seasonal variation of in halocynthia
TRANSCRIPT

Vol. 175: 201-213,1998 p-- -- -
MARINE ECOLOGY PROGRESS SERIES Mar Ecol Prog Ser Published December 17
Seasonal variation of in situ feeding rates b y the temperate ascidian Halocynthia papillosa
Marta Ribes*, Rafel Coma, Josep-Maria Gili
Institut de Ciencies del Mar (CSIC), Passeig Joan de Borbo sln, E-08039 Barcelona, Spain
ABSTRACT: The natural diet and prey capture rate of the temperate ascidian Halocynthia papillosa were studied in the field in a Western Mediterranean population. In sltu grazing rates on dissolved organic carbon, detrital particulate organic carbon, procaryotes, pico- and nanoeucaryotes, phyto- plankton and ciliates were examined through a year cycle. The natural diet of the species included detrital organic matter, heterotrophic bacteria, Prochlorococcus sp., Synechococcus sp. (cyanobactena type), protozoa and phytoplankton with a mean size range from 0.6 Z? 0.3 (heterotrophic bacteria) to 70 * 22 pm (pennate diatoms). Specific clearance rates varied seasonally and exhibited a pattern of increase with temperature increase in which temperature explained 55 % of the variance in clearance rate throughout the year. Annual variation in prey concentration did not affect specific clearance rate Ingestion rate of the species showed a marked seasonal pattern which was different for the 2 main food sources: detrital and live particles. Thus, while the highest ingestion of detrital particles was in spring, the highest values of live particle ingestion occurred during summer and fall. Overall, an H. papillosa specimen of mean size 0.25 g AFDW (ash-free dry weight) was estimated to ingest an annual mean of 1305 * 496 pg C g AFDW-' h-' and 84 + 16 pg N g AFDW" h-' Carbon from detrital origin accounted for 92 k 2 % of the total ingested carbon, while ingestion of live carbon accounted for 8 -r- 2 %. However. the seasonal variation of ingested nitrogen from live particles explained 91 % of the gonadal develop- ment variance for the year, suggesting that live particles are likely to be of more significance in the diet of the species than particles from detrital origm. Feeding rates are discussed in relation to seasonal changes In food availability and production of the species (i.e. growth and reproduction). The results suggest that factors other than food availability determine the seasonal dynamics of the species.
KEY WORDS Diet . Feeding behavior . Prey capture . Ascidians . Mediterranean
INTRODUCTION
Benthic suspension feeders account for an important fraction of the biomass and production in coastal marine ecosystems (Cloern 1982, Officer et al. 1982, Hily 1991). Recently, several studies have indicated that these organisms play an important role in plank- ton-benthos coupling (e.g. Jerrgensen 1990, Gili & Coma 1998). Ascidians, together with sponges and molluscs, are among the groups better represented in hard substrate communities in littoral zones such as tropical areas (Koike & Suzuki 1996), temperate seas (Ramos 1988, Turon 1990a, Petersen & Riisgdrd 1992), kelp beds (Newel1 et al. 1982), and Antarctic waters (Gerdes et al. 1992).
In general, growth and reproduction of benthic sus- pension feeders in tropical, temperate and polar seas are characterized by strong seasonality (e.g. Millar 1971, Clarke 1987, Harrison & Wallace 1990). In the case of ascidians in the Western Mediterranean, 2 characteristic patterns have been distinguished. First, several colonial ascidians have been found to concen- trate growth and reproduction around the winter sea- son (i.e. autumn, winter and spring; Turon 1988, Turon & Becerro 1992), with non-feeding periods exhibited by some species during the warm period (Turon 1988, 1992). Second, the few solitary ascidians which have been studied in contrast show maximum reproductive activity during, or at the end of, the summer period (Becerro & Turon 1992). This fact is surprising because, in seasonal environments, the highest new production (i.e. growth and reproduction) usually occurs during
O Inter-Research 1998 Resale of full article notpermitted

202 Mar Ecol Prog Ser 175: 201-213, 1998
periods of no limitation by external factors (i.e. temper- ature and food availab~lity), and the Western Mediter- ranean in summer is character~zed by oligotrophy and low seston concentration (Zabala & Ballesteros 1989) However, a lack of knowledge of diet and grazing rate in nature has become a barrier to the understanding of the factors that determine the dynamics of these species.
Ascidians, as sessile suspension feeders, are nutri- tionally dependent on the surrounding water which provides them with their resources. As a group, they are considered to be non-selective filter feeders able to capture particles from 0.5 to 100 pm (Fiala-Medioni 1987), with high retention efficiency for particles larger than 0.6 pm (Fiala-Medioni 1978a, Randlov & Riisggrd 1979, Stuart & Klumpp 1984). The filtration rates of as- cidians have been shown to be dependent on external factors such as temperature, oxygen tension, and food concentration, as well as on morphological factors such as body size (Fiala-Medioni 1978c, 1979a, b, Robbins 1983, Klumpp 1984, Petersen & Riisgard 1992). How- ever, most of these studies have been carried out under laboratory conditions and with simplified diets (Fiala- Medioni 1978c, Ra.ndl0v & RiisgArd 1979, Robbins 1983, Petersen & RiisgArd 1992, Petersen et al. 1995). These works have provided an understanding of the physical and biological factors that affect feeding activity of as- cidi.ans. However, the difficu1.t~ of estimating feeding in nature and, its seasonal variation from laboratory results (Jargensen 1975, Okamura 1990, Vogel 1994) points to the need for studies focused on the examination of nat- ural feeding rates.
Halocynthia papillosa (Linnaeus) is one of the most common solitary species on the rocky littoral of the North Western Mediterranean (Fiala-Medioni 1972, Turon 1990a). H. papillosa is an hermaphroditic spe- cies that reproduces once per year in late summer. Al- though it has continuous vitellogenesis throughout the year, gonad development is higher from March to September for ovaries and from July to September for testes (Becerro & Turon 1992). The main objectives of this study were to determine the role of DOC (dissolved organic carbon), detrital POC (particulate organic carbon) and live carbon from different taxa as food sources for the ascidian, and to contribute to the knowledge of factors that determine the temporal vari- ability in production of benthic suspension feeders living in seasonal environments. The natural diet and grazing rates of H. papillosa were exanlined by in s i tu incubations with the whole natural spectrum of food sources (DOC, detrital POC, procaryotes, pico- and nanoeucaryotes, phytoplankton and ciliates). Feeding on these potential food sources was assessed season- ally in order to cover the natural range of food abun- dance and composition.
MATERIAL AND METHODS
Feeding experiments were conduced in situ using continuous flow incubation chambers at the Medes Islands Marine Reserve (NW Mediterranean Sea, 42" 3' N, 3" 13'E) from October 1995 to November 1996. Incubations were carried out in hemispherical UV- transparent Plexiglas chambers approximately 3 1 in volume. The chambers (1 experimental, with ascidian [Fig. l a ] , and 1 control [Fig. lb]) were placed on the sea bottom at 15 m depth and water was recirculated with a pump at a speed of 1.2 cm S-' (this flow be- comes turbulent inside the chambers) (Fig. lc) . Plastic fasteners with hooks securely anchor the chambers (see Fig. Id). The plastic fasteners were also used to attach the chambers to the bottom by means of elastic bands between them and permanent nails located on the bot- tom. Close to the pump propeller, a submersible polaro- graphic oxygen and temperature probe (Clarke-type oxygen probe, WTW EOT-196) was connected (Fig. l e ) . Oxygen concentration and temperature during the in- c.ubations were monitored in both chambers. Halocyn- thia papillosa specimens were removed from their natural substratum, and attached to PVC posts (2 cm in diameter, 2.5 cm tall) with an inert cement 1 mo be- fore the experiments and kept in their natural environ- ment with conspecifics. All specimens were selected to have a sim.ilar size (0.25 i 0.08 g ash-free dry welght, AFDW; 0.33 i 0.1 l g dry weight, DW; 0.09 + 0.03 g DW without tunic) in order to avoid the effect of size on the feeding behavior (Robbins 1983, Klumpp 1984, Petersen & Riisgdrd 1992). At the beginning of each ex- periment, an H. papillosa specimen was placed on the
Fig. 1. General view of the experimental set-up for incubat~on expenments. (a) Experimental chamber with the ascidian specimen, (b) control chamber, (c) flow pumps with oxygen electrodes (e) , (d) plastic fasteners with hooks to securely anchor the chambers, I f ) PVC piece c0nnectin.y inlet and outlet apertures of the chambers. When PVC piece ( f ) is not connected it allows outside water to circulate throughout the
chambers

Ribes et al.: In situ feedin .g by a temperate ascidian 203
base of the experimental chamber and both experi- mental and control chambers were closed while allow- ing outside water to circulate throughout the chambers (Fig. If). An acclimatization period of 2 h was left to avoid the lag-phase phenomenon (Petersen & Riisgard 1992). After this acclimatization, 3 replicate water samples of 500 m1 were collected from both chambers (initial water samples) and the flow of both chambers was forced to recirculate within the system. Water samples were preserved for further analysis (see be- low). Two hours later, 3 replicate water samples were collected again from both chambers (final water sam- ples). Grazing was calculated from the decrease in prey concentration in the experimental chamber relative to the control chamber. The potential prey items included: procaryotes (heterotrophic bacteria, Synechococcus sp., and Prochlorococcus sp.),.. pico- and nanoeucary- otes, ciliates, phytoplankton (diatoms and dinoflagel- lates), DOC and detrital POC. Water volume used for the analysis of DOC, POC, procaryotes, and pico- and nanoeucaryotes was previously screened by a 100 pm net to avoid larger plankters. Five experiments were carried out each season, and therefore a total of 20 ex- periments were performed throughout the year cycle.
To quantify heterotrophic bacteria, Prochlorococcus sp., Synecl~ococcus sp., and pico- and nanoeucaryotes we used flow cytometry. Two m1 water samples from the incubation chambers were preserved for flow cytometry by standard protocols (Campbell et al. 1994), frozen in liquid nitrogen, and stored at -80°C or in dry ice. Samples were analyzed using a Coulter EPICS 753 flow cytometer (Coulter Electronics Corpo- ration, Hialeah, Florida) equipped with two 5 W argon lasers and a Micro-Sampler-Delivery-System. The flow cytometer was set up for UV (220 mW) and 488 nm (1 W) colinear analysis. Hoechst 33342 (DNA-specific fluorochrome) was used to stain DNA according to Monger & Landry (1993). Five parameters were col- lected in list mode and analyzed with custom-designed software (CYTOPC by Daniel Vaulot): red fluores- cence (from chlorophyll a), orange fluorescence (from phycoerythrin), blue fluorescence (from DNA stained with Hoechst 33342), forward- and right-angle light scatter signals (FALS and RALS). For statistical pur- poses sample size for analysis was chosen to provide more than 10 000 counts sample-', then 1 m1 of sample was analyzed for picoeucaryotes and nanoeucaryotes and 100 p1 for heterotrophic bacteria, Prochlorococcus sp., and Synechococcus sp.
In order to measure cell sizes (length and width) of heterotrophic bacteria, Synechococcus sp., and pico- and nanoeucaryotes, subsamples of 20 m1 were stained with DAPI and passed through a 0.2 pm filter. For heterotrophic bacteria size estimation, over 100 cells were measured using software for image analysis as
descnbed in Massana et al. (1997). For Synechococcus sp. and pico- and nanoeucaryote size estimation, cell length and width were measured with a micrometer. Picoeucaryotes included naked flagellates smaller than 2 pm and nanoeucaryotes included naked flagellates ldryer than 2 pm as wcl! as small dinoflagellates (unable to be distinguished with an inverted micro- scope) and coccolithophores (unable to be counted with acid Lugol's solution) (see below). It was not possible to measure Prochlorococcus sp. due to the difficulty of observing these cells with epifluorescence microscopy, so a mean size of 0.7 pm, as calculated for this species in the Mediterranean by Vaulot et al. (1990), was used.
To quantify phytoplankton and ciliate cell numbers, 350 m1 water samples were preserved with acid Lugol's solution ( l % final concentration). Subsamples of 100 m1 were settled in settling chambers, and the major groups of nano- and microphytoplankton were quantified under an inverted microscope. The micro- scope was connected to a color CCD video camera and a video recorder. Images of the organisms for measure- ment were recorded on tape and digitized with a framegrabber. Size of the organisms was determined by image analysis software (NIH [National Institute of Health]-image). For each subsample 20 individuals of the most common groups were measured. Volumes were est~mated from length and width measurements assuming ellipsoidal or cylindrical shapes (Edler 1979).
POC was measured by filtering 60 m1 water samples onto pre-combusted glass fiber filters (Whatman GF/F 1825 025). Filters were then frozen in liquid nitrogen and kept at -80°C until analysis. Prior to analysis, filters were dried at 60°C for 24 h and exposed to HC1 vapors for 48 h in order to destroy inorganic material. Then filters were dried again and analyzed with a C:H:N autoanalyser (Perkin-Elmer 240). POC mea- surements included both detrital and live carbon. Detrital POC was estimated as the difference between total POC (C:H:N analysis) and total live carbon (esti- mated from cell counts and cell measurements).
For DOC, 20 m1 water samples were filtered through pre-combusted glass fiber filters (Whatman GF/F 1825 025). The filtered water was stored in glass tubes at -20°C until analysis. Analysis was conducted by high- temperature catalytic oxidation with an autoanalyser (Shimadzu TOC-5000).
Depletion rates of the plankton were calculated as- suming exponential growth and clearance of prey (Frost 1972, Saiz 1993, Ribes 1998). The prey growth rate k (h-') is computed thus:
where CO and C, are the prey concentrations in the chamber at the initial time to and at the final time t , .

204 Mar Ecol Prog Ser
The clearance rate F (volume swept clear biomass-l time-') is computed as follows:
where V is the volun~e of the chamber, b is the ascidian biomass (g AFDW) and g is the grazing coefficient (h- ' ) , computed as:
where k, is the prey growth rate in the control cham- ber, and k, is the apparent growth in the chamber with the ascidian. Finally, the ingestion rate, I (prey in- gested biomass-' time-'), is:
where C is the prey concentration. The significance of predation on each kind of prey
was tested by comparing growth rates of prey in control and experimental chambers with a 2-tailed Wilcoxon test (Sokal & Rohlf 1981). Cell biovolume was calculated from length and wi.dth by approximation to the nearest geometric shape. Carbon content was then estimated from literature conversion factors as follows: heterotrophic bacteria, 0.22 pg C pm-3 (Fry 1988); Pro- chlorococcus, 0.133 pg C pm-3 (Simon & Azam 1989); Synechococcus, 0.357 pg C pm-3 (mean value of: Bjornsen 1986, Kana & Glibert 1987, Verity et al. 1992); pico- and nanoeucaryotes, pg C = 0.433 X (pm")0-863 (Verity et al. 1992); phytoplankton, pg C cell-' = 0.109 X (pm3)0.9" (Montagnes et al. 1994); ciliates, 0.19 pg C pm-3 (Putt & Stoecker 1989).
Feces were obtained from inside the chamber in sev- eral experiments. They were collected with a Pasteur pipette and frozen in liquid nitrogen. Ten pieces of feces from different days were dehydrated in graded ethanol. Afterwards, they were dried by the critical point method (using CO2 as transition fluid), mounted on aluminum stubs and coated with gold in a sputter coater. Observation were done with a Hitachi S-570 scanning electron microscope (SEM). Halocynthia papillosa dry weight was determined for all incubated specimens by drying at 90°C for 24 h and ash-free dry weight was determined by combllstion at 450°C fc: 5 h. The relationship between total dry weight and dry weight without test was established by dissecting 10 specimen.s and drying separately the test and the man- tle at 90°C for 24 h. In average, dry weight without test was a 27 r 6% of tota.1 dry weight.
Multiple regression, analysis was used in order to establish the percentage of the variance of the esti- mated cl.earance rates that could be explained by the independent factors which were monitored during each experiment, which were water temperature ("C, recorded using a WTW oxygen electrode model EOT 196) and initial food concentration (cells ml-' for the
live particles and pg C 1-' for detrital particles). A back- ward stepwise procedure was used to exclude vari- ables that were not relevant (Sokal & Rohlf 1981). Vari- ables were square root transformed when normality (Kolmogorov-Smirnov test) and/or the heteroscedas- ticity (Levene's test) requirements were not fulfilled.
In order to determine whether or not the species exhibits seasonality in growth, Halocynthia papillosa growth rate was estimated from changes in area of the individuals over time based on photographic monitor- ing. Large H. papillosa specimens are highly sensitive to small perturbations, such as the approach of a cam- era, which produce a reaction of contraction. Thus, it is difficult to photograph specimens in a state of full expansion. Small specimens of the species were observed to be much less sensitive to the same kind of perturbations. Therefore, 11 H. papillosa individuals smaller than 1 cm were mapped in the studied area. These individuals weie photographed every 3 to 4 mo: March 1997, June 1997, October 1997 and February 1998. Size (mm2) of the individuals viewed from the side was determined using image analysis software (NIH-image). Growth rate (Gr, d-') was calculated by Eq. (5) as the change in area of every individual during each time interval:
where A. and AI are the ascidian areas at the initial time to and at the final time t,.
RESULTS
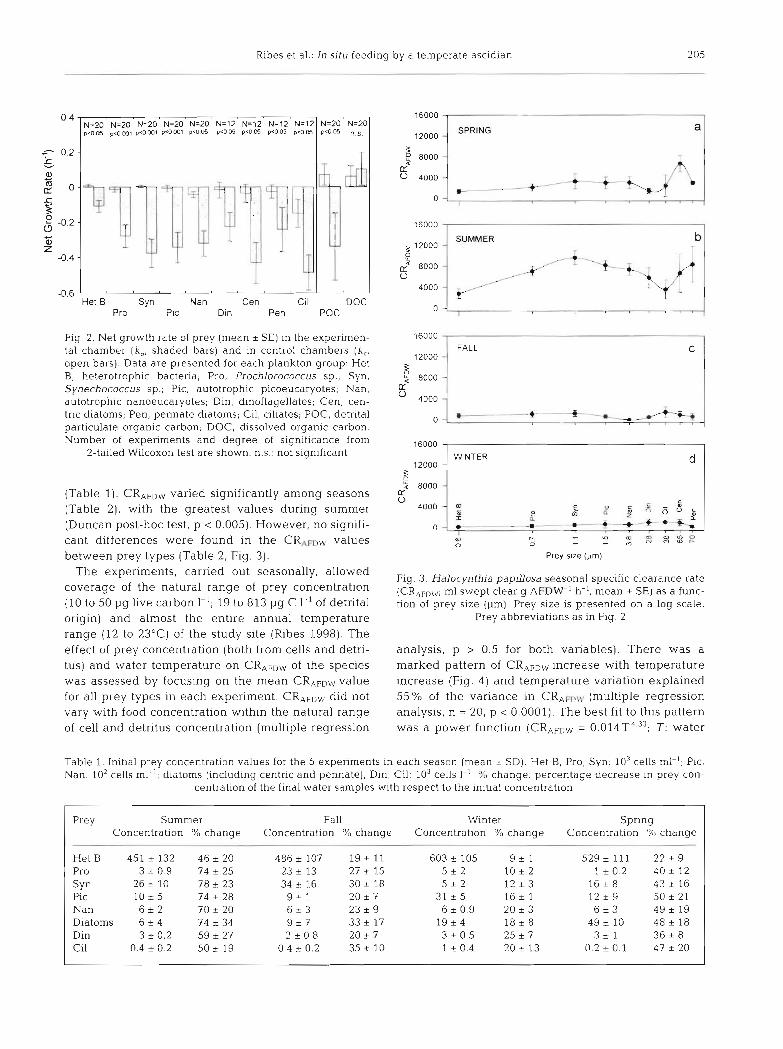
All sources of POC <l00 pm, both live (i.e. recogniz- able cells) and detrital, that were potentially available as food as well as DOC were considered in this study. The taxa distinguished within the live carbon cate- gory were: heterotrophic bacteria, Prochlorococcus sp., Synechococcus sp., pico- and nanoeuca.ryotes, phytoplankton (diatoms and dinoflagellates) and cili- ates. Overall, the prey growth rates calculated in the control and the experimentai chambers showed signif- icantly that Halocynthia papillosa fed on all live carbon groups as well as detrital POC (Fig. 2); however, no significant decrease in DOC was detected..
Ash-free dry weight specific clearance rate values (CRAFDLV, m1 swept clear g AFDW-l h-', hereafter spe- cific clearance rate) (i.e. the volume of water filtered by the ascidian to retain the observed decrease in number of cells, assuming a 100 % retention efficiency) for each season and type of prey are shown in Fig. 3. Mean per- centage decrease in prey concentration during the experiments was 65.6 + 12.3 in summer, 25.9 + 6.3 in fall, 16.3 i 5.6 m winter, and 41.9 + 9.4 in spring
Ribes et al.: In situ feeding by a temperate ascidian 205
4000 -0.6
Het B S Y ~ Nan Cen Cil DOC Pro Plc Din Pen POC 0
Fig. 2. Net growth rate of prey (mean i SE) in the experimen- tal chamber (k,, shaded bars) and in control chambers (k,, open bars). Data are presented for each plankton group: Het B, heterotrophic bacteria; Pro, Prochlorococcus sp.; Syn. Synechococcus sp.; Pic, autotrophic picoeucaryotes, Nan, autotrophic nanoeucaryotes, Din, dinoflagellates; Cen, cen- tric diatoms, Pen, pennate diatoms; Cil, ciliates; POC, detrital particulate organic carbon; DOC, dissolved organic carbon. Number of experiments and degree of significance from
2-tailed Wilcoxon test are shown. n.s.: not significant
(Table 1). CRAFDW varied significantly among seasons (Table 2), with the greatest values during summer (Duncan post-hoc test, p < 0.005). However, no signifi- cant differences were found in the CRtFDW values between prey types (Table 2, Fig. 3).
The experiments, carried out seasonally, allowed coverage of the natural range of prey concentration (10 to 50 pg live carbon 1-'; 19 to 813 pg C 1-' of detrital origin) and almost the entire annual temperature range (12 to 23OC) of the study site (Ribes 1998). The effect of prey concentration (both from cells and detn- tus) and water temperature on CRAFDGV of the species was assessed by focusing on the mean CRAFDw value for all prey types in each experiment. CRAFD\," did not vary with food concentration within the natural range of cell and detritus concentration (multiple regression
SPRING
WINTER 12000
B 3 8000 I
Prey slze (pm)
Fig. 3. Halocynthia paplllosa seasonal specific clearance rate (CRAFDW; m1 swept clear g AFDW-' h-', mean * SE) as a func- tion of prey size (pm). Prey size is presented on a log scale.
Prey abbreviations as in Fig. 2
analysis, p > 0.5 for both variables). There was a marked pattern of CRAFDw increase with temperature increase (Fig. 4) and temperature variation explained 55 % of the variance in CRAFDIP (multiple regression analysis, n = 20, p < 0.0001). The best fit to this pattern was a power function (CRAFDLV = 0 . 0 1 4 T ~ . ~ ~ ; T: water
Table 1. Initial prey concentration values for the 5 experiments in each season (mean i SD). Het B, Pro, Syn: 103 cells ml-l; Pic, Nan: 10' cells ml-l; diatoms (including centric and pennate), Din, Cil: 103 cells 1-' :':, change: percentage decrease in prey con-
centration, of the final water samples with respect to the initial concentration
Prey Summer Concentration % change
HetB 451 * 132 4 6 i 2 0 Pro 3 * 0.9 74 2 25 S Y ~ 2 6 + 1 0 7 8 2 2 3 Pic 10 i 5 74 * 28 Nan 6 i 2 70 * 20 Diatoms 6 + 4 74 r 34 Din 3 + 0.2 59 k 27 Cil 0.4 * 0.2 50 + 19
Fall Winter Concentration % change Concentration % change
486 2 107 19 t 11 603 + 105 9 2 1 23 13 27 t 15 5 2 2 1 0 2 2 34 16 30 * 18 5 2 2 1 2 2 3 9 2 1 20 * 7 31 2 5 16 * 1 6 2 3 23 + 9 6 2 0.9 20 * 3 9 2 7 33 t 17 1 9 2 4 18 + 8 2 0.8 2 0 + 7 3 + 0.5 25 * 7
0.4 .- 0.2 35 t 10 1 + 0.4 20 13
Spring Concentration % change
5292111 2 2 2 9 1 k 0 . 2 4 0 t 12
1 6 2 8 43 t 16 12 i 9 50 i 2 1 6 2 3 49 + 19
4 9 2 1 0 4 8 i 1 8 3 2 1 36 2 8
0 2 2 0 . 1 4 7 2 2 0
206 Mar Ecol Prog Ser 175. 201-213, 1998
Temperature
6000 CR,, l -
Dec Jan Feb Mar Apr May Jun Jul Aug Sep Od Nov
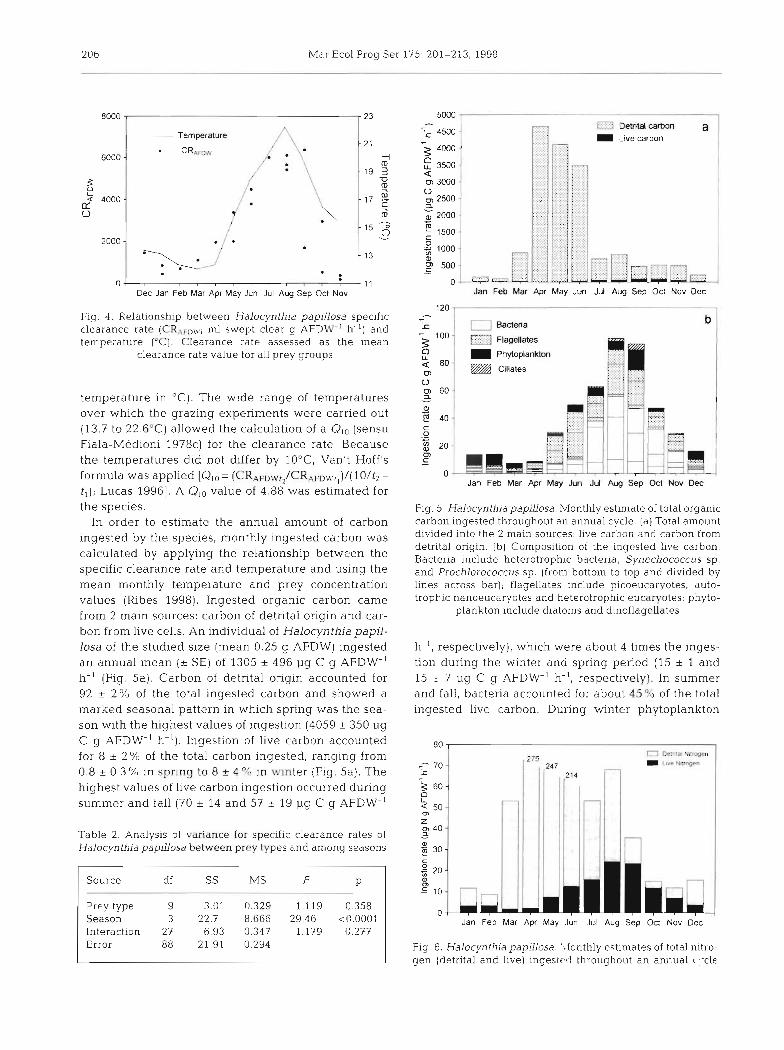
Fig. 4. Relationship between Halocynthia papillosa specific clearance rate (CRAFD\q; ml swept clear g AFDW-' h-') and temperature ('C). Clearance rate assessed as the mean
clearance rate value for a1.l prey groups
temperature in "C). The wide range of temperatures over which the grazing experiments were carried out (13.7 to 22.6'C) allowed the calculation of a Qlo (sensu Fiala-Medioni 1978c) for the clearance rate. Because the temperatures did not differ by 10°C, Van't Hoff's formula was applied [Qlo = (CRAFDWt,/CRAFUWr,)/[10/f2 -
t , ); Lucas 19961. A Q,o value of 4.88 was estima.ted for the species.
In order to estimate the annual amount of carbon ingested by the species, monthly ingested carbon was calculated by applying the relationship between the specific clearance rate and temperature and using the mean monthly temperature and prey concentration values (Ribes 1998). Ingested organic carbon came from 2 main sources: carbon of detrital origin and car- bon from live cells. An individual of Halocynthia papil- losa of the studied size (mean 0.25 g AFDW) ingested a n annual mean (* SE) of 1305 + 496 pg C g AFDW-' h-' (Fig. 5a). Carbon of detrital origin accounted for 92 + 2 % of the total ingested carbon and showed a marked seasonal pattern in which spring was the sea- son with the highest values of ingestion (4059 + 350 pg C g AFDW-l h-'). Ingestion of live carbon accounted for 8 + 2% of the total carbon ingested, ranging from 0.8 + 0..3Oh in spring to 8 +. 4l4 in winter (Fig. 5a). The highest values of live carbon ingestion occurred during summer and fall (70 + 14 and 57 + 19 pg C g AFDW-'
Table 2. Analysis of variance for specific clearance rates of Halocynthia papiUosa between prey types and among seasons
Source d f SS MS F P
Prey type 9 3.01 0.329 1.119 0.358 Season 3 22.7 8.666 29.46 <0.0001 Interaction 27 6.93 0.347 1.179 0.277 Error 88 21.91 0.294
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
120,
m Phytoplanklon
Ciliates
" , . . . . . . , . Jan Feb Mar Apr May .& ~i A& S& 0d NW Dec
Fig. 5. Halocynthia papiuosa. Monthly estimate of total organic carbon ingested throughout an annual cycle. (a) Total amount divided into th.e 2 main sources: live carbon and carbon from detrital origin. (b) Composition of the inyccsted live carbon. Bacteria include heterotrophic bacteria, Synechococcus sp. and Prochlorococcus sp. (from bottom to top and divided by lines across bar); flagellates include picoeucaryotes, auto- trophic nanoeucaryotes and heterotrophic eucaryotes; phyto-
plankton include diatoms and dinoflagellates
h-', respectively), which were about 4 times the inges- tion during the winter and spring period (15 * 1 and 15 + 7 pg C g AFDW-' h-', respectively). In summer and fall, bacteria accounted for about 45 % of the total ingested live carbon. During winter phytoplankton
Jan Feb Mar Apr May Jun Jul Aug Sep Od Nov Dec
Fig. 6. Halocynthia papillosa. ::lonthly estimates of total nitro- gen (detrital and Live) ingesird throughout an annual I :cle
Ribes et al.. In situ feeding b y a temperate ascidian
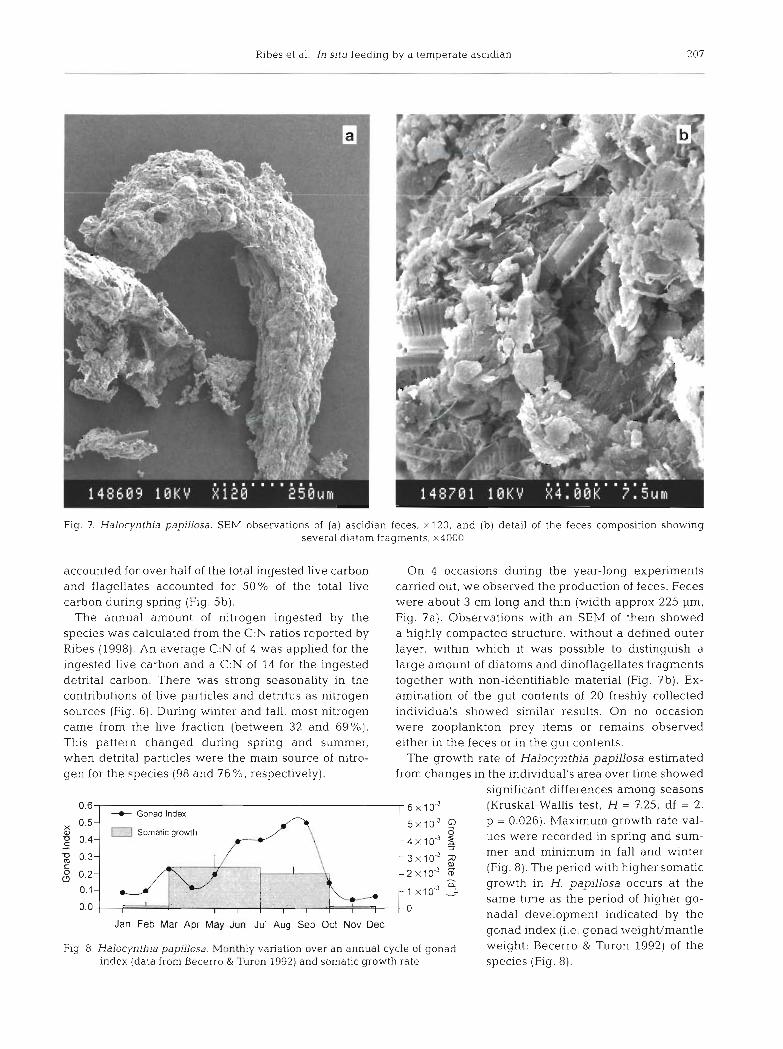
Fig. 7. Halocynthia papillosa. S E M observations of (a) ascidian feces, x120, and (b) detail of the feces composition showlng several diatom fragments, x4000
accounted for over half of the total ingested live carbon and flagellates accounted for 50% of the total live carbon d.uring spring (Fig. 5b).
The annual amount of nitrogen ingested by the species was calculated from the C:N ratios reported by Ribes (1998). An average C:N of 4 was applied for the ingested live carbon and a C:N of 14 for the ingested detrital carbon. There was strong seasonality in the contributions of live particles and detritus as nitrogen sources (Fig. 6). During winter and fall, most nitrogen came from the live fraction (between 32 and 69%). This pattern changed during spring and summer, when detrital particles were the main source of nitro- gen for the species (98 and 76%, respectively).
On 4 occasions during the year-long experiments carried out, we observed the production of feces. Feces were about 3 cm long and thin (width approx 225 pm, Fig. ?a). Observations with an SEM of them showed a highly compacted structure, without a defined outer layer, within which it was possible to distinguish a large amount of diatoms and dinoflagellates fragments together with non-identifiable material (Fig. 7b). Ex- amination of the gut contents of 20 freshly collected individuals showed similar results. On no occasion were zooplankton prey items or remains observed either in the feces or in the gut contents.
The growth rate of Halocynthia papillosa estimated from changes in the individual's area over time showed
significant differences among seasons 0.6 6 X 1 o - ~ (Kruskal-Wallis test, H = 7.25, df = 2, - Gonad Index , 0.5- - 10~3 F p = 0.026). Maximum growth rate val-
G) 0 Somatic growth o F 0.4- - X 10-3 ues were recorded in spring and sum- - - 3x10.3
mer and minimum in fall and winter (Fig. 8). The period with higher somatic -
4 growth in H. papillosa occurs at the same time as the period of higher go- nadal development indicated by the
Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec gonad index (i.e. gonad weight/mantle
Fig. 8. Halocynthia papillosa. Monthly variation over an annual cycle of gonad weight; Becerro & Turon 1992) of the index (data from Becerro & Turon 1992) and somatic growth rate species (Fig. 8).

208 Mar Ecol Prog Ser
DISCUSSION
The natural diet of Halocynthia papillosa was highly heterogeneous, including live and detrital organic car- bon. In size terms, this heterogeneous diet included the capture of a broad size range of food sources, from 0.6 (heterotrophic bacteria) to 70 pm (pennate diatoms). The smallest particle size captured by Halo- cynthia papillosa is in accordance with the size of the branchial filters (0.36 to 1.16 pm in diameter, Turon 1990b). In general, ascidians have been considered as non-selective suspension feeders for particles between 0.5 and 100 pm (Fiala-Medioni 1987). These potential food sources are retained by the mucous net secreted by the endostyle in the pharyngeal cavity and after- wards transported as a food cord to the intestine (Fiala- Medioni 1978b, Flood & Fiala-Medioni 1981). It has been pointed out that large particles (>l00 pm) usually do not reach the brachial cavity because of retention by the oral tentacles (Millar 1971, Klumpp 1984). How- ever, some studies which examined the gut contents of ascidians have shown that some species consume a variety of invertebrate larvae and eggs (Young 1988, Bingham & Walters 1989). Although this work was mainly focused on the study of prey items smaller than 100 pm, examination of the feces of the species showed a large amount of phytoplankton remains as well as non-identifiable material, but no zooplankton remains. Furthermore, no invertebrate larvae or egg remains were observed in the examination of the gut contents of the individuals examined in this study. These obser- vations do not disprove that H. papillosa could be feed- ing on zooplankton to some extent.
The marked seasonal variation in the CRAFDw of the species prompted the question of which environmental cues might influence its variability. As mentioned in the 'Introduction', temperature, food concentration, body size and oxygen tension have been suggested as the main factors that affect filtration rate (Fiala- Medioni 1978a, b, c, Robbins 1983, Klumpp 1984, Pe- tersen & Riisgdrd 1992). We did not examine the effect of oxygen tension on filtration rate of Halocynthia papillosa because a.11 experiments were performed in the field, where oxygen concentration was always fully saturated or slightly supersaturated. Oxygen concen- tration during the incubation experiments was never allowed to decrease by more than 10% of the initial oxygen concentration, a fall in magnitude which appears not to affect the behavior of the ascidian (Fiala-Medioni 1979a, Crisp 1984). Because specimens of similar size (0.25 + 0.08 g AFDW) were used In the experiments, the effect of body size on filtration rate was not examined. The experiments covered the natural range of prey concentration (10 to 50 pg llve carbon 1-', 19 to 813 pg C 1-' of detrital origin, Rlbes
1998), and within this range H. papillosa CRAFDw was not significantly affected by food concentration. These results agree with previous studies carried out with graphite particles (Jsrgensen 1955) and with food that resembles the composition of natural seston (Klumpp 1984). However, in laboratory studies. some authors have reported that food concentration affects filtration rate in ascidians, with a switching off of filtering at very low food concentrations and a satiation point at very high food concentrations (Fiala-Medioni 1979b, Robbins 1983, Klurnpp 1984, Petersen & Riisgdrd 1992). Our results indicate that, in nature, there is a wide range of food concentrations over which filtration rate remains constant, although H. papillosa may exhibit different feeding behaviors at extreme food concentra- tions. Temperature appears to be the factor that best explains the CRAFow seasonality. This result agrees with several previous laboratory studies (Holmes 1973, Fiala-Medioni 1978c, Robbins 1983, Petersen & Riis- gdrd 1992). The Qlo estimated for H. papillosa repre- sents a 5-fold increase in clearance rate over the tem- perature range of 13.7 to 22.6"C (Qlo = 4.88). This value is within the spread of values described for ascidian species studied to date, which ranges from 0.36 to 8.34 (Holmes 1973, Fiala-Medioni 1978c, Robbins 1983, Petersen & Riisgzrd 1992). Our data suggest that the optimal temperature value for filtration activity of H. papillosa corresponds with the maximum temperature of the water in the annual cycle at the study site (23°C). However, temperatures associated with maximum filtration activity may differ greatly between ascidian species (Fiala-Medioni 1 9 7 8 ~ ) .
The best fit of the relationship between clearance rate and temperature was a power function. In a previ- ous study Petersen & Riisgdrd (1992) found a linear relationship between clearance rate and temperature. This discrepancy could be related to several facts. Petersen & RiisgArd (1992) examined the effect of tem- perature on clearance rate under laboratory conditions and with a rather fast temperature change (i.e. hours or a few days). Our experiments represent a different approach to the effect of temperature on clearance rate because they were carried out over a year cycle with specimens that were always kept in their environment. In sublittoral temperate ecosystems, such as the one studied, temperature changes do not occur within a few days but over a longer time period, such as through the seasons. At this time scale, the organisms are sub- jected to physiological changes related to their cycle of production (i.e. growth and reproduction). In Halocyn- thia papillosa, the period of highest effect of tempera- ture on clearance rate coincides with that of the high- est gonadal development, which occurs during the warmest period of th.e year (Becerro & Turon 1992). Thls coincidence is because temperature is one of the

Ribes et al.: In situ feed ling by a temperate ascidian 209
main factors regulating reproduction in this species (Becerro & Turon 1992), as has been, in general, claimed for marine invertebrates (Orton 1920). We also observed that annual variation in growth rate suggests periods of highest growth correlate with those of highest gonadal development (Fig. 5 ) . Therefore, our results are difficult to compare with those of Petersen & Riisgard (1992) because they are subject to very differ- ent experimental conditions. Although a linear func- tion might be expected from the relationship between temperature and clearance rate when all other para- meters (such as ascidian size, reproduction period, etc.) remain constant, our data probably shows the added effect of both temperature and new production on clearance rate.
Although the positive effect of temperature on fil- tration rate appears to be a widespread characteristic of ascidian species, the mechanism that produces it is unclear. Some authors conducting laboratory experi- ments with other suspension feeders have interpreted low filtration rates at low temperatures as an energy- saving adjustment. This serves to reduce the high costs of filter feeding during winter in temperate seas, when food concentrations are low (Newel1 & Bayne 1980). This appears not to be the case in Halocynthia papillosa, because, although low filtration rates have been observed at low temperatures, the concentration of phytoplankton and detritus were high during the winter period (Ribes 1998). It has also been pointed out that the ascidian ciliary pharyngeal pump oper- ates in the absence of physiological mechanisms to regulate water pumping, as is the case with suspen- sion-feeding bivalves ( J~rgensen et al. 1990). In this sense, Petersen & RiisgArd (1992) suggested that the increase of clearance rate in ascidians with tempera- ture increase could correspond to a decrease in water viscosity.
Halocynthia paplllosa ingestion rate also varied sea- sonally. The positive correlation between temperature and clearance rates together with the variation in food concentration throughout the year (Ribes 1998) ex- plains the marked seasonality observed in the inges- tion rate of the species. Maximum ingestion rates ob- served in spring were associated with the relatively high clearance rate and with the high abundance of seaweed detrital POC in the water column during this period (Ballesteros 1991, Ribes 1998). During the sum- mer period, clearance rate increased but available POC decreased due to sedimentation (Ribes 1998). Thus, total ingestion rate decreased but the quality of the ingested material increased because sedimenta- tion is more important for detrital POC than for live carbon. The low ingestion rate values during the winter months were mainly due to the decrease in clearance rate.
Table 3 shows CRAFDW values for different species and habitats. The wide range of CRAFDw reported for Halocynthia papillosa in this study was mainly due to variation in CRAFDW over the year. CRAFDw depends on variables such as body size (Klumpp 1984), tempera- ture (Fia-la-Medioni 1978c) and the methods used to obtain the CRAFDw values (Fiala-Medioni 1978a), mak- ing comparison between species difficult. However, in general, Mediterranean species appear to exhibit CR,4FDw values similar to those of estuarine and fjord species (Table 3). The lower CRAFDw values reported for tropical species may be related to the fact that they are harboring symbiotic algae. These symbiotic ascid- ian species obtain an important portion of their carbon gain from the photosynthetically fixed carbon (Koike et al. 1993, Koike & Suzuki 1996). Thus, they may only need to process a smaller water volume in order to cover their requirements.
Although the role of detritus in benthic invertebrate nutrition has been widely debated (Levinton et al. 19841, the fate and role of detritus derived from benthic marine macrophytes is an issue of considerable impor- tance in coastal trophic dynamics (Tenore et al. 1982). Models of energy flow derived from estuarine systems suggest that detrital pathways are, by far, the domi- nant route by which seagrass production is utilized by higher trophic levels (e.g. Phillips 1984, Thayer et al. 1984). In general, attention has been focused on detri- tus produced by marine angiosperms (such as marsh and eelgrasses) and utilized by deposit-feeding inver- tebrates (Valiela 1995 and citations therein). However, detritus of macroalgal origin constitutes one of the main sources of the organic carbon pool in the study area (Ballesteros 1991, Ribes 1998), as has been reported in general for coastal waters (Mann 1982). Halocynthia papillosa filtered all items at equal rates, in accord with previous results obtained by Robbins (1983, 1984) that ascidian diets mirror the suspended food con~position. Because detrital POC was always about an order of magnitude higher than live POC (Ribes 1998), organic carbon from detrital origin was the main source of ingested carbon throughout the year. Several studies have shown the importance of the origin of detritus for macroconsumers (Tenore et al. 1982, Crosby et al. 1989, Charles et al. 1996). In con- trast with that of fecal pellets and vascular plants detritus, much of the potential energy of seaweed detritus is readily available (i.e. assimilable) to macro- consumers without the degradation of microbial or- ganisms. Thus, although assimilation rates in ascidians have been shown to be much higher for phytoplankton cells (go%, Fiala-Medioni 1973; 75%, Klumpp 1984) than for organic carbon of detrital origin (42 %, Klumpp 19841, using the assimilation rates from Klumpp (1984) for both carbon sources, the annual mean organic

210 Mar Ecol Prog Ser 175. 201-213, 1.998
Table 3. Clearance rate values (volumes in ml) reported for different species of ascidians and different habitats. DW. dry weight; DW,.,: dry weight without tunic. Indirect methods included: clearance (CR), flowtneter and hot film. Direct method: constant level
tank. Methods terminology after Riisgard & Larsen (1995)
Species m1 g DW-' h-' m1 g DW,, ' h-' Collection site Observations Source
Aplidiuni altarium 2600 Ascidiella aspersa Ascidiella dspersd Ascidiella aspersa Ascidiella aspersa Ciona in testinales Ciona in testinales Ciona intestinales Ciona in testinales Ciona intestinales Clavelina lepadjformis Didemnum molle 2000 Didemnum cf. 3600 albopunctatum Halocynthia papillosa Halocynthia papillosa 162-4823 Lissoclinum bistratum 180 Lissoclinum voeltzkowi 70 Microcosm us sa batieri Phallusia mammillata Phallusia mammillata Phallusia mamnullata Pyura stolonifera Styela clava Styela clava Styela cla va Styela cla va Styela plica ta Styela plicata
Fijian seagrass CR, 25-27°C Southampton estuary, UK Flowmeter, 10°C Southampton estuary, UK Flowmeter, 15°C Southampton estuary, UK CR, 15°C Limfjord, Denmark CR, 20°C Mediterranean CR, 15-16°C Mediterranean CR, 15°C Mediterranean Hot film, 15°C Danish fjord CR, 15'C Limfjord, Denmark CR, 20°C Mediterranean CR, 15-16°C Fijian seagrass CR, 25527°C Fijian seagrass CR, 25-27°C
Mediterranean CR, 15-16°C Mediterranean CR, 12-22°C Fijian seagrass CR, 2 5 2 7 ° C Fijian seagrass CR, 25-27°C Mediterranean CR, 15-16°C Mediterranean CR, 15°C Mediterranean CR, 15°C Mediterranean Hot film, 15°C South Africa CR, 14°C Southampton estuary, UK Flowmeter, 10°C Southampton estuary, UK Flowmeter, 15°C Southampton estuary, UK CR, 15°C North Atlantic Direct method, 15°C Mediterranean CR, 1.5"C Mediterranean Hot film, 15°C
Koike & Suzuki (1996) Holmes (197 3) Holmes (1973) Holmes (19731 Randlov & FhsgBrd (1979) Fiala-Medioni (1978a) Fiala-Medioni (1978a) Fiala-Medionl (1978a) Petersen & hsydrd (1992) Randlov & RLisgard (1979) Fiala-Medloni (1974) Koike & Suzuki (1996) Koike & Suzuki (1996)
Fiala-Medioni (1974) This study Koike & Suzuki (1996) Koike & Suzuki (1996) Fiala-Medioni (1974) Fiala-Medioni (1973) Fiala-Medioni (1978a) Fiala-Mediom (1978a) Klumpp (1984) Holmes (1973) Holmes (1973) Holmes (1973) Riisgard (1988) Fiala-Medlonl (1978a) Fiala-Medioni (1978a)
carbon from detrital origin assimilated by H, papillosa was an order of magnitude higher than that obtained from live carbon (detrital origin: 714 pg C g AFDW-' h-', live carbon: 21 pg C g AFDW-' h-'). Nevertheless, because of its high C:N ratio, the detrital pool is likely to represent a relatively poor nitrogen source for the species in comparison with live carbon (Tenore et al. 1982, Seiderer & Newel1 1985). In this sense, the annual mean consumption of nitrogen from live parti- cles ingested by H. papillosa was of the same order of magnitude as that obtained from detrital particles (live particles [mean + SE]: 10 * 2 pg N g AFDW-l h-'; detri- tal particles: 74 + 29 pg N g AFDW-' h-'). Furthermore, in some periods, such as fall, the importance of live particles as a source of nitrogen was higher than that of detrital particles. Thus, in the feeding ecology of H. papillosa, carbon obtained from procaryotes, pico- and nanoeucaryotes, phytoplankton and ciliates may be an essential source of nitrogen and others nutrients neces- sary for growth and reproduction, especially in certain periods of the year.
Annual variation in growth rate suggests periods of highest growth correlate with those of highest gonadal development. In order to assess the qualitative impor- tance of the detrital and live particles, the seasonal
pattern of both resources as a source of nitrogen was compared to that of gonadal development (but not to growth due to the wide sampling intervals). The sea- sonal variation of nitrogen of detrital origin did not significantly correlate with gonadal development dur- ing the year (multiple regression analysis, p = 0.177). However, the seasona.1 variation of ni.trogen from live particles explained 91 % of gonadal development vari- ance (multiple regression analysis, p < 0.0001). These results suggest that live particles are likely to be of more significance in the diet of th.e species than par- ticles from detrital origin. These results agree with those obtained by Seiderer & Newel1 (1988) for the ascidian Pyura stolonifera in kelp beds. Thus, the main fraction of the particulate material present in the water column appeared to be relatively poorly exploited by some ascidian species, which has been suggested to be due to the lack of the necessary digestive enzymes (Seiderer & Newell 1988).
This study has reported the diet and natural grazing rate of Halocynthia papillosa over an annual cycle, and has shown that growth and reproduction of the species mainly occur during spring and summer This fact is surprising because, in seasonal environments, the highest new production (i.e. growth and reproduction)

Ribes et al.: In situ feedi .ng by a temperate ascidian 211
usually occurs during periods of no limitation by ex- ternal factors (i.e. temperature and food availability), and the Western Mediterranean in summer is charac- terized by oligotrophy and low seston concentration (Zabala & Ballesteros 1989). However, different rea- sons suggest that food availabi!ity dees not s e e 3 to be limiting. First, it has been recently shown that oligo- trophy and low seston during the summer period in the Mediterranean entails mainly detrital POC, but the abundance of live particles remains rather constant through the year (Ribes 1998). Because the ingestion of live particles appears to be the main determinant of growth and reproduction, food availability should not significantly affect the species' secondary production. Furthermore, because food intake of the species was mainly dependent on CRAFDiV seasonality (i.e. maxi- mum CRApDW during sprlng and summer), our work indicates that other factors (such as larval survival, competition, predation) rather than food concentration appear to be determining the seasonal dynamic in growth and reproduction of the species.
Acknotvledgements. We especially thank Mike1 Zabala and Enric Saiz for their help with this study. The manuscript was improved by the comments of Xavier Turon. Joan-Domenec Ros, Quim Garrabou and 3 anonymous reviewers. We thank Emma Cebrian for her help with phytoplankton counts. The flow cytometry measurements were carried out at the Univer- sity of Hawai'i with the assistance of Hector Nolla. Jose- Manuel Fortufio assisted during SEM observations. POC and DOC analyses were provided by the Scientist Technical Ser- vice (University of Barcelona) with the assistance of I. Casals, P. Fernandez and L. Balart. Support for this work was pro- vided by a F.P.I. fellowship from 'Ministerio de Educacion y Ciencia' to M.R., by a MEC research contract to R.C., by a CICYT grant PB94-0014-C02-01 and by the MAST-III- ELOISE European Union METRO h4ED Project.
LITERATURE CITED
Ballesteros E (1991) Structure and dynamics of North-West- ern Mediterranean phytobenthic communities a concep- tual model. In: Ros J , Prat N (eds) Homage to Ramon Mar- galef or. Why there is such pleasure in studying nature. Oecol Aquat 10:223-242
Becerro MA, Turon X (1992) Reproductive cycles of the ascid- ians Microcosmus sabatieri and Halocynthia papillosa in the Nortwestern Mediterranean. PSZN I: Mar Ecol 13: 363-373
Bingham BL, Walters LJ (1989) Solitary ascidians as predators of invertebrate larvae: evidence from gut analyses and plankton samples. J Exp Mar Biol Ecol 131:147-159
Bjarnsen PK (1986) Automatic determination of bacterio- plankton biomass by image analysis. Appl Environ Micro- bio151:1119-1204
Campbell L, Nolla H, Vaulot D (1994) The importance of Prochlorococcus to community structure in the central North Pacific Ocean. Limnol Oceanogr 39:954961
Charles F, Gremare A, Amouroux JM (1996) Ingestion rates and absorption efficiencies of Abra ovata (Mollusca: Bi- valvia) fed on macrophytobenthic detritus. Estuar Coast
Shelf Sci 42:83-102 Clarke A (1987) Temperature, latitude and reproductive
effort. Mar Ecol Prog Ser 3839-99 Cloern J E (1982) Does the benthos control phytoplankton bio-
mass in South San Francisco Bay? Mar Ecol Prog Ser 9: 191-202
Crisp DJ (1984) Energy flow measurements. In: Holme NA, McIntyre AD (eds) Methods for the study of marine ben- thos. Blackwell Scientific Publications, Oxford, p 284-372
Crosby MP, Langdon CJ, Newel1 RIE (1989) Importance of the refractory plant material to the carbon budget of the oyster Crassostrea virginica. Mar Biol 100:343-352
Edler L (1979) Recommendations for marine biological studies in the Baltic Sea. Phytoplankton and chlorophyll. Baltic Mar Biol5:5-38
Fiala-Medioni A (1972) Les peuplements sessiles des fonds rocheux de la region de Banyuls-sur-Mer: ascidians- bryozoaires (2eme partie et fin). Vie Milieu 23:143-182
Fiala-Medioni A (1973) Ethologie alimentaire d'invertebres benthiques filtreurs (ascidians). 1. Dispositif experimental. Taux de filtration et de digestion chez Phallusia mainmil- lata. Mar Biol23:137-145
Fiala-Medioni A (1974) Ethologie alirnentaire d'invertebres benthiques filtreurs (ascidians). 11. Variations des taux de filtration et de digestion en function de l'esphce. Mar Biol 28:199-206
Fiala-Medioni A (1978a) Filter-feeding ethology of benthic invertebrates (ascidians). IV. Pumping rate, filtration rate, filtration efficiency. Mar Biol48:243-249
Fiala-Medioni A (1978b) A scanning electron microscope study of the branchial sac of benthic filter-feeding inverte- brates (ascidians). Acta Zool 59:l-9
Fiala-Medioni A (1978~) Filter-feeding ethology of benthic invertebrates (ascidians). V. Influence of temperature on pumping, filtration and digestion rates and rhythms in Phallusia mammillata. Mar Biol 48:251-259
Fiala-Medioni A (1979a) Effects of oxygen tension on pump- ing, filtration and oxygen uptake in the ascidian Phallusia mamm~llata. Mar Ecol Prog Ser 1:49-53
Fiala-Medioni A (197913) Influence de la concentration algale du milieu sur le pompage, la filtration et I'absorption de Pl~allusia mammillata (Cuvier, 1815) (Ascidie simple). Ann Inst Oceanogr Paris 55:155-162
Fiala-Medioni A (1987) Lower chordates. In: Pandian TJ, Vernberg FJ (eds) Animal energetics, Vol 2. Academic Press, San Diego, p 323-356
Flood PR, Fiala-Medioni A (1981) Ultrastructure and histo- chemistry of the food trapping mucous film in benthic filter-feeders (asc~dians). Acta Zool 62:53-65
Frost BW (1972) Effects of size and concentration of food par- ticles on the feeding behaviour of the marine planktonic copepod Calanuspac~ficus. Limnol Oceanogr 17.805-815
Fry JC (1988) Determination of biomass. In: Austin B (ed) Methods in aquatic bacteriology. Wiley and Sons, New York, p 27-72
Gerdes D, Klages M. Arntz WE, Herman RL, Galeron J , Hain S (1992) Quantitative investigations on macrobenthos communities of the southern Weddell Sea shelf based on multibox core samples. Polar Biol 12:291-301
G*li JM, Coma R (1998) Benthic suspension feeders: their paramount role in littoral marine food webs. Trends Ecol Ev01 13:316-321
Harrison PS, Wallace CC (1990) Reproduction, dispersal and recruitment of scleractinian corals. In: Dubinsky Z (ed) Ecosystems of the world 25. Coral reefs. Elsevier, Amster- dam, p 133-204
Hily C (1991) Is the activity of benthic suspension feeders a

Mar Ecol Prog Ser 175: 201 -213, 1998
factor controhng the water quality in the Bay of Brest? Mar Ecol Prog Ser 69: 179- 188
Holmes N (1973) Water transport in the ascidians Styela clava (Herdman) and Ascidiella aspersa (Miiller). J Exp Mar Biol E C O ~ 11:l-13
Jsrgensen CB (1955) Quantitative aspects of filter-feeding in invertebrates. Biol Rev 30:391-454
Jsrgensen CB (1975) Comparative physiology of suspension feeding. Annu Rev Physiol37:57-59
Jsrgensen CB (1990) Bivalve filter feeding: hydrodynamics, bioenergetics, physiology and ecology. Olsen & Olsen, Fredensborg
Jerrgensen CB, Larsen PS, Riisgdrd HU (1990) Effects of tem- perature on the mussel pump. Mar Ecol Prog Ser 64:89-97
Kana T, Glibert PM (1987) Effect of irradiances up to 2000 pE m-2 s ' ~ on marine Synechococcus WH 7803-1 Growth, pig- mentation, and cell composition. Deep-Sea Res 34: 479-516
Klumpp DW (1984) Nutritional ecology of the ascidian Pyura stolonifera: influence of body size, food quantity and qual- ity on filter-feeding, respiration, assimilation efficiency and energy balance. Mar Ecol Prog Ser 19:269-284
Koike 1, Suzuki T (1996) Nutritional diversity of symbiotic ascidians in a Fijian seagrass meadow. Ecol Res 11: 381 -386
Koike I, Yamamuro M, Pollard PC (1993) Carbon and nitrogen budgets of two ascidians and their symbionts, Prochloron, in a tropical seagrass meadow. Aust J Mar Freshwat Res 44:173-182
Levinton JS, Bianchi TS, Stewart S (1984) What is the role of particulate organic matter in benthic invertebrate nutri- tion? Bull Mar Sci 35:270-282
Lucas A (1996) Bioenergetics of aquatic animals. Taylor & Francis, London
Mann KH (1982) Ecology of coastal waters. A system approach. Blackwell Scientific Publications, Oxford
~Massana R, Gasol JM, Bjornsen PK, Blackburn N, Hagstrom A, Hietanen S, Hygum J , Kupannen BH, Pedros-Alio C (1997) Measurement of bacterial size via Image analysis of epifluorescence preparations: description of an inexpen- sive system and solutions to some of the most common problems. Sci Mar 61:397-407
Millar RH (1971) The biology of ascidians. Adv Mar Biol 9: 1-100
Monger BM, Landry MR (1993) Flow cytometric analysis of marine bacteria with Hoechst 33342. Appl Environ Micro- bio159:905-911
Montagnes DJ, Berges SJA, Harrison PJ, Taylor FJR (1994) Estimating carbon, nitrogen, protein and chlorophyll a from volume in marine phytoplankton. Limnol Oceanogr 39: 1044-1060
Newell RC, Bayne BL (1980) Seasonal changes in the physio- logy, reproductive condition and carbohydrate content of the cockle Cardium (Cerastoderma) edule (Bivalvia: Cardildae). Mar Biol 56:ll-19
NeweLl RC, Fleld JG, Griffiths CL (1982) Energy balance and significance of micro-organisms in a kelp bed community. Mar Ecol Prog Ser 8:103-113
Officer CB, Smayda TJ, Mann R (1982) Benthic filter feedlng: a natural eutrophication control. Mar Ecol Prog Ser 9. 203-210
Okamura B (1990) Behavioural plasticity in the suspension feeding of benthic animals. In: Hughes RN (ed) Behav- ioural mechanisms of food selection. Springer-Verlag, Berlin, p 637-660
Orton JH (1920) Sea temperature, breeding and distribution in marine animalb. J Mar Biol Assoc UK 12:339-366
Petersen JK, Riisgard HU (1992) Filtration capacity of the ascidian Ciona intestinalis and its grazing impact in a shallow fjord. Mar Ecol Prog Ser 88:9-17
Petersen JK, Schou 0, Thor P (1995) Growth and energetics in the ascidian Ciona intestinalis. Mar Ecol Prog Ser 120: 175-184
Philhps RC (1984) The ecology of eelgrass meadows in the Pacific Northwest: a community profile. US Fish Wildl Serv, FSW/OBS-84/24
Putt M, Stoecker DK (1989) An experimentally determined carbon volume ratio for marine 'oligotrichous' ciliates from estuarine and coastal waters. Limnol Oceanogr 34: 1097-1 103
Ramos AA (1988) Ascidias litorales del Mediterrdneo Iberico: faunistica, ecologia y biogeografia. PhD thesis, Univ of Barcelona
Randlsv A, Riisgdrd HU (1979) Efficiency of particle retention and filtration rate in four specles of ascidians. Mar Ecol Prog Ser 1:55-59
Ribes M (1998) Feeding activity and diet of benthic suspen- sion feeders related to metabolic requirements and seston composition. PhD thesis, Unversity of Barcelona
Ribes M, Coma R, Gili JM (1998) Heterotrophic feeding by gorgonian corals with symbiotic zooxanthella. Limnol Oceanogr 43:1170-1179
Riisgird HU (1988) The ascidian pump: properties and energy cost. Mar Ecol Prog Ser 47:129-t34
Robbins IJ (1983) The effects of body size, temperature, and suspension density on the filtration and ingestion of inor- ganic particulate suspensions by ascidians. J Exp Mar Biol Ec01 70:65-78
Robbins IJ (1984) The regulation of ingestion rate, at high suspended particulate concentrations, by some phl.eo- branchiate ascidians. J Exp Mar Rio1 Ecol 82:l.-10
Saiz E (1993) Sources of variability in zooplankton feeding experiments: the importance of accurate determination of algal growth rates. Sci Mar 57:23-29
Seiderer LJ, Newell RC (1985) Relat~ve significance of phyto- plankton, bacteria and plant detritus as carbon and nitro- gen resources for the kelp bed filter-feeder Choromytilus meridionalis. Mar Ecol Prog Ser 22:127-139
Seiderer U, Newell RC (1988) Exploitation of phytoplankton as a food resource by the kelp bed ascidian Pyura slolo- nifera. Mar Ecol Prog Ser 50:107-115
Simon M, Azam F (1989) Protein content and protein synthe- sis rates of planktonic marine bacteria. Mar Ecol Prog Ser 51:201-213
Sokal RR, Rohlf FJ (1981) Biometry. 2nd edn. Freeman, San Francisco
Stuart V, Klumpp DW (1984) Evidence for food-resource par- titioning b;kelp-bed filter feeders. Mar Ecol Prog se; 16. 27-37
Tenore KR, Camm.en L, Findlay SEG, Phillips N (1982) Per- spectives of research on detntus: do factors controlling the availability of detritus to macroconsumers depend on its source? J Mar Res 40:473-490
Thayer GW, Kenworthy WJ, Fonseca MS (1984) The ecology of eelgrass meadows of the Atlantic coast. a community profile. US Fish Wildl Serv, FSWIOBS-84/02
Turon X (1988) The ascidians of Tossa de Mar (NE Spain) 11.- Biological cycles of the colonial species. Cah Biol Mar 29: 407-418
Turon X (1990a) Distribution and abundance of ascidians from a locality on the Northeast Coast of Spain. PSZN I: Mar Ecol 11:291-308
Turon X (1990b) Estructura de 10s filtros branquiales de ascidias (Tunicados). Misc Zoo1 14:125-133

Ribes et al.: In situ feeding by a temperate ascidian 213
Turon X (1992) Penods of non-feeding in Polysjmcraton lacaze~ (Ascidiacea: Didemnidae): a rejuvenative process? Mar Biol 11 2:647-655
Turon X, Becerro MA (1992) Growth and survival of several ascidian species from the northwestern Mediterranean Mar Ecol Prog Ser 82:235-247
Valiela I (1995) Marine ecological processes. Springer-Verlag, New York
Vaulot D, Partensky F, Neveus J, Mantoura RFC, Llewellyn C (1990) Winter presence of prochlorophytes in surface waters of the northwestern Mediterranean Sea. Limnol Oceanogr 35:1156-1164
Editorial responsibility: Roger Hughes (Contributing Editor), Bangor, Gwynedd, Wales
Verity PG, Robertson CY, Tronzo CR, Andrews MG, Nelson JR . Sieracki ME (1992) Relationships between cell volume and the carbon and nitrogen content of marine photo- synthetic nanoplankton. Limnol Oceanogr 37:1434-1446
Vogel S (1994) Life in moving fluids. The physical biology of flow. Princeton University Press, Pr~nceton, NJ
Yoi~nrj CM (1988) Ascidian cannibalism correlates with larval behaviour and adult distribution. J Exp Mar Biol Ecol 117: 9-26
Zabala M, Ballesteros E (1989) Surface-dependent strategies and energy flux in benthic marine communities or, why corals do not exist in the Mediterranean. Sci Mar 53:3-17
Submitted: June 1, 1998; Accepted: September 10, 1998 Proofs received from author(s): November 26, 1998