univerzitet u tuzli „istraživanja stabilnosti proteina ... •biofarmaceutici su proteini...
TRANSCRIPT

Univerzitet u Tuzli
Farmaceutski fakultet
Doktorski studij
Školska: 2014/2015
„Istraživanja stabilnosti proteina
(toplotnom denturacijom, pH
denaturacijom, denaturacijom
ureom) na rekombinantnim
proteinima“.
Dr.sc. Aida Smajlović, docent

• Biofarmaceutci savremeni način liječenja, naročito
malignih i tumorskih oboljenja.
• Biofarmaceutici su uglavnom proteinske strukture,
koja je jako nestabilna, čak i pri malim promjenama
uslova (pH, tamperatura, prisustvo nekih otapala,...),
kada može doći do promjene konformacijskog stanja.

• Biofarmaceutici su proteini (antitijela), nukleinske kiseline
(DNA, RNA, antisense lijekovi, oligonukleotidi) koji se koriste
za terapijske ili in vivo dijagnostičke svrhe.
• Modifikatori biološkog odgovora (engl. biological
modifiers)
• Prirodne supstance identične humanim koje mogu
modifikovati normalan imuni odgovor organizma.
• U biofarmaceutike spadaju: proteini, antitijela, enzimi,
interleukini, hormoni, vakcine...

•FDA (Food and Drug Administration) Biofarmaceutici
se definišu kao biotehnološki proizvodi odnosno bilo
koji virus, terapijski serum, toksin, antitoksin ili analogni
proizvod koji se može primijeniti u prevenciji, tretmanu
ili liječenju bolesti ili povreda kod ljudi .
• Većina biofarmaceutika proizvedena je genetskim
inženjeringom; prva takva supstanca koja je
administrirana u terapeutske svrhe je biosintetisani
humani inzulin, pripremljen metodama rekombinantne
tehnologije,1982. godine.

Razvoj biofarmaceutika
Proizvodnja
biofarmaceutika je
najvažniji
sektor u biotehnološkoj
industriji i jedna
je od najbrže razvijajućih
industrija
visoke tehnologije.
•Prvi primjeri
biotehnoloških
lijekova bili su
jednostavni
proteini –
„replacement
therapy“.
Kasnije je razvoj usmjeren
ka molekularnoj medicini i
otkrivanju većeg broja
gena koji su povezani sa
malignim oboljenjima.
Otkriveni su i klonirani neki
kancer-determinišući geni,
zbog čega se biotehnološki
lijekovi sve više koriste kao
zamjena za hemoterapiju.
Trenutno su u toku
istraživanja na više
od 100 potencijalno
terapijskih gena.
•Kasnih 90-tih
godina više od 350
biotehnoloških
lijekova bilo je u
različitim fazama
razvoja u više od
140 farmaceutskih i
biotehnoloških
kompanija.
•Tada se procijenilo da će
pacijenti oboljeli od
hemofilije, ozbiljne sepse,
ulcera kože,reumatoidnog
artritisa i brojnih kancera
imati koristi u narednim
godinama kada lijekovi
prođu kroz klinička
ispitivanja i dobiju dozvolu
za puštanje u promet.

• Neki od ovih lijekova već su na tržištu (Refacto za
hemofiliju, Fuzeon za terapiju HIV-a, Kineret za
hemoterapiju, Xigris za teške sepse).
• Genetski inženjering (tehnologija rekombinantne DNA)-
vještačko obrazovanje novih kombinacija nasljednog
materijala.
• Genetički kod je univerzalan i može se prenositi iz
jednog organizma u drugi čime se dobija organizam sa
drugačijom kombinacijom gena.
• Takva DNA je hibridna (rekombinantna) - u prirodi se
nikada ne nalazi.

• Genetski inženjering ima za cilj:
-stvaranje novih farmaceutika odnosno
biofarmaceutika (lijekova);
-stvaranje sigurnijih i/ili efikasnijih verzija već postojećih
farmaceutika odnosno biofarmaceutika;
-proizvodnju supstanci identičnih konvencionalno-
napravljenim farmaceuticima, u cilju uštede.
• Primjer biofarmaceutika koji je efikasan u tretmanu
kancera dojke je trastuzumab (Herceptin®),
monoklonsko antitijelo protiv Her2 receptora za tirozin
kinazu, koji je pre-eksprimiran u oko 1/3 tumora dojke.


Klasifikacija biofarmaceutika
•Biofarmaceutike možemo svrstati u dvije skupine:
biofarmaceutike prve generacije i biofarmaceutike
druge generacije.
•Biofarmaceutici prve generacije -
uglavnom kopije
endogenih proteina ili
antitijela, proizvedeni
rekombinantnom DNA tehnologijom.
•Biofarmaceutici druge
generacije - proteini ili
antitijela čije su fizičko-
hemijske karakteristike
dodatno poboljšane
inženjeringom.

Klasifikacioni sistem biofarmaceutika-BCS
(Biopharmaceutical Classification System) uzima u
obzir tri glavna faktora: rastvorljivost, topivost i
intestinalnu apsorpciju lijekova nakon oralne
primjene po trenutnom oslobađanju doze iz čvrste
oralne forme.
•Prema BCS, ljekovite supstance su klasifikovane u
različite grupe:
•1.klasa (visoka topivost i visoka permeabilnost)
•2.klasa (niska topivost, visoka permeabilnost)
•3.klasa (visoka topivost, niska permeabilnost)
•4.klasa (niska topivost i permeabilnost).

Vrste biofarmaceutika
•Biofarmaceutici se danas razvijaju kako bi se pomoću njih borilo sa
karcinomom, virusnim infekcijama, dijabetesom, hepatitisom, multiplom
sklerozom. Prema tome, oni su grupisani u slijedećih sedam glavnih
kategorija:
•Citokini
•Enzimi
•Hormoni
•Faktori zgrušavanja
•Vakcine
•Monoklonska antitijela
•Peptidni terapeutici .
•Trodimenzionalna struktura humanog hormona rasta koji se može svrstati u
biofarmaceutike prikazana je na slici.


Stabilnost biofarmaceutika
• Stabilnost lijekova se definiše kao stepen u kome
dozirni oblik zadržava sve svoje strukturne
karakteristike u navedenim granicama, kroz cijeli
period čuvanja i upotrebe koje je posjedovao i u
vrijeme izrade.
• Cilj ispitivanja stabilnosti ljekovitih preparata je da se
odredi vremenski period u kojem je lijek stabilan,
odnosno za koje vrijeme izgubi najviše 10% dejstva
aktivnog principa, skladišten pod normalnim uslovima.

• Stabilnost: hemijska, fizička, mikrobiološka.
• Na hemijsku stabilnost utiču: pH, temperatura,
svjetlost, priroda rastvarača, prisustvo kiseonika, joni
teških metala, prisustvo peroksida...
• Biotehnološki lijekovi su uglavnom proteini- imaju
nestabilnu strukturu i osjetljivi su na promjene in vivo i
in vitro okruženja.
• Relativno male promjene nekih faktora mogu
narušiti strukturu proteina. Ti faktori su:
- temperatura
- koncentracija soli
-denaturirajuća sredstva (urea, GdnHCl)
- pH

• Terapijska efikasnost biofarmaceutika određena je
njihovom proteinskom trodimenzionalnom strukturom
koja je ovisna o interakcijama između aminokiselina.
•Trodimenzionalna struktura se održava slabim,
nekovalentnim vezama i veoma je osjetljiva čak i na
male promjene u temperaturi ili pH.
•Može doći do denaturacije ili čak razgradnje u
organizmu i eliminacije.
• Proteini mogu umjesto nativnog stanja prilikom svijanja
zauzeti formu agregata ili amiloida,što je nepoželjno.

•Neophodne su mjere za određivanje i poboljšanje
stabilnosti- proteini se koriste u terapeutske svrhe!
•Najpoznatije metode koje se bave studijama stabilnosti
biofarmaceutika su:
Metoda cirkularnog dihroizma
Diferencijalna skenirajuća kalorimetrija
UHPLC ( Ultra-High Performance Liquid Chromatography)
Fluorescentna spektroskopija

•Primarna struktura proteina - podrazumijeva redoslijed vezivanja
aminokiselina u peptidnom lancu.
•Sekundarna struktura- podrazumijeva položaj koji u prostoru zauzimaju atomi
peptidnih veza u molekuli.
•Tercijarna struktura- prostorni raspored svih atoma u molekuli proteina
predstavlja
•Kvaternarna struktura- proteini čija se molekula sastoji iz više od jednog
polipeptidnog lanca.

Stabilnost biofarmaceutika tokom skladištenja
• Tokom skladištenja biofarmaceutika može doći do: racemizacije,
izomerizacije, oksidacije i agregacije.
Biofarmaceutici se skladište na temperaturi između 2 i 8 ºC
• Da bi se spriječilo stvaranje agregata - pažljivo se biraju rastvarači i
pomoćne supstance- ne smiju imati negativno dejstvo po biofarmaceutik.
• Sporije rastvaranje – smanjenje agregacije
• Protresanje oblika- uzrokuje pjenjenje- smanjuje se međuprostor između
vode i zraka.
• Mehanički stres - tokom procesiranja, rukovanja, transporta - uzrokuje
agregaciju biofarmaceutika.
• Jako je bitno: pažljivo rukovanje, transport,
skladištenje - mogu se spriječiti mehanički stresovi -
povećati i održati stabilnost biofarmaceutika.

Metode za određivanje stabilnosti
biofarmaceutika
• Stabilnost: - hemijska ( stabilnost kovalentnih veza )
- konformacijska ( stabilnost svijenog
proteina )
• Definicija: promjena slobodne energije u reakciji
promjene proteina iz nativnog u denaturisano stanje:
G° (T) = -RT ln K(T)

1. Cirkularni dihroizam (CD)
• CD- mjeri razlike u apsorpciji lijevo i desno kružno
polarizovane svjetlosti.
• Mjerenje optičke aktivnosti asimetričnih molekula.
• Mjeri se u dva područja:
- daleki UV CD spektar ( 170-250nm )
- bliski UV CD spektar ( 250- 320 nm )

• Mjerenje CD spektra radi se na temperaturi od 23°C, pomoću
spektropolarimetra.
• on se sastoji od: izvora monohromatskog linearno - polarizovanog
svjetla, optičkog sistema koji ga prevodi u desno i lijevo cirkularno
polarizovanu svjetlost (CPL), detektora i računarskog dijela uređaja
koji izračunava razliku između apsorbovane lijevo cirkularnopolarizovane svjetlosti (LCLP) i desno cirkularno polarizovane
svjetlosti (DCLP).

- ispitivanje sekundarne i tercijarne strukture proteina,
- poređenje strukture proteina iz različitih izvora ili strukture
mutiranih i nativnih proteina,
- ispitivanje uticaja temperature, pH i drugih uvjeta u okolini
ili interakcije sa drugim molekulama (protein-protein ili
protein-ligand) na konformaciju poteina,
- ispitivanje stabilnosti konformacije u stresnim uvjetima (pH,
T, denaturirajući agensi), pronalaženje uslova koji bi
obezbjedili više temperature topljenja i reverzibilnost
termalne denaturacije, dakle uslova koji općenito
povećavaju stabilnost,
- utvrđivanje optimalnih uslova za proizvodnju proteina
CD je metoda naročito pogodna za:

Određivanje sekundarne strukture proteina metodom
cirkularnog dihroizma
• Sekundarna struktura se može odrediti CD
spektroskopijom u "dalekom-UV" spektralnom području
(170-250 nm).
• Hromofora je peptidna veza, a signal nastaje kada se
ona nalazi u normalnom, svijenom stanju.

• Daleki-UV CD spektri zahtijevaju 20-200 μL otopine uzorka koja
sadrži 1 mg/mL do 50 g/mL proteina, u puferu.

Određivanje tercijarne strukture biofarmaceutika
metodom cirkularnog dihroizma
• CD spektar proteina u "bliskom -UV spektru" (250-350 nm)
može biti osjetljiv na određene aspekte tercijarne
strukture.
•Na tim talasnim dužinama hromofore su aromatične
aminokiseline i disulfidne veze, a snimljeni CD signali
odnosno spektri osjetljivi su na ukupnu tercijarnu strukturu
proteina.

• Signali u regiji 250-270 nm mogu se pripisati
fenilalaninskim aminokiselinskim ostacima, signali
270-290 nm mogu se pripisati tirozinskim, a oni u
regiji 280-300 nm mogu se pripisati triptofanskim
aminokiselinskim ostacima.
• Signal u bliskoj- UV CD regiji je dosta slabiji nego u
dalekoj -UV CD regiji.
• Bliski -UV CD spektri zahtijevaju veće količine
proteina za određivanje njihove tercijarne strukture.

Upoređivanje konformacijskih stanja
biofarmacutika pomoću CD

2. Diferencijalna skenirajuća kalorimetrija (DSC)
•Parametar koji karakteriše stabilnost sistema (npr.,
svijeno stanje proteina odnosno odvijeno stanje
proteina) je ravnotežna konstanta (K) ili slobodnaenergija (ΔG°) koja predstavlja zbir entalpije (ΔH°) i
entropije (ΔS°).
•Ovi parametri su ovisni o temperaturi kroz promjenu
toplotnog kapaciteta (ΔCp).
•ΔH° i ΔCp se računaju indirektno pomoću van’t Hoff-
ove metode,ili se mjere direktno kalorimetrijom.

•DSC- direktno mjerenje termodinamičkih
parametara koji karakterišu biomolekule.
• Diferencijalna skenirajuća kalorimetrija
(DSC) je tehnika koja registruje energiju
(energetski fluks) potrebnu za održavanje
nulte temperaturne razlike između
ispitivanog uzorka i referentnog materijala,
pri unaprijed definisanoj brzini grijanja
(hlađenja), uz pretpostavku da se oba
materijala nalaze pod istim uslovima.

• DSC uređaj može raditi u dva režima: izotermskom i adijabatskom
(dinamičkom).
Izotermski metod: temperatura
ispitivanog uzorka i
temperatura referentnog
materijala održavaju jednakim
tokom zagrijavanja, variranjem
snage u oba segmenta peći,
preko koje se dobija promjena
entalpije ili toplotnog
kapaciteta u uzorku u odnosu
na referentni materijal.
Adijabatski režim rada podrazumjeva
da DSC kalorimetar obezbjeđuje
konstantnost toplotnog fluksa
između uzorka i referentnog
materijala. U ovom slučaju, promjena
entalpije ili toplotnog kapaciteta u
uzorku uzrokuje razliku temperature
u odnosu na referentni uzorak, a
mjeri se preko razlike napona između
uzorka i referentnog materijala.

Termodinamički parametri
• DSC-može pratiti termički inducirane prijelaze, a
posebno konformacijske prijelaze bioloških
makromolekula. (npr. između svijenog i odvijenog stanja
proteina ili između pojedinačne i dvostruke uzvojnice
DNA).

•DSC mjeri višak toplinskog kapaciteta molekule koju
ispitujemo (Cp) u funkciji temperature.
• Integracija Cp naspram T krive doprinosi promjenientalpije (ΔH°m).
• DSC je jedina metoda za direktno određivanjepromjene entalpije ΔH°m

DSC i svijanje proteina / stabilnost
• Protein u denaturisanom stanju ima veći toplotni
kapacitet nego onaj u nativnom- ΔCp odvijenog
proteina skoro uvijek pozitivan !
• Na svijanje biofarmaceutika utiču: hidrofobne
interakcije, vodikove veze, konformacijska
entropija, temperatura, pH...
• Temperatura taljenja (Tm)- indikator
termostabilnosti; veći Tm- protein je termostabilniji!
• Uticaj pH na stabilnost- maximalna termička
stabilnost je u blago kiselim uslovima! (studija na
osnovu pH 2-hidroksipropil-b-ciklodekstrin
(HPbCD) saharoze )

3. UHPLC ( Ultra-High Performance Liquid
Chromatography)
• HPLC odnosno tečna hromatografija visokih performansi ili
tečna hromatografija pod visokim pritiskom - oblik kolonske
hromatografije koji se koristi za radzvajanje komponenti iz
smjese na osnovu hemijskih interakcija između supstance
koja se analizira i stacionarne faze u koloni .
• Princip rada HPLC-a je forsiranje prolaska analizirane
supstance (ili smjese) kroz kolonu (cijev punjena
materijalom sitnih čestica) pumpanjem tečnosti (mobilna
faza) pod visokim pritiskom kroz kolonu (stacionarna
faza).

•Vrijeme zadržavanja zavisi od: prirode supstance
koja se analizira, stacionarne faze i sastava mobilne
faze.
• Vrijeme za koje se supstanca eluira (dođe do kraja
kolone)- retenciono vrijeme i karakteristično je za
određenu supstancu.
• Veći pritisak- veća brzina razdvajanja.
• Rastvarači: voda, metanol, organski rastvarači (u
vodu se može dodati i pufer).

Hromatografija na normalnim fazama
• Stacionarna faza- polarna ; mobilna faza- nepolarna
• Supstanca koja se analizira- polarna!
• Veća polarnost- veća jačina adsorpcije i duže vrijeme
zadržavanja.
Hromatografija na obrnutim fazama
• Stacionarna faza- nepolarna; mobilna faza- polarna
• Najčešća stacionarna faza je silikatna, tretirana sa RMe2SiCl,
gdje je R alkilna grupa ravnog lanca kao C18H37 ili C8H17.
• Dodatkom polarnih rastvarača u mobilnu fazu- povećava se
vrijeme zadržavanja.
• Dodatkom hidrofobnih interakcija- smanjuje se vrijeme zadržavanja.

Uređaj za HPLC se sastoji od sljedećih komponenti:
•Rezervoar mobilne faze
•Pumpa
•Injektor
•Kolona
• Detektor
• Ultra HPLC koristi isti princip metode separacije kao i konvencionalna
HPLC, ali razlika je što UHPLC koristi kolone sa česticama manjeg promjera
od 2 μm.
• Manje čestice omogućavaju: veću efikasnost kolone, povećanje masene
osjetljivosti, analitičke rezolucije te brzine.
•Zahtijeva veći pritisak- samim tim i kvalitetnije pumpe.

• Otkriće RNA interferencije (RNAi) kao mehanizma za
selektivno utišavanje ekspresije messenger RNA (mRNA)
obezbjedilo je ogroman potencijal u okviru
biomedicinskih istraživanja i razvoja lijeka.
• Upotreba malih posredničkih RNA (siRNA) postala je
glavni fokus industrijskih i akademskih laboratorija.
• Jedan od glavnih izazova- učinkovita sistemska
dostava siRNA na ciljnim stanicama.
• Formulacije bazirane na liposomima u lipidnim
nanočesticama (LNP) najviše obećavaju i naširoko se
koriste kao strategija za in vivo isporuku siRNA.

•UHPLC- ključna uloga u karakterizaciji hemijskih
svojstava LNP formulacija!
•UHPLC- visoka propusnost i vrhunska učinkovitost
razdvajanja- brzo i pouzdano određivanje RNA, lipida i
srodnih formulacija u složenim frakcijama.
•Učinkovita i sigurna isporuka siRNA na ciljne
stanice- glavna prepreka u ostvarivanju velikih
mogućnosti siRNA kao terapeutika.
•Katjonski liposomi na bazi lipida u nanočesticama
(LNP)- najefikasniji u riješavanju tih problema.
•LNP- in vivo dostavljanje- poboljšavaju farmakokinetička svojstva sintetičkih
oligonukleotida,štiti od enzimske degradacije.

UHPLC u farmaceutskoj analizi
• UHPLC- uveden je 2004. godine i naširoko je
prihvaćena i popularna metoda u mnogim industrijama.
• UHPLC u odnosu na konvencionalnu HPLC ima
slijedeće prednosti:
Povećana hromatografska separacija
Znatno brža separacija
Povećana masena osjetljivost
Malo odlaganje volumena (brzo ponovno dovođenje u
ravnotežu kolone za gradijent analizu)
Brz odgovor detektora za efikasno eluiranje pikova na
hromatogramu
Korištenje submikroskopskih čestica
• UHPLC je najsavremenija metoda za farmaceutsku
analizu i jako je pogodna za brzo i ultra-brzo odvajanje!

4. Fluorescentna spektroskopija
• Fluorescencija-pojava koja nastane kada supstanca
koja je izložena elektromagnentom zračenju emitira
elektromagnetno zračenje veće talasne dužine od onog
kojim je izložena.
•Kod većine proteina nosioc fluorescencije je triptofan.
• Nukleinske kiseline ne fluoresciraju. Međutim, one se
mogu proučavati fluorescentnom spektroskopijom
ukoliko se vežu za neku pogodnu fluorescentnu
molekulu, tzv. sondu .

Horizontalnim linijama označeni su različiti vibracioni nivoi
(v=0,1,2,3...) osnovnog elektronskog stranja (S0), kao i vibracioni nivoi ekscitiranog elektronskog stanja (S1').

• Razlika u energiji odnosno talasnoj dužini (predstavljena kao hνEX - hνEM)
se naziva Stokes-ov pomak (promjena).
• Stokes-ova pomak (promjena) je fundamentalna za osjetljivost
fluorescentne metode.

• Iako postoje 3 aminokiseline koje posjeduju intrinzna fluorescentna
svojstva (Trp, Tyr, Phe) samo tirozin (Tyr) i triptofan (Trp) koriste se tokom
istraživanja svijanja proteina jer su njihovi kvantni prinosi (emitirani
fotoni/ekscitirani fotoni) dovoljno veliki da bi dali zadovoljavajući fluorescentni
signal.
• Ti aminokiselinski ostaci se mogu upotrijebiti za praćenje svijanja proteina,
jer njihova fluorescentna svojstva (kvantni prinosi) su osjetljiva na okolinu, a
ona se mijenja tokom svijanja ili odvijanja proteina.
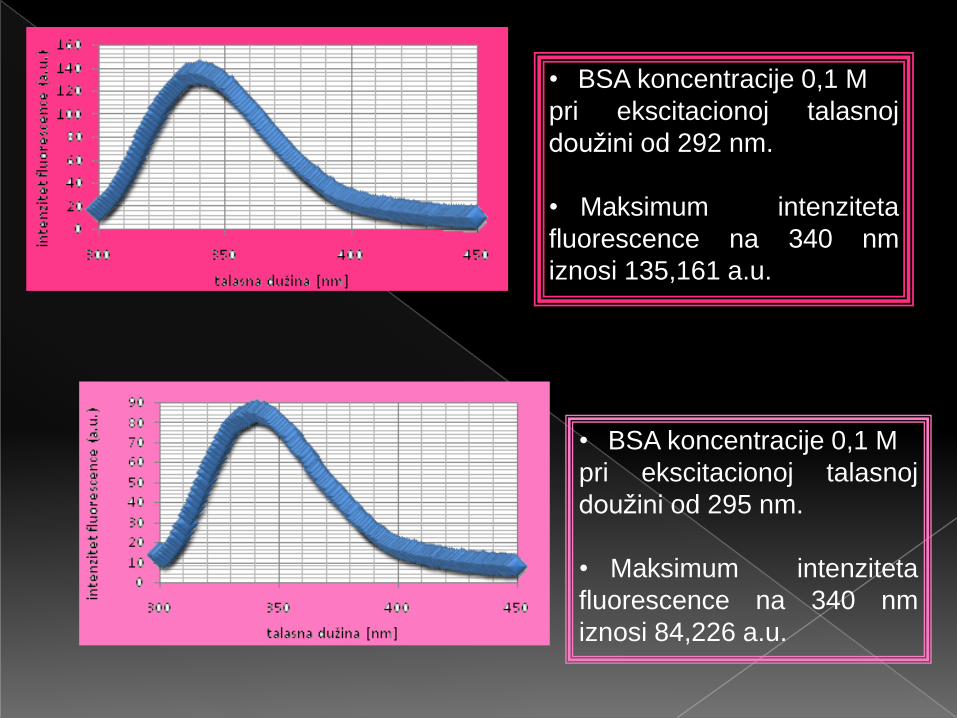
• BSA koncentracije 0,1 M
pri ekscitacionoj talasnoj
doužini od 292 nm.
• Maksimum intenziteta
fluorescence na 340 nm
iznosi 135,161 a.u.
• BSA koncentracije 0,1 M
pri ekscitacionoj talasnoj
doužini od 295 nm.
• Maksimum intenziteta
fluorescence na 340 nm
iznosi 84,226 a.u.

• BSA koncentracije 0,2 M
pri ekscitacionoj talasnoj
doužini od 292 nm.
• Maksimum intenziteta
fluorescence na 340 nm iznosi
180,108 a.u.
• BSA koncentracije 0,2 M
pri ekscitacionoj talasnoj
doužini od 295 nm.
• Maksimum intenziteta
fluorescence na 340 nm
iznosi 120,087 a.u.

• Emisijski fluorescentni spektri nativnog goveđeg serum albumina
(C=0,2M), djelimično denturisanog u 1 – 5 M urei i potpuno
denaturisanog u 6M urei, snimljeni u području 300-450 nm pri
ekscitaciji od 292 nm nakon ravnotežne denaturacije od 40 min.

• Emisijski fluorescentni spektri nativnog goveđeg serum albumina
(C=0,2M), djelimično denturisanog u 1 – 5 M urei i potpuno
denaturisanog u 6M urei, snimljeni u području 300-450 nm pri
ekscitaciji od 295 nm nakon ravnotežne denaturacije od 40 min.

Procjena konformacijske stabilnosti pomoću
fluorescentne spektroskopije triptofana
• Fluorescencija svijenih proteina predstavlja
mješavinu fluorescencija iz pojedinih aromatskih
aminokiselinskih ostataka.
• Većina emisija fluorescencija dolazi od triptofanskih
ostataka, rijeđe od tirozinskih i fenilalaninskih.
• Značajne su i disulfidne veze.
• Triptofan ima talasnu dužinu sa maksimalnom
apsorpcijom od 280 nm i emisiju pika u rasponu od 300
do 350 nm, zavisno od polarnosti lokalnog okruženja.
• Na fluorescenciju triptofana utiče i blizina drugih a.k.
ostataka kao npr. asperginskih i glutaminskih,koji mogu
uzrokovati gašenje fluorescencije triptofana.

•Triptofan – relativno rijetka aminokiselina u proteinima (jedan do
nekoliko triptofanskih ostataka).
•Ako protein koji sadrži 1 triptofanski ostatak denaturira-pojava
crvenog pomaka spektra. (zbog izlaganja triptofana vodenomokruženju)
•Dodatak površinski-aktivne tvari (npr. tenzida) proteinu koji sadrži
triptofan-izaziva plavi pomak spektra. (zbog ugrađivanja triptofana
u vezikule ili micele tenzida)

Dužina života
Absorbancija Fluorescencija
Talasna dužina Apsorpcija Talasna dužina Kvant
Triptofan 2.6 280 5,600 348 0.20
Tirozin 3.6 274 1,400 303 0.14
Fenilalanin 6.4 257 200 282 . 0.04
•Fluorescentna spektroskopija triptofana predstavlja
dobar, brz i ekonomičan način za provjeru termičke
konformacijske stabilnosti različitih formulacija,
kakve su i biofarmaceutici.

Karakterizacija rastvora trastuzumaba (Herceptin®)
pomoću fluorescentne spektroskopije
• Terapeutski imunoglobulin trastuzumab (Herceptin®)
bi trebao biti topiv u 0.9% NaCl, a uočeno je i otapanje
u 5% dekstrozi.
• Nakon analize određenom metodom separacije, ne
može se uočiti razlika između trastuzumaba
otopljenog u NaCl-u i trastuzumaba otopljenog u
dekstrozi.
• U 5% dekstrozi se uočavaju agregati.

•Fluorescentna spektroskopija prati promjene u
strukturi (naročito tercijarnoj) proteina-
biofarmaceutika.
•Analiza pokazuje: dolazi do porasta intenziteta
fluorescencije i porasta anizotropija uzorka antitijela,
što uzrokuje njegovo vezivanje za trastuzumab
otopljen u dekstrozi- što uzorkuje stvaranje agregata.

Spektar anizotropije uzorka
antitijela u prisustvu
trastazumaba otopljenog u
0.9%NaCl-u (jasna linija) i
trastazumaba otopljenog u 5%
dekstrozi (isprekidana linija).
Fluorescentni emisioni spektar
uzorka antitijela u prisustvu
trastazumaba otopljenog u 0.9%
NaCl-u ( jasna linija) i 5%
dekstrozi (isprekidana linija).

Biofarmaceutici su proteini (antitijela), nukleinske kiseline (DNA,RNA)
koji se koriste kao lijekovi. Dobijaju se procesima biotehnologije
korištenjem živih organizama.
Posljednjih godina se zapaža veliki porast ovih lijekova na tržištu i
smatra se, da će se ta tendencija za inovacijom i nastaviti.
Koriste se u terapiji raznih teških oboljenja a naročitu primjenu su
našli u liječenju hroničnih inflamatornih oboljenja kao što je
reumatoidni artritis.
S obzirom na proteinsku strukturu- nestabilni su i skloni promjeni svog
osnovnog stanja.

Velika pažnja se posvećuje proučavanju stabilnosti
biofarmaceutika te načinu poboljšavanja iste.
Metode koje se uspješno koriste za proučavanje
stabilnosti biofarmaceutika su: cirkularni dihroizam,
diferencijalna skenirajuća kalorimetrija, UHPLC,
fluorescentna spektroskopija i druge.
Jako je važno pažljivo rukovanje i skladištenje
biofarmaceutika radi očuvanja stabilnosti
biofarmaceutika, i spriječavanja pojave mehaničkih
stresova koje je narušavaju.

Tačkaste mutacije kod rekombinantnih proteina
P74S(E31) i H75W(Y31) ugrađene su na sredini
treće petlje.

-45
-25
-5
15
35
55
75
95
115
135
250 270 290 310
λ[nm]
deg
cm
2 d
mo
l-1
stB(E31)
P74S(E31)
H75W
▄ Spektri u bliskom UV području, koji pojašnjavaju asimetričnu okolinu
aromatskih aminokiselinskih ostataka, pokazali su da oba mutanta
P74S(E31) i H75W(Y31) imaju istu poziciju tirozinskog pika u bliskom
UV području kod 275 nm.
▄ Rekombinantni mutant H75W(Y31) izo-oblika divljeg-tipa (wt) čovječijeg
stefina B (Y31) ima dodatni triptofanski pik kod 254 nm.
▄ CD spektar mutanta P74S(E31) u bliskom-UV području jasno pokazuje
da njegova mutacija ne mjenja tercijarnu strukturu u odnosu na incijalni
protein tj. divlji-tip (wt) čovječijeg stefina B, tj. ne opažaju se promjene na
rigidnoj okolini aromata

-6000
-4000
-2000
0
2000
4000
6000
8000
195 205 215 225 235 245
λ[nm]
deg
cm
2 d
mo
l-1
stB(E31)
P74S(E31)
H75W
▄ Male razlike u dalekim-UV CD
spektrima posljedica su različite
distribucije oligomera u uzorcima
stefina B (E31) i njegovog mutanta
P74S(E31).
stB(E31)
-7000
-4000
-1000
2000
5000
195 205 215 225 235 245
λ[nm]
de
gc
m2 d
mo
l-1
stB(E31)
stB(E31)-M
stB(E31)-D
Protein %M %D %T %O
stB(E31) 32 33,49 23,54 11,05
P74S(E31) 2,17 73,23 15,92 8,68

Stabilnost proteina
Metode za određivanje
stabilnosti proteina
Denaturacija ureom Toplotna denaturacija
ΔG°N-U
cm
m
ΔH
Tm
termodinamički parametri termodinamički parametri
pH denaturacija

Denaturacija ureom
86420
0
-2000
-4000
-6000
Koncentracija uree [M]
Mo
larn
a e
lip
tiè
no
st
[de
g c
m2
/ d
mo
l]
P74S(E31)
86420
0
-2000
-4000
-6000
Koncentracija uree [M]
Mo
larn
a e
lip
tiè
no
st
[md
eg
cm
2 /
dm
ol]
H75W
86420
0
-2000
-4000
-6000
Koncentracija uree [M]
Mo
larn
a e
lip
tiè
no
st
[de
g c
m2
/ d
mo
l]
stB(E31)
Protein ΔGºN-U(kJ/mol) m (kJ/mol) cm (M)
wt stefin B (E31) -31,29 ± 3,9 6,5 ± 0,8 4,9 ± 0,1
P74S(E31) mutant -40,53 ± 4,7 7,7 ± 0,9 5,3 ± 0,1
H75W(Y31) mutant -20,79 ± 3,1 5,4 ± 0,8 4,3 ± 0,1
▄ Veća vrijednost kooperativnosti (m) tačkastog mutanta P74S(E31) u
odnosu na divlji-tip (wt) čovječijeg stefina B (E31) i tačkasti mutant
H75W(Y31), najvjerovatnije je posljedica dimerizacije.

▄ Denaturacijske krive prikazuju zavisnost udjela odvijenog proteina od
koncentracije denaturirajućeg sredstva (uree), a izračunate su korištenjem dvo-
stepene aproksimacije (Santoro i Bolen, 1988):
Δε={[(ΔεN+mN x [D])+(ΔεU+mU x [D]) x exp-(ΔG˚N-U/RT+mG x [D]/RT)]/
[1+exp-(ΔG˚N-U/RT+mG x [D]/RT)]}
-Δε je vrijednost signala u mdeg pri izabranoj talasnoj dužini,
-ΔεN i ΔεU su vrijednosti signala pri određenoj talasnoj dužini za svijeni i odvijeni
protein,
-mN i mU predstavljaju nagib bazne crte u pred- i post-tranzicijskom dijelu krive,
-mG nagib pravca u tranzicijskom području,
-[D] je koncentracija denaturirajućeg sredstva,
-ΔG˚N-U slobodna energija odvijanja ekstrapolirana na koncentraciju
denaturirajućeg sredstva 0.
▄ Na osnovu gore navedene jednačine izračunati su udjeli odvijenog proteina u
funkciji koncentracije uree:
fU=(ΔεN- Δε)/(ΔεN- ΔεU)

▄ Ravnotežne denaturacije ureom, pokazuju da se divlji-tip (wt)
čovječijeg stefina B i tačkasti mutanati stefina B P74S(3E31) i
H75W(Y31) odvijaju kooperativno (sekundarna i tercijarna
struktura se odvijaju istovremeno) .
▄ Ovo je jedna od osobina proteina koji se svijaju dvostepeno.
86420
1
0,8
0,6
0,4
0,2
0
koncentracija uree [M]
ud
io o
dvije
no
g p
rote
ina
(fu
) stB(E31)
86420
1
0,8
0,6
0,4
0,2
0
koncentracija uree [M]
ud
io o
dvije
no
g p
rote
ina
(fu
) P74S(E31)
86420
1
0,8
0,6
0,4
0,2
0
koncentracija uree [M]
ud
io o
dvije
no
g p
rote
ina
(fu
) H75W

Toplotna denaturacija
▄ CD spektri snimljeni u dalekom UV području, za divlji-tip (wt) čovječijeg
stefina B u temperaturnom području od 313 K do 363 K, a za mutantne
proteine P74S(E31) i H75W(Y31) u temperaturnom području od 298 K do 368
K, na talasnim dužinama 222 nm i 210 nm, radi određivanja temperature
neophodne za potpuno odvijanje proteina.
▄ CD signali na talasnoj dužini od 222 nm i 210 nm korišteni su za analizu
krivih odvijanja proteina dobivenih toplotnom denaturacijom proteina.
▄ Krive toplotne denaturacije su prilagođene i date kao modifikovana forma
Van´t Hoff-ove jednačine, koja istovremeno prilagođava nativnu i
denaturisanu baznu liniju, kao i tranzicijski region.
▄ Na osnovu modifikovane forme Van´t Hoff-ove jednačine određene su
vrijednosti temperature taljenja Tm i promjene entalpije ∆H za divlji-tip (wt)
rekombinantnog čovječijeg stefina B, tačkasti mutant P74S(E31) i tačkasti
mutant H75W koji su podvrgnuti toplotnoj denaturaciji (Karantzeni i sar.,
2003):
∆ε = (mnT + bn) + (mdT + bd) [K / (1+K)]
gdje je
K = exp[-∆H(1 – T/Tm) / RT]

B
380360340320300
-24000
-28000
-32000
-36000
-40000
Temperatura [K]
md
eg
(2
10
nm
) 210 nm
H75W
C
380360340320
-40000
-44000
-48000
-52000
-56000
Temperatura [K]
md
eg
(2
10
nm
)
210 nm
stB(E31)
A
380360340320300
-30000
-35000
-40000
-45000
-50000
Temperatura [K]
md
eg
(2
10
nm
)
210 nm
P74S(E31)
380360340320
-50000
-52000
-54000
-56000
Temperatura [K]
md
eg
(2
22
nm
) 222 nm
stB(E31)
380360340320300
-48000
-50000
-52000
Temperatura [K]
md
eg
(2
22
nm
)
222 nm
P74S(E31)
380360340320300
-35000
-40000
-45000
-50000
-55000
Temperatura [K]
md
eg
(2
22
nm
) 222 nm
H75W
A
B
C
210 nm 222 nm
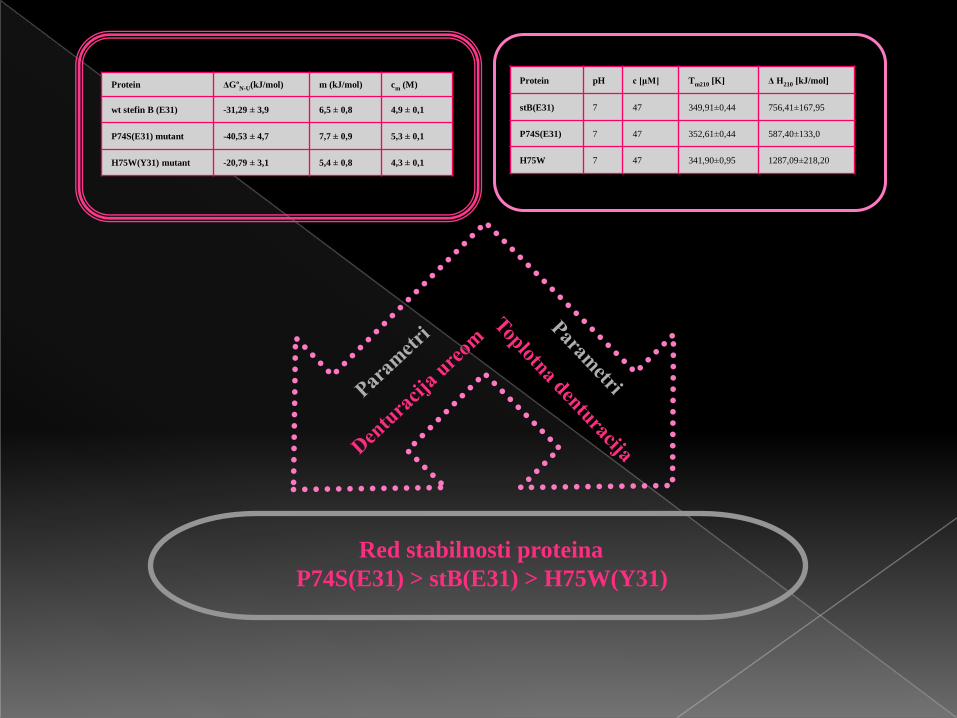
Protein ΔGºN-U(kJ/mol) m (kJ/mol) cm (M)
wt stefin B (E31) -31,29 ± 3,9 6,5 ± 0,8 4,9 ± 0,1
P74S(E31) mutant -40,53 ± 4,7 7,7 ± 0,9 5,3 ± 0,1
H75W(Y31) mutant -20,79 ± 3,1 5,4 ± 0,8 4,3 ± 0,1
Protein pH c [μM] Tm210 [K] Δ H210 [kJ/mol]
stB(E31) 7 47 349,91±0,44 756,41±167,95
P74S(E31) 7 47 352,61±0,44 587,40±133,0
H75W 7 47 341,90±0,95 1287,09±218,20
Red stabilnosti proteina
P74S(E31) > stB(E31) > H75W(Y31)

Strukturna karakterizacija intermedijata svijanja
stB(Y31)
-30
0
30
60
90
120
150
250 270 290 310
λ[nm]
de
gc
m2 d
mo
l-1
stB(Y31); pH=6,04
stB(Y31); pH=4,8
Ravnotežni spektri izo-oblika divljeg-
tipa (wt) čovječijeg stefina B (Y31) pri
pH 6,04 (nativan protein) i pH 4,8
(nepotpunog ponovnog svijanja u
0,015 M acetatnom puferu 0,15 M
NaCl, pH 4,8).
▄ CD spektar intermedijata svijanja sličan spektru nativnog
proteina, ali je nešto nižeg intenziteta
▄ Pokazuje vrijednost amplitude približno 66% od amplitude
tercijarne strukture nativnog proteina .
▄ Dakle, radi se o intermedijatu „nativnom stanju sličnom“, koji
nastaje u uslovima nepotpunog refoldinga kod pH 4,8 (0,015 M
acetatnom puferu 0,15 M NaCl), tj. uslovima pH u kojima se
opaža nastajanje fibrila izo-oblika divljeg-tipa (wt) čovječijeg
stefina B.

stB(Y31)
-5000
-3000
-1000
1000
195 205 215 225 235 245
λ [nm]
deg
cm
2 d
mo
l-1
stB(Y31)
Daleki UV CD spektar od 250 do 195
nm izo-oblika divljeg-tipa (wt)
čovječijeg stefina B (Y31) snimljen
nakon denaturacije u 0,02 M TEA
puferu i nepotpunog ponovnog
svijanja u 0,015 M acetatnom
puferu 0,15 M NaCl, pH 4,8.
▄ „Nativnom stanju sličan“ intermedijat, nastao u
blago kiseloj sredini kod pH 4,8 (0,015 M acetatnom
puferu 0,15 M NaCl, pH 4,8) imao je u potpunosti
uspostavljenu sekundarnu strukturu.

Kinetika svijanja izo-oblika divljeg-tipa (wt)
čovječijeg stefina B (Y31) i njegovog mutanta
H75W(Y31)
Metode za određivanje kinetičkih faza
svijanja i konstanti brzine svijanja proteina
metoda ANS (1-anilino-
naftalen 8–sulfonat)
fluorescence
metoda ANS (1-anilino-
naftalen 8–sulfonat)
fluorescence
metoda Trp fluorescence
metoda ThT fluorescence
H75W(Y31) stB(Y31)

Kinetika svijanja izo-oblika divljeg-tipa (wt)
čovječijeg stefina B (Y31) praćena ANS
fluorescencom kod pH 4,8
6040200
9,2e+005
8,8e+005
8,4e+005
8e+005
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)
ANS fluorescenca; 3 mM ANS; pH=4,8; 36°C; k2
stB(Y31)
A2
80006000400020000
1,4e+006
1,2e+006
1e+006
8e+005
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)
ANS fluorescenca; 3 mM; pH=4,8; 36°C; k3
stB(Y31)
A3
8642
9,2e+005
8,8e+005
8,4e+005
8e+005
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)
ANS fluorescenca; 3 mM ANS; pH=4,8; 36°C; k1
stB(Y31)
A1
▄ U trećoj kinetičkoj fazi svijanja,
koja se odvijala u intervalu od 60 do
7000 s, sa konstantom brzine k3,
došlo je do porasta ANS
fluorescence i stvaranja „nativnom
stanju sličnih“ intermedijata.

1512963
92000
88000
84000
80000
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)
ANS fluorescenca; 3 mM ANS; pH=5,8; 36°C; k1
stB(Y31)
80604020
90000
87000
84000
81000
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)
ANS fluorescenca; 3 mM ANS; pH=5,8; 36°C; k2
stB(Y31)
9006003000
96000
92000
88000
84000
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)
ANS fluorescenca; 3 mM ANS; pH=5,8; 36°C; k3
stB(Y31)
80006000400020000
96000
93000
90000
87000
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)
ANS fluorescenca; 3 mM ANS; pH=5,8; k4
stB(Y31)
▄ Kod pH 5,8 čovječiji stefin B (Y31) se svijao preko dva intermedijata, prvog
kratkoživućeg (15 s) i intermedijata 2 (900 s) nakon čega se u 7000 s
uspostavila nativna konformacija.
Kinetika svijanja izo-oblika divljeg-tipa (wt) čovječijeg stefina B (Y31) praćena
ANS fluorescencom kod pH 5,8

6040200
1,8e+005
1,6e+005
1,4e+005
1,2e+005
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)
ANS fluorescenca; 3 mM ANS; pH=7,0; 36°C; k3
stB(Y31)
80006000400020000
1,6e+005
1,4e+005
1,2e+005
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)
ANS fluorescenca; 3 mM ANS; pH=7,0; 36°C; k4
stB(Y31)
A1
A2
▄ Slična kinetika svijanja opažena je
i kod pH 7,0 s tim da su se
konformacije intermedijata
uspostavljale brže, a nativna
konformacija se počela
uspostavljati već nakon 60 s.
▄ Sa povećanjem pH od 4,8 na 5,8
odnosno 7,0 povećale su se
konstante brzina nastajanja
intermedijata svijanja, a smanjilo
se vrijeme njihovog života.
Kinetika svijanja izo-oblika divljeg-tipa (wt) čovječijeg
stefina B (Y31) praćena ANS fluorescencom kod pH
7,0

▄ ANS fluorescencom, Trp fluorescencom i ThT fluorescencom praćena
kinetika svijanja mutanta H75W(Y31) je pokazala da se protein kod pH 4,8 svijao u
jednoj kinetičkoj fazi do intermedijata, na osnovu čega se može zaključiti da se
njegov mehanizam svijanja razlikuje od mehanizma svijanja izo-oblika divljeg-
tipa (wt) čovječijeg stefina B (Y31) kod ovog pH.
80006000400020000
2e+005
1,6e+005
1,2e+005
80000
40000
Vrijeme [s]
Inte
nzit
et A
NS
flu
ore
scen
ce (
"a.u
.")
ANS fluorescenca; 3 mM ANS; pH=4,8; 36°C; k1
H75W
80006000400020000
1000
800
600
Vrijeme [s]
Inte
nzit
et T
rp f
luo
resc
ence
("a
.u."
)
Trp fluorescenca; pH=4,8; 36°C; k1
H75W
80006000400020000
52000
48000
44000
40000
36000
Vrijeme [s]
Inte
nzi
tet
ThT
flu
ore
scen
ce (
"a.u
.")
ThT fluorescenca; 6 M urea; pH=4,8; 36°C; k1
H75WT
rp f
luo
rescen
ca
Kinetika svijanja mutanta
H75W(Y31) kod pH 4,8

80006000400020000
6000
4000
2000
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)
ANS fluorescenca; 3 mM ANS; pH=5,8; 36°C; k1
H75W
80006000400020000
1000
800
600
400
Vrijeme [s]
Inte
nzi
tet
Trp
flu
ore
scen
ce (
"a.u
.")
Trp fluorescenca; pH=5,8; 36°C; k1
H75W
A
B
▄ ANS fluorescencom i Trp
fluorescencom praćena
kinetika svijanja mutanta
H75W(Y31) je pokazala da
se protein kod pH 5,8 svijao u
jednoj kinetičkoj fazi do
intermedijata što je slično
kinetici svijanja ovog proteina
kod pH 4,8.

20161284
1640
1600
1560
1520
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)ANS fluorescenca; 3 mM ANS; pH=7,0; 36°C; k1
H75W
8642
1920
1900
1880
1860
Vrijeme [s]
Inte
nzi
tet
Trp
flu
ore
scen
ce (
"a.u
.")
Trp fluorescenca; 3 mM ANS; pH=7,0; 36°C; k1
H75W
25002000150010005000
1650
1600
1550
1500
1450
1400
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)
ANS fluorescenca; 3 mM ANS; pH=7,0; 36°C; k2
H75W
12010080604020
1850
1800
1750
1700
Vrijeme [s]
Inte
nzi
tet
Trp
flu
ore
scen
ce (
"a.u
.")
Trp fluorescenca; 3 mM ANS; pH=7,0; 36°C; k2
H75W
6800640060005600
1480
1440
1400
1360
Vrijeme [s]
Inte
nzi
tet
AN
S f
luo
resc
ence
("a
.u."
)
ANS fluorescenca; 3 mM ANS; pH=7,0; 36°C; k3
H75W
80006000400020000
2000
1800
1600
Vrijeme [s]
Inte
nzi
tet
Trp
flu
ore
scen
ce (
"a.u
.")
Trp fluorescenca; 3 mM ANS; pH=7,0; 36°C; k3
H75W
▄ Kod pH 7,0 mutant H75W se svijao u tri kinetičke faze do
uspostavljanja strukture intermedijata 2, analogno svijanju čovječijeg
stefina B (Y31) na pH 4,8.

Puno zdravlja, ljubavi, sreće i uspjeha na predstojećim
ispitnim rokovima u novoj 2015. godini