the soluble antigens of clostridium œdematiens type d (cl. hæmolyticum)
TRANSCRIPT

THE SOLUBLE ANTIGENS OF CLOSTRIDI U H QCDEMATIENS TYPE D (CL. H&MOLYTICUM)
C. L. OAKLEV and G. HARRIET WARRACK Bacteriology Department, University of Leeds, and Wellcome
Research Laboratories, Beckenham, Kent
VAWTER and Records (1925-26) isolated an obligate anaerobe bacillus from cases of infectious icterohzmoglobinuria (red water disease) of cattle in the United States ; to judge from their report, the organism was readily distinguishable from Clostridizcm welchii, (21. septicuiri and Cl. chauvai, less readily from Cl. edematiens.
Cultures were lethal to rabbits, guinea-pigs and cattle by intramuscular injection ; the disease produced in cattle closely resembled naturally occurring icterohamoglobinuria. Rabbits could be protected from the lethal effect of intramuscularly injected culture by injection, 16 hr before, of homologous antiserum ; Cl. welchii, C1. septicum, Cl. chauvcei and Cl. cedematiens (presumably type A) antisera were ineffective.
Supernatants from centrifuged 8-20-hr cultures were lethal to rabbits and guinea-pigs by intramuscular or intravenous injection ; supernatants from older cultures were not lethal and produced only “local lesions” on intra- muscular injection. The authors concluded that the pathogenicity of their cultures was due to an unstable hamolytic toxin, and suggested the name Clostridium hcemolyticum bovis for the organism. Hall (1929) suggested that it should be called Bacillus hcemolyticus ; it is now usually called Clostridium hcemolyticum.
Vawter and Records (1931) showed that 15 of 16 strains of Cl. hmmolyticum were antigenically homogeneous in agglutination tests ; the sixteenth was agglutinated by homologous serum only. No strain was agglutinated by sera against GI. welchii, C1. septicurn types I, I1 and 111, GI. chauvmi or Cl. sordellii, and sera against Cl. hcemolyticum failed to agglutinate any of these organisms ; tests against Cl. cedematiens type-A antigens were unsatisfactory. Kreuzer { 1939) found no cross-agglutination between the 0 antigens of single strains of Cl. cedematiens types A and B and Cl. hamolyticum. Turner and Eales (1943), in a comprehensive analysis of the somatic and flagellar antigens of the Cl. cedematiens group, showed that all strains of Cl. odematiens types A, B and C had two somatic antigens in common, one of which was shared with one of the two strains of Cl. hcemolyticum they examined.”
Dernnitz (1934) states that Cl. hamolyticum differs from Cl. novyi (CZ. a d e m t i e n s type A) in not producing “ novytoxin ”, and that its only toxin is .a hemotoxin identical with that of GI. gigas (GI. cedematiens type B).
Weinberg, Nativelle and Pr6vot (1937) state that Sordelli et al. informed them that Cl. welchii antisera protected animals against C1. hmmolyticum infection ; that Cl. hcemolyticum antisera neutralise Cl. welchii “ toxin ” and Cl. cedematiens type-A and B “ toxins ” ; and that Cl. cedematiens type-B, but not type-A, antisera neutralise Cl. hamolyticum toxin-all presumably in lethal tests. Oakley, Warrack and Clarke (1947) showed by assay of antisera that GI. hcemolyticum filtrates contain the p-toxin of Cl. edematiens type B,
* There is, however, considerable doubt that this strain was GI. hcemolyticum (see Smith, 1953).
J. PATH. BACT.-VOL. 78 (1969) 543

544 C. L. O A K L E Y A X D C. HARRIhT’ W A R R A C K
thus confirming the relationship between Cl. hamolyticum and CI. adematiens ; they left the question of classification for future work.
Jasmin (1947) showed that the capacity of Cl. hmmolyticum filtrates to produce death of injected mice, hEmolysis of rabbit red cells, opalescence of human serum and flocculation of egg-yolk emulsions ran parallel ; no serological work was reported.
Bard and McClung (1948) suggested that Gl. kamolyticum filtrates contain, in addition to a ha?molytic lecithinase, a lysolecithin; the argument is that part of the hpmolytic activity is Iost after treatment of the filtrate with a lecithinase-B preparation, which decomposes hwmolytic lysolecithins to non- hzmolytic substances. No quantitative work was reported ; the lecithinase B was a crude preparation and may have contained other active material, e.g. proteinases.
Macfarlane (1950a) showed that Cl. hcenzolyticum filtrates decompose lecithin to phosphorylcholine and a neutral fat, and that this locithinase-C activity is inhibited by Cl. hamolyticum and Cl. cedematiens type-B antisera, but not by Cl. cedematiens type-A or Cl. welchii type-A antisera. The filtrates decomposed sphingomyelin with liberation of water-soluble phosphorus, and there was also evidence for the presence of a lipase. She also found (Macfarlane, 1955) that Cl. edematiens type B and Cl. hmmolyticum, but not C1. ademdiens type A, produce a tropomyosinase (7-antigen).
In this paper we show that the ha~iolytic, necrotising and lethal activities of 01. hemolyticum filtrates are due to the P-lecithinase of Cl. adematiens ; that the filtrates contain, beside the P-lecithinase, another substance (0) produciiig opalescence in egg-yolk emulsions ; and that they contain no detectable Ci. ademutiens a-toxin.
MATERIALS AND METHODS
Sera, filtrates and antisera Strains.
Filtrates.
We had the following strains of Cl. hamolyticum : CN3527-3533 from Dr L. DS. Smith and CN2041 from Dr L. S. McClung.
Strains of Cl. hmmolyticum and Cl. adematiens type B were grown for 24 hr in a papain digest of horse meat containing 10 per cent. saturated sodium sulphate-extract of horse muscle, in 4-litre bottles. The organisins were removed by filtration through Seitz pads ; the filtrates were precipitated by saturation with ammonium sulphate, dried over silica gel, and the dry powder freed from much of the ammonium sulphate by flotation on chloroform. Dried filtrates were dissolved before use in Mj5 sodium acetate-acetic acid buffer pH 6.5 or nutrient broth.
We used 8 Cl. hamolyticum, 6 Cl. edematiens type-B and 4 Cl. edematiens type-A antisera, produced by injecting horses with the appropriate toxic filtrates. Some of the sera were refined by pepsin digestion.
Antisera.
Assay of antisera Antisera were assayed by conventional methods against laboratory standard
antisera, with tho following indicators :-sheep red cells ; horse red cells ; filtered egg-yolk emulsion (lecithovitellin or L.V., Macfarlane, Oakley and Anderson, 1941) ; egg-yolk-agar columns (Oakley et al., 1947) ; guinea-pig skin injected intracutaneously ; mice injected intravenously. For most of the tests serum RX5528 (value 9000 Cl. Qdematiens p-antitoxin units per ml.) was used as standard.
Sheep and horse red cells were about equally affected by Cl. hmmolyticum filtrates ; the hzmolysis has a marked hot-cold character.
Hmmolytic tests.

C L O S T R I D I A L A X T I G E N S 545
900 6900 2700 350
The end-point with sheep cells is rather sharper. Saline suspensions of 6 per cent. sheep or horse red cells were used in the tests. The level of test was 3 p units ; the test-dose contained about 10 indicating doses, and the standard indicating effect was 30 per cent. hzmolysis.
L. 8. tests. Opalescence, flocculation and curd-formation were readily produced in L.V. by Cl. hamolyticum filtrates. The level of test was 3 /3 units, the test-dose contained about 16 indicating doses, and the standard indicating effect was slight opalescence.
TABLE I Comparison of values (units per ml.) obtained for sera in L.V., hamolytic, necrotis-
i n g and lethal tests against Clostridium hzmolyticum jiltrate A'X605 with their C1. cedematiens anti-cc and anti-B ualues
900 6800 2500
270
Serum
9000 1700 3400 4000 5000 3200 4000 3700
Cl. rvdemutiens type-B
EX1117 . . ant isera
9000 1700 3400 3600 5200 3500 4200 3600
RX5325 . T3 . T61 . 2096 . 85CB .
Cl. hcemolyticunz anti- sera
RX552S* . EX1249 . . EX1250 . EX1269 . . EX1287 . EX1292 . EX1321 . EX1325 .
Values against CZ. cedematiens
a-toxin
50 300
7700 3000
900 ...
<0.1 <0.1 <0.1 <0.1 <0.1 <0.1 <0.1 <0.1
&toxin
900 6800 2500 420 670
1600
9000 1800 3400 4250 4300 3400 4800 3450
Values against CZ. hcendytirum filtrate 3 x 6 0 9 in
L.V. tests
480 3700 3200
290 530
1000
9000 1650 2900 4450 5800 4600 6600 3970
hamolytic tests
620 4150 2570
350 3 80
1210
9000 1550 2700 3100 6200 3400 4900 3950
iecrotising I lethal tests j tests
* Standard. Tests carried out from 1949-51.
Egg-agar tests.
Intracutaneous tests.
The test level was 3 /3 units, and the standard indicating effect a slight opalescence well below the upper meniscus of the column.
The lesion produced in guinea-pigs by injection of Cl. hmmolyticum filtrates is a sharply defined yellow or yellowish-green area of necrosis with surrounding erythema and some cedema, well developed in 24 hr ; it closely resembles the lesion provoked by intracutaneous injection of Cl. welchii cc-toxin. The level of test was 50 /3 units (10 p units injected), and the standard indicating effect an area of necrosis c. 5 x 5 mm. a t the final reading of the tests a t 48 hr.
units injected), and the standard indicating effect was death within 48 hr of half the mice injected.
Lethal tests. The level of test was 100 p units (25
RESULTS Intracutaneous and lethal tests
Table I shows that the values of C1. cdematiens tj-pe-B mtisera against Cl. hwmolylicum filtxate NX605 in lethal and intracutaneous
J. PATH. BACP.-VOL. 78 (1959) 2 M
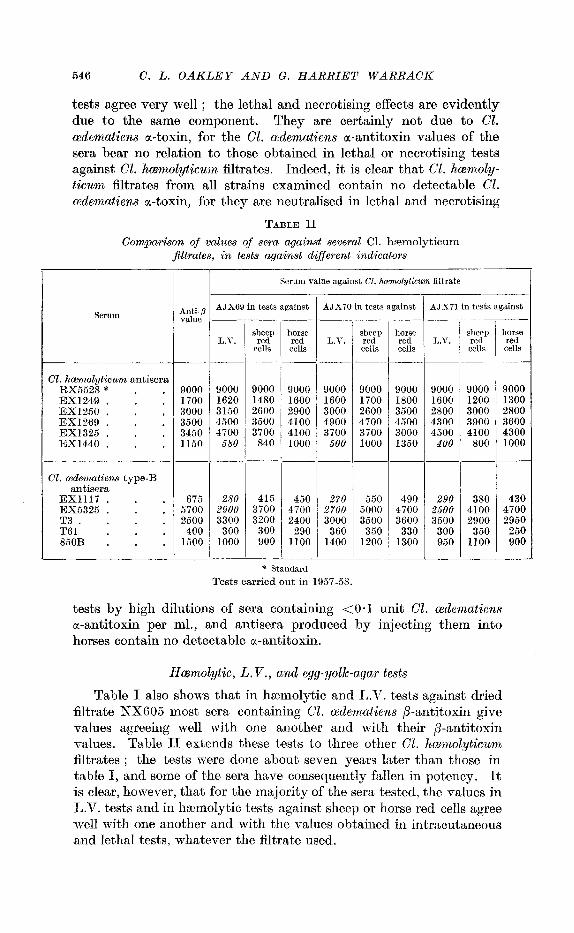
546 C . L. O A K L E Y AND G. H A R R I E T W A R R A C K
L.V.
tests agree very well ; the lethal and necrotising effects are evidently due to the same component. They are certainly not due to Cl. ccdematiens a-toxin, for the Cl. cedematiens a-antitoxin values of the sera bear no relation to those obtained in lethal or necrotising tests against Cl. hcemolyticum filtrates. Indeed, it is clear that C1. hmnzoly- ticunz filtrates from all strains examined contain no detectable Cl. ccdenzatiens a-toxin, for they are neutralised in lethal and necrotising
TABLE I1 Comparison of values of sera against several C1. hzmolyticum
filtrates, in tests against different indicators
sheep red
cells
9000 1620 3150 4500 4700
580
_____
9000 1480 2600 3500 3700 840
Cl. hcemolyticum antisera RX5528* . . EX1249 . .
850B . . . I 1500
9000 1700
Serum value against CZ. hccmolyticum filtrate
EX1250 . , EX1269 . . EX1325 . . EX1440 . .
_ _ _ ~
Cl. mdematiew type-B antisera
EX1117 . . .
AJXB9 in tests against
3000 3500 3450 1150
675 280 2900 3300 300
1000
415 3700 3200 300 900
EX5325 . . T3 . . T61 . .
__ horse red cells
5700 2500 400
9000 1600 2900 4100 4100 1000 __
450 4700 2400 290
1100
AJX70 in tests against
L.V.
__
9000 1600 3000 4900 3700 500 __
270 2700 3000 360
1400
* Standard Tests carried out in 1957-58
__ sheep red cells
~
9000 1700 2600 4700 3700 1000
550 5000 3500 350
1200
__ horse red cells
9000 1800 3500 4500 3000 1350
490 4700 3600 330
1300
AJX71 in tests against
r,.v. __
9000 1600 2800 4300 4500
400
290 2500 3500 300 950
sheep red cells __
9000 1200 3000 3900 4100 800
~
380 4100 2900 350
1100
- liorse red cells __
9000 1300 2800 3600 4300 1000
430 4700 2950 250 900
tests by high dilutions of sera containing <0.1 unit Cl. adematicns a-antitoxin per ml., and antisera produced by injecting them into horses contain no detectable a-antitoxin.
Hcemolytic, L. V., and egg-yolk-agar tests
Table I also shows that in hzmolytic and L.V. tests against, dried filtrate NX605 most sera containing Cl. cpdematiens p-antitoxin give values agreeing well with one another and with their p-antitoxin values. Table I1 extends these tests to three other C1. hwmolyticum filtrates ; the tests were done about seven years later than those in tabla I, and some of the sera have consequently fallen in potency. It is clear, however, that for the majority of the sera tested, the values in L.V. tests and in hzmolytic tests against sheep or horse red cells agree me11 with one another and with the values obtained in intracutaneous and lethal tests, whatever the filtrate used.

CLOSTRIDIAL ANTIGENS 547
1 Values against CZ. hmrno~yticunz I filtrate NX605 in
Three sera, however, give values in L.V. tests against all four filtrates lower than the values in the other tests. These are EX1117 and RX5325 (tables I and 11) and EX1440 (table 11). This suggests that Cl. hcemolyticum filtrates contain, beside the hsmolytic, lethal and necrotising lecithinase p, a substance (possibly the lipasc observed by Macfarlane, 19504, which produces opalescence in egg-yolk emulsions, but has no haemolytic or necrotising powers.
Oakley et al. (1947) had shown that in CZ. adematiens type-A filtrates, which contain a lecithinase ( y ) and a lipase ( E ) , the effects of these two components could be separated in egg-agar columns. We therefore examined two sera showing no discrepancies between the values obtained in L.V. and h8molytic
TABLE I11
Values of sera against C1. haemolytioum filtrate ifi L.V. and egg-agar tests
Egg-yolk-agar tests.
Serum
RX5528 * EX1250 EX1440
EX1117
RX5325
Nature Anti-8 value
Cl. hcemolyticum j 9000 antiserum
3100 , 1 1150
Cl. adenlatiens 900 i 7000 type-B antiserum
L.V.tests
9000
3000 380
440
2500
Egg-agar tests
9000
3000 1300
920
8200
* Standard.
tests, and the three showing gross discrepancies, in egg-yolk-agar tests against filtrate NX605. It is clear from table I11 that if a serum shows no discrepancy between its values determined in L.V. and hsniolytic tests, its value in egg-yolk-agar t'ests agrees with these values ; the value for discrepant sera is raised to that found in hemolytic tests. This confirms the existence in Cl. hmmolyticum filtrates of a non- haeniolytic substance producing opalescence in egg-yolk emulsions.
It was possible that this substance might be the €-antigen of Cl, adematiens type A. A number of C1. cedematiens type-A antisera known to be deficient in €-antibody, as shown by the discrepancy between their estimated values in L.V. and h=molytic tests with horse cells as indicator, were therefore mixed with 6'1. hcemolyticum antisera that did not give discrepant values when tested against GI. hcemolyticurn filtrates. The mixtures were then tested against a Cl. odematiem type-A filtrate (OC 19245) in IA.TT. and hsrnolytic tests against horse red cells, to see whether the values were in any way altered. Table IV shows that no such change occurred; the Cl. hcemolyficum antisera used therefore contained no significant amount of €-antibody, so that it is unlikely that the second opalescing factor

548 C. L. OAKLEY AAVD CJ. IIARRIET WARRACK
in CI. hEmolyticum filtrates is €-antigen. As it appears t o be different from all the other antigens, we suggest it be called 0-antigen.
TABLE IV Tests for the presence of CI. cedcrnatiens €-antibody in
C1. h~molyticum antisera
EX1078 . 28 .
644 . 9783 .
-
-I- - - - - - --'
Value against CI. axhv7tiens typc-A filtrate 0019245 of I
1700 1 1700 620 740 640 i 1000 840 1 1510
1700 i 1700 I 1700 ' 1700 ' 5 80 740 640 i 1000 ' E:i I 1::; 940 1 1510 j 1000 I 1510 ~
Relationship of C1. adeunatiens P-tozirL and C1. uwlchii a-toxin. Since Sordelli et al. had informed Weinberg, Xativelle and Prhvot ( 1 937) that Cl. welchii ailtisera protect animals against Cl. hmmolyticurrz
TABLE V Cornpari.9o.n of C1. wclohii a-antitoxin a d C1. cedematiens ,&antitoxin wli tes of
a number of C1. wclchii t?ype-A and C1. hzmolyticum antisera ________
Rrrirm I
EX902
1 r'x251 ' 7005
7023 7041 7105 7148
IiX5528
EX1249 EX1250 EX1569 EX1287 EX1 292 EX1321
Cl. uximatiens Sature &antitoxin
, value (A)
GI. welchii type- A antisorum
GI. ha?rnal?jticuni antiserum
100
1'. 190 310
3 40 GO
190 - __ .
~ 500
460 320 600 570 480 650
9000 I YO
1700 1 20 3400 10 3500 40
3200 6 0 4800 0.5
4300 j ! _______
0.5
0.26 0.97 0~005 0.07 0.125 0.29
110
85 340 88 72 54
24,000
infection and that Cl. hmmolyticum antisera neutralise Cl. welchii toxin (presumably the a-toxin), we have determined the Cl. welchii a- antitoxin values of our Cl. hcemolyticum antisera and the Cl. mdematiens ,6-antitoxin values of some GI. webhii type-A antisera. The Cl. hmaolyticum antisera had been produced in horses that had not been

CLOSTRIDIAL ANTIGENS 549
injected with any other toxin or toxoid, apart from tetanus toxoid, SO
that apart from accident, it was unlikely that they had any experience of Cl. wclchii a-toxin. The C1. welchii antisera had been obtained from horses that had never been injected with C1. ademutiens toxins or toxoids.
Table B shows that Cl. welchii type-A antisera contain CI. mdema- tiens 13-antitoxin, and that Cl. hcmtolyticum antisera contain C’l. uielchii a-antitoxin. The ratio between the concentrations of the two antitoxins (Cl. ademutiens ,&antitoxin units per ml. jCl. uielchii %-antitoxin units per ml.) covers a wide range--0.005-0.97 for Cl. welchii type-A antisera and 54-24,OOO for Cl. adenzatiens type-13 antisera, so that it seems improbable that the antigenic relationship betvr-een the two toxins can be a close one. However, since tliese sera carnc from horses that had, as far as we know, received only one or the other filtrate, it is unlikely that the finding is fortuitous.
Discussmx What view one takes of the position of Clostridium hcemolyticurn
in the genus Clostridium is very largely a question of convenience. It differs from Cl. cx?detmtiens types A and B only in producing infectious icterohaemoglobinuria in cattle rather than gas gangrene or infectious necrotic hepatitis, and in the fact that the pathogenic effects of cultures in animals may be prevented by previous injection of Cl. hcemolyticum or Cl. cedematiens type-R antiserum, but not by injection of Cl. ademutiens type-A antiserum. On the other hand, Turner and Eales (1943) have shown that one strain of Cl. hmnolyticum and several strains of Cl. edematiens share one somatic antigen (but see p. 543 of this. paper) ; we have shown that they share one soluble antigen (p ) , and Jlacfarlane (1955) that they share another (the txopomyosinase q).
The 13-antigen is a calcium-dependent haeniolytic, necrotising and lethal lecithinase, about equally active against the red cells of horses and sheep (Macfarlane, 1950b). It is produced by Cl. Jmnaolyticu?n in much higher concentration than by Cl. ademutiens type B, and weak C1. odemutiens type-B antisera may therefore fail to neutralise it. The pathogenic effects of cultures or filtrates of C1. humholyticum in cattle are evidently due to the P-lecithinase, whereas those of Cl. ademutiens types A and B are in the main due to the necrotisillg and lethal %-toxin.
From the point of view of the immunologist, it is obviously more convenient to regard C1. hctzmolyticzm as CI. cx?dematiena type D, differing from type A in producing 13, 7, and 0 and no cc, y or E, and from type B in producing no c( or 5 (type B most probably produces a little 0, as type-B antisera often have antibody to it) ; type C is almost certainly non-toxic (table VI). 1f this suggestion is adopted, there need be no confusion over the toxins: which will be called by the same name whichever organism is referred to ; clinical papers could put
J. PATH. BACT.-VOL. 78 (1959) 21\12

650 C. L. O A K L E Y A N D G. HARRIET W A R R A C K
C1. hcernolyticum in brackets after C1. adematiens type D, or in papers where the toxins were not referred to, use the older and more familiar name.
The diagnosis of Cl. ademutiens type D will normally be fairly easy. The scheme proposed by Oakley, Warrack and Clarke (1947, p. 101), may be supplemented by the use of a tube containing 1 ml. of the filtrate and 200 units Cl. ademutiens p-antitoxin. If this amount of antitoxin neutralises the filtrate in L.V. tests and 10 p units do not, the organism is probably Cl. hceinolyticum, a diagnosis which may be confirmed by showing that lesions produced by the filtrate on intra- cutaneous injection into the guinea-pig’s skin are completely neutralised
TABLE VI Soluble antigens of the C1. cedematiens group
1 Presence in filtrates from Cl. cedematiens type .~
Activity Designation
Necrotising, lethal . Haemolytic necrotising lethal
Haemolytic necrotising lecithinase Oxygen-labile hamolysin . Opalescence in egg-yolk emulsion:
? pearly layer Haemolysin . Tropomyosinase . Opalescence in egg-yolk emulsion :
lecithinase
? lipase
++ + + +
-
- - -
+++ + - - -
+ + t r
czdemu- by C1. adenmtiens type-D antisera containing no detectable C1 t iens a-antitoxin. We used this method in showing that the organism isolated by Soltys and Jennings (1950) was Cl. ademutiens type D ; it has given the expected results with all strains of Cl. ademutiens type 13 we have examined.
SUMMARY Clostridium hcemolyticum produces two soluble antigens ; one is
a calcium-dependent necrotising lethal lecithinase, active against sheep and horse red cells, and antigenically indistinguishable from the P-lecithinase of Cl. odenutiens type R ; the other is a substance {Cl. adernatiens &antigen) producing opalescence in egg-yolk emulsions, but possessing no hzemolytic, necrotising or lethal activity. No CZ. adermtiens a-toxin is produced by any strain examined.
It is suggested that in papers on classification or on immunology the organism should be referred to as C1. ademutiens t,ype D.
c We should like to express our thanks to Miss J. Waite for culture filtrates,
and to Miss E. Hart for technical assistance.

CLOS'TRIDIAL AATTIBEA7S
REFERENCES
55L
BARD, R. C., AND MCCLUNC, L. S. 1948. DEMNITZ, A. . . . . . . 1934.
HALL, I. C. . . . . . . . JASMIN, A. M. . . . . . . KREUZER, E. . . . . . . MACFARLANE, MARJORIE G. . .
li
,>
. .
. . MACFARLANE, R. G., OAKLEY,
C. L., AND ANDERSON, C. G. OAKLEY, C. L., WARRACK, G.
HARRIET, AND CLARICE, PATRICIA H.
SMITH, L. DS. . . . . . . SOLTYS, M. A., AND JENNINGS,
TURNER, A. W., AND EALES, A. R.
CATHARINE E.
1929. 1947. 1939. 1950a. 1950b. 1955. 1941.
1947.
1953. 1950.
1943.
J . Bact., 56, 666. Medicine in its chemical aspects,
J . Inf. Dis., 45, 156. Amer. J. Vet. Res., 8, 289. 2. ImmunForsch., 95, 345. Biocliem. J., 47, 267. Ibid., 47, 270. Ibid., 61, 308. This Journal, 52, 99.
Leverkusen, vol. 2, p. 289.
J . Gen. Microbiol., 1, 91
J. Bact., 65, 222. Vet. Rec., 62, 5.
Austral. J. Exp. Biol. Med. Xci., 21, 79.
VAWTER, L. R., AND RECORDS, E. 1925-26. J . Amer. Vet. Med. Assoc., 682 494.
WEINBERG, M., NATIVELLE, R., 1937. Les microbes anaArobies, Paris, 2 , >, 1, 2, 1931. J. Imf . Dis., 48, 581.
AND PREVOT, A. R. p. 293.