universiti putra malaysia the nucleotide...
TRANSCRIPT

UNIVERSITI PUTRA MALAYSIA
THE NUCLEOTIDE SEQUENCE OF THE MATRIX (M) PROTEIN GENE OF NEWCASTLE DISEASE VIRUS (NDV) STRAIN AF 2240
SITI FATHIMAH PUTERY JEMAIN
FSAS 1999 31

THE NUCLEOTIDE SEQUENCE OF THE MATRIX (M) PROTEIN GENE OF NEWCASTLE DISEASE VIRUS (NDV) STRAIN AF 2240
By
SITI FATHIMAH PUTERY JEMAIN
Thesis Submitted in Fulfilment of the Requirements for the Degree of Master of Science in the Faculty of
Science and Environmental Studies Universiti Putra Malaysia
March 1999

ACKNO�EDGEMENTS
Bismillahirrahmanirrahim, I would like to express my most sincere
thanks and appreciation to my Chairperson, Associate Prof. Dr. Khatijah Yusoff,
Department of Biochemistry and Microbiology, Universiti Putra Malaysia, for
her guidance, support, understanding, encouragement, patience and
constructive suggestions throughout the course of study. I also wish to convey
my thanks to Associate Prof. Dr. Abdullah Sipat and Dr. Abd.Rahman Omar for
their support and valuable suggestions during the course of this research.
I would like to convey my heartfelt thanks and gratitude to my lab-mates,
Omeima, Subha, Vijay, Alan, Laili, Filza, Ainu, Wai Kean, Chiew Ling, Majid,
Jeeven, Wei Hong, Chui Fung, Sing King, Siti and Priya for the friendship,
support and encouragement and for making the 2 years stay, an enjoyable one.
My deepest gratitude and thanks especially go to MSMN, SRA, my
parents, brothers and sisters, my beloved husband - abang Mustapha, and the
'Family' especially, mama, ibu, Azu, Nor, kak Yam, Uncle Ali and Dr. Nyonya
for the continued support and prayers.
ii

TABLE OF CONTENTS
Page
ACKNOWLEDGEMENTS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ii LIST OF TABLES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . vi LIST OF FIGURES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . vii LIST OF PLATES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . viii LIST OF ABBREVIATIONS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ix ABSTRACT. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xi ABSTRAK. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xiii
CHAPTER
I
II
III
INTRODUCTION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 LITERATURE REVIEW . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 Newcastle Disease. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
Newcastle Disease Virus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4 Ma trix Protein. . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . .. . . . . .. . . . . ... 7
Sequence Characteristics.. .. . . . . . . . . . . . . . . . . . . . . . . . . . . 7 Functions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9 Physical Location in the Virus Particle. . . . . . . . . . . . 10 Changes During Infection. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
Polymerase Chain Reaction . . . . . . . . . . . . . . . . . " . .. . . . . .. . .. . . . . . 12 RNA (RT) PCR. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13 Contamination. . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . 15
DNA Sequencing. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16 Dideoxy Chain Termination. . . . . . . . . . . . . . . . . . . . . . . . 17 Cycle Sequencing . . . , . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
New Sequence Determination Strategies. . . . . . . . . . . . . . .. . . 20 Summary. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21 Objectives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22 MATERIALS AND METHODS . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 General Procedures. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 Virus Isolate. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 Preparation of RNA Template . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
iii

N
Virus Cultivation. . .. . . . . . . . . . . . . . . . . . . . . . .. . . ...... . . . .. 24 Harvesting Technique.. . . . .. . . . . .. . . . . ..... . ... . . . .. . .. 25 Purification of the Virus . . .. . . . . . . . .. . ... ..... . . .. . . ... 25 Viral RNA Extraction. . . . . . . ... . . . ... . . . . . . ... . .. . . . . .. . . 26
Primers Design and Selection . . . . . . . . . , . . . . . . . . . . . . . .. . . . . . . . .. 27 Reverse Transcription Polymerase Chain Reaction (RT-PCR) . . .. . . . . .. . . . ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . ... . .. . . . . . . . . . . 29 Cloning of RT-PCR Product into Vector pCR™2.1... . . . .. 31
Ligation. . . . . . . . . . . . . . .. . . . . .. . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31 Transformation.... . . . .... . . . . . . . . . . . . . . . . . . .. . .. . . . . . . . .. 32 Competent Cells . .... . . . . . . . . . . . . . . . . . ... . .. . . . ... ... . . . . . 36 Growth of Bacterial Colony. . . . . . . . . . . .. . . . . .. . . . . . . . . 37 Plasmid Extraction.. . . . . . . . . . . .. . . . . . .. . . . . ... . . . . .. . . . . . 37 Restriction Enzyme Analysis...... . . . . ... . . . . .. . . . . . . . 39 Plasmid PCR. . . . ... . . . ... . . . . .. . . .. .... . . .. . . . . . . . .. . . . ... 39
Colony Hybridisation.'... . . . . .. . . . . . . . . . . .. . . . ... . . . . . . ... . ... .. 40 Pro be Labeling. . . . . . . . . . . . . . . . . .. . . . . . . . .. . .. . . . . .. . . . . . . 40 Colony Lifts ... . . .... . . .. . . . . ... . . . .. . . . . . . . .. . . . . . , . . . . . , . 41 Hybridisation.. . . . .. . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . 42 Stringency Washes . . . , . . . .. , . . . . . . . .. . . . . . . . .. . . . . .. . . . . . 43 Colorimetric Detection with NBT and BCIP. . . . 43
Cycle Sequencing. . . . .. . . . . . . . . . . . . ... . . . . . . . . . . ... . . . . . . . . . . . . . .. 44 Computer Analysis of Nucleotide Sequence. . . . . . . . . . . .. . . 46
RESULTS AND DISCUSSION . . . .. . . . .. . . . . . .. . . . . . . . . . . . . .. . . 47 Amplification of the Matrix (M) Protein Gene. . . . .. . . . . . .. 47 Ligation and Transformation. . . . ... . . . . . .. . . . . . . . . ... . .. . . . . . . . 51 Colony Hybridisation. ... . .. . . . ... . . . . . . ... . . . .. . . . . . . . . .. . . . . . . . 54 Restriction Enzyme Analysis. . . . ... . . . . . . . . . . ... . . . .. . . . . . . . .. 56 Plasmid PCR. . . . ... .. . . .. . . . . .. . . . . . . . .. . ... . . . .. . . ... . . . . . .. . . . . .. 60 Sequencing of the M Gene.... . .. . . . . . . . . .. . . . . . . ..... . .. . . . . . . . 62
Problems Encountered.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63 Sequence Analysis. . . . . . . . . . . . . . . .... . . . .. ... .. . . ... . . .. . . . .. . . ... 65
FASTA Search Results.. . .. . . . . . . . . .... . . . . .. . . . .. . . . . . . . 65 Physical Properties of the M Protein. . . . . . . . . . . . . . . . . . . . . . . . . 68
The Amino Acid Composition and Molecular Weight. .. . . ... . .... .. . . .. . .. . . . . . . . .. . . . ... . . . . . . . . . . . . . . . .. 68
Structural Properties of the M Protein. . . . . . . . . . . . . . . . . . . . ... 73 Hydrophobicity Patterns . . ... . . . . . . .... . . . . . ... . . . . . . .. 73
Secondary Structure. . .... . . . . . .. . .. . . . . . . . . . . . . . .. . . . . . . . . . . . . . .. 77 Non-graphical Prediction. .. ... .. . . . . . . . . . ... . . .. . . . . . . 78
Phylogenetic Analysis. . ... . . .. . . .. . .. . . . . . . . .. . . . . . . .... . . . . . . . . 82
iv

v
Distance Matrix Methods. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82 Maximum Likelihood Method. . . . . . . . . . . . . . . . . . . . . . . . . 85 Consensus Tree. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 86
NDV Phylogenetic Grouping. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87 The Grouping of Strain AF 2240 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . , . 90 Multiple Alignment of the Amino Acids Sequence. . . . . . . . 90 Comparison Between the Amino Acid of the M Protein of AF 2240 and the M Protein of Other Strains of NDV and Other Members of the Paramyxoviridae Family. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 100 Restriction Enzyme Sites on the M gene. . . . . . . . . . . . . . . . . . . . . . 101
CONCLUSION . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 104
REFERENCES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 107
APPENDICES. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . 114
Appendix A : Hydropathy Plot for M Protein of Strain Turkey IND. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 115
Hydropathy Plot for M Protein of Strain Largo. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 116
Hydropathy Plot for M Protein of Strain Hertz/33. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 117
Appendix B: Evolutionary tree of 13 NDV Strains. . . . . . . . . 118 Appendix C: Nucleotide Sequence of the M Gene of
NDV strain AF 2240 Submitted to the GenBank . . . . . . . . . . . . . . . , . . . . . . . . . . . . . . . . . . . . . . . . . . . 119
VITA........................................................................... 121
v

LIST OF TABLES
Table Page
1 Function of the NDV Proteins. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
2 Potential Sources of Nucleic Acid Contamination in PCR. .. . . . . . . . . 16
3 Primers for RT-PCR and Sequencing. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
4 Universal Primers used for Sequencing......... . .......... . . . . . ...... . . . 28
5 Features of pCR™2.1. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . 35
6 Results from the FASTA Program. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 66
7 Results from the BLASTP Program. . . . . . . . . . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . 102
8 List of Enzymes that are Able to Cut the M Gene. . . . . . . . . . . . . .. . . . . . . .. 103
9 List of Enzymes that are Not Able to Cut the M Gene. . . . . . . . . . . . .. . ... 103
vi

LIST OF FIGURES
Figure Page
1 Schematic Diagram of a Typical Paramyxovirus Particle. . . . . . . . . . . 6
2 The Concept Behind the TA Cloning® Method . . .. . . . . . . . . . . . . . . . . . . . . . . 33
3 The pCR™2.1 Vector. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 34
4 The Nucleotide Sequence of the M Gene of Strain AF 2240. . . . . . . . . 69
Sa Hydropathy Profile of the M Gene Amino Acid Residues of NDV Strain AF 2240. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74
5b Hydropathy Profile of the M GeneAmino Acid Residues of NDV Strain Fontana . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 75
6 Non-graphical Representation of AF 2240 M Protein. . . . . . . . . . . . . . . . . 80
7 Consensus Tree Generated by BOOTSTRAP Resamplings (1000 bootstrap reiterations) Using the ECONSENSUS Programs. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88
8 Multiple Sequence Alignment of 13 NDV Strains Using the PREITYPLOT Program . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91
9 Hydropathy Plot for M Protein of Strain Turkey /ND. . . . . . . . . . . . . . . .. 115
10 Hydropathy Plot for M Protein of Strain Largo... . . . . . . . . . . . . . . . . . . . . . . 116
11 Hydropathy Plot for M Protein of Strain Hertz/33. . . . . . . . . . . . . . . . . . . . . 117
12 EvolUtionary Tree of 13 NDV Strains Based on the HN Gene. . . . . . .. 118
vii

LIST OF PLATES
Plate
1 RT-PCR Amplification of the M Gene of NDV
Page
Strain AF 2240 ................................................................... 49
2 The Colonies on the LB Plate with X-GaljIPTG....... .......... ......... 53
3 Colony Hybridisation of the Putative Recombinants................... 55
4 Eco RI Enzyme Analysis of Putative Recombinant Plasmid 01 and P5a.......................................................................... 57
5 Eco RI Enzyme Analysis of Plasmids 01,P5a and P5a'.................. 59
6 PCR of the Insert in the Recombinant Plasmid........................... 61
7 Sequence Ladders of the Cloned RT-PCR Product of the M Gene... 64
viii

LIST OF ABBREVIATIONS
The following abbreviations were used in the text:
bp
kDa
kb
min
s
v/v
w/v
1-11
NDV
RT-PCR
Mbp
M
st
NDU 25828
NDU 25829
NDU 25830
base pair
degrees Centigrade
kilodalton
kilo basepair
minute
second
volume per volume
weight per volume
microlitre
Newcastle disease virus
reverse transcription polymerase chain reaction
mega base pair
matrix
strain
NDV strain Bl NDV strain Fontana
NDV strain Hertz
ix

NDU 25831 NDV strain Kimber
NDU 25832 NDV strain Largo
NDU25833 NDV strain La Sota
NDU 25834 Queensland/V 4
NDU 25835 Texas/GB
NDU 25836 Turkey/ND
NDU25837 NDV strain Ulster
NDU 25838 NDV strain VGGA
VMAT_NDVA NDV strain Australia/Victoria/32
VMAT_NDVB NDV strain Beudette C/45
x

Abstract of thesis presented to the Senate of Universiti Putra Malaysia in fulfillment of the requirements for the degree of Master of Science.
THE NUCLEOTIDE SEQUENCE OF THE MATRIX (M) PROTEIN GENE OF NEWCASTLE DISEASE VIRUS (NDV) AF 2240
By
SITI FATHIMAH PUTERY JEMAIN
March 1999
Chairperson: Associate Professor Khatijah Yusoff, Ph.D.
Faculty: Science and Environmental Studies
The complete nucleotide sequence of the matrix (M) protein gene of the
local Newcastle disease virus (NDV) strain AF 2240 was determined. Based on
consensus primers, several segments of the M gene were reverse transcribed
and amplified by the polymerase chain reaction. The RT-PCR products were
cloned into a plasmid vector pCR™2.1. The DNA inserts in the clones were then
cycle-sequenced. The start and polyadenylation signals have been identified.
Assuming that the M gene starts and terminates at these sequences, the M gene
is 1223 nucleotides long and encodes an open reading frame of 364 amino acids,
corresponding to a polypeptide calculated molecular weight of 40 kDa. The M
protein, is both hydrophobic and basic. Phylogenetic analysis shows that the
strain has a close relationship with the North American Fontana strain. The M
protein amino acid sequence also shows similarities with members of the
xi

Rubulavirus genus such as the Simian virus 5, the human parainfluenza virus
type 4 and the mumps virus. Similarities of the amino acid sequence also exist
between the local strain and other members of the Paramyxovirus and
Morbillivirus genus.
xii

Abstrak tesis yang dikemukakan kepada Senat Dniversiti Putra Malaysia sebagai memenuhi keperluan untuk ijazah Master Sains.
JDJUKAN NUKLEOTIDA GEN PROTEIN MATRIKS (M) VIRUS PENYAKIT SAMPAR AYAM (NOV) STRAIN AF 2240
Oleh
SITI FATHIMAH PUTERY JEMAIN
Mac 1999
Pengerusi: Profesor Madya Khatijah Yusoff, Ph.D.
Fakulti : Sains dan Pengajian Alam Sekitar
Gen protein matriks (M) virus penyakit sampar ayam (NDV) strain AF
2240, yang merupakan strain tempatan, belum di ketahui jujukan
nukleotidanya. Oleh itu kajian ini dijalankan untuk mengenalpasti jujukan
nukleotida protein M tersebut. Produk RT-PCR gen M diklonkan ke dalam
vektor plasmid pCR™2.1. Jujukan produk RT-PCR gen M yang telah diklonkan
tadi ditentukan melalui proses penjujukan berulang. Kodon-kodon permulaan
dan isyarat poliadenilat telah dikenalpasti. Dengan mengambil kira kodon-
kodon ini, jujukan nukleotida protein M didapati mempunyai 1223 pasangan
bes dan berupaya untuk mengkodkan protein yang mempunyai 364 asid-asid
amino. Anggaran berat molekular protein M ialah lebih kurang 40 kDa. Secara
keseluruhannya, jujukan asid amino protein M adalah bersifat hidrofobik dan
basik. Analisis filogenetik, menunjukkan bahawa strain AF 2240 mempunyai
xiii

pertalian yang rapat dengan strain NDV daripada Amerika Utara, iaitu strain
Fontana. Jujukan asid amino protein M mempunyai persamaan dengan ahli
ahli genus Rubulavirus yang lain, iaitu virus Simian 5, virus parainfluenza
manusia jenis 4 dan virus penyakit begok dan juga ahli genus Paramyxovirus
dan Morbillivirus.
xiv

CHAPTER I
INTRODUCTION
Newcastle disease or 'penyakit sampar ayam' is endemic in
Malaysia and has always been a serious threat to the livestock industry.
This disease is caused by the Newcastle disease virus (NDV) which
belongs to the Rubulavirus genus within the Paramyxoviridae family. The
disease currently has a worldwide distribution with a wide host range
in which all orders of birds have been reported to be infected by NDV
(Brandly, 1964). From the Office International Des Epizooties (O.I.E)
homepage (http://ss.niah.affrc.go.jp/OIE/yb95/yb95b.html), it was
shown that there has been 11 reported outbreaks of ND in Malaysia and
frequent outbreaks also occur in Indonesia, the Philippines, India, South
Korea and other developing countries from January to August 1995.
The techniques used to eradicate the virus are (1) destruction of
the entire chicken population and disinfection of the affected area with
viricidal agents, (2) employing quarantine procedures and (3)

2
vaccination of the chickens (Spradbrow, 1987). Vaccination has so far given an
adequate protection against NDV, at least for chickens reared commercially.
Two types of vaccines are currently being used against Newcastle disease; a live
attenuated vaccine (for example, strains Bl and La Sota) usually administered in
drinking water and the inactivated vaccine (for example, strain Ulster) usually
administered by intra-muscular injection. In addition, researchers in UPM has
used the lentogenic strain V 4 (UPM) as a live attenuated vaccine and in the form
of food pellet (Ideris et al., 1990).
The rapid advance in recombinant DNA technology has given birth to a
new generation of genetically engineered vaccine that has several advantages
over the traditional vaccines. These advantages include (1) greater safety to
workers and the environment because only the usage of isolated genes (cloned
into specific vectors) are involved, (2) vaccine associated complications are
reduced because non-essential viral components are not present in the vaccine,
(3) purified proteins are more stable than virus particles and (4) cheaper
production cost (Obijeski, 1985).
The local velogenic-viscerotropic NDV virus reference strain AF 2240, has
been studied in UPM for the past few years. This strain has the potential to be

3
developed as a subunit vaccine capable of protecting poultry against a wider
range of NDV isolates. Being heat resistant, its transportation and handling will
be cheaper and easier as it will limit the number of essential 'cold-chains'
involved in maintaining its effectiveness. Unlike the haemagglutinin-
neuraminidase (HN) protein and the fusion (F) protein genes of strain AF 2240
(Tan et al., 1995; Yusoff et al., 1993) which have been extensively studied, very
little work has been done on the matrix (M) protein gene. The M protein gene
encodes the M protein, which plays an important role in the construction of the
virus as well as the regulation of the viral RNA synthesis (Tanabayashi et al.,
1990; Seal, 1996). It is hoped that knowledge of the M gene sequence will give a
better understanding of its structure and function.
This thesis reports on the nucleotide sequence of the M gene of NDV
strain AF 2240.

CHAPTER II
LITERATURE REVIEW
Newcastle Disease
Newcastle disease which causes 100% mortality in birds
(Brandly, 1964), was discovered in 1926 near Batavia (Jakarta) . A similar
outbreak was also reported in Newcastle-on-Tyne (hence, the name of
the disease) and in Korea in that same year. By 1933, researchers in
England, Philippines, Indonesia and India, through cross-immunity
tests, agreed that the same virus named as the Newcastle disease virus
caused these outbreaks.
Newcastle Disease Virus
The Newcastle disease virus, the human parainfluenza virus
types 2, 4a and 4b, the Simian virus 5 and mumps virus are grouped
under the genus Rubulavirus (Lamb and Kolakofsky, 1996).
4

5
The Paramyxoviridae family was reclassified in 1993 into two subfamilies: the
Paramyxovirinae and the Pneumovirinae. The Paramyxovirinae contains three genera,
Parainjluenzavirus, Rubulavirus and Morbillivirus. The sub-family Pneumovirinae
contains the genus Pneumovirus. The new classsification is based on morphological
criteria, genomic organisation, biological activities of proteins, and the sequence
relationship of the encoded proteins (Lamb and Kolakofsky, 1996).
Members of this family of viruses are very pleomorphic, with a diameter of
about 100 to 500 nm, and are capable of acquiring a variety of shapes ranging from
circular, spherical, flattened, and often in filamentous forms. The virion is enveloped
with a lipid bilayer membrane, derived from the plasma membrane of the host cell,
which gives it a certain flexibility to exist in these various morphological forms suited
to the pH and salinity of the surrounding environment (Waterson, 1964). The
haem agglutinin-neuraminidase (HN) and fusion (F) glycoproteins protrude from the
lipid bilayer membrane. They are visible as spike-like structures under the electron
microscope, ranging from about 8 to 20 nm in length and spaced 6 to 10 nm apart
Below the lipid bilayer membrane is a shell of protein known as the matrix (M) protein. Protected by the lipid and protein layers, is the long helical nucleocapsid 13
to 18 nm in diameter with the "herring-bone" structure quite unique to NDV,
containing the genome (Figure. 1) (Alexander, 1988; Samson, 1988).

NP Protei n
RNA Genome
P Protei n
M = matrix protein F = fusion protein consists of 2 disulphide-linked subunits (Fl+F2) HN = haemagglutinin-neuraminidase protein NP = nucleoprotein L = large protein P = nucleocapsid associated protein
Figure 1: Schematic Diagram of a Typical Paramyxovirus Particle.
6
( www.tulane.edu)

7
The genome of NDV is a non-segmented, single stranded linear RNA of
negative polarity with a molecular weight of 5.2 to 5.7xl06 daltons which is
approximately 15 kilobase (kb) long. It codes for the following six structural
proteins nucleocapsid protein (NP), nucleocapsid associated protein (P), matrix
protein (M), fusion protein (F), haemagglutinin-neuraminidase (HN) protein
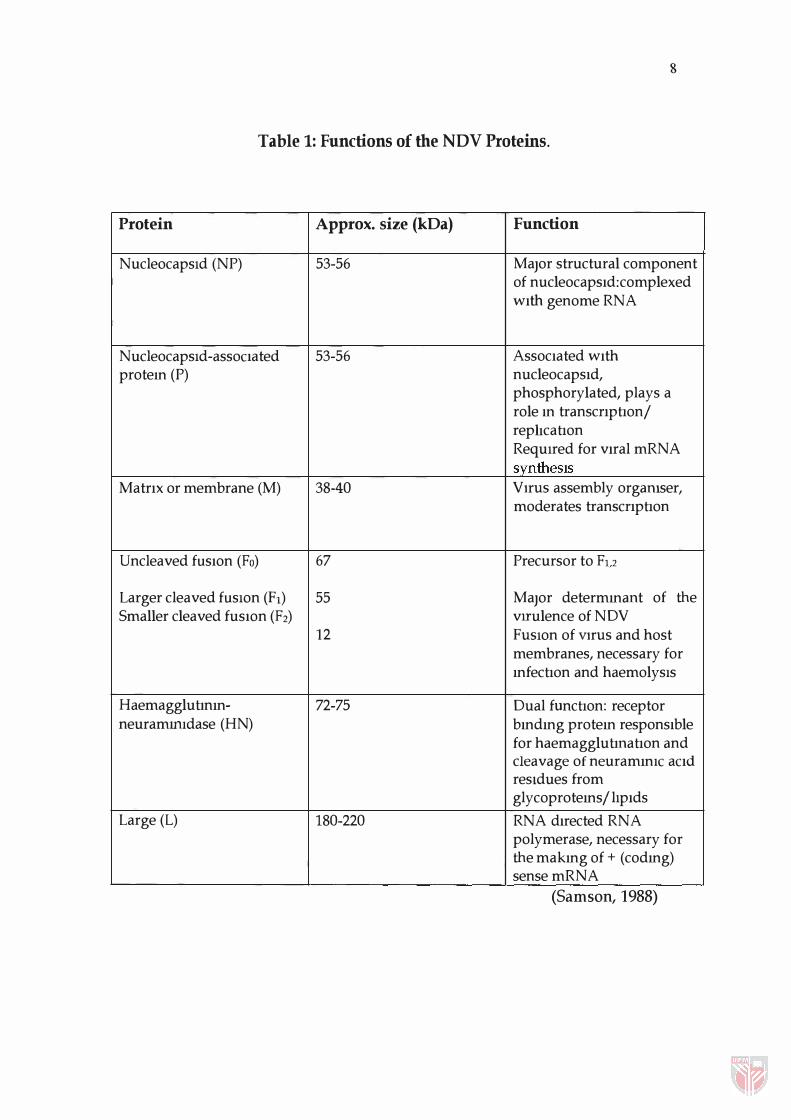
and large (L) polymerase protein (Samson, 1988) (Table 1).
Matrix (M) Protein
Sequence Characteristics
The matrix (M) protein is a non-glycosylated, non-phosphorylated,
hydrophobic and highly basic protein of 364 amino acids (Chambers et aI., 1986;
McGinnes and Morrison, 1987; Seal, 1996). Most of its positively charged basic
amino acids lie in the C-terminal portion of the molecule. It is reported that the
amino acid sequence contains five cysteine residues (McGinnes and Morrison,
1987). The positions of glycine, proline and paired basic amino acid residues (R
K, K-K, K-R, R-R) of the M gene have been shown to be conserved in NDV,
8
Table 1: Functions of the NDV Proteins.
Protein Approx. size (kDa) Function
NucleocapsId (NP) 53-56 Major structural component of nucleocapsId:complexed WIth genome RNA
NucleocapsId-assoCIated 53-56 AssocIated WIth protem (P) nucleocapsId,
phosphorylated, plays a role m transcnphon/ rephcahon RequIred for vIral mRNA synthesIS
MatrIx or membrane (M) 38-40 VIrUS assembly organiser, moderates transcnphon
Uncleaved fusIOn (Fa) 67 Precursor to F1,2
Larger cleaved fusIOn (Fl) 55 Major determmant of the Smaller cleaved fusIOn (F2) vrrulence of NDV
12 FUSIOn of VIrUS and host membranes, necessary for mfechon and haemolysis
Haemaggluhnm- 72-75 Dual funchon: receptor neurammidase (HN) bmdmg protem responsIble
for haemaggluhnahon and cleavage of neurammiC aCId resIdues from glycoprotems / lIpIds
Large (L) 180-220 RNA dIrected RNA polymerase, necessary for the makmg of + (codmg) sense mRNA
(Samson, 1988)

9
Send ai, measles and canine distemper viruses (Bellini et al., 1986; Chambers et al.,
1986; McGinnes and Morrison, 1987).
The M protein is one of the main structural elements of the virus. However,
the functioning of this M protein is suggested to be mediated by its overall
characteristics (basic and hydrophobic), rather than by any particular conserved
sequences (Bellini et al., 1986; Chambers et al., 1986; Spriggs et al., 1987; Elango 1989;
Limo and Yilma., 1990; Tanabayashi et ai ., 1990; Randhawa et al., 1996).
Functions
Firstly, the M protein seems to be a controlling factor in RNA synthesis and
has a key role in virus assembly by locating nucleocapsid structures beneath those
regions of the plasma membrane in which the F and HN glycoproteins are
attached. This was proven when paramyxoviruses that produced defective M
proteins were unable to produce virus particles (Peeples and Bratt, 1984). This was
perfectly demonstrated in vitro when the Sendai virus nucleocapsid would not

10
form a complex with the viral glycoproteins unless the M protein was added.
Secondly, it could be involved in moderating the virion RNA polymerase activity
possibly in interactions with cellular actin (negatively charged) which drives the
budding process. Thirdly, it could be involved in protein kinase activity
(Blumberg et al., 1984). Analysis of chimeric M proteins indicates that mutations
in the amino-terminal and the carboxyl-terminal regions of the M protein all
abrogate nucleocapsid binding. This suggests that the M protein conformation
is important for interaction with the viral nucleocapsid (Hirano et al., 1993).
Physical Location in the Virus Particle
In electron micrographs of virions (Lamb and Kolakofsky, 1996), an
electron-dense layer is observed underlying the viral lipid bilayer. This is
thought to represent the location of the M protein. Fractionation studies of the
virus indicate that this protein is peripherally associated with membranes and is
not an intrinsic membrane protein (Faaberg and Peeples, 1988). The M protein
probably contains amphipathic oc-helices that insert themselves into the inner
leaflet of the lipid bilayer to coat this surface and organise its contacts with the