takuya nakazato 2,6 and elizabeth a. housworth 3 – 5 - umdrive
TRANSCRIPT

American Journal of Botany 98(1): 88–98. 2011.
88
American Journal of Botany 98(1): 88–98, 2011; http://www.amjbot.org/ © 2011 Botanical Society of America
Geography plays an important role in various evolutionary processes in many, if not most, natural populations. For exam-ple, geographic features such as mountains, continents, and riv-ers serve as physical barriers to gene fl ow among populations (e.g., Knowlton et al., 1993 ; Duvernell and Turner, 1998 , 1999 ) and/or induce ecological isolation due to differences in envi-ronmental conditions between populations (e.g., Macnair, 1983 ; Hawthorne and Via, 2001 ; Wilding et al., 2001 , reviewed by Coyne and Orr [2004] ). In many species, geographic distance between populations alone has signifi cant effects on patterns of dispersal and gene fl ow (e.g., Handley et al., 2007 ; Platt et al., 2010 ). Understanding how geography affects adaptation demo-graphic history, species divergence, and patterns of dispersal and hybridization is an important step toward deciphering the mechanisms of evolutionary processes in nature ( Manel et al., 2003 ; Givnish, 2010 ; Hickerson et al., 2010 ). Here, we investi-gate roles of the complex geographic features of the Andes in the demographic history of two closely related wild tomato spe-cies that inhabit on the opposite sides of the mountain chain.
Wild and cultivated tomatoes form a small monophyletic clade consisting of 13 closely related species within the large and diverse genus Solanum ( Peralta et al., 2008 ). All members of the tomato clade are diploids (2 n = 24; Peralta and Spooner, 2001 ;
Nesbitt and Tanksley, 2002 ) that share a high degree of ge-nomic synteny ( Chetelat and Ji, 2007 ) and are to some degree intercrossable ( Rick, 1979 ). The center of diversifi cation of wild tomato species (and of the genus) is from Ecuador to Chile, along the Andean mountains and the adjacent Pacifi c coast. The recent rapid uplift of the Andes since the Eocene (~55 million years ago [Ma]; Campbell, 1975 ; Stibane, 1975 ) has created a dramatic altitudinal cline (sea level to > 6500 m a.s.l.) and cor-respondingly diverse climate zones ranging from temporal des-ert along the western coast to seasonable highlands in the Andes to tropical rainforests in the Amazon basin all within 500 km ( Young et al., 2002 ). Despite the recent divergence of the to-mato group ( < 6 Ma; Rodriguez et al., 2009 ), each species dis-plays a distinct geographical distribution pattern and habitat use across this environmentally diverse region ( Rick, 1973 , 1978 , 1979 ; Taylor, 1986 ; Smith and Peralta, 2002 ; Nakazato et al., 2010 ). They are also morphologically diverse, and some traits are likely adaptive responses to their habitats ( Rick, 1973 , 1976 ; Patterson et al., 1978 ; Rick, 1978 ; Vallejos, 1979 ; Bloom et al., 2004 ; Nakazato et al., 2008 ), suggesting that abiotic ecological conditions likely play a critical role in their phenotypic evolu-tion and divergence. This combination of close genetic relation-ships among species and high ecological and phenotypic divergence makes wild tomato species ideal for studying local adaptation and ecological divergence.
The study species, Solanum lycopersicum L. ( SL ) and S. pimpinellifolium L. ( SP ) are genetically closely related sister species of red-fruited tomatoes (~0.8% silent site divergence, ~1.3 Ma divergence; Nesbitt and Tanksley, 2002 ). Solanum ly-copersicum is a large-fruited species that includes the cultivated tomato, while SP is a small-fruited species, often referred to as
1 Manuscript received 25 July 2010; revision accepted 1 November 2010. The authors thank the Tomato Genetics Resource Center for the sample
seeds and geographic information of the tomato accessions and two anonymous reviewers for insightful suggestions.
6 Author for correspondence (e-mail: [email protected])
doi:10.3732/ajb.1000272
SPATIAL GENETICS OF WILD TOMATO SPECIES REVEALS ROLES OF THE ANDEAN GEOGRAPHY ON DEMOGRAPHIC HISTORY1
Takuya Nakazato 2,6 and Elizabeth A. Housworth 3 – 5
2 Department of Biological Sciences, The University of Memphis, 3774 Walker Avenue, Memphis, Tennessee 38152 USA; 3 Department of Mathematics, 831 East 3rd Street, Indiana University, Bloomington, Indiana 47405 USA; 4 Department of
Biology; and 5 Department of Statistics, Indiana University, Bloomington, Indiana, USA
• Premise of the study : Understanding the demographic history of natural populations in relation to the geographic features in their habitats is an important step toward deciphering the mechanisms of evolutionary processes in nature. This study investi-gates how the complex geographic and ecological features of the Andes play a role in demographic history, species divergence, dispersal patterns, and hybridization in wild tomato species.
• Methods : We investigated spatial genetics of two closely related wild tomato species, Solanum lycopersicum and S. pimpinel-lifolium , by integrating amplifi ed fragment length polymorphism (AFLP) marker data and geographic information system (GIS)-derived geographic and climatic data.
• Key results : The two species represent genetically distinct lineages largely separated by the Andes, but hybridize extensively in central to northern Ecuador, likely mediated by the transitional climatic conditions between those of the two species. Sola-num lycospericum has likely experienced a severe population bottleneck during the colonization of the eastern Andes followed by a rapid population expansion.
• Conclusions : The study demonstrates that the evolutionary patterns of the two wild tomatoes, including demographic history, dispersal patterns, interspecifi c divergence, and hybridization, are intimately related to the complex geographic and ecological features of the Andes. Integrating genetic data across the genome and GIS-derived environmental data can provide insights into the patterns of complex evolutionary processes in nature.
Key words: AFLP; Andes; demography; geographic information system; hybridization; Solanaceae; Solanum ; spatial genet-ics; speciation; tomato.

89January 2011] Nakazato and Housworth — Spatial genetics of wild tomatoes
by geography and climatic conditions. Results from this study will help us better understand the interaction of geographic and ecological factors in adaptation, divergence, and hybridization in nature.
MATERIALS AND METHODS
Study samples — We obtained seeds of the study accessions representing 14 SL and 19 SP populations (Appendix 1), from the Tomato Genetics Re-source Center (TGRC) at the University of California at Davis. We chose study populations that spanned each species ’ known geographic range in South America based on the coordinates of the original collection localities available at TGRC. Wild SL populations are closely related to the domesticated tomato, and SP populations often occur near tomato cultivations. Because domesticated tomatoes are often traded over long distances, which can strongly affect phylo-geographic inferences, we carefully chose wild populations that are unlikely to have been cultivated on the basis of their descriptions available at TGRC. Therefore, phylogeographic relationships should not be obscured. Seeds were germinated and plants were grown in the University of Memphis greenhouse according to the recommendations by TGRC. Before the present study, 10 plants from each population were grown to maturity and mass-pollinated to ensure that their offspring are in Hardy – Weinberg equilibrium. Ten to twelve of the resulting seeds from each population were chosen, and genomic DNA was extracted from each seedling using a MagAttract 96 Miniprep Core Kit (Qiagen, Valencia, California, USA) for AFLP genotyping.
AFLP genotyping — AFLP genotyping was conducted as in Nakazato et al. (2008) using eight EcoR I- Msp I primer combinations with selective extensions of ACG (NED dye)-AGT, AAC (NED)-ATC, ACT (FAM)-AGT, ATG (FAM)-ATC, ACC (VIC)-AGT, ATC (VIC)-ATC, ACA (PET)-AGT, ATA (PET)-ATC. PCR products were run on a 3730 DNA Analyzer (Applied Biosystems, Foster City, California, USA) at the Molecular Institute at Indiana University. All unambiguously scorable fragments between 50 – 500 bp, segregating in at least one individual, were scored manually as present (1) or absent (0) using the GeneMapper software (Applied Biosystems).
Genetic analyses — Wright ’ s fi xation indices ( F ST ’ s) and expected heterozy-gosity under Hardy – Weinberg equilibrium (Nei ’ s gene diversity) were esti-mated using the program AFLP-SURV version 1.0 ( Vekemans, 2002 ) based on all the scored AFLPs using the Bayesian method with uniform prior distribu-tion, assuming Hardy – Weinberg equilibrium within populations. Pairwise F ST values among accessions were estimated based on 1000 permutations. Analysis of molecular variance (AMOVA) and estimation of pairwise population geo-graphic distances were conducted using the program genalex 6 ( Peakall and Smouse, 2006 ). All Mantel tests were conducted using the zt program ( Bonnet and van de Peer, 2002 ) with 30 000 randomizations. Because these tests were used to test for isolation by distance (IBD), they were treated as one-tailed. Population structure analyses were conducted using the STRUCTURE soft-ware ( Pritchard et al., 2000 ) with 100 000 replicates and 10 000 burn-in periods using the recessive alleles model without assigning each population to pre-defi ned clusters. The analysis was repeated for two, three, and four clusters ( K ) with three independent runs each. A Fitch – Margoliash phylogeny was con-structed based on pairwise F ST values using the FITCH program of PHYLIP ( Felsenstein, 1989 ). We estimated the supports for internal branches using the CONSENSE program of PHYLIP based on 1000 pairwise F ST tables by boot-strapping both AFLP markers and individuals within populations. All other statistical analyses were conducted using the program SPSS version 14.0 (SPSS, Chicago, Illinois, USA).
RESULTS
Patterns of AFLP polymorphism — A total of 127 polymor-phic AFLP loci were found from eight primer combinations with 112 (88.2%) and 124 (97.6%) segregating within SL and SP , respectively. In contrast, an ongoing AFLP analysis of self-incompatible wild tomato species, S. peruvianum and S. pen-nellii , using the same eight primer combinations, found 427 polymorphic loci among 19 populations and 1050 loci among
the currant tomato. Despite their close genetic relationship, they are geographically, ecologically, and morphologically distinct ( Nakazato et al., 2008 , 2010 ). Solanum lycopersicum largely occurs in the warm and wet environments of the edge of the Amazon basin (mean annual precipitation of 2061 mm at known population localities) on the eastern Andes and is widely dis-tributed in other tropics and subtropics of the world, whereas SP is restricted to the temperate desert environments of the west coast (mean of 289 mm; Nakazato et al., 2010 ). The dramatic precipitation difference between the eastern and western Andes results from the fact that the moisture of the easterly trade winds from the Amazon basin precipitates as it ascends the Andes, causing high precipitation on the eastern slopes and hyper-arid conditions on the western slopes of the Andes (the rain shadow effect). Consequently, SP and other tomato species in the west-ern Andes largely rely on the water from the numerous rivers fl owing toward the Pacifi c Ocean and from the heavy mist during the winter months ( Cooper, 1971 ). Rigorous phylogenetic anal-yses (e.g., Rodriguez et al., 2009 ) have shown that SL and SP are nested within other wild tomato species, which are all re-stricted to the western Andes. Therefore, the preference of SL for the wet environments of the eastern Andes is most certainly a derived state, and SL has likely undergone a recent drastic habitat shift during the colonization of the eastern Andes fol-lowed by a rapid population expansion throughout South/Cen-tral America and beyond.
Studying the evolutionary modes and tempo of SL and SP has until recently been diffi cult for two reasons. First, their close relationship has yielded insuffi cient genetic polymor-phisms, resulting in low resolution of the species relationships and the population structure within each species. Second, it has been diffi cult to study environmental conditions at natural pop-ulations because detailed fi eld environmental data are generally not available at their locations. The recent maturation of cost-effi cient markers, however, has enabled detection of a large number of markers from many samples simultaneously, allow-ing detailed investigation of population structure. A rapid prog-ress has also been made in studying environmental conditions of natural populations. The steady improvements of remote sensing technologies and rapidly accumulating GIS-derived en-vironmental data provide information about geographic fea-tures and terrestrial environmental variation at both global and local scales. Recently, students of evolutionary biology and ecology have begun to integrate genetic data across the genome and detailed geographic data to form the fi elds of landscape genetics (reviewed by Manel et al. [2003] ) and spatial genetics (reviewed by Guillot et al. [2009] ). Such integrative approaches allow us not only to effi ciently, objectively, and accurately characterize natural populations and their habitats, but also to analyze biotic and abiotic factors simultaneously to understand how they interact to drive complex evolutionary processes such as adaptation and speciation.
Here, we analyze AFLP genotypes spanning the genome of natural populations of SL and SP to study their population struc-ture as well as patterns of evolutionary processes including de-mographic history, species divergence, and hybridization. The goals of this study were to investigate (1) the genetic structure within and between species and their dispersal modes, (2) the genetic consequences of the colonization of the eastern Andes by SL , and (3) the extent of interspecifi c hybridization. By inte-grating geographic and climatic information of these species from the previous study ( Nakazato et al., 2010 ), we investigated how population structure and introgression may be infl uenced
90 American Journal of Botany [Vol. 98
unlinked, the genome-wide excess of rare alleles in SL is likely due mostly to demographic history. Possible population expan-sion was also inferred from the allele frequency distribution of multilocus sequences in closely related wild tomato species, S. peruvianum and S. chilense ( Arunyawat et al., 2007 ), suggest-ing that population bottlenecks and expansions may be com-mon among tomato species.
The mean expected heterozygosity (Nei ’ s gene diversity, H S ) based on Hardy – Weinberg equilibrium among SL populations (0.212) was signifi cantly lower than that among SP populations (0.239, Welch ANOVA, F 1, 25.56 = 4.626, P = 0.041). Higher genetic diversity in SP than in SL was also observed in allozyme polymorphisms ( Rick and Fobes, 1975 ). This difference in ge-netic diversity between species may refl ect that (1) SL is pre-dominantly autogmous, whereas outcrossing rates vary among SP populations ( Rick, 1950, 1958 ; Rick et al., 1977, 1978 ) and (2) the genetic diversity within SL was dramatically reduced during the colonization of the eastern Andes. In both species, the expected heterozygosity was not randomly distributed across the geographic range. Values tended to be highest in northwest-ern Peru and Ecuador and declining in the north and south of these regions, particularly in SL ( Fig. 2 ), similar to the results based on allozyme polymorphisms in SP ( Rick et al., 1977 ).
Wright ’ s fi xation indices among populations of SL , SP , and both species combined were 0.291, 0.295, and 0.397, respec-tively, all deviating substantially from the random expectation ( P < 0.01). These high F ST values are consistent with previous F ST estimates among SP populations based on nuclear genes (0.467; Caicedo and Schaal, 2004 ) and among SL , SP , and S. cheesmaniae based on AFLPs (0.462; Nuez et al., 2004 ). Much lower values were obtained based on AFLPs (0.052 and 0.023 within SL and SP , respectively; Nakazato et al., 2008 ). How-ever, these unusually low F ST values were likely an artifact of spurious autoapomorphies resulting from automatic scoring of polymorphic AFLP bands. Although the phylogenetic relation-ships among populations are unlikely affected by such random scoring errors, population differentiation measures such as F ST are likely severely underestimated. In general, high F ST values in this and previous studies suggest that populations of both species are highly fragmented with little gene fl ow.
The partitions of genetic variance between species, among populations, and within populations based on AMOVA were 41.76, 38.77, and 19.47%, respectively, which signifi cantly de-viated from the random expectation ( P < 0.01). The high between-species and among-population genetic variances indicate that not only are populations within each species fragmented, but the two species are also genetically distinct, consistent with the conclusion from a previous allozyme study ( Rick et al., 1974 ).
Clustering populations using the STRUCTURE software identifi ed two clusters ( K = 2) with the highest likelihood, largely corresponding to the two species ( Fig. 3 ). Increasing K simply subdivided each species and progressively decreased the likelihood values (data not shown). Therefore, we consid-ered only the results from K = 2 runs for the subsequent analyses. The generally large difference in the Q value partitions between SL and SP clearly indicates that these species are largely geneti-cally distinct and that there is little gene fl ow between species. Also, there is generally little within-population genetic varia-tion, consistent with the large F ST values for both species (see above). However, some populations showed intermediate geno-types; that is some SL populations genetically look more like SP , and vice versa, (i.e., SP 1256, SL 1231, SL 1385, and SP 2852), suggesting allele sharing between species.
16 populations, respectively (T. Nakazato, R. Franklin, and E. Housworth, unpublished data). Hence, the genetic diversity in SL and SP appears to be relatively low among wild tomato spe-cies, likely due to the self-compatible reproductive system. The correlation between AFLP fragment size and marker frequency was nonsignifi cant ( r = − 0.031, P = 0.726), indicating no sig-nifi cant fragment size homoplasy, which can be problematic for marker types with numerous fragments such as AFLP ( Vekemans et al., 2002 ). Therefore, the majority of AFLP markers in this study likely represent unique fragments.
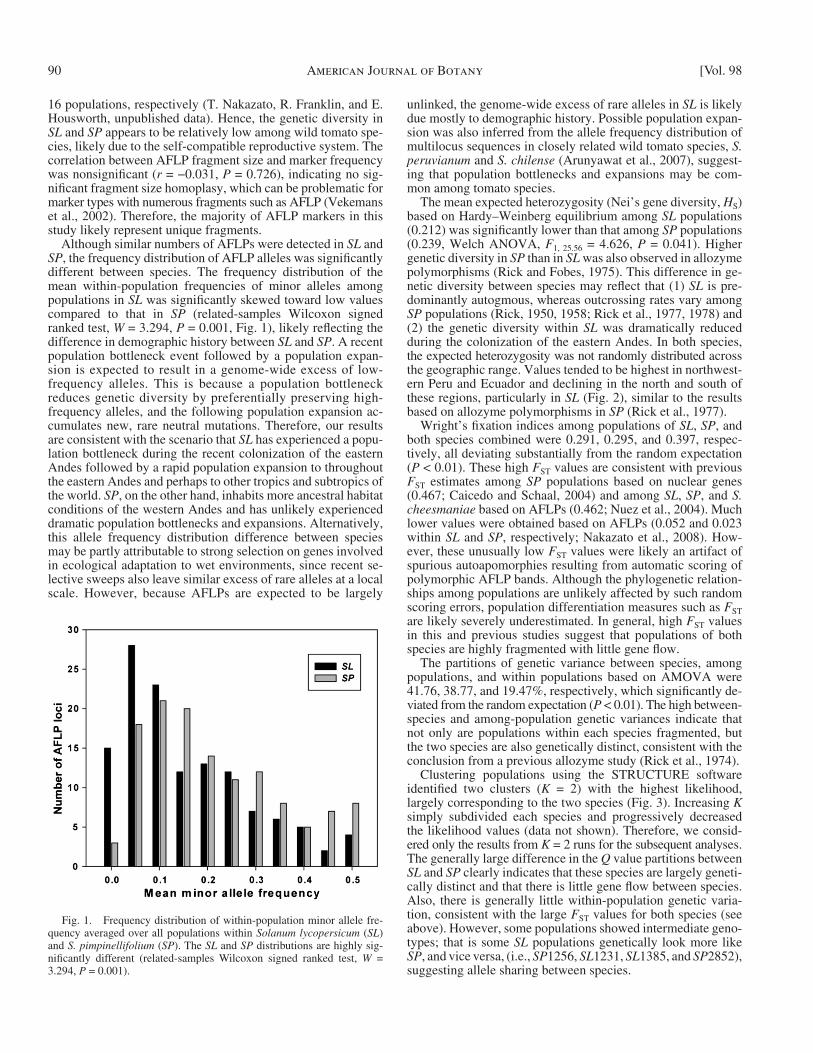
Although similar numbers of AFLPs were detected in SL and SP , the frequency distribution of AFLP alleles was signifi cantly different between species. The frequency distribution of the mean within-population frequencies of minor alleles among populations in SL was signifi cantly skewed toward low values compared to that in SP (related-samples Wilcoxon signed ranked test, W = 3.294, P = 0.001, Fig. 1 ), likely refl ecting the difference in demographic history between SL and SP . A recent population bottleneck event followed by a population expan-sion is expected to result in a genome-wide excess of low-frequency alleles. This is because a population bottleneck reduces genetic diversity by preferentially preserving high-frequency alleles, and the following population expansion ac-cumulates new, rare neutral mutations. Therefore, our results are consistent with the scenario that SL has experienced a popu-lation bottleneck during the recent colonization of the eastern Andes followed by a rapid population expansion to throughout the eastern Andes and perhaps to other tropics and subtropics of the world. SP , on the other hand, inhabits more ancestral habitat conditions of the western Andes and has unlikely experienced dramatic population bottlenecks and expansions. Alternatively, this allele frequency distribution difference between species may be partly attributable to strong selection on genes involved in ecological adaptation to wet environments, since recent se-lective sweeps also leave similar excess of rare alleles at a local scale. However, because AFLPs are expected to be largely
Fig. 1. Frequency distribution of within-population minor allele fre-quency averaged over all populations within Solanum lycopersicum ( SL ) and S. pimpinellifolium ( SP ). The SL and SP distributions are highly sig-nifi cantly different (related-samples Wilcoxon signed ranked test, W = 3.294, P = 0.001).

91January 2011] Nakazato and Housworth — Spatial genetics of wild tomatoes
isolation by distance (IBD), consistent with previous studies ( Caicedo and Schaal, 2004 ; Nakazato et al., 2008 ). Therefore, both species appear to show a classic stepping-stone mode of dispersal. The F ST -based Fitch – Margoliash phylogeny ( Fig. 4 ) clearly illustrates that SL and SP are distinct lineages. Although many branches within each species are only weakly supported due to the lack of suffi cient informative polymorphisms, the divergence of SL and SP is strongly supported. Despite the clear genetic distinctness between species, several populations are positioned between the SL and SP clades ( SP 1256, SL 1231, SL 1385, and SP 2852). The geographic locations of these popu-lations, however, suggest that they are likely interspecifi c hy-brids (see below). Strong IBD in SP is also apparent when the phylogeny is superimposed on the population localities ( Fig. 4 ), where there is a strong association between phylogenetic rela-tionships and geographic distances, particularly in central Peru. In SL , on the other hand, such association is much less pro-nounced, although some regional clustering of genetically closely related populations is apparent.
Geographic distribution patterns of within-population allele frequencies provide insights on how mutations arise and spread as well as how hybrid zones infl uence gene fl ow between species. Geographic distribution patterns of within-species AFLP loci in this study can be broadly categorized into four types on a visual basis, similar to those in Rick et al. (1977) : simple clines, single-peaked clines, rare alleles, and no gen-eral patterns.
Pattern 1. Simple clines — Forty-three markers (33.9%) showed a clear divergence in allele frequency between species, but showed intermediate frequencies in the hybrid zone in Ecua-dor. A previous study found a similarly large fraction of al-lozyme markers with simple-cline patterns ( Rick et al., 1977 ). This pattern can result from the fi xation of neutral alleles within a species by genetic drift followed by introgression in Ecuador and can be accomplished most easily if alleles arose in the bot-tleneck populations of SL during the colonization of the eastern Andes. Alternatively, such divergence of allele frequencies be-tween species can be achieved by selection induced by the envi-ronmental differences on the opposite sides of the Andes. The Ecuadorian populations show intermediate allele frequencies possibly because the climate is intermediate between those of the eastern and western Andes allowing SL and SP alleles to coexist (see Discussion). Given the large number of unlinked loci with this distribution pattern, it is tempting to speculate that the former hypothesis predominates, but it is possible that a large fraction of the genome is under environment-induced selection.
Pattern 2. Single-peaked clines — Another 43 markers (33.9%) showed regional peaks in allele frequency and rapid frequency decrease over short geographic distances. This pat-tern is most consistent with the fi xation of derived alleles in single populations by genetic drift, followed by the spread of these alleles to nearby populations, although we cannot rule out the possibility of local environment-induced selection. Of those 43 markers, 40 (93.0%) markers had peaks within SP rather than within SL , which may refl ect the fact that SL has experi-enced a recent bottleneck event and, therefore, has not had enough time to fi x neutral mutations within populations.
Pattern 3. Rare alleles — Twelve markers (9.4%) showed high allele frequencies in a single population or in few popu-lations that do not commonly occur in other populations.
Geographic distribution of AFLP genotypes — Sharing of alleles between species can result from either incomplete lin-eage sorting or introgression. Incomplete lineage sorting results from random segregation of ancestral alleles among descendent lineages. The fact that only some populations show genome-wide allele sharing strongly suggests that introgression rather than incomplete lineage sorting is largely responsible for the allele sharing between species. Furthermore, the geographic distribution of the proportions of species-specifi c alleles ( Q scores; Fig. 4 ) clearly indicates that recombinant genotypes are concentrated in central to northern Ecuador. This association between the occurrence of interspecifi c recombinant genotypes and geographic locations strongly suggests extensive hybrid-ization between species in this region. One notable exception to this pattern is seen in the southernmost SL population, SL 1385. Given the highly recombinant nature and extremely low within-population genetic diversity ( H S = 0.167), it is likely an early-generation hybrid population, resulting either from hybridization between an SP individual from the western Andes and a local SL individual or from long-distance dispersal of a hybrid individual.
The correlation between pairwise F ST ’ s and geographic dis-tances was highly signifi cant for SP ( r = 0.50, P = 0.000067) and marginally signifi cant for SL ( r = 0.33, P = 0.055), indicating
Fig. 2. Geographic distribution of expected heterozygosity (Nei ’ s gene diversity, H S ) in Solanum lycopersicum and S. pimpinellifolium as-suming Hardy – Weinberg equilibrium.

92 American Journal of Botany [Vol. 98
linked to genes under strong selection induced by local envi-ronments or reproductive incompatibility. Finer scale analy-ses of the hybrid zone using a large number of recombinant individuals as well as physically mapped markers (e.g., SNPs) may in the future sort out these alternatives.
DISCUSSION
Divergence of SL and SP — AFLP data among SL and SP populations in this study clearly showed that they are geneti-cally distinct lineages and that gene fl ow between species is minimal, except in central to northern Ecuador, where they oc-cur in sympatry. Divergence of the two species is also refl ected in the substantial differences in their morphology, including a greater seed mass and seedling size, larger leaves, and faster growth of SL compared to SP ( Nakazato et al., 2008 ). Because the two species are allopatric in most of their geographic ranges, geographic isolation imposed by the Andes mountain chain clearly contributes to the genetic divergence between these spe-cies. However, the recent analysis of 11 GIS-derived environ-mental variables of wild tomato species ( Nakazato et al., 2010 ) strongly suggests that the difference in climatic conditions, par-ticularly precipitation, has likely played a critical role in species divergence. At the same time, the study found no sig-nifi cant correlation between pairwise species range overlap and their genetic divergence (age – range correlation; Barraclough et al., 1998 ; Berlocher, 1998 ; Barraclough and Vogler, 2000 ; Berlocher and Feder, 2002 ; Fitzpatrick and Turelli, 2006 ) ex-pected for purely geographic modes (allopatric or sympatric) of speciation. Instead, tomato species appear to be largely allopat-ric regardless of the degree of interspecifi c genetic divergence. Therefore, the ecological, morphological, and genetic diver-gence of SL and SP is certainly attributable to the presence of
This pattern is most consistent with the scenario in which newly arising neutral mutations are fi xed within populations by genetic drift, but have not migrated to other populations. Fixation of neutral mutations within populations is likely fre-quent in wild tomatoes since population sizes are generally small ( < 100), and migration is limited as indicated by large F ST values.
Pattern 4. No clear patterns — Twenty-nine markers (22.8%) showed various allele frequencies among populations with no clear geographic patterns. This lack of clear geographic patterns in allele frequencies may be caused by old neutral polymorphisms segregating among populations by incomplete lineage sorting between species, or, alternatively, AFLP frag-ment homoplasy.
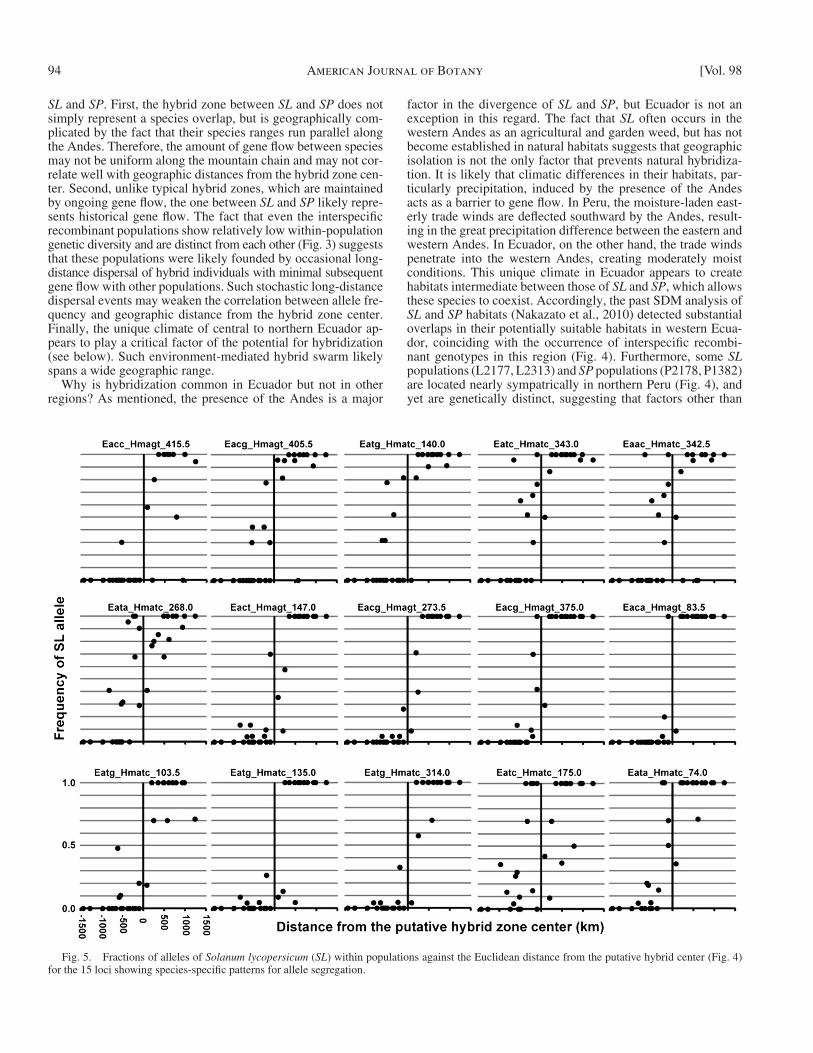
When loci show simple cline patterns (pattern 1), the change in species-specifi c allele frequencies across the hy-brid zone provides information about the cline width and possibly the strength of species-specifi c selection acting on and around these marker loci, although interpreting the pat-tern is complicated by the small number of interspecifi c re-combinant genotypes and complex geographic distribution of these species. We focused on the 15 loci (11.81% of 127 markers, a subset of pattern 1) that show species-specifi c al-lele segregation patterns ( > 90% mean allele fi xation among “ pure ” SL [ SL 1286, 2308, 2618, 2792] and SP [ SP 1478, 1578, 1597, 1629, 2178]). These loci showed allele fre-quency clines across the putative hybrid zone center in Ec-uador ( Fig. 5 ), indicating introgression of alleles between species. On a visual basis, the cline widths for most loci are ~500 km, quite extensive for bee-pollinated species. How-ever, some loci, namely Eaca_Hmagt_83.5, and Eatg_Hmatc_135.0 showed truncated patterns of allele frequency changes, suggesting narrow cline widths. These loci may be
Fig. 3. Q values of individuals within populations estimated using STRUCTURE with K = 2. Values were averaged over three independent runs.

93January 2011] Nakazato and Housworth — Spatial genetics of wild tomatoes
artifi cial crosses (reviewed by Rick and Butler [1956] ). Sola-num pimpinellifolium is often observed around agricultural ar-eas along cultivated tomatoes, and bees (particularly species of Augochlora and Exomalopsis ) are so common that cross polli-nation and introgression of alleles are inevitable ( Rick, 1950 ; Rick et al., 1978 ). Although natural hybridization among wild tomato species other than between SL and SP is extremely rare at most ( Taylor, 1986 ), signifi cant gene fl ow between closely related wild tomato species, S. peruvianum and S. chilense , has been detected from multilocus sequencing data ( Stadler et al., 2005 , 2008 ; Arunyawat et al., 2007 ).
The simple allele frequency clines in many loci in Ecuador ( Fig. 5 ) are indicative of introgression of alleles between SL and SP . However, these clines differ from those of typical hybrid zones in, for example, Bombina toad species in Croatia ( Szymura, 1993 ) and European house mice ( Teeter et al., 2008 ) in two ways: (1) the clines are not always smooth, and allele frequencies of nearby populations can be quite different, and (2) the cline widths are roughly an order of magnitude greater (~500 km as opposed to the typical 10 – 30 km). There are sev-eral possible reasons for the nontypical hybrid zone structure in
the Andes, but is unlikely driven purely by geographic modes of speciation. Instead, adaptation to different ecological niches created by the recent uplift of the Andes likely plays a major role in their divergence.
Hybridization between SL and SP — Our results clearly indi-cated that extensive allele sharing is occurring in central to northern Ecuador between otherwise distinct SL and SP . Such association between the occurrence of interspecifi c recombi-nant genotypes and geography is a strong indication of natural hybridization. Previously, a similar pattern of introgression be-tween these species was observed in the geographic distribution of allozymes ( Rick et al., 1974 ). More recently, unusually high sequence diversity among 50 nuclear genes was observed among Ecuadorian SL accessions ( Labate et al., 2009 ). Past fi eld observations are also consistent with extensive natural hy-bridization in this region. For example, Rick (1958) observed that morphological characters intermediate of SL and SP are particularly common in Ecuadorian populations. Natural hy-bridization between these two species is not surprising because very little, if any, reproductive barrier has been observed in
Fig. 4. Geographic distribution of mean within-population Q scores (pie charts based on Fig. 3 ) for Solanum lycopersicum ( SL ) and S. pimpinellifolium ( SP ) populations along with the Fitch – Margoliash phylogeny. The trees are connected by the arrows. The distribution is superimposed over the potentially suitable habitats of SL and SP predicted by species distribution modeling (SDM) at the threshold of equal sensitivity and specifi city (from Nakazato et al., 2010 ). The red dot on the map indicates the putative hybrid zone center used for calculation of the geographic distance between each population locality and the approximate hybrid canter.
94 American Journal of Botany [Vol. 98
factor in the divergence of SL and SP , but Ecuador is not an exception in this regard. The fact that SL often occurs in the western Andes as an agricultural and garden weed, but has not become established in natural habitats suggests that geographic isolation is not the only factor that prevents natural hybridiza-tion. It is likely that climatic differences in their habitats, par-ticularly precipitation, induced by the presence of the Andes acts as a barrier to gene fl ow. In Peru, the moisture-laden east-erly trade winds are defl ected southward by the Andes, result-ing in the great precipitation difference between the eastern and western Andes. In Ecuador, on the other hand, the trade winds penetrate into the western Andes, creating moderately moist conditions. This unique climate in Ecuador appears to create habitats intermediate between those of SL and SP , which allows these species to coexist. Accordingly, the past SDM analysis of SL and SP habitats ( Nakazato et al., 2010 ) detected substantial overlaps in their potentially suitable habitats in western Ecua-dor, coinciding with the occurrence of interspecifi c recombi-nant genotypes in this region ( Fig. 4 ). Furthermore, some SL populations (L2177, L2313) and SP populations (P2178, P1382) are located nearly sympatrically in northern Peru ( Fig. 4 ), and yet are genetically distinct, suggesting that factors other than
SL and SP . First, the hybrid zone between SL and SP does not simply represent a species overlap, but is geographically com-plicated by the fact that their species ranges run parallel along the Andes. Therefore, the amount of gene fl ow between species may not be uniform along the mountain chain and may not cor-relate well with geographic distances from the hybrid zone cen-ter. Second, unlike typical hybrid zones, which are maintained by ongoing gene fl ow, the one between SL and SP likely repre-sents historical gene fl ow. The fact that even the interspecifi c recombinant populations show relatively low within-population genetic diversity and are distinct from each other ( Fig. 3 ) suggests that these populations were likely founded by occasional long-distance dispersal of hybrid individuals with minimal subsequent gene fl ow with other populations. Such stochastic long-distance dispersal events may weaken the correlation between allele fre-quency and geographic distance from the hybrid zone center. Finally, the unique climate of central to northern Ecuador ap-pears to play a critical factor of the potential for hybridization (see below). Such environment-mediated hybrid swarm likely spans a wide geographic range.
Why is hybridization common in Ecuador but not in other regions? As mentioned, the presence of the Andes is a major
Fig. 5. Fractions of alleles of Solanum lycopersicum ( SL ) within populations against the Euclidean distance from the putative hybrid center ( Fig. 4 ) for the 15 loci showing species-specifi c patterns for allele segregation.

95January 2011] Nakazato and Housworth — Spatial genetics of wild tomatoes
generally dispersed over wide geographic areas, gene fl ow be-tween populations via pollen dispersal is likely a rare event.
Although populations of SL and SP appear to have equally limited gene fl ow, within-population genetic diversity was sig-nifi cantly higher in SL than in SP . This difference may be partly due to the mating system difference between species. Although both species are self-compatible, SL is largely autogamous, and stigmas are often submerged within the anther tube ( Rick, 1950, 1976 ), whereas exerted stigmas are common among SP popula-tions, resulting in various outcrossing rates ( Rick, 1950, 1958 ; Rick et al., 1977, 1978 ). Although higher selfi ng rates per se do not result in low within-population genetic diversity, founding of populations by a small number of highly homozygous indi-viduals via seed dispersal causes a severe genetic bottleneck, which is likely a common occurrence in wild tomato species.
Interestingly, within-population diversity decreased with in-creasing distances from the northwestern Peru – Ecuador region in both species. This may be partly due to the introgression of alleles between species upon interspecifi c hybridization in Ecuador. However, Rick et al. (1977) found that not only heterozygosity, but also allele diversity, stigma exertion, and cross pollination rates decreased as populations move further away from the northwestern Peru – Ecuador region, that is, pop-ulations increasingly have more selfi ng habits with latitude. Therefore, the heterozygosity gradient we observed is not caused by simple introgression of alleles between species but by other evolutionary factors. Such a heterozygosity gradient can be explained by the “ abundant center ” model of species distribution ( Hengeveld and Haeck, 1982 ; Brown, 1984 ; Lawton, 1993 ; Sagarin and Gaines, 2002 ), where population size, fre-quency, and density are proposed to generally decrease as pop-ulations move farther away from the geographical center of diversity. This isolation of peripheral populations likely leads to less-diverse pollinators ( Aizen and Feinsinger, 1994 ; Groom, 1998 ), reduced outcrossing due to lower density of conspecifi c pollen sources ( Byers, 1995 ; Agren, 1996 ), and therefore, pro-motes the evolution of self-fertilization ( Cruden and Lyon, 1989 ; Lloyd, 1992 ). Consistent with the abundant center model, Jenkins (1948) and Rick and Fobes (1975) have suggested that the Peru – Ecuador region served as a center of diversity of SL leading to domestication.
Limitations of the study and future directions — The present study demonstrates the power of integrating polymorphism data spanning the genome and geographic data to investigate how population structure of wild species and the mode of their evo-lution are infl uenced by factors such as geographic distances and climate. This approach is particularly powerful in wild to-mato species because of a number of natural accessions with geographic location information, whose seeds are readily ac-cessible from seed banks.
However, the approach used in this study has several limita-tions. First, even with the cost-effi cient AFLP genotyping, only a relatively small number of accessions (33 accessions, 378 in-dividuals) could be analyzed due to the cost, labor, and space limitations. These limitations prevented us from obtaining a more complete picture of between- and within-species genetic structure, particularly within the hybrid zone. Second, the dom-inant and anonymous nature of the AFLP markers prevented us from forming a detailed picture of polymorphism patterns in their genomes. Although AFLPs allowed us to screen a large number of genomic fragments, only a small number of markers
geographic distance are maintaining the species integrity. In-deed, SL populations, L2177 and L2313 receive higher precipi-tation (1165 and 965 mm, respectively) than SP populations, P2178 and P1382 (913 and 815 mm, respectively). Therefore, precipitation again appears to play an important role in ecologi-cal isolation between species despite their close geographic proximity, although more sympatric populations need to be sampled in the future to understand the nature of this barrier to gene fl ow.
Although our results suggest that SL and SP are largely ge-netically distinct and hybridize in Ecuador, they do not help us answer how the “ hybrid zone ” was formed. In particular, we do not know whether the hybrid zone represents a zone of second-ary contact between well-differentiated SL and SP or a gradual transition from the coastal type to the Amazonian type medi-ated by a climatic (particularly precipitation) gradient. Given the extensive introgression between species occurring over an unusually wide geographic area, the hybrid zone is likely quite old. Therefore, it is tempting to speculate that the hybrid zone represents a transitional area rather than a secondary contact zone. The sympatric region in the northern Peru, on the other hand, shows little evidence of introgression and may represent a “ tension zone ” as a result of a secondary contact between spe-cies. Again, further studies are needed to understand the nature of these hybrid zones.
Dispersal patterns of SL and SP — As in our previous study ( Nakazato et al., 2008 ), we found that IBD is much stronger in SP than in SL , indicating that SP exhibits a stronger stepping-stone dispersal pattern than SL . This difference in dispersal pattern is likely related to the differences in their habitats. Because SP is restricted to the western Andes, populations are largely bound to a series of river valleys along the otherwise hyper-arid tem-perate deserts ( Rick, 1978 ; Taylor, 1986 ). Therefore, the strong IBD in SP is consistent with the view that SP populations dis-perse by occasional long-distance dispersal events to nearby river valleys followed by long isolation from other populations. Solanum lycospersicum , on the other hand, inhabits more eco-logically continuous habitats along the edge of tropical rain for-ests. This may make successful long-distance seed dispersal more frequent and likely weakens the correlation between ge-netic and geographic distances among populations. It is not clear, however, what agents are responsible for their seed dis-persal because virtually nothing is known about the modes and tempo of seed dispersal in wild tomato species. However, the fact that seeds of many wild tomato species need to be chemi-cally treated to germinate (TGRC) suggests that ingestion by birds and animals is critical for tomato reproduction and likely plays a signifi cant role in seed dispersal. In addition, because SL is closely related to the domesticated tomato, we cannot rule out the possibility that some SL populations may have been traded by long-distance transportation for cultivation, which weakens IBD among populations.
Although successful seed dispersal may be more frequent in SL than in SP , gene fl ow among populations appears to be limited in both species as indicated by equally high F ST values. This is not surprising, given that tomato species in general are bee-pollinated ( Rick, 1950 ; Rick et al., 1978 ; Chetelat et al., 2009 ; Albrecht et al., 2010 ), and therefore, pollen dispersal is likely limited to short geographic distances. In the case of the carpenter bee, Xylo-copa fl avorufa (DeGeer), foraging range was found to be up to 6 km, but most activities occur within plant patches and rarely between ( Pasquet et al., 2008 ). Because tomato populations are

96 American Journal of Botany [Vol. 98
Cruden , R. W. , and D. L. Lyon . 1989 . Facultative xenogamy: Examination of a mixed mating system. In J. H. Bock and Y. B. Linhart [eds.], The evolutionary ecology of plants, 171 – 208. Westview Press, Boulder, Colorado, USA.
Duvernell , D. D. , and B. J. Turner . 1998 . Evolutionary genet-ics of Death Valley pupfi sh populations: Mitochondrial DNA se-quence variation and population structure. Molecular Ecology 7 : 279 – 288 .
Duvernell , D. D. , and B. J. Turner . 1999 . Variation and divergence of Death Valley pupfi sh populations at retrotransposon-defi ned loci. Molecular Biology and Evolution 16 : 363 – 371 .
Felsenstein , J. 1989 . Phylogeny inference package (version 3.2). Cladistics 5 : 164 – 166 .
Fitzpatrick , B. M. , and M. Turelli . 2006 . The geography of mam-malian speciation: Mixed signals from phylogenies and range maps. Evolution 60 : 601 – 615 .
Givnish , T. J. 2010 . Ecology of plant speciation. Taxon 59 : 1326 – 1366 . Groom , M. J. 1998 . Allee effects limit population viability of an annual
plant. American Naturalist 151 : 487 – 496 . G uillot, G. , R . L eblois , A . C oulon , and A . C . F rantz . 2009 . Statistical
methods in spatial genetics. Molecular Ecology 18 : 4734 – 4756 . Handley , L. J. L. , A. Manica , J. Goudet , and F. Balloux . 2007 .
Going the distance: Human population genetics in a clinal world. Trends in Genetics 23 : 432 – 439 .
Hawthorne , D. J. , and S. Via . 2001 . Genetic linkage of ecological specialization and reproductive isolation in pea aphids. Nature 412 : 904 – 907 .
Hengeveld , R. , and J. Haeck . 1982 . The distribution of abundance I: Measurements. Journal of Biogeography 9 : 303 – 316 .
Hickerson , M. J. , B. C. Carstens , J. Cavender-Bares , K. A. Crandall , C. H. Graham , J. B. Johnson , L. Rissler , et al. 2010 . Phylogeog-raphy ’ s past, present, and future: 10 years after. Molecular Phylogenetics and Evolution 54 : 291 – 301 .
Jenkins , J. A. 1948 . The origin of the cultivated tomato. Ecological Botany 2 : 379 – 392 .
Knowlton , N. , L. A. Weigt , L. A. Solorzano , D. K. Mills , and E. Bermingham . 1993 . Divergence in proteins, mitochondrial DNA, and reproductive compatibility across the Isthmus of Panama. Science 260 : 1629 – 1632 .
Labate , J. A. , L. D. Robertson , and A. M. Baldo . 2009 . Multilocus sequence data reveal extensive departures from equilibrium in domes-ticated tomato ( Solanum lycopersicum L.). Heredity 103 : 257 – 267 .
Lawton , J. H. 1993 . Range, population abundance and conservation. Trends in Ecology & Evolution 8 : 409 – 413 .
Lloyd , D. G. 1992 . Self fertilization and cross fertilization in plants II: The selection of self fertilization. International Journal of Plant Sciences 153 : 370 – 380 .
Macnair , M. R. 1983 . The genetic control of copper tolerance in the yel-low monkey fl ower, Mimulus guttatus. Heredity 50 : 283 – 293 .
Manel , S. , M. K. Schwartz , G. Luikart , and P. Taberlet . 2003 . Landscape genetics: Combining landscape ecology and population genetics. Trends in Ecology & Evolution 18 : 189 – 197 .
Nakazato , T. , M. Bogonovich , and L. C. Moyle . 2008 . Environmental factors predict adaptive phenotypic differentiation within and between two wild Andean tomatoes. Evolution 62 : 774 – 792 .
Nakazato , T. , D. Warren , and L. C. Moyle . 2010 . Ecological and geographic modes of species divergence in wild tomatoes. American Journal of Botany 97 : 680 – 693 .
Nesbitt , T. C. , and S. D. Tanksley . 2002 . Comparative sequencing in the genus Lycopersicon : Implications for the evolution of fruit size in the domestication of cultivated tomatoes. Genetics 162 : 365 – 379 .
Nuez , F. , J. Prohens , and J. M. Blanca . 2004 . Relationships, ori-gin, and diversity of Galapagos tomatoes: Implications for the con-servation of natural populations. American Journal of Botany 91 : 86 – 99 .
Pasquet , R. S. , A. Peltier , M. B. Hufford , E. Oudin , J. Saulnier , L. N. Paul , J. T. Knudsen , et al. 2008 . Long-distance pollen fl ow assess-ment through evaluation of pollinator foraging range suggests transgene
could be analyzed because of the close genetic relationship among accessions of the two species. With the available geno-typing technologies, it is not feasible to scan the genome to de-tect loci under selection. To better understand the number and locations of loci under selection during the divergence of these species, we are conducting a genome-wide survey using sec-ond-generation-based restriction-site-associated DNA (RAD) sequencing ( Baird et al., 2008 ).
LITERATURE CITED
Agren , J. 1996 . Population size, pollinator limitation, and seed set in the self-incompatible herb Lythrum salicaria. Ecology 77 : 1779 – 1790 .
Aizen , M. A. , and P. Feinsinger . 1994 . Habitat fragmentation, native insect pollinators, and feral honey bees in Argentine Chaco Serrano. Ecological Applications 4 : 378 – 392 .
Albrecht , E. , M. Escobar , and R. T. Chetelat . 2010 . Genetic diversity and population structure in the tomato-like nightshades Solanum lyco-persicoides and S. sitiens. Annals of Botany 105 : 535 – 554 .
Arunyawat , U. , W. Stephan , and T. Stadler . 2007 . Using multilo-cus sequence data to assess population structure, natural selection, and linkage disequilibrium in wild tomatoes. Molecular Biology and Evolution 24 : 2310 – 2322 .
Baird , N. A. , P. D. Etter , T. S. Atwood , M. C. Currey , A. L. Shiver , Z. A. Lewis , E. U. Selker , et al. 2008 . Rapid SNP discovery and genetic mapping using sequenced RAD markers. PLoS ONE 3 : e3376 .
Barraclough , T. G. , and A. P. Vogler . 2000 . Detecting the geograph-ical pattern of speciation from species-level phylogenies. American Naturalist 155 : 419 – 434 .
Barraclough , T. G. , A. P. Vogler , and P. H. Harvey . 1998 . Revealing the factors that promote speciation. Philosophical Transactions of the Royal Society of London, B, Biological Sciences 353 : 241 – 249 .
Berlocher , S. H. 1998 . Can sympatric speciation via host or habitat shift be proven from phylogenetic and biogeographic evidence? In D. J. Howard and S. H. Berlocher [eds.], Endless forms: Species and speciation, 99 – 113. Oxford University Press, New York, New York, USA.
Berlocher , S. H. , and J. L. Feder . 2002 . Sympatric speciation in phy-tophagous insects: Moving beyond controversy? Annual Review of Entomology 47 : 773 – 815 .
Bloom , A. J. , M. A. Zwieniecki , J. B. Passioura , L. B. Randall , N. M. Holbrook , and D. A. St. Clair . 2004 . Water relations under root chilling in a sensitive and tolerant tomato species. Plant, Cell & Environment 27 : 971 – 979 .
Bonnet , E. , and Y. van de Peer . 2002 . zt: A software tool for simple and partial Mantel tests. Journal of Statistical Software 7 : 1 – 12 .
Brown , J. H. 1984 . On the relationship between abundance and distribu-tion of species. American Naturalist 124 : 255 – 279 .
Byers , D. L. 1995 . Pollen quantity and quality as explanations for low seed set in small populations exemplifi ed by Eupatorium (Asteraceae). American Journal of Botany 82 : 1000 – 1006 .
Caicedo , A. L. , and B. A. Schaal . 2004 . Population structure and phy-logeography of Solanum pimpinellifolium inferred from a nuclear gene. Molecular Ecology 13 : 1871 – 1882 .
Campbell , C. J. 1975 . Ecuador. In R. W. Fairbridge [ed.], The encyclo-pedia of world regional geology, 261 – 270. Dowden Hutchinson & Ross, Stroudsburg, Pennsylvania, USA.
Chetelat , R. T. , and Y. Ji . 2007 . Cytogenetics and evolution. In K. Razdan and A. K. Mattoo [eds.], Genetic improvement of solanaceous crops, 77 – 112. Science Publishers, Enfi eld, New Hampshire, USA.
Chetelat , R. T. , R. A. Pertuz é , L. Fa ú ndez , E. B. Graham , and C. M. Jones . 2009 . Distribution, ecology and reproductive biology of wild tomatoes and related nightshades from the Atacama Desert region of northern Chile. Euphytica 167 : 77 – 93 .
Cooper , A. J. 1971 . The native habitat of the tomato. Annual Report, Glasshouse Crops Research Institute, West Sussex, UK, 123 – 129.
Coyne , J. A. , and H. A. Orr . 2004 . Speciation. Sinauer, Sunderland, Massachusetts, USA.

97January 2011] Nakazato and Housworth — Spatial genetics of wild tomatoes
Rick , C. M. , R. W. Zobel , and J. F. Fobes . 1974 . Four peroxidase loci in red-fruited tomato species: Genetics and geographic distribu-tion. Proceedings of the National Academy of Sciences, USA 71 : 835 – 839 .
Rodriguez , F. , F. N. Wu , C. An é , S. Tanksley , and D. M. Spooner . 2009 . Do potatoes and tomatoes have a single evolutionary his-tory, and what proportion of the genome supports this history? BMC Evolutionary Biology 9 : 191 .
Sagarin , R. D. , and S. D. Gaines . 2002 . The ‘ abundant centre ’ distribu-tion: To what extent is it a biogeographical rule? Ecology Letters 5 : 137 – 147 .
Smith , S. D. , and I. E. Peralta . 2002 . Ecogeographic surveys as tools for analyzing potential reproductive isolating mechanisms: An example using Solanum juglandifolium Dunal, S. ochranthum Dunal, S. lycoper-sicoides Dunal, and S. sitiens I. M. Johnston. Taxon 51 : 341 – 349 .
Stadler , T. , U. Arunyawat , and W. Stephan . 2008 . Population ge-netics of speciation in two closely related wild tomatoes ( Solanum Section Lycopersicon ). Genetics 178 : 339 – 350 .
Stadler , T. , K. Roselius , and W. Stephan . 2005 . Genealogical foot-prints of speciation processes in wild tomatoes: demography and evi-dence for historical gene fl ow. Evolution 59 : 1268 – 1279 .
Stibane , F. R. 1975 . Colombia. In R. W. Fairbridge [ed.], The encyclo-pedia of world regional geology, 245 – 250. Dowden Hutchinson & Ross, Stroudsburg, Pennsylvania, USA.
Szymura , J. M. 1993 . Analysis of hybrid zones with Bombina. In R. G. Harrison [ed.], Hybrid zones and the evolutionary process, 261 – 289. Oxford University, New York, New York, USA.
Taylor , I. B. 1986 . Biosystematics of the tomato. In J. G. Atherton and J. Rudich [eds.], The tomato crop: A scientifi c basis for improvement, 1 – 34. Chapman and Hall, London, UK.
Teeter , K. C. , B. A. Payseur , L. W. Harris , M. A. Bakewell , L. M. Thibodeau , J. E. O ’ Brien , J. G. Krenz , et al . 2008 . Genome-wide patterns of gene fl ow across a house mouse hybrid zone. Genome Research 18 : 67 – 76 .
Vallejos , C. E. 1979 . Genetic diversity of plants for response to low temperatures and its potential use in crop plants. In J. M. Lyons, D. Graham, and J. K. Raison [eds.], Low temperature stress in crop plants: The role of the membrane, 473. Academic Press, New York, New York, USA.
Vekemans , X. 2002 . AFLP-SURV, version 1 [computer program]. Universit é Libre de Bruxelles, Bruxelles, Belgium.
Vekemans , X. , T. Beauwens , M. Lemaire , and I. Roldan-Ruiz . 2002 . Data from amplifi ed fragment length polymorphism (AFLP) mark-ers show indication of size homoplasy and of a relationship between degree of homoplasy and fragment size. Molecular Ecology 11 : 139 – 151 .
Wilding , C. S. , R. K. Butlin , and J. Grahame . 2001 . Differential gene exchange between parapatric morphs of Littorina saxatilis detected using AFLP markers. Journal of Evolutionary Biology 14: 611 – 619.
Young , K. R. , C. U. Ulloa , J. L. Luteyn , and S. Knapp . 2002 . Plant evolution and endemism in Andean South America: An introduction. Botanical Review 68 : 4 – 21 .
escape distances. Proceedings of the National Academy of Sciences, USA 105 : 13456 – 13461 .
Patterson , B. D. , R. Paull , and R. M. Smillie . 1978 . Chilling resis-tance in Lycopersicon hirsutum Humb. and Bonpl.: A wild tomato with a wide altitudinal distribution. Australian Journal of Plant Physiology 5 : 609 – 617 .
Peakall , R. O. D. , and P. E. Smouse . 2006 . genalex 6: Genetic analy-sis in Excel: Population genetic software for teaching and research. Molecular Ecology Notes 6 : 288 – 295 .
Peralta , I. E. , and D. M. Spooner . 2001 . Granule-bound starch syn-thase (GBSSI) gene phylogeny of wild tomatoes ( Solanum L. sec-tion Lycopersicon [Mill.] Wettst. subsection Lycopersicon ). American Journal of Botany 88 : 1888 – 1902 .
Peralta , I. E. , D. M. Spooner , and S. Knapp . 2008 . Taxonomy of wild tomatoes and their relatives ( Solanum sect. Lycopersicoides , sect. Juglandifolia , sect. Lycopersicon ; Solanaceae). Systematic Botany Monographs 84 : 1 – 186 .
Platt , A. , M. Horton , Y. S. Huang , Y. Li , A. E. Anastasio , N. W. Mulyati , J. Agren , et al . 2010. The scale of population structure in Arabidopsis thaliana. PLoS Genetics 6 : e1000843 .
Pritchard , J. K. , M. Stephens , and P. Donnelly . 2000 . Inference of population structure using multilocus genotype data. Genetics 155 : 945 – 959 .
Rick , C. M. 1950 . Pollination relations of Lycopersicon esculentum in native and foreign regions. Evolution 4 : 110 – 122 .
Rick , C. M. 1958 . The role of natural hybridization in the derivation of cultivated tomatoes of western South America. Economic Botany 12 : 346 – 367 .
Rick , C. M. 1973 . Potential genetic resources in tomato species: Clues from observations in native habitats. In A. M. Srb [ed.], Genes, enzymes, and populations, 255 – 269. Plenum Press, New York, New York, USA.
Rick , C. M. 1976 . Natural variability in wild species of Lycopersicon and its bearing on tomato breeding. Agraria 30 : 249 – 259 .
Rick , C. M. 1978 . Tomato germplasm resources. In Proceedings of 1st International Symposium on Tropical Tomato, Asian Vegetable Research and Development Center, Shanhua, Taiwan, 214 – 224.
Rick , C. M. 1979 . Biosystematic studies in Lycopersicon and closely re-lated species of Solanum . In J. G. Hawkes, R. N. Lester, and A. D. Skelding [eds.], The biology and taxonomy of the Solanaceae, 667 – 679. Academic Press, London, UK.
Rick , C. M. , and L. Butler . 1956 . Cytogenetics of the tomato. Advances in Genetics 8 : 267 – 382 .
Rick , C. M. , and J. F. Fobes . 1975 . Allozyme variation in the cultivated tomato and closely related species. Bulletin of the Torrey Botanical Club 102 : 376 – 384 .
Rick , C. M. , J. F. Fobes , and M. Holle . 1977 . Genetic variation in Lycopersicon pimpinellifolium : Evidence of evolutionary change in mating systems. Plant Systematics and Evolution 127 : 139 – 170 .
Rick , C. M. , M. Holle , and R. W. Thorp . 1978 . Rates of cross pollina-tion in Lycopersicon pimpinellifolium : Impact of genetic variation in fl oral characters. Plant Systematics and Evolution 129 : 31 – 44 .

98 American Journal of Botany
Species Accession Country Province Location Longitude Latitude Altitude (m a.s.l.)
SL 1231 Ecuador Napo Tena − 77.817 − 0.983 540 SL 1286 Peru Junin San Martin de Pangoa − 74.467 − 11.167 738 SL 1385 Peru Cusco Quince Mil − 70.760 − 13.229 666 SL 1421 Ecuador Napo Santa Cecilia − 76.993 − 0.085 337 SL 1540 Colombia Cauca Cali to Popayan − 75.621 2.444 746 SL 2126 Ecuador Zamora-Chinchipe El Dorado − 78.697 − 3.931 825 SL 2136 Ecuador Santiago-Morona Bella Union − 78.224 − 3.006 373 SL 2177 Peru Cajamarca San Ignacio − 79.001 − 5.146 1283 SL 2308 Peru San Martin San Francisco − 76.333 − 6.733 325 SL 2313 Peru Amazonas Jumbilla − 77.783 − 5.908 2128 SL 2618 Peru Huanuco Santa Lucia − 75.950 − 9.217 748 SL 2789 Colombia Antioquia Canaveral − 75.300 6.950 1532 SL 2792 Colombia Antioquia Tamesis − 75.716 5.668 1595 SL 2843 Peru San Martin Moyobamba − 76.967 − 6.050 880 SP 412 Ecuador Los Rios Pichilingue − 79.483 − 1.100 68 SP 1245 Ecuador El Oro Santa Rosa − 79.967 − 3.458 8 SP 1246 Ecuador Loja La Toma − 79.360 − 3.990 1253 SP 1256 Ecuador Guayas Naranjal − 79.617 − 2.667 24 SP 1263 Ecuador Guayas Barranco Chico − 79.817 − 2.067 6 SP 1382 Peru Amazonas Chachapoyas-Balsas − 77.850 − 6.217 2234 SP 1416 Ecuador Pichincha Las Delicias − 79.333 − 0.267 329 SP 1469 Peru Lambayeque El Pilar − 79.790 − 5.860 128 SP 1478 Peru Piura Santo Tome − 80.018 − 5.211 112 SP 1578 Peru La Libertad Santa Marta − 79.551 − 7.336 9 SP 1579 Peru Lambayeque Colegio Punto Cuatro − 79.870 − 6.590 31 SP 1592 Peru La Libertad Moche − 79.040 − 8.147 0 SP 1597 Peru Ancash Rio Casma − 78.340 − 9.470 36 SP 1629 Peru Lima Mirafl ores to Costa Verde − 77.033 − 12.117 92 SP 1630 Peru Ica Fundo La Palma − 76.121 − 13.495 14 SP 1683 Peru Piura Miramar − 81.110 − 4.870 8 SP 2178 Peru Cajamarca Tororume − 78.722 − 5.647 536 SP 2646 Peru Piura Chalaco − 79.800 − 5.050 2085 SP 2852 Ecuador Manabi Cristo Rey de Charapoto − 80.483 − 0.833 175
Appendix 1. Geographic information of the study accessions obtained from the Tomato Genetics Resource Center (TGRC) at the University of California at Davis. SL = Solanum lycopersicum L.; SP = Solanum pimpinellifolium L.