targeting syk activated b-cells in murine and human
TRANSCRIPT

1
Targeting Syk Activated B-cells in murine and human chronic graft-versus-host disease
Ryan Flynn1, Jessica L. Allen2, Leo Luznik3, Kelli P. MacDonald4, Katelyn Paz1,
Kylie A. Alexander4, Ante Vulic3, Jing Du1, Angela Panoskaltsis-Mortari1, Patricia A. Taylor1, Jonathan C. Poe2, Jonathan S. Serody5, William J. Murphy6, Geoffrey R. Hill4, Ivan Maillard7, John Koreth8, Corey S. Cutler8, Robert J. Soiffer8, Joseph
H. Antin8, Jerome Ritz8, Nelson J. Chao2, Raphael A. Clynes*9, Stefanie Sarantopoulos*2 and Bruce R. Blazar*1.
1Division of Blood and Marrow Transplantation, University of MN, Masonic Cancer Center and Department of Pediatrics, Minneapolis, MN; 2Division of Hematological Malignancies and Cellular Therapy and Duke Cancer Institute, Duke University Department of Medicine and Duke Cancer Institute, Durham, NC; 3Sidney Kimmel Comprehensive Cancer Center and Department of Oncology, The Johns Hopkins University, Baltimore, MD; 4Department of Immunology, Queensland Institute of Medical Research, Brisbane, Australia; 5Lineberger Comprehensive Cancer Center Division of Hematology/Oncology, University of North Carolina, Chapel Hill, NC; 6Department of Dermatology, School of Medicine, University of California, Davis, Sacramento, CA; 7Life Sciences Institute and Department of Internal Medicine, Division of Hematology-Oncology, University of Michigan, Ann Arbor, MI; 8Dana Farber Cancer Institute, Boston, MA; 9Columbia University Medical Center, Columbia University. *Equal contributors
Short title: Chronic GVHD suppression by Syk inhibition
Correspondence: Bruce R. Blazar, M.D.; MMC 109, University of Minnesota 420 Delaware St. SE; Minneapolis, MN 55455 USA; E-mail: [email protected] Tel: 612-626-1926 Fax: 612-624-3913 Word No: Abstract: 200; Text: 3889; Figures: 7; Supplemental Figures 1; Supplemental Table 1; References: 37; Category: TRANSPLANTATION
Blood First Edition Paper, prepublished online April 7, 2015; DOI 10.1182/blood-2014-08-595470
Copyright © 2015 American Society of Hematology
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

2
Key Points
1) Syk is required for increased B cell activation and cGVHD generation and
maintenance.
2) The Syk inhibitor Fostamatinib can treat murine cGVHD and increase
human cGVHD B cell death.
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

3
Abstract
Novel therapies for chronic graft-versus-host disease (cGVHD) are needed.
Aberrant B cell activation has been demonstrated in mice and humans with
cGVHD. Having previously found that human cGVHD B cells are activated and
primed for survival, we sought to further evaluate the role of the spleen tyrosine
kinase (Syk) in cGVHD in multiple murine models and human peripheral blood
cells. In a murine model of multi-organ system, non-sclerodermatous disease
with bronchiolitis obliterans where cGVHD is dependent on antibody and
germinal center (GC) B cells, we found that activation of Syk was necessary in
donor B cells but not T cells for disease progression. BM-specific Syk deletion in
vivo was effective in treating established cGVHD as was a small molecule
inhibitor of Syk, fostamatinib which normalized GC formation and decreased
activated CD80/86+ dendritic cells. In multiple distinct models of sclerodermatous
cGVHD, clinical and pathological disease manifestations were not eliminated
when mice were therapeutically treated with fostamatinib, though both clinical
and immunological effects could be observed in one of these scleroderma
models. We further demonstrated that Syk inhibition was effective at inducing
apoptosis of human cGVHD B cells. Together these data demonstrate a
therapeutic potential of targeting B-cell Syk signaling in cGVHD.
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

4
Introduction
The development of cGVHD is a major complication following allogeneic
hematopoietic stem cell transplantation. Discovery of new therapies has been
limited by the absence of murine models that closely represent the clinical human
disease and pathogenesis1, 2. Herein we use multiple mouse models to assess
the effectiveness of therapies during cGVHD. We utilize a mouse model of
antibody-dependent, multi-organ system cGVHD, previously demonstrated to
mimic several aspects of human cGVHD pathology, with the exception of
sclerodermatous cGVHD3, 4and three models of sclerodermatous cGVHD with
skin manifestations5, 6.
While the exact mechanisms of cGVHD remain unknown, recent studies
have elucidated a role for antibody production by B-cells. The increase of B-cells
in the germinal center (GC) has been shown to be necessary for the
development of an antibody-facilitated multi-organ system cGVHD model that
includes bronchiolitis obliterans (BO)3 Spleen tyrosine kinase (Syk) is activated
by B-cell receptor (BCR) engagement. After antigen-BCR engagement, Syk is
phosphorylated at Y348, allowing for B-cell survival and proliferation. Increased
proximal BCR signaling was recently described in cGVHD patient B cells.7
Syk also affects myeloid cell [macrophages; neutrophils and dendritic cells
(DCs)] including phagocytosis, signal transduction via activating FcRs, and
antigen-uptake, internalization and upregulation of costimulatory molecules in
DCs.8 Syk also has been demonstrated to have an impact on cell-migration of
monocytes.9 Given the function of Syk in myeloid cells and the critical role of
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

5
macrophages and DCs in cGVHD5,10, Syk inhibition may have additional
advantages of treating cGVHD via macrophage and DC effects.
We sought to determine whether B-cell Syk activation was a critical
component in cGVHD pathophysiology. First, we demonstrated that Syk was
hyper-responsive in B-cells during cGVHD and that Syk was necessary in B- but
not T-cells for murine cGVHD progression. Next, we demonstrated that the
deletion of Syk in donor BM cells in vivo in mice with established cGVHD with BO
was able to reverse disease, a finding that was phenocopied by the in vivo
molecular inhibition of Syk with fostamatinib. Inhibition of Syk decreased the
frequency of GCs and expression of the activation costimulatory molecules CD80
and CD86 on CD11c+ cells in vivo. In multiple distinct models of sclerodermatous
cGVHD, clinical and pathological disease manifestations were not eliminated
when mice were therapeutically treated with fostamatinib, though both clinical
and immunological effects could be observed in one of the sclerodermatous
cGVHD models. Human cGVHD B cells had increased death when treated with
fostamatinib, demonstrating BCR-activated B cells can be preferentially targeted.
These data together suggest that Syk could be a novel therapeutic for cGVHD
patients.
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

6
Materials and Methods
Mice
C57Bl/6 (B6; H2b) mice were purchased from the National Cancer Institute.
B10.BR (H2k) mice were purchased from Jackson Laboratories. Female C57Bl/6
(B6) (H-2b, CD45.2), and B6D2F1 (H-2b/d, CD45.2) mice were purchased from
the Animal Resources Center. BALB/c, B10.D2-Thy1.2 and B10-.D2-Thy1.1 mice
were propagated in the animal facility at the Johns Hopkins University Cancer
Research Building I. The Syk fl/fl x ERT2-cre mice were provided by RAC from
Columbia University. Deletion of Syk (Syk KO) occurred with administration of
tamoxifen (Sigma) PO for 5 days and confirmed by Western Blot. Mice were
housed in a specific pathogen-free facility and used with the approval of each
institution’s animal care committee.
BMT
For model 1 (systemic cGVHD with BO, MHC disparate), B10.BR recipients were
conditioned with cyclophosphamide and total body irradiation (8.3 Gy).
Recipients received 10x106 T cell depleted (TCD) bone marrow (BM) cells from
B6 wildtype (WT) or BM cells from donors that were induced to delete Syk by
tamoxifen administration. Cohorts received no supplemental T cells or purified
splenic T-cells (0.1x106) from WT vs KO donors3. In one experiment, as
indicated, mice were given donor BM from Syk fl/fl x ERT2-cre or WT mice with
or without purified T cells; on D28 post-BMT, mice were given tamoxifen to delete
Syk from donor BM cells. For model 2 (sclerodermatous cGVHD, semi-allogeneic
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

7
recipients), on day -1, B6D2F1 recipient mice received total body irradiation (11
Gy; 137Cs source), split into 2 doses separated by 3 hours to minimize
gastrointestinal toxicity5. On day 0, recipients underwent transplantation with 5 x
106 BM with or without 1 x 106 purified T cells from B6 mice. Non-GVHD control
groups were injected with TCD BM grafts. For model 3 (sclerodermatous cGVHD
with G-CSF mobilized splenocytes, MHC disparate), on day –1, mice received
total-body irradiation (1Gy; 137 Cs source), delivered as in model 2. On day 0, B6
mice each received 25 × 106 donor splenocytes from G-GSF–mobilized BALB/c
donors. Spleen were depleted of T cells as previously described for non-GVHD
control groups11. Transplanted mice were monitored daily, and those with GVHD
clinical scores of ≥610, 11 were sacrificed, and the date of death registered as the
next day in accordance with institutional animal ethics committee guidelines. For
model 4 (sclerodermatous cGVHD, minor histocompatibility antigen disparate), a
single lethal irradiation dose of 7.75 Gy was administered using a 137Cs irradiator.
Animals were reconstituted with 107 B10.D2-Thy1.2 TCD BM cells (107) alone or
supplemented 1.8 × 106 CD4+ and 0.9 × 106 CD8+ Thy1.1+ T cells, reflecting T
cells found in 1.2 × 107 B10.D2 donor splenocytes, a dose that reproducibly
induces GVHD6. Purified populations of donor T cells were obtained using T cell
isolation kits (Dynabeads, Invitrogen, Carlsbad, CA).
Where indicated, cGVHD recipients were given R788 (30
mg/Kg/animal/twice PO daily) or 0.1% carboxymethylcellulose vehicle from d28-
56 (BO model 1), D14-28 (B6 into B6D2F1, scleroderma, model 2), days 7-21
(G-CSF mobilized Balb/c splenocytes into B6 recipients, scleroderma model 3),
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

8
or B10.D2 into Balb/c (experiment 1, D21-35; experiment 2, D14-28). Both pro-
drug, R788, and its active metabolite, R406 (both known as fostamatinib) were
kindly provided by Rigel Pharmaceuticals. Lung function was assessed as
previously described.12
Patient Samples
Samples were obtained from patients following written informed consent in
accordance with the Declaration of Helsinki. Viably frozen peripheral blood
samples from patient with either active clinical manifestations of cGVHD or no
active cGVHD were randomly selected from cell banks for study (patient
characteristic details are shown in (Supplemental Table 1). The Institutional
Review Boards at the University of North Carolina Chapel Hill, Duke University
Medical Center, and the Dana-Farber Cancer Institute approved all studies.
Analysis of Syk Activation and In Vitro Effects of Syk Inhibition
For analysis of Syk phosphorylation in murine cGVHD, B-cells were purified from
murine cGVHD spleens or BM only controls on D60 in the cGVHD/BO model and
activated by 5 μg/mL anti-IgM Fab (Jackson Research), fixed with BD Cytofix
Fixation Buffer and permeabilized with BD Perm Buffer III. Cells were
subsequently stained with anti-Syk Y348 PE (eBioscience) as previously
described13.
For analysis of the effects of in vitro Syk inhibition, cryopreserved PBMCs
from cGVHD or non-cGVHD control patients were thawed and allowed to rest
overnight. The Syk inhibitor, R406, (0.01 or 0.1 μM) or DMSO control (0.1%) for
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

9
48 hours at 37°C. Apoptosis was measured by gating on CD19+ or CD19- cells
and co-staining with Annexin V and 7-AAD.
Flow Cytometry
GC B cells and CD11c+ cells were labeled with anti-CD19 (Clone: eBio1D3), anti-
CD11c (Clone: N418), anti-CD80 (Clone:16-10A1), anti-CD86 (Clone: GL1), anti-
MHCII (Clone: M5/114.15.2), anti-GL7 (Clone: GL-7) (eBioscience), anti-CD4
(clone: RM4-5) anti-CD95 (Clone: Jo2) (BD Biosciences), anti-F4/80 (Clone:
BM8), anti-CXCR4 (Clone: L276F12), anti-Ly6G (Clone: 1A8), anti-CD3
(Clone:145-2C11), anti-Ly6C (Clone: HK1.4), anti-CD8 (Clone: 53-6.7), and anti-
CD11b (Clone: M1/70), (biolegend) were analyzed on BD LSRFortessa. Ficolled
human PBMC were stimulated and stained for Annexin V as previously
described.14, 15
Frozen tissue preparation, pathology and immunofluorescence Organs
harvested were embedded in Optimal Cutting Temperature compound, snap
frozen, and stored at -80°. GC detection and pathologic scoring were
accomplished as previously described4. At various times post transplantation,
GVHD target tissues were harvested, fixed in 10% formalin for 24hrs, embedded
in paraffin, and processed to generate 5µm-thick sections. H&E sections of skin
were examined in a blinded fashion using a semi quantitative scoring system for
GVHD as previously published.4, 16 Samples were scored from 0 to 4 for
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

10
epidermal and dermal inflammation, dermal fibrosis and subcutaneous fibrosis,
epidermal apoptosis, and loss of subcutaneous fat.
Statistics
Group comparisons of pathology, pulmonary function tests, cell counts and flow
cytometry data were analyzed by Student t-test or one-way ANOVA.
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

11
Results
Syk phosphorylation is increased during murine cGVHD and is necessary
in donor BM derived cells for the development and maintenance of cGVHD.
To determine if Syk activation is important in murine cGVHD, purified B
cells (Figure 1) from day 60 cGVHD spleens were assayed for Syk
phosphorylation. Purified B cells from cGVHD mice were activated with anti-IgM
and the amount of phosphorylated Syk at Y348 was measured. There was an
increase in the percentage pSyk in both BCR stimulated B-cells and unstimulated
B-cells in cGVHD (Figure 1). These data extend a recent report that human B-
cells have increased pSyk during cGVHD7.
To test whether Syk is necessary for cGVHD progression, mice were
transplanted with WT or induced Syk KO BM with WT T-cells and analyzed for
disease on day 60. Mice receiving Syk KO BM and WT T-cells did not develop
cGVHD pulmonary dysfunction compared to those receiving WT BM and T-cells
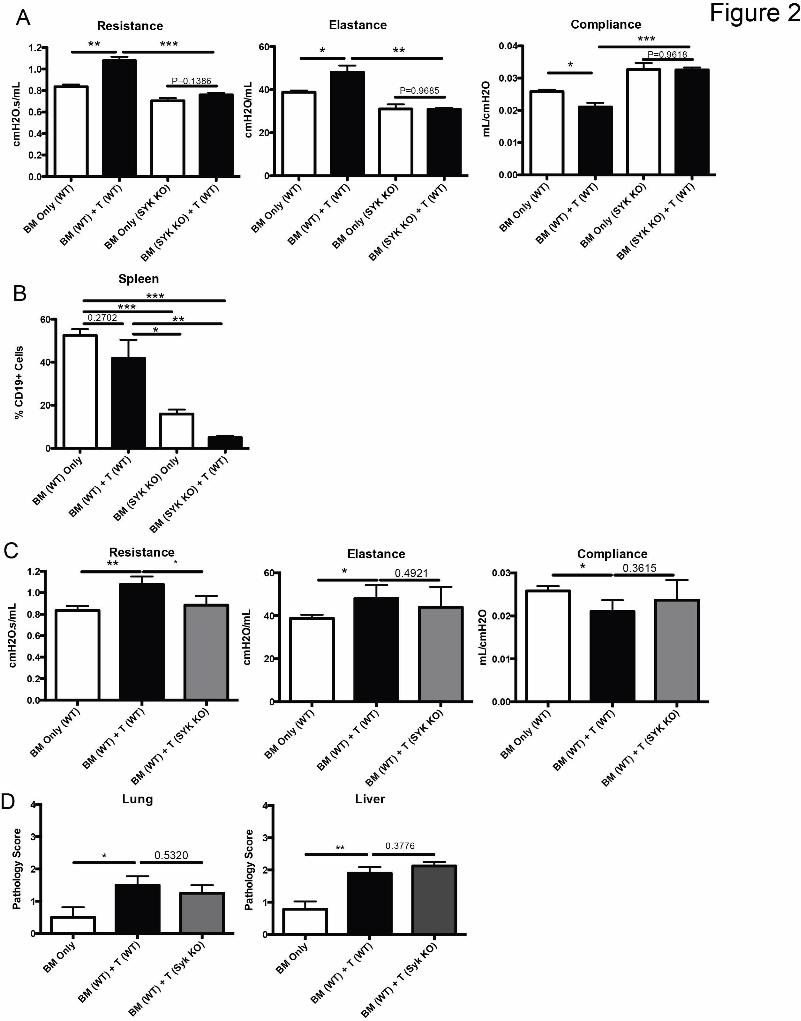
(Figure 2 A). Since Syk is necessary for the proliferation of B-cells following
antigen stimulation17, we analyzed the spleens of transplanted mice to determine
if there was a defect in the maintenance of Syk KO BM-derived B-cells. We found
that total B-cell frequency in mice receiving Syk KO BM was decreased 8-fold
(Figure 2B). These data are consistent with the dependency of activated B-cells
on Syk for proliferation and survival, and a requirement for activated donor-
derived BM B-cells in cGVHD pathogenesis3, 4.
In contrast to Syk dependence in BM derived cells, the addition of Syk KO
donor T-cells did not attenuate the functional manifestations of BO (with the
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

12
exception of resistance) compared to Syk WT T cell transplanted mice (Figure 2
C). In addition cGVHD pathology in the lungs and liver were similar in recipients
of Syk WT vs KO T cells (Figure 2D). While T cell effects after Syk inhibition were
previously described in acute18 and sclerodermatous cGVHD19, in this systemic
cGVHD model with BO and without scleroderma, donor T-cell Syk was not fully
required for cGVHD development.
To determine whether Syk function in BM-derived B cells was essential for
sustaining cGVHD in mice with established disease, recipients were given donor
BM from Syk fl/fl x ERT2-cre or WT mice with or without purified T cells. On D28
post-BMT, mice were given tamoxifen. Induced deletion of Syk in donor BM cells
significantly improved all pulmonary function parameters to levels of non-cGVHD
controls (Figure 3).
Therapeutic pharmacological inhibition of Syk decreases pathology in a
murine BO model of cGVHD but had little clinical therapeutic effect in
scleroderma models of cGVHD.
Fostamatinib is a potent small molecule inhibitor of Syk. Studies in
rheumatoid arthritis have demonstrated efficacy with fostamatinib in randomized
phase II clinical trials.20, 21 Fostamatinib was notably safe in patients treated for
non-Hodgkin’s lymphoma, including some who had received prior autologous
HCT.22. Additionally, Leonhardt et. al.18 demonstrated that Syk inhibition
decreased costimulatory molecules on antigen-presenting cells and increased
survival of mice during acute GVHD while preserving anti-tumor and anti-viral
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

13
immunity. In addition, other studies have demonstrated the importance of BCR
signaling for the development of murine cGVHD.23
Having demonstrated the requirement for Syk in donor BM cells in mice
with established cGVHD and BO, we next sought to determine whether drug
treatment with fostamatinib in mice with established cGVHD would be similarly
efficacious as Syk deletion in donor BM cells.3, 4 Mice receiving fostamatinib
beginning on D28 had restoration of pulmonary function, similar to the healthy
transplanted BM only controls (Figure 4 A). Improvement in pulmonary function
correlated with a reduction in cGVHD pathology in the lung (Figure 4B). The
number of GC reactions in the spleen was decreased in fostamatinib versus
vehicle treated cGVHD mice (Figure 4C). This was highlighted by a decrease in
frequency of splenic GC B-cells in fostamatinib treated cGVHD mice, matching
BM only controls (Figure 4D). These data demonstrate the importance of Syk in
B cell signaling during active disease.
Previous reports demonstrated a decrease in activation of CD11c+ cells18
in mice with acute GVHD treated with fostamatinib. In the BO cGVHD model we
found increased expression of costimulatory molecules CD80 and CD86 on
CD11c+ spleen cells compared to BM only transplanted mice. When these
cGVHD mice are treated with fostamatinib beginning D28, we found a significant
decrease in the frequency of CD11c+ cells expressing CD80 and CD86 though
without affecting MHC class II expression (Figure 4 E-G). These data highlight a
potentially beneficial alternative effect of Syk inhibition CD11c+ myeloid cells
during cGVHD.
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

14
In order to determine whether the effects of Syk inhibition attenuate
murine sclerodermatous cGVHD in our hands, we utilized three different cGVHD
scleroderma models involving lethally irradiated recipients: B6 into B6D2F1
model (model 2); G-CSF mobilized BALB/c splenocytes into B6 (model 3), and a
minor-mismatch model of B10.D2 into Balb/c (model 4). In the B6 into F1 model
(fostatinib, D14-28) and in the G-CSF-mobilized Balb/c into B6 models (fostatinib,
D7-21), the frequency of splenic CD19+ B cells in vehicle and fostatinib treated
cGVHD were >10- and 5-fold fold lower than non-cGVHD, BM only controls in
both the B6 into B6D2F1 and G-CSF Balb/c into B6 models, respectively (Figure
5 A and Supplemental Figure 1A). Neither a significant beneficial clinical
therapeutic nor pathological effect as reflected by total skin inflammation score
was seen in either model (Figure 5 B and Supplemental Figure 1B). However, we
found a significant decrease in peripheral blood CD11b+F4/80+ and Ly6Chi
monocytes numbers in the B6 into B6D2F1 model after treatment with
fostamatinib in the B6 into B6D2F1 model (Figure 5 C-E), which was also seen in
the G-CSF mobilized Balb/c into B6 model but did not reach statistical
significance (Supplemental Figure 1 C-E).
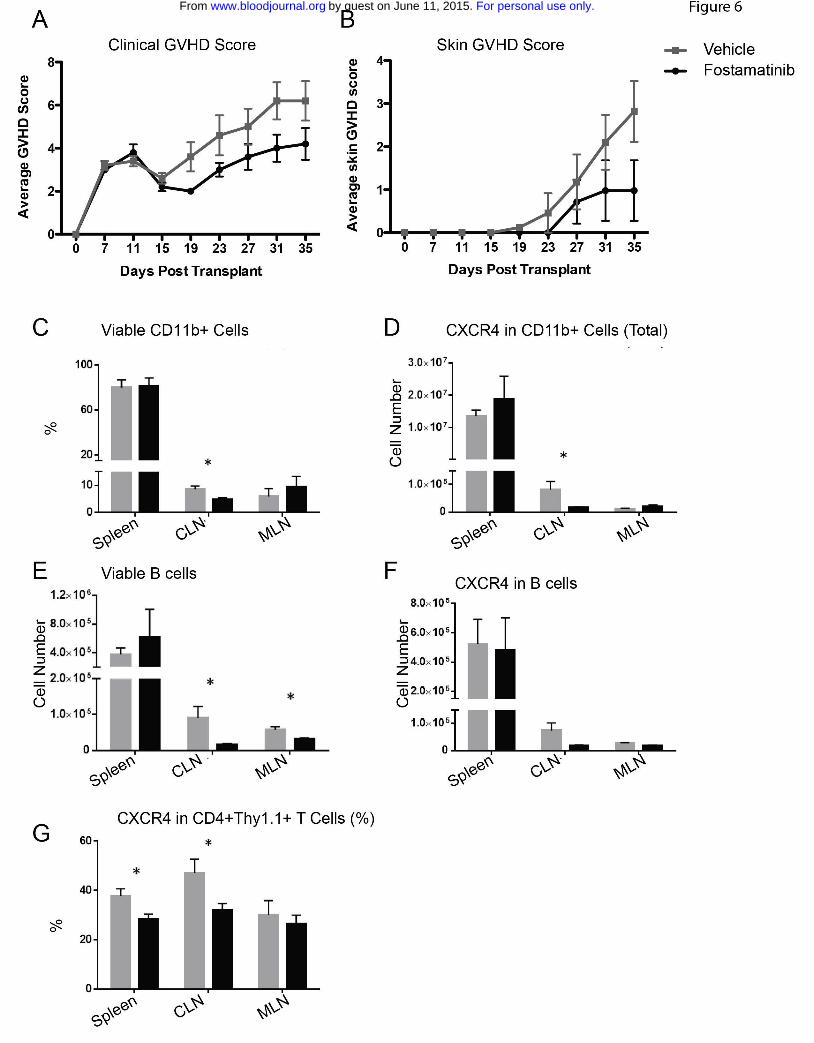
In the B10.D2 into Balb/c model of scleroderma, fostamatinib treatment
from D.21-35 did not impact skin disease or clinical score (not shown). However,
beginning fostatinib treatment 1 week earlier on D14, as used by Huu et. al.19, did
significantly improve these parameters on D19 and there was a reduction in the
both mean skin score from 3 to 1 and the GVHD clinical score from 6 to 4 in the
fostatinib vs vehicle treated controls on D35, the end of the study (Figure 6 A and
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

15
B). In the latter experiment, there also was a decrease in total numbers of viable
cervical and mesenteric lymph node (but not splenic) B cells and frequency of
CD11b+ cervical lymph node cells. CXCR4+ T cells in the spleen and cervical
lymph node were also decreased in fostamatinib vs vehicle treated cGVHD mice
(Figure 6 C-G), suggesting an impairment to the overall immune response and
migration of T cells and myeloid cells to tissues. Taken together, these data
suggest that early post-transplant treatment of sclerodermatous cGVHD may
attenuate disease progression.
Fostamatinib increases apoptosis in B cells purified from cGVHD patients.
To determine if human cGVHD B-cells are more susceptible to Syk blockade,
human peripheral blood (Supplementary Table 1) were treated in vitro with R406,
the active form of R788. B cells from patients with active cGVHD had increased
apoptotic and total cell death compared to patients with inactive or no cGVHD
after R406 (Figure 7). These data, consistent with work by Allen et. al,7 reveal
that R406 preferentially kills cGVHD B cells via apoptosis. Global targeting of B
cells with rituximab has been met with mixed success possibly due to altered B
cell homeostasis perpetuated in some patients.24, 25 We now demonstrate that
constitutively activated B cells can be selectively targeted in cGVHD by Syk
inhibition (Figure 7B). Consistent with the lack of a requirement for Syk in donor
T cells in the cGVHD BO model, fostamatinib did not induce apoptosis in the
non-B cell population (Figure 7C). In 3 of samples from each patient group, T-cell
specific anti-CD3 antibody was included to stimulate T-cell proliferation in the
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

16
presence of R406; no increase in T-cell death was observed under these
conditions. Together, these data point to the therapeutic potential of fostatinib
and pSyk targeting in cGVHD patients.
Discussion
Here, we demonstrated that pSyk was upregulated in murine cGVHD B
cells in a multi-organ system model of cGVHD with BO that is dependent upon
donor T cell support of antibody isotype switching and immunoglobulin
deposition. Such cGVHD B cells were hyper-responsive to B cell activation
signals in vitro. Syk function in donor BM-derived B cells but not T cells was
required for cGVHD generation and Syk deletion in donor BM cells in mice with
established cGVHD on D28 post-BMT reversed disease. A small molecule
inhibitor of pSyk, fostamatinib that has been tested in the clinic for autoimmune
diseases, was able to treat established cGVHD when therapy was initiated on
D28. In cGVHD patients, previous studies have shown that cGVHD B cells have
high levels of pSyk and in the current study, we now show that B cells from
patients with active cGVHD but not B cells from patients without active cGVHD
was induced into apoptosis in vitro by the active metabolite of fostamatinib.
Whereas fostamatinib was highly effective in reversing cGVHD in the BO model,
clinical and immunological effects in sclerodermatous cGVHD appeared to be
more modest under the conditions tested.
It is unclear if B cells are intrinsically or extrinsically affected during
cGVHD. B cells might increase their activation state based on pathogen
activating molecular patterns released during conditioning treatment and
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

17
activating toll-like receptors on B cells. In contrast there is potential that
increased activating factors such as B cell activating factor (BAFF) are causing B
cells to increase response from factors such as Syk since BAFF receptor may be
reliant on Syk for its signaling.26 The requirement of Syk in BM derived B cells for
cGVHD development in the BO model also allowed us to elucidate a potential
mechanism of disease. BCR stimulation with antigen leads to phosphorylation of
proximally located Syk to initiate down-stream signaling for B cell survival and
proliferation. Syk signaling is necessary for survival of B cells during the GC
reaction. The decrease in B cells after Syk-deficient BM transplant in the BO
model suggests a mechanistic link between BCR-antigen engagement that
requires Syk for B cell promotion and perpetuation. The absence or relatively
modest effects of fostamatinib in sclerodermatous cGVHD is consistent with the
findings that there were few splenic B cells present in our studies in fostamatinib
or vehicle treated sclerodermatous cGVHD mice when analyzed 4-5 weeks post-
transplant. These B cell data contrast to the significantly increased splenic GC B
cell frequency seen in cGVHD mice with BO.3, 4
Studies of cGVHD patient peripheral blood have demonstrated an
important role of B cells in development of disease. The production of auto- and
allo-antibodies has previously been associated with the development of cGVHD
in patients.27-30 The dysregulation of B cells could be due to the significant
increase in BAFF present during cGVHD.15, 31 B cells in patients with cGVHD
were more sensitive to Syk inhibition compared to B cells from patients without
active cGVHD, consistent with our previous finding that these cells have
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

18
increased activation of Syk and the proximal BCR signalosome7. We
demonstrated that these cells had an increase in cell death rates compared to
controls. Like BCR-activated B cell lymphomas32, only those cGVHD B cells
activated through the BCR pathway are presumably susceptible to killing at low
drug concentration. Together these data demonstrate the relative selective ability
of fostamatinib to target the activated B cells in cGVHD patients.
The role of Syk in donor T cells was demonstrated to be of little
importance in the development of murine cGVHD in the BO model. However in
the minor antigen-disparate sclerodermatous cGVHD model, we did find a
decrease in the expression of CXCR4 on T cells when mice with established
clinical cGVHD were treated with R788, consistent with Huu et al.19 who also
demonstrated increased pSyk in T cells on day 14 post-transplant also showed a
reduction in proliferation of T cells from cGVHD mice when exposed in vitro to
the active fostamatinib metabolite, R406 and mice (4-6 per group) treated with
fostamatinib in vivo exhibited ~1 week shift in cGVHD onset and progression
when treatment was initiated on day 14 post-transplant, resulting in significantly
lower average skin scores and body weight between days 21-42 post-
transplant.19 The decrease in migration of T cells to effector tissues could have a
beneficial effect on the development of disease. Thus, while treatment did not
fully restore health in our study in the B10.D2 into Balb/c mode, there was a
decrease in the progression of disease associated with a lowered frequency of
CXCR4+ T cells that may contribute to disease reduction.
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

19
Despite the apparent lack of Syk dependency of donor T cells in cGVHD
with BO, we demonstrate that the Syk inhibitor fostamatinib is capable of
blocking the cGVHD GC response that is dependent upon T follicular helper cells
and GC B cells.3 These data point to a T cell independent effect of fostamatinib
in abrogating the cGVHD GC response, which is essential for cGVHD generation
and progression in the BO model.
In addition to targeting B cells during cGVHD, there is evidence that
antigen-presenting cells are susceptible to fostamatinib treatment. For example,
in an acute GVHD model, Leonhardt et. al.18 showed that fostamatinib reduced
costimulatory molecule expression and disrupted cytoskeletal organization, along
with blocking T cell proliferation and migration, resulting in acute GVHD
amelioration with preservation of graf-versus-leukemia and anti-viral immunity.
Consistent with other reports that inhibition of Syk can decrease costimulatory
molecules expression.19, 33-35 we demonstrated in vivo decrease in the frequency
of activated (CD80+, CD86+) DCs in mice with cGVHD and BO.
Although we could not discern effects of fostamatinib on the total skin
inflammatory scores in either MHC-disparate models of sclerodermatous
cGVHD, in the B6 into B6D2F1 model, fostamatinib significantly decreased the
absolute number of CD11b+F4/80+ monocytes and the inflammatory Ly6Chi but
not the Ly6Clo non-inflammatory subset in peripheral blood. These data are
interesting in light of the role of Syk in macrophages and monocytes and our
recent study implicating monocytes and macrophages in cGVHD generation and
maintenance in the two scleroderma cGVHD models as well as in the BO model5.
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

20
Notably, there was a significant decrease in CD11b+ cells in the cervical lymph
node of fostamatinib treated mice in the minor antigen disparate B10.D2 into
Balb/c sclerodermous cGVHD model. Monocyte and macrophages may
contribute to cGVHD in these models by antigen-presentation capacity or the
elaboration of pro-fibrogenic proteins such as TGFb5. Thus, the impact of
fostamatinib in cGVHD models may depend in part as to the stage of disease
progression as well as the dominant underlying mechanisms for disease
pathogenesis.
In summary, our data clearly indicate by biochemical and genetic and
pharmacological approaches a role of Syk in donor BM-derived B cells in a multi-
organ system model of cGVHD with BO. Further we have translated these
findings into a therapeutic using a pharmacological small molecule Syk inhibitor,
fostamatinib, that has progressed through phase II clinical trials20, 21 Fostamatinib
ameliorated disease in mice with cGVHD and BO and has biological effects in
some models of scleroderma. These preclinical murine and human data point to
consideration of clinical trials of fostamatinib or other Syk inhibitor alone or in
combination with other similar agents23 for the treatment of cGVHD.
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

21
Acknowledgements. We thank Rigel Pharmaceuticals for providing
Fostamatinib (R788/R406). Supported in part by National Institutes of Health
National Cancer Institute grants P01 CA142106-06A1 and 5P01-CA047741-20,
National Institutes of Allergy and Infectious Diseases grants P01 AI 056299 and
T32 AI 007313 and K08HL107756 and Leukemia and Lymphoma Society
Translational Research Grants 6458-15 and 6462-15.
Author contributions
RF designed experiments, performed experiments, and wrote the paper. JLA, LL,
KPM, KP, JP, KAA, JD, and JCP designed and performed experiments. AP-M
performed histological analyses, discussed experimental design, and edited the
paper. PAT performed experiments and edited the paper. JSS, WJM, GRH,
KPM, LL, IM, JK, CSC, RJS, and JR designed experiments and edited the paper;
NJC and RAC provided reagents, discussed experiments and edited the paper,
SS and BRB designed experiments and edited the paper.
Conflict of interest
No conflicts of interest are known.
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

22
Literature Cited
1. Schultz KR, Miklos DB, Fowler D, Cooke K, Shizuru J, Zorn E et al. Toward
biomarkers for chronic graft-versus-host disease: National Institutes of
Health consensus development project on criteria for clinical trials in chronic
graft-versus-host disease: III. Biomarker Working Group Report. Biol Blood
Marrow Transplant 2006; 12(2): 126-137. e-pub ahead of print 2006/01/31;
doi: 10.1016/j.bbmt.2005.11.010
2. Shlomchik WD, Lee SJ, Couriel D, Pavletic SZ. Transplantation's greatest
challenges: advances in chronic graft-versus-host disease. Biol Blood Marrow
Transplant 2007; 13(1 Suppl 1): 2-10. e-pub ahead of print 2007/01/16; doi:
10.1016/j.bbmt.2006.10.020
3. Flynn R, Du J, Veenstra RG, Reichenbach DK, Panoskaltsis-Mortari A, Taylor
PA et al. Increased T follicular helper cells and germinal center B cells are
required for cGVHD and bronchiolitis obliterans. Blood 2014; 123(25): 3988-
3998. e-pub ahead of print 2014/05/14; doi: 10.1182/blood-2014-03-
562231
4. Srinivasan M, Flynn R, Price A, Ranger A, Browning JL, Taylor PA et al. Donor
B-cell alloantibody deposition and germinal center formation are required
for the development of murine chronic GVHD and bronchiolitis obliterans.
Blood 2012; 119(6): 1570-1580. e-pub ahead of print 2011/11/11; doi:
10.1182/blood-2011-07-364414
5. Alexander KA, Flynn R, Lineburg KE, Kuns RD, Teal BE, Olver SD et al. CSF-1-
dependant donor-derived macrophages mediate chronic graft-versus-host
disease. J Clin Invest 2014; 124(10): 4266-4280. doi: 10.1172/JCI75935
6. Radojcic V, Pletneva MA, Yen HR, Ivcevic S, Panoskaltsis-Mortari A, Gilliam AC
et al. STAT3 signaling in CD4+ T cells is critical for the pathogenesis of
chronic sclerodermatous graft-versus-host disease in a murine model. J
Immunol 2010; 184(2): 764-774. e-pub ahead of print 2009/12/10; doi:
10.4049/jimmunol.0903006
7. Allen JL, Tata PV, Fore MS, Wooten J, Rudra S, Deal AM et al. Increased BCR
responsiveness in B cells from patients with chronic GVHD. Blood 2014. e-
pub ahead of print 2014/02/18; doi: 10.1182/blood-2013-10-533562
8. Leibundgut-Landmann S, Osorio F, Brown GD, Reis e Sousa C. Stimulation of
dendritic cells via the dectin-1/Syk pathway allows priming of cytotoxic T-
cell responses. Blood 2008; 112(13): 4971-4980. doi: 10.1182/blood-2008-
05-158469
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

23
9. Gevrey JC, Isaac BM, Cox D. Syk is required for monocyte/macrophage
chemotaxis to CX3CL1 (Fractalkine). J Immunol 2005; 175(6): 3737-3745.
10. Anderson BE, McNiff JM, Jain D, Blazar BR, Shlomchik WD, Shlomchik MJ.
Distinct roles for donor- and host-derived antigen-presenting cells and
costimulatory molecules in murine chronic graft-versus-host disease:
requirements depend on target organ. Blood 2005; 105(5): 2227-2234. e-
pub ahead of print 2004/11/04; doi: 10.1182/blood-2004-08-3032
11. Burman AC, Banovic T, Kuns RD, Clouston AD, Stanley AC, Morris ES et al.
IFNgamma differentially controls the development of idiopathic pneumonia
syndrome and GVHD of the gastrointestinal tract. Blood 2007; 110(3): 1064-
1072. doi: 10.1182/blood-2006-12-063982
12. Panoskaltsis-Mortari A, Tram KV, Price AP, Wendt CH, Blazar BR. A new
murine model for bronchiolitis obliterans post-bone marrow transplant.
American journal of respiratory and critical care medicine 2007; 176(7): 713-
723. e-pub ahead of print 2007/06/19; doi: 10.1164/rccm.200702-335OC
13. Beers SA, Chan CH, James S, French RR, Attfield KE, Brennan CM et al. Type II
(tositumomab) anti-CD20 monoclonal antibody out performs type I
(rituximab-like) reagents in B-cell depletion regardless of complement
activation. Blood 2008; 112(10): 4170-4177. e-pub ahead of print
2008/06/28; doi: 10.1182/blood-2008-04-149161
14. Pullerits R, Brisslert M, Jonsson IM, Tarkowski A. Soluble receptor for
advanced glycation end products triggers a proinflammatory cytokine
cascade via beta2 integrin Mac-1. Arthritis and rheumatism 2006; 54(12):
3898-3907. e-pub ahead of print 2006/11/30; doi: 10.1002/art.22217
15. Allen JL, Fore MS, Wooten J, Roehrs PA, Bhuiya NS, Hoffert T et al. B cells from
patients with chronic GVHD are activated and primed for survival via BAFF-
mediated pathways. Blood 2012; 120(12): 2529-2536. doi: 10.1182/blood-
2012-06-438911
16. Hill GR, Crawford JM, Cooke KR, Brinson YS, Pan L, Ferrara JL. Total body
irradiation and acute graft-versus-host disease: the role of gastrointestinal
damage and inflammatory cytokines. Blood 1997; 90(8): 3204-3213. e-pub
ahead of print 1997/10/24;
17. Craxton A, Jiang A, Kurosaki T, Clark EA. Syk and Bruton's tyrosine kinase are
required for B cell antigen receptor-mediated activation of the kinase Akt. J
Biol Chem 1999; 274(43): 30644-30650. e-pub ahead of print 1999/10/16;
18. Leonhardt F, Zirlik K, Buchner M, Prinz G, Hechinger AK, Gerlach UV et al.
Spleen tyrosine kinase (Syk) is a potent target for GvHD prevention at
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

24
different cellular levels. Leukemia 2012; 26(7): 1617-1629. e-pub ahead of
print 2012/02/04; doi: 10.1038/leu.2012.10
19. Le Huu D, Kimura H, Date M, Hamaguchi Y, Hasegawa M, Hau KT et al.
Blockade of Syk ameliorates the development of murine sclerodermatous
chronic graft-versus-host disease. Journal of dermatological science 2014;
74(3): 214-221. doi: 10.1016/j.jdermsci.2014.02.008
20. Weinblatt ME, Kavanaugh A, Burgos-Vargas R, Dikranian AH, Medrano-
Ramirez G, Morales-Torres JL et al. Treatment of rheumatoid arthritis with a
Syk kinase inhibitor: a twelve-week, randomized, placebo-controlled trial.
Arthritis Rheum 2008; 58(11): 3309-3318. e-pub ahead of print 2008/11/01;
doi: 10.1002/art.23992
21. Genovese MC, Kavanaugh A, Weinblatt ME, Peterfy C, DiCarlo J, White ML et
al. An oral Syk kinase inhibitor in the treatment of rheumatoid arthritis: a
three-month randomized, placebo-controlled, phase II study in patients with
active rheumatoid arthritis that did not respond to biologic agents. Arthritis
Rheum 2011; 63(2): 337-345. doi: 10.1002/art.30114
22. Herlands RA, Christensen SR, Sweet RA, Hershberg U, Shlomchik MJ. T cell-
independent and toll-like receptor-dependent antigen-driven activation of
autoreactive B cells. Immunity 2008; 29(2): 249-260. e-pub ahead of print
2008/08/12; doi: 10.1016/j.immuni.2008.06.009
23. Dubovsky JA, Flynn R, Du J, Harrington BK, Zhong Y, Kaffenberger B et al.
Ibrutinib treatment ameliorates murine chronic graft-versus-host disease. J
Clin Invest 2014; 124(11): 4867-4876. doi: 10.1172/JCI75328
24. Sarantopoulos S, Stevenson KE, Kim HT, Washel WS, Bhuiya NS, Cutler CS et
al. Recovery of B-cell homeostasis after rituximab in chronic graft-versus-
host disease. Blood 2011; 117(7): 2275-2283. e-pub ahead of print
2010/11/26; doi: 10.1182/blood-2010-10-307819
25. Cutler C, Kim HT, Bindra B, Sarantopoulos S, Ho VT, Chen YB et al. Rituximab
prophylaxis prevents corticosteroid-requiring chronic GVHD after allogeneic
peripheral blood stem cell transplantation: results of a phase 2 trial. Blood
2013; 122(8): 1510-1517. e-pub ahead of print 2013/07/19; doi:
10.1182/blood-2013-04-495895
26. Schweighoffer E, Vanes L, Nys J, Cantrell D, McCleary S, Smithers N et al. The
BAFF receptor transduces survival signals by co-opting the B cell receptor
signaling pathway. Immunity 2013; 38(3): 475-488. e-pub ahead of print
2013/03/05; doi: 10.1016/j.immuni.2012.11.015
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

25
27. Tazzari PL, Gobbi M, Zauli D, Tassinari A, Crespi C, Miserocchi F et al. Close
association between antibodies to cytoskeletal intermediate filaments, and
chronic graft-versus-host disease. Transplantation 1987; 44(2): 234-236.
28. Svegliati S, Olivieri A, Campelli N, Luchetti M, Poloni A, Trappolini S et al.
Stimulatory autoantibodies to PDGF receptor in patients with extensive
chronic graft-versus-host disease. Blood 2007; 110(1): 237-241.
29. Miklos DB, Kim HT, Miller KH, Guo L, Zorn E, Lee SJ et al. Antibody responses
to H-Y minor histocompatibility antigens correlate with chronic graft-versus-
host disease and disease remission. Blood 2005; 105(7): 2973-2978. e-pub
ahead of print 2004/12/23; doi: 10.1182/blood-2004-09-3660
30. Zorn E, Miklos DB, Floyd BH, Mattes-Ritz A, Guo L, Soiffer RJ et al. Minor
histocompatibility antigen DBY elicits a coordinated B and T cell response
after allogeneic stem cell transplantation. J Exp Med 2004; 199(8): 1133-
1142. e-pub ahead of print 2004/04/21; doi: 10.1084/jem.20031560
31. Sarantopoulos S, Stevenson KE, Kim HT, Cutler CS, Bhuiya NS, Schowalter M
et al. Altered B-cell homeostasis and excess BAFF in human chronic graft-
versus-host disease. Blood 2009; 113(16): 3865-3874.
32. Chen L, Monti S, Juszczynski P, Daley J, Chen W, Witzig TE et al. SYK-
dependent tonic B-cell receptor signaling is a rational treatment target in
diffuse large B-cell lymphoma. Blood 2008; 111(4): 2230-2237.
33. Christensen SR, Shlomchik MJ. Regulation of lupus-related autoantibody
production and clinical disease by Toll-like receptors. Seminars in
immunology 2007; 19(1): 11-23. e-pub ahead of print 2007/02/06; doi:
10.1016/j.smim.2006.12.005
34. Lin YC, Huang DY, Chu CL, Lin WW. Anti-inflammatory actions of Syk
inhibitors in macrophages involve non-specific inhibition of toll-like
receptors-mediated JNK signaling pathway. Molecular immunology 2010;
47(7-8): 1569-1578. e-pub ahead of print 2010/02/09; doi:
10.1016/j.molimm.2010.01.008
35. Yan J, Harvey BP, Gee RJ, Shlomchik MJ, Mamula MJ. B cells drive early T cell
autoimmunity in vivo prior to dendritic cell-mediated autoantigen
presentation. J Immunol 2006; 177(7): 4481-4487. e-pub ahead of print
2006/09/20;
36. Allen JL WJ, Fore M, Bhuiya NS, Armistead P, Coghill JM, Gabriel D, Roehrs PA,
Sharf A, Irons RN, Hoffert T, Richards K, Shea TC, Baldwin AS, Serody J and
Sarantopoulos S. B Cells from patients with chronic GVHD signal via the AKT-
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

26
driven survival and metabolic fitness pathway. In: The American Society of
Bone Marrow Transplantation. San Diego, CA: Elsevier, 2012.
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

27
Figure Legends
Figure 1: Syk activation during cGVHD. B cells were purified from the spleens
of healthy control (BM Only) or cGVHD (BM + T) mice and stimulated with anti-
IgM at 5 μg/mL. Splenocytes from B10.BR mice transplanted with B6 BM and low
numbers of T cells were analyzed for phosphorylated Syk at Y348 after ex vivo
stimulation by 5 μg/mL of anti-IgM* p < 0.5; Error bars represent SEM, n=8
representative data from 2 experiments.
Figure 2: Presence of Syk in BM derived B cells but not donor T cells is
necessary for development of cGVHD pathology in BO model. A) Day 60
pulmonary function tests of mice transplanted with taxomifen induced Syk KO
BM and WT T cells. B) Frequency of B cells in transplanted mice on D60 after
transplant. C) Pulmonary function tests from mice transplanted with WT BM and
either WT T cells or Syk KO T cells. D) Pathology scores from lung and liver of
mice transplanted with Syk KO T cells. * p < 0.5; ** p < 0.01. Error Bars
represent SEM, Representative data from 3 independent experiments with n=8
per group.
Figure 3: Genetic deletion of Syk during active disease prevents
pathogenic pulmonary function. Mice were transplanted with either WT or Syk
fl/fl x ERT2-Cre BM. On D8 mice were treated with Tamoxifen for 5 days to
delete Syk in donor BM derived cells. A) Pulmonary function tests of mice on day
60. * p < 0.5; ** p < 0.01. Error Bars represent SEM, n=10 per group.
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

28
Figure 4: Inhibition of Syk by R788 decreases cGVHD in murine BO model.
A) Day 56 pulmonary function tests on mice treated with 30 mg/Kg of R788 twice
daily from D28-56. B) GVHD pathology scores from the lungs of mice on D60. C)
Number of GCs present in situ in spleens of mice. D) Frequency of GC B cells
(gated on CD19+GL7+CD95hi) present in the spleens of transplanted mice on day
60. E-G) Frequency of CD11c cells expressing CD80, CD86 and MHC Class II in
the spleens of transplanted mice on day 60. * p < 0.5; ** p < 0.01; *** p< 0.001.
Error Bars represent SEM, Representative data from 3 independent experiments
with n=8 per group.
Figure 5: Fostamatinib does not alter skin inflammation in B6 into B6D2F1
model of sclerodermatous GVHD but decreases peripheral blood Ly6Chi
monocytes. A) Frequency of CD19+ B cells in spleens of mice on day 21 in
vehicle treated (grey bars), R788 treated (black bars) and non-GVHD controls
(white bars). B) Total inflammation score. C) Absolute number of CD11b+ F4/80+
peripherial blood monocytes. Peripherial monocytes were examines for D) Ly6C
high or E) Ly6C low expression. * p < 0.5; ** p < 0.01; Error bars represent SEM,
n=6 per group.
Figure 6: Fostamatinib has little clinical therapeutic benefit in the B10.D2
into Balb/c sclerodermatous cGVHD model but decreases expression of
CXCR4 on CD11b+ and CD4+ cells. A) Clinical GVHD scores or b) Skin GVHD
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

29
Scores in vehicle treated (grey squares) or R788 treated (black circles) in B10.D2
into Balb/c model. C) Frequency of viable CD11b+ cells and D) expression of
CXCR4 on CD11b+ cells. E) Absolute number CD19+ B cells and F) expression
of CXCR4 on B cells G) expression of CXCR4 on CD4+ cells. * p < 0.5; Error
bars represent SEM. n=5 per group.
Figure 7: Increased apoptosis in human B cells when treated with R788. (A)
Representative flow plots of human PBMC with or without cGVHD. Consistent
with previous work36 unmanipulated, untreated cGVHD B cells had superior
survival compared to B cells from patients without disease. Peripheral blood
mononuclear cells from patients without cGVHD (n = 6, open circles) and with
cGVHD (n = 6, filled squares) treated with R406 (0, 0.01, and 0.1 μM) as
indicated for 48 h. Apoptotic B cells were defined as CD19+ Annexin V+ 7AAD-
cells (B) or as CD19 negative non-B cells (C). Fold increase in apoptosis by
R406 divided by PBS is depicted. Data are median +/- range pooled from 2
independent experiments. * p < 0.5; ** p < 0.01; *** p < 0.001.
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom

doi:10.1182/blood-2014-08-595470Prepublished online April 7, 2015;
H. Antin, Jerome Ritz, Nelson J. Chao, Raphael A. Clynes, Stefanie Sarantopoulos and Bruce R. BlazarWilliam J. Murphy, Geoffrey R. Hill, Ivan Maillard, John Koreth, Corey S. Cutler, Robert J. Soiffer, JosephVulic, Jing Du, Angela Panoskaltsis-Mortari, Patricia A. Taylor, Jonathan C. Poe, Jonathan S. Serody, Ryan Flynn, Jessica L. Allen, Leo Luznik, Kelli P. MacDonald, Katelyn Paz, Kylie A. Alexander, Ante graft-versus-host diseaseTargeting Syk Activated B-cells in murine and human chronic
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
digital object identifier (DOIs) and date of initial publication. indexed by PubMed from initial publication. Citations to Advance online articles must include final publication). Advance online articles are citable and establish publication priority; they areappeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet
Copyright 2011 by The American Society of Hematology; all rights reserved.Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society of
For personal use only.on June 11, 2015. by guest www.bloodjournal.orgFrom