role of a-type potassium currents in excitability, network synchronicity, and epilepsy
TRANSCRIPT

Role of A-Type Potassium Currents in Excitability,Network Synchronicity, and Epilepsy
Erik Fransen1,2* and Jenny Tigerholm1,2
ABSTRACT: A range of ionic currents have been suggested to beinvolved in distinct aspects of epileptogenesis. Based on pharmacologi-cal and genetic studies, potassium currents have been implicated, inparticular the transient A-type potassium current (KA). Epileptogenic ac-tivity comprises a rich repertoire of characteristics, one of which issynchronized activity of principal cells as revealed by occurrences offor instance fast ripples. Synchronized activity of this kind is particularlyefficient in driving target cells into spiking. In the recipient cell, thissynchronized input generates large brief compound excitatory postsy-naptic potentials (EPSPs). The fast activation and inactivation of KA leadus to hypothesize a potential role in suppression of such EPSPs. In thiswork, using computational modeling, we have studied the activation ofKA by synaptic inputs of different levels of synchronicity. We find thatKA participates particularly in suppressing inputs of high synchronicity.We also show that the selective suppression stems from the current’sability to become activated by potentials with high slopes. We furthershow that KA suppresses input mimicking the activity of a fast ripple.Finally, we show that the degree of selectivity of KA can be modified bychanges to its kinetic parameters, changes of the type that are producedby the modulatory action of KChIPs and DPPs. We suggest that thewealth of modulators affecting KA might be explained by a need tocontrol cellular excitability in general and suppression of responses tosynchronicity in particular. We also suggest that compounds changingKA-kinetics may be used to pharmacologically improve epileptic status.VVC 2009 Wiley-Liss, Inc.
KEY WORDS: epileptogenesis; fast ripples; synchronicity; dendriticpotentials; transient A-type potassium current; Kv4.2
INTRODUCTION
In epileptogenesis, a range of ionic currents have been implicated, forexample potassium currents (Johnston et al., 2000; Birnbaum et al.,2004; Timofeev et al., 2004). It is for instance, well known that applica-tion of the K-channel blocker 4-AP induces seizures (Voskuyl and Albus,1985), but as glutamate also is released the interpretation is complicated.
However, more specific support that in particular thetransient A-type potassium current (KA) may beinvolved comes from studies where seizures wereinduced by application of the selective KA blockerscorpion toxin PiTx-Ka (Juhng et al., 1999). Further-more, in the methylazoxymethanol model of corticalmalformations and epilepsy (Castro et al., 2001), thehippocampal heterotopia showed loss of the KA-cur-rent. Moreover, the heterotopia also lacked expressionof functional Kv4.2 potassium channels, which is oneof the genes coding for KA currents in brain (Yuanet al., 2005; Chen et al., 2006). Finally, temporal lobeepilepsy has been reported in a patient with a geneticKv4.2 truncation mutation (Singh et al., 2006).
KA has fast activation and relatively fast inactiva-tion. As viewed from the soma, it belongs to thethreshold currents activated near the spiking thresholdand in this respect it produces the well known delayof an action potential. KA is present in pyramidal celldendrites (Hoffman et al., 1997; Korngreen and Sak-mann, 2000), where it affects the backward spread ofaction potentials, as well as integration of excitatorypostsynaptic potentials (EPSPs) (Hoffman et al.,1997; Ramakers and Storm, 2002; Kim et al., 2007).Importantly, it also has the property to suppress rapidlarge compound EPSPs (Urban and Barrionuevo,1998). Such EPSPs may be present during fast ripples.Fast ripples are discrete high frequency oscillations(HFOs) in the range 200–600 Hz, which have beenrecorded in human mesial temporal epilepsy (Braginet al., 1999), and will be further discussed below. Epi-leptogenic activity and in particular fast ripples arecharacterized by spiking at high synchronicity levels(Bragin et al., 2000, 2002a, 2005; Lasztoczi et al.,2004). Such activity will at target cells produce syn-chronous compound EPSPs characterized by fast risetimes, large amplitudes, and short duration. Wehypothesized that the properties of KA, with its fastactivation, would be suitable to suppress this voltageprofile more than one from less synchronous input.Less synchronized activity would due to its higherDC-level, and smaller and slower EPSP-fluctuationsalso trigger the relatively slower KA inactivation andthereby prevent the current from suppressing theEPSP. In vivo, action potentials most commonly arisefrom brief dendritic depolarizations reflecting synchro-nous synaptic inputs (Azouz and Gray, 1999, 2003).KA may therefore reduce how effective synchronousinput is in generating action potentials in a neuron.
1Department of Computational Biology, School of Computer Scienceand Communication, Royal Institute of Technology, Stockholm, Sweden;2 Stockholm Brain Institute, Royal Institute of Technology, Stockholm,SwedenAdditional Supporting Information may be found in the online version ofthis article.Grant sponsor: Swedish VR; Grant number: 621-2007-4223; Grant spon-sor: NIH; Grant number: MH061492-06A2.*Correspondence to: Erik Fransen, Department of Computational Biology,School of Computer Science and Communication, Royal Institute ofTechnology, AlbaNova University Center, SE-106 91 Stockholm, Sweden.E-mail: [email protected] for publication 15 July 2009DOI 10.1002/hipo.20694Published online 23 September 2009 in Wiley InterScience (www.interscience.wiley.com).
HIPPOCAMPUS 20:877–887 (2010)
VVC 2009 WILEY-LISS, INC.

Availability and integrity of dendritic KA currents may thus bean important component regulating recruitment of neuronsinto seizure activity.
Fast ripples are field oscillations between 200 and 600 Hz witha broad peak in the power spectrum around, e.g., 250 Hz (Engelet al., 2009) or 350 Hz (Dzhala and Staley, 2004). These events,recorded from human focal epileptic brain, occur mostly in regionsof primary epileptogenesis and rarely in regions of secondaryspread (Jirsch et al., 2006). The phenomena is suggested to dependon simultaneous onset of activity in excitatory synaptic circuits(Bragin et al., 2002, 2002b; Grenier et al., 2003; Dzhala and Sta-ley, 2004). It is evident that principal neurons cannot sustain firingat these high rates, as found by Lasztoczi et al. (2004) and dis-cussed by Foffani et al. (2007) and one suggestion has been thatthere may exist populations of neurons firing at lower subhar-monics and that the populations alternate and thereby togethergenerate this high frequency (Staley, 2007). Moreover, the fre-quency of fast ripples may not be the main factor distinguishing afast ripple from, e.g., a ripple (Engel et al., 2009) and frequenciesof fast ripples may reach into the interval commonly attributed toripples. Furthermore, increases in synchronicity appear to precedeincreases in frequency (e.g., transitions into fast ripples) shortlybefore onset of epileptogenic activity (Lasztoczi et al., 2004). More-over, neurons are more synchronized in fast ripples than in ripples(Lasztoczi et al., 2004; Engel et al., 2009). We have for these rea-sons focused on the increase in synchronicity and studied whetherKA is differentially more activated by fast ripples than ripples. Toevaluate whether such a dynamic perspective would work quantita-tively, we performed biophysical simulations. It may be noted thatwe will not deal with the synchronizing mechanism per se, inwhich inhibition from interneurons have been suggested to play amajor role, but investigate the response of a neuron to synchron-ized input, and thereby the mechanism of recruitment of principalneurons into seizure activity.
METHODS
Neuron Model
The model used in this work was based on the model byMigliore et al. (1999), which was downloaded from theModelDB (Hines et al., 2004) database http://senselab.med.yale.edu/modeldb. This model has been used by Hoffman et al.(1997), Migliore et al. (1999), Poolos et al. (2002) in their workon CA1 pyramidal cell dendrite processing, specifically the roleof the KA and h-currents in backpropagating action potentials.During initial tests, it was found that the Na-current conduct-ance had to be reduced in the dendritic compartments by 1/8and enhanced in the soma compartments by 5.6 (giving a rela-tive conductance difference of 45) to ensure that synapticallyinduced action potentials were induced in the soma and not inthe dendrites. This is consistent with combined experimentaland computational findings where an �50 times higher Na-channel density at the site of initiation of action potentials hasbeen found (Meeks and Mennerick, 2007; Kole et al., 2008).
Moreover, to simplify our analysis of dendritic processing, wedid not make the distinction as in the original model betweendistal and proximal characteristics of KA, but used only the distalmodel throughout the dendrite. In a subsequent sensitivity testwhere effects of additional currents were analyzed, the influenceof another threshold current, the h-current, as well as the influ-ence of a persistent sodium current, was studied. The model hada resting potential of 265 mV, which well corresponds to valuesfound in vivo (Crochet et al., 2006). The EPSP amplitude in thesoma remaining for a synaptic input at 171 lm was 41%. Thiscorresponds well to experimental findings of 38% remainder forlacunosum moleculare inputs on CA1 pyramidal cells (Inoueet al., 2001) and 78–29% remainder in CA1 cells for inputs at100 and 300 lm (Magee, 2000).
Model of Synaptic Input
CA1 pyramidal cells have several thousand synapses distrib-uted over the dendritic tree. The proportion of neurons, andthus also synapses, that is active at any point in time is gener-ally low, and this is particularly true for hippocampus (Buzsakiet al., 1983; Frank et al., 2001). Thus, ten synaptic inputswere included in the neuron model, representing the synapsesthat become activated at a particular period in time in a re-stricted part of the dendrite. For each synapse, we generatedPoisson distributed synaptic events at 12 Hz. The simulationwas run for 1.5 s leading to 17 synaptic inputs for the case offull synchronicity. Synaptic inputs of kainate/AMPA-type had arise time constant of 1.5 ms and a decay time constant of2.5 ms and were positioned on the apical dendrite 171 lmfrom the soma. Experimentally, EPSP amplitudes as measuredin the soma vary greatly and range from 0.13 to 2.8 mV(Markram et al., 1997) and from 0 to 4 mV (Thomson et al.,1995). We used inputs generating 3.1 mV in the soma, whichthus were 7.5 mV in the dendrite at the point of input (leadingto the reduction factor 0.41 discussed above). For the case withKA present, the synaptic inputs were increased 41% to compen-sate for the hyperpolarizing effect of KA. The different levels ofsynchronicity were modeled according to Charcos Llorens andFransen (2004). Briefly, different inputs were separated with aPoisson distribution within a time window, and the relative sizeof the window in relation to the base frequency of 12 Hzdetermines the degree of synchronicity.
Model of Fast Ripples
Although much is known about the phenomenology of fastripples, the mechanisms behind them are not known. Wetherefore modeled them following two different experimentalmodels, one in vivo and one in vitro.
Case 1
The model is based on studies of fast ripples generated in aslice preparation (Dzhala and Staley, 2004). To model fastripple input to the target neuron, the model uses a sequence ofinterspike interval (ISI) intervals from data of spontaneously
878 FRANSEN AND TIGERHOLM
Hippocampus
generated burst-like discharges in individual neurons evoked inhigh potassium concentration (Dzhala and Staley, 2004) withthe following consecutive intervals: 3.8, 4.5, 4.9, 6.8, 7.2, 8.6,9, 10, 10, 10 ms. The corresponding ISI series from spontane-ous burst-type intervals for control levels of potassium concen-tration were as follows: 5, 7, 9, 10, 10, 10, 10, 10, 10, 10 ms(Dzhala and Staley, 2004). This input was used to representnormal spiking patterns not associated with epileptogenic activ-ity. If KA works selectively to suppress epileptogenic activity, itis not expected to be significantly activated by this kind ofinput. A real neuron is assumed to have many such inputs ofthe type described above spread over the dendritic tree. In themodel, ten synaptic inputs were provided at five different loca-tions of the dendritic tree. In the control case, the ten synaptictime series were evenly spaced with a time delay of 2.5 ms,leading to the average delay between bursts of 6 ms as reportedby Dzhala and Staley (2004); see also Supporting InformationFigure 1. For the fast ripple the delay was decreased to 0.15ms, consistent with the observed decrease toward zero delays inhigh potassium (Dzhala and Staley, 2004).
Case 2
Input from a ripple versus input from a fast ripple. Ripples inawake animals range from 80 to 200 Hz (Ylinen et al., 1995)and are, contrary to fast ripples, associated with normal brain ac-tivity. A ripple episode commonly consist of some 4–16 cycles ofoscillations (Ylinen et al., 1995, our estimate). A neuron partici-pates (spikes) on average in 11% of the cycles, and at most in
40% (Ylinen et al., 1995). Focusing on the neurons making upthe core of a ripple, we have assumed neurons participate in25% of the cycles and produce a total of four spikes each.Assuming individual neurons fire at 50 Hz during the ripple, a25% involvement gives an ensemble frequency of 100 Hz con-
taining 16 cycles if neurons fire in four groups 90 degrees out ofphase. The target neuron was subjected to 50 synaptic inputs,10 each on five different dendrite locations, with the temporalstructure just described; see Supporting Information Figure 2.Moreover, the 10 inputs at a site were evenly spaced by 1.1 ms(Ylinen et al., 1995). Fast ripples are more synchronized than
ripples (Bragin et al., 2002a; Lasztoczi et al., 2004; Engel et al.,
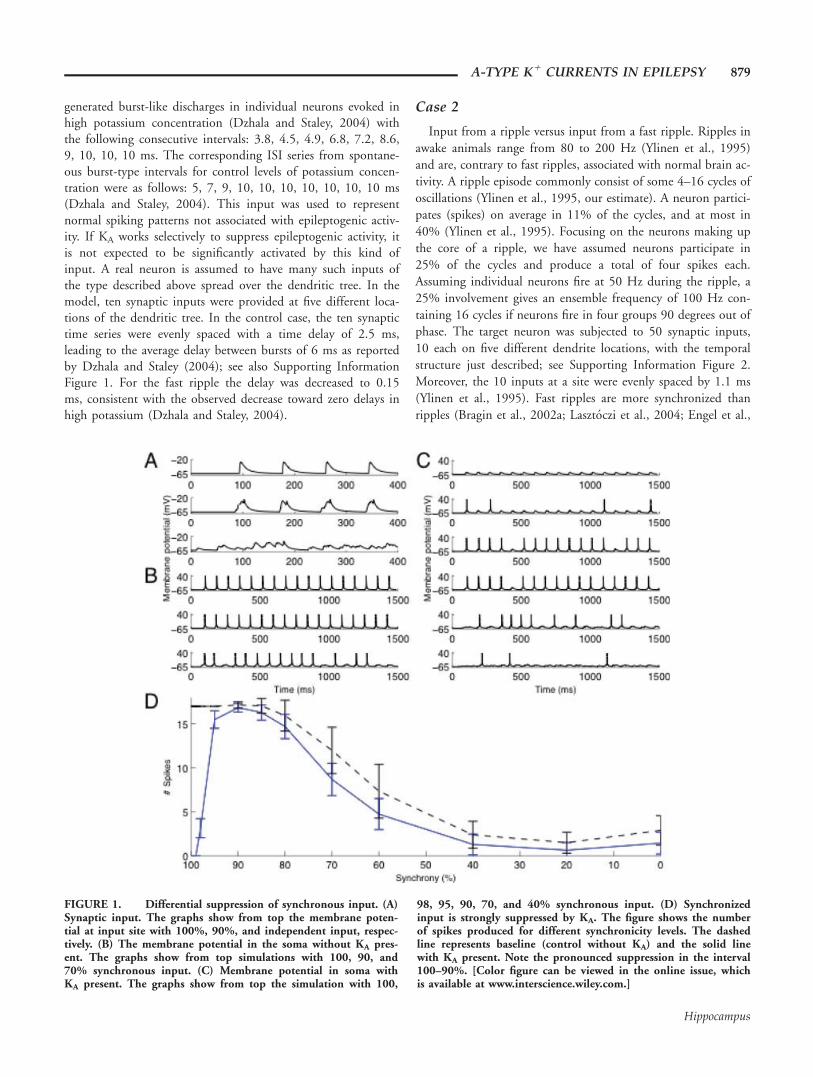
FIGURE 1. Differential suppression of synchronous input. (A)Synaptic input. The graphs show from top the membrane poten-tial at input site with 100%, 90%, and independent input, respec-tively. (B) The membrane potential in the soma without KA pres-ent. The graphs show from top simulations with 100, 90, and70% synchronous input. (C) Membrane potential in soma withKA present. The graphs show from top the simulation with 100,
98, 95, 90, 70, and 40% synchronous input. (D) Synchronizedinput is strongly suppressed by KA. The figure shows the numberof spikes produced for different synchronicity levels. The dashedline represents baseline (control without KA) and the solid linewith KA present. Note the pronounced suppression in the interval100–90%. [Color figure can be viewed in the online issue, whichis available at www.interscience.wiley.com.]
A-TYPE K1 CURRENTS IN EPILEPSY 879
Hippocampus

2009). Assuming a four times higher frequency during a fast ripplecompared to a ripple (e.g., 400 Hz and 100 Hz respectively) a spac-ing of 1.1 ms in a ripple would correspond to a spacing of 0.27 ms
in a fast ripple. Thus, to test the synchronicity aspect of a fast rip-ple, a ‘‘synchronized’’ ripple was used. In this case, the time lagbetween the 10 spikes within one cycle of a ripple was reduced to0.3 ms. We subsequently studied whether KA would be more acti-vated and thereby to a larger extent suppress activity of a fast ripple.
RESULTS
To investigate the potential capacity of KA to differentiallysuppress EPSPs depending on their level of synchronicity, theneuron model was subjected to 11 runs each of a particular levelof synchronicity. In Table 1, we show the median time betweenEPSPs as a function of the synchronicity level. We measured thenumber of spikes produced by the input as a measure of KA sup-pression. The rationale behind this choice is that, even thoughwe are considering KA as a postsynaptic dendritic excitability fac-tor, network activity mediated by action potentials is the ulti-mate component of epileptogenic activity. The neuronal trans-formation of inputs into spiking is therefore a key element.
Differential Suppression
The effect on the postsynaptic dendritic membrane forinputs of three different levels of synchronicity in the absenceof KA is shown in Figure 1A. It can be seen that with decreas-ing levels of synchronicity, amplitude and rate of rise of thecompound EPSP decrease but variability increases. In Figure1B, we show the soma membrane spiking pattern at these levels
of synchronicity in the absence of KA. In Figure 1C KA is pres-ent. The different panels show number of spikes produced andone can infer how many are suppressed at different levels ofsynchronicity. In Figure 1D, we summarize the neuronresponse with and without KA present. Error bars are based on20 repetitions of the procedure with different starting randomseeds. In general, without KA, higher levels of synchronicitygenerate more spikes, consistent with the general properties ofa neuron as discussed in the Introduction. But, with KA pres-ent, there is a substantial suppression of efficacy of inputs forhigh degrees of synchronicity in the interval 90–100%. Thelevel 90% corresponds to EPSPs separated by a median of0.7 ms and an interval ranging from 0.1 to 2.2 ms (10 and90% of the events, respectively). The particular shape of thecurve, particularly of the peak, depends on the synapticstrength, which is further studied in the Supporting Informa-tion. Thus, as can be seen from Figure 1, while KA, as a potas-sium current, will always be suppressive, it has a selective effecton the synchronous input that otherwise is so powerful in acti-vating a neuron.
KA Current Amplitude and Activation
To further analyze the contribution of KA in this differentialsuppression, we show in Figure 2 the activation, inactivation,and current through KA at two different levels of synchronicity.We compare two cases, one fully synchronized and the other at80% level of synchronicity. For comparison, the two instancesshown produced the same peak potential (Fig. 2A) around17 ms. As can be seen, KA gives more current (second trace),opens more (third trace), and is less inactivated (bottom trace),at the higher level of synchronicity. Note that this is true eventhough the case of less synchronized input leads to spiking ofthe cell (around 18 ms). The figure also shows that this is adynamic effect caused by the interaction between the activationand the inactivation dynamics. More specifically, this is seen byreplacing the instantaneous activation and inactivation withtheir steady-state values (dashed traces) computed using themembrane potential and steady-state activation/inactivationcurves (shown in Supporting Information Fig. 3). Usingsteady-state values, differences in kinetics between activationand inactivation are removed. This can be seen most clearly forthe relatively slower inactivation (trace D, in particular between
FIGURE 2. Activation of KA by synchronized versus semi-synchronized input. Synchronous input (100%), in grey, activatesKA more than semisynchronous input (80%), in black. The dashedlines represent values of KA steady-state activation and inactivationat the membrane potentials dictated by A. (A) Membrane potentialin the soma. Inset shows initial slope of EPSP more clearly. (B)Current through KA at input site. Note the difference in currentaround 6 ms. (C) Inactivation of KA at input site. The interval 2–10 ms shows that the effect seen in B originates from the dynamicaspects of KA. (D) Activation of KA at input site. Note the differ-ence in inactivation around time of input 2–10 ms. [Color figurecan be viewed in the online issue, which is available atwww.interscience.wiley.com.]
TABLE 1.
Time Between Inputs
100% 90% 70% 40% Independent
Median 0 0.67 2.01 2.68 6.71
10% 0 0.096 0.29 0.57 0.96
90% 0 2.15 6.45 12.9 21.5
The table shows time (in ms) between synaptic inputs for a select number ofsynchronization levels of Figure 1. In the table, Median refers to the medianvalue in the distribution of input intervals produced, 10% refers to the shortestintervals, and 90% covers almost all the intervals produced.
880 FRANSEN AND TIGERHOLM
Hippocampus

4 and 10 ms), which at steady-state balances out activation(trace C) yielding very little current (trace B). Further analysisof the dynamics of KA confirmed that the inactivation is partic-ularly important in the suppression; it must be fast enough,but not as fast as the activation. This influence of the inactiva-tion time constant is further studied in relation to modulatoryeffects on KA described below.
Slope of Compound EPSP
One major difference between the fully synchronized andthe one at the 80% level of synchronicity is the slope (deriva-tive) of the rising phase of the EPSP [Figs. 1A and 2A (inset)].To further analyze the influence of the derivative on thedifferential activation of KA, we conducted voltage clampsimulations providing triangular ramps as inputs to the locationof the synaptic input (Fig. 3A). To enable comparisons of casesof different derivatives, we kept the area of the triangular vol-tages constant, as higher potential and longer time always givemore activation and more current. As can be seen in Figure3B, higher derivatives produced higher activation of KA. Thesedifferences can be seen both in the amplitude of the currentand in the integral of the current. Thus, in general, inputs ofhigher synchronicity produce compound EPSPs with higherderivatives, which as shown here activate KA more.
Spatial Extent of KA Activation
In addition to its sensitivity to the derivative of the voltage,KA as a voltage-gated channel is sensitive to membrane poten-tial. EPSPs propagating in the dendrite decay with distancefrom the point of synaptic input, and we were interested tostudy the spatial region of activated KA currents. In Figure 4A,we show the membrane potential at input site. The EPSPappears around the time point 10 ms and the backpropagating
action potential around 20 ms. In Figure 4B, we show the cur-rent of KA at several locations around the point of synapticinput. It can be seen that the peak of KA current is relativelylocal, about a third of the peak remaining 46 lm more proxi-mally, but that points even 100 lm away are activated. In thefigure, around the time point 20 ms, it can also be seen howKA is affected by a backpropagating action potential. Eventhough the backpropagating action potential amplitude and therate of rise are larger than those for the EPSP, the backpropa-gating action potential is preceded by the EPSP that producesa partial inactivation of KA.
Selectivity of the Suppression ofSynchronous Input
KA is modulated by a range of factors, affecting the conduct-ance of the current as well as its activation and inactivation char-acteristics, and we will return to this in the Discussion section.One potential consequence of such changes is changes in thesuppression profile of different synchronicity levels. We havestudied the effects on the inactivation time constant produced byauxillary subunits of KA, KChIPs and DPPs (Nakamura et al.,2001a; Jerng et al., 2004, 2007; Gutman et al., 2005). In Figure5, we show the effect on suppression by using different KA
inactivation time constants. As can be seen in Figure 5, both theselectivity of suppression with regard to high levels of synchro-nicity (interval of high suppression, position of the peak ofactivation) and the degree of suppression of lower levels of syn-chronicity (level of suppression at independent input, rate ofdecline at intermediate levels of input) are attenuated. We willreturn to the modulatory action on inactivation kinetics in ouranalysis of suppression of fast ripples below. Furthermore,
FIGURE 3. Sensitivity to voltage slope. The top graph showsvoltage clamp command of different initial slope but same area.The bottom graph shows the corresponding current through theKA channel. Note the difference in the peak current at 25 ms. Thelegend indicates the integral of the current for each protocol.[Color figure can be viewed in the online issue, which is availableat www.interscience.wiley.com.]
FIGURE 4. Distance-dependent KA activation. The top graphshows the membrane potential at input site (171 lm from soma)with a 92% level of synchronous input. The first event is theEPSP and second is the backpropagating action potential. The bot-tom graph shows the current through the KA channel. The currentwas measured at five different locations: 125, 164, 171, 247, and340 lm from the soma. [Color figure can be viewed in the onlineissue, which is available at www.interscience.wiley.com.]
A-TYPE K1 CURRENTS IN EPILEPSY 881
Hippocampus

auxillary subunits as well as modulators like PKA and PKC pro-duce shifts in the steady-state activation and to a lesser extentalso the inactivation curves (Hoffman and Johnston, 1998;Nakamura et al., 2001a; Jerng et al., 2004, 2007; Gutman et al.,2005). When auxillary subunits produce a hyperpolarizing shift,the produced KA current is increased (data not shown). Con-versely, PKA and PKC produce shifts in the depolarizing direc-tion, leading to an effective reduction of KA current and as aconsequence a loss of differential suppression (data not shown).The results thus show that the suppressive characteristics of KA
may be changed and may potentially be adjusted to the func-tional demands of the network.
Suppression of Epileptogenic Activity
As discussed in the Introduction, experimental observationsindicate the appearance of fast ripples at or before the onset ofseizures. We have investigated to what extent KA is able to sup-press a fast ripple. In the first study, we generated input to aneuron from in vitro data on spontaneous bursts in individualneurons (Dzhala and Staley, 2004) generated either at controllevels or at high concentrations of potassium, the latter beingused as a model of epiletogenic activity. Figure 6 shows theresult comparing activity produced under control levels ofpotassium to ‘‘fast ripples’’ generated under high levels of potas-sium, the latter case tested both with and without KA present.As can be seen, KA does not suppress input under the controlcondition, but it does so in the fast ripple case. The fast ripplecase without KA shows that without KA the cell spikes as underthe control condition. Thus, a neuron containing KA would toa lesser extent participate in the fast ripple event, and spread ofthe fast ripple event might therefore be reduced or prevented.
In the second case, we started from a model based on invivo data on ripples (Ylinen et al., 1995). From this, we gener-ated fast ripples by increasing the synchronicity, consistent withthe observation by Bragin et al. (2002a), Lasztoczi et al. (2004)and Engel et al. (2009). We tested both inputs at a frequencyof 50 and 100 Hz, effectively representing oscillations of higherharmonics like 100 and 200 Hz as suggested by Staley (2007)and discussed in the Introduction. As can be seen in Figure 7,the control case with input replicating a ripple does not getsuppressed, but input from the more synchronized fast ripple issuppressed if KA is present. These results are also consistentwith the findings of Figure 1 showing differential suppressionfor comparable spike ISIs in the window 90% (no suppres-sion)–100% (total suppression). We were also interested tostudy the modulatory influence by, e.g., KChIPs and DPPs on
FIGURE 5. The selectivity to synchronized input varies whenKA dynamics is modified. The inactivation time constant waschanged representing modulatory influences of, e.g., auxillary pro-teins. As in Figure 1, the solid black curve is the result with standardvalues of KA. The dashed dark grey, medium grey, light grey, black,dark grey and light grey curves represent decreasing inactivation timeconstant in steps of 1 ms. Dashed black and solid grey curves repre-sent increasing inactivation time constant in steps of 1 ms. Note thesignificant difference in reduction of synchronized input, effectivelycovering the interval 95–80%. [Color figure can be viewed in theonline issue, which is available at www.interscience.wiley.com.]
FIGURE 6. KA reduces response to fast ripple input. The firstcolumn represents the membrane potential of the soma. Thesecond column represents the current through the KA channelmeasured at the location of the synaptic input. The third columnrepresents the gating variables of the KA channel. The black linerepresents activation and the grey line inactivation. (A) Fast ripplewith no KA present. Input represents higher synchronicity levelspresent under high potassium concentration conditions. (B) Fast
ripple with KA present. Note the reduction in number of actionpotentials generated due to the increase in KA current. The plot ofgating variables shows that this input leads to a larger degreeof activation, particularly at its peaks, and to a lesser degree ofinactivation, particularly for later inputs. (C) Control input repre-senting lower synchronicity present in control concentration ofpotassium. [Color figure can be viewed in the online issue, whichis available at www.interscience.wiley.com.]
882 FRANSEN AND TIGERHOLM
Hippocampus

the suppression of fast ripples found. In Table 2, we summarizeour findings. Enhanced modulation (reduced inactivation timeconstant) leads to a reduced current as the inactivation kineticsapproaches that of activation (as discussed in Fig. 2) andthereby to reduced suppression of fast ripples. Decreasedmodulation on the other hand leads to increased current and asa result suppression that also affects ripple activity.
As mentioned, the genesis of fast ripple events is not known.If a group of neurons would receive several inputs within asmall window of time (due to random fluctuations or someparticular process), it is conceivable that they might spike andthat the synchronous activity thereby could spread to otherneurons. We investigated whether KA would oppose such a sin-gle synchronous input. Analyzing the data from Figure 1 at thesynchronicity level of 98%, the probability of the first spike ofthe input is 0.15, the probability of a spike following the sec-ond input is 0.35, and for the third it is 0.3. Thus, the firstinput is more suppressed than the following two. Thus, KA isnot only able to suppress trains of inputs, but also single ones.KA may therefore also be a part in preventing the initiation offast ripples and seizures.
DISCUSSION
In this work, we have shown that the KA current selectivelysuppresses synchronized inputs. This property may be particu-larly important in the context of epiletogenesis, wheresynchronized activity, e.g., in the form of fast ripples isobserved at the onset of seizure activity. Preventing the spreadof highly synchronized input may be an important mechanismin the control of seizures.
Neocortical Pathological Ripples
We have here used a model of a CA1 pyramidal cell.However, KA is also present in neocortical pyramidal cells(Korngreen and Sakmann, 2000; Schaefer et al., 2003; Yuanet al., 2005). Moreover, although there are differences betweenthe pathological high frequency ripples observed in neocortexcommonly denoted ‘‘neocortical ripples’’ compared to thoseobserved in hippocampal structures termed ‘‘fast ripples,’’ thereare also commonalities. Grenier et al. (2003) present evidencethat neocortical ripples occur at the time of transition to ictalevents. There is furthermore a strong correlation between neu-ronal excitation and the intensity of neocortical ripples (Grenieret al., 2003). More specifically, neocortical ripples duringseizures involve coordinated action potentials in a majority ofneurons synchronized on the millisecond scale (Grenier et al.,
FIGURE 7. KA reduces response to fast ripple input. The firstthree columns correspond to the simulation when the input fre-quency was 50 Hz and the fourth column a case with 100 Hz input.The first column represents the membrane potential of the soma.The second column represents the current through the KA channelmeasured at the location of the synaptic input. The third columnrepresents the gating variables of the KA channel. The black line
represents activation and the grey line inactivation. (A) Fast rippleinput with no KA present. (B) Fast ripple input with KA present.Note the significant reduction in spike activity. It can be seen thatthe inactivation is lower than that under the control case C, partic-ularly in its troughs. (C) Ripple input. KA does not suppress inputrepresenting ripple activity. [Color figure can be viewed in theonline issue, which is available at www.interscience.wiley.com.]
TABLE 2.
Effects on Fast Ripples From Changes in Inactivation Kinetics
50 Hz 100 Hz
2 c 1 2 c 1
Without KA 4 4 4 4 4 4
Fast ripple input 0 0 3 0 0 3
Ripple input 3 4 4 2 4 4
Number of spikes produced following fast ripple input (Case 2).c, control, standard value of the inactivation time constant, i.e., same data as infirst and fourth columns of Figure 7; 1, enhanced modulation by reducing theinactivation time constant by 4 ms representing enhanced KChIP/DPP modula-tion; 2, reduced modulation by increasing the inactivation time constant by 4ms representing reduced KChIP/DPP modulation.
A-TYPE K1 CURRENTS IN EPILEPSY 883
Hippocampus

2003). Our results based on hippocampal fast ripples maytherefore be of more general nature.
Separate Roles for Inhibition and KA
In this work, the focus is on the spread of synchronousactivity among excitatory neurons. We have thus studied theresponse of a neuron to synchronized input and have not dealtwith the synchronizing mechanism per se, in which inhibitionfrom interneurons has been suggested to play a major role(Magloczky and Freund, 2005). Moreover, activity of interneur-ons may also play a minor role in the generation of ripples andfast ripples. More specifically, during ripples, interneurons werereported to fire at higher frequencies than that of the ripples,and there was no clear modulation of firing by ripples (Grenieret al., 2003), e.g., the interneurons are not directly pacing theripples. On the other hand, neocortical ripples duringnonseizure states depend on IPSPs (Grenier et al., 2003). Thus,while inhibition from interneurons may play a major roleduring seizures, synchronized activity of principal cells may bemore important for the initial spread of activity.
Synchronization in Nonrhythmic Events
Synchronous activity is not only present during oscillations,but may also be present during nonrhythmic events relevantfor epilepsy. Large field potential spikes, for instance, reflectepileptic network activity (Ulbert et al., 2004). Comparisons ofhuman intracranial EEG recordings and spontaneous events inin vitro slice preparations taken from the same patient showthey have spike and waves events resembling each other (Cohenet al., 2002). In these slices, intracellular recordings show highdegrees of synchronization during the spike and wave events.Thus, in both rhythmic and nonrhythmic events, synchronizedneuronal firing is present and this synchronous input may beparticularly effective in recruiting additional neurons.
Amplitude of Compound EPSP
Our results show that the differential suppressive effect ofKA increases with increasing EPSP amplitude. CompoundEPSPs generated by multiple inputs in concert may indeedreach large amplitudes. For instance, amplitudes of sharp wavesof 8 mV have been recorded intracellularly (Maier et al.,2003). In pathological circuits comprising the start of epilepticactivities, synaptic efficacies may be even larger. KA may there-fore be particularly important in preventing hyperexcitabilityunder those circumstances.
Role of Modulators of KA
A number of different substances modulate KA, for instance,PKA (Hoffman and Johnston, 1998), PKC (Hoffman andJohnston, 1998), MAPK (Yuan et al., 2002), arachidonic acid(AA) (Colbert and Pan, 1999; Ramakers and Storm, 2002), ad-enyl cyclase activating peptide (Han and Lucero, 2005, 2006),and frequentin (Nakamura et al., 2001b); for a review, seeJerng et al. (2004). Furthermore, potassium channels colocalize
with membrane proteins, notably the Kv channel-interactingproteins (KChIP) and dipeptidyl peptidase-like proteins (DPP),e.g., DPP6 and DPP10; for a review, see Jerng et al. (2004).The presence of KChIPs and DPPs in neurons enhances thesurface expression of KA. Moreover, it has been shown thatKChIPs produce substantial changes in KA characteristics, e.g.,shifting the steady-state activation and inactivation curves 10sof millivolts as well as changing inactivation kinetics(Nakamura et al., 2001a; Gutman et al., 2005; Jerng et al.,2007). Moreover, DPPs produce a hyperpolarizing shift ofsteady-state curves and a more rapid inactivation kinetics(Jerng et al., 2007). There may be several reasons for thiscomplex regulation of KA: (1) General excitability, the controlof the threshold for action potential generation, latency ofspike, or number of spikes produced by somatic input; (2)backpropagation of action potentials, specifically a control ofspread of backpropagating action potentials; (3) interactionwith incoming EPSPs and potentially synaptic plasticity; (4)modulation of efficacy of synchronized input, in particular,modulating the degree of specificity of this suppression, asshown in Figure 5. Our results also suggest that effects fromPKA and PKC would work in the detrimental direction,whereas KChIPs and DPPs would enhance the suppressiveeffects of KA on hypersynchronous activity.
A Vicious Circle Involving KA
We have in this work discussed how reduction of KA may bea risk factor in epilepsy. However, the opposite may also betrue. In hippocampal tissue after pilocarpine induced seizures,CA1 pyramidal neuron dendrites showed decreased availabilityof the A-type potassium channel due to transcriptionaland post-translational processes (Bernard et al., 2004). Further-more, after convulsant drug pentylenetetrazole treatment(Tsaur et al., 1992; Francis et al., 1997), Kv1.2 and Kv4.2mRNAs were reduced. Thus, there seems to be a bidirectionalrelationship between seizures on the one hand and expressionand electrophysiological properties of KA on the other, andthus a possibility of a vicious circle.
Anticonvulsants and KA
Preventing or reversing the vicious circle of KA run-downmay be a key objective in treatment of seizures. Indeed, severalanticonvulsants have been shown to enhance KA currents:lamotrigine (Zona et al., 2002, but see Huang et al. 2004),valproate (Walden et al., 1993), and carbamazepine (Zonaet al., 1990). Conversely, GABAB activation reduces potassiumcurrents by shifting their activation in the positive direction(Saint et al., 1990). This interaction could work via the sup-pressive effect of AA on KA (Ramakers and Storm, 2002) acti-vated by phospholipase A2 activation downstream of GABAB.This suppression of KA could be one explanation for less goodresults of GABA-enhancing compounds on some patients.GABAB activation could on the one hand lead to a beneficialdirect hyperpolarization of the neuron, but on the other handalso an indirect enhancement of temporal summation capacity
884 FRANSEN AND TIGERHOLM
Hippocampus

and an increased sensitivity to synchronous activation. In thiswork, we have investigated the potential link between KA andneural responsiveness to synchronous synaptic input. Based onthe results in this work, the prediction would be a decrease insynchronicity following administration of these compounds.Indeed, lamotrigine, which enhances KA (Zona et al., 2002),reduces population spikes (Langoscha et al., 2000), and popula-tion spikes have been suggested to be a sign of neuronal syn-chrony (Andersen et al., 1971). Furthermore, interictal spikesare reduced by lamotrigine (Marciani et al., 1996) and theaddition of lamotrigine decreases EEG ictal and interictalabnormalities (Akman and Holmes, 2003). A second antiepi-lepticum, valproate, has been shown to reduce the frequency ofextracellularly recorded spontaneous interictal bursts (Albus andWilliamson, 1998).
Properties of Potassium Current Enhancers
Observations in epileptic patients of increased excitabilitydue to compromized potassium currents have led to the recentdevelopment of potassium current enhancers [Retigabine andFlupirtine enhancing the M-type (KCNQ/Kv7) channel,Nicorandil enhancing an adenosine triphosphate-sensitivepotassium (K1ATP) channel, and Chlorzoxazone as an openerof a calcium-sensitive potassium current], and with their antie-pileptic properties evaluated in Kobayashi et al. (2008).However, K-current-enhancing drugs may not always givethe desired effect. Resting membrane potential changes inducedby a K-current enhancer may be counteracted by homeostaticmechanisms (Marder and Goaillard, 2006) either reducingother K-currents or enhancing depolarizing currents. Like-wise, influences on average firing frequency may be balancedout in the long run. Based on the work presented here, phar-macological interventions that affect the neuron response tohighly synchronized inputs but less so for intermediate orunsynchronized inputs are suggested.
Acknowledgments
We thank Asad Rustum and Vicente Charcos Llorens fortheir work during their Masters Thesis projects.
REFERENCES
Akman CI, Holmes GL. 2003. The effect of lamotrigine on the EEGs ofchildren and adolescents with epilepsy. Epilepsy Behav 4:420–423.
Albus H, Williamson R. 1998. Electrophysiologic analysis of theactions of valproate on pyramidal neurons in the rat hippocampalslice. Epilepsia 39:124–139.
Andersen P, Bliss TVP, Skrede KK. 1971. Unit analysis of hippocam-pal population spikes. Exp Brain Res 13:208–221.
Azouz R, Gray CM. 1999. Cellular mechanisms contributing toresponse variability of cortical neurons in vivo. J Neurosci19:2209–2223.
Azouz R, Gray CM. 2003. Adaptive coincidence detection anddynamic gain control in visual cortical neurons in vivo. Neuron37:513–523.
Bernard C, Anderson A, Becker A, Poolos NP, Beck H, Johnston D.2004. Acquired dendritic channelopathy in temporal lobe epilepsy.Science 305:532–535.
Birnbaum SG, Varga AW, Yuan LL, Anderson AE, Sweatt JD,Schrader LA. 2004. Structure and function of Kv4-family transientpotassium channels. Physiol Rev 84:803–833.
Bragin A, Engel J Jr, Wilson CL, Fried I, Mathern GW. 1999. Hippo-campal and entorhinal cortex high-frequency oscillations (100–500Hz) in human epileptic brain and in kainic acid-treated rats withchronic seizures. Epilepsia 40:127–137.
Bragin A, Wilson CL, Engel J Jr. 2000. Chronic epileptogenesisrequires development of a network of pathologically intercon-nected neuron clusters: A hypothesis. Epilepsia 41 (Suppl 6):S144–S152.
Bragin A, Wilson CL, Staba RJ, Reddick M, Fried I, Engel J Jr.2002a. Interictal high-frequency oscillations (80–500 Hz) in thehuman epileptic brain: Entorhinal cortex. Ann Neurol 52:407–415.
Bragin A, Mody I, Wilson CL, Engel J Jr. 2002b. Local generation offast ripples in epileptic brain. J Neurosci 22:2012–2021.
Bragin A, Azizyan A, Almajano J, Wilson CL, Engel J Jr. 2005. Analy-sis of chronic seizure onsets after intrahippocampal kainic acidinjection in freely moving rats. Epilepsia 46:1592–1598.
Buzsaki G, Lai-Wo L, Vanderwolf CH. 1983. Cellular bases ofhippocampal EEG in the behaving rat. Brain Res Rev 6:139–171.
Castro PA, Cooper EC, Lowenstein DH, Baraban SC. 2001. Hippo-campal heterotopia lack functional Kv4.2 potassium channels inthe methylazoxymethanol model of cortical malformations and epi-lepsy. J Neurosci 21:6626–6634.
Charcos Llorens V, Fransen E. 2004. Intrinsic desynchronization prop-erties of neurons containing dendritic rapidly activating K-currents.Neurocomputing 58–60:137–143.
Chen X, Yuan LL, Zhao C, Birnbaum SG, Frick A, Jung WE,Schwarz TL, Sweatt JD, Johnston D. 2006. Deletion of Kv4.2gene eliminates dendritic A-type K1 current and enhances induc-tion of long-term potentiation in hippocampal CA1 pyramidalneurons. J Neurosci 26:12143–12151.
Cohen I, Navarro V, Clemenceau S, Baulac M, Miles R. 2002. Onthe origin of interictal activity in human temporal lobe epilepsy invitro. Science 298:1418–1421.
Colbert CM, Pan E. 1999. Arachidonic acid reciprocally alters theavailability of transient and sustained dendritic K(1) channelsin hippocampal CA1 pyramidal neurons. J Neurosci 19:8163–8171.
Crochet S, Fuentealba P, Cisse Y, Timofeev I, Steriade M. 2006. Syn-aptic plasticity in local cortical network in vivo and its modulationby the level of neuronal activity. Cereb Cortex 16:618–631.
Dzhala VI, Staley KJ. 2004. Mechanisms of fast ripples in the hippo-campus. J Neurosci 24:8896–8906.
Engel J Jr, Bragin A, Staba R, Mody I. 2009. High-frequencyoscillations: What is normal and what is not? Epilepsia 50:598–604.
Foffani G, Uzcategui YG, Gal B, Menendez de la Prida L. 2007.Reduced spike-timing reliability correlates with the emergence offast ripples in the rat epileptic hippocampus. Neuron 55:930–941.
Francis J, Jugloff DG, Mingo NS, Wallace MC, Jones OT, BurnhamWM, Eubanks JH. 1997. Kainic acid-induced generalized seizuresalter the regional hippocampal expression of the rat Kv4.2 potas-sium channel gene. Neurosci Lett 232:91–94.
Frank LM, Brown EN, Wilson MA. 2001. A comparison of the firingproperties of putative excitatory and inhibitory neurons from CA1and the entorhinal cortex. J Neurophysiol 86:2029–2040.
Grenier F, Timofeev I, Steriade M. 2003. Neocortical very fast oscilla-tions (ripples, 80–200 Hz) during seizures: Intracellular correlates.J Neurophysiol 89:841–852.
A-TYPE K1 CURRENTS IN EPILEPSY 885
Hippocampus

Gutman GA, Chandy KG, Grissmer S, Lazdunski M, McKinnon D,Pardo LA, Robertson GA, Rudy B, Sanguinetti MC, Stuhmer W,Wang X. 2005. International Union of Pharmacology. LIII. No-menclature and molecular relationships of voltage-gated potassiumchannels. Pharmacol Rev 57:473–508.
Han P, Lucero MT. 2005. Pituitary adenylate cyclase activatingpolypeptide reduces A-type K1 currents and caspase activity incultured adult mouse olfactory neurons. Neuroscience 134:745–756.
Han P, Lucero MT. 2006. Pituitary adenylate cyclase activating poly-peptide reduces expression of Kv1.4 and Kv4.2 subunits underlyingA-type K(1) current in adult mouse olfactory neuroepithelia. Neu-roscience 138:411–419.
Hines ML, Morse T, Migliore M, Carnevale NT, Shepherd GM.2004. ModelDB: A database to support computational neuro-science. J Comput Neurosci 2004 17:7–11.
Hoffman DA, Johnston D. 1998. Downregulation of transient K1channels in dendrites of hippocampal CA1pyramidal neurons byactivation of PKA, PKC. J Neurosci 18:3521–3528.
Hoffman DA, Magee JC, Colbert CM, Johnston D. 1997. K1 chan-nel regulation of signal propagation in dendrites of hippocampalpyramidal neurons. Nature 387:869–875.
Huang CW, Huang CC, Liu YC, Wu SN. 2004. Inhibitory effect oflamotrigine on A-type potassium current in hippocampal neuron-derived H19-7 cells. Epilepsia 45:729–736.
Inoue M, Hashimoto Y, Kudo Y, Miyakawa H. 2001. Dendriticattenuation of synaptic potentials in the CA1 region of rat hippo-campal slices detected with an optical method. Eur J Neurosci13:1711–1721.
Jerng HH, Pfaffinger PJ, Covarrubias M. 2004. Molecular physiologyand modulation of somatodendritic A-type potassium channels.Mol Cell Neurosci 27:343–369.
Jerng HH, Lauver AD, Pfaffinger PJ. 2007. DPP10 splice variants arelocalized in distinct neuronal populations and act to differentiallyregulate the inactivation properties of Kv4-based ion channels. MolCell Neurosci 35:604–624.
Jirsch JD, Urrestarazu E, LeVan P, Olivier A, Dubeau F, Gotman J.2006. High-frequency oscillations during human focal seizures.Brain 129:1593–1608.
Johnston D, Hoffman DA, Poolos NP. 2000. Potassium channels anddendritic function in hippocampal pyramidal neurons. Epilepsia41:1072–1073.
Juhng KN, Kokate TG, Yamaguchi S, Kim BY, Rogowski RS, Blaus-tein MP, Rogawski MA. 1999. Induction of seizures by thepotent K1 channel-blocking scorpion venom peptide toxinstityustoxin-K(alpha) and pandinustoxin-K(alpha). Epilepsy Res34:177–186.
Kim J, Jung SC, Clemens AM, Petralia RS, Hoffman DA. 2007. Reg-ulation of dendritic excitability by activity-dependent trafficking ofthe A-type K1 channel subunit Kv4.2 in hippocampal neurons.Neuron 54:933–947.
Kobayashi K, Nishizawa Y, Sawada K, Ogura H, Miyabe M. 2008.K(1)-channel openers suppress epileptiform activities induced by4-aminopyridine in cultured rat hippocampal neurons. J PharmacolSci 108:517–528.
Kole MH, Ilschner SU, Kampa BM, Williams SR, Ruben PC, StuartGJ. 2008. Action potential generation requires a high sodium chan-nel density in the axon initial segment. Nat Neurosci 11:178–186.
Korngreen A, Sakmann B. 2000. Voltage-gated K1 channels in layer5 neocortical pyramidal neurones from young rats: Subtypes andgradients. J Physiol 3:621–639.
Langoscha JM, Zhoua X-Y, Grunzeb H, Waldena J. 2000. Newinsights into the mechanisms sites of action of lamotrigine. Neuro-psychobiology 42 (Suppl 1):26–27.
Lasztoczi B, Antal K, Nyikos L, Emri Z, Kardos J. 2004. High-fre-quency synaptic input contributes to seizure initiation in the low-[Mg21] model of epilepsy. Eur J Neurosci 19:1361–1372.
Magee J. 2000. Dendritic integration of excitatory synaptic input. NatRev Neurosci 1:181–190.
Magloczky Z, Freund TF. 2005. Impaired and repaired inhibitory cir-cuits in the epileptic human hippocampus. Trends Neurosci28:334–340.
Maier N, Nimmrich V, Draguhn A. 2003. Cellular and networkmechanisms underlying spontaneous sharp wave-ripple complexesin mouse hippocampal slices. J Physiol 550(Pt 3):873–887.
Marciani MG, Spanedda F, Bassetti MA, Maschio M, Gigli GL, Mat-tia D, Bernardi G. 1996. Effect of lamotrigine on EEG paroxysmalabnormalities and background activity: A computerized analysis. BrJ Clin Pharmacol 42:621–627.
Marder E, Goaillard J. 2006. Variability, compensation and homeo-stasis in neuron and network function. Nat Rev Neurosci 7:563–574.
Markram H, Lubke J, Frotscher M, Roth A, Sakmann B. 1997. Physi-ology and anatomy of synaptic connections between thick tuftedpyramidal neurones in the developing rat neocortex. J Physiol500:409–440.
Meeks JP, Mennerick S. 2007. Action potential initiation andpropagation in CA3 pyramidal axons. J Neurophysiol 97:3460–3472.
Migliore M, Hoffman DA, Magee JC, Johnston D. 1999. Role of anA-type K1 conductance in the back-propagation of action poten-tials in the dendrites of hippocampal pyramidal neurons. J ComputNeurosci 7:5–15.
Nakamura TY, Nandi S, Pountney DJ, Artman M, Rudy B, CoetzeeWA. 2001a. Different effects of the Ca(21)-binding protein,KChIP1, on two Kv4 subfamily members, Kv4.1 and Kv4.2. FEBSLett 499:205–209.
Nakamura TY, Pountney DJ, Ozaita A, Nandi S, Ueda S, Rudy B,Coetzee WA. 2001b. A role for frequenin, a Ca21-binding pro-tein, as a regulator of Kv4 K1-currents. Proc Natl Acad Sci USA98:12808–12813.
Poolos N, Migliore M, Johnston D. 2002. Pharmacological upregula-tion of h-channels selectively reduces the excitability of pyramidalneuron dendrites. Nat Neurosci 5:767–774.
Ramakers GM, Storm JF. 2002. A postsynaptic transient K(1) currentmodulated by arachidonic acid regulates synaptic integration andthreshold for LTP induction in hippocampal pyramidal cells. ProcNatl Acad Sci USA 99:10144–10149.
Saint DA, Thomas T, Gage PW. 1990. GABAB agonists modulate atransient potassium current in cultured mammalian hippocampalneurons. Neurosci Lett 118:9–13.
Schaefer AT, Helmstaedter M, Sakmann B, Korngreen A. 2003.Correction of conductance measurements in non-space-clampedstructures: 1. Voltage-gated K1 channels. Biophys J 84:3508–3528.
Singh B, Ogiwara I, Kaneda M, Tokonami N, Mazaki E, Baba K,Matsuda K, Inoue Y, Yamakawa K. 2006. A Kv4.2 truncationmutation in a patient with temporal lobe epilepsy. Neurobiol Dis24:245–253.
Staley KJ. 2007. Neurons skip a beat during fast ripples. Neuron55:828–830.
Thomson AM, West DC, Deuchars J. 1995. Properties of single axonexcitatory postsynaptic potentials elicited in spiny interneurons byaction potentials in pyramidal neurons in slices of rat neocortex.Neuroscience 69:727–738.
Timofeev I, Grenier F, Steriade M. 2004. Contribution of intrinsicneuronal factors in the generation of cortically driven electro-graphic seizures. J Neurophysiol 92:1133–1143.
Tsaur ML, Sheng M, Lowenstein DH, Jan YN, Jan LY. 1992. Differ-ential expression of K1 channel mRNAs in the rat brain anddown-regulation in the hippocampus following seizures. Neuron8:1055–1067.
Ulbert I, Magloczky Z, Eross L, Czirjak S, Vajda J, Bognar L, Toth S,Szabo Z, Halasz P, Fabo D, Halgren E, Freund TF, Karmos G.
886 FRANSEN AND TIGERHOLM
Hippocampus

2004. In vivo laminar electrophysiology co-registered with histol-ogy in the hippocampus of patients with temporal lobe epilepsy.Exp Neurol 187:310–318.
Urban NN, Barrionuevo G. 1998. Active summation of excitatorypostsynaptic potentials in hippocampal CA3 pyramidal neurons.Proc Natl Acad Sci USA 95:11450–11455.
Voskuyl RA, Albus H. 1985. Spontaneous epileptiform discharges inhippocampal slices induced by 4-aminopyridine. Brain Res342:54–66.
Walden J, Altrup U, Reith H, Speckmann EJ. 1993. Effects of val-proate on early and late potassium currents of single neurons. EurNeuropsychopharmacol1 3:137–141.
Ylinen A, Bragin A, Nadasdy Z, Jando G, Szabo I, Sik A, Buzsaki G.1995. Sharp wave-associated high-frequency oscillation (200 Hz) inthe intact hippocampus: Network and intracellular mechanisms.J Neurosci 15(1 Pt 1):30–46.
Yuan LL, Adams JP, Swank M, Sweatt JD, Johnston D. 2002. Proteinkinase modulation of dendritic K1 channels in hippocampusinvolves a mitogen-activated protein kinase pathway. J Neurosci22:4860–4868.
Yuan W, Burkhalter A, Nerbonne JM. 2005. Functional role of thefast transient outward K1 current IA in pyramidal neurons in (rat)primary visual cortex. J Neurosci 25:9185–9194.
Zona C, Tancredi V, Palma E, Pirrone G, Avoli M. 1990. Potassiumcurrents in rat cortical neurons in culture are enhanced by the anti-epileptic drug carbamazepine. Can J Physiol Pharmacol 68:545–547.
Zona C, Tancredi V, Longone P, D’Arcangelo G, D’Antuono M,Manfredi M, Avoli M. 2002. Neocortical potassium currents areenhanced by the antiepileptic drug lamotrigine. Epilepsia 43:685–690.
A-TYPE K1 CURRENTS IN EPILEPSY 887
Hippocampus