two pore types in the inner-wall endothelium of schlemm's canal
TRANSCRIPT

Two Pore Types in the Inner-Wall Endothelium ofSchlemm's Canal
C. Ross Ethier,1 Fides M. Coloma,1 Arthur J. Sit,2 and Mark Johnson2
PURPOSE. It has been reported that fixation conditions significantly influence the apparent poredensity in the inner-wall endothelium of Schlemm's canal. In the present study, the manner inwhich fixation conditions affect the two subtypes of inner-wall pores, intracellular pores andintercellular (or border) pores, was investigated.
METHODS. Outflow facility was measured in enucleated human eyes. Eyes were fixed under"constant flow" or constant pressure conditions, microdissected to expose the inner wall ofSchlemm's canal, and prepared for scanning electron microscopy. The density and diameter of thetwo subtypes of pores in the inner wall were measured.
RESULTS. Intracellular pore density decreased with increasing postmortem time (P < 0.001) andincreased with increasing volume of fixative passed through the outflow pathway (P < 0.001),whereas border pore density showed no dependence on these parameters (P > 0.25 and P > 0.15,respectively). Border pore density increased with increasing fixation pressure (P < 0.005), eventhough intracellular pore density showed no such dependence (P > 0.4). No correlation was foundbetween outflow facility and the predictions of Poiseuille's law, Sampson's law, or the funnelingtheory for the hydraulic conductivity of the intracellular pores (P > 0.35) or the border pores (P >0.1).
CONCLUSIONS. The intracellular and border pores form two morphologically and functionally distinctpopulations in the inner wall of Schlemm's canal. The dependence of intracellular pore density onpostmortem time and on volume of fixative passed through the outflow pathway suggests thatthese pores are artifacts of tissue fixation or processing conditions. That border pores do notdepend on such conditions and that their presence is correlative with perfusion pressure suggeststhat this population may be nonartifactual. New histologic techniques for examining the inner wallof Schlemm's canal are necessary to determine the in vivo state of inner-wall pores and how theyinfluence outflow facility. (Invest Ophthalmol Vis Set. 1998;39:204l-2048)
The cause of the elevated aqueous humor outflow resis-tance characteristic of primary open-angle glaucoma1 isunknown. Among the difficulties in localizing the patho-
logic condition responsible for outflow obstruction is the in-ability to determine the locus of outflow resistance, even inhealthy eyes. An early2 candidate site for generating aqueousoutflow resistance has been the inner wall of Schlemm's canal.When eyes are fixed at pressure, it has been observed that the
From the 'Department of Mechanical Engineering and Ophthal-mology, University of Toronto, Canada; and the 2Department of Me-chanical Engineering, Massachusetts Institute of Technology, Cam-bridge.
Supported in part by Grant EY09699 from the National Institutesof Health, Bethesda, Maryland; Grant MA-10051 from the MedicalResearch Council of Canada, Ottawa, Ontario, Canada, and the NaturalSciences and Engineering Research Council of Canada, Ottawa, On-tario, Canada (partial support for AJS).
Presented in part at the annual meeting of the Association ofResearch in Vision and Ophthalmology, Fort Lauderdale, Florida, May1997.
Submitted for publication February 12, 1997; revised June 20,1997; accepted July 9, 1998. The delay in publication was caused by anoversight during the editorial transition.
Proprietary interest category: N.Reprint requests: C. Ross Ethier, Department of Mechanical and
Industrial Engineering, University of Toronto, 5 King's College Road,Toronto, Ontario, M5S 3G8 Canada.
inner wall of Schlemm's canal contains giant vacuoles protrud-ing from the endothelial layer into the lumen of Schlemm'scanal.3"5 Associated with these giant vacuoles but also occur-ring in other areas6 are pores passing through the inner wall.However, the pioneering hydrodynamic studies of Bill andSvedbergh7 and confirmation of their results by other investi-gators8'9 showed that the inner-wall pores could be responsi-ble for, at most, a modest fraction of total outflow resistance.Simply put, there were just too many holes in the inner-wallendothelium.
Since that time, investigations have focused on the possi-bility that the juxtacanalicular connective tissue, and in partic-ular, its extracellular matrix, generates the bulk of aqueousoutflow resistance.10"12 Experimental evidence has confirmedthat a significant fraction of outflow resistance is generated bythe juxtacanalicular connective tissue.13 However, this resultleaves unexplained the experimental observations14'5 that anincrease in the pore density of the inner-wall endothelium isassociated with an increase of outflow facility and that returnof the pore density to its baseline value is associated with rapidrestoration of outflow facility.
Using perfusion studies in combination with scanningelectron microscopy (EM), we have recently found that thedensity of pores in the inner-wall endothelium depends onfixation conditions.16 Specifically, in fresh human eyes (per-fused less than 20 hours after death) total pore density in-
Investigative Ophthalmology & Visual Science, October 1998, Vol. 39, No. 11Copyright © Association for Research in Vision and Ophthalmology 2041

2042 Ethler et aJL IOVS, October 1998, Vol. 39, No. 11
creased with the total volume of fixative that passed throughthe inner wall. This suggests that the in vivo state of theinner-wall pores may differ from that seen in fixed specimensand thus potentially reopens questions about the role (if any)that the inner wall plays in influencing aqueous outflow resis-tance.
During the course of the previous study l6, we observedtwo distinct pore types in the inner-wall endothelium ofSchlemm's canal: pores passing through cells (intracellularpores) and pores passing between cells (intercellular or "bor-der" pores). The latter type of pore is consistent with obser-vations of a paracellular pathway for aqueous humor drainage,described by Epstein and Rohen.17
The goal of the present study was to characterize morecompletely these two types of pores in the inner-wall endothe-lium. In particular, in view of our previous work showingfixation-induced changes in pore density, we sought to deter-mine what effect perfusion and fixation conditions had on eachtype of pore. This was achieved by examining the inner-wallendothelium of eyes perfused and fixed under a variety ofconditions, including constant-flow fixation.
METHODS
Most of the experimental methods used in the present studywere identical with those previously reported,16 and thereforewill be summarized only briefly here. The same two series ofpostmortem normal human eyes were used in the presentstudy as were used in our previous work. The measured pa-rameters for each eye are defined in Table 1 and summarized inTable 2.
Eyes were excluded if donors had had ophthalmic surgery,diabetes, or a history of ocular disease. In series A, 21 eyes (10pairs plus a single eye) were perfused with Dulbecco's phos-phate-buffered saline (Life Technologies, Grand Island, NY)with 5.5 mM glucose added at a constant flow rate of 2 /xl/min,and were then subjected to anterior chamber exchange andfixation under constant-flow conditions. More specifically, an-terior chamber exchange and fixation using modified Kar-novsky's fluid (2.5% glutaraldehyde and 2% paraformaldehydein Sorenson's phosphate buffer [pH 7.3]) was carefully per-formed at the constant pressure recorded at the end of theconventional perfusion, that is, just before anterior chamberexchange. This produced approximately equal fixation flowrates (but different pressures) in all eyes. In series B, 13 eyeswere perfused at a constant pressure of 8 mm Hg and werethen subjected to anterior chamber exchange. In 8 of the 13eyes, subsequent constant pressure fixation was performed at8 mm Hg; in the remaining 5 eyes, the fixations were per-formed at an elevated pressure that was constant during thefixation procedure (range, 9-46 mm Hg).
At the conclusion of fixation, the inner wall of Schlemm'scanal was prepared for scanning EM, and pore densities anddiameters were analyzed using previously described meth-ods.16 Briefly, probable pores were identified from montages,and the original samples were then rescanned at magnificationX 10,000 to verify that each putative pore was nonartifactual.Openings in the endothelium were identified as artifactualtears and disregarded if the edges were rough, torn, irregular,or notched. Border pores were defined to be those poresobserved by scanning EM to intersect with a cell margin,
TABLE 1. Nomenclature for Parameters Measured in thePresent Study
Measured Quantity Symbol Units
Patient agePostmortem time to the
start of the experimentTime between patient
death and enucleationIntracellular pore density of
the inner wallendothelium
Border pore densityAverage intracellular pore
diameterAverage border pore
diameterOutflow facility before
fixationOutflow facility after
fixation (series A only)Fixation pressureFlow rate of buffer through
outflow pathway beforefixation
Volume of buffer perfusedthrough outflow systembefore fixation
Volume of fixative perfusedthrough outflow system(series A only)
Time that fixative wasperfused through theoutflow system
Age
PM
E
* i
A
c
IOP
years
hours
hours
pores/mm2
pores/mm2
jam
jam
/xl/min/mm Hg
jui/min/mm Hgmm Hg
Q jLtl/min
ml
ml
mm
whereas intracellular pores were those pores that did notintersect a cell margin. A small percentage of the pores (2%-3%) could not be definitively classified and were not includedin either category. A typical image of the inner-wall endothe-lium with the two pore types identified (intracellular andborder) is shown in (Figure 1); note the irregular margins ofthe artifactual pore.
Because pores are typically elliptical, the major and minoraxes were measured directly from the screen of the micro-scope at X 10,000. Length measurements were calibrated usingtwo scanning EM calibration grids (SIRA Institute, Chislehurst,England) with densities of 19.7 lines/mm and 2160 lines/mm.Because circular pores will appear elliptical when viewed intilted section, the pore diameter was assumed to be equal tothe major axis of the pore; however, all results reported in thisarticle were also verified using the geometric mean of themajor and minor axes as the pore diameter.
Statistical Methods and Quantitative Analysis
The measured parameters for each eye included patient age(Age: in years), postmortem time to the start of the experiment(PM: in hours), time between patient death and enucleation (E:hours), intracellular pore density of the inner-wall endothelium(N{. pores per square millimeter), border pore density (NB:pores per square millimeter), average intracellular pore diam-eter (£>{. in micrometers), average border pore diameter (Z)B: inmicrometers), outflow facility before fixation (C: in microliters

IOVS, October 1998, Vol. 39, No. 11 Two Pore Types in the Inner Wall of Schlemm's Canal 2043
TABLE 2. Summary of Experimental Data for Each Eye
Eye
Q5Q6Q7Q8Q9Q10Q13Q14Q15Q16Q17Q18Q19Q20Q21Q23Q24Q25Q26Q27Q28258262264270274276242244246250251268282
Age(y)
868671715597976262939365655085859191757585797574858786779081858476
PM(h)
14.3314.3325.8325.8324.7524.7513.4313.4334.334.317.817.8
8.338.3327.9826.9826.9810.5810.58
20.2520.2525342723251731272527131918
IOP (mmHg)
434111.89.42.93.118.6
25.510.2
10.528.721.410.2
8.33.54.47.517.2
21.915.19.494625827198888888
C(/Lil/min/mm Hg)
0.0500.0500.1700.2100.7000.6500.1100.080
0.1950.1900.0700.0900.2000.2400.640
0.4250.268
0.1150.0900.1300.2100.340.070.150.260.080.120.180.050.170.14
0.190.170.12
cF(fd/min/mm Hg)
0.0250.0300.0800.1100.4000.7500.050
0.0450.0650.120
0.0450.0530.0800.110
0.2750.1310.0750.0350.0350.0800.070
N
I
753.61255.21060.022636
1236.51063.12234.8893.0
259.5161.7620.2
1210.92166.81698.6
55.3136.4
798.91458.41612.1
750.5410.1834.1
465.31277.2456.2660.9149.3724.6105.1883.4
800.9857.5473.7838.4
(pores/mm2)
B
613.3614.0
394.5620.1
1305.5478.4622.8
284.9363.3452.9772.8341.0493.0369.2118.260.2
130.3234.9204.3205.6
377.3419.4487.6
470.3199.5360.468.6140.136.2177.0274.2283.2163.6138.6
Total
1417.82014.21485.4
2935.72542.01562.8
2958.31275-1622.9614.61446.6
156312670.82091.4
173.5211.5960.21704.81838.0961.2
791.51279.6988.61804.1686.81042.7230.8864.7149.8
1060.31105.11140.8
643.9990.0
I
0.951.340.81
1.130.820.841.020.811.361.080.870.74
0.790.97
1.631.210.881.24
1.290.920.96
0.951.020.740.870.911.100.740.541.210.780.700.87
0.99
2>0un)
B
1.691.72
1.391.851.391.441.701.54
1.451.441.241.481.141.582.362.64
1.671.711.521.111.641.761.60
1.891.451.881.98
1.591.96
2.531.181.171.571.67
Total
1.311.52
0.971.321.111.021.20
1.031.411.34
1.090.910.861.08
2.131.651.031.311.330.971.281.241.34
1.051.081.271.360.870.90
1.430.890.821.061.10
T(min)
11511533833846013428393982586224027093706506181180181753451201506012010060606060906013090
(ml)
0.1150.1380.318
0.3550.5240.3100.258
0.1050.066
0.3250.08
0.2830.2830.098
0.7250.3630.114
0.1490.170
0.0910.272
See Table 1 for definition of symbols. Blank cells indicate that no data were measured.
per minute per millimeter of mercury), outflow facility afterfixation (CF: microliters per minute per millimeter of mercury;series A only), fixation pressure (IOP: in millimeters of mer-cury), flow rate of buffer through outflow pathway beforefixation (Q: in microliters per minute), volume of buffer per-fused through outflow system before fixation (FB: in millili-ters), volume of fixative perfused through outflow system (FF:in milliliters; series A only), and time that fixative was perfusedthrough the outflow system (7": in minutes) (Table 2).
We also included one derived parameter in the data anal-yses. Because we did not measure VF in series B, we denned anew parameter VF' (in milliliters) = 0.5 • Q • T, which is anestimate of the volume of fixative perfused through the out-flow system based on the observation that outflow facilityrapidly decreases by 50% during fixation16. Because VF, VF',and T were closely related and thus not statistically indepen-dent, we used only one of these parameters in any regressionanalysis. If one of these parameters yielded a statistically sig-nificant correlation, all were investigated to determine whichproduced the lowest P.
We first performed linear regression analyses relating out-flow facility to pore density and pore density to fixation flow
rate and fixation pressure. We then performed multivariateregression analyses (Systat for Macintosh, version 5.2.1; SPSS,Chicago, IL) examining the correlation of three of the variables(outflow facility, pore density, and pore diameter) with theremaining variables measured in series A and B. Variables thatwere not plausibly connected to the dependent variable werenot included in the correlation (e.g., fixation time could notaffect prefixation outflow facility), nor were variables thatcould be directly used to calculate the dependent variable(e.g., flow rate and pressure were not both included in corre-lations for outflow facility). We used an automated forwardstepwise regression as implemented in the software (Systat)using a tolerance (overall P) of 0.01 and P for adding variablesto the correlation of 0.01. In this scheme, at each step of theiteration, the most statistically significant variable not yet in-cluded in the correlation was added to the correlation whileensuring that the tolerance did not exceed 0.01. The moststatistically significant variable was chosen by computingwhich of the remaining variables not yet added to the correla-tion would have the lowest individual P (constrained to be lessthan 0.01) if it were added to the correlation. When no furthervariables could be added without having the tolerance exceed

2044 Ethier et al. IOVS, October 1998, Vol. 39, No. 11
FIGURE 1. Scanning electron photomicrographs of pores in inner-wall endothelium. (A) Overview of section of inner wall. (B)High-power view of intercellular or border pore (B) from boxed region in (A). (C) High-power view of intracellular pore I andartifactual pore A. Arrows in (B) refer to cellular margins. Note that the (B) pore is intersected by a cellular margin on each edge.An overview micrograph of the region that contains (C) was published in reference 16. Scale bar, (A) 10 /xm; (B, C) 1 ̂ ,m.
IOVS, October 1998, Vol. 39, No. 11 Two Pore Types in the Inner Wall of Schlemm's Canal 2045
2500 n
2000-
1500-
5 1000Q
£ 500-
8 ooo
• ' 19 •' 1 '
10 20 30Post Mortem Time (hours)
40
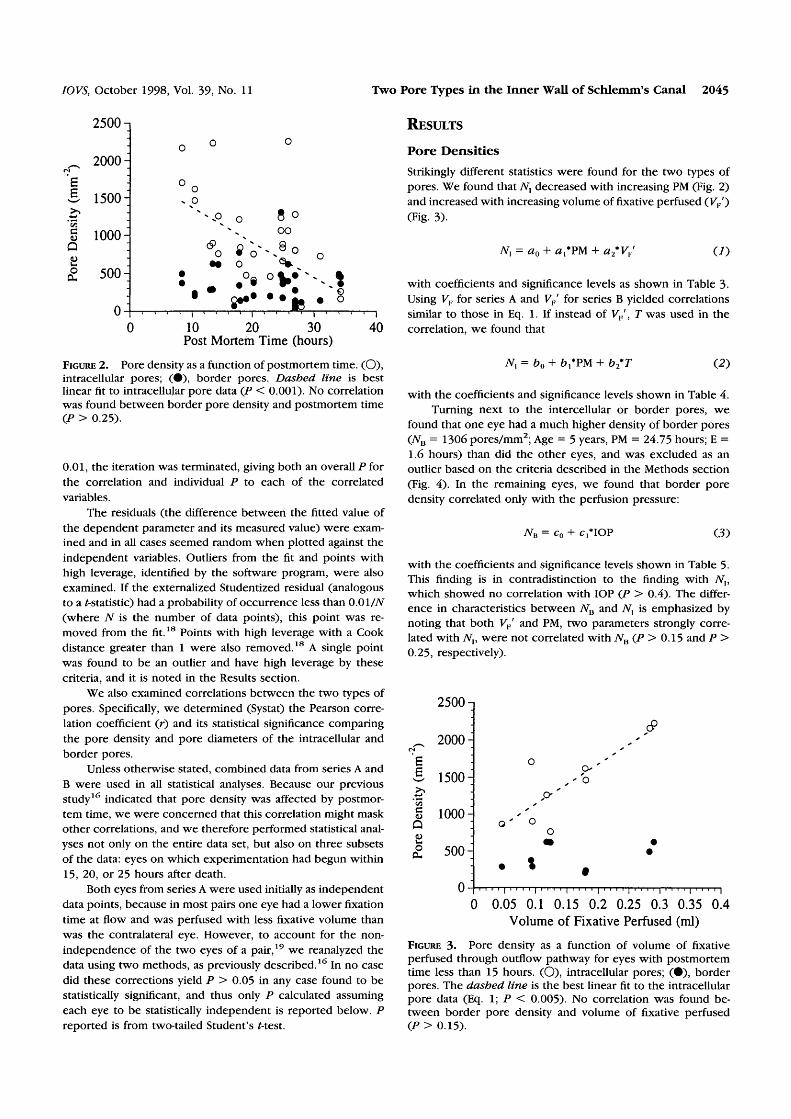
FIGURE 2. Pore density as a function of postmortem time. (O),intracellular pores; (•), border pores. Dashed line is bestlinear fit to intracellular pore data (P < 0.001). No correlationwas found between border pore density and postmortem time(P > 0.25).
0.01, the iteration was terminated, giving both an overall P forthe correlation and individual P to each of the correlatedvariables.
The residuals (the difference between the fitted value ofthe dependent parameter and its measured value) were exam-ined and in all cases seemed random when plotted against theindependent variables. Outliers from the fit and points withhigh leverage, identified by the software program, were alsoexamined. If the externalized Studentized residual (analogousto a ̂ -statistic) had a probability of occurrence less than 0.0\/N(where N is the number of data points), this point was re-moved from the fit.18 Points with high leverage with a Cookdistance greater than 1 were also removed.18 A single pointwas found to be an outlier and have high leverage by thesecriteria, and it is noted in the Results section.
We also examined correlations between the two types ofpores. Specifically, we determined (Systat) the Pearson corre-lation coefficient (f) and its statistical significance comparingthe pore density and pore diameters of the intracellular andborder pores.
Unless otherwise stated, combined data from series A andB were used in all statistical analyses. Because our previousstudy16 indicated that pore density was affected by postmor-tem time, we were concerned that this correlation might maskother correlations, and we therefore performed statistical anal-yses not only on the entire data set, but also on three subsetsof the data: eyes on which experimentation had begun within15, 20, or 25 hours after death.
Both eyes from series A were used initially as independentdata points, because in most pairs one eye had a lower fixationtime at flow and was perfused with less fixative volume thanwas the contralateral eye. However, to account for the non-independence of the two eyes of a pair,19 we reanalyzed thedata using two methods, as previously described.16 In no casedid these corrections yield P > 0.05 in any case found to bestatistically significant, and thus only P calculated assumingeach eye to be statistically independent is reported below. Preported is from two-tailed Student's £-test.
RESULTS
Pore Densities
Strikingly different statistics were found for the two types ofpores. We found that iV, decreased with increasing PM (Fig. 2)and increased with increasing volume of fixative perfused (F,/)(Fig. 3).
TV, = a0 + #,*PM + a2*VF' (/)
with coefficients and significance levels as shown in Table 3-Using VF for series A and VP' for series B yielded correlationssimilar to those in Eq. 1. If instead of VF , T was used in thecorrelation, we found that
TV, = b0 + + b2*T (2)
with the coefficients and significance levels shown in Table 4.Turning next to the intercellular or border pores, we
found that one eye had a much higher density of border pores(NB = 1306 pores/mm2; Age = 5 years, PM = 24.75 hours; E =1.6 hours) than did the other eyes, and was excluded as anoutlier based on the criteria described in die Methods section(Fig. 4). In the remaining eyes, we found that border poredensity correlated only with the perfusion pressure:
7VB = c0 + c,*IOP (3)
with the coefficients and significance levels shown in Table 5.This finding is in contradistinction to the finding with TV,,which showed no correlation with IOP (P > 0.4). The differ-ence in characteristics between 7VB and JV, is emphasized bynoting that both VF' and PM, two parameters strongly corre-lated with Nv were not correlated with 7VB (P > 0.15 and P >0.25, respectively).
cQ
2000:
1500:•
1000:
500:
0-
o
• *
. . . . 1 . . . . 1
•
o
9*''- b
fn i | i n
•
0 0.05 0.1 0.15 0.2 0.25 0.3 0.35 0.4Volume of Fixative Perfused (ml)
FIGURE 3- Pore density as a function of volume of fixativeperfused through outflow pathway for eyes with postmortemtime less than 15 hours. (O), intracellular pores; (•), borderpores. The dashed line is the best linear fit to the intracellularpore data (Eq. 1; P < 0.005). No correlation was found be-tween border pore density and volume of fixative perfused(P > 0.15).

2046 Ethier et al.
TABLE 3. Coefficients
PostmortemTime (h)
All data<25 hours<20 hours<15 hours
for
n
3419149
Equation 1
ao
1830 ±1870 ±1710 ±580 ±
270320400230
a1
-43.2 (P < 0.001)-75.9 (P< 0.001)-82.9 (P < 0.005)
NS (P = 0.078)
IOVS, October 1998,
NS (P = 0.44)2630 (P < 0.005)4450 (P < 0.001)5450 (P < 0.005)
Vol. 39, No. 11
P*
<0.001<0.001<0.00005<0.005
NS, not significant at P — 0.01. Values for a0 are means ± SE and means for «, and a2, with statistical significance in parentheses.* Probability for the overall correlation.
We also examined the Pearson correlation coefficient (r)between Nt and 7VB. For the data set as a whole (n = 33; outlierfrom Nu population excluded), the two types of pore densitiescorrelated with each other (r = 0.47; P < 0.01). For the datasubsets (PM < 15, 20, or 25 hours), no statistically significantcorrelation was found (P > 0.15).
No statistically significant correlations were found be-tween age and Nx (P > 0.07) or 7VB (P > 0.04, outlier exclud-ed), although P approached statistical significance. When datafrom both 5-year-old eyes were excluded, P was much higher(P > 0.14 and P > 0.2, respectively). Figure 5 shows poredensity as a function of age for the adult eyes.
Pore SizesWith the major axis of the pore as the pore diameter, Dx was0.97 ± 0.23 ixm (n = 34; mean ± SD) and Da was 1.64 ± 0.36/xm (n = 34); with the geometric mean of the major and minoraxes of the pore as the pore diameter, Dl was 0.75 ±0.19 ju-m(n = 34) and £>B was 1.13 ± 0.26 /Ltm (n = 34). We alsoexamined the correlation between the diameter of the poresand the other variables. The only statistically significant corre-lation was between the pore size of the intracellular pores andthat of the border pores (whole data set: r = 0.48, P < 0.005,n = 34; PM < 25 hours: r = 0.59, P < 0.01, n = 19).
Outflow Facility
Prefixation C correlated with patient age and with fixationpressure, as previously reported.16 Similarly, outflow facilityafter fixation correlated with patient age. Including the poredensity and pore diameters of the intracellular pores and bor-der pores with these correlations showed one further correla-tion:
C = d0 d2*Age
with the coefficients shown in Table 6. Da in Equation 4 isbased on the pore major axis; a similar, but statistically less
significant result (P < 0.02 for d3), was found using the meanpore diameter (see Methods section). A statistically significantcorrelation was found only for the data set as a whole, and nosuch correlation (involving pore densities or diameters) wasfound for the data subsets involving shorter PM (P > 0.5).
Several theories have been proposed to relate the numberand size of the pores of the inner-wall endothelium to the flowresistance generated by this cell layer. These include Poi-seuille's law,7 Sampson's law,7 and the theory of funneling,20
which predict a scaling relationship of C with pore numberand size as N • D4, N • D3, and TV • D, respectively. Whenexcluding the single outlier for Dn (see earlier explanation), nocorrelation was found between C and any of these combina-tions in intracellular pores (P > 0.4) or in border pores(P > 0.1).
DISCUSSION
Pores passing through the inner-wall endothelium ofSchlemm's canal were first identified by Holmberg.4 Thesepores usually were associated with giant vacuoles but alsowere seen occasionally in flat regions of the inner wall.6 Usingscanning EM (Fig. 1), we have found that the inner-wall endo-thelium has two distinct types of pores: an intracellular porepopulation (pores passing through the inner wall endothelialcells) and an intercellular (or border) pore population (porespassing between the cells). In primate eyes, Epstein and Ro-hen17 identified a pathway between inner-wall endothelialcells through which an undetermined fraction of aqueous hu-mor passes, and we postulate that the intercellular pores seenin our study corresponded to the luminal opening of theparacellular pathway identified by Epstein and Rohen.
The major conclusion of the present study is that the twopore types show markedly different dependencies on impor-tant experimental parameters, such as IOP, Vl7, and PM. Inparticular, we found that the intracellular pore density was
TABLE 4. Coefficients for Equation 2
PostmortemTime (h)
All data<25 hours<20 hours<15 hours
341914
9
1830 ± 2701870 ± 3201750 ± 350540 ± 230
-43.2 (P < 0.001)-77.1 (P< 0.001)-91.1 (P< 0.001)
NS (P = 0.090)
NS (P = 0.48)2.72 (P < 0.01)4.80 (P < 0.0005)5.77 (P < 0.005)
<0.001<0.001<0.00002<0.005
NS, not significant at P = 0.01. Values for b0 are means ± SE and means for bl and b2, with statistical significance in parentheses.• Probability for the overall correlation.

IOVS, October 1998, Vol. 39, No. 11 Two Pore Types in the Inner Wall of Schlemm's Canal 2047
cQ
&
1600n
1400:
1200:
1000:
800^
600:
400:
200:
0-0 10 20 30
IOP (mm Hg)
40 50
FIGURE 4. Pore density as a function of fixation pressure. (O),intracellular pores; (•), border pores. The solid line is the bestlinear fit to the border pore data (P < 0.005). No correlationwas found between intracellular pore density and fixationpressure (P > 0.4).
observed to decrease with PM and to increase with the Vp (Eq.1); in contrast, NB did not depend on these parameters, butinstead increased with IOP (Eq. 3). Thus, these two types ofpores seem to be functionally and morphologically distinct.
The dependence of TV, on PM and total VF and the absenceof dependence on flow-related variables such as IOP and VF
suggests that intracellular pores may be fixation artifacts. Sim-ilar logic may suggest that border pores are not fixation arti-facts. However, at present, it is impossible to conclude defin-itively that either one (or both) pore types are artifacts. Ifintracellular pores were artifacts not present in vivo, a calcu-lation using the methodology of Bill and Svedbergh7 indicatesthat border pores would generate most of the known aqueousoutflow resistance. Clearly, determination of the actual in vivopore distribution in the inner wall of Schlemm's canal is re-quired.
How PM and VF affect intracellular pores and how fixationpressure affects border pores is unclear. We have previouslyhypothesized16 that the effect of PM on pore density may becaused by loss of inner-wall endothelial cell viability, thusdecreasing the cellular capability to form pores. The effect ofVF was attributed to pores continuing to form during thefixation process; thus, rather than a "snapshot" of the innerwall, a "time exposure" image was seen. The increase in DB
with increasing IOP may have been caused by increased stress
TABLE 5. Coefficients for Equation 3
8.64 (P < 0.005)NS (P = 0.035)NS (P = 0.031)NS(P= 0.164)
NS, not significant at P = 0.01. Values for d0 are means ± SE andmeans for d{ and d2, with statistical significance for the overall corre-lation in parentheses.
PostmortemTime (h)
All data<25 hours<20 hours<15 hours
n
3318149
Co
200 ± 50360 ± 50370 ± 60410 ± 60
£
lsit
y
<uQ
Pore
1600-
1400 T
1200 T
1000-
800^
600-
400^
200 T
0-
oo
°*
40 50 60 70 80 90 100
Age (years)
FIGURE 5- Pore density as a function of age for adult eyes. (O),intracellular pores; (•), border pores. No correlation wasfound between age and intracellular pore density (P > 0.2) orborder pore density (P > 0.14).
on the intercellular junctions, promoting pore formation. Toinvestigate these hypotheses, improved methods of studyingthe inner wall may be required. It is interesting that poredensity and the pore diameter of the intercellular and borderpores correlated with each other (at least for the longer PM),which suggests that there may be some interaction betweenthe pore types. However, the nature of this interaction (if any)is unclear at present.
We have previously proposed20 that a hydrodynamic in-teraction between the pores in the inner-wall endothelium andthe juxtacanalicular connective tissue could lead to an en-hanced effective flow resistance of the juxtacanalicular region(the funneling effect). Other theories of how inner-wall poresmay influence outflow facility have also been proposed: shortpores passing through an otherwise impermeable inner wall7
(inner-wall flow resistance given by Sampson's law), and longpores7 (inner-wall flow resistance given by Poiseuille's law).Each of these theories predicts relationships between outflowfacility and inner-wall pore characteristics.
However, when we previously used data for all pores,16
we did not find support for any of these predicted relation-ships. The present study also found no such correlations,considering either the intracellular pores or for the borderpores individually. In the case of Sampson's law or Poiseuille'slaw, in which the pores are themselves entirely responsible forgenerating flow resistance, this absence of correlation arguesstrongly against inner-wall pores playing a major role in deter-mining flow resistance. However, in the case of the funnelingtheory, such a conclusion is less clear cut, because aqueousoutflow resistance in that case is primarily determined by thehydraulic conductivity of the extracellular matrix in the juxta-canalicular connective tissue, and pores act only to modulateflow resistance in the juxtacanalicular connective tissue. Forthis reason, a strong statistical correlation between outflowfacility and inner-wall pore characteristics may be difficult toobtain, without also determining the hydraulic conductivity ofthe juxtacanalicular connective tissue.
In this discussion, we have implicitly assumed that thepores are short straight-through openings in the inner wall ofSchlemm's canal. Under high magnification, it is possible to see

2048 Ethler et al.
TABLE 6. Coefficients for Equation 4
IOVS, October 1998, Vol. 39, No. 11
PostmortemTime (h)
All data
n
34 0.502
do
± 0.076
i
-0.00396 (P< 0.01)
d2
-0.00585 (P< io-8) 0. 124
d,
0.005)
P*
<10~9
Values for d0 is means ± SE and means for dlt d2, and rf3, with statistical significance in parentheses.* Probability for the overall correlation.
an amorphous porous material through the mouth of some ofthe larger pores. Such material is probably juxtacanaliculartissue, and this would seem to support the assumption that thepores are short passages. However, in their study of paracellu-lar pores, Epstein and Rohen17 noted that some intercellularpores can be long and tortuous, eventually enlarging at theiropening to the lumen of Schlemm's canal. If our border porescorrespond to these paracellular pathways, it is possible thatthe structure that we see as a pore in the inner-wall endothe-lium may not entirely reflect the geometry and flow resistanceof the underlying flow channel. If this were the case, it wouldrequire further rethinking of our concept of a "pore" in thecontext of Schlemm's canal, and could explain the absence ofcorrelation between outflow facility and the predictions of thehydrodynamic models investigated.
A current controversy regarding microvascular hydraulicpermeability also involves two types of endothelial pores (or"gaps" in their terminology): those passing between the cellsand those passing through the cells. Although it is widelybelieved that microvascular permeability is controlled by theintercellular pathway, Neal and Michel21 have shown thatwhen an ionophore is used to increase microvascular perme-ability, this increase is associated with an increased number ofintracellular pores, with little or no change in the intercellularpathway. These changes in permeability and pore numberwere reversed by perfusion with ionophore-free solutions. It ispossible that the two types of pores we have investigated inthe inner-wall endothelium of Schlemm's canal are related tothose involved in fluid transport across the vascular endothe-lium.
Summarizing, we have found two types of pores in theinner-wall endothelium that correlate very differently with fix-ation parameters. Further study, probably using new tech-niques that show better the in vivo state of the inner-wall ofSchlemm's canal, is needed to determine what role these twotypes of pores play in influencing outflow facility.
References
1. Grant WM. Clinical measurements of aqueous outflow. Arch Oph-thalmol. 1951;46:113-131.
2. Seidel E. Experimented Untersuchungen uber die Quelle und denVerlauf der intracoluaren Saftstromung. Graefes Arch Clin ExpOphtbalmol. 1918;95:1.
3. Garron LK, Fenney ML, Hogan MJ McEwen WK. Electron micro-scopic studies of the human eye. Am J Ophthalmol. 1958;40:27-35.
4. Holmberg A. The fine structure of the inner wall of Schlemm'scanal. Arch Ophthalmol. 1959;62:956-958.
5. Tripathi RC. Ultrastructure of Schlemm's canal in relation to aque-ous outflow. Exp Eye Res. 1968;7:335-34l.
6. Inomata H, Bill A, Smelser GK. Aqueous humor pathwaysthrough the trabecular meshwork and into Schlemm's canal incynomologus monkey (Macca Irus). Am J Ophthalmol. 1972;73:760-789.
7. Bill A, Svedbergh B. Scanning electron microscopic studies of thetrabecular meshwork and the canal of Schlemm: an attempt tolocalize the main resistance to outflow of aqueous humor in man.Acta Ophthalmol. 1972;5O:295-32O.
8. Moseley H, Grierson I, Lee WR. Mathematical modelling of aque-ous humor outflow from the eye through the pores in the lining ofSchlemm's canal. Clin Phys Physiol Meas. 1983;4:47-63.
9. Grierson I, Lee WR, Mosley H, Abraham S. The trabecular wall ofSchlemm's canal: a study of the effects of pilocarpine by scanningelectron microscopy. Br J Ophthalmol. 1979;63:9-l6.
10. Ethier CR, Kamm RD, Palaszewski BA, Johnson MC, RichardsonTM. Calculations of flow resistance in the juxtacanalicular mesh-work. Invest Ophthalmol Vis Sci. 1986;27:1741-1750.
11. Alexander JP, Samples JR, Van Burkirk EM, Acott TS. Expression ofmatrix metalloproteinases and inhibitor by human trabecularmeshwork. Invest Ophthalmol Vis Sci. 1991;32:172-180.
12. Seiler T, Wollensak J. The resistance of the trabecular meshwork toaqueous humor outflow. Graefes Arch Clin Exp Ophthalmol.1985;223:88-91.
13. Maepea O, Bill A. Pressures in the juxtacanalicular tissue andSchlemm's canal in monkeys. Exp Eye Res. 1992;54:879-883.
14. Hamanaka T, Bill A. Morphological and functional effects ofNa2EDTA on the outflow routes for aqueous humor in monkeys.Exp Eye Res. 1987;44:171-190.
15. Hamanaka T, Bill A. Effects of alpha-chymotrypsin on the outflowroutes for aqueous humor. Exp Eye Res. 1988;46:323-34l.
16. Sit AJ, Coloma FM, Ethier CR, Johnson M. Factors affecting thepores of the inner-wall endothelium of Schlemm's canal. InvestOphthalmol Vis Sci. 1997;38:1517-1525.
17. Epstein DL, Rohen JW. Morphology of the trabecular meshworkand inner-wall endothelium after cationized ferritin perfusion inthe monkey eye. Invest Ophthalmol Vis Sci. 1991 ;32:160-171.
18. Weisberg S. Applied Linear Regression. New York: John Wiley &Sons; 1985:114-121.
19. Glynn RJ, Rosner B. Accounting for the correlation betweenfellow eyes in regression analysis. Arch Ophthalmol. 1992;110:381-387.
20. Johnson M, Shapiro A, Ethier CR, Kamm RD. Modulation of out-flow resistance by the pores of the inner-wall endothelium. InvestOphthalmol Vis Sci. 1992;33:1670 -1675.
21. Neal CR, Michel CC. Trancellular gaps in microvascular walls offrogs and rat when permeability is increased by perfusion with theionophore A23187. J Physiol. 1995;488:427-437.