no se ven afectados. Por otro lado, en mucosa gástrica de controles sanos, la actividad de tPA es baja
en comparación con pacientes con cáncer gás-trico asociándose también con una baja sobrevi-da.(80-82) Tanto PAI-1 como uPA son factores de pronóstico independientes y parecen estar asociados a la diferenciación del tumor en este tipo de cáncer.
Carcinoma Colorectal•
Al comparar muestras de tejido de colon nor-mal con muestras tumorales se han encontrado diferencias en los niveles de proteicos de PAI-1 y uPA, reportándose niveles más altos en los tumo-res. También se han encontrado niveles altos de uPAR que correlacionan con metástasis e indican una baja sobrevida, y su presencia es un factor pronóstico independiente.(83, 84) Así mismo, se ha visto que tanto PAI-1 como uPA son factores de pronóstico independientes.(85;86)
Cáncer de Esófago•
En tejidos de adenocarcinoma de esófago los ni-veles de las proteínas uPA y PAI-1 también se ven incrementados, lo cual ha sido relacionado con el grado del tumor, (87) diagnóstico y sobre-vida desfavorable.(88)
Cáncer de Pulmón•
En el cáncer de pulmón aunque los niveles de las proteínas uPA, uPAR y PAI-1 son más altos en comparación con el tejido de pulmón normal, los pronósticos de sobrevida cambian dependien-do del tipo de células afectadas. En el carcinoma de pulmón de células no pequeñas (NSCLC) no se ha encontrado relación entre los niveles de estas proteínas y la sobrevida, sin embargo, en adenocarcinoma y cáncer de pulmón escamoso el incremento de PAI-1 y uPAR está relacionado con una baja sobrevida. (89-91)
Cáncer de Ovario•
Así como en los tipos tumorales antes mencio-nados, en cáncer de ovario se han encontrado

180 Activación del Sistema Plasminógeno-Plasmina
niveles altos de PAI-1 y proteínas involucradas en el sistema de activación plasminógeno/plasmina, principalmente con las proteínas PAI-1 y uPA. Te-cimer y colaboradores identifican que niveles altos de PAI-1 pueden relacionarse a una recurrencia del tumor además de que su presencia también correlaciona con una baja sobrevida. (92, 93)
Cáncer de Mama•
En este tipo de cáncer Janicke y colaboradores, en 1993, reportaron niveles altos de PAI-1 y que ésto funciona como un factor independiente de mal pronóstico. Así mismo, los niveles altos de PAI-1 se han relacionado con cáncer de tipo más agresivo (nódulo positivo y negativo).(94)
Por otro lado, Leissner encontró que los niveles elevados PAI-1 en cáncer de mama representan un factor pronóstico independiente para el desa-rrollo de metástasis y una menor sobrevida.(95) así como un mayor riesgo de regresión.(96) Sin embargo, los pacientes con niveles elevados de PAI-1 responden mejor a la quimioterapia en comparación con aquellos que tienen niveles mas bajos de esta proteína.(97)
Algunos estudios de inmunolocalización sugieren que PAI-1 es sintetizado como un mecanismo de protección hacia el tumor ya que al inhibir uPA se evita la degradación de la MEC y por lo tanto la migración. (96) Por otro lado se ha sugerido que la producción de PAI-1, está directamente relacionada con angiogénesis y la neovasculariza-ción a favor del tumor. (94, 98)
La interacción de uPAR-uPA-PAI-1-LRP induce el de-cremento de la migración celular en cáncer de mama, además de que la expresión de uPA y PAI-1 se ha iden-tificado en cáncer de mama in situ e in vivo (99)
Cáncer Cérvico-Uterino•
En cuanto a la concentración de las proteínas se ha encontrado que en extracto de muestras de cáncer cérvico-uterino la concentración del los
antígenos uPA y PAI-1 es mayor en comparación con las lesiones de bajo grado y normales. Es im-portante notar que aunque la concentración del antígeno PAI-1 está incrementada en el tumor, no se encontró un incremento significativo en la acti-vidad de esta proteína, ésto concuerda con lo re-portado por Kobayashi y colaboradores, quienes encontraron que la mayor parte de esta proteína se encuentra en su forma latente. (100, 101) Sin embargo, la sobre-expresión de PAI-1 también se ha identificado como un factor independiente para una baja en la sobrevida de las pacientes.(102)
Conclusiones•
La alteración en la expresión la proteína de PAI-1 puede llevar a la pérdida de las interacciones de la MEC con la célula, lo cual provoca cambios en las vías de señalización y en la morfología celular. Esta pérdida de homeostasis afecta los procesos biológicos del tejido y puede generar diferentes tipos de patologías, algunas las cuales están direc-tamente relacionadas con procesos de fibrosis, cáncer, problemas vasculares como aterosclerosis, infarto al miocardio, trombosis venal entres otras. En cuanto al cáncer, de manera general existen reportes contradictorios con respecto a la función inhibitoria que ejerce PAI- 1. Sin embargo, el au-mento en los niveles de esta proteína en diferentes tipos de cáncer tiene un gran impacto en relación a la sobrevida y a la posible agresividad de la mis-ma, además de que se ha visto que PAI-1 participa activamente en procesos de angiogénesis y migra-ción celular. Por todo lo anterior, es importante el estudio detallado de los factores que controlan la expresión y actividad de esta proteína.
Referencias•
1. Zern MA, Reid LM. Extracellular matrix, chemistry, biology and pathology with emphasis on the liver. 1993•2. Wiman B. Plasminogen activator inhibitor 1 (PAI-1) in plasma: its role in thrombotic disease. Thromb Haemost 1995; 74: 71-76•3. Paramo J, Alfaro M, Rocha E. Postoperative chan-ges in the plasmatic levels of tissue-type plasminogen activator and its fast-acting inhibitor-relationship to deep vein thrombosis and influence of prophylaxis. Thromb Haemost 1985; 54: 713-716•

181Nolasco et al, Cancerología 2 (2007): 171-183
4. Goto D, Fujii S, Kaneko T et al. Intracellular signal transduction modulating expression of plasminogen activator inhibitor-1 in adipocytes. Biochemical Phar-macology 2003; 65:1907-1914•5. Sakkinen PA, Wahl P, Cushman M et al. Cluste-ring of procoagulation, inflammation, and fibrinólisis variables with metabolic factors in insulin resistance syndrome. Am J Epidemiol 2000; 152: 897-907•6. Bodary P, Wickenheiser K, Eitzman D. Recent ad-vances in understanding endogenous fibrinolysis: im-plications for molecular-based treatment of vascular disorders. Expert Rev Mol Med 2002; 4:1-10•7. Sternlicht MD, Dunning AM, Moore DH et al. Prog-nostic Value of PAI1 in Invasive Breast Cancer: Evidence That Tumor-Specific Factors Are More Important Than Genetic Variation in Regulating PAI1 Expression. Cancer Epidemiol Biomarkers Prev 2006;15: 2107-2114•8. Yurchenco PD, Birk DE, Robert M. Extracellular Ma-trix Assembly and Structure. Academic Press ed. 1994•9. Streuli C. Extracellular matrix remodelling and cellular diffe-rentiation. Current Opinion in Cell Biology 1999; 11: 634-640•10. Streuli C. Extracellular matrix remodelling and cellular diffe-rentiation. Current Opinion in Cell Biology 1999; 11: 634-640•11. Gravilovicc J, Murphy G. The role of plasminogen activator in cell-mediated collagen degradation. Cell Biol Int 1989; 13: 367-375•12. Knudsen BS, Silverstein R, Leung LK. Binding of plasminogen to extracellular matrix. J Biol Chem 1986; 104: 1085-1096•13. Gordon AP, Strickland S. Interactions of heparin with plasminogen activator and plasminogen: effects on the acti-vation of plasminogen. Biochemistry 1986; 25: 4033-4040•14. Roldan A, Cubellis M, Masucci M. Cloning and ex-pression of the receptor for human urokinase plasmino-gen activator,a central molecule in cell surface, plasmin dependent proteolysis. EMBO J 1990; 9: 467-474•15. Ichinose A, Fujikawa K, Suyama T. The activation of prourokinase by plasma kallikrein and its inactivation by thrombin. J Biol Chem 1986; 261: 3486-3489•16. Carmeliet P, Moons L, Lijnen R et al. Urokinase-gene-rated plasmin activates matrix metalloproteinase during aneurysm formation. Nat Genet 1997; 17: 439-444•17. Sternlicht MD, Werb Z. How Matrix Metalloproteinases regulate cell behavior. Annu Rev Cell Dev 2001; 17: 516•18. Liu Y. Regulation of the plasminogen activator sys-tem in the ovary. Biol Signals Recept 1999; 8: 160-177•19. Andreasen P, Kjoller L, Christensen L et al. The urokinasa type plasminogen activator system in can-cer metastasis: a review. Int J Cancer 1997; 72: 1-22.20. Bachmann F, Kruithof E. Tissue plasminogen activator, chemical and physiological aspects. Semin Thromb Hemost 1984; 10: 17•21. Madison EL. Fibrinolysis 8. 1994•22. Lijnen H, Hoef BV, Nelles L et al. Plasminogen activation with single-chain urokinase-type plasmino-gen activator (scu-PA). Studies with active site muta-genized plasminogen (Ser740----Ala) and plasmin-re-sistant scu-PA (Lys158----Glu) , Mar 1990. J Biol Chem 1990; 265: 5232-5236.23. Vassalli J, Sappino A, Belin D. The plasminogen activator/plasmin system. 1991. Clin Invest 1991; 88: 1067-1072•
24. Holmes WE, Pennica D, Blaber M et al. Cloning and expression of the gene for pro-urokinase in Es-cherichia coli. Bio/Technology 1985; 3: 923-929•25. Appella E, Robinson E, Ullrich S et al. The recep-tor-binding sequence of urokinase. A biological func-tion for the growth-factor module of proteases. J Biol Chem 1987; 262: 4437-4440•26. Potempa J, Korzus E. The serpin superfamily of proteases inhibitor: Structure, function, and regula-tion. J Biol Chem 1994; 269: 15957-15960•27. Zhou A, Huntingto J, Pannu N et al. How vitronec-tina binds PAI-1 to modulate fibrinolysis and cell migra-tion. Nature Structural Biology 2003;10: 541-544•28. McMahon GA, Petitclerc E, Stefansson S et al. Plasmi-nogen activator inhibitor-1 regulates tumor growth and angiogenesis. J Biol Chem 2001; 276: 33964-33968•29. Catalano RD, Johnson MH, Campbell EA et al. Inhi-bition of Stat3 activation in the endometrium prevents implantation: A nonsteroidal approach to contraception. Proc Natl Acad Sci USA 2005; 102: 8585-8590•30. Burysek L, Syrovets T, Simmet T. The Serine Protease Plasmin Triggers Expression of MCP-1 and CD40 in Human Primary Monocytes via Activation of p38 MAPK and Janus Kinase (JAK)/STAT Signaling Pathways. J Biol Chem 2002; 277: 33509-33517•31. Koshelnick Y, Ehart M, Hufnagl P et al. Urokinase Receptor Is Associated with the Components of the JAK1/STAT1 Signaling Pathway and Leads to Activa-tion of This Pathway upon Receptor Clustering in the Human Kidney Epithelial Tumor Cell Line TCL-598. J Biol Chem 1997; 272: 28563-28567•32. Blasi F, Carmeliet P. uPAR: a versatile signalling or-chestrator. Nat Rev Mol Cell Biol 2002; 3: 932-943•33. Tang H, Kerins DM, Hao Q et al. The urokinase-type plasminogen activator receptor mediates tyrosine phosphorylation of focal adhesion proteins and activa-tion of mitogen-activated protein kinase in cultured en-dothelial cells. J Biol Chem 1998; 273: 18268-18272•34. Yebra M, Goretzki L, Pfeifer M et al. Urokinase-type plasminogen activator binding to its receptor stimulates tumor cell migration by enhancing integrin-mediated sig-nal transduction. Exp Cell Res 1999; 250: 231-240•35. Nguyen DHD, Webb DJ, Catling AD et al. Uroki-nase-type Plasminogen Activator Stimulates the Ras/Extracellular Signal-regulated Kinase (ERK) Signaling Pathway and MCF-7 Cell Migration by a Mechanism That Requires Focal Adhesion Kinase, Src, and Shc. RA-PID DISSOCIATION OF GRB2/SOS-SHC COMPLEX IS ASSOCIATED WITH THE TRANSIENT PHOS-PHORYLATION OF ERK IN UROKINASE-TREATED CELLS. J Biol Chem 2000; 275: 19382-19388•36. Webb D, Thomas K, Gonias S. Plasminogen Activator Inhibitor 1 Functions as a Urokinase Response Modifier at the Level of Cell Signaling and Thereby Promotes MCF-7 Cell Growth. Journal of Cell Biology 2001; 152: 741-751•37. Navaneetha KR, lO G-PS, Harold AC. Urokinase Receptor Is a Multifunctional Protein: Influence of Re-ceptor Occupancy on Macrophage Gene Expression. J Clin Invest 2005; 96: 465-474•38. Nykjaer A, Conese M, Christensen E et al. Recycling

182 Activación del Sistema Plasminógeno-Plasmina
of the urokinase receptor upon internalization of the uPA:serpin complexes. EMBO J 1997; 16: 2610-2620•39. Czekay R, Kuemmel T, Orlando R et al. Direct binding of occupied urokinase receptor (uPAR) to LDL receptor-related protein is required for endo-cytosis of uPAR and regulation of cell surface uroki-nase activity. Mol Biol Cell 2001; 12: 1467-1479•40. Deng G, Curriden S, Wang S et al. Is plasmi-nogen activator inhibitor-1 the molecular switch that governs urokinase receptor-mediated cell adhesion and release? J Cell Biol 1996; 134: 1563-1571•41. Waltz DA, Natkin LR, Fujita RM et al. Plasmin and Plas-minogen Activator Inhibitor Type 1áPromote Cellular Mo-tility by Regulating the Interaction between the Urokinase Receptor and Vitronectin. J Clin Invest 1997; 100: 58-67•42. Deng G, Curriden S, Hu G et al. Plasminogen activator in-hibitor-1 regulates cell adhesion by binding to the somatome-din B domain of vitronectin. J Cell Physiol 2001; 189: 23-33•43. Bale M, Wohlfahrt L, Mosher D et al. Identifi-cation of vitronectin as a major plasma protein ad-sorbed on polymer surfaces of different copolymer composition. Blood 1989; 74: 2698-2706•44. Czekay R, Aertgeerts K, Curriden S et al. Plasminogen activator inhibitor-1 detaches cells from extracellular matri-ces by inactivating integrins. J Cell Biol 2003; 160: 781-791•45. Ugwu F, Van Hoef B, Bini A et al. Proteolytic Cleava-ge of Urokinase-Type Plasminogen Activator by Stromely-sin-1 (MMP-3). Biochemistry 1998; 37: 7231-7236•46. Declerck P, De Mol M, Vaughan D et al. Identification of a conformationally distinct form of plasminogen activator inhibitor-1, acting as a noninhibitory substrate for tissue-type plasminogen activator. J Biol Chem 1992; 267: 11693-11696•47. Mottonen J, Strand A, Symersky J et al. Structural basis of latency in plasminogen activator inhibitor-1. Nature 1992; 355: 270-273•48. Hekman C, Loskutoff D. Endothelial cells produce a latent inhibitor of plasminogen activators that can be activa-ted by denaturants. J Biol Chem 1985; 60: 11581-11587•49. Frixen UH, Nagamine Y. Stimulation of urokinase-type plas-minogen activator expression by blockage of E-cadherin-depen-dent cellcell adhesion. Cancer Res 1993; 53: 3618-3623•50. Samarakoon R, Higgins PJ. MEK/ERK pathway mediates cell-shape-dependent plasminogen activator inhibitor type 1 gene expression upon drug-induced disruption of the microfilament and microtubule net-works. J Cell Sci 2002; 115: 3093-3103.51. Dennler S, Itoh S, Vivien D et al. Direct binding of Smad3 and Smad4 to critical TGF beta-inducible ele-ments in the promoter of human plasminogen activator inhibitor-type 1 gene. EMBO J 1998; 17:3091-3100•52. Pandey M, Loskutoff D, Samad F. Molecular me-chanisms of tumor necrosis factor-á-mediated plasmi-nogen activator inhibitor-1 expression in adipocytes. FASEB J 2005; 19: 1317-1319•53. Hamdan AD, Quist WC, Gugne JB et al. Angioten-sinconverting enzyme inhibition suppresses plasminogen activator-1 expression in the neointima of balloon-inju-red rat aorta. Circulation 1996; 93: 1073-1078•54. Motojima M, Kakuchi J, Yoshioka T. Association of TGF-[beta] signaling in angiotensin II-induced PAI-1
mRNA upregulation in mesangial cells: role of PKC. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research 1999; 1449: 217-226•55. Grohe C, Kahlert S, Lobbert K et al. Angioten-sin converting enzyme inhibition modulates cardiac fibroblast growth. J Hypertens 1998; 16: 377-384•56. Motojima M, Ando T, Yoshioka T. Sp1-like ac-tivity mediates angiotensin-II-induced plasminogen-activator inhibitor type-1 (PAI-1) gene expression in mesangial cells. Biochem J 2000; 349: 435-441•57. Juhan-Vague I, A.Mavri, Morange P.E. Plasminogen acti-vator inhibitor-1, inflammation, obesity, insulin resistance and vascular risk. J Thromb Haemost 2003; 1: 1575-1579•58. Carmeliet P, Collen D. Molecular genetics of the fibrinolytic and coagulation systems in haemostasis, thrombogenesis, restenosis and atherosclerosis. Curr Opin Lipidol 1997; 8: 118-125•59. Spiegelman B, Flier J. Adipogenesis and obesity: rounding out the big picture. Cell 1996; 87: 377-389•60. Alessi M, Peiretti F, Morange P et al. Production of plasminogen activator inhibitor 1 by human adipose tissue: possible link between visceral fat accumulation and vascular disease. Diabetes 1997; 46: 860-867•61. Samad F, Loskutoff D. Tissue distribution and re-gulation of plasminogen activator inhibitor-1 in obese mice. Mol Med 1996; 2: 568-582•62. Juhan-Vague I, Alessi M. PAI-1, obesity, insulin resistance and risk of cardiovascular events. Thromb Haemost 1997; 78: 656-660•63. Sakamoto T, Woodcock-Mitchell J, Fuji S et al. Augmentation of expression of plasminogen activator inhibitor type-1 in adipocytes and its potential role in acceleration of vasculopathy associated with obesity. Circulation 1997; 96 (suppl I): I-546•64. Nagi D, Yudkin J. Effects of metformin on insulin re-sistance, risk factors for cardiovascular disease, and plas-minogen activator inhibitor in NIDDM subjects: a study of two ethnic groups. Diabetes Care 1993; 16: 621-629•65. Gertler JP, Abbott WM. Prothrombotic and fibri-nolytic function of normal and perturbed endothe-lium. J Surg Res 1992; 52: 89-95•66. Brown SL, Sobel BE, Fujii S. Attenuation of the synthesis of plasminogen activator inhibitor type 1 by niacin. A potential link between lipid lowering and fibrinolysis. Circulation 1995; 92: 767-772•67. Janand-Delenne B, Chagnaud C, Raccah D et al. Visceral fat as a main determinant of plasminogen ac-tivator inhibitor 1 level in women. Int J Obes Relat Metab Disord 1998; 22: 312-317•68. Oh CK, Ariue B, Alban R et al. PAI-1 promo-tes extracellular matrix deposition in the airways of a murine asthma model. Biochem Biophys Res Com-mun 2002; 294: 115511-60•69. Eitzman DT, McCoy RD, Zheng X et al. Bleomycin-in-duced Pulmonary Fibrosis in Transgenic Mice That either Lack or Overexpress the Murine Plasminogen Activator Inhibitor-1 Gene. J Clin Invest 1996; 97: 232-237•70. Eitzman DT, McCoy RD, Zheng X et al. Bleomycin-in-duced Pulmonary Fibrosis in Transgenic Mice That either Lack or Overexpress the Murine Plasminogen Activator

183Nolasco et al, Cancerología 2 (2007): 171-183
Inhibitor-1 Gene. J Clin Invest 1996; 97: 232-237•71. Oikawa T, Freeman M, Lo W et al. Modulation of plasminogen activator inhibitor-1 in vivo: a new mechanism for the anti-fibrotic effect of renin-angio-tensin inhibition. Kidney Int 1997; 51: 164-172•72. Cho SH, Ryu CH, Oh CK. Plasminogen Activator Inhibitor-1 in the Pathogenesis of Asthma. Experi-mental Biology and Medicine 2004; 229: 138-146•73. Pepper M.S, Sappino A.P, Stocklin R., Stocklin R., Stoc-klin R., Montesano R, Montesano R, Orci L, and Vassalli J.D. pregulation of urokinase receptor expression on mi-grating endothelial cells. J.Cell Biol. 1993; 122: 673-684•74. Soff GA, Sanderowitz J, Gately S et al. Expression of plasminogen activator inhibitor type 1 by human prostate carcinoma cells inhibits primary tumor growth, tumor-as-sociated angiogenesis, and metastasis to lung and liver in an athymic mouse model. J Clin Invest 1995; 96: 2593-2600•75. Stefansson S, Petitclerc E, Wong MKK et al. Inhi-bition of Angiogenesis in vivo by Plasminogen Activa-tor Inhibitor-1. J Biol Chem 2001; 276: 8135-8141•76. Bajou K, Noel A, Gerard R et al. Absence of host plasminogen activator inhibitor 1 prevents cancer inva-sion and vascularization. Nat Med 1998; 4: 923-928•77. Rakica JM, Maillardb C, Jostb M et al. Role of plasminogen activator-plasmin system in tumor angio-genesis. CMLS, Cell Mol Life Sci 2003; 60: 463-473•78. Tsuojiya, H, Katsuo, E, Sunayama, C, Tomita, K, Ueda, Y., and Binder, B. R. The antibody to plasmi-nogen activator inhibitor-1 suppresses pulmonary metastases of human fibrosarcoma in athymic mice. Gen.Diagn.Pathol. 1995; 141: 41-48•79. McMahon GA, Petitclerc E, Stefansson S et al. Plasmi-nogen activator inhibitor-1 regulates tumor growth and angiogenesis. J Biol Chem 2001; 276: 33964-33968•80. Nekarda H, Schmitt M, Ulm K et al. Prognostic impact of urokinase-type plasminogen activator and its inhibitor PAI-1 in completely resected gastric can-cer. Cancer Res 1994; 54: 2900-2907•81. Heiss M, Allgayer H, Gruetzner K et al. Clinical value of extended biologic staging by bone marrow micrometastases and tumor-associated proteases in gastric cancer. Ann Surg 1997; 226:7336-7344•82. Ganesh S, Sier C, Heerding M et al. Prognostic value of the plasminogen activation system in patients with gastric carcinoma. Cancer 1996; 77:1035-1043•83. Berger DH. Plasmin/plasminogen system in co-lorectal cancer. World J Surg 2002; 26: 767-771•84. Miseljic S, Galandiuk S, Myers S et al. Expression of uro-kinase-type plasminogen activator and plasminogen activator inhibitor in colon disease. J Clin Lab Anal 1995; 9: 413-417•85. Ganesh S, Sier C, Heerding M et al. Contribution of plasminogen activators and their inhibitors to the sur-vival prognosis of patients with Dukes’ stage B and C colorectal cancer. Br J Cancer 1997; 75: 1793-1801•86. Ganesh S, Sier C, Griffioen G et al. Prognostic rele-vance of plasminogen activators and their inhibitors in colorectal cancer. Cancer Res 1994; 54: 4065-4071•87. Nekarda H, Schlegel P, Schmitt M et al. Strong prog-nostic impact of tumor-associated urokinase-type plasmi-nogen activator in completely resected adenocarcinoma
of the esophagus. Clin Cancer Res 1998; 4: 1755-1763•88. Sakakibara T, Hibi K, Kodera Y et al. Plasmino-gen Activator Inhibitor-1 as a Potential Marker for the Malignancy of Esophageal Squamous Cell Carci-noma. Clin Cancer Res 2004; 10: 1375-1378•89. Salden M, Splinter T, Peters H et al. The urokina-se-type plasminogen activator system in resected non-small-cell lung cancer. Ann Oncol 2000; 11: 327-332•90. Pedersen H, Brunner N, Francis D et al. Prognos-tic impact of urokinase, urokinase receptor, and type 1 plasminogen activator inhibitor in squamous and large cell lung cancer tissue. Cancer Res 1994; 54: 4671-4675•91. Pedersen H, Grondahl-Hansen J, Francis D et al. Uroki-nase and plasminogen activator inhibitor type 1 in pulmo-nary adenocarcinoma. Cancer Res 1994; 54: 120-123•92. Abendstein B, Daxenbichler G, Windbichler G et al. Pre-dictive value of uPA, PAI-1, HER-2 and VEGF in the serum of ovarian cancer patients. Anticancer Res 2000; 20: 569-572•93. Hoffmann G, Pollow K, Weikel W et al. Urokinase and plasminogen activator-inhibitor (PAI-1) status in primary ovarian carcinomas and ovarian metastases compared to benign ovarian tumors as a function of histopathological parameters. Clin Chem Lab Med 1999; 37: 47-54•94. Janicke F, Schmitt M, Pache L et al. Urokinase plasmi-nogen activator (uPA) and its inhibitor PAI-1 are strong and independent prognostic factors in node negative breast cancer. Breast Cancer Res Treat 1993; 24: 195-208•95. Leissner P, Verjat T, Bachelot T et al. Prognostic significance of urokinase plasminogen activator and plasminogen activator inhibitor-1 mRNA expression in lymph nodeand hormone receptor-positive breast cancer. BMC Cancer 2006; 6: 216•96. Luqmani YA, Temmim l, Parkar AH et al. Clinical implications of urokinase and tissue type plasminogen activators and their inhibitor (PAI-1) in breast cancer tissue. Oncology Reports 2002; 9: 645-651•97. Harbeck N, Kates R, Look M et al. Enhanced benefit from adjuvant chemotherapy in breast cancer patients classified high-risk according to urokinase-type plasmino-gen activator (uPA) and plasminogen activator inhibitor type 1 (n = 3424). Cancer Res 2002; 62: 4617-4622•98. Grondahl-Hansen J, Peters H, van Putten W et al. High levels of urokinase type plasminogen activator and it’s inhibi-tor PAI-1 in cytoslic extracts of breast carcinomas are associa-ted with poor prognosis. Cancer Res 1993; 53: 2513-2521•99. Chazaud B, Ricoux R, Christov C et al. Promigratory Effect of Plasminogen Activator Inhibitor-1 on Invasive Breast Cancer Cell Populations. Am J Pathol 2002; 160: 237-246•100. Daneri Navarro A, Macias LG, Oceguera VA et al. Urokinase-type Plasminogen Activator and Plas-minogen Activator Inhibitors (PAI-1 and PAI-2) in Ex-tracts of Invasive Cervical Carcinoma and Precursor Lesions. Eur J Cancer 1998; 34: 566-569•101. Kobayashi H, Fujishiro S, Terao T. Impact of uro-kinase-type plasminogen activator and its inhibitor type 1 on prognosis in cervical cancer of the uterus. Cancer Res 1994; 54: 6539-6548•102. Hazelbag S, Kenter G, Gorter A et al. Pronostic relevance of TGR- and PAI-1 in cervical cancer. Int J Cancer 2004; 112: 1020-1028•