report - circulation researchcircres.ahajournals.org/content/circresaha/early/2009/06/11/... ·...
TRANSCRIPT

Report
Avoidance of Transient Cardiomyopathy inCardiomyocyte-Targeted Tamoxifen-InducedMerCreMer Gene Deletion Models
Norimichi Koitabashi, Djahida Bedja,Ari L. Zaiman, Yigal M. Pinto,Manling Zhang, Kathleen L. Gabrielson,Eiki Takimoto, David A. Kass
Cardiac myocyte targeted MerCreMer transgenic miceexpressing tamoxifen-inducible Cre driven by the�-myosin heavy chain promoter are increasingly used tocontrol gene expression in the adult heart. Here, we showtamoxifen-mediated MerCreMer (MCM) nuclear trans-location can induce severe transient dilated cardiomyop-athy in mice with or without loxP transgenes. The car-diomyopathy is accompanied by marked reduction ofenergy/metabolism and calcium-handling gene expres-sion (eg, PGC1-�, peroxisome proliferator-activated �,SERCA2A), all fully normalized with recovery. MCM-negative/flox-positive controls display no dysfunctionwith tamoxifen. Nuclear Cre translocation and equallyeffective gene knockdown without cardiomyopathy isachievable with raloxifene, suggesting toxicity is not sim-ply from Cre. Careful attention to controls, reducedtamoxifen dosing and/or use of raloxifene is advised withthis model.
The Cre-loxP system is widely used for selective cell-targeted manipulation of gene expression1 and has been
further enhanced by generating tamoxifen-responsive fusionproteins for conditional Cre induction.2,3 Targeted cells con-stitutively express Cre flanked by mutated estrogen receptorligand-binding domains (MerCreMer [MCM]) insensitive toendogenous estrogen but sensitive to tamoxifen (TAM).MCM is cytoplasmic via binding to heat shock protein 90complex, but this complex dissociates on TAM-Mer binding,whereupon the MCM targeting sequence sends the constructto the nucleus for Cre-mediated excision of loxP flankedsequences.1 Sohal et al linked MCM with an �-myosin heavychain (Myh6) promoter to create cardiomyocyte-specific gene
targeting.3 However, Cre recombinase displays dose-dependent cytotoxicity impairing growth and causing DNAfragmentation,4,5 and a recent review raised a caution thatTAM-stimulated MCM in adult hearts may also adverselyinfluence heart function.6 Here, we report on these cardiaceffects and provide methods to avoid them.
Materials and MethodsMyh6-MerCreMer�/� transgenic mice (no. 005650, Jackson Labs,Bar Harbor, Me) were used. Myh6-MCM�/�/no-flox were gener-ated by mating to C57Bl/6 mice. Two strains with floxed allelescoding for either R2 (Tgfbr2fl/fl) or R1 (Alk5fl/fl) transforminggrowth factor (TGF)� receptors (both on C57Bl/6 backgrounds)were crossed with Myh6-MCM�/� mice to study gene knock-down. Cardiac function was assessed by serial echocardiographyand invasive pressure–volume analysis. Gene expression wasdetermined by real-time PCR, gene knockdown was determinedby analysis of mRNA and TGF�-stimulated Smad2 phosphory-lation, and nuclear Cre was determined by immunohistochemistryand immunoblot. Details are provided in the Online Data Supple-ment, available at http://circres.ahajournals.org.
ResultsIn both MCM�/�/Tgfbr2fl/fl and MCM�/�/Alk5fl/fl mice,TAM administered at 20 mg/kg body weight (BW) IP for 5days (proposed dose3) was insufficient for gene and func-tional knockdown (latter assessed by suppression of TGF�-stimulated Smad2 phosphorylation; Figure 1A and 1B; FigureI, A and B). Increasing the dose to 80 mg/kg BW per day for5 days (IP) resulted in 60% mortality by 6 days after TAMtreatment because of severe cardiomyopathy (Online FigureII). Oral delivery of the same dose for 7 days was tolerated(no mortality) and effective at gene and functional TGF�-receptor knockdown attained (Figure 1A and 1B; OnlineFigure I, A and B). However, a marked though reversibledilated cardiomyopathy (Figure 1C and Online Figure I) wasalso observed in both floxed/MCM�/� models and MCM�/�
mice without a floxed transgene. MCM-negative controlswith or without floxed genes (eg, MCM�/�/Tgfbr2fl/fl) devel-oped no myopathy at any TAM dose. Cardiac-depressionpeaked �3 days after terminating TAM (day 10 of protocol)with fractional shortening declining from 61�1% to26.5�5% (P�0.01) and end-diastolic dimension increasing(3.2�0.1 to 4.1�1.4 mm; P�0.01) in MCM�/� mice (withor without floxed alleles; n�19). In vivo pressure–volumeanalysis in MCM�/�/no flox mice confirmed marked tran-sient systolic and diastolic depression (Figure 1D and OnlineTable I), with full recovery observed by day 28 (3 weeks afterstopping TAM). MCM�/� controls had no TAM-induceddysfunction. Myocardium displayed patchy interstitial mono-nuclear infiltration at day 10 (mild myocarditis) that resolvedby day 28 and no myocyte hypertrophy (Online Figure III).
TAM/MCM-induced cardiomyopathy was accompanied bymarked changes in stress response, energy/metabolism, andcalcium-handling genes (Figure 2 and Online Figure V). Natri-uretic peptide expression (Nppa and Nppb) rose markedly inMCM�/� versus MCM�/� by day 10 and then returned tonormal, although �-myosin heavy chain (Myh7), which typi-cally rises with cardiac stress, was unchanged. Peroxisome
Original received April 1, 2009; revision received May 26, 2009;accepted June 1, 2009.
From the Divisions of Cardiology (N.K., M.Z., E.T., D.A.K.) andPulmonary and Critical Care Medicine (A.L.Z.), Department of Medi-cine; and Division of Comparative Medicine (D.B., K.L.G.), JohnsHopkins Medical Institutions, Baltimore, Md; and University of Amster-dam (Y.M.P.), Heart Failure Research Center, The Netherlands.
This manuscript was sent to David Eisner, Consulting Editor, forreview by expert referees, editorial decision, and final disposition.
Correspondence to: David A. Kass, MD, Division of Cardiology,Johns Hopkins Medical Institutions, Ross Research Building, Room 858,720 Rutland Ave, Baltimore, MD 21205. E-mail [email protected]
(Circ Res. 2009;105:00-00.)© 2009 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.orgDOI: 10.1161/CIRCRESAHA.109.198416
1
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from

proliferator-activated receptor (PPAR)�, PPAR�-coactivator(PGC)1�, and transcription factor A-mitochondrial (TFAM)genes, which are centrally involved with coordinating mitochon-drial function, energetics, and metabolism7 and suspected toplay a key role in dilated human cardiomyopathy,8 alldeclined substantially with cardiac depression and then fullyrecovered to normal levels. Lastly, both sarcoplasmic retic-ular ATPase and phospholamban expression declined tran-siently, correlating with cardiac function. Although these
changes are observed with various cardiac failure condi-tions, the insult in this instance started in the nucleus andits striking reversibility unusual. Although Cre toxicitymight be suspected, reversibility would be less anticipatedfrom DNA fragmentation, particularly in differentiatedtissue without a high rate of cell regeneration.
Raloxifene (RAL) is an alternative selective estrogenreceptor modulator with similarities but also differenceswhen comparing TAM-regulated transcription. This may be
Figure 2. Influence of tamoxifen on gene transcription in MCM�
hearts. Stress response (A) and mitochondrial/bioenergetic regu-latory (B) and calcium-handling (C) gene expression in MCM�/�
or MCM�/� mice (neither with floxed allele). No Tx indicates noTAM; Day 10 and Day 28, as previously defined with TAM Tx.Data normalized as in Figure 1A (n�4 each group). *P�0.01 vsDay10 MCM�/�; †P�0.05 vs No Tx MCM�/� (1-way ANOVA,Tukey test).
Figure 1. Tamoxifen-mediated cardiomy-opathy in MCM mice. A, Expression ofTgfbr2 normalized to Gapdh and then todata in MCM�/� mice. Gene knockdownin MCM�/�/Tgfbr2fl/fl is not achieved at20 mg/kg per day IP for 5 days (recom-mended dose3) but required 80 mg/kgper day PO for 7 days (n�4 to 6 eachgroup,*P�0.05 vs MCM�/�). B, Repre-sentative immunoblot for Smad2 phos-phorylation stimulated by recombinanthuman TGF�1 (rhTGF�: 5 ng/mL, 30 min-utes) in cardiomyocytes from MCM�/�/Tgfbr2fl/fl mice. Summary data in OnlineFigure I. C, Effective TAM-induced geneknockdown is accompanied by transient,reversible cardiomyopathy. FS indicatesfractional shortening; EDD, end-diastolicdimension. *P�0.05 vs MCM�/�/Tgfbr2fl/fl
(n�7 for each group). See also OnlineFigure II and Online Table I. D, In vivopressure–volume loops in MCM�/� vsMCM�/� mice (no floxed allele) at base-line and day 10 and 28 of protocol (TAM,80 mg/kg per day for 7 days) in MCM�/�
mice. Bottom, Results for contractility(preload recruitable stroke work [PRSW]and end-systolic elastance [Ees] andrelaxation [Tau]). *P�0.05 vs other groups(1-way ANOVA, Tukey test; n�4 for eachgroup).
2 Circulation Research July 2, 2009
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from

attributable to differential binding to estrogen-related recep-tors (eg, ERR�)9 and/or recruitment of different coactiva-tors.10 Because RAL interacts with Mer, albeit at lowerbinding affinity,11 we tested whether RAL could induce geneknock-down without cardiomyopathy. Because of poor solu-bility, DMSO was required for IP dosing, limiting the dose to�40 mg/kg BW per day, which was suboptimal for geneknockdown in MCM�/�/Alk5fl/fl. However, higher oral doseswere feasible, tolerated, and effective. Myh6-MCM�/� micefed 160 mg/kg BW per day RAL PO displayed effective geneknockdown but without cardiac dysfunction (Figure 3A and3B; Online Figure VI). Nuclear Cre targeting was similarwith TAM or RAL treatments, as shown by histochemistry(Figure 3C) and nuclear fraction immunoblot (Online FigureVII). Stress, metabolic, and Ca2�-handling gene changes (eg,Figure 2) were not observed (data not shown). Importantly,gene knockdown efficacy with RAL was similar as with 80mg/kg BW per day TAM, although required longer exposure(21 days) (Figure 3B and Online Figures VIII and IX).Recombination (lox-P site excision) was observed earlierwith RAL (7 days), although at lower levels. Both RAL andTAM resulted in a similar �10% decline in BW during thefirst week that subsequently recovered (n�6 to 7/group;P�0.95, 2-way ANOVA). Lower oral TAM dose (20 mg/kgBW per day, one-third previously reported12) for 21 days alsoinduced effective gene knockdown without dysfunction (On-line Figure V).
DiscussionOur study did not precisely define the mechanism for TAM-MCM cardiac effects, and such analysis falls outside thescope and intent of this report. However, the data raise a
novel hypothesis that the cardiotoxicity is not simply attrib-utable to Cre. First, the striking reversibility is difficult toreconcile with mechanisms of cell damage attributed to Cre,namely targeting pseudo loxP sites to cause DNA fragmen-tation, cell growth arrest, and/or death.4,5 Second, the findingthat both TAM and RAL induced similar nuclear Cre local-ization and gene suppression, yet with striking differences incardiac phenotype, further questions a Cre-toxicity mecha-nism. An alternative relates to the specific nature of theligand–MCM complex. In addition to recruiting differentnuclear coactivators that can differentially target transcrip-tion,10 TAM but not RAL can inhibit ERR�, which, alongwith ERR�, plays a central role in bioenergetic regulation.13
These differences could alter nuclear interactions that dependor are independent of Cre recombinase. Although TAMexposure at the same dose was not toxic, the MCM constructincreases nuclear levels �4-fold, which could amplify inter-actions. Although similar energy/metabolic changes oftenaccompany pathological cardiac stress,7,8 here, the triggeringmechanism involved altered nuclear signaling; therefore,these hibernation-like reversible changes may indeed beprimary. Further studies are needed to clarify this hypothesis.Lastly, it remains possible that differences in the time courseand/or nuclear Cre exposure between TAM and RAL playsome role, and the finding that lower prolonged dosing ofTAM was also effective without myopathy might suggestthis. However, this could also reflect less nuclear exposure toTAM-MCM.
Our results have implications for existing and ongoingresearch with the MerCreMer model. Studies lacking Myh6-MCM�/�-flox� controls should be viewed cautiously, partic-ularly if a significant cardiac phenotype is found within the 1
Figure 3. RAL-induced gene knockdowndoes not affect cardiac function. A,M-mode echocardiograms from MCM�/�/Alk5fl/fl treated with TAM (80 mg/kg perday for 7 days) or RAL (160 mg/kg perday for 21 days). B, Top, Fractional short-ening in mice with or without MCM trans-gene and TAM vs RAL dosing. Bottom,Gene knockdown with each treatment(n�4 to 6/group). *P�0.01 vs MCM�/�/Alk5fl/fl. C, RAL and TAM equally induceCre localization to myocyte nuclei. Confo-cal immunofluorescence of Cre in myo-cytes isolated from MCM�/�/flox�/� miceon finishing 7 days of TAM or 21 days ofRAL treatment, as in Figure 3A and 3B.Low-power (�100) and higher-power(�630) images are shown (Cre, Acn:sar-comeric �-actinin; DAPI nuclear staining).
Koitabashi et al Cardiomyopathy in MerCreMer Mouse 3
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from

to 2 weeks after starting TAM. In such models, gene deletionwithout cardiodepression (eg, using RAL or longer-termlow-dose TAM) is required. Both TAM and RAL dosing mayneed to be individualized depending on the floxed gene(perhaps related to gene accessibility and/or expression rate).For TAM, care to include MCM� controls and providesufficient recovery time is strongly advised. Although RALavoids the myopathy, the dose required was fairly high in thepresent floxed models (lower doses might work for othermodels). However, mice treated mice with �10� this dosefor up to 3 months had no systemic limiting effects.14 BecauseRAL may have antihypertrophic effects when given chroni-cally,15 MCM� controls are also advised.
Sources of FundingSupported by National Heart, Lung, and Blood Institute grantsHL-59480, HL-77180, and HL-98297 (to D.A.K.); an AmericanHeart Association Fellowship Award (to M.Z.); and an AmericanHeart Association Scientist Development Grant (to E.T.).
DisclosuresNone.
References1. Glaser S, Anastassiadis K, Stewart AF. Current issues in mouse genome
engineering. Nat Genet. 2005;37:1187–1193.2. O’Neal KR, Agah R. Conditional targeting: inducible deletion by Cre
recombinase. Methods Mol Biol. 2007;366:309–320.3. Sohal DS, Nghiem M, Crackower MA, Witt SA, Kimball TR, Tymitz
KM, Penninger JM, Molkentin JD. Temporally regulated and tissue-specific gene manipulations in the adult and embryonic heart using atamoxifen-inducible Cre protein. Circ Res. 2001;89:20–25.
4. Silver DP, Livingston DM. Self-excising retroviral vectors encoding theCre recombinase overcome Cre-mediated cellular toxicity. Mol Cell.2001;8:233–243.
5. Schmidt-Supprian M, Rajewsky K. Vagaries of conditional gene tar-geting. Nat Immunol. 2007;8:665–668.
6. Molkentin JD, Robbins J. With great power comes great responsibility:using mouse genetics to study cardiac hypertrophy and failure. J Mol CellCardiol. 2009;46:130–136.
7. Huss JM, Kelly DP. Nuclear receptor signaling and cardiac energetics.Circ Res. 2004;95:568–578.
8. Sihag S, Cresci S, Li AY, Sucharov CC, Lehman JJ. PGC-1alpha andERRalpha target gene downregulation is a signature of the failing humanheart. J Mol Cell Cardiol. 2009;46:201–212.
9. Greschik H, Flaig R, Renaud JP, Moras D. Structural basis for thedeactivation of the estrogen-related receptor gamma by diethylstilbestrolor 4-hydroxytamoxifen and determinants of selectivity. J Biol Chem.2004;279:33639–33646.
10. Shang Y, Brown M. Molecular determinants for the tissue specificity ofSERMs. Science. 2002;295:2465–2468.
11. Paulmurugan R, Gambhir SS. An intramolecular folding sensor forimaging estrogen receptor-ligand interactions. Proc Natl Acad Sci U S A.2006;103:15883–15888.
12. Kiermayer C, Conrad M, Schneider M, Schmidt J, Brielmeier M. Opti-mization of spatiotemporal gene inactivation in mouse heart by oralapplication of tamoxifen citrate. Genesis. 2007;45:11–16.
13. Dufour CR, Wilson BJ, Huss JM, Kelly DP, Alaynick WA, Downes M,Evans RM, Blanchette M, Giguere V. Genome-wide orchestration ofcardiac functions by the orphan nuclear receptors ERRalpha and gamma.Cell Metab. 2007;5:345–356.
14. Cohen IR, Sims ML, Robbins MR, Lakshmanan MC, Francis PC, LongGG. The reversible effects of raloxifene on luteinizing hormone levelsand ovarian morphology in mice. Reprod Toxicol. 2000;14:37–44.
15. Ogita H, Node K, Liao Y, Ishikura F, Beppu S, Asanuma H, Sanada S,Takashima S, Minamino T, Hori M, Kitakaze M. Raloxifene prevents cardiachypertrophy and dysfunction in pressure-overloaded mice. Hypertension.2004;43:237–242.
KEY WORDS: mouse � cardiac � conditional knockout � Cre recombinase� ventricular function � tamoxifen � raloxifene
4 Circulation Research July 2, 2009
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from

L. Gabrielson, Eiki Takimoto and David A. KassNorimichi Koitabashi, Djahida Bedja, Ari L. Zaiman, Yigal M. Pinto, Manling Zhang, Kathleen
MerCreMer Gene Deletion ModelsAvoidance of Transient Cardiomyopathy in Cardiomyocyte-Targeted Tamoxifen-Induced
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2009 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
published online June 11, 2009;Circ Res.
http://circres.ahajournals.org/content/early/2009/06/11/CIRCRESAHA.109.198416.citationWorld Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org/content/suppl/2009/06/15/CIRCRESAHA.109.198416.DC1Data Supplement (unedited) at:
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on July 1, 2018http://circres.ahajournals.org/
Dow
nloaded from

1
On-Line Supplements and Figures
Supplemental Materials and Methods:
Mice: Myh6-MerCreMer+/+ transgenic mice (B6/129-TG Myh5-cre/Esr1)1Jmk/J; #005650, Jackson
Labs, Bar Harbor, ME)) were used. Myh6-MCM+/-/no-flox were generated by mating these to
C57Bl/6 mice. TGFBR2fl/fl (gift of Hal Moses)1 and Alk5fl/fl (2) (both on C57Bl/6 background) were
mated to the Myh6-MCM+/- to derive Myh6-MCM+/-/tgfbr2fl/fl or Myh6-MCM+/-/alk5fl/fl, respectively.
Genotype was confirmed with primers to Cre and tgfbr2 or alk5 floxed alleles.
Tamoxifen/Raloxifene administration: For IP injection, 10mg tamoxifen free base (T5648, Sigma)
was dissolved in 1mL peanut oil (Sigma) and then sonicated. 50uL (0.5mg tamoxifen) was injected
daily (20mg/kg/day, i.p.). Raloxifene was dissolved in warmed dimethyl sulfoxide (50mg/mL),
then diluted ten-fold (40mg/kg/day, i.p.). For oral dosing, drugs were mixed in soft diet (Bioserve,
MA), with daily intake ~4g/day (e.g. 500mg/kg tamoxifen-food = 80mg/kg/day, 1000 mg/kg
raloxifene-food = 160mg/kg/day; 25 gm mouse).
Western blot/Smad2 phosphorylation analysis Isolated myocytes suspended in Tyrode’s
solution were aliquoted and a sub-fraction stimulated by recombinant human TGFβ1(Sigma). After
30minutes incubation with gentle rotation at room temperature, cells were centrifuged, the pellet
lysed (Cell Signaling Technology), and lysates subjected to Western blot using a NuPage system
(Invitrogen) and probed with Phospho Smad2/3 antibody (Cell Signaling). Membranes were then
reprobed with total Smad2 antibody or GAPDH (Cell Signaling). Band intensity was quantified by
NIH Image J software. For Cre or MerCreMer fusion protein detection, we used rabbit polyclonal

2
anti-Cre antibody (Novagen).
RNA analysis Total RNA was prepared using TRIzol reagent (Invitrogen) from left ventricular
tissue or isolated myocytes. Messenger RNA was analyzed by quantitative real-time polymerase
chain reaction (PCR) using either SYBR green or Taqman probe method3. Complementary DNAs
(cDNAs) were synthesized by standard techniques by using the TaqMan Reverse Transcription
Reagent (Applied Biosystems) with random hexamer primers. Real-time PCR reactions were
performed, recorded, and analyzed using the ABI PRISM 7900 (Applied Biosystems, Foster City,
California). Tgfbr2 and Alk5 were assessed using each targeted exon specific primer-probe from
Applied Biosystems: Tgfbr2, Mm00436973_m1; Alk5, Mm01353995_m1, and normalized with
Gapdh, Mm99999915_g1. For tissue, the Nppa (atrial natriuretic peptide: ANF), Nppb (B-type
natriuretic peptide: BNP), Myh7 (beta myosin heavy chain: βMHC), Atp2a2 (Sarcoplasmic
reticulum calcium ATPase 2a: SERCA2a), Pln (phospholamban) and gapdh mRNA levels were
quantified by using SYBR green PCR primers or TaqMan primer-probe method as previously
described4. Following primers were also used for SYBR gree real-timer RT-PCR; Ppargc1a
(peroxisome proliferative activated receptor gamma coactivator 1 alpha: PGC1α), Forward: 5’-
ATAGAGTGTGCTGCTCTGGTTGGT-3’, Reverse: 5’-TGGTCGCTACACCACTTCAATCCA-
3’; Ppara (peroxisome proliferator activated receptor alpha: PPARα), Forward: 5’-
GAAACGGAAACGGCCTCATGTGTT -3’, Reverse: 5’-AGAGTACAATCGCTTGCTGTCCCA-
3’; Tfam(mitochondrial transcription factor A), Forward: 5’-
TGAAAGCTTCCAGGAGGCAAAGGA-3’, Reverse: 5’-ACTTCAGCCATCTGCTCTTCCCAA-
3’. The specificity of the SYBR green assays was confirmed by dissociation curve analysis.
Genomic DNA analysis in cardiomyocytes Genomic DNA was extracted from isolated myocytes
using DNeasy tissue kit (Qiagen) according to the manufacturer’s protocol. Genotyping PCR for

3
alk5 excised allele was performed with the triple PCR method using primers as follows; ALK5-
flox:Prim1, fw ACC CTC TCA CTC TTC CTG AGT; ALK5-flox:Prim2, Rev ATG AGT TAT
TAG AAG TTG TTT; ALK5-flox:Prim3, Rev GGA ACT GGG AAA GGA GAT AAC. This PCR
gives a band(s) for each genotype as follows; wild type, 150bp; alk5+/fl, 150+250bp; alk5fl/fl,
250bp; Cre excised allele, 350bp2.
Immunostaining and confocal microscopy Cardiac myocytes were isolated and stained with anti-
Cre antibody (1:1000, Novagen) and anti-sarcomeric α actinin antibody (1:500, Sigma). Confocal
microscopic analysis was performed as previously described3 (RGS paper).
Nuclear protein extraction Nuclear protein was fractionated by NE-PER Nuclear and Cytoplasmic
Extraction Reagent (Pierce) according to the manufacturer’s protocol. Anti-Histon H3 antibody
(Cell Signaling) was used as a nuclear extract marker.
Echocardiography In vivo cardiac morphology was assessed by transthoracic echocardiography
(Acuson Sequoia C256, 13MHz transducer; Siemens) in conscious mice. M-mode LV end-systolic
and end-diastolic dimensions were averaged from 3-5 beats. LV percent FS and LV mass were
calculated as described previously3;5. Studies and analysis were performed by investigators blinded
to genotype or treatments.
In vivo hemodynamics In vivo LV function was assessed by PV catheter as described
previously3;6. Briefly, mice were anesthetized with 1%–2% isoflurane, 750–100 mg/kg urethane i.p.,
5–10 mg/kg etomidate i.p., and 1–2 mg/kg morphine i.p.; were subjected to tracheostomy; and were
ventilated with 6–7 μl/g tidal volume and 130 breaths/min. Volume expansion (12.5% human
albumin, 50–100 μl over 5 min) was provided through a 30-gauge cannula via the right external
jugular vein. The LV apex was exposed through an incision between the seventh and eighth ribs,

4
and a 1.4-Fr PV catheter (SPR 839; Millar Instruments Inc.) was advanced through the apex to lie
along the longitudinal axis. Absolute volume was calibrated, and PV data were measured at steady
state and during transient reduction of venous return by occluding the inferior vena cava with a 6-0
silk snare suture. Data were digitized at 2 kHz, stored to disk, and analyzed with custom software.
From the 10–15 successive cardiac cycles during the inferior vena cava occlusion, the end-systolic
PV relation slope (i.e., end-systolic elastance) and stroke work–end-diastolic volume relation (i.e.,
preload recruitable stroke work were derived.
Statistics All values are expressed as mean ± SEM. Group comparisons were performed by 1-way
ANOVA or non-paired 2-tailed Student’s t test. Sample sizes and individual statistical results for
all analyses are provided in the figures, supplemental figures, and supplemental tables.

5
References
1. Chytil A, Magnuson MA, Wright CV, Moses HL. Conditional inactivation of the TGF-beta type II receptor using Cre:Lox. Genesis. 2002;32:73-75.
2. Larsson J, Goumans MJ, Sjostrand LJ, van Rooijen MA, Ward D, Leveen P, Xu X, ten Dijke P, Mummery CL, Karlsson S. Abnormal angiogenesis but intact hematopoietic potential in TGF-beta type I receptor-deficient mice. EMBO J. 2001;20:1663-1673.
3. Takimoto E, Koitabashi N, Hsu S, Ketner EA, Zhang M, Nagayama T, Bedja D, Gabrielson KL, Blanton R, Siderovski DP, Mendelsohn ME, Kass DA. Regulator of G protein signaling 2 mediates cardiac compensation to pressure overload and antihypertrophic effects of PDE5 inhibition in mice. J Clin Invest. 2009;119:408-420.
4. Tsujita Y, Muraski J, Shiraishi I, Kato T, Kajstura J, Anversa P, Sussman MA. Nuclear targeting of Akt antagonizes aspects of cardiomyocyte hypertrophy. Proc Natl Acad Sci U S A. 2006;103:11946-11951.
5. Takimoto E, Champion HC, Belardi D, Moslehi J, Mongillo M, Mergia E, Montrose DC, Isoda T, Aufiero K, Zaccolo M, Dostmann WR, Smith CJ, Kass DA. cGMP catabolism by phosphodiesterase 5A regulates cardiac adrenergic stimulation by NOS3-dependent mechanism. Circ Res. 2005;96:100-109.
6. Pacher P, Nagayama T, Mukhopadhyay P, Batkai S, Kass DA. Measurement of cardiac function using pressure-volume conductance catheter technique in mice and rats. Nat Protoc. 2008;3:1422-1434.

Alk5/Gapdh
Fold
incr
ease
MCM+/-/Alk5fl/fl
MCM-/-/Alk5fl/fl
0
0.4
0.8
1.2
*
20mg/kgi.p. 5d
80mg/kgp.o. 7d
80mg/kgp.o. 7d
%
0
100
200
%Increase of pSmad2 by rhTGFβ
*
*MCM+/-/Tgfbr2fl/fl
MCM-/-/Tgfbr2fl/fl
20mg/kgi.p. 5d
80mg/kgp.o. 7d
80mg/kgp.o. 7d
A B
Online Figure I

0
20
40
60
80
100
MCM+/-/flox-/- (N=9)MCM-/-/flox-/- (N=5)
TAM 80mg/kg IP/5days
Sur
viva
l (%
)
Online Figure II
DeathFS 61.5%EDD 3.4mm
FS 40.3%EDD 3.4mm
FS 13.7%EDD 5.1mm
Baseline Day3 Day5 Day8A
B
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Days
Online Figure III
0
20
40
60
Baseline Day10 Day14 Day28
FS(%
)
*
*
2
3
4
5
MCM+/-/ALK5fl/fl
MCM-/-/ALK5fl/fl
Baseline Day10 Day14 Day28
ED
D (m
m) *
A BBaseline Day10 Day28
MCM flox
+/- -/-
+/- fl/fl
-/-fl/fl
Tgfbr2
+/- fl/fl
-/-fl/fl
Alk5

X200; Scale bar 50µm
MCM(-/-) / flox-/-
Day10 Day28
MCM(+/-) / flox-/-
Day10
All mice treated with tamoxifen 80 mg/kg/day P.O. for 7 days.
Online Figure IV

2
2.5
3
3.5
4
4.5
Baseline Day10 Day28E
DD
(mm
)0
10
20
30
40
50
60
70
Baseline Day10 Day28
RAL160mg/kg-21d p.o.TAM20mg/kg-21d p.o.
RAL320mg/kg-7d p.o.
FS(%
)
Online Figure V

MCM+/-/flox-/-MCM-/-/flox-/-Control Lysate
100kD
35
MerCreMer
Cre
A
IB:Histone H3
IB:GAPDH
IB:Cre
IB:GAPDH
IB:Cre
B MCM+/-/flox-/-
MCM-/-/flox-/-
Drug (-) TAM RAL
Online Figure VI

Drug(-)
TAM 80mg/kg
p.o.7d
500400
300
200
Cre excised allele
Recombinant allele
500400
300
600
bp
MerCreMerControl
exon IIIneo
exon IV
Primer1 Primer2 Primer3
Cre recombinase250bp
exon IV
350bp
LoxP
A
B
MCM-/-/Alk5fl/fl
MCM+/-/Alk5fl/fl
RAL160mg/kg
p.o.7d
TAM20mg/kgp.o.21d
RAL160mg/kg
p.o.21d
Online Figure VII

MCM+/-/Alk5fl/fl
MCM-/-/Alk5fl/fl
FS (%
)0
20
40
60
*
TAM80mg/kgp.o 7d
TAM20mg/kgp.o. 21d
RAL160mg/kgp.o. 21d
Alk
5/G
apdh
Nor
mal
ized
to M
CM
-/-
0
0.4
0.8
1.2
* **
Online Figure VIII

HR (/min)
ESP (mmHg)
EDP (mmHg)
CO (mL/min)
EF (%)
Ea (mmHg/uL)
dP/dtmax (mmHg/sec)
Parameter
Systolic indeces
PWRMax/EDV (mmHg/uL)
Diastolic indeces
-dP/dtmin (mmHg/sec)
Tau (S) (msec)
PFR/EDV (/sec)
MCM(-)+TAMDay10N=3
MCM+TAMDay10N=4
424+37458+35
85.1+3.2 75.7+3.4
7.5+2.3 10.5+2.0
7.4+1.7 5.4+0.8
6.7+1.5 5.3+1.1
21.7+3.8*60.0+8.8
7711+1144 6467+543
6000+510 3477+366 §
28.7+9.9 6.6+1.1*
6.4+0.4 9.1+0.7*
19.9+4.0 9.8+0.8*
Supplemental Table: Hemodynamic parameters and indices derived from PV loop analysis
HR, heart rate; ESP, end-systolic pressure; EDP, end-diastolic pressure; EDV, end-diastolic volume; ESV, end-systolic volume; CO, cardiac output; Ea, arterial elastance; EF, ejection fraction; dPdtmax, peak rate of pressure rise; PWRmax/EDV, peak power index; -dP/dtmin, peak rate of pressure decline; Tau (S), relaxation time constant calculated by Suga method; PER/EDV, peak filling rate/end-diastolic volume, * p<0.05 vs other two groups, †p<0.005 vs other two groups, ‡ p<0.0o5 vs MCM+TAM; §p=0.06 vs MCM(-) +TAM; 1-way ANOVA; tukey test for multiple comparisons.
EDV (μL)
ESV (μL)
31.0+7.5 57.0+5.6*
13.5+5.0 44.5+4.7†
574+18 *
92.1+6.0
4.8+2.0
11.0+0.7‡
5.0+0.3
70.0+8.1
11033+425 *
9152+890*
41.1+11.3
4.3+0.4*
34.3+4.6*
27.4+3.6
8.9+3.3
MCM+TAMDay28N=3
14

15
Online Figure Legends:
Online Figure I. A) Gene expression changes in Alk5 in MCM+/-Alk5fl/fl mice treated with
tamoxifen TAM) at two dosing regimens. Data are as displayed in Fig 1A; normalized to Gapdh and
to results in MCM-/- controls. Only the higher TAM dose yielded effective gene knockdown. This
result was similar to that in the other flox’d model shown in Fig 1A,B. B) Summary data for Smad
phosphorylation assay results shown in Fig 1B. (n=4/group, *p<0.05 vs MCM-/-).
Online Figure II A, An example of tamoxifen-mediated cardiomyopathy in MCM+/-/Tgfbr2fl/fl
mouse. This mouse was administered TAM 80mg/kg i.p. injection for only 2 days and then followed
by echocardiography. Marked cardiac dysfunction developed and the animal succumbed at day 8. B,
Kaplan-Meier analysis of MCM-/-/flox-/- versus MCM+/-/flox-/- mice after receiving the same i.p. dose
of tamoxifen for only 2 days.
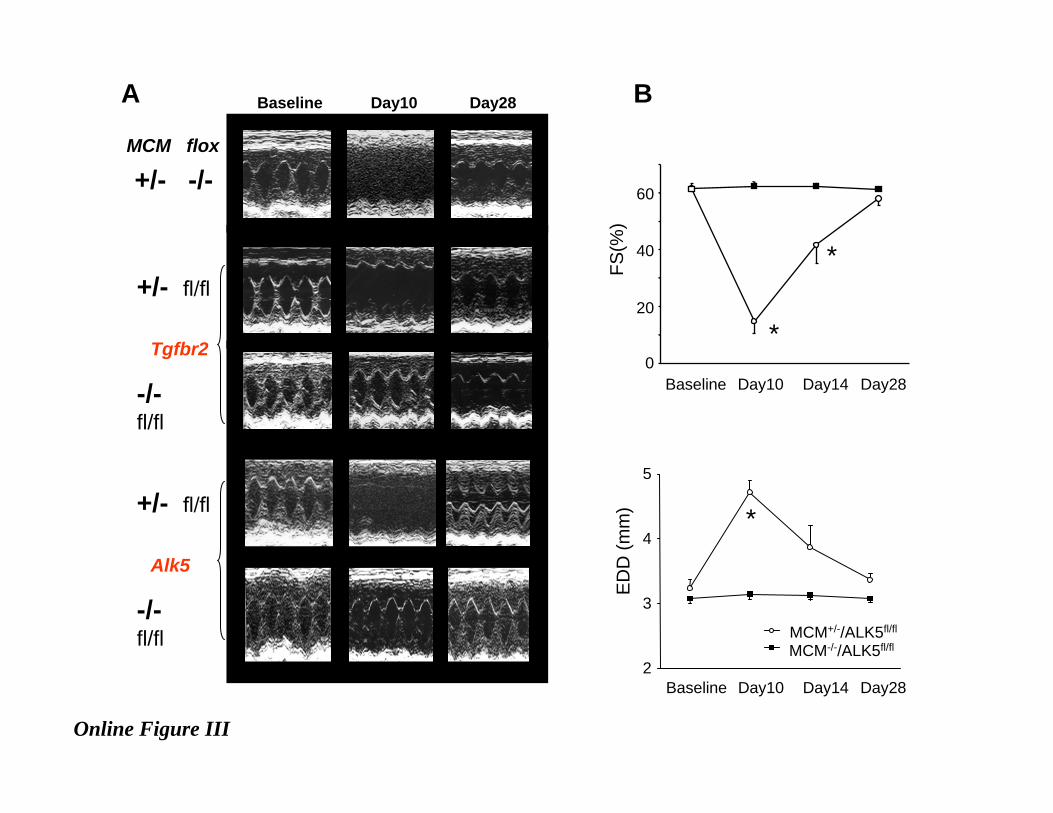
Online Figure III . A, Representative M-mode echocardiographic images showing the temporal
changes of cardiac function before and after oral tamoxifen treatment (80mg/kgBW/day for 7days).
Results are shown at baseline, day 10 (3-days after terminating tamoxifen treatment; and peak of
cardiac depression), and day 28 (21 days after terminating tamoxifen). Reversible dilated
cardiomyopathy was observed in all models that included the heterozygote merCremer (MCM)
transgene, with or without a flox’d gene target. MCM-negative controls did not develop dysfunction.,
(B) Summary data for fractional shortening (FS) and left ventricular end-diastolic dimension (EDD)
before and after tamoxifen administration; additional time point at day 14 is shown. (N=5 of each
group. *P<0.05 vs MCM-/-/Alk5fl/fl.

16
Online Figure IV
Hematoxylin/eosin stained myocardium from mice treated with oral tamoxifen (80 mg/kg/day for 7
days). All mice received this treatment. Mice lacking the MCM transgene show normal histology.
Those with MCM+/- (no flox’d gene) show patchy inflammation with mononuclear infiltrates at day
10 (3 days after completing tamoxifen) – middle pane - , and this essentially resolved with little
residual damage when examined at day 28 (same time when functional recovery was documented).
These results were typical of samples from 3-4 separate hearts for each condition.
Online Figure V
Temporal changes in fractional shortening (FS) and left ventricular end-diastolic dimension (EDD)
during raloxifene (RAL) or tamoxifen (TAM) treatment in MCM+/-/Alk5fl/fl mice. RAL
320mg/kgBW-7d p.o for 7days treatment; RAL160mg/kgBW-21d p.o.; TAM 20mg/kg-21d p.o for
21days. N=5 of each group. None of these dosing regimens induced changes in ventricular function.
Online Figure VI A) Western blot for Cre showing constitutive expression of MerCreMer
expression in cardiac myocardium (whole tissue lysates) in MCM+/- but not MCM-/- left ventricles.
The ~100kDa band represents the MerCreMer fusion protein. Cell lysate of COS7 cells transfected
with Cre-adenovirus (Cell Biolabs, Inc) is used as a control. GAPDH is shown as a loading control.
B) Western blot of nuclear extract from isolated cardiomyocytes in MCM-/-/flox-/- or MCM+/-/flox-/-
mice immediately after completing 7day TAM (80mg/kg/d) or 21day RAL (160mg/kg/d) treatments.
Nuclear localization of Cre was detected similarly with both treatments (top bands). Histone H3 is
used as a marker for nuclear fraction, and GAPDH as a negative control (used for cytosolic fraction).
Online Figure VII Recombination of the targeted allele in MCM+/-/alk5fl/fl mouse. A) A scheme
showing Alk5 recombinant allele (upper) and Cre excised allele (lower). PCR primers are indicated

17
by small arrow heads B) Genotyping by PCR of genomic DNA extracted from isolated
cardiomyocytes. Upper panel shows the triple primer PCR for detecting the Cre excised allele using
the primers shown in A. Lower panel shows genotyping for MerCreMer. Seven-day raloxifene
treatment showed some induction of the Cre-excised allele, but longer exposure was required to
achieve levels similar to TAM. Part of this may relate to a fall in food intact with RAL-mixed food
during the initial week (possibly related to taste) which rose after that.
Online Figure VIII
Smad2 phosphorylation by rhTGFβ: 5ng/mL, 30min stimulation in cardiomyocytes from MCM-
/+/Alk5fl/fl treated with 21d RAL. Representative immunoblot (left) and summary data for %rise in
pSmad2 (right) are shown (n=4 for each group, †p<0.01 vs No Tx group).
Online Figure IX
Summary data for fractional shortening (FS) and Alk5 gene expression (normalized to GAPDH, and
then to MCM(-/-) data) showing efficacy of low dose sustained tamoxifen (20 mg/kg/d for 21 days)
without significant cardiac changes. There was somewhat greater variance in the cardiac response to
TAM versus 21d raloxifene (RAL) treatment, however. Data not presented in Fig 3B is denoted by
the dashed box.