unidad 4 metabolismo de los carbohidratos. fátima canul manzanero
TRANSCRIPT

*INSTITUTO TECNOLOGICO DE MERIDA*
MATERIA
BIOQUÍMICA.
UNIDAD 4
METABOLISMO DE CARBOHIDRATOS.
NOMBRE DEL PROFESOR
PROF. SARA ALICIA GONZALEZ NOVELO.
ALUMNO
FATIMA CANUL MANZANERO.
FECHA DE ENTREGA
JUEVES 25 DE ABRIL DE 2013.

Introducción.
En el presente trabajo nos centraremos específicamente en hacer un trabajo escrito de todo aquello que involucre y refiere al metabolismo de los carbohidratos, a partir de investigaciones y análisis sobre el tema.
Como primer punto se expondrá la vía degradativa de Carbohidratos, en el cual identificaremos las enzimas, coenzimas, cofactores que en ella actúan, así como el balance energético total y el mecanismo de reacción durante el catabolismo.
La comparación de la glucólisis y la gluconeogénesis, en el cual se identificará los puntos de reacción para que la vía sea espontánea o térmicamente posible.
También los mecanismos de degradación, síntesis y regulación del glucógeno, así como mencionar las rutas de esta vía, las enzimas que en ella actúan y los procesos energéticos.
Las pentosas fosfato, ubicando sus productos que se obtendrán en relación a las 3 fases del metabolismo intermediario, y como este es precursor de otras vías metabólicas importantes.
Para finalizar se identificarán las 2 fases, productos, sustratos y balance general del ciclo de Calvin.

Vía degradativa de Carbohidratos.
La vía degradativa de Carbohidratos tiene lugar por medio de la Glucólisis y se realiza a partir de 10 reacciones enzimáticas que se explicaran a continuación.
Se puede entender que la glucólisis transcurre en dos fases.
La primera fase comprende de la reacción 1-5, en donde la glucosa es fosforilada para dar lugar a dos triosas de gliceraldehído 3 fosfato. En este proceso se dará un consumo de 2 ATP.
La segunda fase comprende de la reacción 6-10, en donde las dos triosas de gliceraldehído 3 fosfato se convertirán a piruvato, dando lugar a la producción de 4 ATPs. El rendimiento neto de la glucólisis será de 2 ATPs por cada molécula de glucosa.
Reacciones.
1.-Primera reacción es la transferencia de un grupo fosforilo de ATP a la glucosa para formar Glucosa 6 fosfato, esta reacción es catalizada por la enzima Hexoquinasa. Necesita la ayuda del cofactor Mg2+. Consumo del primer ATP.
Glucosa + ATP Glucosa 6 fosfato + ADP+ H+
Hexoquinasa.
Mg 2+
2.- Segunda reacción es la isomerización de la Glucosa 6 fosfato a Fructosa 6 fosfato que va a ser catalizada por la enzima Fosfohexoisomerasa. Esta reacción no necesita ayuda de ningún cofactor.
Glucosa 6 fosfato Fructosa 6 fosfato.
3.- Tercera reacción, fosforilación de la Fructosa 6 fosfato para formar Fructosa 1-6 bifosfato. La reacción será catalizada por la enzima Fosfofructoquinasa. Necesita la ayuda del cofactor Mg2+. En esta reacción se dará el consumo del segundo ATP.
Fosfofructoquinasa.
Fructosa 6 fosfato + ATP Mg2+ Fructosa 1-6 bifosfato + ADP+ H+

4.- Cuarta reacción que consiste en el rompimiento de la Fructosa 1-6 bifosfato en dos triosas: el gliceraldehído 3 fosfato y la Dihidroxiacetona fosfato. Esta reacción es catalizada por la enzima Aldolasa.
Fructosa 1-6 bifosfato Gliceraldehído 3 fosfato + Dihidroxiacetona fosfato.
Aldolasa.
5.- Quinta reacción, interconversión de las triosas fosfato, la Dihidroxiacetona fosfato es isomerizada en Gliceraldehído 3 fosfato, para la continuación en la ruta metabólica. La enzima que cataliza dicha reacción es la Triosa fosfato isomerasa.
Dihidroxiacetona fosfato Gliceraldehído 3 fosfato.
Triosa fosfato isomerasa.
6.- Sexta reacción es la oxidación y fosforilación del gliceraldehído 3 fosfato con ayuda de coenzima NAD+ y Pi que es catalizada por la enzima gliceraldehído 3 fosfato deshidrogenasa.
Gliceraldehído 3 fosfato + NAD+ +Pi 1-3 bifosfoglicerato + NADH + H+
Gliceraldehído 3 fosfato deshidrogenasa.
7.- Séptima reacción, producción del primer ATP, se transfiere un fosfato al ADP para generar así el primer ATP. Obtiene 3 fosfoglicerato a partir del 1-3bifosfoglicerato cuya enzima que interviene es la Fosfogliceratoquinasa. Interviene el cofactor Mg 2+.
1-3 fosfoglicerato + ADP Mg2+ 3 fosfoglicerato + ATP
Fosfogliceratoquinasa
8.- Octava reacción, conversión del 3 fosfoglicerato a 2 fosfoglicerato, es catalizado por Fosfoglicerato mutasa, que ayudará al cambio de posición del grupo funcional de un carbono a otro. No interviene cofactor.
3 Fosfoglicerato fosfoglicerato mutasa 2 fosfoglicerato.

9.- Novena reacción, deshidratación del 2 fosfoglicerato para obtener fosfoenolpiruvato. Esta reacción será catalizada por la enzima Enolasa que promoverá la eliminación de una molécula de h2o.
2 fosfoglicerato fosfoenolpiruvato + h2o
Enolasa.
10.- Décima reacción, en la cual se va a dar una desfosforilación del fosfoenolpiruvato para obtener así piruvato y ATP. A través de la enzima Piruvato Quinasa. Interviene el cofactor Mg2+.
Mg2+
Fosfoenolpiruvato + ADP+ H+ piruvato quinasa Piruvato + ATP.
Balance Global
De la glucólisis obtendremos el siguiente balance energético global, esto se dará por cada molécula de glucosa.
Glucosa + 2 NAD+ + 2 ADP + 2 Pi 2 piruvato + 2 NADH + 2 H+ + 2 ATP + 2 H2O.
En la glucólisis se genera energía por medio de la producción de ATP, y se sabe también que parte de la energía se tiene en el piruvato
Podemos dividir la ecuación de la glucólisis en 2 procesos:
La primera que es la conversión de glucosa en piruvato, que es Exergónica:
Glucosa + 2NAD piruvato+ 2 NADH + 2H ⁺
ΔG₁= -146 kJ/mol
Formación de ATP a partir de ADP y Pi, que es Endergónica:
2ADP + 2Pi 2ATP + 2H₂O
ΔG₂= 2(30.5 kJ/mol= 61.0 kJ/mol

Comparación de Glucolisis y Gluconeogénesis reconociendo los puntos de reacción que permiten a esta vía ser espontánea ó termodinámicamente favorable.
Glucólisis. Gluconeogénesis.Degradación de los carbohidratos. Síntesis de glucosa a partir de
precursores no glucídicos.La glucosa se convierte en Piruvato. Piruvato se convierte a glucosa.
Las reacciones de la glucólisis tienen lugar en el citosol de la célula.
Principal lugar de la Gluconeogénesis tienen lugar en el hígado y el riñón.Las reacciones se llevan a cabo en el citosol excepto de la piruvato carboxilasa(Mitocondria)
La Velocidad de reacción en la vía glucolítica va a estar determinada por la concentración de la glucosa.
La intensidad de la Gluconeogénesis va a estar determinada por la concentración del lactato y otros precursores.
Consta de 10 reacciones, donde se consideran 2 etapas:1.- Con un gasto de energía(consumo de ATP)2.- Producción de energía.
Consta de 11 reacciones, en donde 3 de estas reacciones son irreversibles y son las que difieren con la glucólisis.
Se obtiene una producción neta de 2 moléculas de ATP al final de la vía glucolítica.
Tiene un consumo de 4 ATPs la vía.
El balance energético de la glucólisis es:Glucosa + 2 NAD+ + 2 ADP + 2 Pi 2 piruvato + 2 NADH + 2 H+ + 2 ATP + 2 H2O.
Balance energético de la Gluconeogénesis es:2Piruvato+4ATP+2GTP+2NADH+6H2o
Glucosa+4ADP+2GDP+6Pi+2NAD+2H
Permite la entrada de otros azúcares a la vía glucolítica como son: Fructosa, Galactosa y Manosa.
Los precursores más importantes de la gluconeogénesis son: Lactato, aminoácidos y el glicerol.
Tiene como puntos de control enzimas que catalizan reacciones irreversibles como son:*Hexoquinasa.*Fosfofructoquinasa.*Piruvatoquinasa.
Tiene como puntos de control enzimas que catalizan reacciones como:*Piruvato Carboxilasa.*Fructosa 1-6 bifosfatasa.*Fosfoenolpiruvatocarboxilasa.

Los puntos de control de la vía glucolítica serán activadas e inhibidas según cada enzima.Hexoquinasa: Inhibe Glucosa6fosfato.
Fosfofructoquinasa: Inhibe ATP, Citrato. Activa: AMPPiruvato Quinasa: Inhibe: ATP, alanina. Activa: Fructosa1-6 bifosfato.
Los puntos de control de la vía de la gluconeogénesis serán activados e inhibidos según cada enzima.Piruvato Carboxilasa: Inhibe: ADP
Activa: Acetil Coa.Fructosa 1-6bifosfatasa: Inhibe: AMP
Activa: Citrato.Fosfoenolpiruvatocarboxilasa: Inhibe: ADP
Es una reacción Exergónica. Reacción Exergónica.
La conversión de glucosa en dos moléculas de piruvato liberan una energía libre de -96 KJ/mol-1
La energía libre real para la formación del piruvato a partir de glucosa es de -84 KJ/mol-1

Degradación, Síntesis y regulación del Glucógeno.
El glucógeno es un polímero muy grande y ramificado de residuos de glucosa, que puede ser roto para dar lugar a moléculas de glucosa cuando el consumo de energía es necesario.
El glucógeno es una forma de almacenamiento de glucosa que tiene facilidad de movilización, en los vertebrados el glucógeno se encuentra principalmente en el hígado y el músculo esquelético; puede representar hasta el 10% del peso del hígado y del 1 al 2% del peso del músculo. El glucógeno está presente en el citoplasma en forma de gránulos citosólicos.
En gran medida el glucógeno en el músculo se encuentra allí para proporcionar una fuente de energía rápida para el metabolismo tanto aeróbico como anaeróbico. En cambio el glucógeno que se encuentra en el hígado sirve como almacén de glucosa para otros tejidos cuando no hay glucosa disponible en el organismo.
Los mecanismos de almacenamiento y movilización de glucógeno son los mismos en el músculo que en el hígado, aunque difieren un poco en cuanto a las enzimas que actúan en cada uno de ellos. El glucógeno se degrada en el intestino lo que implica un conjunto de enzimas diferentes hidrolíticas que convierten el glucógeno en glucosa libre.
El metabolismo del glucógeno consiste en la liberación y el almacenamiento de glucosa de forma regulada.
Degradación del glucógeno.
Se puede decir que la degradación del glucógeno consta de 3 pasos:
1.- Liberación de Glucosa 1-fosfato a partir del Glucógeno.
2.- Remodelación del glucógeno para que la degradación pueda continuar.
3.- conversión de glucosa 1-fosfato a glucosa 6-fosfato.
La glucosa 6-fosfato que proviene de la degradación que antes se mencionó, puede tener 3 destinos distintos.
1.- Ser el sustrato inicial de la glucólisis.
2.- Convertirse en glucosa libre y liberarse en el torrente sanguíneo.
3.- Puede procesarse por la vía de las pentosas fosfato para generar NADPH y derivados de la ribosa.

La degradación eficiente del glucógeno para suministrar glucosa 6-fosfato para su metabolismo posterior, requiere la actividad de 4 enzimas.
Uno para degradar el glucógeno. Dos para remodelar la molécula de glucógeno de forma que permanezca como un sustrato apto para la degradación. Uno para transformar el producto de la ruptura del glucógeno en una forma apropiada para su metabolismo posterior.
Síntesis del glucógeno.
La síntesis de glucógeno tiene lugar de forma diferente a la de su degradación. La UDP-glucosa, el intermediario activado en la síntesis del glucógeno, se forma a partir de glucosa 1-fosfato y UTP. La glucógeno sintasa cataliza la transferencia de glucosa desde la UDP-glucosa hasta el grupo hidroxilo del C-4 de un residuo terminal de una molécula de glucógeno en crecimiento. La síntesis se inicia por la glucogenina, una proteína capaz de autoglucosilarse y que contiene un oligosacárido unido a un residuo. Una enzima ramificante convierte algunos enlaces sigma 1-4 en enlaces sigma 1-6 para aumentar el número de terminaciones de modo que el glucógeno pueda producirse y degradarse rápidamente.
Regulación del Glucógeno.
La síntesis y degradación del glucógeno están reguladas cuidadosamente para que pueda disponerse de suficiente glucosa para las necesidades energéticas del organismo. La glucogénesis y la Glucogenólisis están controladas principalmente por tres hormonas: Insulina, Glucagón y adrenalina.
La adrenalina y el glucagón estimulan la degradación del glucógeno e inhiben su síntesis por incremento del nivel citoplasmático de AMP cíclico, el cual activa a la proteína quinasa A. La proteína quinasa A activa la degradación del glucógeno por unión de un fosfato a la fosforilasa quinasa e inhibe la síntesis del glucógeno por fosforilación de la glucógeno sintasa.
Las acciones de movilización del glucógeno debidas a la PKA son invertidas por la proteína fosfatasa 1, enzima regulado por varias hormonas. La Adrenalina inhibe a esta fosfatasa al bloquear su unión a las moléculas de glucógeno y al activar a un inhibidor.
La insulina desencadena una fosforilación y la inactivación del glucógeno sintasa quinasa.
Entonces la síntesis de glucógeno decrece a causa de la adrenalina y aumenta a causa de la insulina.

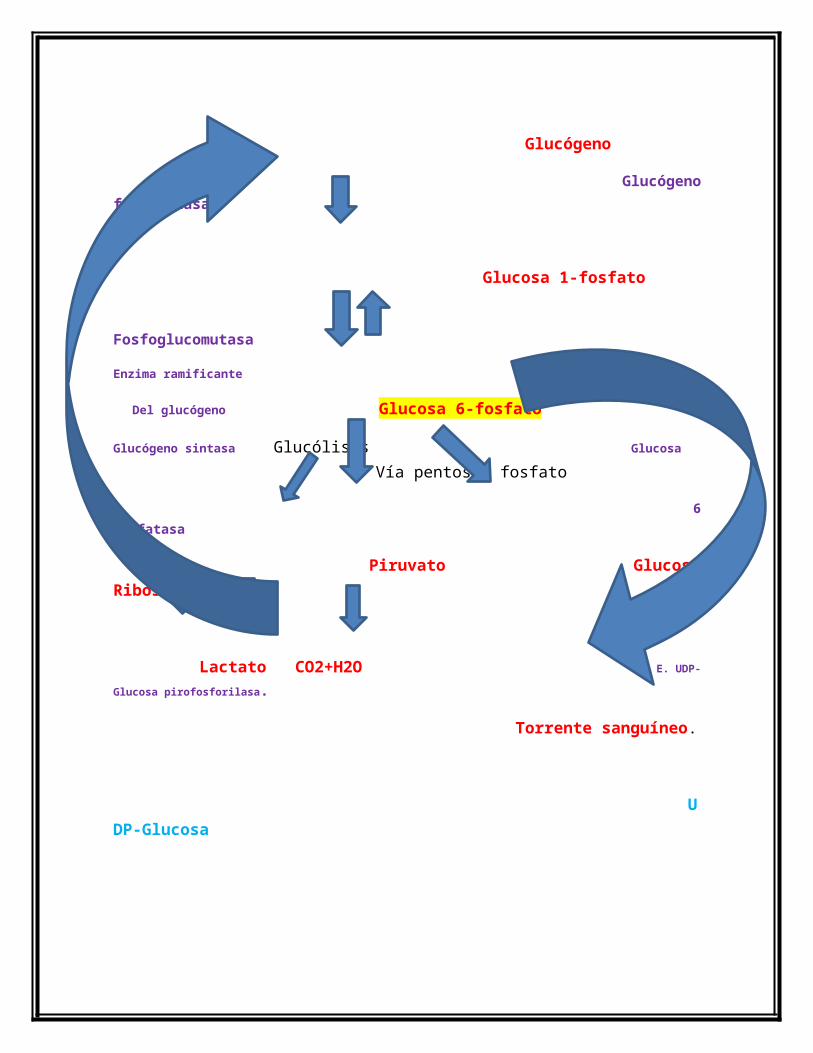
Diagrama de la Glucogenólisis y Glucogénesis.
Glucógeno
Glucógeno fosforilasa
Glucosa 1-fosfato
Fosfoglucomutasa
Enzima ramificante
Del glucógeno Glucosa 6-fosfato
Glucógeno sintasa Glucólisis Glucosa Vía pentosas fosfato
6 fosfatasa
Piruvato Glucosa Ribosa+ NADPH
Lactato CO2+H2O E. UDP-Glucosa pirofosforilasa.
Torrente sanguíneo.
UDP-Glucosa
Vía de las Pentosas fosfato.

Es una ruta metabólica secundaria de la glucosa cuya principal función es generar energía, pero no como ATP, si no en forma de poder reductor: NADPH + H+
Tiene funciones principales cuantitativamente hablando:
1. Producir poder reductor (NADPH + H+)
2. Producir Pentosas: Ribosa-5-P: pentosa-P más importante porque a partir de ella formamos ácidos nucleicos (RNA, DNA), ATP, FAD, NAD.
De manera que a partir de la ribosa-5-P no solo formamos ácidos nucleicos sino también multitud de nucleótidos importantísimos en el metabolismo.
También tiene otras funciones secundarias:
1. Producir CO2 (necesario para la formación de AG)
2. Degradar glucosa (ruta del catabolismo de la glucosa)
3. Degradar pentosas.
Características de la vía de las pentosas fosfato:
- Es la vía multifuncional por excelencia en el organismo y muy especializada en función de las necesidades de las células.
- Se produce en el citosol.
- Localización, atendiendo a las necesidades de las células:
-Se produce en pequeñas cantidades en todas las células (todas las células necesitan ribosa).
-Se producirá de manera cuantitivamente mayoritaria en todas aquellas células que necesiten mucho NADPH.
-Se produce de forma esencial en los eritrocitos, la vía de las pentosas será de las únicas rutas metabólicas que se den en el eritrocito.
-Atendiendo a la función principal del poder reductor (NADPH), es decir, la síntesis de lípidos, esta vía se producirá también en el tejido adiposo y en hígado
En resumen, la vía de las pentosas fosfato se produce en pequeñas cantidades en todas las células.
División de la vía de las pentosas fosfato en 2 ramas.

1) Rama oxidativa.
Es totalmente irreversible y es dónde se produce todo el reductor (NADPH) de la vía, y donde se genera ya la primera pentosa.
2) Rama no oxidativa
Es totalmente reversible y tiene como principal función la interconversión de monosacáridos-fosfato.
Rama oxidativa de la vía de las pentosas fosfato Reacciones:
Rama oxidativa
Parte de glucosa-6-fosfato (G6P) que en la 1ª reacción oxida su grupo aldehído (des hidrogenación) a carbonilo dando lugar a la (delta) δ-lactona del 6-fosfogluconato, y (puesto que es una oxidación) el NADP+ se reduce a NADPH + H+
Enzima: *Glucosa-6-fosfato deshidrogenasa* (G6P DH), cuyo inhibidor será el NADPH, es decir, cuando haya mucha cantidad de NADPH la enzima G6P DH se inhibe. Es la enzima (limitante) más importante de la vía ya que cataliza la 1ª reacción irreversible. Activador (+) hormonal: la insulina aumenta los niveles de la G6P DH
En una 1ª oxidación el ácido 6-fosfoglucónigo se va a formar ciclado, por eso se genera la δ-lactona del 6-fosfogluconato, que es muy inestable, por lo que se rompe inmediatamente para dar el 6-fosfogluconato
.
2. La 2ª reacción consiste en una 2ª oxidación de la δ-lactona del 6-fosfogluconato para forma el 6-fosfogluconato (6-P-gluconato), con la adición de 1 molécula de H2O.
Enzima: Lactonasa
3.-Última reacción de la rama oxidativa: el 6-fosfogluconato se descarboxila a ribulosa-5-fosfato (cetosa) formando la 1ª pentosa de la vía. Es una reacción similar a la 1ª y se genera una 2ª molécula de poder reductor (NADPH + H+). Además se genera también CO2 (producto secundario de la vía).
Enzima: 6-Fosfogluconato deshidrogenasa (6-P-gluconato DH)
En resumen, la rama oxidativa consiste en una doble oxidación, 1º de G6P a 6-P-gluconato, que después se oxida y descarboxila a ribulosa-5-P. Ya que es la fase irreversible (única irreversible de toda la vía), es la rama que controla toda la ruta (fase limitante), y en concreto, sobre todo la primera enzima, la G6P DH, que va a ser la mayor reguladora de la vía.

Balance material y energético de la Rama Oxidativa:
Producción a partir de 1G6P de 1 pentosa (cetosa), la 1ª de la vía: ribulosa-5-P.
Liberación de 1 CO2 y de 2 moléculas de poder reductor (2 NADPH + H+).
Rama no oxidativa
Recordemos que una cetosa es un monosacárido que tiene un grupo cetona (-CO) Dihidroxiacetona = cetosa más simple; y una aldosa es un monosacárido que tiene un grupo aldehído (-CHO) gliceraldehído = aldosa más simple.
Función: interconversión de unos monosacáridos-fosfato en otros, respondiendo siempre a un patrón común:
Para que se inicie la 2ª fase de la vía de las pentosas, es decir, la rama no oxidativa, necesitaré una cetosa y una aldosa que siempre seguirán ese mecanismo, y que pueden intercambiar 2 ó 3 carbonos.
La rama oxidativa ha terminado en una cetosa (ribulosa-5-P), pero esa no nos sirve de forma directa para comenzar la fase no oxidativa, si no que a partir de ella obtendré una cetosa y una aldosa que serán las que inicien esta 2ª rama.
- Reacciones:
1. Una cetosa-fosfato se va a transformar en una aldosa-fosfato, a la vez que una aldosa-P se trasforma en una cetosa-P.
Cuando el intercambio es de 2C enzima: TRANSCETOLASA (que necesita TPP - Pirofosfato de Tiamina - como CoE para funcionar)
Cuando el intercambio es de 3C enzima: TRANSALDOLASA (NO necesita ninguna CoE)
La CETOSA y la ALDOSA nos las proporciona la Ribulosa-5-P (pentosa producto final de la rama oxidativa), que es una cetosa, y se puede transformar fácilmente en otra cetosa, la Xilulosa-5-P.
Ribulosa-5-P y Xilulosa-5-P son epímeros entre sí, es decir, solo se diferencian en la posición del –OH en un carbono.
Enzima: EPIMERASA, Isómeros, una un grupo cetona y otra un grupo aldehído.
También nos puede dar una Ribosa-5-P, que es la importante.
Enzima: ISOMERASA
Esta fase también es conocida como etapa de conexión.

2. La XILULOSA-5-P y la RIBOSA-5-P iniciarán la Rama No Oxidativa. TRANSCETOLASA (TPP) 2C
Xilulosa-5-P (cetosa 5C) cede 2C = G3P (aldosa 3C)
Ribosa-5-P (aldosa 5C) acepta 2C = sedoheptulosa-7-P (cetosa de 7 C no importante en humanos, tan solo como simple intermediario)
3. TRANSALDOLASA 3C
G3P (aldosa 3C) acepta 3C = *F6P* (producto final)
La fructosa se queda como producto final pero por el otro lado la rama continúa.
Sedoheptulosa-7-P (cetosa 7C) cede 3C = eritriosa-4-P (aldosa 4C no importante en humanos)
4. TRANSCETOLASA 2C
Eritriosa-4-P (aldosa 4C) acepta 2C = *F6P* (producto final)
*Xilulosa-5-P* (cetosa5C) cede 2C = *G3P* (producto final)
En resumen la rama no oxidativa de la Vía de las Pentosas Fosfato se trata de la interconversión de monosacáridos: una cetosa en una aldosa y una aldosa en una cetosa. La Cetosa cediendo y aldosa aceptando, 2 (transcetolasa) ó 3 C (transaldolasa). Todas las reacciones serán reversibles.
Es una vía que se puede parar en cualquier punto, por lo que es adaptable. De manera que por eso es multifuncional y muy especializada, ya que dependiendo de lo que necesite la célula se adapta.
Balance material y energético de la Rama No Oxidativa:
Partimos de 1 Ribosa-5-P y 2 Xilulosas (otra al final)
1 Ribosa-5-P + 2 Xilulosas-5-P 2 F6P + 1
La Vía de las Pentosas Fosfato en función de las necesidades de la célula (evidencia de su multifuncionalidad) presenta 4 modalidades distintas que clasificamos en 2 grupos según su frecuencia:
- Poco frecuentes:
1.- Cuando la célula requiere cantidades de ribosa-5-P y de poder reductor (NADPH) equilibradas: realiza la rama oxidativa y se para, formando ribosa.
2. Cuando la célula requiere mucha ribosa-5-P y no poder reductor para formar ácidos nucleicos, solo en el momento de la división celular.

- Frecuentes:
3.- Cuando la célula requiere elevadas cantidades de NADPH simplemente como agente reductor en general (más poder reductor que ribosa): la vía de las pentosas fosfato se combina con la gluconeogénesis y da lugar al ciclo de las pentosas fosfato.
4.- Cuando la célula requiere mucho poder reductor (NADPH + H+) como agente reductor y además para sintetizar lípidos, por lo que además requiere energía (ATP) = *modalidad más frecuente de las 4* que adquirirá entrando en la glucólisis.
Pentosas-fosfato.
Es una de las modalidades de la Vía de las Pentosas más frecuente en la célula, y será muy activa en eritrocitos, ya que son los que necesitan mantener muy estables (mediante el poder reductor) sus membranas (lípidos reducidos) para un adecuado transporte.
En resumen, cuando la célula necesita mucho más poder reductor la vía se produce en su totalidad (rama oxidativa y rama no oxidativa).


Ciclo de Calvin.
Los organismos fotosintéticos que utilizan el ciclo de Calvin sintetizan la glucosa a partir de dióxido de carbono y de agua, y utilizan la luz solar como fuente de energía. El ciclo de Calvin introduce en la biosfera la totalidad de los átomos de carbono que se utilizarán como combustible y como esqueleto carbonado de las biomoléculas.
El ATP y el NADPH formados en las reacciones de la fase luminosa de la fotosíntesis se utilizan para convertir el CO2 en hexosas y otros compuestos orgánicos.
La fase oscura de la fotosíntesis, es llamada Ciclo de Calvin.
El ciclo de Calvin presenta 3 etapas:
1.- Fijación del CO2 sobre la Ribulosa 1,5 bisfosfato para formar dos moléculas de 3 fosfoglicerato.
2.- la reducción del 3 fosfoglicerato para formar hexosas.
3.- La regeneración de la Ribulosa 1,5 bifosfato de tal modo que se pueda fijar mas CO2.
Estas reacciones tienen lugar en el estroma de los cito plastos, los organelos foto sintetizadores.
El primer paso del ciclo de Calvin es la fijación del CO2. La molécula de CO2 se condensa con la Ribulosa 1,5 bifosfato para formar un compuesto de 6 carbonos que se hidroliza rápidamente en dos moléculas de 3-fosfoglicerato.
Los pasos para convertir 3 fosfoglicerato en fructosa 6-fosfato se dan de manera muy parecida a los de la gluconeogénesis. El cambio que se presenta es que la enzima gliceraldehído 3-fosfato deshidrogenasa del cloroplasto es específica para NADPH en vez de serlo para NADH.
Por medio de reacciones mas complejas va a tener lugar la regeneración de la Ribulosa 1,5 bifosfato a partir de la fructosa 6 fosfato, gliceraldehído 3 fosfato e hidroxiacetona fosfato.
Por cada molécula de CO2 que se convierte en una hexosa se consumen 3 moléculas de ATP y dos moléculas de NADPH.
La actividad del ciclo de Calvin va a depender de las condiciones ambientales, la tiorredoxina reducida, formada en la transferencia de electrones desde la ferredoxina, producida por la luz, activa las enzimas del ciclo de Calvin, mediante reducción de sus puentes disulfuro.

La Elevación del Ph y del nivel de Mg2+ provocada por la luz en el estroma tiene un papel importante en la estimulación de la carboxilación de la Ribulosa 1,5 bifosfato por la rubisco.
La producción neta de carbono en el ciclo de Calvin es de una molécula de glieraldehido-3-fosfato, las moléculas de3-G-P restantes se procesan para generar 3 moléculas de ribulosa-1,5-bifosfato; 2 moléculas de isomerizan para formar 2 de hidroxiacetona fosfato.
Una molécula de Dihidroxiacetona se combina con una tercera molécula de gliceraldehido-3-fosfato para formar fructosa-2,6-bifosfato.
La fructosa-2,6-bifosfato se combina nuevamente con una molécula de gliceraldehido-3-fosfato para formar xilulosa-5-fosfato y eritrosa-4-fosfato.
La eritrosa-4-fosfato se une a una hidroxiacetona para formar sedoheptulosa-1,7-bifosfato, la cual se hidroliza para formar sedoheptulosa-7-fosfato.
Una quinta molécula de gliceraldehido-3-fosfato se une con la sedoheptulosa-7-fosfato para formar ribosa-5-fosfato y una segunda molécula de xilulosa-5-fosfato, estas 3 moléculas se convierten ribulosa-5-fosfato y en un último paso las 3 moléculas de Ri-5-P se fosforilan a expensas de 3 moléculas de ATP para formar3 moléculas de ribulosa-1,5-bifosfato. La molécula restante de gliceraldehido-3fosfato es utilizada dentro del cloroplasto en la síntesis de almidón o se exporta al citoplasma en donde puede emplearse en la síntesis de sacarosa o algún otro metabolito.
La ecuación neta del ciclo de Calvin es:
3CO₂ + 6NADPH + 9ATP Glieraldehido-3-fosfato + 6 NADPH⁺ +9ADP + 8Pi
En donde por cada 3 moléculas de dióxido de carbono que se incorporan en moléculas de carbohidratos, existe una ganancia neta de una molécula de gliceraldehido-3-fosfato. La fijación de 6CO₂ en una molécula de hexosa tiene lugar a expensas de 12 NADPH y 18 ATP.

Conclusión.
Podemos concluir que el metabolismo de los carbohidratos es uno de los más importantes pues ellos nos ayudan a que nuestro organismo actúe de la mejor manera posible y lleva a cabo todas sus funciones.
Es a partir de este metabolismo en el cual los organismos obtienen la energía necesaria para sus funciones, también de la relación que van teniendo cada uno de ellos, la glucolisis con la Glucogenólisis, el ciclo de Calvin con el ciclo de las pentosas.
Es muy frecuente que los productos de los metabolismos sean los reactivos de otros y es así de como todos se relacionan.

Bibliografía.
Principios de bioquímica. David Lehninger Nelson; Michael M. Cox.
Bioquímica. Jeremy M. Berg; Lubert Stryer; John Tymoczko.
http://www.itescam.edu.mx/principal/sylabus/fpdb/recursos/r80485.PDF