the inhibition of human plasma acetylcholinesterase by some naturally occurring compounds
TRANSCRIPT

Gen. Pharmac., 1978, Vol. 9, pp. 21 to 24. Peroamon Press. Printed in Great Britain
THE INHIBITION OF HUMAN PLASMA ACETYLCHOLINESTERASE BY SOME NATURALLY
OCCURRING COMPOUNDS*
T. T. NGO, G. TUNNICLIFF, C. F. YAM, M. CHARBONNEAU and A. BARBEAU
Clinical Research Institute of Montreal, and Department of Medicine, University of Montreal, Montreal, Quebec, Canada H2W 1R7
(Received 23 June 1977)
Abstract--1. Thiamine, choline, and carnitine each inhibited the human plasma acetylcholinesterase. Thiamine hydrochloride produced a mixed type of inhibition with a Ki of 1.5 mM and an ctKi of 6.5 mM. Choline bromide gave rise to a competitive inhibition of the enzyme, with a K~ of 5.2 mM. DL-Carnitine hydrochloride also inhibited the acetylcholinesterase in a competitive manner, with a K~ of 4.1 mM. The methyl ester of carnitine was eight times as potent as free carnitine (K~ = 0.5 mM):
2. Metrazol produced a mixed type of inhibition of the enzyme. It was less potent than the other inhibitors (Ki = 17 mM, ctKi = 11 mM).
3. No differences emerged in the activity of plasma acetylcholinesterase between Friedreich's ataxia patients and a control group of subjects. The mean values for the enzyme were 2.06 and 2.25/~moles/min per ml of plasma, respectively.
4. The results demonstrate that although Friedreich's ataxia might be a disease in which the acetylcho- line system is not functioning adequately, this is not reflected in the activity of plasma ACHE. However, since thiamine, choline and carnitine are able to inhibit the enzyme, they might be able to provide therapeutic value in the treatment of Friedreich's ataxia or any other cholinergic-deficiency disease.
INTRODUCTION
There is some evidence that the cholinergic system may be involved in the etiology of Friedreich's ataxia and Huntington's chorea. Both the oxidation of pyru- vate and the activity of pyruvate dehydrogenase (PDH) have been reported to be reduced in Fried- reich's patients (Kark et al., 1974; Barbeau et al., 1976; Blass et al., 1976). Furthermore, Gibson and colleagues (1975) have demonstrated that if PDH is inhibited by certain drugs, brain acetylcholine syn- thesis is concomitantly reduced. This link between pyruvate oxidation and acetylcholine metabolism is further strengthened by the findings that rats made thiamine deficient possess a reduced PDH activity and a decrease in acetylcholine levels in the brain (Dreyfus & Hauser, 1965; Reinauer et al., 1968; Cheney et al., 1969; Heinrich et al., 1973). Moreover, this deficiency of thiamine is accompanied by an ataxia. It should be pointed out, however, that Reynolds and Blass (1975) could not confirm that brain acetylcholine is affected by thiamine deficiency, nor could Heinrich et al. (1973) show that brain PDH activity was decreased in such animals. It is pertinent to note that physostigmine, an acetylcholinesterase (ACHE) inhibitor, is reported to be beneficial in the treatment of Friedreich's ataxia (Kark et al., 1975).
In the post-mortem brain of patients with Hunting- ton's disease, choline acetyltransferase activity has been shown to be substantially reduced (McGeer et al., 1973; Bird & Iversen, 1974). A decrease in choline levels has also been reported in the cerebrospinal fluid
* Quebec Cooperative Study of Friedreich's ataxia. 21
of patients with this disease (Stahl & Swanson, 1974; Aquilonius et al., 1972). As with Friedreich's ataxia, physostigmine has been used with some success to treat people suffering from Huntington's chorea (Klawans & Rubovits, 1972; Tarsy et al., 1974).
In light of the foregoing, we speculated that, in addition to physostigmine, substances that inhibit the breakdown of acetylcholine might have therapeutic value in the treatment of Friedreich's ataxia and Huntington's disease. It is known that substances that contain a quaternary nitrogen are almost invariably able to inhibit the esterase (Kitz, 1964). We are par- ticularly interested in finding compounds naturally occurring in the body that possess this ability to in- hibit ACHE. Thiamine, choline and carnitine should
f i t this role as each possesses such a quaternary nitrogen group. Indeed, it has already been shown that thiamine and choline can inhibit the serum enzyme and that isolated from nervous tissue of the electric eel (Glick & Antopol, 1939; Alspach & Ingra- ham, 1977; Adam & Whittaker. 1950).
This paper describes our attempts to study the mechanism of action of thiamine and other sub- stances on the activity of human plasma ACHE, and our conclusions concerning their use as potential therapeutic agents.
METHODS
Chemicals
Thiamine hydrochloride, choline bromide, OL-camitine hydrochloride and acetylthiocholine chloride were obtained from the Sigma Chemical Co., St. Louis, MO, U.S.A.

22 T.T. NGO, G. TUNNICLIFF, C. F. YAM, M. CHARBONNEU and A. BARBEAU
Source of enzyme For the inhibitor studies, blood was collected from nor-
mal subjects, both male and female, who were between the ages of 22 and 38. For the study of AChE in Fried- reich's ataxia, blood was taken from a sample of patients, both male and female, whose ages ranged from 8 to 35. Control patients' ages ranged from 21 to 53 years. In every case blood was collected in heparinized tubes and centrifuged at 2000 rev/min in a Beckman centrifuge (JA-20 rotor) for 15 min at 4°C. The resulting plasma was the source of the enzyme.
Assay of AChE One milliliter of plasma was diluted to 50 ml with 0.1 M
sodium phosphate buffer, pH 7.5. This diluted plasma was used to measure the activity of the AChE by the procedure of Ellman et al. (1961). One milliliter of enzyme was added to 1 ml acetylthiocholine and 1 ml of 5,5'-dithiobis (2-nitro- benzoic acid). The latter two substances were dissolved in the buffer described above. The increase in absorbance of the solution was monitored at 412 nm in a Beckman Acta M VI spectrophotometer at 30°C. Further details of each assay are given in the Results section and in the legends for the figures.
Preparation of the methyl ester of carnitine The methyl ester of carnitine was prepared according
to the standard method of amino acid carboxylic methyla- tion (Greenstein & Winitz, 1961). The ester was thoroughly dried under high vacuum before use.
R E S U L T S
AChE activity in patients with Friedreich's ataxia
When the activity of plasma AChE was measured in Friedreich's pat ients and compared to the enzyme in control group, no significant differences were observed (Fable 1).
Table I. AChE activity in the serum of Friedreich's patients and control subjects
Enzyme activity n (itmoles/min per ml) +S.E.
Friedreich's 13 2.06 0.45 Controls 13 2.25 0.67
0.6
' b tO 2 30
0.4.
b 0.2
Fig. 1. Inhibition of plasma AChE by thiamine hydrochlo- ride recorded as a Lineweaver and Burk plot. The enzyme was measured in the absence (O) or presence (A, 2 mM; va, 4 mM) of thiamine over a substrate concentration (S)
range of 0.033q3.25 mM
Inhibition Of AChE Each of the compounds tested inhibi ted the plasma
enzyme either competitively or with a mixed type of inhibition. In addition, there were differences in the potencies of the inhibitors.
The enzyme was assayed over a substrate concen- t rat ion range of 0.03343.2 m M in the absence or pres- ence of th iamine hydrochloride. When the results were plotted by the method of Lineweaver & Burk (1934) a mixed type of inhibi t ion was observed (Fig. 1) with a Ki of 1.5 m M and ~K~ of 6.5 mM. This con- firms the results of Alspach & Ingraham (1977) who demonstra ted that electric eel AChE was inhibi ted by thiamine with a mixed type of inhibition.
ES
E
E + P
ESI
Scheme 1.
The mixed inhibi t ion can be represented by Scheme 1, where E = enzyme, S = substrate, I = inhibitor, Ks are various equil ibrium constants, k c = catalytic constant , ~ = a proport ional i ty factor. The inhibitor, when it binds to the free enzyme, causes a change in substrate binding, hence Ks is changed to ~Ks. Similarly the enzyme-subst ra te binds the inhibi tor with an affinity (~Ki) which differs from that of the free enzyme (K0. The rate equation, in the presence of mixed inhibitor , is:
gmax I-S]
K, 1 + K i J + 1 + [S]
where Vm,, = kc[-E]. The double-reciprocal plots of equat ion (1) at
several fixed concentrat ions of inhibitors give a family of straight lines intersecting to the left of 1/V axis and above the 1/S axis for 7 > 1, and below the 1/S axis for ~ < 1. K i was determined from the secondary plot of the slopes of the Lineweaver-Burk plot against the inhibi tor concentrat ion, and 0tK~ was determined from the secondary plot of the intercepts of the Lineweaver-Burk plot against the inhibi tor concen- tration. When the experiment was repeated using cho- line bromide in place of the thiamine, a competit ive inhibi t ion with respect to substrate was noted (Fig. 2). The K~ was calculated as 5.2 mM. oL-Carnit ine hydrochlor ide gave rise to a very similar inhibit ion of the enzyme (K~ = 4.1 mM) (Fig. 3). We thought that the structure of the carni t ine molecule might not
Inhibition of plasma AChE 23
(15
t13
0 . 2
Y Fig. 2. Inhibition of the plasma esterase by choline bro- mide plotted by the procedure of Lineweaver and Burk. The enzyme was assayed in the absence (0) or presence (A, 2 mM; F-I, 4 mM) of choline over a substrate concen-
tration (S) range of 0.033 mM-0.25 mM.
f.O
[ ]
0.8
0.6
O4
O.2
i i t
0 I0 20 30
Fig. 4. Inhibition of plasma AChE by metrazol. Data plot- ted by the method of Lineweaver and Burk. The esterase was assayed in the absence (0) or presence (A, 3 mM; [-1,
6 mM} of the drug.
be the most advantageous for its inhibition of the AChE due to an internal neutralization of the positive charge of the quaternary ammonium group by the negative charge of the carboxylic group. Conse- quently, we synthesized OL-camitine methyl ester and carried out a similar inhibition study. Esterification of the carboxylic group by methanol eliminates its
3
~v 2
A
zx
' 'o ' ' 'o 0 fO 2 30 40 5
Fig. 3. Inhibition of the plasma enzyme by carnitine hydro- chloride and camitine methyl ester. The data are recorded as a Lineweaver and Burk plot. The enzyme was deter- mined in the absence (0) or presence of carnitine (0.6 mM) or carnitine methyl ester (/x, 3 mM; ,t, 6 mM) over a sub-
strate concentration (S) range of 0.02 to 0.25 mM.
negative charge, thus making the internal charge neu- tralization impossible. As can be seen from Fig. 3 the methyl ester was much more potent in its inhibi- tion of the enzyme than carnitine itself. The K i (0.5 mM) was almost 8-fold that of carnitine.
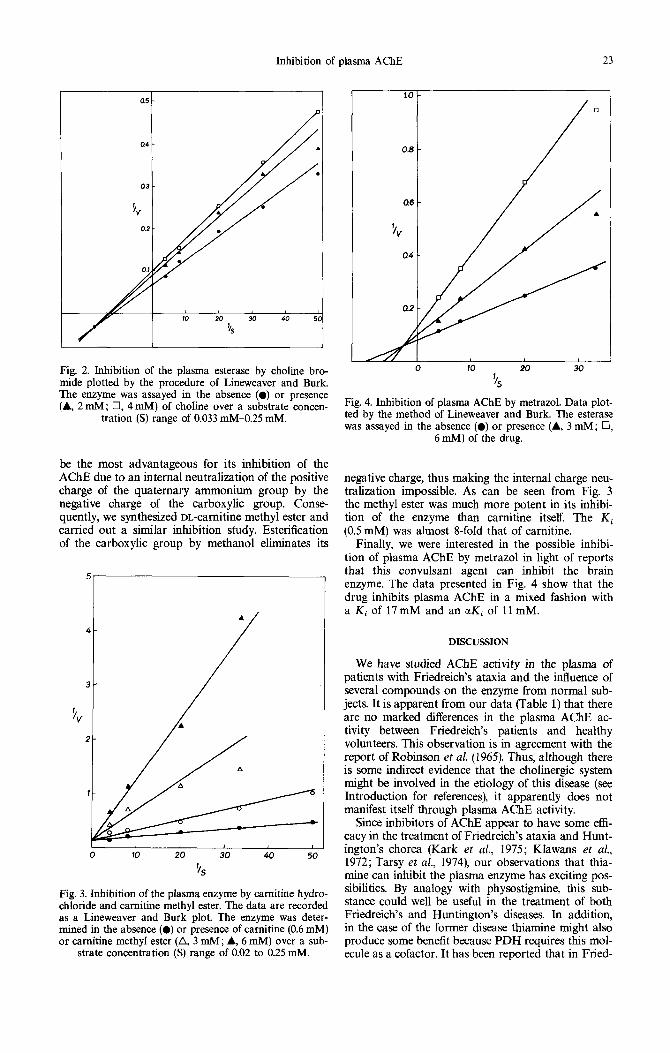
Finally, we were interested in the possible inhibi- tion of plasma AChE by metrazol in light of reports that this convulsant agent can inhibit the brain enzyme. The data presented in Fig. 4 show that the drug inhibits plasma AChE in a mixed fashion with a Ki of 17mM and an ctKi of l l m M .
D I S C U S S I O N
We have studied AChE activity in the plasma of patients with Friedreich's ataxia and the influence of several compounds on the enzyme from normal sub- jects. It is apparent from our data (Fable l) that there are no marked differences in the plasma AChE ac- tivity between Friedreich's patients and healthy volunteers. This observation is in agreement with the report of Robinson et al. (1965). Thus, although there is some indirect evidence that the cholinergic system might be involved in the etiology of this disease (see Introduction for references), it apparently does not manifest itself through plasma AChE activity.
Since inhibitors of AChE appear to have some effi- cacy in the treatment of Friedreich's ataxia and Hunt- ington's chorea (Kark et al., 1975; Klawans et aL, 1972; Tarsy et al., 1974), our observations that thia- mine can inhibit the plasma enzyme has exciting pos- sibilities. By analogy with physostigmine, this sub- stance could well be useful in the treatment of both Friedreich's and Huntington's diseases. In addition, in the case of the former disease thiamine might also produce some benefit because PDH requires this mol- ecule as a cofactor. It has been reported that in Fried-

24 T . T . NGO, G. TUNNICLIFF, C. F. YAM, M. CHARBONNEU and A. BARBEAU
reich's ataxia, in addit ion to a reduction in pyruvate oxidation, P D H activity is also decreased in this dis- ease.
As with thiamine, choline might offer benefit to pat ients with Friedreich's or Hunt ing ton ' s diseases. Choline not only inhibits plasma AChE (Fig. 2) but also is a precursor of acetylcholine and so will tend to enhance acetylcholine levels by two mechanisms: inhibi t ion of degradat ion and stimulation of synthesis. It has already been demonstra ted that dietary choline can increase brain acetylcholine concentra t ion in rat (Cohen & Wur tman, 1975).
The inhibi t ion of AChE by carnit ine (Fig. 3) may not reflect the true potency of the molecule since the quaternary a m m o n i u m group of carni t ine is internally being neutral ized by the negative charge of its own carboxylic group, thus rendering the molecule incap- able of interacting with the anionic site of the ACHE. When carnit ine methyl ester was used to inhibi t the estcrasc, the Kg was reduced by 86%. The molecule in the ester form contains a quaternary a m m o n i u m group that is free to interact with the anionic site of the enzyme.
Finally, metrazol was included in this study, not because of any potential therapeutic value, but because it contains no quaternary ni trogen yet it has been reported that it can inhibi t brain ACHE. We were interested to see if it had a similar effect on the plasma enzyme. As can be seen in Fig. 4 the drug acts as a mixed inhibi tor of the esterase. However, the K i values were higher than that of the other in- hibi tors tested. Since metrazol can induce seizures, it has been suggested that, as acetylcholine is an exci- ta tory neurotransmit ter , these convulsions could be related to an increase in brain levels of this substance (Mahon & Brink, 1970). It is probable tha t the inhibi- t ion of plasma AChE by this convulsant drug that we have observed is merely a reflection of what is occurring centrally and is unrelated to the seizures tha t can be induced by metrazol.
REFERENCES
ADAM D. H. & WHITTAKER V. P. (1950) The cholinesterases of human blood--II. The forces acting between enzyme and substrate. Biochim. biophys. Acta 4, 543 558.
ALSPACH J. D. & INGRAHAM L. L. (1977) Inhibition of ace- tylcholinesterase by thiamine. A structure-function study. J. reed. Chem. 20, 161-164.
AQUILONIUS S. M., NYSTROM B., SCHUBERTH J. ~,~ SUND- WALL H. (1972) Cerebrospinal fluid choline in extrapyra- midal disorders. J. Neurol., Neurosurg. Psychiat. 35, 720-725.
BARBEAU A., BUTTERWORTH R. F., NGO T. T., BRETON G., MELANCON S., SHAPCOTT D., GEOFFROY G. & LEMIEUX B. (1976) Can. J. Neurol. Sci. 3, 379 388.
BIRD E. D. & IVERSZN L. L. (1974) Huntington's chorea. Post-mortem measurement of glutamic acid decarboxy- lase, choline acetyltransferase and dopamine in basal ganglia. Brain 97, 457472.
BLASS J. P., KARK R. A. P. & MENON K. (1976) Dehydro- genase deficiency in Friedreich's ataxia. New Engl. J. Med. 295, 62 67.
CHENEV D. L., GUBLER C. J. & JAUSSI A. W. (1968) Produc- tion of acetylcholine in rat brain following thiamine deprivation treatment with thiamine antagonists. J. Neurochem. 16, 128~1291.
COHEN E. L. & WURTMAN R. J. (1975) Brain acetytcholine: increase after systemic choline administration. Life Sci. 16, 1095-1102.
DREYFUSS P. M. & HAUSER G. (1965) The effect of thiamine deficiency on the pyruvate decarboxylase system of the central nervous system. Biochim. biophys. Acta 104, 78-84.
ELLMAN G. L., COURTNEY K. D., ANDRES V. & FEATHER- STONE R. M. (1961) A new and rapid colorimetric deter- mination of acetylcholinesterase activity. Biochem. Phar- mac. 7, 88-95.
GIBSON G. E., JOPE R. & BLASS J. P. (1975) Decreased synthesis of acetylcholine accompanying impaired oxi- dation of pyruvic acid in rat brain minces. Biochem. J. 148, 17 23.
GLICK D. & ANTOPOL W. (1939) The inhibition of choline esterase by thiamine (vitamin B). J. Pharmac. exp. Ther. 65, 389-394.
GREENSTEIN J. P. & WINITZ M. (1961) Chemistry of the Amino Acids, pp. 926. John Wiley, New York.
HEINRICH C. P., STADLER H. & WEISER H. (1973) The effect of thiamine deficiency on the acetylcoenzyme A and ace- tylcholine levels in the rat brain. J. Neurochem. 21, 1273 1281.
KARK R. A. P., BLASS J. P. & ENGEL W. K. (1974) Pyruvate oxidation in neuromuscular diseases---evidence of a gen- etic defect in two families with the clinical syndrome of Friedreich's ataxia. Neurology 24, 964~971.
KARK R. A. P., BLASS J. P., SPENCE M. A., MENON N. K. & HARRIS S. E. (I.975) Pharmacologic and enzymatic studies in Friedreich's ataxia. Neurosci. Abstr., 5th Annual Meeting of the Society for Neuroscience. Vol. 1, p. 327.
KITZ R. J. (1964) The chemistry of anticholinesterase ac- tivity. Acta anaesth, scand. 8, 197 218.
KLAWANS H. L. & RUBOVITS R. (1972) Central cholinergic anticholinergic antagonism in Huntington's chorea. Neurology 22, 107 116.
MCGEER P. L., MCGEER E. G. & FIB1GER H. C. (1973) Cholineacetylase and glutamic acid decarboxylase in Huntington's chorea. Neurology 23, 912~17.
MAHON P. J. & BRINK J. J. (1970) Inhibition of acetylcho- linesterase in vitro by pentylenetetrazol. J. Neurochem. 17, 949~53.
REINAUER H., GRASSOW G. 8z, HOLLMAN S. (1968) Aktivit~its- anderungen der Py~'uvdehydrogenase im Thiaminno- mangel. Hoppe-Seyler's Z. physiol. Chem. 349, 969 978.
REYNOLDS S. F. 8z. BLASS J. P. (1975) Normal levels of acetyl coenzyme A and of acetylcholine in the brain of thiamin-deficient rats. J. Neurochem. 24, 185-186.
ROBINSON N., PHILLIPS B. M. & CUMINGS J. N. (1965) Serum enzymes in Friedreich's ataxia. Brain 88, 131 136.
STAHL W. L. & SWANSON P. D. (1974) Biochemical abnor- malities in Huntington's chorea brains. Neurology 24, 813 819.
TARSY D., LEOPOLD N. & SAX D. S. (1974) Physostigmine in choreiform movement disorders. Neurology 24, 28-33.