universidade federal do cearÁ faculdade de … · figura 1 aspecto morfológico do h. capsulatum...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO CEARÁ
FACULDADE DE MEDICINA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS MÉDICAS
ÂNGELA DONATO MAIA MALAQUIAS
EFEITO INIBITÓRIO IN VITRO DE MILTEFOSINA FRENTE À CEPAS
DE HISTOPLASMA CAPSULATUM VAR. CAPSULATUM E SPOROTHRIX SPP.
FORTALEZA
2013

ÂNGELA DONATO MAIA MALAQUIAS
EFEITO INIBITÓRIO IN VITRO DE MILTEFOSINA FRENTE A CEPAS DE
HISTOPLASMA CAPSULATUM VAR. CAPSULATUM E SPOROTHRIX SPP.
Dissertação submetida ao Curso de Pós-
Graduação em Ciências Médicas, da
Faculdade de Medicina, da Universidade
Federal do Ceará, como requisito parcial para
obtenção do grau de Mestre.
Orientadora: Profª. Drª. Raimunda Sâmia
Nogueira Brilhante
Coorientador: Prof. Dr. José Júlio Costa
Sidrim
FORTALEZA
2013

Dados Internacionais de Catalogação na Publicação
Universidade Federal do Ceará
Biblioteca de Ciências da Saúde
M197e Malaquias, Ângela Donato Maia.
Efeito inibitório in vitro de miltefosina frente a cepas de Histoplasma capsulatum var.
capsulatum e sporothrix spp./ Ângela Donato Maia Malaquias. – 2013.
86 f. : il.
Dissertação (Mestrado) – Universidade Federal do Ceará. Faculdade de Medicina. Programa de
Pós-graduação em Ciências Médicas, Fortaleza, 2013.
Orientação: Profa. Dra. Raimunda Sâmia Nogueira Brilhante.
1. Histoplasma. 2. Sporothrix. 3. Antifúngicos. 4. Testes de Sensibilidade Microbiana. I.
Título.
CDD 616.9041

ÂNGELA DONATO MAIA MALAQUIAS
EFEITO INIBITÓRIO IN VITRO DE MILTEFOSINA FRENTE A CEPAS DE HISTOPLASMA
CAPSULATUM VAR. CAPSULATUM E SPOROTHRIX SPP.
Dissertação submetida ao Curso de Pós-
Graduação em Ciências Médicas, da
Faculdade de Medicina, da Universidade
Federal do Ceará, como requisito parcial para
obtenção do grau de Mestre.
Aprovada em:
BANCA EXAMINADORA
________________________________________________________________
Profa. Dra. Raimunda Sâmia Nogueira Brilhante (Orientadora)
Universidade Federal do Ceará
_________________________________________________________________
Prof. Dr. Marcos Fábio Gadelha Rocha
Universidade Estadual do Ceará
________________________________________________________________
Profa. Dra. Tereza de Jesus Pinheiro Gomes Bandeira
Faculdade Christus

Dedico este trabalho primeiramente a Deus, minha
fortaleza. Ao meu amado esposo Rodrigo Malaquias, meu
maior incentivador e companheiro de todos os momentos.
Aos meus pais Nerges Maia e Fátima Donato por me
ensinarem o verdadeiro valor de cada atitude que
tomamos na vida. As minhas queridas irmãs que sempre
me deram força na busca desse sonho. Amo vocês.

AGRADECIMENTOS
À Deus, por me ensinar que tudo na vida tem o seu momento. Por me mostrar que na vida
pode haver perdas, mas que na hora certa chegam grandes alegrias e vitórias. Obrigada pela
conclusão de mais uma fase de minha vida e por estar sempre ao meu lado guiando e
iluminando meus passos.
À banca examinadora, por terem aceitado participar da avaliação desse trabalho.
Ao Centro Especializado em Micologia Médica (CEMM), da Universidade Federal do Ceará.
De modo especial, à profª Drª Raimunda Sâmia Nogueira Brilhante, minha orientadora, por
todos os ensinamentos, pela compreensão, pela confiança, competência e incentivos
constantes. Obrigada por toda a dedicação e orientação durante meu mestrado.
Ao Prof. Dr. José Júlio Costa Sidrim, pelos ensinamentos e pela oportunidade de trabalhar no
Centro Especializado em Micologia Médica.
Ao Prof. Dr. Marcos Fábio Gadelha Rocha, exemplo de profissionalismo, agradeço por sua
imensa dedicação, orientação e pelas críticas construtivas durante a correção dos artigos
científicos que muito contribuíram para a minha formação profissional.
À Profa Dra. Rossana de Aguiar Cordeiro, pelo apoio, incentivo, esclarecimento de dúvidas e
ensinamentos transmitidos.
À Profa Dra Tereza Bandeira pelo apoio, incentivo e por transmitir paz em seus gestos e
palavras.
Ao prof. Dr. André Jalles Monteiro, pelo estudo estatístico realizado neste trabalho.
Ao Programa de Pós-Graduação em Ciências Médicas (PPGCM), da Faculdade de Medicina,
da Universidade Federal do Ceará.
À Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico - FUNCAP,
pelo apoio financeiro.
As secretárias do programa de pós-graduação em Ciências Médicas, Ivone Mary Fontenele de
Sousa e Rita de Cássia Antunes, por tudo que fizeram por mim.

A todos os companheiros do laboratório (Iniciação científica, Mestrandos e Doutorandos),
pelas conversas, pela ajuda e pelo incentivo durante a realização desse trabalho, em especial a
Érica Pacheco Caetano, pelos ensinamentos, pelo auxílio e pelo acolhimento, sem a qual a
realização desse trabalho teria se tornado bem mais difícil. A querida amiga Rita Amanda
Chaves que sempre se fez presente nos momento de alegrias e tristezas e não mediu esforços
para me ajudar na conclusão do meu mestrado. A Natalya Fechine, pela amizade, carinho e
ajuda nos experimentos. Jakelyne Marques que sempre esteve disposta a me ajudar, pela
amizade e companheirismo. A Débora Castelo-Branco pela troca de conhecimentos,
esclarecimento de dúvidas e por toda a força e apoio nos momentos difíceis da minha vida.
Devo parte dessa vitória a vocês.
Aos técnicos do CEMM Terezinha de Jesus Santos Rodrigues e Daniel Texeira de Lima por
todo o auxílio durante a realização desse trabalho.
Aos meus pais Nerges Andrade Maia e Maria de Fátima Rodrigues Donato, por serem o
alicerce da minha vida. Por todo o carinho, dedicação, compreensão e por ajudarem a me
tornar a cada dia uma pessoa melhor.
As minhas amadas irmãs, Ana Cristina e Alexsandra, que sempre estiveram ao meu lado.
Obrigada pela confiança, amizade e por tudo que nos une me dando força nos momentos mais
difíceis. Querida irmã Alexsandra, serás lembrada eternamente. Inesquecível irmã. Te amo.
Aos meus preciosos avós Djalma e Izete Donato, Marcondes e Miriam Maia por me
ensinarem o valor do estudo e serem um modelo para mim de moral e caráter.
Ao meu amado esposo Rodrigo Malaquias que esteve comigo a cada passo dessa jornada. Seu
caráter e coragem de encarar as dificuldades da vida me estimulam e incentivam. Te admiro
ainda mais pela compreensão, pelos conselhos, carinho e amor.
Ao meu querido filho Bento Gabriel, que ainda em meu ventre, já é a pessoa mais amada e
esperada desse mundo. Você foi enviado por Deus para encher nossas vidas de alegrias.
A minha sogra Luzia Freire, uma segunda mãe que Deus colocou em minha vida. Obrigada
por tudo.

RESUMO
Histoplasma capsulatum var. capsulatum e Sporothrix spp. são fungos dimórficos,
patogênicos, causadores da histoplasmose e da esporotricose, respectivamente, sendo capazes
de acometer o homem e outros animais. Embora a terapia antifúngica comumente utilizada
seja eficiente para o tratamento dessas micoses, algumas desvantagens podem ser apontadas,
como a necessidade de tratamento por tempo prolongado, bem como, a elevada toxicidade
apresentada pela anfotericina B. Ademais, casos de refratariedade e recidivas já foram
descritos. Esse cenário tem impulsionado a busca por novos agentes que apresentem atividade
antifúngica. Alguns estudos tem demonstrado que determinadas drogas já empregadas
clinicamente no tratamento de doenças de etiologia não-fúngica podem apresentar potencial
antifúngica relevante. A miltefosina, originalmente desenvolvida como agente anticâncer e
atualmente utilizada no tratamento da Leishmaniose, tem demonstrado uma importante
atividade antifúngica contra diferentes patógenos fúngicos. Baseado no exposto, o presente
trabalho teve por objetivo avaliar a atividade antifúngica in vitro da miltefosina ante aos
fungos dimorficos H. capsulatum var. capsulatum e as espécies do complexo S. schenckii,
incluindo S. brasiliensis, S. schenckii, S. globosa e S. mexicana, por meio da técnica de
microdiluição em caldo padronizada pelo CLSI. Os resultados obtidos mostraram que a
miltefosina possui efeito inibitório in vitro frente H. capsulatum var. capsulatum e Sporothrix
spp. sendo capaz de inibir o crescimento de todas as cepas analisadas. Os valores de
Concentração Inibitória Mínima (CIM) obtidos variaram de 0,25 a 2 µg mL-1
para H.
capsulatum var. capsulatum na fase filamentosa e de 0,125 a 1 µg mL-1
na fase leveduriforme.
Para Sporothrix spp. na fase filamentosa o intervalo de CIM foi de 0,25 a 2 µg mL-1
. Os
valores de Concentração Fungicida Mínima (CFM) foram ≤ 4 µg mL-1
para ambas as espécies
analisadas. Assim, o presente estudo demonstrou pela primeira vez a relevante atividade
antifúngica da miltefosina ante H. capsulatum var. capsulatum e Sporothrix spp. Vale
ressaltar, que mais estudos ainda são necessários para avaliar a aplicabilidade da miltefosina
in vivo e validar a sua utilização como uma alternativa para subsidiar o tratamento de
pacientes com histoplasmose e esporotricose.
Palavras-chave: Histoplasma. Sporothrix. Antifúngicos. Testes de Sensibilidade Microbiana.

ABSTRACT
Histoplasma capsulatum var. capsulatum and Sporothrix spp. are dimorphic fungi,
pathogenic, causing histoplasmosis and sporotrichosis, respectively, being able to affect
humans and other animals. Although the commonly used antifungal therapy is effective for
the treatment of such fungal infections, some drawbacks may be mentioned, as the need for
treatment for prolonged periods, as well as by the high toxicity of amphotericin B. Moreover,
refractory cases and recurrences have been described. This scenario has driven the search for
new agents that have antifungal activity. Some studies have shown that certain drugs already
used clinically in the treatment of non-fungal etiology diseases may have significant
antifungal potential. The miltefosine, originally developed as anticancer agent and currently
used in the treatment of leishmaniasis, has demonstrated a significant antifungal activity
against various fungal pathogens. Based on the above, this study aimed to evaluate the
antifungal activity in vitro of miltefosine against the dimorphic fungi H. capsulatum var.
capsulatum and S schenckii complex species including S. brasiliensis, S. schenckii, S. globosa
and S. mexican, by broth microdilution technique standard by CLSI. The results showed that
miltefosine inhibitory effect in vitro against H. capsulatum var. capsulatum and Sporothrix
spp. being able to inhibit the growth of all strains analyzed. The Minimum Inhibitory
Concentration (MIC) values obtained ranged from 0.25 to 2 mg.L-1
for H. capsulatum var.
capsulatum the filamentous phase and from 0.125 to 1 ug.mL-1
in the yeast phase. For
Sporothrix spp. filamentous phase in the MIC range was from 0.25 to 2 mg.L-1
. The
Minimum Fungicidal Concentration (MFC) values were ≤ 4 mg L-1
for both species analyzed.
Thus, the present study demonstrated for the first time the relevant antifungal activity of
miltefosine against H. capsulatum var. capsulatum and Sporothrix spp. It is noteworthy that
more studies are necessary to evaluate the applicability of miltefosine in vivo and validate its
use as an alternative to support the treatment of patients with histoplasmosis and
sporotrichosis.
Keywords: Histoplasma. Sporothrix. Antifungal Agents. Microbial Sensitivity Tests.

LISTA DE FIGURAS
Figura 1 Aspecto morfológico do H. capsulatum A. Macroscopia da fase filamentosa em Agar
batata, demonstrando colônia algodonosa branca; B. Micromorfologia da fase filamentosa,
mostrando hifas septadas e macroconídeos tuberculados; C. Macromorfologia da fase
leveduriforme em àgar BHI com sangue de carneiro 10%, mostrando colônias de coloração
creme com aspecto membranoso; D. Micromorfologia da fase leveduriforme, mostrando
células ovais uninucleadas e estrutura leveduriforme com brotamento Fonte: CEMM, 2011.
................................................................................................................................................ 17
Figura 2 Aspecto morfológico de Sporothrix schenckii. A. Macromorfologia na fase
filamentosa em Agar batata, mostrando colônia de branca a creme, de textura lisa, com
aspecto coreáceo; B. Micromorfologia, mostrando hifas finas e delicadas, com conídios nas
extremidades dos conidióforos assemelhando-se a rosetas (Fonte: CEMM, 2013); C.
Macromorfologia da fase leveduriforme em àgar sabouraud dextrose, mostrando colônia de
cor creme e aspecdto cremoso; D. Micromorfologia da fase leveduriforme, mostrando células
leveduriformes ovaladas (Fonte: CDC, 2013). ....................................................................... 18
Figura 3 Fórmula estrutural da miltefosina ........................................................................... 36

LISTA DE QUADROS E TABELAS
Quadro 1 Relação dos Isolados Clínicos de Histoplasma capsulatum, fase filamentosa,
estocados na micoteca do CEMM ......................................................................................... 44
Quadro 2 Relação dos Isolados Clínicos de Histoplasma capsulatum, fase leveduriforme,
utilizados na pesquisa ............................................................................................................ 46
Quadro 3 Relação das espécies do complexo Sporothrix schenckii estocados na micoteca do
CEMM .................................................................................................................................... 47
Quadro 4 Concentração Inibitória Mínima (CIM) (µg/mL) da droga Miltefosina (MIL), frente
as cepas de Histoplasma capsulatum var. capsulatum na fase filamentosa. ........................ 76
Quadro 5 Concentração Inibitória Mínima (CIM) (µg/mL) da droga Miltefosina (MIL), frente
as cepas de Histoplasma capsulatum var. capsulatum na fase Leveduriforme. ................... 78
Quadro 6 Concentração Inibitória Mínima (CIM) (µg/mL) da droga Miltefosina (MIL), frente
as cepas de Sporothrix spp. .................................................................................................. 79
Tabela 1 Atividade antifúngica in vitro de Miltefosina (MIL), frente as cepas de Histoplasma
capsulatum na fase filamentosa e Leveduriforme e Sporothrix spp.. ................................... 54

LISTA DE ABREVIATURAS
AFST-EUCAST Antifungal Susceptibility Testing Subcommittee of the European Committee on
Antimicrobial Susceptibility AIDS Acquired Immune Deficiency Syndrome
AMB Anfotericina B
ANVISA Agência Nacional de Vigilância Sanitária
ATCC American Type Culture Collection
BHI Brain Heart Infusion
CDC Centers for Disease Control And Prevention
CEMM Centro Especializado em Micologia Médica
CFM Concentração Fungicida mínima
CIM Concentração Inibitória Mínima
CLSI Clinical and Laboratory Standards Institute
COX Citocromo c oxidase
DMSO Dimetilsulfóxido
DNA Ácido Desoxirribonucléico
E-test Epsilometer Strip Test
EUCAST European Committee on Antimicrobial Susceptibility Testing
FDA Food and Drug Administration
HE Hematoxilina-Eosina
HIV Human Immunodeficiency Viru
ID Imunodifusão Dupla
IFN-γ Interferon gama
IL Interleucina
INF-γ Interferon-gama
ITC Itraconazol
MIL Miltefosina
µg Microgramas
mL Mililitros
MOPS Ácido 3-[N-morfolino] propanosulfônico
NB-3 Nível de biossegurança 3
OMS Organização Mundial de Saúde
PCR Polymerase Chain Reaction

RFC Reação de Fixação do Complemento
PKB Proteína-quinase B
RIA Radioimunoensaio
RPMI Roswell Park Memorial Institute
Th1 T-helper1
TNF-α Fator de necrose tumoral alfa

SUMÁRIO
1 INTRODUÇÃO ................................................................................................................. 14
2 REVISÃO DE LITERATURA ......................................................................................... 16
2.1 H. capsulatum var. capsulatum e Sporothrix spp. ........................................................ 16
2.1.1 Classificação Taxonômica ..................................................................................................... 16
2.1.2 Morfologia ........................................................................................................................ 17
2.1.3 Aspectos ecológicos ............................................................................................................. 19
2.2 Aspectos Gerais da Histoplasmose e da Esporotricose ............................................... 20
2.2.1 Epidemiologia ..................................................................................................................... 20
2.2.2 Patogênese e Imunidade ............................................................................................ ... 22
2.2.3 Quadros Clínicos ........................................................................................................... 24
2.2.4 Diagnóstico .................................................................................................................... 26
2.2.5 Tratamento, Controle e Prevenção ................................................................................ 29
2.3 Drogas Antifúngicas Clássicas e Testes de Sensibilidade in vitro ................................................. 31
2.3.1 Busca de novos agentes antifúngicos ....................................................................................... 36
2.3.1.1 Miltefosina ....................................................................................................................... 36
3 PERGUNTAS DE PARTIDA ........................................................................................... 39
4 HIPÓTESES ...................................................................................................................... 40
5 OBJETIVOS ........................................................................................................................ 41
5.1 Objetivo Geral .................................................................................................................. 41
5.2 Objetivos Específicos ....................................................................................................... 41
6 MATERIAL E MÉTODOS ................................................................................................... 42

6.1 Microrganismos ................................................................................................................ 42
6.2 Agentes antimicrobianos .................................................................................................. 49
6.3 Preparo do inóculo para teste de sensibilidade a drogas .................................................. 50
6.4 Testes de Sensibilidade in vitro ......................................................................................... 50
6.5 Análise estatística ............................................................................................................. 52
7 RESULTADOS .................................................................................................................. 53
8 DISCUSSÃO ...................................................................................................................... 55
9 CONCLUSÕES .................................................................................................................. 60
REFERÊNCIAS ................................................................................................................... 61
ANEXO A Quadros ............................................................................................................... 75
ANEXO B Corante, meios de cultura e soluções .................................................................. 82

14
1 INTRODUÇÃO
Diferentes tipos de infecções fúngicas, tornaram-se, nos últimos anos, um importante
problema de saúde publica (YAN et al., 2011). A histoplasmose e a esporotricose são doenças
causadas pelos fungos Histoplasma capsulatum var. capsulatum e Sporothrix spp.,
respectivamente. São fungos dimórficos, patogênicos sendo capazes de acometer o homem e
outros animais (SILVEIRA et al., 2009; MADRID, 2011b).
A frequência das micoses profundas e oportunistas vem aumentando no mundo inteiro
(BERGOLD; GEORGIADIS, 2004). O arsenal terapêutico disponível para micoses invasivas
é limitado e embora a terapia antifúngica comumente utilizada seja em geral eficiente no
tratamento dessas micoses, algumas desvantagens podem ser apontadas, como a necessidade
de tratamento por tempo prolongado, bem como, a elevada toxicidade apresentada pela
anfotericina B (WHEAT et al., 2007; KAUFFMAN et al., 2007). Além disso, casos de
refratariedade e recidivas já foram descritos para essas doenças (SCHUBACH et al., 2004;
WHEAT et al., 2006; KAUFFMAN et al., 2007).
Até a década de 1970, o tratamento das infecções fúngicas profundas e oportunistas
era constituído por um pequeno número de drogas de baixo espectro ou alta toxicidade
(MARTINEZ, 2006). Com a descoberta dos triazólicos e das equinocandinas, uma melhora
significativa na terapia dessas infecções foi observada, devido à elevada eficácia in vivo e
baixa toxicidade desses compostos (CAILLOT et al., 2007; PASQUALOTTO; DENNING,
2008). No entanto, a terapia atual disponível para o tratamento das micoses profundas ainda
está restrita a poucas classes de drogas (GONZÁLEZ, 2009). Neste contexto, pesquisas têm
sido realizadas no sentido de investigar novos agentes que não são utilizados na terapia
antifúngica, mas que podem apresentar um amplo espectro de ação frente a espécies fúngicas.
Alguns estudos tem demonstrado que determinadas drogas já empregadas clinicamente no
tratamento de doenças de etiologia não-fúngica podem apresentar atividade antifúngica
relevante (BRILHANTE et al., 2010; CORDEIRO et al., 2011a).
A miltefosina foi originalmente desenvolvida como agente anticâncer, contudo,
estudos posteriores constataram sua relevante atividade contra a Leishmania spp. e
Trypanosoma cruzi (CROFT; BARRETT; URBINA, 2005; SOTO; SOTO, 2006, LUNA et
al., 2009). Atualmente, também tem sido relatada uma importante atividade antifúngica contra
diferentes patógenos fúngicos, tais como leveduras dos gêneros Candida spp. e Cryptococcus

15
spp. e a fungos filamentosos como dermatófitos, Aspergillus spp., Fusarium spp. e
Scedosporium spp. (WIDMER et al., 2006; TONG et al., 2007).
Diante do exposto, este trabalho teve como objetivo avaliar pela primeira vez, a
atividade inibitória in vitro da miltefosina frente aos fungos dimórficos H. capsulatum var.
capsulatum e o complexo Sporothrix schenckii, através da determinação da Concentração
Inibitória Mínima por meio do ensaio de microdiluição em caldo. É importante ressaltar que
anteriormente nenhum estudo havia sido realizado sobre o potencial antifúngico da
miltefosina frente a estes patógenos.

16
2 REVISÃO DE LITERATURA
2.1 Histoplasma capsulatum e Sporothrix spp.
2.1.1 Classificação Taxonômica
Taxonomicamente, as espécies H. capsulatum e S. Schenckii são eucariotos
pertencentes ao Filo Ascomycota, o qual está contido no Reino Fungi. H. capsulatum está
classificado na Classe Eurotiomycetes, Ordem Onygenales, Família Ajellomycetaceae. O
Gênero Histoplasma,inclui a Espécie Histoplasma capsulatum, cuja forma teleomórfica
denomina-se Ajellomyces capsulatus (LACAZ et al., 2002; KASUGA et al., 2003).
A espécie H. capsulatum é dividida em três variedades: H. capsulatum var.
capsulatum, que é considerada o agente etiológico da histoplasmose clássica, de distribuição
universal. H. capsulatum var. duboisii que causa a histoplasmose africana e está restrita a uma
parte do continente africano e H. capsulatum var. farciminosum que não foi encontrada
parasitando o homem, tendo sido descrita apenas causando linfangite epizoótica em equinos
(LACAZ et al., 2002). O presente trabalho dará enfoque à variedade capsulatum, a qual será
identificada apenas como H. capsulatum, sem fazer menção à variedade deste ponto em
diante.
A espécie S. schenckii está classificada taxonomicamente como pertencente ao Reino
Fungi, Filo Ascomycota, Classe Pyrenomycetes, Ordem Ophiostomatales e a Família
Ophiostomatacea (GUARRO; GENÉ; STCHIGEL, 1999). Considerava-se que S. schenckii
era única espécie do gênero que causava doença em humanos (ARRILLAGA-MONCRIEFF
et al., 2009). Contudo, numerosos estudos moleculares com esta espécie foram efetuados, os
quais evidenciaram a existência de um complexo de espécies filogenéticas, geneticamente
diferentes. Um estudo utilizando uma sequência de análise multilocus, investigou a estrutura
populacional de S. schenckii e revelou a existência de espécies filogenéticas compondo um
complexo (MARIMON et al., 2006). Assim, novas espécies além de S. schenkii foram
estabelecidas como pertencentes ao gênero Sporothrix, sendo demoninadas como, S.
brasiliensis, S.globosa, S. mexicano, S. albicans e S. luriei (MARIMON, et al., 2007;
MARIMON, et al., 2008). Mais recentemente foram incluídas mais três outras espécies ao
complexo S. schenckii. S. stylites, S. humicola, e S. lignivora, as quais não estão associadas a
infecções e, portanto classificadas como ambientais. (DE MEYER et al., 2008). O presente
trabalho dará enfoque aos membros do complexo S. schenckii, o qual será referido apenas

17
como S. schenckii deste ponto em diante.
2.1.2 Morfologia
H. capsulatum e S. schenckii são fungos dimórfico térmicos, que em sua fase
saprofítica, na natureza ou em cultura a 25°C apresenta-se na forma filamentosa. Entretanto,
quando parasitando os tecidos de um hospedeiro e/ou em culturas mantidas a 37 °C,
apresenta-se sob a forma de levedura (GUIMARÃES; NOSANCHUK; ZANCOPÉ-
OLIVEIRA, 2006; MADRID et al. 2011a; RODRIGUES; HOOG; CAMARGO, 2012).
Na forma filamentosa H. capsulatum se caracteriza macromorfologicamente por
colônias brancas de textura algodonosa, com micélio aéreo (Figura 1A). Microscopicamente,
o microrganismo apresenta hifas hialinas, septadas e ramificadas, e dois tipos de conídios
como estruturas de propagação, os microconídios que são estruturas ovaladas, com paredes
lisas, medindo em torno de 2 a 4 µm, que constituem a forma infectante, e os macroconídios
ou conídios tuberculados que são estruturas redondas ou piriformes, que possuem parede
espessa, cobertos por projeções espiculadas e medem em torno de 8 µm a 15 µm (Figura ex
1B) (KAUFFMAN, 2006; AIDÉ, 2009; FERREIRA; BORGES, 2009). Na fase parasitaria,
sob a forma leveduriforme, é caracterizado macromorfologicamente pela presença de colônias
de aspecto úmido e cremosas (Figura 1C). Microscopicamente, o microrganismo apresenta
células ovais uninucleadas medindo cerca de 3 a 5 µm de diâmetro (Figura 1D) (FERREIRA;
BORGES, 2009).
Figura 1. Aspecto morfológico de H. capsulatum
A. Macromorfologia da fase filamentosa em ágar batata, demonstrando colônia algodonosa branca; B.
Micromorfologia da fase filamentosa, mostrando hifas septadas e macroconídios tuberculados; C.
Macromorfologia da fase leveduriforme em ágar BHI com sangue de carneiro 10%, mostrando colônias de
coloração creme com aspecto membranoso; D. Micromorfologia da fase leveduriforme, mostrando células ovais
uninucleadas e estrutura leveduriforme com brotamento.
Fonte: CEMM, 2011.

18
Por sua vez, S. schenckii na fase filamentosa exibe colônias que variam da cor branca
ao creme, de textura lisa, com aspecto membranoso, coreácea, com superfície sulcada (Figura
2A). Posteriormente, as colônias podem desenvolver a coloração castanha escura a negra
(LACAZ; PORTO; MARTINS, 1991; LOPES-BEZERRA; SCHUBACH; COSTA, 2006),
embora, algumas cepas apresentem colónias escuras desde o início do crescimento
(ALMEIDA-PAES et al., 2009). Microscopicamente, a fase filamentosa revela hifas hialinas,
septadas, ramificadas e muito delicadas, medindo de 1,5 a 2,0 μm de espessura. Os conídios,
que podem medir de 2 a 6 μm, dispõem-se em cachos terminais, assemelhando-se à flor de
margarida, na extremidade do conidióforo (Figura 2B) (MARQUES-EVANGELISTA, 2010).
Na fase leveduriforme, em cultura a 37°C as colônias de S. schenckii apresentam aspecto
cremoso, são escurecidas ou creme e com superfície irregular (Figura 2C). (TRILLES et al.,
2005; MADRID et al., 2010; BARROS; PAES; SCHUBACH, 2011). Microscopicamente, as
leveduras exibem gemulação única e variam de forma e tamanho, podendo ser ovais ou
redondas, com 2 a 6 μm de diâmetro, ou em formato de charuto, medindo 2x3 a 3x10 μm
(Figura 2D) (MARQUES-EVANGELISTA, 2010).
A melanina é o pigmento responsável pela coloração escurecida observada nas
colônias de S. schenckii e é um importante fator de patogenicidade do fungo (ALMEIDA-
PAES et al., 2009). Nas hifas, existem conídios pigmentados, com paredes celulares grossas,
que variam em número, o que determina a intensidade da cor da colônia (NOBRE et al.,
2008). Apesar das colônias na forma leveduriforme não apresentarem grande variações de cor
entre cepas, sendo sempre de coloração creme, foi demonstrada uma fina camada de melanina
nas células leveduriformes (MORRIS-JONES et al., 2003).
Figura 2. Aspecto morfológico de Sporothrix schenckii.
A. Macromorfologia na fase filamentosa em ágar batata, mostrando colônia de cor branca a creme, de textura
lisa, com aspecto coreáceo; B. Micromorfologia, mostrando hifas finas e delicadas, com conídios nas
extremidades dos conidióforos assemelhando-se a rosetas; C. Macromorfologia da fase leveduriforme em ágar
Sabouraud dextrose, mostrando colônia de cor creme e aspecto cremoso; D. Micromorfologia da fase
leveduriforme, mostrando células leveduriformes ovaladas.
Fonte: CDC, 2013; CEMM, 2013.

19
A reversão in vitro da fase filamentosa para a fase leveduriforme de H. capsulatum e
S. schenckii pode ser obtida com a utilização de meios de cultura suplementados, tal como
ágar BHI (Brain Heart Infusion) acrescido de sangue de carneiro. Outros fatores como
aeração, tensão de CO2 e pH também podem ser importantes para facilitar o processo de
transição morfológica, contudo, vale ressaltar que a temperatura é um fator determinante no
dimorfismo, sendo essencial incubação das culturas em uma faixa de temperatura de 35-37°C
(LOPES-BEZERRA; SCHUBACH; COSTA, 2006). Algumas cepas do complexo S.
schenckii, especialmente à espécie S. globosa, não crescem acima de 37 °C e, portanto
requerem temperaturas de conversão mais baixas (MARIMON et al., 2007).
2.1.3 Aspectos ecológicos
Na sua fase saprofítica, H. capsulatum é um fungo geofílico, encontrado em solos
contaminados com excrementos de aves, morcegos ou pássaros, com elevado teor de
compostos nitrogenados, com pH ácido e com alta umidade. Assim, o fungo pode ser isolado
de locais onde o solo possa estar enriquecido com excretas desses animais como: cavernas,
minas, parques públicos, ocos de árvores, galinheiros, forros de casas e edificações antigas
(AIDÉ, 2009; FERREIRA; BORGES, 2009; NORMAN et al., 2009). Atividades como
limpeza de sótãos ou celeiros, demolição de prédios antigos e revolvimento de solos podem
estar relacionadas a casos de histoplasmose (KAUFFMAN, 2009). Correntes de ar carreiam
os conídios por quilômetros de distância, expondo indivíduos que não estavam próximos de
áreas contaminadas (DEUS FILHO et al., 2009). Vários animais são suscetíveis à
histoplasmose, desta forma, H. capsulatum já foi isolado de vísceras de quirópteros, cães,
gatos, ratos e marsupiais de vida silvestre (SILVA-VERGARA et al., 2001). Os morcegos
são suscetíveis à infecção e participam ativamente da epidemiologia da histoplasmose.
Devido à sua capacidade migratória, esses animais podem disseminar o fungo de um local a
outro, liberando em suas excretas formas viáveis do fungo, ajudando na propagação da doença
(WANKE; LAZÉRA, 2004; KAUFFMAN, 2009; ALLTON et al., 2010). Acreditava-se as
aves não albergavam o microrganismo devido a sua elevada temperatura corporal (39-40°C),
entretanto, em 2011, um caso de co-infecção com H. capsulatum e Candida albicans em um
papagaio (Eclectus roratus) foi relatado, mostrando que as aves, embora raramente, também
podem ser acometidas (QUIST et al., 2011).
A fase saprofítica de S. schenkii também é geofílica, sendo a presença de matéria
orgânica no solo fundamental para o desenvolvimento do micélio. O fungo pode ser

20
encontrado em solos abundantes em celulose, pH 3,5 a 9,4 e em ambientes de clima tropicais
e subtropicais, com umidade relativa não inferior a 92% e temperaturas em torno de 30
°C(BARROS; PAES; SCHUBACH, 2011). Normalmente, a infecção com S. schenckii ocorre
devido à inoculação acidental ou traumática do agente na pele, por espinhos ou material
vegetal o que, provavelmente, justifica a maior frequência da enfermidade em jardineiros,
agricultores, floristas, manipuladores de sementes, mineiros, trabalhadores de alvenaria,
laboratoristas em microbiologia ou ainda, tosadores e tratadores de animais, bem como, em
médicos veterinários que são vítimas ocasionais de arranhaduras e mordeduras, sendo desta
forma, considerada uma micose ocupacional (MORRIS-JONES, 2002; LOPES-BEZERRA;
SCHUBACH; COSTA, 2006; LARSSON, 2011; CARRADA, 2012). Pode acometer o
homem e uma grande variedade de animais, sendo os felinos os mais frequentemente
infectados pelo agente. Há, relatos de infecções em mamíferos, como primatas, bovinos,
camelídeos, murinos, caprinos, suínos, golfinhos, em aves e em artrópodes que carreiam o
microrganismo (abelhas, pulgas, formigas) que, eventualmente, inclusive, podem servir de
fonte de infecção ao homem e outros animais (LARSSON, 2011). Atualmente a caça de tatus
também tem sido relacionada a casos de esporotricose (ALVES et al., 2010). Embora, S.
schenckii não tenha sido achado na epiderme dos tatus, o fungo foi encontrado sobre a grama
seca utilizada nos ninhos desses animais (BARROS; PAES; SCHUBACH, 2011).
2.2 Aspectos Gerais da Histoplasmose e da Esporotricose
2.2.1 Epidemiologia
Atualmente, a histoplasmose está distribuída em todo o mundo. É uma micose
sistêmica prevalente nas Américas, com predominância de casos em algumas áreas dos
Estados Unidos como as regiões centrais e sul do país, ao longo dos vales dos rios Mississipi
e Ohio. Países da América Central (México, Honduras, Guatemala, Nicarágua, Panamá) e
América do Sul (Venezuela, Colômbia, Peru, Brasil, Argentina e Uruguai) também
apresentam alta prevalência dessa micose. A doença também tem sido relatada na África, Ásia
e na Europa, onde foram diagnosticados casos esporádicos e autóctones (ROSSINI,
GOULART, 2006; FERREIRA; BORGES, 2009; BRILHANTE et al., 2012 ).
No Brasil, a ocorrência da histoplasmose se dá através da observação de casos clínicos
autóctones, seja em casos isolados ou sob a forma de microepidemias (CHANG et al., 2007).
Já foram registrados microepidemias em vários estados brasileiros: Rio Grande do Sul, Rio de
Janeiro, São Paulo, Distrito Federal, Minas Gerais, Paraíba, Amazonas, Bahia, Maranhão e

21
Piauí (AIDÉ, 2009; DEUS FILHO et al., 2009). No Ceará, estudos realizados com 378
pacientes HIV positivos, no período compreendido entre 1995 a 2004, a histoplasmose
disseminada foi detectada em quase 44% dos pacientes e apresentou alta taxa de mortalidade
(DAHER et al., 2007). Ainda no Estado do Ceará, um estudo realizado por Brilhante et al.,
2012, no período compreendido entre 2006 a 2010, encontraram 208 casos de histoplasmose
associada à pacientes HIV positivos, atendidos em um hospital de referência de Fortaleza.
A esporotricose por sua vez, é uma micose subaguda ou crônica, com distribuição
ubiquitária, cosmopolita, hoje evidenciada, principalmente, no meio urbano superando a
ocorrência rural. Também é caracterizada como uma micose universal, relatada nos cinco
continentes, onde tem maior prevalência em zonas com condições climáticas tropicais e
subtropicais. Embora seja prevalente nos EUA, tem maior importância epidemiológica nas
Américas Central e do Sul, especialmente no México (região central) e no Brasil.
(LARSSON, 2011; BARROS; PAES; SCHUBACH, 2011). Centenas de casos foram
diagnosticados nos últimos anos na Austrália, Brasil, Uruguai, China, Colômbia, Estados
Unidos, Índia, Espanha, Itália, Marrocos, Japão, México, Peru, Turquia e Venezuela
(MADRID et al., 2011 B). Mas as principais áreas de endemicidade estão localizadas no
Japão, Índia, México, Brasil, Uruguai e Peru (BARROS; PAES; SCHUBACH, 2011).
A esporotricose, atualmente é considerada a micose subcutânea mais frequente no
Brasil, devido ao aumento progressivo da transmissão zoonótica da doença na região
metropolitana do Rio de Janeiro (SILVA et al., 2012). Desde 1998, o Instituto de Pesquisa
Clínica Evandro Chagas – Fiocruz vem constatando um crescimento exponencial de casos de
esporotricose em humanos, felinos e caninos, onde o Rio de Janeiro é o estado brasileiro com
o maior número de registros, atingindo proporções epidêmicas (BARROS et al., 2010;
NUNES et al., 2011). No Rio Grande do Sul, muitos casos humanos estão conectados com as
atividades ocupacionais (GOMES et al., 2012). Em outros estados, como Espírito Santo, Mato
Grosso, Minas Gerais, São Paulo, Paraná e Santa Catarina, a esporotricose tem sido
ocasionalmente descrita em cães e gatos (MADRID et al., 2011b). No entanto, existem poucas
referências na literatura sobre a esporotricose em outras áreas geográficas do país. No
Nordeste brasileiro apenas dois relatos de caso foram relatados, incluindo o acometimento de
um cão na cidade de Mossoró, RN e um gato no município de Itaporanga, estado da Paraíba.
(FILGUEIRA, 2009; NUNES et al., 2011).

22
2.2.2 Patogênese e Imunidade
A histoplasmose é adquirida através da inalação de partículas infectantes, formadas
por fragmentos de hifas e microconídios, presentes em solo contaminado com H. capsulatum.
Ao alcançar os alvéolos pulmonares o fungo estimula uma resposta inflamatória no
hospedeiro, sendo fagocitado por macrófagos alveolares (NOSANCHUK; GRACSER, 2008).
A conversão do microrganismo para a fase leveduriforme se dá no interior dos macrófagos, no
parênquima pulmonar, sendo transportado por via linfática até os linfonodos hilo-
mediastinais, onde forma um complexo pulmonar ganglionar primário e ganham acesso à
circulação sanguínea para disseminação a vários órgãos, tais como fígado, baço e medula
óssea. A partir daí, é a resposta imunológica do hospedeiro contra a infecção que vai
determinar a extensão da doença (ROSSINI; GOULART, 2006; FERREIRA; BORGES,
2009). Após a segunda ou terceira semana do início da infecção primária, a imunidade celular,
do tipo T-helper 1 (Th1), é ativada no organismo do hospedeiro, que reconhecem os antígenos
da parede celular fúngica e as proteínas do choque térmico, produzindo INF-γ e outras
citocinas, como IL-2 e IL-12, que ativam os macrófagos, os quais adquirem a capacidade de
lisar leveduras intracelulares do H. capsulatum. Desta forma, em hospedeiros
imunocompetentes, os macrófagos ativados exibem atividade microbicida, ocorrendo uma
intensa reação granulomatosa, seguida de cicatrização, fibrose e calcificação. Esse tipo de
reação imune leva a cura da infecção primária tornando o hospedeiro resistente às novas
infecções, que eventualmente aconteçam. Se a infecção não for controlada pode se disseminar
para outros órgãos, como fígado e baço, sendo capaz de sobreviver durante muitos anos e
reativar-se durante um quadro de imunodepressão (ROSSINI; GOULART, 2006;
FERREIRA; BORGES, 2009; KAUFFMAN, 2009).
Vários autores têm demonstrado que H. capsulatum possui diversos genes que
desempenham um papel importante na virulência do patógeno, dentre os quais têm destaque o
gene Ryp1 fundamental no processo de transição dimórfica; o gene CBP1, expresso apenas na
fase leveduriforme, codificante da proteína de ligação ao cálcio CBP (calcium-binding
protein) essencial na sobrevivência e patogenicidade, o gene YPS3 que participa na
disseminação do H. capsulatum para sítios extrapulmonares e o gene α-(1,3)-glucana sintetase
codificante do polímero da parede celular presente na fase leveduriforme (KLEIN,
TEBBETS, 2007; HOLBROOK, ROPPLEYE, 2008; WEBSTER, SIL, 2008). Ademais,
estudos realizados por Nosanchuk et al. (2002) indicam que os conídios e leveduras de
Histoplasma capsulatum podem produzir melanina ou compostos semelhantes à melanina in

23
vitro e in vivo (Nosanchuk et al.,2002). A melanina é um importante fator de virulência em
fungos patogênicos, como exemplos o Cryptococcus neoformans, Sporothrix schenckii, dentre
outros (ROMERO-MARTINEZ et al, 2000; DUIN; CASADEVALL; NOSANCHUK, 2002),
que ajuda na proteção do microrganismo contra agressões ambientais e terapias
antimicrobianas. Assim, acredita-se que este pigmento pode ter um papel semelhante na
patogênese da histoplasmose (NOSANCHUK et al.,2002).
Na esporotricose, a infecção geralmente se inicia com uma lesão traumática por
vegetais ou decorrentes de ferimentos obtidos durante brigas entre animais. Após penetrar
através da pele e converter para forma de levedura, o fungo pode ficar localizado no tecido
subcutâneo ou prolongar ao longo dos vasos linfáticos adjacentes. Mais raramente, pode se
disseminar por via hematogênica (STALKUP, BELL, ROSEN, 2002). Além de lesões
cutâneas, o fungo pode ser ocasionalmente, inalado, infectar os pulmões e depois disseminar-
se para outros órgãos (REIS et al., 2009). O tempo de transição micélio-levedura dura em
torno de 13 dias, sendo que 24 - 48hs após a inoculação da fase filamentosa do fungo há o
desaparecimento das hifas e inicia-se a conversão dos conídios para leveduras (CORREA et
al., 1991). Desta forma, o período de incubação para o desenvolvimento da infecção por
esporotricose é maior quando um ferimento é produzido por uma planta contaminada por S.
schenckii, pois o inóculo fúngico na sua forma filamentosa precisará converter para forma de
levedura para que então ocorra a sua multiplicação. Já na contaminação por contato com um
animal portador de lesões ulceradas, um grande número de células leveduriformes é
transferido diretamente para o tecido lesionado. (CRUZ, 2010; CRUZ, 2013).
S. schenckii possui diversos fatores de virulência que aumentam suas chances de
sobrevivência no hospedeiro, levando assim ao desenvolvimento da doença. Dentre os
principais, se destacam a presença de enzimas extracelulares, termotolerância, moléculas
constituites da parede celular e a presença de grânulos de melanina (LOPES-BEZERRA;
SCHUBACH; COSTA, 2006; Madrid et al., 2010). Esses fatores auxiliam na invasão tecidual
e interferem na eficácia da resposta imune, propiciando a instalação do microorganismo no
tecido e permitindo sua transição para a forma leveduriforme (REIS et al., 2009). Estudos in
vitro demonstraram que fagócitos ingerem e fagocitam até duas vezes mais conídios hialinos
do que pigmentados, indicando que a melanina contribui para a sobrevivência do fungo no
hospedeiro (ROMERO-MARTINEZ et al., 2000). Além disso, as melaninas suprimem a
produção de citocinas pró-inflamatórias, interferem na ativação do complemento e na ação de
drogas antifúngicas. No ambiente, protege os conídios contra a radiação ultravioleta,

24
temperaturas extremas e dissecação (MADRID, 2007). Enzimas extracelulares como as
fosfatases ácidas, produzidas pelo fungo, também parecem desempenhar papel importante na
interação parasito-hospedeiro interferindo na dinâmica entre células leveduriformes de S.
schenckii com macrófagos (NASCIMENTO; ALMEIDA, 2005).
2.2.3 Quadros Clínicos
A gravidade e evolução das manifestações clínicas tanto na histoplasmose quanto na
esporotricose estão na dependência de fatores como estado imunológico do hospedeiro, a
carga do inóculo inicial, virulência da cepa, da profundidade do trauma no caso da
esporotricose, dentre outros (AIDÉ, 2009; ARRILLAGA-MONCRIEFF et al., 2009;
FERNANDES, MATHEWS, BEZERRA, 1999).
Na histoplasmose as apresentações clínicas, podem variar desde uma infecção
assintomática até uma doença disseminada. Cerca de 90 a 95% dos casos de histoplasmose
são assintomáticos, não sendo observada nenhuma sintomatologia clínica. Em paciente
imunocompetentes, a doença regride de modo espontâneo, ocasionando apenas quadro
semelhante a uma infecção viral respiratória (ROSSINI; GOULART, 2006; KAUFFMAN,
2009; AIDÉ, 2009). Quando se desenvolve, a histoplasmose pode ser clinicamente
classificada em pulmonar aguda, pulmonar crônica ou disseminada (SEGURO; SILVEIRA,
2008). A doença pulmonar aguda é geralmente regressiva, contudo, quando uma grande
quantidade de conídios é inalada pode provocar um sério quadro pulmonar agudo, cujos
sintomas chegam a desaparecer após duas a quatro semanas, sem a necessidade de tratamento.
A sintomatologia mais comum consiste em febre, calafrios, cefaléia, mialgias, tosse, dispnéia,
dor torácica, dor retro-esternal e prostração intensa (ROSSINI; GOULART, 2006;
FERREIRA; BORGES, 2009; AIDÉ, 2009).
Já a histoplasmose pulmonar crônica acomete, principalmente, indivíduos tabagistas,
com mais de 50 anos e portadores de enfisema pulmonar, favorecendo a colonização do H.
capsulatum nessas lesões, podendo apresentar os seguintes sintomas: febre baixa vespertina,
perda de peso, sudorese noturna, dor torácica e tosse com expectoração hemoptoica
(COUPPIÉ et al., 2006; ROSSINI; GOULART, 2006; FERREIRA; BORGES, 2009; AIDÉ,
2009). Por fim, a histoplasmose disseminada é a forma clínica mais grave, ocorre
principalmente em pessoas com imunidade celular baixa, pacientes com neoplasias
hematológicas, transplantados ou usuários de drogas imunossupressoras (MUKHERJEE et
al., 2010; SEGURO; SILVEIRA, 2008). Os pacientes HIV positivos são os mais afetados,

25
muito embora um pequeno número de pessoas sadias ou que apresentam alguma forma de
imunodepressão chegam a desenvolver a histoplasmose disseminada. Os sintomas mais
observados são o aumento do fígado, baço e linfonodos, e mais raramente a produção de
úlceras na boca e no intestino. Este quadro clínico pode ser fatal em 90% dos casos, sem
tratamento adequado (FERREIRA; BORGES, 2009).
Quanto aos quadros clínicos de esporotricose, atualmente a classificação mais adotada
é baseada na revisão publicada por Lopes-Bezerra et al. (2006) a qual será adotada a seguir. A
forma cutânea fixa apresenta-se como uma lesão única ou poucas lesões no local da
inoculação, frequentemente ulcerativa, com bordas eritematosas. A morfologia pode ser
vegetativa verrucosa, com placa infiltrada, sem envolvimento linfático (BARROS; PAES;
SCHUBACH, 2011). Alguns autores relatam a forma cutânea fixa como a principal clínica de
apresentação em crianças (TLOUGAN et al., 2009). A forma cutânea-linfocutânea é a forma
mais frequente da doença, estando presente em geral, em mais de 75% dos casos (RAMOS-E-
SILVA, et al., 2007). A lesão inicial começa com uma lesão nodular ou ulcerada no local da
inoculação do fungo, normalmente localizada nas extremidades, especialmente nas mãos e
antebraços e segue uma trajetória linfática caracterizada por lesões nodulares com aspecto de
uma ulcera ou de um pequeno nódulo subcutâneo. Esta lesão, exercendo pressão sob a pele,
provoca isquemia sob a epiderme, evolui para gomas, ulcera, e libera uma secreção purulenta.
Esta descrição clínica levou a nomear a doença como linfangite ascendente nodular (LOPES-
BEZERRA; SCHUBACH; COSTA, 2006; BARROS; PAES; SCHUBACH, 2011).
A forma cutânea disseminada é bem menos frequente que as anteriores e a forma
clínica mais grave, a qual ocorre como resultado de uma disseminação hematogênica do sítio
de inoculação primário ou do linfonodo. É caracterizada pela extensão das lesões a grandes
áreas da pele, atingindo principalmente hospedeiros imunocomprometidos e pode se
apresentar de diversas formas, dentre elas sinusite e meningite (DE ARAUJO, MARQUES,
KERDEL, 2001; BARROS; PAES; SCHUBACH, 2011). A forma mucosa é incluída por
alguns autores como uma variante da forma cutânea. Na mucosa nasal, as lesões,
frequentemente, envolvem o septo com drenagem de sangue e desprendimento de crostas. Na
conjuntiva, as lesões granulomatosas são acompanhadas de material seropurulento e
vermelhidão (SCHUBACH, et al., 2005). Eritema nodoso e eritema multiforme foram
relatados em casos de esporotricose por transmissão zoonótica. Estas condições parecem estar
associadas com reação de hipersensibilidade, o que resulta de uma exposição contínua a
grandes quantidades de fungos e reinfecções subclínicas (GUTIERREZ-GALHARDO, et al.,

26
2002; GUTIERREZ-GALHARDO et al., 2005). Em gatos com esporotricose, o nariz é a
região mais afetada e sinais respiratórios são comuns. Como os proprietários brincam com
seus animais em estreito contato, a transmissão é facilitada (GREMIÃO et al., 2009;
BARROS; PAES; SCHUBACH, 2011).
As formas extracutâneas são raras, embora tenha se tornado mais freqüentes após o
aparecimento da AIDS. Entre as formas extracutâneas, a osteoarticular e o comprometimento
pulmonar são as manifestações mais comuns, mas também há relatos de casos de
disseminação hematogênica grave com envolvimento de múltiplos órgãos (LOPES-
BEZERRA; SCHUBACH; COSTA, 2006; BARROS; PAES; SCHUBACH, 2011). Após a
pele, o tecido ósseo é o mais afetado. A forma osteoarticular pode ocorrer por contiguidade ou
disseminação hematogênica. As lesões podem variar de grandes a pequenos granulomas com
lesões líticas idênticas a osteomielite (MORRIS-JONES, 2002; KOHLER; HAMDAN;
FERRARI, 2007). Por fim, a esporotricose pulmonar primária, resultante da inalação do
fungo, é geralmente associada à doença pulmonar obstrutiva crônica, alcoolismo, uso crônico
de corticóides e doenças imunossupressoras. A apresentação clínica é semelhante ao da
tuberculose, e o diagnóstico muitas vezes é tardio, devido à raridade de envolvimento
pulmonar. Esporotricose sistêmica é extremamente rara e sempre associada com deficiência
do sistema imunitário (BARROS; PAES; SCHUBACH, 2011). Relatos sobre meningite
associadas a infecção por Sporothrix não são frequente, e estes casos são frequentemente
associados com deficiência imunológica. (BARROS; PAES; SCHUBACH, 2011).
2.2.4 Diagnóstico
O diagnóstico da histoplasmose e da esporotricose pode ser obtido através da
correlação de dados clínicos, epidemiológicos e laboratoriais. A análise laboratorial inclui:
testes micológicos, histopatológicos, imunológicos e moleculares (AIDÉ, 2009; SILVIA et
al., 2012). Vale ressaltar que o diagnóstico laboratorial é indispensável para a confirmação
dos casos de suspeitos. Assim, o diagnóstico clínico baseado apenas na patogenia e nas
alterações patológicas deve ser confirmado pela observação microscópica de estruturas
leveduriformes no material coletado, bem como, no isolamento e identificação do agente
etiológico envolvido no processo (AIDÉ, 2009; CRUZ, 2013).
Os testes micológicos incluem simultaneamente a realização de exame direto e cultura.
Na histoplamose as amostras clínicas, mais utilizadas incluem medula óssea, sangue
periférico, escarro, líquor e biopsias de tecidos (ROSSINI; GOULART, 2006). Dentre as

27
técnicas utilizadas para o diagnóstico da histoplasmose a cultura fúngica continua sendo o
“padrão ouro” e o exame direto, apesar de rápido e de baixo custo, é uma técnica de baixa
sensibilidade para detecção de H. capsulatum, devido às pequenas dimensões do patógeno
(COUPPIÉ et al., 2006; ROSSINI; GOULART, 2006). Os meios de cultura Ágar Sabouraud
dextrose ou ágar BHI, suplementados ou não com cloranfenicol e cicloheximida podem ser
utilizados. O fungo cresce, em cerca de 1 a 3 semanas, apresentando colônias brancas,
algodonosas, com micélio aéreo que tende a escurecer com o passar do tempo (ROSSINI;
GOULART, 2006).
Já nos casos de suspeita de esporotricose, as amostras clínicas comumente utilizadas
incluem secreção, crostas e biopsias das lesões (NOBRE et al., 2001). Embora sempre seja
recomendada a obtenção de amostras para o cultivo micológico, a ausência de isolamento não
exclui a presença do S. schenckii, e assim a histopatologia (associada a técnicas de coloração
especial) pode torna-se uma opção extremamente viável para a confirmação da enfermidade
(NUNES et al., 2011). O isolamento geralmente é realizado em ágar Sabouraud dextrose,
acrescido de cicloeximida e para caracterizar os aspectos macroscópicos e microscópicos das
colônias utiliza-se agar BHI ou agar batata. O cultivo demanda entre dez e 14 dias para
propiciar ou afastar o diagnóstico etiológico (LARSSON, 2011). A possibilidade de se
estabelecer o diagnóstico, através de isolamento do agente, oscila entre 34% e 94% dos casos,
na dependência da origem do material colhido (decalque de garras – 39,5%; cavidade oral –
42%; coágulo sanguíneo, de animais em fungemia - 34% e vias aéreas anteriores – 94%)
(SCHUBACH et al., 2003a).
O exame histopatológico dos casos de histoplamose se dá através da visualização de
macrófagos parasitados com células leveduriformes em cortes histológicos corados por
hematoxilina-eosina (HE), Giemsa ou coloração da prata-metenamina de Grocott-Gomori. As
formas leveduriformes aparecem como corpúsculos esféricos ou ovalados, rodeados por um
halo claro delimitado por uma parede celular muito fina e hialina (ZANCOPÉ-OLIVEIRA;
MUNIZ; WANKE, 2005). O exame pode ser realizado a partir de várias amostras de tecidos
como pulmão, medula óssea, fígado e linfonodos revelando a presença de granulomas
epitelóides, onde o microrganismo, assim como no exame direto, pode ser vistos no interior
de macrófagos (AIDÉ, 2009; FERREIRA; BORGES, 2009).
Na esporotricose, o exame histopatológico permite o estabelecimento do diagnóstico
etiológico em 95% a 100% dos casos (LARSON, 2002; SCHUBACH et al., 2003a). O

28
citodiagnóstico, a partir de material (exsudado, aspirado, decalcado, biopsiado) submetido às
colorações de Gram, Wright, Giemsa, Rosenfeld, permite que se evidencie, principalmente
em gatos, um excesso de formas leveduriformes, arredondadas, ovaloides, em forma de
“charuto”, com 3-5 µm de diâmetro e 5-9 µm de comprimento (LARSSON, 2011). A
realização do diagnóstico diferencial é indispensável, pois as estruturas leveduriformes de S.
schenckii podem ser confundidas com o patógeno causador da Leishmaniose tegumentar
americana (SCHUBACH et al., 2003b; MELLO; SCHUBACH; MADEIRA, 2011). Embora a
leishmaniose seja muito mais prevalente em cães, o diagnóstico diferencial em felinos
também deve ser levado em consideração (SCHUBACH et al., 2003b; SOUZA et al., 2005;
SOUZA et al., 2009).
Os testes sorológicos são auxiliares ao diagnóstico definitivo e na histoplasmose
apresentam sensibilidade superior a 90%, quando corretamente executados (AIDÉ, 2009).
Para tanto, várias técnicas têm sido utilizadas, tais como Reação de Fixação do Complemento,
Imunodifusão Dupla e Radioimunoensai. Por mostrar-se mais sensível, no teste de fixação do
complemento, são utilizados antígenos extraídos da fase leveduriforme de H. capsulatum. O
teste de Imunodifusão dupla é muito empregado no diagnóstico de micoses sistêmicas e
conhecido mundialmente por ser uma prova de rápida e fácil execução, mostrando-se mais
específica do que a Reação de Fixação do Complemento (ROSSINI; GOULART, 2006;
KAUFFMAN, 2006). Outra técnica utilizada é o Radioimunoensaio, o qual apresenta bastante
sensibilidade para detecção de antígenos circulante de H. capsulatum. No entanto, não é
muito adotado nos laboratórios de rotina, devido a sua complexidade técnica (ROSSINI;
GOULART, 2006; COUPPIÉ et al., 2006). Na esporotricose as técnicas sorológicas mais
empregadas, incluem as reações de fixação do complemento, imunodifusão,
imunofluorescência indireta e principalmente a soro aglutinação, a qual é a técnica mais
sensível, específica e de fácil execução (LARSSON, 2011). Os exames sorológicos podem ser
úteis auxiliando o diagnóstico da esporotricose, especialmente, para as formas atípicas e
extracutâneas (LOPES-BEZERRA; SCHUBACH; COSTA, 2006).
Os métodos moleculares são considerados excelentes ferramentas para o diagnóstico
de histoplasmose, capazes de confirmar os dados dos testes sorológicos de detecção de
antígenos (GUIMARÃES; NOSANCHUK; ZANCOPÉ-OLIVEIRA, 2006). Atualmente, as
técnicas disponíveis e mais utilizadas são a hibridização de nucleotídeos e PCR, que trazem
como vantagem a diminuição do tempo de espera dos resultados, apresentando maiores
índices de sensibilidade e especificidade (ZANCOPÉ-OLIVEIRA; MUNIZ; WANKE, 2005).

29
2.2.5 Tratamento, Prevenção e Controle
O tratamento da histoplasmose depende da severidade dos sintomas clínicos e das
condições gerais do indivíduo, incluindo seu estado imunológico (WHEAT et al., 2007).
Dentre as opções terapêuticas disponíveis, a anfotericina B continua sendo a droga de escolha
para a terapêutica da histoplasmose principalmente nos quadros clínicos mais graves, apesar
do seu efeito nefrotóxico (KAUFFMAN, 2009). Nos últimos anos, com a disponibilidade dos
derivados azólicos a partir da década de 1980, vários estudos têm demonstrado a eficácia
desses medicamentos no tratamento dessa infecção fúngica, inclusive na forma disseminada,
mostrando-se como uma importante alternativa para o esquema terapêutico da histoplasmose
(KAUFFMAN, 2007; FERREIRA; BORGES, 2009).
O cetoconazol foi o primeiro derivado azólico utilizado como alternativa a
anfotericina B para o tratamento da histoplasmose em pacientes imunocompetentes (WHEAT;
KAUFFMAN, 2003). No entanto, hoje a droga raramente é empregada para a terapia da
doença, especialmente em pacientes com AIDS, devido a sua fraca absorção diante da
redução do ácido gástrico, provocando baixa resposta ao tratamento (KAUFFMAN, 2007;
KAUFFMAN, 2009). Dentre os demais derivados azólicos, o itraconazol é o mais efetivo,
sendo considerada a droga de primeira escolha no tratamento da histoplasmose pulmonar
aguda ou pulmonar cavitária crônica (WHEAT et al., 2007, KAUFFMAN, 2009). Ademais,
muitos autores têm sugerido o itraconazol como fármaco de escolha para o tratamento de
histoplasmose disseminada em pacientes com AIDS (KAUFFMAN, 2007; FERREIRA;
BORGES, 2009).
Alguns pacientes podem apresentar intolerância ao itraconazol, necessitando do uso de
terapias alternativas, como o voriconazol e posacanazol. O fluconazol também tem sido
utilizado com sucesso contra a histoplasmose, mas apresenta menor eficácia quando
comparado ao itraconazol e a anfotericina B, além disso, existem relatos de resistência
resultando em falhas terapêuticas (WHEAT et al., 2007). Até o momento não há trabalhos na
literatura que relatem sobre o tratamento da histoplasmose com equinocandinas em seres
humanos. Alguns autores chegaram a demonstrar sua ineficácia quando comparada com a
anfotericina B, no tratamento da doença em modelo murino, não sendo então recomendado
seu uso na terapia da histoplasmose (WHEAT et al., 2007; KAUFFMAN, 2009).
Na esporotricose, por sua vez a escolha terapêutica baseia-se na condição clínica do
indivíduo, a extensão das lesões cutâneas, a avaliação das interações medicamentosas, eventos

30
adversos e envolvimento sistêmico (LOPES-BEZERRA; SCHUBACH; COSTA, 2006,
CORDEIRO, et al., 2011). O tratamento da esporotricose pode ser complicado, especialmente
pela longa duração, desistência por parte dos pacientes e diagnósticos tardios (GOMES et al.,
2012). A solução saturada de iodeto de potássio é, tradicionalmente, usada no tratamento da
esporotricose desde o início do século XX, com taxas de sucesso terapêutico, variando de 80 a
100%. A escolha do iodeto de potássio se baseou nas séries de casos relatados na literatura, na
experiência do serviço e no baixo custo financeiro (CORDEIRO et al., 2011; BARROS;
PAES; SCHUBACH, 2011). Ainda hoje, o iodeto de potássio é a terapêutica de primeira
escolha para o tratamento das esporotricoses cutâneo-linfática, cutânea localizada e em muitos
pacientes da forma cutânea disseminada. No entanto, na forma extracutânea e em muitos
casos da forma cutânea disseminada, este agente não apresenta o mesmo padrão de eficácia
(SIDRIM; ROCHA, 2004).
O azólico itraconazol emergiu como uma alternativa terapêutica eficaz, com reduzidos
efeitos colaterais em relação aos fármacos tradicionalmente utilizados na terapia da
esporotricose, sendo atualmente o fármaco de primeira escolha para o tratamento das formas
cutâneas e linfocutâneas dessa micose. É uma das drogas utilizadas em pacientes
imunocomprometidos, que apresentam formas clínicas mais extensas e com acometimento
sistêmico (LOPES-BEZERRA; SCHUBACH; COSTA, 2006, KAUFFMAN, 2007, SILVA et
al., 2012). A administração do fármaco deve ser mantida por no mínimo um mês após a cura
clínica (SILVA et al., 2012). Relatórios sobre o sucesso do tratamento com fluconazol podem
ser encontrados na literatura, mas esta não é a droga de primeira escolha para o tratamento da
doença (LOPES-BEZERRA et al., 2006). O fluconazol é menos efetivo que o itraconazol,
sendo utilizado em pacientes que não toleram ou apresentam interações medicamentosas com
o itraconazol (KAUFFMAN et al., 2007).
A anfotericina B é indicado para o tratamento de formas clínicas moderadas a graves
em indivíduos imunodeprimidos e aqueles que não respondem aos fármacos descritos
anteriormente (LOPES-BEZERRA; SCHUBACH; COSTA, 2006). Mesmo apresentando
eficaz atividade sobre S. schenckii, os efeitos adversos, como a hipocalemia, hipertermia,
anorexia, náuseas, vômitos, cefaléia, mialgia e flebite, limitam o seu uso (BUSTAMANTE;
CAMPOS, 2004). A terbinafina não é formalmente indicado para o tratamento da
esporotricose, mas relatos demonstraram sucesso terapêutico (LOPES-BEZERRA;
SCHUBACH; COSTA , 2006). A terbinafina é um antifúngico do grupo das alilaminas, que
além de apresentar efeitos tóxicos reduzidos, demonstrou ser bem efetivo e tolerado pelos

31
pacientes quando comparado ao itraconazol, tornando-se, uma opção para o tratamento da
esporotricose cutânea (MEINERZ et al., 2007, FRANCESCONI et al., 2011 ).
Neste contexto, as medidas de prevenção da histoplasmose têm como finalidade evitar
a exposição a áreas potencialmente contaminadas com o fungo, tais como, locais com fezes de
morcegos como grutas, furnas, porões ou com fezes de aves como galinheiros e ainda locais
de construção ou escavação em que haja revolvimento de terra. É fundamental o uso de
máscaras apropriadas para os indivíduos com risco de exposição a locais suspeitos ou
sabidamente contaminados. Até o momento, não há vacina disponível contra H. capsulatum.
(KAUFFMAN, 2009). Quanto a esporotricose, as medidas de prevenção devem ser adotadas
de maneira a diminuir os riscos de transmissão e evitar a contaminação do meio ambiente
(SILVA et al., 2012). A esporotricose é uma doença com risco de transmissão para o médico
veterinário e seus auxiliares, por contato com exsudato, secreções, excreções, arranhaduras e
mordeduras de gatos infectados durante atendimento clínico (SILVA et al., 2012). A
cremação e o programa de controle da população de animais que vivem soltos nas ruas como
os cães, gatos e pombos deveriam ser obrigatórios nas secretarias de Saúde dos Estados e
Municípios. Os animais que morrem em ambiente já colonizado por S. schenkii também
contribuem para o aumento da população fúngica presente no solo, as plantas passarão a ter
uma carga maior de células fúngicas em suas superfícies aumentando assim a probabilidade
de transmissão da esporotricose (CRUZ, 2013).
2.3 Drogas Antifúngicas Clássicas e Testes de Sensibilidade in vitro
Diferentes tipos de infecções fúngicas, tornaram-se, nos últimos anos, um importante
problema de saúde publica, devido ao aumento de sua incidência, principalmente, em
pacientes imunodeprimidos como pacientes portadores do vírus HIV, com doenças
hematológicas, transplantados, dentre outros (YAN et al., 2011). Atualmente a terapia atual
disponível para o tratamento das micoses profundas é restrita a poucas classes de drogas
(GONZÁLEZ, 2009). Dentre essas drogas, a anfotericina B, os derivados azólicos e as
equinocandinas têm sido os principais fármacos utilizados na terapia clínica para o tratamento
das infecções fúngicas profundas e oportunistas, os quais se diferenciam quanto aos seus
mecanismos de ação (FIANCHI et al., 2007; PETRIKKOS; SKIADA, 2007).
A anfotericina B é um derivado poliênico produzido pelo actinomiceto Streptomyces
nodosus. É o agente antifúngico mais utilizado no tratamento das micoses sistêmicas, sendo
considerado o medicamento antimicótico “padrão ouro”, apesar de sua elevada toxicidade

32
(FILIPPIN; SOUZA, 2006). Pode ter ação fungicida ou fungistática dependendo da
sensibilidade do fungo e da concentração alcançada no sítio da infecção (CATALÁN;
MONTEJO, 2006). Possui afinidade pelo ergosterol da membrana celular fúngica,
componente ao qual vai fixar-se, levando a alterações na estrutura da membrana,
provavelmente pela formação de poros que levam a sua despolarização e um aumento da
permeabilidade, resultando no extravasamento de nucleotídeos, proteínas e de eletrólitos do
meio intracelular para o meio extracelular, ocasionando a morte do microrganismo. Os danos
que a droga causa na membrana dependem de uma série de fatores, tais como a fase de
crescimento da célula, a dose e a via de administração da droga (CATALÁN; MONTEJO,
2006). Devido à semelhança entre as estruturas moleculares do ergosterol das células fúngicas
e do colesterol presente nas células dos mamíferos, a anfotericina B têm dificuldades em
diferenciá-los, podendo ligar-se também ao colesterol, o que pode explicar o efeito tóxico do
fármaco (HUANG et al., 2002).
Essa droga possui um amplo espectro de ação antifúngica, sendo ativo contra H.
capsulatum, Sporothrix schenckii, Coccidioides spp., Paracoccidioides brasiliensis,
Aspergillus spp., Blastomyces dermatitidis, espécies de Candida, Cryptococcus neoformans, ,
Mucor spp. e Rhizopus spp. Algumas espécies de Candida, como C. lusitaniae, C.
guilliermondii, C. tropicalis podem apresentar resistência pré-clínica e/ou uma elevada
concentração inibitória mínima (CIM) para esse fármaco (WANKE; LAZÉRA, 2004;
CATALÁN; MONTEJO, 2006). Essa resistência se deve a uma diminuição na síntese de
ergosterol, sendo substituído por outros esteróis que fazem parte da composição da membrana
fúngica e possuem baixa afinidade pela AMB (THOMPSON; CADENA; PATTERSON,
2009).
Os efeitos adversos observados com o uso da anfotericina B podem ocorrer de forma
imediata durante a infusão, com manifestação de febre, calafrios, taquicardia, hipertensão
arterial, náuseas, vômitos e taquipneia, assim como tardios, como a nefrotoxicidade e a
anemia, necessitando da correção de doses e dos intervalos da administração e mesmo da
suspensão da terapia (MARTINEZ, 2006). Diferentes formulações da AMB foram
desenvolvidas, tais como anfotericina B complexo lipídico, anfotericina B dispersão coloidal
e anfotericina B lipossomal, com o intuito de melhorar sua tolerabilidade, especialmente,
diminuir a nefrotoxicidade sem alterar a eficácia (CATALÁN; MONTEJO, 2006). Embora
essas formulações apresentem uma menor nefrotoxicidade que a AMB convencional ainda

33
são mais nefrotóxicas que os triazóis e as equinocandinas (CHAPMAN; SULLIVAN;
CLEARY, 2008).
Os derivados azólicos são compostos totalmente sintéticos, que são utilizados na
terapia antifúngica, e podem ser divididos em dois grupos: os imidazólicos e os triazólicos
(MARTINEZ, 2006; CHAPMAN; SULLIVAN; CLEARY, 2008). Os imidazólicos possuem
um anel pentagonal formado por três átomos de carbono e dois átomos de nitrogênio e são
representados pelo miconazol e o cetoconazol. A partir do núcleo imidazólico geraram os
triazólicos que apresentam dois átomos de carbono e três átomos de nitrogênio, representados
pelo itraconazol e o fluconazol (CATALÁN; MONTEJO, 2006). O mecanismo de ação dos
derivados azólicos fundamenta-se na inibição da biossíntese do ergosterol em diferentes
etapas, atuam inibindo a enzima do citocromo P450 dos fungos, o que acarreta bloqueio na
demetilação do lanosterol e síntese de ergosterol, e altera a permeabilidade da membrana e a
viabilidade fúngica. Agem também modificando a síntese de lipídeos e inativando enzimas do
processo oxidativo dos fungos (MARTINEZ, 2006; CATALÁN; MONTEJO, 2006;
CHAPMAN; SULLIVAN; CLEARY, 2008). A deficiência de ergosterol provoca acúmulo de
compostos metilesteróis, levando ao desenvolvimento de uma membrana com propriedades
alteradas, não satisfazendo as funções fundamentais para o desenvolvimento fúngico
(BERGOLD; GEORGIADIS, 2004).
Os triazólicos são os antifúngicos mais utilizados e têm atividade contra muitos
patógenos fúngicos, com nenhum dos graves efeitos nefrotóxicos observados com a
administração de anfotericina B. Os imidazólicos, tais como o cetoconazol, vêm sendo
substituídos pelos triazólicos, por apresentarem uma melhor segurança e eficácia clínica. Os
derivados azólicos exibem amplo espectro de ação in vitro frente às principais espécies de
fungos patogênicos, incluindo leveduras dos gêneros Candida, Cryptococcus e os fungos
filamentosos – Aspergillus spp., Penicillium spp., Acremonium spp., Sporothrix schenckii,
bem como também isolados de Fusarium spp., Scedosporium apiospermum dentre outros
(CUENCA-ESTRELLA et al., 2006). O espectro de ação dos triazólicos inclui os agentes das
principais infecções fúngicas sistêmicas atuais: P. brasiliensis, H. capsulatum, C. immitis, B.
dermatitidis, C. neoformans e a maioria das espécies de Candida, sendo que C. krusei e C.
glabrata são menos sensíveis (MARTINEZ, 2006). O itraconazol, por exemplo, é fungistático
contra espécies de Candida, enquanto o voriconazol e o posaconazol são fungicidas contra
espécies de Aspergillus (CHAPMAN; SULLIVAN; CLEARY, 2008).

34
As equinocandinas são constituídas por um grupo semi-sintético de agentes
antifúngicos, que foram isolados, pela primeira vez em 1974, de culturas de Aspergillus spp.
(GOBERNADO; CANTÓN, 2008). A primeira aprovação comercial pelo FDA deu-se apenas
em 2001, com a caspofungina, seguida da micafungina em 2005 e anidulafungina em 2006.
Atualmente, as equinocandinas são consideradas um importante grupo de drogas no
tratamento da candidíase superficial e invasiva e de aspergiloses (CHAPMAN; SULLIVAN;
CLEARY, 2008). O mecanismo de ação destas drogas é caracterizado pela inibição não
competitiva e irreversível de (1,3)-β-D-glucano sintase, enzima responsável pela síntese do
(1,3)-β-D-glucano, um importante polissacarídeo da parede celular da maioria dos fungos
patogênicos, responsável pela rigidez da parede, assim como a integridade osmótica,
crescimento e divisão da célula (CHANDRASEKAR; SOBEL, 2006). Na ausência deste
polissacarídeo, as células fúngicas sofrem danos na estrutura e integridade da parede celular,
provocando assim a morte do microrganismo (GOBERNADO; CANTÓN, 2008). As
equinocandinas apresentam atividade fungicida contra Candida spp. e efeito fungistático
contra Aspergillus spp. O espectro de ação da caspofungina inclui também outras espécies
fúngicas, tais como: Scedosporium apiospermum, B. dermatitidis, Coccidioides spp. e H.
capsulatum (CHAPMAN; SULLIVAN; CLEARY, 2008). No entanto, as equinocandinas não
possuem atividade in vitro frente à C. neoformans, Trichosporon asahii, Rhizopus spp. e
Fusarium spp. (CATALÁN; MONTEJO, 2006).
Com o aumento da prevalência das micoses e da variabilidade de drogas para o
tratamento das micoses, a padronização de métodos de testes de sensibilidade in vitro se fez
cada vez mais necessária (ODDS et al., 1998). Os testes de sensibilidade antifúngica, in vitro,
visam avaliar o potencial terapêutico do medicamento, comprovar a sensibilidade do fungo
frente aos antifúngicos e obter informações para o controle da terapia antifúngica, a fim de
tornar a escolha da terapêutica mais adequada e eficiente. Atualmente, as técnicas adotadas
para a análise da atividade de agentes antifúngicos seguem normas de referência padronizadas
pelos órgãos internacionais European Committee on Antimicrobial Susceptibility Testing
(EUCAST) e Clinical and Laboratory Standards Institute (CLSI) (HOSPENTHAL;
MURRAY; RINALDI, 2004), os quais propõem como principais métodos a disco-difusão, a
diluição em ágar e a diluição em caldo. Estes métodos permitem aferir a sensibilidade e
identificar a concentração inibitória mínima (CIM) da droga, capaz de inibir o crescimento do
microrganismo em estudo (STOPPA et al., 2009). Apesar de não haver uma relação direta
entre a sensibilidade in vitro com a in vivo, os resultados encontrados in vitro podem prever a

35
resposta clínica ou, pelo menos, a falha terapêutica (REX et al., 2001; HOSPENTHAL;
MURRAY; RINALDI, 2004).
O comitê europeu AFST-EUCAST - Antifungal Susceptibility Testing Subcommittee
of the European Committee on Antimicrobial Susceptibility desenvolve metodologias padrões
para testes de sensibilidade antifúngica, com a finalidade de alcançar maior reprodutibilidade
e exatidão na determinação dos valores de CIM. O CLSI é uma organização qualificada para a
criação de normas de referência que garantem a padronização técnica dos testes de
sensibilidade antifúngica, tratando de assuntos como a seleção e preparação dos agentes
antifúngicos, a interpretação dos testes, bem como, o uso de testes de controle de qualidade,
facilitando assim a comunicação e padronização entre os diversos laboratórios na utilização
dessas técnicas (HOSPENTHAL; MURRAY; RINALDI, 2004).
A norma M27-A aprovada em 1997, é destinada a testes de microdiluição em caldo
para avaliar a sensibilidade in vitro de leveduras, como Candida spp., e Cryptococcus
neoformans. Em 2002, este documento foi revisto e atualizado para o documento M27-A2
(CLSI, 2002a). Neste mesmo ano, foi publicado o documento M38-A, que se destina à
realização de testes de macrodiluição para avaliar a sensibilidade in vitro de fungos
filamentosos, incluindo espécies de Aspergillus spp., Fusarium spp., Rhizopus arrhizus,
Pseudallescheria boydii (Scedosporium apiospermum) e Sporothrix schenckii (CLSI, 2002b).
Em 2008, foi publicada uma atualização dos documentos anteriores, sendo o M27-A2
substituído pelo M27-A3. E o M38-A substituído pelo M38-A2, o qual incluiu a padronização
do teste para os gêneros Trichophyton, Microsporum, Epidermophyton e Scedosporium
prolificans, bem como, para espécies de demáceos e zigomicetos (CLSI, 2008a; CLSI,
2008b). Ambos os documentos M27-A2 e M38-A apresentam critérios a serem adotados para
a realização dos testes de sensibilidade, como: uso de RPMI (Roswell Park Memorial
Institute) – 1640, tamponado para pH 7,0 com ácido o 2-[N-morfolino]-propanossulfônico
(MOPS; Sigma) 0,165M, como meio de cultivo, tamanho e preparo do inóculo fúngico,
tempo e temperatura de incubação, leitura visual dos tubos ou placas de acordo com o teste e
determinação dos pontos de corte. Condições de ensaio para as drogas antifugicas tais como
anfotericina B, terbinafina, derivados azólicos, equinocandinas, dentre outros também estão
mencionados nestes documentos (CLSI, 2002a; 2002b).

36
2.3.1 Busca de novos agentes antifúngicos
Embora a terapia antifúngica comumente utilizada seja de modo geral eficiente para o
tratamento das micoses, algumas desvantagens podem ser apontadas, como a necessidade de
tratamento por tempo prolongado, bem como, a elevada toxicidade apresentada pela
anfotericina B (WHEAT et al., 2007; KAUFFMAN et al., 2007). Além disso, casos de
refratariedade e recidivas já foram descritos (SCHUBACH et al., 2004; WHEAT et al., 2006;
KAUFFMAN et al., 2007). Este cenário tem impulsionado a busca por novos agentes que
apresentem potencial antifúngico. Alguns estudos tem demonstrado que determinadas drogas
já empregadas clinicamente no tratamento de doenças de etiologia não-fúngica podem
apresentar atividade antifúngica relevante (BRILHANTE et al., 2010; CORDEIRO et al.,
2011).
2.3.1.1 Miltefosina
A miltefosina é uma molécula anfifílica, sua porção hidrofílica é constituída por um
grupo fosfocolina e sua porção hidrofóbica consiste em uma cadeia alquílica hexadecílica.
Este fármaco é um alquil-fosfolipídio análogo da fosfocolina que foi inicialmente
desenvolvido como agente anti-câncer. Posteriormente, foi constatada sua atividade frente à
Leishmania spp. e Trypanosoma cruzi, sendo aprovada para o tratamento da leishmaniose
desde 2002. Atualmente é o único agente oral usado para o tratamento de todos os tipos de
Leishmaniose, apresentando taxa de cura de 98% (KAMINSKY, 2002; AGRESTA et al.,
2003; CROFT; SEIFERT; DUCHENE, 2003; DE CASTRO et al., 2004; MISHRA;
SAXENA; SINGH, 2007; DORLO et al., 2012). Mais recentemente a miltefosina também
mostrou ação antifúngica relevante, in vitro, frente a leveduras dos gêneros Candida spp. e
Cryptococcus spp., bem como, frente a fungos filamentosos, tais como, dermatófitos,
Aspergillus spp., Fusarium spp. e Scedosporium spp. (WIDMER et al., 2006; TONG et al.,
2007).
Figura 3. Formula estrutural da miltefosina
Fonte: DORLO et al., 2012.

37
Atualmente, diferentes produtos patentiados da miltefosina estão disponíveis no
mercado, tais como Impavido W (produto farmacêutico humano sólido oral), Miltex W
(produto farmacêutico humano líquido tópico) e Milteforan W (produto farmacêutico
veterinário oral líquido). A miltefosina (Impavido W) foi aprovado para o tratamento da
Leishmaniose visceral no Nepal, e ambas Leishmaniose visceral e Leishmaniose cutânea na
Argentina, Bangladesh, Bolívia, Colômbia, Equador, Alemanha, Guatemala, Honduras, Índia,
México, Paquistão, Paraguai e Peru. Além disso, a miltefosina foi designada como um
medicamento órfão para o tratamento para leishmaniose visceral, tanto pela Agência Européia
de Medicamentos, em 2002 e pelo Food and Drug Administration (FDA) nos EUA, em 2006.
Recentemente, a droga também foi adicionada à Lista Modelo da OMS de Medicamentos
Essenciais (DORLO et al., 2012).
A miltefosina pertence a classe de tensoativos zwitteriônicos, sendo estruturalmente
semelhantes aos fosfolipídios presentes nas membranas biológicas (CASTRO et al., 2001;
MACDONALD et al., 1991). Desta forma, possui a capacidade de interagir com a membrana
de células tumorais e de protozoários, atingindo rapidamente outras membranas subcelulares,
sendo capaz de afetar o metabolismo em diferentes níveis (HUANG et al., 2005; MARCO et
al., 2009). Segundo Moreira et al. (2013), a miltefosina penetra na membrana através do seu
componente lipídico e interage com as cadeias laterais hidrofóbicas na interface lipídeo-
proteína. Através de uma ação de detergente, a miltefosina pode formar estruturas
semelhantes a micelas com as proteínas e, assim, favorecer conformações mais expandidas e
dinâmicas. Proteínas com maior hidrofobicidade pode fazer com que os grupos hidrófilos de
miltefosina penetrem na celula e, assim, levar à ruptura da mesma (MOREIRA et al., 2013).
(MOREIRA et al., 2013; DORLO et al., 2012).Portanto, a membrana celular parece ser um
dos sítios de ação primário deste fármaco (WIEDER et al., 1999).
Em estudos realizados por Zuo et al. (2011), foi demonstrado em Saccharomyces
cerevisiae a miltefosina é rapidamente absorvida e penetra na membrana mitocondrial interna,
onde interrompe o potencial de membrana, inibindo a enzima citocromo c oxidase (COX),
induzindo a apoptose celular. Outro mecanismo proposto de ação antifúngica está relacionado
com a inibição da fosfolipase B extracelular, produzida por diversos fungos, sendo importante
para a sobrevivência dos mesmos e para a disseminação da doença (WIDMER et al., 2006).
No entanto, concentrações elevadas de miltefosina foram necessárias para estabelecer apenas
uma inibição mínima da fosfolipase B e, portanto, não é provável que este é o alvo principal
da miltefosina (OBANDO et al., 2007).

38
Ainda em estudos realizados por Zuo et al. (2011), dois marcadores intrínsecos de
apoptose, como a ativação das caspases e fragmentação do DNA nuclear, foram procurados
em células de levedura tratadas com miltefosina e não tratadas, utilizando citometria de fluxo
e microscopia de fluorescência. A fragmentação do DNA e ativação de caspases celulares foi
evidente em culturas celulares tratadas com miltefosina. (THEVISSEN et al., 2008). Os genes
envolvidos na morte celular programada em S. cerevisiae foram também identificados como
reguladores de apoptose em células de mamíferos, o que indica que o mecanismo básico de
apoptose está de fato presente e funcional. Ademais, em experimentos realizados por
Khademvatan; Gharavi; Saki (2011) foram analisadas as atividades dos genes metacaspases e
proteína PARP em Leishmania infantum, onde mostrou que a miltefosina cliva a protéina
PARP (ADP-ribose) uma enzima de reparação do DNA e ativa as caspases um interuptor da
função central para desencadear a apoptose celular. As caspases induzem uma cascata de
reações que conduzem finalmente a morte celular. A participação das metacaspases em
leveduras tem sido estudada e revelou que as mesmas desempenham um papel crucial na
indução da morte celular por apoptose em resposta a vários genes de stress, envelhecimento e
deteorização de algumas funções biológicas (KHADEMVATAN; GHARAVI; SAKI, 2011).
Os mecanismos de ação baseados na indução de apoptose e ruptura da membrana
celular são consistentes com o amplo espectro de atividade da miltefosina contra fungos
patogénicos e protozoários, bem como, ante alguns tipos de células tumorais (MOREIRA et
al., 2013). Contudo, apesar de várias hipóteses potenciais para o mecanismo de ação da
miltefosina, já terem sido sugeridos nos últimos anos, nenhum mecanismo foi identificado
definitivamente. A multiplicidade de mecanismos potenciais propostos e estudos
contraditórios pode indicar que miltefosina possui mais de um local de ação molecular
(DORLO et al., 2012).
Embora da capacidade antifúngica da miltefosina tenha sido previamente
demonstrada, nenhum estudo foi realizado sobre seu potencial frente a esses dois fungos
dimórficos. Baseado no exposto, e devido ao constante aumento de casos de histoplasmose e
esporotricose em diversas regiões do Brasil, e a consequente busca por novas drogas para o
tratamento dessas micoses, houve um interesse em avaliar pela primeira vez, a atividade
inibitória in vitro da miltefosina ante a Histoplasma capsulatum var. capsulatum e a diferentes
espécies do complexo S. schenckii.

39
3 PERGUNTAS DE PARTIDA
1. A miltefosina apresenta atividade antifungica in vitro frente aos fungos dimórficos
Histoplasma capsulatum var. capsulatum e Sporothrix spp.?
2. A miltefosina apresenta potencial antifúngico semelhante ante aos fungos dimórficos
Histoplasma capsulatum var. capsulatum e Sporothrix spp.?

40
4 HIPÓTESES
1. A miltefosina é capaz de inibir o crescimento de Histoplasma capsulatum var.
capsulatum e Sporothrix spp in vitro.
2. A miltefosina apresenta potencial antifúngico similar ante Histoplasma capsulatum
var. capsulatum e Sporothrix spp. não apresentando diferença significativa nos valores
Concentração Inibitoria Mínima e Concentração Fungicida Mínima obtidos para estes
fungos.

41
5 OBJETIVOS
5.1 Objetivo geral
Avaliar o efeito inibitório in vitro de miltefosina frente a cepas de e Histoplasma
capsulatum var. capsulatum e Sporothrix spp.
5.2 Objetivos específicos
1. Determinar a concentração inibitória mínima (CIM) in vitro, frente a cepas de
Histoplasma capsulatum var. capsulatum e Sporothrix spp.
2. Determinar a concentração fungicida mínima (CFM) in vitro, frente a cepas de
Histoplasma capsulatum var. capsulatum e Sporothrix spp.

42
6 MATERIAL E MÉTODOS
6.1 Microrganismos
Para realização deste estudo, foi utilizado um total de 52 cepas de H. capsulatum var.
capsulatum na fase filamentosa (50 humanas e 2 animais) e 15 cepas na fase leveduriforme
(13 humanas e 2 animais). Todos os isolados são pertencentes ao Centro Especializado em
Micologia Médica (CEMM) da Universidade Federal do Ceará (UFC). Quanto às espécies do
complexo Sporothrix schenckii, foram utilizadas 62 cepas na fase filamentosa, incluindo
espécies de S. brasiliensis (n=48), S. schenckii (n=6); S. mexicana (n=5) e S. globosa (n=3).
Deste total, estão presentes isoaldos de origem de humana (n=25), origem animal (n=34) e
origem ambiental (n=3), todos cedidos pelo Laboratório de Micologia Médica Molecular da
Universidade Federal de São Paulo. Todos os procedimentos da pesquisa envolvendo cepas
de H. capsulatum var. capsulatum foram realizados em cabine de segurança biológica classe
II em laboratório de nível de biossegurança 3 (NB3), em virtudo da elevada virulência deste
fungo.
As culturas de H. capsulatum var. capsulatum na fase filamentosa encontravam-se
estocadas em ágar batata dextrose (Difco, Detroit, E.U.A.) acrescido de glicerol a 10%
(Cromoline Química Fina Ltda, São Paulo) a -20°C ou em solução salina estéril a 4°C, as
quais foram previamente identificadas por meio de análise microscópica direta e cultura, e por
técnicas imunológicas e moleculares. As cepas retiradas do estoque foram recuperadas
mediante semeadura em ágar batata, as quais foram submetidas à análise macroscópica,
levando-se em consideração relevo, textura e pigmentação das colônias após 7-10 dias de
incubação a 25-28ºC, e análise microscópica, através da observação de fragmentos da colônia
em microscópio óptico, a partir da montagem de lâmina e lamínula em lactofenol azul-
algodão. As culturas foram consideradas positivas para H. capsulatum var. capsulatum pela
presença de hifas hialinas finas e macroconídios tuberculados (WANKE; LAZÉRA, 2004).
Para a obtenção das culturas de H. capsulatum var. capsulatum na fase leveduriforme, a
conversão da fase filamentosa para a fase leveduriforme foi realizada através do cultivo das
culturas em ágar Sabouraud ou BHI suplementados com sangue de carneiro a 10%, incubadas
a 35°C e mantidas por repiques semanais, sendo analisada posteriormente a macromorfologia
e a micromorfologia (BRILHANTE et al., 2010).
As culturas de S. schenckii na fase filamentosa encontravam-se estocadas em ágar
batata dextrose (Difco, Detroit, EUA), acrescido de glicerol 10% (Cromoline Química Fina

43
Ltda, São Paulo) a -20°C ou em solução salina estéril a 4°C. As cepas previamente
identificadas e cedidas pelo Laboratório de Micologia Médica Molecular da Universidade
Federal de São Paulo – UNIFESP, foram submetidas a análise micológica para avaliação de
pureza e viabilidade. As cepas retiradas do estoque foram recuperadas mediante semeadura
em ágar batata, as quais foram submetidas à análise macroscópica, levando-se em
consideração relevo, textura e pigmentação das colônias após 7-10 dias de incubação a 25-
28ºC. Uma análise microscópica foi realizada através da observação de fragmentos da colônia
em microscópio óptico, a partir da montagem de lâmina e lamínula em lactofenol azul-
algodão. As culturas foram consideradas positivas para S. schenckii, na presença de hifas fina
e delicadas, hialinas, septadas com conídios dispostos nas extremidades dos conidióforos
assemelhando-se a rosetas. (MARQUES-EVANGELHISTA, 2010).
A origem de cada isolado de H. capsulatum incluído neste estudo encontra-se descrita
nos Quadros 1 e 2. Quanto às espécies do complexo Sporothrix schenckii, a origem de todas
as cepas utilizadas encontram-se descritas no Quadro 3.
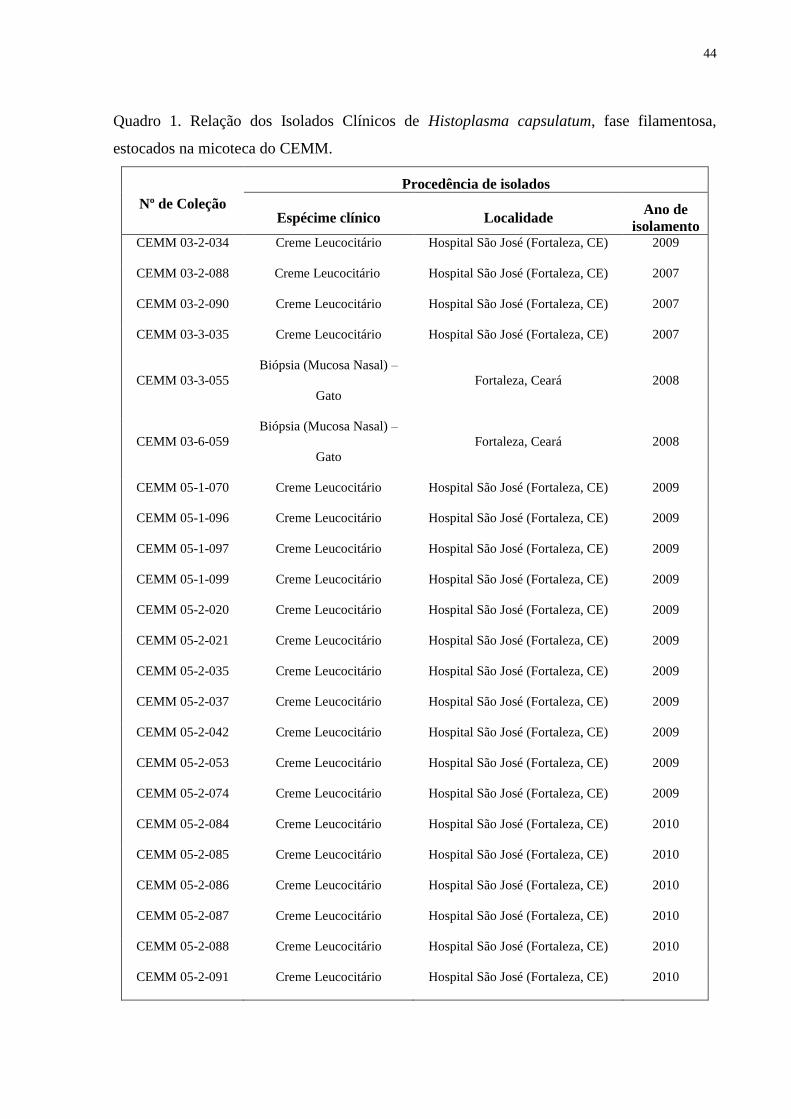
44
Quadro 1. Relação dos Isolados Clínicos de Histoplasma capsulatum, fase filamentosa,
estocados na micoteca do CEMM.
Nº de Coleção
Procedência de isolados
Espécime clínico Localidade Ano de
isolamento CEMM 03-2-034 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 03-2-088 Creme Leucocitário Hospital São José (Fortaleza, CE) 2007
CEMM 03-2-090 Creme Leucocitário Hospital São José (Fortaleza, CE) 2007
CEMM 03-3-035 Creme Leucocitário Hospital São José (Fortaleza, CE) 2007
CEMM 03-3-055
Biópsia (Mucosa Nasal) –
Gato
Fortaleza, Ceará 2008
CEMM 03-6-059
Biópsia (Mucosa Nasal) –
Gato
Fortaleza, Ceará 2008
CEMM 05-1-070 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-1-096 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-1-097 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-1-099 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-020 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-021 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-035 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-037 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-042 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-053 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-074 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-084 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-2-085 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-2-086 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-2-087 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-2-088 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-2-091 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010

45
Nº de Coleção
Procedência de isolados
Espécime clínico Localidade Ano de
isolamento CEMM 05-2-093 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-3-014 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-3-015 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-3-016 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-3-017 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-3-018 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-3-019 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-3-023 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-3-025 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-3-040 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-3-042 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-3-044 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-3-045 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-4-015 Biopsia São Paulo (SUDESTE) 2007
CEMM 05-4-024 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-4-026 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-4-028 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-5-011 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-6-010 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-6-011 Medula Óssea Hospital São José (Fortaleza, CE) 2009
CEMM 05-6-012 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-6-013 Creme Leucocitário Hospital São José (Fortaleza, CE) 2012
CEMM 05-6-014 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-6-017 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-6-022 Creme Leucocitário Hospital São José (Fortaleza, CE) 2012
CEMM 05-6-026 Creme Leucocitário Hospital São José (Fortaleza, CE) 2012
CEMM 05-6-027 Creme Leucocitário Hospital São José (Fortaleza, CE) 2012
CEMM 05-6-030 Creme Leucocitário Hospital São José (Fortaleza, CE) 2012
CEMM 05-6-066 Creme Leucocitário Hospital São José (Fortaleza, CE) 2012

46
Quadro 2. Relação dos Isolados Clínicos de Histoplasma capsulatum, fase leveduriforme,
utilizados na pesquisa.
Nº de Coleção
Procedência de isolados
Espécime clínico Localidade Ano de
isolamento
CEMM 03-2-034 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 03-2-090 Creme Leucocitário Hospital São José (Fortaleza, CE) 2007
CEMM 03-3-035 Creme Leucocitário Hospital São José (Fortaleza, CE) 2007
CEMM 03-3-055 Biópsia (Mucosa Nasal) -
Gato Fortaleza, Ceará 2008
CEMM 03-6-059 Biópsia (Mucosa Nasal) -
Gato Fortaleza, Ceará 2008
CEMM 05-1-097 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-035 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-037 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-042 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-053 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-074 Creme Leucocitário Hospital São José (Fortaleza, CE) 2009
CEMM 05-2-087 Creme Leucocitário Hospital São José (Fortaleza, CE) 2010
CEMM 05-4-028 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-5-011 Creme Leucocitário Hospital São José (Fortaleza, CE) 2011
CEMM 05-6-027 Creme Leucocitário Hospital São José (Fortaleza, CE) 2012

47
Quadro 3. Relação das espécies do complexo Sporothrix schenckii estocados na micoteca do
CEMM.
Código
UNIFESP Espécie Nº de Coleção
Procedência de isolados
Isolado Localidade Ano de
isolamento
Ss 01 S. schenckii CEMM 05-3-048 Gato SP 2004
Ss 05 S. brasiliensis CEMM 05-3-049 Gato MG 2004
Ss 53 S. brasiliensis CEMM 05-3-050 Gato RS 2004
Ss 54 S. brasiliensis CEMM 05-3-051 Gato RS 2004
Ss 151 S. brasiliensis CEMM 05-3-052 Cão RS 2006
Ss 152 S. brasiliensis CEMM 05-3-053 Gato RS 2006
Ss 153 S. brasiliensis CEMM 05-3-054 Gato RS 2006
Ss 154 S. brasiliensis CEMM 05-3-055 Gato RS 2006
Ss 155 S. brasiliensis CEMM 05-3-056 Gato RS 2006
Ss 156 S. brasiliensis CEMM 05-3-057 Gato RS 2006
Ss 157 S. brasiliensis CEMM 05-3-058 Gato RS 2006
Ss 171 S. brasiliensis CEMM 05-3-059 Gato SP 2010
Ss 172 S. brasiliensis CEMM 05-3-060 Gato PR 2010
Ss 173 S. brasiliensis CEMM 05-3-061 Gato PR 2010
Ss 174 S. brasiliensis CEMM 05-3-062 Gato PR 2010
Ss 226 S. brasiliensis CEMM 05-3-063 Gato SP 2010
Ss 227 S. brasiliensis CEMM 05-3-064 Cão SP 2010
Ss 245 S. brasiliensis CEMM 05-3-065 Gato RJ 2010
Ss 246 S. brasiliensis CEMM 05-3-066 Gato RJ 2010
Ss 247 S. brasiliensis CEMM 05-3-067 Gato RJ 2010
Ss 248 S. brasiliensis CEMM 05-3-068 Gato RJ 2010
Ss 249 S. brasiliensis CEMM 05-3-069 Gato RJ 2010
Ss 250 S. brasiliensis CEMM 05-3-070 Gato RJ 2010
Ss 251 S. brasiliensis CEMM 05-3-071 Gato RJ 2010
Ss 252 S. brasiliensis CEMM 05-3-072 Gato RJ 2010

48
Código
UNIFESP Espécie Nº de Coleção
Procedência de isolados
Isolado Localidade Ano de
isolamento
Ss 253 S. brasiliensis CEMM 05-3-073 Gato RJ 2010
Ss 254 S. brasiliensis CEMM 05-3-074 Gato RJ 2010
Ss 255 S. brasiliensis CEMM 05-3-075 Gato RJ 2010
Ss 256 S. brasiliensis CEMM 05-3-076 Gato RJ 2010
Ss 257 S. brasiliensis CEMM 05-3-077 Gato RJ 2010
Ss 258 S. brasiliensis CEMM 05-3-078 Gato RJ 2010
Ss 259 S. brasiliensis CEMM 05-3-079 Gato RJ 2010
Ss 260 S. brasiliensis CEMM 05-3-080 Gato RS 2010
Ss 264 S. brasiliensis CEMM 05-3-081 Gato RJ 2010
Ss 65 S. brasiliensis CEMM 05-3-082 Humano RJ 2004
Ss 66 S. brasiliensis CEMM 05-3-083 Humano RJ 2004
Ss 67 S. brasiliensis CEMM 05-3-084 Humano RJ 2004
Ss 68 S. brasiliensis CEMM 05-3-085 Humano RJ 2004
Ss 69 S. brasiliensis CEMM 05-3-086 Humano RJ 2004
Ss 70 S. brasiliensis CEMM 05-3-087 Humano RJ 2004
Ss 71 S. brasiliensis CEMM 05-3-088 Humano RJ 2004
Ss 72 S. brasiliensis CEMM 05-3-089 Humano RJ 2004
Ss 73 S. schenckii CEMM 05-3-090 Humano RJ 2004
Ss 74 S. brasiliensis CEMM 05-3-091 Humano RJ 2004
Ss 75 S. schenckii CEMM 05-3-092 Humano RJ 2004
Ss 76 S. brasiliensis CEMM 05-3-093 Humano RJ 2004
Ss 77 S. brasiliensis CEMM 05-3-094 Humano RJ 2004
Ss 78 S. schenckii CEMM 05-3-095 Humano RJ 2004
Ss 79 S. brasiliensis CEMM 05-3-096 Humano RJ 2004
Ss 80 S. schenckii CEMM 05-3-097 Humano RJ 2004
Ss 177 S. brasiliensis CEMM 05-3-098 Humano RJ 2009

49
Código
UNIFESP Espécie Nº de Coleção
Procedência de isolados
Isolado Localidade Ano de
isolamento
Ss 178 S. brasiliensis CEMM 05-3-099 Humano RJ 2009
Ss 181 S. mexicana CEMM 05-3-100 Vegetal México 2005
Ss 182 S. mexicana CEMM 05-3-048 Solo México 2009
Ss 243 S. schenckii CEMM 05-3-049 Solo Brasil 2010
Ss 265 S. brasiliensis CEMM 05-3-050 Humano MG 2010
Ss 06 S. globosa CEMM 05-3-051 Humano MG 2004
Ss 41 S. globosa CEMM 05-3-052 Humano RS 2004
Ss 49 S. globosa CEMM 05-3-053 Humano RS 2004
Ss 131 S. mexicana CEMM 05-3-054 Humano RS 2004
Ss 132 S. mexicana CEMM 05-3-055 Humano RS 2004
Ss 133 S. mexicana CEMM 05-3-056 Humano RS 2004
6.2 Agentes antimicrobianos
Para os testes de sensibilidade foi utilizada uma solução comercial de miltefosina
(MIL) (Sigma Chemical Corporation, EUA). Para o preparo da solução-estoque, o fármaco foi
diluído segundo o fabricante em água destilada estéril. As soluções estoque dos antifúngicos
anfotericina B (AMB) (Sigma Chemical Corporation, USA) e itraconazol (ITC) (Janssen
Pharmaceutica, Belgium) foram testadas como controle. Estes fámacos foram diluídos em
DMSO a 100%, tal como recomendado pelo CLSI (CLSI, 2008a; CLSI, 2008b). Estas
soluções também foram estocadas em freezer a -20 oC até o momento do uso. Posteriormente,
todos os agentes antimicrobianos foram diluídos em RPMI 1640 com L-glutamina e sem
bicarbonato de sódio (Sigma), tamponado a pH 7.0 com ácido 3-[N-morfolino]-
propanosulfônico – MOPS 0,165 M (Sigma) (CLSI, 2008a; CLSI, 2008b). Para ensaio de
microdiluição em caldo, a solução do composto encerrava uma concentração quatro vezes
maior que a maior concentração-teste inicial. Os intervalos de concentração testados para cada
droga isolada foram: MIL (0,0312 a 16 µg/mL), AMB (0,0312 a 16µg/mL), ITC (0,0312 a 16
µg/mL) para H. capsulatum var. capsulatum em ambas as fases e MIL (0,0312 a 16 µg/mL),
AMB (0,0312 a 16µg/mL), ITC (0, 0312 a 16 µg/mL) para Sporothrix spp. na fase
filamentosa.

50
6.3 Preparo do inóculo para teste de sensibilidade
Os inóculos das cepas de H. capsulatum var. capsulatum e Sporothrix spp. na fase
filamentosa foram preparados a partir de culturas previamente crescidas em meio ágar BHI
(Brain Heart Infusion) (Himedia, India) e incubadas por 6-7 dias a 25-28°C (BRILHANTE et
al., 2010; ALVARADO-RAMIREZ; TORRES-RODRIGUEZ, 2007). Para H. capsulatum
var. capsulatum na fase leveduriforme, as cepas foram cultivadas em ágar BHI suplementado
com sangue de carneiro a 10% e incubadas por 7 dias a 35 °C (BRILHANTE et al., 2010).
O procedimento seguiu através da adição de 1 mL de solução salina 0,9% estéril nas
culturas fúngicas e realização de leves raspagens na superfície do micélio com o auxílio de
uma alça microbiológica, a fim de obter uma suspenção livre de fragmentos de meio de
cultura. As suspensões foram transferidas para um tubo de ensaio estéril contendo 4 mL de
solução salina 0,9%. Vale ressaltar que o preparo do inóculo para as cepas de H. capsulatum
na fase leveduriforme deu-se através da transferência de uma pequena quantidade da colônia
fúngica para 4 mL de solução salina 0,9% com o auxílio de uma alça microbiológica, sendo
levadas ao agitador magnético por 10 segundos.
Em seguida, cada suspensão fúngica foi ajustada a escala 0,5 de McFarland para H.
capsulatum var. capsulatum em ambas as fases (BRILHANTE et al., 2010) e escala 2 de
McFarland para Sporothrix spp. (ALVARADO-RAMIREZ; TORRES-RODRIGUEZ, 2007)
e submetidas a homogenização manual sendo deixadas em repouso por 5 minutos. As
suspensões de H. capsulatum var. capsulatum em ambas as fases foram então submetidas à
leitura em espectrofotômetro a um comprimento de onda de 530 nm e transmitância ajustada
para 95%, sendo diluídas na proporção de 1:10 em meio RPMI 1640 para obtenção de uma
concentração final de 0,5 x 103 a 2,5 x 10
4 UFC mL
-1 (BRILHANTE et al., 2010). Para
Sporothrix spp. as suspensões foram diluídas da mesma forma e foi realizada a quantificação
de UFC mL-1
em placas de ágar batata, para obtenção de uma concentração final de 1 x 105 a
5 x 105 UFC mL
-1 (ALVARADO-RAMIREZ; TORRES-RODRIGUEZ, 2007).
6.4 Testes de Sensibilidade in vitro
A atividade antifúngica in vitro foi determinada pelo método de microdiluição em
caldo, de acordo com o protocolo descrito nos documentos M27-A3 com adaptações para H.
capsulatum var. capsulatum em ambas as fases, e M38-A2 com adaptações para Sporothrix
spp., ambos documentos padronizados pelo Clinical and Laboratory Standards Institute -

51
CLSI (CLSI, 2008a; CLSI, 2008b). A miltefosina foi testada frente a cepas de H. capsulatum
var. capsulatum, na fase filamentosa e na fase leveduriforme e as cepas de Sporothrix spp.
Com relação ao controle de qualidade do teste, foram utilizadas as cepas Candida
parapsilosis ATCC 22019 e Candida krusei ATCC 6258, de acordo com o documento M27-
A3 do CLSI (CLSI, 2008a).
No ensaio de microdiluição, primeiro foram distribuídos 100 µL do meio RPMI 1640
tamponado a pH 7,0 com MOPS 0,165 M em cada poço da microplaca contendo 96 poços
com fundo arredondado em forma de U. Em seguida, foi realizada uma diluição em série das
drogas, adicionando 100 µL ao primeiro poço, a partir do qual, após homogeneização, foram
transferidos 100 µL ao segundo poço, sendo este procedimento repetido sucessivamente nos
demais poços até os poços da décima coluna, onde 100 µL foram então retirados e
descartados. As concentrações das drogas testadas variaram conforme apresentadas
anteriormente. Em seguida, foram adicionados aos poços 100 µL do inóculo fúngico
previamente preparado. Controles positivos de crescimento e esterilidade foram incluídos para
cada observação testada. Os procedimentos foram realizados em duplicata.
A leitura resultados para a determinação das CIMs foi realizada visualmente após 4 e 7
dias de incubação a 35°C para H. capsulatum var. capsulatum em fase leveduriforme e em
fase filamentosa, respectivamente e 3 dias de incubação a 35°C para Sporothrix spp.
(BRILHANTE et al., 2010; ALVARADO-RAMIREZ; TORRES-RODRIGUEZ, 2007). As
concentrações inibitórias mínimas (CIM) para MIL foram definidas como a concentração
mais baixa de droga capaz de inibir 80% do crescimento dos fungos (WIDMER et al., 2006),
quando comparado com o controlo de crescimento, mas as concentrações mínimas que
provocaram 50% e 100% de inibição do crescimento fúngico foram também observadas.
Como para as drogas de controle, os CIMs para AMB foram definidos como as mais baixas
concentrações capazes de inibir 100% de crescimento de fungos, enquanto que para ITC, os
CIMs foram definidos como a concentração mais baixa de droga capaz de inibir 50% de
crescimento fúngico, quando comparado com o respectivo controle crescimento
(BRILHANTE et al., 2010). Após a leitura da CIM uma alíquota de 100 µL, dos poços que
não apresentaram crescimento fúngico visível, foi transferida para tubos contendo ágar batata
e incubados por 7 dias a 35 °C. A Concentração Fungicida Mínima (CFM) foi determinada
como a menor concentração capaz de inibir em 100% o crescimento de colônias fúngicas
(CANTON et al., 2003).

52
6.5 Análise estatística
Os valores de CIM obtidos (inibição de 80%) e os valores de CFM para miltefosina
contra H. capsulatum na fase filamentosa e leveduriforme e as espécies Sporothrix foram
comparados por análise de variância (ANOVA). Resultados com P <0,05 foram considerados
estatisticamente significativos.

53
7 RESULTADOS
A Miltefosina foi capaz de inibir o crescimento de todas as cepas testadas, com CIMs
≤ 2 µg mL-1
e CFMs ≤ 4 µg mL-1
(Tabela 1). Para H. capsulatum var. capsulatum na fase
filamentosa os valores de CIM variaram de 0,25 a 2 µg mL-1
(média geométrica 0, 75 g mL-
1) e valores de CFM variando de 0,5 a 4 µg mL
-1 (média geométrica 1,80 g mL-1
). Todos os
dados estão apresentados no Quadro 4 – Anexo A. Para H. capsulatum var. capsulatum na
fase Leveduriforme os valores de MIC variando de 0,125 a 1 µg mL-1
(média geométrica 0,34
g mL-1) e valores de CFM variando de 0,25 a 4 µg mL
-1 (média geométrica 0,95 g mL-1
).
Todos os dados estão apresentados no Quadro 5 – Anexo A. Os MICs e CFMs frente a H.
capsulatum var. capsulatum na fase filamentosa foram estatisticamente superiores aos frente
as cepas de H. capsulatum var. capsulatum na fase leveduriforme (CIM: P = 0,0002; CFM: P
= 0,0015). Apresentado na Tabela 1. Para Sporothrix spp., o intervalo de CIM foi de 0,25 a 2
µg mL-1
(média geométrica 0,67 g mL-1) e os valores de CFM obtidos foram de 1 a 4 µg
mL1(média geométrica 1,81 g mL-1
). Todos os dados estão apresentados no Quadro 6 –
Anexo A.
Foram observadas diferenças estatisticamente significativas entre os CIMs frente as
diferentes espécies Sporothrix (P <0,01), mas não entre CFM. A Miltefosina apresentou
valores de CIM frente S. globosa superiores, quando comparado com S. brasiliensis (P
<0,0001), Sporothrix schenckii (P = 0,0009) e S. mexicana (P = 0,0085) e os valores de CIM
frente S. mexicana foram superiores aos de S. brasiliensis (P = 0,0417). Além disso, os
valores de CIM e CFM de miltefosina frente Sporothrix spp. foram semelhantes aos de
Histoplasma capsulatum na fase filamentosa, mas maiores do que H. capsulatum na fase
leveduriforme (CIM: P = 0,0021; CFM: P = 0,0017). Com relação ao controle de qualidade
das drogas, as cepas padrão Candida parapsilosis ATCC 22019 e Candida krusei ATCC
6258 apresentaram CIM para MIL, AMB e ITC de 1, 1 e 0,25 µg mL-1
, respectivamente, de
acordo com os dados descritos na literatura (Tabela 1).

54
Tabela 1. Atividade antifúngica in vitro de miltefosina frente H. capsulatum na fase filamentosa e leveduriforme
e Sporothrix spp.
Espécies n
Droga
MIL
CIM (µg. mL-1
) CFM
(µg. mL-1
) 50% Inibição 80% Inibição 100% Inibição
Histoplasma capsulatum (Fase filamentosa)
52 0.125 (8)a
0.25 (8) 0.5 (8) 0.5 (6)
0.25 (9) 0.5 (9) 1 (9) 1 (5)
0.5 (32) 1 (32) 2 (32) 2(32)
1 (3) 2 (3) 4 (3) 4 (9)
Histoplasma capsulatum (Fase leveduriforme)
15 0.0625 (1) 0.125 (1) 0.25 (1) 0.25 (1)
0.125 (8) 0.25 (8) 0.5 (8) 0.5 (3)
0.25 (4) 0.5 (4) 1 (4) 1 (7)
0.5 (2) 1 (2) 2 (2) 2 (4)
Sporothrix brasiliensis 48 0.125 (7) 0.25 (7) 0.5 (7) 1 (19)
0.25 (21) 0.5 (21) 1 (21) 2 (21)
0.5 (20) 1 (20) 2 (20) 4 (8)
Sporothrix schenckii 6 0.25 (2) 0.5 (2) 1 (2) 1 (1)
0.5 (4) 1 (4) 2 (4) 2 (4)
4 (1)
Sporothrix mexicana 5 0.25 (2) 0.5 (2) 1 (2) 1 (1)
0.5 (2) 1 (2) 2 (2) 2 (3)
1 (1) 2 (1) 4 (1) 4 (1)
Sporothrix globosa 3 0.5 (1) 1 (1) 2 (1) 2 (1)
1 (2) 2 (2) 4 (2) 4 (2) Legenda: MIL: Miltefosina, CIM: Concentração inibitória mínima, CFM: Concentração fungicida mínima, M: fase micelial, Y: fase leveduriforme, ªNúmero de cepas
testadas. Cepas controle: Candida Krusei ATCC 6258 (Anfotericina B: 1 µg/mL; Itraconazol: 0,25 µg/mL; MIL: 1 µg/mL); Candida parapsilosis ATCC 22019
(Anfotericina B: 1 µg/mL; Itraconazol: 0,25 µg/mL; MIL: 1 µg/mL).

55
8 DISCUSSÃO
Devido ao pequeno número de drogas antifúngicas disponíveis, muitos estudos estão
sendo realizados no sentido de ampliar o arsenal terapêutico antifúngico (BERGOLD;
GEORGIADIS, 2004). Apesar do desenvolvimento de novos fármacos, o aumento no número
de casos de micoses profundas e oportunistas tem sido motivo de preocupação para a
comunidade médica e científica (ESPINEL-INGROFF, 2009). Desta forma, vários
pesquisadores têm procurado por novos compostos e moléculas ativas com propriedades
antifúngicas que possam substituir ou vir a subsidiar a terapia convencional atual
(SCHUBACH et al., 2004; WHEAT, 2006; KAUFFMAN et al., 2007). Neste contexto,
estudos recentes tem investigado potencial antifúngico de fármacos não antifúngicos
comerciamente disponíveis, tais como agentes imunossupressores, antiparasitários,
antineoplásicos e antibacterianos (AFELTRA; VERWEIJ, 2003; AFELTRA et al., 2004;
WIDMER et al., 2006).
Tais estudos podem revelar-se como uma alternativa para o tratamento de infecções
fúngicas não responsivas ao tratamento convencional ou quando o patógeno apresenta
resistência (CUENCA-ESTRELLA et al., 2006; O’SHAUGHNESSY et al., 2006; CHOU,
2006; KAUFFMAN et al., 2007). Deste modo, estudos recentes tem investigado a atividade
inibitória contra fungos de “drogas não antifúngicas”, tanto isoladamente como em
combinação com agentes antifúngicos clássicos (BRILHANTE et al., 2010; CORDEIRO et
al., 2011; VENTURINE et al., 2011). Vários fármacos antineoplásicos citotóxicos têm
mostrado atividade antiparasitária e antifúngica. A miltefosina é um alquil-fosfolipídio,
desenvolvida como agente anti-câncer que apresenta ação antifúngica, in vitro, frente a
leveduras dos gêneros Candida spp. e Cryptococcus spp. e frente a fungos filamentosos, tais
como, dermatófitos, Aspergillus spp., Fusarium spp. e Scedosporium spp. (WIDMER et al.,
2006; TONG et al., 2007)
No presente estudo, a atividade antifungica in vitro da droga miltefosina, foi
investigado pela primeira vez ante aos fungos dimorficos H. capsulatum var. capsulatum e
Sporothrix spp. Desta forma, os resultados obtidos mostraram que a milteforsina foi capaz de
inibir todas as cepas testadas de ambos os gêneros fúngicos testados. Ademais, foi possível
observar que os intervalo de CIM encontrados para miltefosina (0,125 ≤ MIC ≤ 2 mg. ML-1)
ante a esses fungos dimórficos são semelhantes ou inferiores aos reportados por outras
espécies de fungos. Como relatado por Tong et al. (2007) para diferentes espécies de

56
dermatófitos, o intervalo de CIM para miltefosina ante as cepas dos gêneros Trichophyton,
Epidermophyton e Microsporum foi de 0,25 ≤ CIM ≤ 2 µg. mL-1
. Valores CIM mais elevados
foram apresentados por Widmer et al. (2006) contra Cryptococcus spp., (0,25 ≤ CIM ≤ 4 µg
mL-1
), Candida spp. (1 ≤ CIM ≤ 8 µg mL-1
), bem como, frente a diferentes espécies do gênero
Aspergillus (2 ≤ CIM ≤ 16 µg mL-1
). Tais relatos, ressaltam a relevância de nossos resultados
quanto ao potencial inibitório da miltefosina ante aos agentes patogênicos H. capsulatum var.
capsulatum e Sporothrix spp. É interessante mencionar que as CIMs e CFMs de miltefosina
obtidas nesta pesquisa (0,125 - 4 µg mL-1
), estão dentro do nível de concentração plasmática
atingível in vivo, a qual apresenta um valor médio de 30,8 µg mL-1
em seres humanos
(DORLO et al., 2008).
Em H. capsulatum, foram observadas diferenças significativas entre a fase filamentosa
e a fase leveduriforme, tanto nos valores de CIMs quanto nos CFMs obtidos para miltefosina,
onde os valores estatisticamente superiores (CIM: P=0,0002; CFM: P=0.0015) foram
observados para a fase filamentosa. Embora existam poucos dados disponíveis na literatura,
outros trabalhos também relataram valores de CIM maiores para fase filamentosa (ANDREU
et al., 2003). Nakai et al., em 2003, demonstraram que fluconazol apresentou CIMs
significativamente menores para fase leveduriforme quando comparadas as CIMs da fase
filamentosa. Contudo o mesmo não foi observado para os antifúngicos anfotericina B e
itraconazol, nos quais não foram observadas diferenças significativas entre as duas fases
(NAKAI et al., 2003). Por tratar-se da forma parasitária do microrganismo, grande parte dos
pesquisadores adotam os testes de sensibilidade com H. capsulatum na fase leveduriforme
(CONNOLLY et al., 1999; WHEAT et al., 2006), entretanto ainda não existem estudos que
definam em testes in vitro que fase morfológica o patógeno, melhor apresenta correlação com
a resposta terapêutica.
Em Sporothrix spp., foram observadas diferenças estatisticamente significativas entre
os CIMs frente as diferentes espécies complexo (P <0,01), mas não entre os valores de CFM.
A Miltefosina apresentou valores de CIM frente S. globosa superiores, quando comparado
com S. brasiliensis, Sporothrix schenckii e S. mexicana. Ademais, os valores de CIMs e
CFMs obtidos para miltefosina frente Sporothrix spp. foram semelhantes aos da fase
filamentosa de H. capsulatum, mas superiores aos de levedura de H. capsulatum (CIM: P=
0,0021; CFM: P= 0,0017). Em estudos realizados por Trilles et al. (2005), onde as CIMs
obtidas tanto para terbinafina como para o itraconazol, estudando a fase filamentosa do S.
schenckii, foram maiores em relação os obtidos na fase leveduriforme. Kohler et al. (2006)

57
avaliando a sensibilidade de isolados clínicos de casos de esporotricose humana e animal,
utilizando as duas fases do agente, resultaram em valores de CIMs maiores quando aplicado o
teste de sensibilidade com a fase filamentosa do S. schenckii. Segundo os autores, essa
diferença entre as fases filamentosa e leveduriforme frente as provas de suscetibilidade, geram
a necessidade de novos estudos, que estabeleçam critérios diferenciados para a realização das
provas in vitro para S. schenckii.
Considerando que a miltefosina é um fármaco que induz apoptose celular, acredita-se
que este fármaco possua ação fungicida e não apenas fungistática. Os resultados de CFMs
obtidos no presente estudo corroboram com esta especulação, uma vez que, os valores da
CFMs encontrados foram aproximados aos valores das CIMs em todos os isolados de H.
capsulatum var. capsulatum e de Sporothrix spp. analisados neste estudo. Ademais, segundo
Vila et al. (2013), um efeito fungicida é considerado significativo quando o valor de CFM foi
≤ 4 vezes os valores de CIM. Acima desse valor, o efeito antifúngico é considerado
fungistático. Em experimentos realizados por Vila et al. (2013), também relataram que a
miltefosina é capaz de reduzir a formação de biofilme e a viabilidade do biofilme maduro de
C. albicans. A atividade antibiofilme dos alquilfosfolípidos é uma descoberta importante uma
vez que estes compostos parecem ser ativos tanto em células planctônicas e em biofilme
(VILA et al.,2013).
In vivo, a atividade antifúngica da miltefosina foi confirmada por Widmer et al. (2006)
frente a espécie Cryptococcus neoformans, em modelo murino de criptococose disseminada.
Mais recentemente, Schubitz et al. (2011) e Wiederhold et al. (2013), demonstraram eficácia
limitada da miltefosina em modelos murinos de coccidioidomicose e criptococose
(meningoencefalite e infecção disseminada). Os autores afirmam que estes resultados
discrepantes podem estar associados a farmacocinética da droga e ao estado imune do
hospedeiro animal (SCHUBITZ et al., 2011; WIEDERHOLD et al., 2013). Na prática clínica,
Kesson et al. (2009) afirma que a miltefosina pode ser considerada uma terapia de resgate
para infecções fúngicas invasivas, para quais existem hoje poucos agentes efetivos. A terapia
antifúngica ótima para a infecção invasiva por Scedosporium prolificans ainda não foi
determinada. Em um relato de caso, a miltefosina em combinação com voriconazol e
terbinafina foi utilizada com sucesso para tratar uma criança que desenvolveu uma infecção
severa com S. prolificans osteomielite. Mesmo fazendo uso da terapia antifúngica com
caspofungina, terbinafina e voriconazol e terapia adjuvante com interferon-g, a infecção não
foi contida. Apenas após o início da terapia com a miltefosina, houve uma normalização

58
gradual dos níveis de marcadores inflamatórios no sangue e os resultados das culturas
cirúrgicas tornaram-se negativos. Embora a utilização de vários agentes terapêuticos torne
difícil atribuir a resposta terapêutica observada a miltefosina sozinha, a progressão da doença
foi invertida apenas após o seu início, e várias culturas de tecidos infectados tornaram-se
negativas no espaço de três semanas (KESSON et al., 2009).
Várias hipóteses potenciais para o mecanismo de ação da miltefosina surgiram nos
últimos anos, no entanto, nenhum mecanismo foi identificado definitivamente. Alvos
moleculares similares foram identificados para Leishmania e células de câncer humano, e tais
observações apontam que a atividade da miltefosina está essencialmente ligada a apoptose e a
perturbação de vias de sinalização celular dependentes de lipídeo (DORLO et al., 2012). Tem
sido proposto que a miltefosina usa jangadas lipídicas para interiorização em células
cancerosas para inibir a síntese de fosfocolina e proteína-quinase B (PKB), a qual é
supostamente suficiente para induzir a apoptose (MOREIRA et al., 2013; DORLO et al.,
2012). Segundo Moreira et al. (2013), a miltefosina penetra na membrana através do seu
componente lipídico e interage com as cadeias laterais hidrofóbicas na interface lipídeo-
proteína. Proteínas com maior hidrofobicidade podem fazer com que os grupos hidrófilos de
miltefosina penetrem na membrana e, assim, levar à ruptura da mesma.
Em estudos realizados por Zuo et al. (2011), foi demonstrado em Saccharomyces
cerevisiae que a miltefosina é absorvida rapidamente e penetra na membrana mitocondrial
interna, onde interrompe o potencial de membrana e faz com que ocorra a inibição dose
dependente de citocromo c oxidase (COX), levando a apoptose celular. Ainda em estudos
realizados por Zuo et al. (2011), dois marcadores intrínsecos de apoptose, como a ativação
caspase e fragmentação do DNA nuclear, foram procurados em células de S. cerevisiae
tratadas com miltefosina. A fragmentação do DNA e ativação de caspases celulares foi
evidente. Estas observações são sinais típicos de apoptose em leveduras (THEVISSEN et al.,
2008).
Em experimentos realizados por Khademvatan; Gharavi; Saki (2011) foram analisadas
as atividades dos genes metacaspases e proteína PARP em Leishmania infantum, onde
mostrou que a miltefosina cliva a protéina PARP (ADP-ribose) uma enzima de reparação do
DNA e ativa as caspases, um interuptor da função central para execução da apoptose. As
caspases induzem uma cascata de reações que conduzem finalmente a morte celular por
apoptose. Considerando as semelhanças estruturais e moleculares entre essas células, é

59
plausível supor que a ação da miltefosina em fungos pode estar essencialmente ligada a
indução de apoptose celular. Os experimentos realizados por Zuo et al. (2011) e
Khademvatan; Gharavi; Saki (2011) em que comprovam a fragmentação do DNA e ativação
de caspases celulares no tratamento com miltefosina são sinais típicos que este fármaco induz
a apoptose em fungos e parasitas Leishmania. Contudo, não se pode dizer com certeza se este
é o mecanismo real de ação da miltefosina, pois a multiplicidade de mecanismos potenciais
propostos podem indicar que miltefosina tem mais de um local de ação molecular (DORLO et
al., 2012).
Por fim, considerando as semelhanças clínicas entre a leishmaniose, e as micoses
histoplasmose e esporotricose, o parasitismo intracelular de seus agentes etiológicos
(WHEAT, 2003; CORDEIRO et al., 2011c; MIRANDA et al., 2011), a similaridade das
concentrações inibitórias mínimas da miltefosina in vitro ante Leishmania spp., H.
capsulatum var. capsulatum e Sporothrix spp. (MAIA et al., 2013), bem como, o sucesso no
tratamento de Leishmaniose cutânea, subcutânea e visceral (SUNDAR et al., 2002;
WIEDERHOLD et al., 2013), é possível inferir que essa droga representa uma perspectiva
interessante para subsidiar a terapia convencional atual de casos refratários ou recidivos de
infecções fúngicas causadas por estes patógenos. Vale salientar que, mais estudos são
necessários para avaliar a aplicabilidade da miltefosina in vivo e validar a sua utilização como
uma alternativa para o tratamento de histoplamose e esporotricose.

60
9 CONCLUSÕES
1. A miltefosina inibiu o crescimento in vitro de todas as cepas de H. capsulatum em ambas
as fases avaliadas. Valores de CIMs inferiores foram encontrados para miltefosina frente ao
H. capsulatum em fase de levedura.
2. A miltefosina inibiu o crescimento in vitro de todas as cepas de Sporothrix spp. avaliadas.
Valores de CIMs superiores foram observados frente S. globosa quando comparados com as
outras espécies do complexo S. schenckii.
3. Valores de CIMs e CFMs de Sporothrix spp. frente a miltefosina foram similares aos de H.
capsulatum na fase filamentosa e superiores aos de H. capsulatum na fase leveduriforme
4. Todos os valores de CFMs obtidos para miltefosina frente aos fungos H. capsulatum e
Sporothrix spp. foram ≤ 4 vezes os valores de CIMs encontrados, apontando uma ação
fungicida da droga.

61
REFERÊNCIAS
AFELTRA, J.; VERWEIJ, P. E. Antifungal activity of nonantifungal drugs. European
Journal Clinical Microbiology Infectious Diseases, v. 22, n. 7, p. 397-407, 2003.
AFELTRA, J; VITALE, RG; MOUTON, JW; VERWEIJ, PE. Potent synergistic in vitro
interaction between nonantimicrobial membrane-active compounds and itraconazole against
clinical isolates of Aspergillus fumigatus resistant to itraconazole. Antimicrobial Agents and
Chemotherapy, v. 48, n. 4, p. 1335-1343, 2004.
AGRESTA, M.; D’ARRIGO, P; FASOLI, E.; LOSI, D.; PEDROCCHI-FANTONI, G.;
RIVA, S.; SERVI, S.; AND TESSARO, D. Synthesis and antiproliferative activity of
alkylphosphocholines. Chemistry and physics of lipids, v. 126, n. 2, p. 201–210, 2003.
AIDÉ, M.A. Histoplasmosis. Jornal Brasileiro de Pneumologia, v. 35, n. 11, p. 1145-1151,
2009.
ALLTON, D.R.; RIVARD, R.G.; CONNOLLY, P.A.; McCALL, S.; DURKIN, M.M.;
BOYD, T.M; FLANAGAN, J.P.; WHEAT, L.J.; HOSPENTHAL, D.R. Detection of Latin
American strains of Histoplasma in a murine model by use of a commercially available
antigen test. Clinical and Vaccine Immunology, v. 17, n. 5, p. 802-806, 2010.
ALMEIDA-PAES, R.; FRASES, S.; MONTEIRO, P. C. F.; GUTIERREZ-GALHARDO, M.
C.; ZANCOPÉ-OLIVEIRA, R. M.; NOSANCHUK, J. D. Growth conditions influence
melanization of Brazilian clinical Sporothrix schenckii isolates. Microbes Infection, v. 11,n.
5 p. 554–562, 2009.
ALVARADO-RAMÍREZ, E., AND TORRES-RODRÍGUEZ, J. M.. In vitro susceptibility of
Sporothrix schenckii to six antifungal agents determined using three different methods.
Antimicrobial Agents and Chemotherapy v. 51, p. 2420–2423, 2007.
ALVES, S. H.; BOETTCHER, C. S.; OLIVEIRA, D. C.; TRONCO-ALVES, G. R.;
SGARIA, M. A.; THADEU, P.; OLIVEIRA, L. T.; SANTURIO, J. M. Sporothrix Schenckii
associated with armadillo hunting in Southern Brazil: epidemiological and antifungal
susceptibility profiles. Revista da Sociedade Brasileira de Medicina Tropical, v. 43, n. 5, p.
523–525, 2010.
ANDREU, C. M. F.; LEÓN, A. M.; MEDINA, Y. E.; MACHÍN, G. M.; LANCHA, M. R. P.;
ZARAGOZÍ, M. T. I. Sensibilidad in vitro de Histoplasma capsulatum var. capsulatum frente
a anfotericina B, ketoconazol, itraconazol y fluconazol. Revista Cubana de Medicina
Tropical, v. 55, n. 2, p. 76-82, 2003.
ARRILLAGA-MONCRIEFF, I; CAPILLA, J.; MAYAYO, E.; MARIMON, R.; MARINÉ,
M.; GENÉ, J.; CANO, J. AND GUARRO J. Different virulence levels of the species of
Sporothrix in a murine model. Clinical Microbiology Infection, v.15, n.7, p. 651–655, 2009.
BARROS, M. B. L.; PAES, R. A.; SCHUBACH, A. Sporothrix schenckii and Sporotrichosis.
Clinical Microbiology Reviews, v. 24, n. 4, p.633-654, 2011.

62
BARROS, M.B.L.; SCHUBACH, T.P.; COLL, J.O.; GREMIÃO, I. D.; WANKE, B.;
SCHUBACH, A. Esporotricose: a evolução e os desafios de uma epidemia. Revista
Panamericana de Salud Pública, v. 27, n. 6, p. 455–460, 2010.
BERGOLD, A. M.; GEORGIADIS, S. Novidades em fármacos antifúngicos: uma revisão.
Visão Acadêmica, v. 5, n. 2, p. 159-172, 2004.
BRILHANTE, R. S. N.; FECHINE, M. A. B.; CORDEIRO, R. A.; ROCHA, M. F. G.;
RIBEIRO, J. F.; MONTEIRO, A. J.; LIMA, R. A. C.; MESQUITA, J. R. L.; CAMARGO, Z.
P.; SIDRIM, J. J. C. In vitro effect of sulfamethoxazole-trimethoprim against Histoplasma
capsulatum var. capsulatum. Antimicrobial Agents Chemotherapy, v. 54, n. 9, p. 3978-
3979, 2010.
BRILHANTE, R.S.; FECHINE, M.A.; MESQUITA, J.R.L.; CORDEIRO, R.D.; ROCHA,
M.F.; MONTEIRO, A.J.; DE LIMA, R.A.; CAETANO, E.P.; PEREIRA, J.F.; CASTELO-
BRANCO, D.S.C.M.; DE CAMARGO, Z.P.; SIDRIM, J.J. Histoplasmosis in HIV- positive
patients in Ceará, Brazil: clinical-laboratory aspects and in vitro antifungal susceptibility of
Histoplasma capsulatum isolates. Transactions Royal Society of Tropical Medicine
Hygiene, v. 106, n. 8, p. 484–488, 2012.
BUSTAMANTE, B.; CAMPOS, P. E. Sporotrichosis: a forgotten disease in the drug rsearch.
Expert review of anti-infective therapy, v. 2, n. 1, p. 85-94, 2004.
CAILLOT, D.; THIÉBAUT, A.; HERBRECHT, R.; BOTTON, S.; PIGNEUX, A.;
BERNARD, F.; LARCHÉ, J.; MONCHECOURT, F.; ALFANDARI, S.; MAHI, L.
Liposomal amphotericin B in combination with caspofungin for invasive aspergillosis in
patients with hematologic malignancies: a randomized pilot study (Combistrat trial).
American Cancer Society, v. 110, n. 12, p. 2740-2746, 2007.
CANTÓN, E.; PEMÁN, J.; VIUDES, A.; QUINDÓS, G.; GOBERNADO, M.; ESPINEL-
INGROFF, A. Minimum fungicidal concentrations of amphotericin B for bloodstream
Candida species. Diagnnotic Microbiology and Infections Diasease, v. 45, n. 3, p. 203-206,
2003.
CARRADA, B. T. New observations on the epidemiology of sporotrichosis and Sporothrix
schenckii complex. Revista Latino-americana de Patologia Clínica, v. 59, n. 2, p. 88-100,
2012.
CASTRO, B.; GAMEIRO, P.; LIMA, J. L. F. C.; MATOS, C.; REIS, S. Interaction of drugs
with hexadecylphosfocoline micels. Derivative spectroscopy, acid-base and solubility studies.
Material Science and Engineering, v. 18, n. esp., p. 71-78, 2001.
CATALÁN, M.; MONTEJO, J. C. Antifúngicos sistémicos. Farmacodinamia y
farmacocinética. Revista Iberoamericana de Micologia, v. 23, p. 39-49, 2006.
CHANDRASEKAR, P. H.; SOBEL, J. D. Micafungin: A new echinocandin. Clinical
Infectious Diseases, v. 42, n. 8, p. 1171-1178, 2006.

63
CHANG, M.R.; TAIRA, C.L.; PANIAGO, A.M.M.; TAIRA, D.L.; CUNHA, R.V.; WANKE,
B. Study of 30 cases of histoplasmosis observed in Mato Grosso do Sul State, Brazil. Revista
Instituto de Medicina Tropical de São Paulo, v. 49, n. 1, p. 37-39, 2007.
CHAPMAN, S. W.; SULLIVAN, D. C.; CLEARY, J. D. In search of the holy grail of
antifungal therapy. Transactions of The American Clinical and Climatological
Association, v. 119, p. 197-216, 2008.
CHOU, T. Theoretical Basis, Experimental Design, and Computerized Simulation of
Synergism and Antagonism in Drug Combination Studies. Pharmacological Reviews, v. 58,
n. 3, p. 621-681, 2006.
CLINICAL AND LABORATORY STANDARDS INSTITUTE (CLSI.). Método de
referência para testes de diluição em caldo para determinação da sensibilidade de
leveduras à terapia antifúngica: norma aprovada – M27-A2. 2. ed. Pennsylvania, Estados
Unidos, 2002a.
CLINICAL AND LABORATORY STANDARDS INSTITUTE (CLSI). Método de
referência para testes de diluição em caldo para determinação da sensibilidade a terapia
antifúngica dos fungos filamentosos: norma aprovada – M38-A. Pennsylvania, Estados
Unidos, 2002b.
CLINICAL AND LABORATORY STANDARDS INSTITUTE (CLSI). Reference Method
for Broth Diluition Antifungical Suscetibility Testing of Yeasts: approved Standard. 3nd
ed. Wayne, PA, 2008a.
CLINICAL AND LABORATORY STANDARDS INSTITUTE (CLSI). Reference method
for broth dilution antifungal susceptibility testing of filamentous fungi. 2nd ed. Wayne,
PA, 2008b.
CONNOLLY, P.; WHEAT, J.; SCHNIZLEIN-BICK, C.; DURKIN, M.; KOHLER, S.;
SMEDEMA, M.; GOLDBERG, J.; BRIZENDINE, E.; LOEBENBERG, D. Comparison of a
89 new triazole antifungal agent, Schering 56592, with itraconazole and amphotericin B for
treatment of histoplasmosis in immunocompetent mice. Antimicrobial Agents and
Chemotherapy, v. 43, n. 2, p. 322-328, 1999.
CORDEIRO, R. A.; COELHO, C. G. V.; BRILHANTE, R. S. N.; SIDRIM, J. J.; CASTELO-
BRANCO, D. S.; MOURA, F. B.; ROCHA, M. F. Serological evidence of Histoplasma
capsulatum infection among dogs with leishmaniasis in Brazil. Acta Tropica, v. 119, n. 2-3,
p. 203-205, 2011c.
CORDEIRO, R. A.; MARQUES, F. J. F.; BRILHANTE, R. S. N.; SILVA, K. R. C.;
MOURÃO, C. I.; CAETANO, E. P.; FECHINE, M. A. B.; RIBEIRO, J. F.; CASTELO-
BRANCO, D. S. C. M; LIMA, R. A. C.; MESQUITA, J. R. L.; MONTEIRO, A. J.; ROCHA,
F. A. C.; ROCHA, M. F. G.; SIDRIM, J. J. C. Synergistic Effect of Antituberculosis Drugs
and Azoles In Vitro against Histoplasma capsulatum var. capsulatum. Antimicrobial Agents
and Chemotherapy. v. 55, n. 9, p. 4482-4484, 2011a.

64
CORREA, B.; GAMBALE, W.; PAULA, C. R.; PALAZZO, S. Morphogenesis of Sporothrix
schenckii “in vivo” and “in vitro” through the method of viability by fluorescence. Applied
Fluorescence Technology. v. 3, p.1-8, 1991.
COUPPIÉ, P.; AZNAR, C.; CARME, B.; NACHER, M. American histoplasmosis in
developing countries with a special focus on patients with HIV: diagnosis, treatment, and
prognosis. Current Opinion in Infectious Diseases, v. 19, n. 5, p. 443-449, 2006.
CROFT, S.L., BARRETT, M.P., URBINA, J.A. Chemotherapy of trypanosomiases and
leishmaniasis. Trends in Parasitology, v. 21, n. 11, p. 508-512, 2005.
CROFT, S.L.; SEIFERT, K.; DUCHERE, M. Antiprotozoal activities of phospholipid
analogues. Molecular Biochemical Parasitology, v. 126, n. 2, p. 165-172, 2003.
CRUZ, L. C. H. Micologia veterinária. Rio de Janeiro: Revinter; 2010.
CRUZ, L. C. H. Complexo Sporothrix schenckii. Revisão de parte da literatura e
considerações sobre o diagnóstico e a epidemiologia. Revista veterinária Zootecnia, p. 8-28,
2013.
CUENCA-ESTRELLA, M.; GOMEZ-LOPEZ, A.; MELLADO, E.; BUITRAGO, M. J.;
MONZON, A.; RODRIGUEZ-TUDELA, J. L. Head-to-head comparison of the activities of
currently available antifungal agents against 3,378 spanish clinical isolates of yeasts and
filamentous fungi. Antimicrobial Agents and Chemotherapy, v. 50, n. 3, p. 917-921, 2006.
DAHER, E.F.; BARROS, F.A.S.; DA SILVA JÚNIOR, G.B.; TAKEDA, C.F.V.; MOTA,
R.M.S.; FERREIRA, M.T.; MARTINS, J.C.; OLIVEIRA, S.A.J.; GUTIÉRREZ-
ADRIANZÉN, O. A. Risk factors for death in acquired immunodeficiency syndrome-
associated disseminated histoplasmosis. The American Journal of Tropical Medicine
Hygiene, v. 74, n. 4, p. 600-603, 2006.
DAHER, E.F.; SILVA Jr, G.B.; BARROS, F.A.S.; TAKEDA, C.F.V.; MOTA, R.M.S.;
FERREIRA, M.T.; OLIVEIRA, S.A.; MARTINS, J.C.; ARAÚJO, S.M.H.A, GUTIÉRREZ-
ADRIANZZÉN, O.A. Clinical and laboratory features of disseminated histoplasmosis in HIV
patients from Brazil. Tropical Medicine Internacional Health, v. 12, n. 9, p. 1108-1115,
2007.
DE ARAUJO, T.; MARQUES, A. C.; KERDEL, F. Sporotrichosis. International Journal
Dermatology, v. 40, n. 12, p. 737-742, 2001.
DE CASTRO, S. L.; SANTA-RITA, R. M.; URBINA, J. A.; CROFT, S. L. Antiprotozoal
lysophospholipid analogues: a comparison of their activity against trypanosomatid parasites
and tumor cells. Mini Reviews Medicinal Chemistry, v. 4, p. 141–151, 2004.
DE MEYER, E. M.; DE BEER, Z. W.; SUMMERBELL, R. C.; MOHARRAM, A. M.; DE
HOOG, G. S.; VISMER, H. F.; WINGFIELD, M. J. Taxonomy and phylogeny of new wood-
and soil-inhabiting Sporothrixspecies in the Ophiostoma stenoceras-Sporothrix schenckii
complex. Mycologia, v.100, n. 4, p. 647–661, 2008.

65
DEUS FILHO, A.; BODO, W.; CAVALCANTI, S.; AMPARO, M.; SOARES MARTINS,
L.M.; CASTELO BRANCO, A.D. Histoplasmose no Nordeste do Brasil. Relato de três casos.
Revista Portuguesa de Pneumologia, v. 15, n. 1, p. 109-114, 2009.
DORLO, T. P. C.; VAN THIEL, P. P.; HUITEMA, A. D.; KEIZER, R. J.; DE VRIES, H. J.;
BEIJNEN, J. H.; DE VRIES, P. J. Pharmacokinetics of miltefosine in Old World cutaneous
leishmaniasis patients. Antimicrobial Agentes Chemotherapy, v. 52, n. 8, p. 2855-2860,
2008.
DORLO, T. P. C.; BALASEGARAM, M.; BEIJNEN, J. H.; VRIES, P. J. Miltefosine: a
review of its pharmacology and therapeutic efficacy in the treatment of leishmaniasis.
Journal of Antimicrobial Chemotherapy, v. 67, n. 11, p. 2576 – 2597, 2012.
DUIN, D. V.; CASADEVALL, A.; NOSANCHUK, J. D. Melanization of Cryptococcus
neoformans and Histoplasma capsulatum Reduces Their Susceptibilities to Amphotericin B
and Caspofungin. Antimicrobial Agents and Chemotherapy, v. 46, n.11, p. 3394-3400,
2002.
ESPINEL-INGROFF, A. Novel antifungal agents, targets or therapeutic strategies for the
treatment of invasive fungal diseases: a review of the literature (2005-2009). Revista
Iberoamericana de Micología, v. 26, n. 1, p. 15-22, 2009.
FERNANDES, K. S. S.; HERBERT L.; MATHEWS, H. L.; BEZERRA, L. M. L. Differences
in virulence of Sporothrix schenckii conidia related to culture conditions and cell-wall
components. Journal of Medical Microbiology, v. 48, n. 2, p. 195-203, 1999.
FERREIRA, M.S.; BORGES, A. S. Histoplasmosis. Revista da Sociedade Brasileira de
Medicina Tropical, v. 42, n. 2, p. 192-198, 2009.
FIANCHI, L.; PAGANO, L.; CAIRA, M.; NOSARI, A.; VAN LINT, M. T.; CANDONI, A.;
OFFIDANI, M.; ALOISI, T.; IRRERA, G.; BONINI, A.; PICARDI, M.; CARAMATTI, C.;
INVERNIZZI, R.; MATTEI, D.; MELILLO, L.; WAURE, C.; REDDICONTO, G. Fungal
infections in recipients of hematopoietic stem cell transplants: results of the SEIFEM B-2004
study - sorveglianza epidemiologica infezioni fungine nelle emopatie maligne. Clinical
Infectious Diseases, v. 45, n. 9, p. 1161-1170, 2007.
FILGUEIRA, K. D. Esporotricose na espécie canina: relato de um caso na cidade de Mossoró,
RN. Ciência Animal Brasileira, v. 10, n. 2, p. 673-677, 2009.
FILIPPIN, F. B.; SOUZA, L. C. Eficiência terapêutica das formulações lipídicas de
Anfotericina B. Revista Brasileira de Ciências Farmacêuticas, v. 42, n. 2, p. 167-194,
2006.
FRANCESCONI, G.; FRANCESCONI DO VALLE, A.C.; PASSOS, S.L.; DE LIMA
BARROS, M. B. L.; ALMEIDA-PAES, R.; CURI, A. L.; LIPORAGE, J.; PORTO, C. F.;
GALHARDO, M.C. Comparative study of 250 mg/day terbinafine and 100 mg/day
itraconazole for the treatment of cutaneous sporotrichosis. Mycopathologia, v. 171, n. 5, p.
349-354, 2011.

66
GOBERNADO, M.; CANTÓN, E. Anidulafungina. Revista Española de Quimioterapia, v.
21, n. 2, p. 99-114, 2008.
GOMES, A. R; MADRID, I. M.; MATOS, C. B.; TELLES, A. J.; WALLER, S. B.; NOBRE,
M. O.; MEIRELES, M. C. A. Dermatopatias Fúngicas: aspectos clínicos, diagnósticos e
terapêuticos. Acta Veterinaria Brasilica, v. 6, n. 4, p. 272-284, 2012.
GONZÁLEZ, G. M. In vitro activities of isavuconazole against opportunistic filamentous and
dimorphic fungi. Medical Mycology, v. 47, n. 1, p. 71-76, 2009.
GREMIÃO, I. D. F.; SCHUBACH, T. M. P.; PEREIRA, S. A.; RODRIGUES, A. M.;
CHAVES, A. R.; BARROS, M. B. L. Intralesional amphotericin B in a cat with refractory
localised sporotrichosis. Journal of Feline Medicine and Surgery, v. 11, n. 8, p. 720–723,
2009.
GUARRO, J.; GENÉ, J.; STCHIGEL, A. M. Developments in fungal taxonomy. Clinical
Microbiology Reviews, v. 12, n. 3, p. 454–500, 1999.
GUIMARÃES, A. J.; NOSANCHUK, J. D.; ZANCOPÉ-OLIVEIRA, R. M. Diagnosis of
Histoplasmosis. Brazilian Journal of Microbiology, v. 37, n. 1, p. 1-13, 2006.
GUTIERREZ-GALHARDO, M. C.; BARROS, M. B.; SCHUBACH, A. O.; CUZZI, T.;
SCHUBACH, T. M.; LAZÉRA, M. S.; VALLE, A.C. Erythema multiforme associated with
sporotrichosis. Journal of the European Academy of Dermatology Venereology, v. 19, n.
4, p. 507–509, 2005.
GUTIERREZ-GALHARDO, M. C.; SCHUBACH, A. DE O.; BARROS, M. B. L.;
BLANCO, T. C. M.; CUZZI-MAYA, T.; SCHUBACH, T. M. P.; LAZÉRA, M. S.; DO
VALLE, A. C. F. Erythema nodosum associated with sporotrichosis. International Journal
Dermatology, v. 41, n. 2, p. 114–116, 2002.
HOLBROOK, E. D.; RAPPLEYE, C. A. Histoplasma capsulatum pathogenesis: making a
lifestyle switch. Current Opinion in Microbiology, v. 11, n. 4, p. 318-324, 2008.
HOSPENTHAL, D. R.; MURRAY, C. K.; RINALDI, M. G. The role of antifungal
susceptibility testing in the therapy of candidiasis. Diagnostic Microbiology and Infectious
Disease, v. 48, n. 3, p. 153-160, 2004.
HUANG, F.; QU, F.; PENG, Q.; XIA, Y.; PENG, L. Synthesis and characterization of
photolabeling probes of miltefosine. Journal of Fluorine Chemistry, v. 126, n. 5, p. 739-
743, 2005.
HUANG, W.; ZHANG, Z.; HAN, X.; TANG, J.; WANG, J.; DONG, S.; WANG, E. Ion
channel behavior of amphotericin B in sterol-free and cholesterol or ergosterol-containing
supported phosphatidylcholine bilayer model membranes investigated by electrochemistry
and spectroscopy. Biophysical Journal, v. 83, n. 6, p. 3245-3255, 2002.
KAMINSKY, R. Miltefosine zentaris. Current Opinion Investigation Drugs, v. 3, n. 4, p.
550-554, 2002.

67
KASUGA, T.; WHITE, T. J.; KOENIG, G.; MCEWEN, J.; RESTREPO, A.; CASTAÑEDA,
E.; LACAZ, C. S.; HEINS-VACCARI, E. M.; FREITAS, R. S.; ZANCOPÉ-OLIVEIRA, R.
M.; QIN, Z.; NEGRONI, R.; CARTER, D. A.; MIKAMI, Y.; TAMURA, M.; TAYLOR, M.
L.; MILLER, G. F.; POONWAN, N.; TAYLOR, J. W. Phylogeography of the fungal
pathogen Histoplasma capsulatum. Molecular Ecology, v. 12, n. 12, p. 3383-3401, 2003.
KAUFFMAN, C. A. Endemic Mycoses: Blastomycosis, Histoplasmosis, and Sporotrichosis.
Infectious Disease Clinics of North America, v. 20, n. 3, p. 645-662, 2006.
KAUFFMAN, C. A. Histoplasmosis. Clinics in Chest Medicine, v. 30, n. 2, p. 217-225,
2009.
KAUFFMAN, C. A.; BUSTAMANTE, B.; CHAPMAN, S. W.; PAPPAS, P. G. Clinical
Practice Guidelines for the Management of Sporotrichosis: 2007 Update by the Infectious
Diseases Society of America. Clinical Infectious Diseases, v. 45, n. 10, p. 1255–1265,
2007a.
KAUFFMAN, C.A. Histoplasmosis: a clinical and laboratory update. Clinical Microbiology
Reviews, v. 20, n. 1, p. 115-132, 2007b.
KESSON, A. M.; BELLEMORE, M. C.; O’MARA, T. J.; ELLIS, D. H.; SORRELL, T. C.
Scedosporium prolificans osteomyelitis in an immunocompetent child treated with a novel
agent, hexadecylphosphocholine (miltefosine), in combination with terbinafine and
voriconazole: a case report. Clinical Infectious Diseases, v. 48, n. 9, p. 1257 – 1261, 2009.
KHADEMVATAN, S.; GHARAVI, M. J.; SAKI, J. Miltefosine induces metacaspase and
PARP genes expression in Leishmania infantum. The Brazilian Journal of infectious
Diseases, v. 15, n. 5, p. 442-448, 2011.
KLEIN, B. S.; TEBBETS, B. Dimorphism and virulence in fungi. Current opinion in
microbiology, v. 10, n. 4, p. 314-319, 2007.
KOHLER, L. M.; HAMDAN, J. S.; FERRARI, T. C. Successful treatment of a disseminated
Sporothrix schenckii infection and in vitro analysis for antifungal susceptibility testing.
Diagnostic Microbiology Infectious Disease, v. 58, n. 1, p. 117–120, 2007.
KOHLER, L. M.; SOARES, B. M.; DE ASSIS SANTOS, D.; DA SILVA BARROS, M. E.;
HAMDAN, J. S. In vitro susceptibilities isolates of sporothrix schenckii to amfotericin B,
itraconazole, and terbinafine: comparisionof yeast and mycelial forms. Clinical Microbiology
Reviews, v. 52, n. 9, p. 843 – 847, 2006.
LACAZ, C. S.; PORTO, E.; MARTINS, J. E. C. M. Esporotricose e outras micoses gomosas.
In: ______. Micologia médica. São Paulo: Savier, 1991. p. 233-247.
LACAZ, C. S.; PORTO, E.; MARTINS, J. E. C.; HEINS-VACCARI, E. M.; MELO, N. T.
Lacaz: tratado de micologia médica Lacaz. 9. ed. São Paulo: Sarvier, 2002.

68
LARSSON, C. E. Sporotrichosis. Brazilian journal of Veterinary Reserach Animal
Science, v. 48, n. 3, p. 250-259, 2011.
LARSSON, C. E. Sporotrichosis. In: CONGRESS OF THE WORLD SMALL ANIMAL
VETERINARY ASSOCIATION, 27., 2002, Barcelona. Proceedings... Barcelona, 2002. p.
127-128.
LOPES-BEZERRA, L.M.; SCHUBACH, A.; COSTA, R. O. Sporothrix schenckii and
Sporotrichosis. Anais da Academia Brasileira de Ciências, v. 78, n. 2, p. 293-308, 2006.
LUNA KP, HERNÁNDEZ IP, RUEDA CM, ZORRO MM, CROFT SL, ESCOBAR P. In
vitro susceptibility of Trypanosoma cruzi strains from Santander, Colombia, to
hexadecylphosphocholine (miltefosine), nifurtimox and benznidazole. Biomédica, v. 29, n. 3,
p. 448-455, 2009.
MACDONALD, P. M.; RYDALL, J. R.; KUEBLER, S. C. Synthesis and characterization of
a homologus series of zwitterionic surfactants based on phosphocoline. Langmuir, v. 7, n.
11, p. 2602-2606, 1991.
MADRID, I. M.; MATTEI, A. S.; FERNANDES, C. G.; NOBRE, M. O.; MEIRELES, M. C.
A. Epidemiological Findings and Laboratory Evaluation of Sporotrichosis: A Description of
103 Cases in Cats and Dogs in Southern Brazil. Mycopathologia, v. 173, n. 4, p. 265-273,
2011b.
MADRID, I. M.; SANTIN, R.; SCHUCH, L. F. D.; GOMES, A. R.; NOBRE, M. O.;
MEIRELES, M. C. A. Morphological pattern and thermotolerance of clinical and from
environment isolates of Sporothrix schenckii from the south of Brazil. Revista Brasileira de
Ciência Veterinária, v. 18, n. 2/3, p. 57-61, 2011a.
MADRID, I. M.; XAVIER, M. O.; MATTEI, A. S.; FERNANDES, C. G.; GUIM, T. N.;
SANTIN, R.; SCHUCH, L. F. D.; NOBRE, M. O.; MEIRELES, M. C. A. Role of melanin in
the pathogenesis of cutaneous sporotrichosis. Microbes and Infection, v. 12, n. 2, p.162-165,
2010.
MADRID, I. M.; XAVIER, M. O.; MATTEI, A.; CARAPETO, L. P.; ANTUNES, T. A.;
SANTOS, J. R. R.; NOBRE, M. O.; MEIRELES, M. C. A. Esporotricose óssea e cutânea em
canino. Brazilian journal of Veterinary Reserach Animal Science, v. 44, n. 6, p. 441-443,
2007.
MAIA, C.; NUNES, M.; MARQUES, M.; HENRIQUES, S.; ROLÃO, N.; CAMPINO, L. In
vitro drug susceptibility of Leishmania infantum isolated from humans and dogs.
Experimental Parasitolology, v. 135, n. 1, p. 36 – 41, 2013.
MARCO, C.; JIMENEZ-LOPEZ, J. M.; RIOS-MARCO, P. Hexadecylphosphocholine alters
nonvesicular cholesterol traffic from the plasma membrane to the endoplasmic reliculum and
inhibits the synthesis of sphingomyelin in HepG2 cells. International Journal of
Biochemistry Cell Biology, v. 41, n. 6, p. 1296-1303, 2009.

69
MARIMON, R.; CANO J.; GENÉ J.; SUTTON, D. A.; KAWASAKI, M.; GUARRO, J.
Sporothrix brasiliensis, S. globosa, and S. mexicana, three new Sporothrix species of clinical
interest. Journal of Clinical Microbiology, v. 45, n. 10, p. 3198-3206, 2007.
MARIMON, R. J.; GENÉ, J.; CANO, L.; TRILLES, L.; Dos SANTOS LAZERA, M.;
GUARRO, J. Molecular phylogeny of Sporothrix schenckii. Journal of Clinical
Microbiology, v. 44, n. 9, p. 3251–3256, 2006.
MARIMON, R.; SERENA, C.; GENÉ, J.; CANO, J.; GUARRO, J. Susceptibilities of Five
Species of Sporothrix. Antimicrobial Agents and Chemotherapy. v. 52, n. 2, p. 732–734,
2008.
MARQUES-EVANGELISTA, O. M.; ALMEIDA-PAES, R.; DE MEDEIROS-MUÑIZ, M.;
DE LIMA-BARROS, B. M.; GUTIERREZ-GALHARDO, C. M.; ZANCOPE-OLIVEIRA, R.
M. Sporotrichosis Caused by Sporothrix globosa in Rio de Janeiro Brazil: Case report.
Mycopathologia, v. 169, n. 5, p. 359-363, 2010.
MARTINEZ, R. Atualização no uso de agentes antifúngicos. Jornal Brasileiro de
Pneumologia, v. 32, n. 5, p. 449-460, 2006.
MEINERZ, A. R. M.; NASCENTE, P. S.; SCHUCH, L. F. D.; CLEFF, M. B.; SANTIN, R.;
BRUM, C. S.; NOBRE, M. O.; MEIRELES, M. C. A.; MELLO, J. R. B. In vitro
susceptibility of isolates of Sporothrix schenckii to terbinafine and itraconazole. Revista da
Sociedade Brasileira de Medicina Tropical, v.40, n. 1, p. 60–62, 2007.
MELLO, C. X.; SCHUBACH, A. O.; MADEIRA, M. F. Can yeast-like form of Sporothrix
schenckii confuse the direct parasitological diagnosis of American cutaneous leishmaniasis?
Revista da Sociedade Brasileira de Medicina Tropical, v. 44, n. 1, p. 127, 2011.
MIRANDA, L. H. M.; QUINTELLA, L. P.; MENEZES, R. C.; DOS SANTOS, I. B.;
OLIVEIRA, R. V.; FIGUEIREDO, F .B.; LOPES-BEZERRA, L. M.; SCHUBACH, T. M.
Evaluation of immunohistochemistry for the diagnosis of sporotrichosis in dogs. Veterinary
Journal, v. 190, n. 3, p. 408-411, 2011.
MISHRA, J.; SAXENA, A.; SINGH, S. Chemotherapy of leishmaniasis: past, present and
future. Current Medicinal Chemistry, v. 14, n. 10, p. 1153-1169, 2007
MOREIRA, R. A.; MENDANHA, S. A.; HANSEN, D.; ALONSO, A. Interaction of
Miltefosine with the Lipid and Protein Components of the Erythrocyte Membrane Journal of
Pharmaceutical Sciences, v. 102, n. 5, p. 1661-1669, 2013.
MORRIS-JONES, R.; YOUNGCHIM, S.; GOMES, B. L.; AISEN, P.; HAY, R. J.;
NOSANCHUK, J. D.; CASADEVALL, A.; HAMILTON, A. J. Synthesis of melanin-like
pigments by Sporothrix schenckii in vitro and during mammalian infection. Infection
Immunity, v. 71, n. 7, p. 4026-4033, 2003.
MORRIS-JONES, R. Sporotrichosis. Clinical Experimental Dermatology, v. 27, n. 6, p.
427–431, 2002.

70
MUKHERJEE, A.; TANGRI, R.; VERMA, N.; GAUTAM, D. Chronic disseminated
histoplasmosis bone marrow involvement in an immunocompetent patient. Indian Journal
of Hematology and Blood Transfusion, v. 26, n. 2, p. 65-67, 2010.
NAKAI, T.; UNO, J.; IKEDA, F.; TAWARA, S.; NISHIMURA, K.; MIYAJI. M. In vitro
antifungal activity of Micafungin (FK463) against dimorphic fungi: comparison of yeast-like
and mycelial forms. Antimicrobial Agents Chemotherapy, v. 47, n. 4, p. 1376-1381, 2003.
NASCIMENTO, R. C.; ALMEIDA, S. R. Humoral immune response against soluble and
fractionate antigens in experimental sporotrichosis. FEMS Immunology Medical
Microbiology, v. 43, n. 2, p. 241-247, 2005.
NOBRE, M. O.; POTTER-DE-CASTRO, A.; CAETANO, D.; LEONARDO-DE-SOUZA, L.;
ARAUJO-MEIRELES, M. C. Recurrence of sporotrichosis in cats with zoonotic involvement.
Revista Iberoamericana de Micologia, v. 18, n. 3, p. 137-140, 2001.
NOBRE, M.O.; MADRID, I. M.; ANTUNES , T. A.; MARTINS, A. A.; FERNANDES, C.
G.; MATTEI, A. S.; SPANAMBERG, A.; MEIRELES, M. C. A.; FERREIRO, L. Virulence
differences between albino and pigmented Sporothrix schenckii isolates. Journal of
Medical Mycology, v. 18, n. 4, p. 191-197, 2008.
NORMAN, F. F.; MARTÍN-DÁVILA, P.; FORTÚN, J.; DRONDA, F.; QUEREDA, C.;
SÁNCHEZ-SOUSA, A.; LÓPEZ-VÉLEZ, R. Imported histoplasmosis: two distinct profiles
in travelers and immigrants. Journal of Travel Mediocine, v. 16, n. 4, p. 258-262, 2009.
NOSANCHUK, J. D.; GRACSER, A. Histoplasma capsulatum at the host-pathogen
interface. Microbes and Infection, v. 10, n. 9, p. 973-977, 2008.
NOSANCHUK, J. D.; GOMEZ, B. L.; YOUNGCHIM, S.; DIEZ, S.; AISEN, P.;
ZANCOPE-OLIVEIRA, R. M.; RESTREPO, A.; CASADEVALL, A.; HAMILTON, A. J.
Histoplasma capsulatum synthesizes melanin-like pigments in vitro and during mammalian
infection. Infection and Immunity, v. 70, n. 9, p. 5124–5131, 2002.
NUNES, G. D. L.; CARNEIRO, R. S. do.; FILGUEIRA, K. D.; FILGUEIRA, F. G. F.;
FERNANDES, T. H. T. Esporotricose felina no município de Itaporanga, Estado da Paraíba,
Brasil: relato de um caso. Arquivo Ciência Veterinária Zoologia Unipar, Umuarama, v. 14,
n. 2, p. 157-161, 2011.
O’SHAUGHNESSY, E. M.; MELETIADIS, J.; STERGIOPOULOU, T.; DEMCHOK, J. P.;
WALSH, T. J. Antifungal interactions within the triple combination of amphotericin B,
caspofungin and voriconazole against Aspergillus species. Journal of Antimicrobial
Chemoterapy, v. 58, n. 6, p. 1168-1176, 2006.
OBANDO, D.; WIDMER, F.; WRIGHT, L. C.; SORRELL, T. C.; JOLLIFFE, K. A.
Synthesis, antifungal and antimicrobial activity of alkylphospholipids. Bioorganic &
Medicinal Chemistry, v. 15, n. 15, p. 5158–5165, 2007.

71
ODDS, F. G.; GERVEN, F. V.; ESPINEL-INGROFF, A.; BARTLETT, M. S.;
GHANNOUM, M. A. Evaluation of possible correlations between antifungal susceptibilities
of filamentous fungi in vitro and antifungal treatment outcomes in animal infection models.
Antimicrobial Agents and Chemotherapy, v. 42, n. 2, p. 282-288, 1998.
PASQUALOTTO, AC; DENNING, DW. New and emerging treatments for fungal infections.
The journal of Antimicrobial Chemotherapy, v. 61, n. 1, p. i19–i30, 2008.
PETRIKKOS, G.; SKIADA, A. Recent advances in antifungal chemotherapy. International
Journal of Antimicrobial Agents, v. 30, n. 2, p. 108-117, 2007.
QUIST, E. M.; BELCHER, C.; LEVINE, G.; JOHNSON, M.; HEATLEY, J.J.; KIUPEL, M.;
GIRI,D. Disseminated histoplasmosis with concurrent oral candidiasis in na Eclectus parrot
(Eclectus roratus). Avian Pathology, v. 40, n.2, p. 207-211, 2011.
RAMOS-E-SILVA, M.; VASCONCELOS, C.; CARNEIRO, S.; CESTARI, T.
Sporotrichosis. Clinics in Dermatology, v. 25, n. 2, p. 181–187, 2007.
REIS, R. S.; ALMEIDA-PAES, R.; MUNIZ, M. M.; TAVARES, P. M.; MONTEIRO, P. C.;
SCHUBACH, T. M.; GUTIERREZ-GALHARDO, M. C.; ZANCOPÉ-OLIVEIRA, R. M.
Molecular characterisation of Sporothrix schenckii isolates from humans and cats involved in
the sporotrichosis epidemic in Rio de Janeiro, Brazil. Memórias do Instituto Oswaldo Cruz.
v. 104, n. 5, p. 769-774, 2009.
REX, J. H.; PFALLER, M. A.; WALSH, T. J.; CHATURVEDI, V.; ESPINEL-INGROFF,
A.; GHANNOUM, M. A.; GOSEY, L. L.; ODDS, F. C.; RINALDI, M. G.; SHEEHAN, D. J.;
WARNOCK, D. W. Antifungal Susceptibility Testing: Practical Aspects and Current
Challenges. Clinical Microbiology Reviews, v. 14, n. 4, p. 643-658, 2001.
RODRIGUES, A. M.; DE HOOG; S.; DE CAMARGO, Z. P. Emergence of patogenicity in
the Sporothrix schenckii complex. Medical Mycology, v. 51, n. 4, p. 405-412, 2012.
ROMERO-MARTINEZ, R.; WHEELER, M.; GUERRERO-PLATA, A.; RICO, G.;
TORRES-GUERRERO, H. Biosynthesis and functions of melanin in Sporothrix schenckii.
Infection and Immunity, v. 68, n. 6, p. 3696–3703, 2000.
ROSSINI, F.T.; GOULART, L.S. Classic histoplasmosis: Review. Revista Brasileira de
Análise Clínicas, v. 38, n. 4, p. 275-279, 2006.
SCHUBACH T.M.P.; FIGUEIREDO F.B.; OKAMOTO T.; BARBIERE I.; PEREIRA A.S.;
CUZZI-MAYA T.; ANDRADE M.V.; MADEIRA M.F.; LEAL C.A.; SILVA R.M.M.;
SCHUBACH A.O. Leishmaniose tegumentar americana em gato doméstico (Felis catus)
naturalmente infectado no Rio de Janeiro – Relato de isolamento de Leishmania sp. Revista
da Sociedade Brasileira de Medicina Tropical, v. 36, n. 1, p.342, 2003b.
SCHUBACH, A.; BARROS, M. B. L.; SCHUBACH, T. M.; FRANCESCONI-DO-VALLE,
A. C.; GUTIERREZ-GALHARDO, M. C.; SUED, M.; DE MATOS SALGUEIRO, M.;
FIALHO-MONTEIRO, P. C.; REIS, R. S.; MARZOCHI, K. B.; WANKE, B.;
CONCEIÇÃO,-SILVA, F. Primary conjunctival sporotrichosis: two cases from a zoonotic
epidemic in Rio de Janeiro, Brazil. Cornea, v. 24, n. 4, p. 491–493, 2005.

72
SCHUBACH, T. M. P.; SCHUBACH, A. O.; OKAMOTO, T.; BARROS, M. B. L.;
FIGUEIREDO, F. B.; CUZZI, T.; FIALHO-MONTEIRO, P. C.; REIS, R. S.; PEREZ, M. A.;
WANKE, B. Evaluation of an epidemic of sporotrichosis in cats: 347 cases (1998–2001).
Journal of the American Veterinary Medical Association, v. 224, n. 10, p. 1623–1629,
2004.
SCHUBACH, T. M.; SCHUBACH, A.; OKAMOTO, T.; PELLON, I. V.; FIALHO-
MONTEIRO, P. C.; REIS, R. S.; BARROS, M. B.; ANDRADE-PEREZ, M.; WANKE, B.
Haematogenous spread of Sporothrix schenckii in cats with naturally acquired sporotrichosis.
Journal of Small Animal Practice, v.44, n. 9, p. 395-398, 2003a.
SEGURO, F. S.; SILVEIRA, P. A. A. Histoplasmose. Einstein, v. 6, n. 3, p. 378-379, 2008.
SHUBITZ, L. F.; NIX, D. E.; MARTINEZ, M.; CHEN, J. N.; GALGIANI, J. N.; STEVENS,
D. A. Miltefosine (MF) in vitro and in vivo against Coccidioides. In: ANNUAL
COCCIDIOIDOMYCOSIS STUDY GROUP, 55th
, 2011, Davis, California. Proceedings...
Davis, California, 2011.
SIDRIM, J.J.C.; ROCHA, M.F.G. Micologia médica à luz de autores contemporâneos. Rio
de Janeiro: Guanabara-Koogan, 2004.
SILVA, D. T.; MENEZES, R. C.; GREMIÃO, I. D. F.; SCHUBACH, T. M. P.; BOECHAT,
J. S.; PEREIRA, S. A. Esporotricose zoonótica: procedimentos de biossegurança. Acta
Scientiae Veterinariae, v. 40, n. 4, p. 1067, 2012.
SILVA-VERGARA, M. L.; MARTINEZ, R.; BORGES-MALTA, M. E.; LEITE-MAFFEI,
C. M.; RAMIREZ, L. E. Histoplasma capsulatum isolated from Didelphis albiventris
(Marsupialia: Didelphidae) in the State of Minas Gerais, Brazil. Revista Iberoamericana de
Micologia, v. 18, n. 4, p. 180-182, 2001.
SILVEIRA, C. P.; TORRES-RODRÍGUEZ, J. M.; ALVARADO-RAMÍREZ, E.;
MURCIANO-GONZALO, F.; DOLANDE, M.; PANIZO, M.; REVIAKINA, V. MICs and
minimum fungicidal concentrations of amphotericin B, itraconazole, posaconazole and
terbinafine in Sporothrix schenckii. Journal of Medical Microbiology, v. 58, n. 12, p. 1607–
1610, 2009.
SOTO, J.; SOTO, P. Miltefosine: oral treatment of leishmaniasis. Expert Review of Anti-
Infective Terapy, v. 4, n. 2, p. 177-185, 2006.
SOUZA, N.T.; NASCIMENTO, A.C.B.M.; SOUZA, J.O.T.; SANTOS, F.C.G.C.A.;
CASTRO, R.B. Esporotricose canina: relato de caso. Arquvo Brasileiro de Medicina
Veterinária e Zootenia, v.61, n. 3, p. 572-576, 2009.
SOUZA, A. I.; BARROS, E. M. S.; ISHIKAWA, E.; ILHA, I. M. N.; MARIN, G. R. B.;
NUNES, V. L. B. Feline leishmaniasis due to Leishmania (Leishmania) amazonensis in Mato
Grosso do Sul State, Brazil. Veterinary Parasitology, v.128, n. 1/2, p. 41-45, 2005.
STALKUP, J. R., K. BELL, AND T. ROSEN. Disseminated cutaneous sporotrichosis treated
with itraconazole. Cutis, v. 69, n. 5, p. 371–374, 2002.

73
STOPPA, M. A.; CASEMIRO, L. A.; VINHOLIS, A. H. C.; CUNHA, W. R.; SILVA, M. L.
A.; MARTINS, C. H. G. Estudo comparativo entre as metodologias preconizadas pelo CLSI e
pelo EUCAST para avaliação da atividade antifúngica. Química Nova, v. 32, n. 2, p. 498-
502, 2009.
SUNDAR, S.; JHA, T. K.; THAKUR, C. P.; ENGEL, J.; SIDERMANN, H.; FISCHER, C.;
JUNGE, K.; BRYCESON, A.; BERMAN, J. Oral miltefosine for Indian visceral
leishmaniasis.The New England Journal of Medicine, v. 347, n. 22, p. 1739–1746, 2002.
THEVISSEN, K.; MADEO, F.; LUDOVICO, P.; CAMMUE, B.; WINDERICKX, J. Joined
in death: highlights of the Sixth International Meeting on Yeast Apoptosis in Leuven,
Belgium, 30 April-4 May 2008. Yeast, v. 25, n. 12, p. 927–934, 2008.
THOMPSON, G. R.; CADENA, J.; PATTERSON, T. F. Overview of Antifungal Agents.
Clinics in Chest Medicine, v. 30, n. 2, p. 203-215, 2009.
TLOUGAN, B. E.; PODJASEK, J. O.; PATEL, S. P.; NGUYEN, X. H.; HANSEN, R. C.
Neonatal sporotrichosis. Pediatric Dermatology, v. 26, n. 5, p. 563–565, 2009
TONG, Z., WIDMER, F.; SORRELL, T. C.; GUSE, Z.; JOLLIFFE, K. A.; HALLIDAY, C.;
LEE, O. C.; KONG, F.; WRIGHT, L. C.; SHARON, C. A.; CHEN, S. C. A. In vitro activities
of miltefosine and two novel antifungal biscationic salts against a panel of 77 dermatophytes.
Antimicrobiology Agents Chemotherapy, v. 51, n. 6, p. 2219-2222, 2007.
TRILLES, L.; FERNÁNDEZ-TORRES, B.; LAZÉRA, M. S.; WANKE, B.; SCHUBACH, A.
O.; ALMEIDA PAES, R.; ISABEL INZA, I.; GUARRO, J. In Vitro Antifungal
Susceptibilities of Sporothrix schenckii in Two Growth Phases. Antimicrobial Agents and
Chemotherapy, v. 49, n. 9, p. 3952–3954, 2005.
VENTURINI, T.P.; ROSSATO, L.; SPADER, T.B.; TRONCO-ALVES, G.R.; AZEVEDO,
M.I.; WEILER, C.B.; SANTURIO, J.M.; ALVES, S.H. In vitro synergisms obtained by
amphotericin B and voriconazole associated with non-antifungal agents against Fusarium spp.
Diagnostic Microbiology and Infectious Disease, v. 71, n. 2, p. 126–130, 2011.
VILA, T. V. M.; ISHIDA, K.; DE SOUZA, W.; PROUSIS, K.; CALOGEROPOULOU, T.;
ROZENTAL, S. Effect of alkylphospholipids on Candida albicans biofilm formation and
maturation. Journal Antimicrobial Chemotherapy, v. 68, n. 1, p.113 – 125, 2013
WANKE, B; LAZÉRA, M. Histoplasmose clássica e africana. In: SIDRIM, J. J. C.; ROCHA,
M. F. G. Micologia médica à luz de autores contemporâneos. Rio de janeiro: Guanabara
Koogan, 2004. p. 222-251.
WEBSTER, R. H.; SIL, A. Conserved factors Ryp2 and Ryp3control cell morphology and
infections spore formation in the fungal pathogen Histoplasma capsulatum. Proceedings of
the Nathional Academy of Sciences, v. 105, n. 38, p. 14573-14578, 2008.
WHEAT, L. J.; CONNOLLY, P.; SMEDEMA, M.; DURKIN, M.; BRIZENDINE, E.;
MANN, P.; PATEL, R.; MCNICHOLAS, P. M.; GOLDMAN, M. Activity of newer triazoles

74
against Histoplasma capsulatum from patients with AIDS who failed fluconazole. The
journal of Antimicrobial Chemoterapy, v. 57, n. 6, p. 1235-1239, 2006.
WHEAT, L. J. Nonculture diagnostic methods for invasive fungal infections. Current
Infections Disease Reports, v. 9, n. 6, p. 465-471, 2007.
WHEAT, L. J.; FREIFELD, A. G.; KLEIMAN, M. B.; BADDLEY, J. W.; MCKINSEY, D.
S.; LOYD, J. E.; KAUFFMAN, C. A. Clinical practice guidelines for the management of
patients with histoplasmosis: 2007 update by the Infectious Diseases Society of America.
Clinical Infectious Diasease, v. 45, n. 7, p. 807-825, 2007.
WHEAT, L. J.; KAUFFMAN, C. A. Histoplasmosis. Infectious Disease Clinics of North
America, v. 17, n. 1, p. 1–19, 2003.
WIDMER, F., WRIGHT, L. C.; OBANDO, D.; HANDKE, R.; GANENDREN, R.; ELLIS,
D. H.; SORRELL, T. C. Hexadecylphosphocholine (miltefosine) has broad-spectrum
fungicidal activity and is efficacious in a mouse model of cryptococcosis. Antimicrobial
Agents and Chemotherapy, v. 50, n. 2, p. 414–421, 2006.
WIEDER, T.; REUTTER, W., ORFANOS, C. E.; GEILEN, C. C. Mechanisms of action of
phospholipid analogs of anticancer compounds. Progress in Lipid Research, v. 38, n. 3, p.
249-259, 1999.
WIEDERHOLD, N. P.; NAJVAR, L. K.; BOCANEGRA, R.; KIRKPATRICK, W. R.;
SORRELL, T. C.; PATTERSON, T. F. Limited activity of miltefosine in murine models of
cryptococcal meningoencephalitis and disseminated cryptococcosis. Antimicrobial Agents
and Chemotherapy, v. 57, n. 2, p. 745-750, 2013.
YAN, Y.; YU, S.; CHAI, X.; HU, H.; WU, Q. Synthesis and Antifungal Activity of Novel
Triazole Derivatives. Archives of Pharmacal Research, v. 34, n. 10, p. 1649-1656, 2011.
ZANCOPÉ-OLIVEIRA, R. M.; MUNIZ, M. M.; WANKE, B. Histoplasmose. In: COURA, J.
R. Dinâmica das doenças infecciosas e parasitárias. 3. ed. Rio de Janeiro: Guanabara
Koogan, 2005. p. 1208-1221.
ZUO, X.; DJORDJEVIC, J. T.; OEI J, B.; DESMARINI, D.; SCHIBECI, S. D.; JOLLIFFE,
K. A.; SORRELL, T. C. Miltefosine induces apoptosis-like cell death in yeast via cox9p in
cytochrome c oxidase. Molecular Pharmacology, v. 80, n. 3, 476 – 485, 2011.

75
ANEXO A
QUADROS

76
Quadro 4. Concentração Inibitória Mínima (CIM) (µg/mL) da droga Miltefosina (MIL)
frente as cepas de Histoplasma capsulatum var. capsulatum na fase filamentosa.
Nº de Coleção Miltefosina - g mL
-1
CIM 50 % CIM 80 % CIM 100% CFM
CEMM 03-2-034 0,5 1 2 2
CEMM 03-2-088 0,5 1 2 2
CEMM 03-2-090 0,25 0,5 1 2
CEMM 03-3-035 0,5 1 2 2
CEMM 03-3-055 0,5 1 2 2
CEMM 03-6-059 0,5 1 2 2
CEMM 05-1-070 0,5 1 2 2
CEMM 05-1-096 0,5 1 2 2
CEMM 05-1-097 0,5 1 2 4
CEMM 05-1-099 0,5 1 2 2
CEMM 05-2-020 0,125 0,25 0,5 0,5
CEMM 05-2-021 1 2 4 4
CEMM 05-2-035 0,5 1 2 4
CEMM 05-2-037 0,5 1 2 2
CEMM 05-2-042 0,5 1 2 2
CEMM 05-2-053 0,5 1 2 4
CEMM 05-2-074 0,5 1 2 2
CEMM 05-2-084 0,5 1 2 2
CEMM 05-2-085 1 2 4 4
CEMM 05-2-086 0,5 1 2 4
CEMM 05-2-087 0,5 1 2 4
CEMM 05-2-088 1 2 4 4
CEMM 05-2-091 0,5 1 2 2
CEMM 05-2-093 0,25 0,5 1 1
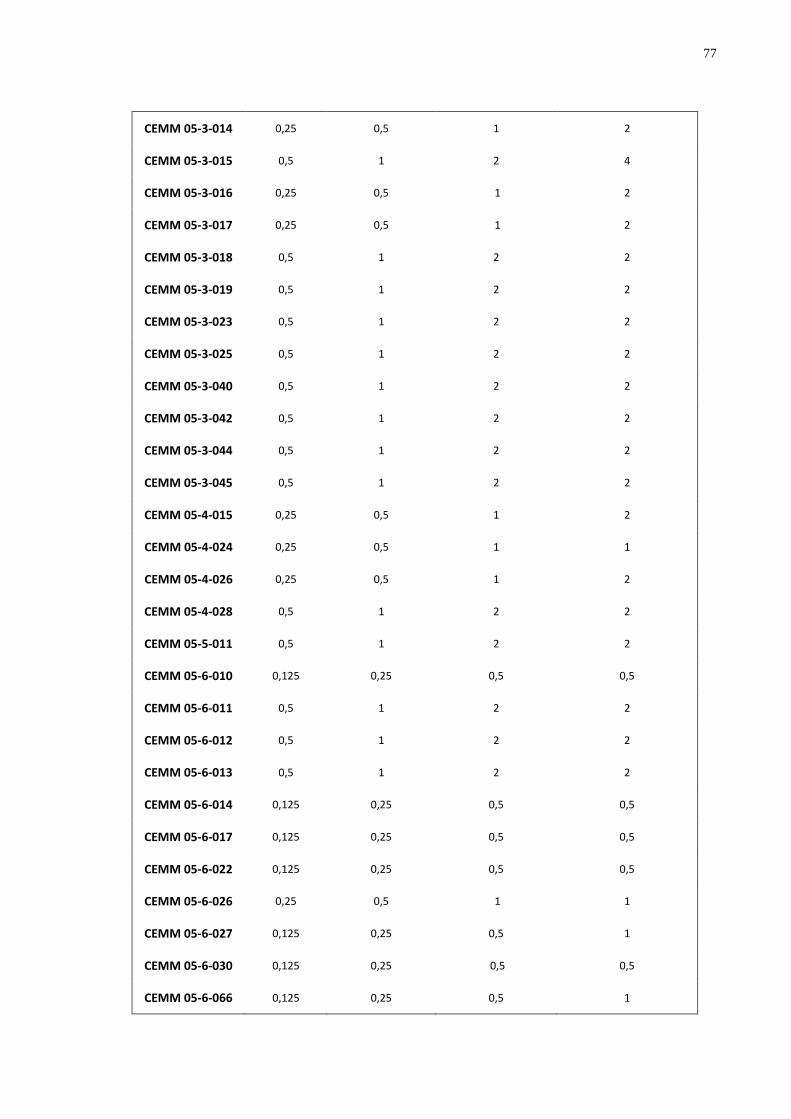
77
CEMM 05-3-014 0,25 0,5 1 2
CEMM 05-3-015 0,5 1 2 4
CEMM 05-3-016 0,25 0,5 1 2
CEMM 05-3-017 0,25 0,5 1 2
CEMM 05-3-018 0,5 1 2 2
CEMM 05-3-019 0,5 1 2 2
CEMM 05-3-023 0,5 1 2 2
CEMM 05-3-025 0,5 1 2 2
CEMM 05-3-040 0,5 1 2 2
CEMM 05-3-042 0,5 1 2 2
CEMM 05-3-044 0,5 1 2 2
CEMM 05-3-045 0,5 1 2 2
CEMM 05-4-015 0,25 0,5 1 2
CEMM 05-4-024 0,25 0,5 1 1
CEMM 05-4-026 0,25 0,5 1 2
CEMM 05-4-028 0,5 1 2 2
CEMM 05-5-011 0,5 1 2 2
CEMM 05-6-010 0,125 0,25 0,5 0,5
CEMM 05-6-011 0,5 1 2 2
CEMM 05-6-012 0,5 1 2 2
CEMM 05-6-013 0,5 1 2 2
CEMM 05-6-014 0,125 0,25 0,5 0,5
CEMM 05-6-017 0,125 0,25 0,5 0,5
CEMM 05-6-022 0,125 0,25 0,5 0,5
CEMM 05-6-026 0,25 0,5 1 1
CEMM 05-6-027 0,125 0,25 0,5 1
CEMM 05-6-030 0,125 0,25 0,5 0,5
CEMM 05-6-066 0,125 0,25 0,5 1

78
Quadro 5. Concentração Inibitória Mínima (CIM) (µg/mL) da droga Miltefosina (MIL)
frente as cepas de de Histoplasma capsulatum var. capsulatum na fase Leveduriforme.
Nº de Coleção Miltefosina - g mL
-1
CIM 50% CIM 80 % CIM 100% CFM
CEMM 03-2-034 0,25 0,5 1 1
CEMM 03-2-090 0,0625 0,125 0,25 0,25
CEMM 03-3-035 0,125 0,25 0,5 0,5
CEMM 03-3-055 0,125 0,25 0,5 1
CEMM 03-6-059 0,125 0,25 0,5 1
CEMM 05-1-097 0,25 0,5 1 2
CEMM 05-2-035 0,5 1 2 2
CEMM 05-2-037 0,125 0,25 0,5 0,5
CEMM 05-2-042 0,25 0,5 1 2
CEMM 05-2-053 0,5 1 2 2
CEMM 05-2-074 0,125 0,25 0,5 0,5
CEMM 05-2-087 0,125 0,25 0,5 1
CEMM 05-4-028 0,25 0,5 1 1
CEMM 05-5-011 0,125 0,25 0,5 1
CEMM 05-6-027 0,125 0,25 0,5 1

79
Quadro 6. Concentração Inibitória Mínima (CIM) (µg/mL) das drogas Miltefosina (MIL)
frente as cepas de Sporothrix spp.
CEPAS ESPÉCIE
Miltefosina - g mL-1
CIM 50% CIM 80% CIM 100% CFM
CEMM 05-3-049 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-050 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-051 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-052 S.brasiliensis 0,5 1 2 2
CEMM 05-3-053 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-054 S.brasiliensis 0,125 0,25 0,5 1
CEMM 05-3-055 S.brasiliensis 0,5 1 2 2
CEMM 05-3-056 S.brasiliensis 0,5 1 2 2
CEMM 05-3-057 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-058 S.brasiliensis 0,125 0,25 0,5 1
CEMM 05-3-059 S.brasiliensis 0,125 0,25 0,5 1
CEMM 05-3-060 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-061 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-062 S.brasiliensis 0,125 0,25 0,5 1
CEMM 05-3-063 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-064 S.brasiliensis 0,5 1 2 4
CEMM 05-3-065 S.brasiliensis 0,25 0,5 1 2
CEMM 05-3-066 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-067 S.brasiliensis 0,125 0,25 0,5 1
CEMM 05-3-068 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-069 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-070 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-071 S.brasiliensis 0,25 0.5 1 2
CEMM 05-3-072 S.brasiliensis 0,5 1 2 4
CEMM 05-3-073 S.brasiliensis 0,25 0,5 1 2
CEMM 05-3-074 S.brasiliensis 0,5 1 2 2

80
CEMM 05-3-075 S.brasiliensis 0,125 0,25 0,5 1
CEMM 05-3-076 S.brasiliensis 0,5 1 2 4
CEMM 05-3-077 S.brasiliensis 0,125 0,25 0,5 2
CEMM 05-3-078 S.brasiliensis 0,5 1 2 4
CEMM 05-3-079 S.brasiliensis 0,5 1 2 2
CEMM 05-3-080 S.brasiliensis 0,5 1 2 2
CEMM 05-3-081 S.brasiliensis 0,5 1 2 4
CEMM 05-3-082 S.brasiliensis 0,5 1 2 2
CEMM 05-3-083 S.brasiliensis 0,5 1 2 2
CEMM 05-3-084 S.brasiliensis 0,5 1 2 2
CEMM 05-3-085 S.brasiliensis 0,5 1 2 2
CEMM 05-3-086 S.brasiliensis 0,25 0,5 1 2
CEMM 05-3-087 S.brasiliensis 0,5 1 2 4
CEMM 05-3-088 S.brasiliensis 0,5 1 2 2
CEMM 05-3-089 S.brasiliensis 0,25 0,5 1 2
CEMM 05-3-091 S.brasiliensis 0,5 1 2 2
CEMM 05-3-093 S.brasiliensis 0,25 0,5 1 4
CEMM 05-3-094 S.brasiliensis 0,5 1 2 2
CEMM 05-3-096 S.brasiliensis 0,25 0,5 1 1
CEMM 05-3-098 S.brasiliensis 0,5 1 2 2
CEMM 05-3-099 S.brasiliensis 0,25 0,5 1 2
CEMM 05-4-003 S.brasiliensis 0,25 0,5 1 4
CEMM 05-3-048 S.schenckii 0,25 0,5 1 1
CEMM 05-3-090 S.schenckii 0,5 1 2 4
CEMM 05-3-092 S.schenckii 0,5 1 2 2
CEMM 05-3-095 S.schenckii 0,25 0,5 1 2
CEMM 05-3-097 S.schenckii 0,5 1 2 2
CEMM 05-4-002 S.schenckii 0,5 1 2 2
CEMM 05-3-100 S.mexicana 0,5 1 2 2

81
CEMM 05-4-001 S.mexicana 0,5 1 2 2
CEMM 05-4-007 S.mexicana 1 2 4 4
CEMM 05-4-008 S.mexicana 0,25 0,5 1 1
CEMM 05-4-009 S.mexicana 0,25 0,5 1 2
CEMM 05-4-004 S.globosa 1 2 4 4
CEMM 05-4-005 S.globosa 0,5 1 2 2
CEMM 05-4-006 S.globosa 1 2 4 4

82
ANEXO B
CORANTES, SOLUÇÕES E MEIOS DE CULTURA

83
1. CORANTES
1.1 Lactofenol azul-algodão
Objetivo
Líquido de montagem utilizado entre lâmina-lamínula para analisar a micromorfologia das
colônias fúngicas.
Composição
Ácido lático 20 g
Fenol 20 g
Glicerina 20 g
Azul-algodão 0,05 g
Água destilada 20 mL
Modo de preparo
1) Fundir os cristais de fenol e juntar com as demais componentes;
2) Aguardar 24 horas e filtrar com o auxílio de papel-filtro;
3) Acondicionar em um frasco âmbar com conta-gotas;
4) Colocar de uma a duas gotas do corante colocando 2 gotas sobre uma lâmina de
microscopia limpa, depositar, em seguida, um pequeno fragmento da colônia fúngica a ser
analisado, sobre o líquido e homogeneizar . Cobrir com uma lamínula e observar ao
microscópio óptico na objetiva de 40x.
2. SOLUÇÕES
2.1 Solução salina
Objetivo
Utilizar para o preparo de inóculo fúngico.
Composição
Cloreto de sódio 0,9 g
Água destilada 100 mL
Modo de preparo
1) Adicionar o cloreto de sódio a água destilada, homogeneizando até sua completa
dissolução;
2) Autoclavar por 15 minutos, a 121°C;
3) Estocar a 4°C.
2.2 MOPS (ácido 2-[N-morfolino] propanosulfônico)

84
Objetivo
Usado para o ajustar o pH de soluções, em especial o meio RPMI 1640.
Composição
MOPS 6,9 g
Água destilada 200 mL
Modo de preparo
1) Dissolver o MOPS em água destilada autoclavada;
2) Armazenar a 4°C em garrafa envolvida com papel alumínio.
2.3 DMSO (dimetil sufóxido)
Objetivo
Usado preparar soluções de algumas drogas antifúngicas.
Composição
DMSO 50 mL
Modo de preparo
1) Autoclavar por 15 minutos, a 121°C;
2) Estocar a 4°C.
3. MEIOS DE CULTURA
3.1 Ágar Batata
Objetivo
Utilizado para realizar repiques e estoque de cepas fúngicas.
Composição
Infusão de batatas 500 mL
Dextrose 10 g
Ágar bacteriológico 15 g
Água destilada 1000 mL
Modo de preparo
1) Cozinhar 250 g de batata-inglesa (Solanum tuberosum) descascadas em 500 mL de água,
por 1h;
2) Filtrar o cozido de batatas através de gaze;
3) Restituir o volume inicial de água (500 mL) e acrescentar 500 mL de água destilada;
4) Adicionar o ágar e a dextrose, até a dissolução completa;

85
5) Autoclavar o meio por 15 minutos a 121°C e distribuir, alíquotas de 4 mL, em tubos de
ensaio;
6) Esperar solidificar, na posição inclinada, de modo a obter uma superfície de
aproximadamente 6 cm.
3.2 Ágar BHI (Brain Heart Infusion)
Objetivo
Usado para isolamento primário de fungos que precisam de meio enriquecido para seu
crescimento, principalmente de fungos dimórficos.
Composição
Cérebro-coração, infusão de sólidos 8,0 g
Hidrolisado péptico de tecido animal 5,0 g
Hidrolisado pancreático de caseína 16,0 g
Cloreto de sódio 5,0 g
Glucose 2,0 g
Fosfato dissódico de hidrogênio 2,5 g
Ágar 13,5 g
Água destilada q.s.p 1000 mL
Modo de preparo
1) Preparar conforme indicação do fabricante, dissolvendo os componentes em água destilada;
2) Autoclavar o meio por 15 minutos a 121°C e distribuir, alíquotas de 4 mL, em tubos de
ensaio;
3) Deixar solidificar, na posição inclinada, de modo a obter uma superfície de
aproximadamente 6 cm.
3.3 Ágar Sabouraud a 2%
Objetivo
É o principal meio de cultura usado na Micologia. Utilizado para o cultivo primário de fungos
no geral (leveduras, filamentosos a alguns dimórficos).
Composição
Peptona 10 g
Dextrose 20 g
Ágar bacteriológico 20 g
Água destilada 1000 mL

86
Modo de preparo
1) Dissolver ágar, peptona e dextrose em água destilada, aquecendo em banho-maria;
2) Autoclavar por 15 minutos, a 121°C;
3) Distribuir alíquotas de 4 mL em tubos de ensaio, deixar solidificar, na posição inclinada, de
modo a obter uma superfície de 6 cm.
3.4 Ágar Sabouraud a 2% ou Ágar BHI com Sangue
Objetivo
Utilizados, especialmente, para a reversão de fungos dimórficos.
Composição
Ágar Sabouraud a 2% ou ágar BHI 50 mL
Sangue de carneiro desfibrinado 5 mL
Modo de preparo
1) Preparar 50 mL de ágar Sabouraud a 2% ou ágar BHI, autoclavar e deixar resfriar, até
atingir a temperatura de 48°C;
2) Adicionar o sangue, homogeneizar e distribuir alíquotas de 4 mL em tubos de ensaio;
3) Deixar solidificar, na posição inclinada, de modo a obter uma superfície de 6 cm.
3.5 RPMI 1640
Objetivo
Meio de cultura padronizado pelo CLSI, para execução de testes de sensibilidade a
antifúngicos, através da técnica de microdiluição ou macrodiluição em caldo .
Composição
RPMI 1640 (com glutamina e sem bicarbonato de sódio) 10,5 g
Água destilada 1000 mL
Modo de preparo
1) Dissolver o meio em água destilada;
2) Ajustar o pH final para 7,0 (com solução de MOPS);
3) Completar o volume com água destilada e esterilizar o meio por filtração em membrana de
0,22 μm de poro, utilizando pressão positiva;
4) Manter sob refrigeração até o momento do uso.