reguladores de la supervivencia celular en … · pacientes con síndrome de ovario poliquístico...
TRANSCRIPT

UNIVERSIDAD DE CHILE
FACULTAD DE MEDICINA
ESCUELA DE POSTGRADO
REGULADORES DE LA SUPERVIVENCIA CELULAR EN ENDOMETRIOS OBTENIDOS DE PACIENTES CON SINDROME DE OVARIO POLIQUISTICO. POTENCIALES CONSECUENCIAS EN EL
DESARROLLO DE HIPERPLASIA ENDOMETRIAL
ALEJANDRA MARIELA VILLAVICENCIO GALDEANO
TESIS PARA OPTAR AL GRADO DE DOCTOR EN CIENCIAS BIOMEDICAS
Director de Tesis: Prof. Dra. Margarita Vega Blanco
2005

UNIVERSIDAD DE CHILE FACULTAD DE MEDICINA
ESCUELA DE POSTGRADO \
INFORME DE APROBACION TESIS DE
DOCTORADO EN CIENCIAS BIOMEDICAS
Se informa a la Comisión de Grados Académicos de la Facultad de Medicina, que la Tesis de Doctorado en Ciencias Biomédicas presentada por el candidato
ALEJANDRA MARIELA VILLAVICENCIO GALDEANO
ha sido aprobada por la Comisión Informante de Tesis como requisito para optar al Grado de Doctor en Ciencias Biomédicas en Examen de Defensa de Tesis rendido el día15 de Noviembre de 2005.
Prof. Dra. Margarita Vega Blanco Director de Tesis
Instituto de Investigaciones Materno Infantil, Facultad de Medicina, Universidad de Chile
COMISION INFORMANTE DE TESIS
PROF. DR. HERNAN LARA PROF. DR. JORGE MARTINEZ
PROF. DR. CLAUDIO LIBERMAN PROF. DR. ENRIQUE CASTELLON Presidente Comisión de Examen

Se quiere más aquello que se ha conseguido con mucho esfuerzo
Aristóteles

A mis padres Oscar y María del Carmen por la vida
y porque nada de esto hubiese sido posible
sin su inmenso amor y apoyo

A mis hermanas Noelia y María Verónica
por haber sido luz en este camino
y a quienes pido sigan acompañándome siempre

Indice
INDICE Página
i AGRADECIMIENTOS………………………………………………………...……….. i
ii FINANCIAMIENTO……………………………………………………………...……..iv
iii PUBLICACIONES Y PRESENTACIONES A CONGRESOS ORIGINADAS DE
ESTA TESIS ……………..……………………………………………..………………….. vi
iv RESUMEN…………………………………………………………………………….....x
v ABSTRACT…………………………………………………………………………….xv
I. INTRODUCCION .................................................................................................. 1
I.1 El Endometrio Humano .................................................................................. 1
I.2 Síndrome de Ovario Poliquístico (SOP) ......................................................... 3
I.3 Hormonas Ováricas ....................................................................................... 6
I.4 Receptores Esteroidales ................................................................................ 6
I.4.1 Receptores endometriales de estrógenos .................................................. 7
I.4.2 Receptores de progesterona en endometrio .............................................. 8
I.4.3 Receptor de andrógenos endometrial ........................................................ 9
I.5 Co-Reguladores y su Acción en Endometrio Humano ................................ 10
I.6 Homeostasis Endometrial ............................................................................ 12
I.6.1 Apoptosis .................................................................................................. 12

Indice
I.6.2 Proliferación .............................................................................................. 15
I.7 Reguladores del Ciclo Celular ...................................................................... 20
I.7.1 Regulación del ciclo celular por estrógenos ............................................. 20
I.7.2 Otros reguladores del ciclo celular ........................................................... 20
II. HIPOTESIS ......................................................................................................... 24
III. OBJETIVOS GENERALES ............................................................................. 24
IV. OBJETIVOS ESPECIFICOS ........................................................................... 25
V. MATERIALES Y METODOS .............................................................................. 27
V.1 Obtención de las Muestras .......................................................................... 27
V.1.1 Controles Sanos (EN) ........................................................................... 27
V.1.2 Pacientes SOP (ESOP) ........................................................................ 27
V.1.3 Pacientes SOP con Hiperplasia Endometrial (ESOPH) ........................ 28
V.1.4 Pacientes no SOP con Hiperplasia Endometrial (EH) ........................... 29
V.2 Estudios Ex Vivo .......................................................................................... 29
V.3 Estudios In Vitro ........................................................................................... 30
V.4 Diseño Experimental .................................................................................... 30
V.5 Cultivos de Explantes de Tejidos ................................................................. 30
V.6 Análisis Morfológico ..................................................................................... 31

Indice
V.7 Estudios de Expresión Génica ..................................................................... 32
V.7.1 Reacción de polimerización en cadena de DNA complementario (RT-
PCR)…………………………………………………………………………………….32
V.8 Estudios de Expresión Proteica ................................................................... 33
V.8.1 SDS-PAGE y Análisis de Western Blot ................................................. 33
V.8.2 Estudio inmunohistoquímico ................................................................. 36
V.9 Estudio de Actividad de Cdk-2 ..................................................................... 38
V.10 Estudios de Apoptosis .............................................................................. 39
V.10.1 Método fluorescente de detección de apoptosis (TUNEL) .................... 39
V.11 Estudios de Proliferación .......................................................................... 39
V.11.1 Expresión de la proteína Ki67 ............................................................... 39
V.12 Análisis Estadístico ................................................................................... 40
VI. RESULTADOS ................................................................................................ 41
Vi.1 Características de las Pacientes Estudiadas ............................................... 41
VI.2 Estudios Ex Vivo .......................................................................................... 42
VI.2.1 Estudio histológico de los endometrios de las pacientes estudiadas .... 42
VI.2.2 Expresion génica y proteica de los receptores esteroidales en el
endometrio de las mujeres en estudio ................................................................. 44
VI.2.3 Expresión proteica del co-activador AIB1 ............................................. 50

Indice
VI.2.4 Expresión de la enzima P450 aromatasa ............................................... 51
VI.2.5 Estudios de Proliferación ...................................................................... 52
VI.2.6 Estudios de Apoptosis .......................................................................... 54
VI.2.7 Estudio de la onco-proteína Ras ........................................................... 58
VI.2.8 Estudio de la proteína supresora de tumores PTEN ............................. 61
VI.2.9 Expresión y actividad de Akt ................................................................. 64
VI.2.10 Reguladores del ciclo celular ............................................................. 66
Vi.3 Estudios In Vitro ........................................................................................... 78
VI.3.1 Evaluación morfológica del tejido endometrial posterior al cultivo ........ 78
VI.3.2 Acción in vitro de concentraciones elevadas de androstenediona,
estradiol y/o insulina sobre la expresión del co-activador AIB1 .......................... 78
VI.3.3 Acción in vitro de concentraciones elevadas de androstenediona,
estradiol y/o insulina sobre la expresión de reguladores del ciclo celular .......... 80
VII. DISCUSION ..................................................................................................... 82
VIII. CONCLUSIONES .......................................................................................... 101
IX. PROYECCIONES .......................................................................................... 103
X. ANEXOS ........................................................................................................... 104
X.1 Consentimiento Informado para la Obtención de Muestras de Tejido
Endometrial ........................................................................................................... 104

Indice
XI. BIBLIOGRAFIA ............................................................................................. 106

Agradecimientos
i
i. AGRADECIMIENTOS
A Dios y su Madre, por acompañarme siempre y por haberme permitido alcanzar
este logro.
A la Dra. Margarita Vega por su cariño, confianza, y apoyo. Por haberme
incentivado y ayudado a crecer científicamente, y por haber estado conmigo de una
forma tan especial en todo este tiempo. De todo corazón, muchas gracias Margarita
por tanto.
A Carla Avellaira y Ketty Bacallao, por tener siempre una sonrisa y una palabra de
aliento, por ser excelentes compañeras de trabajo, y sobretodo, por ser grandes
amigas dentro y fuera del laboratorio.
Con especial cariño quiero agradecer a Dora Arrieta, por su gran amor e
incondicionalidad.
A Alfredo Mac-Mahon, gracias por haber estado conmigo en esta etapa,
acompañándome y ayudándome en los momentos más difíciles. Pero principalmente
gracias por entenderme y amarme.

Agradecimientos
ii
Agradezco a todos mis amigos por su confianza, paciencia, y compañía a lo largo de
este camino.
Quiero agradecer al IDIMI y a cada una de las personas que lo integran o integraron.
Gracias por haber sido mi segunda casa en Chile y por el aporte que me brindaron.
En forma especial agradezco a Ingrid Espinoza, Alejandra Estévez, Jaime Albornoz
Fernando Gabler, Ariel Fuentes, Rosita Smith, Mabel Anido, María Cecilia Johnson,
Angélica Boric, Simón Urquiaga, Cita Lorca, Luigi Devoto, Olguita Castro, Germán
Iñiguez, y Andrea Castro por su ayuda a lo largo de todos estos años.
A todas las mujeres que generosamente donaron parte de su endometrio para la
realización de este estudio, en forma especial a Flor González y a mamá.
Al Dr. Ricardo Deis y a todos los miembros del IMBECU, por haber confiado en mí.
A Carlos Tellería y Alejandra Goyeneche por su generosidad y valioso apoyo
durante mi estadía en su laboratorio.
A CONICET, PROGRESAR y al Centro FONDAP de Estudios Moleculares de la
Célula, por haberme financiado durante el desarrollo de mi doctorado.

Agradecimientos
iii
A los miembros de la comisión examinadora por todos sus consejos, comentarios
y sugerencias.
Agradezco al Dr. Jorge Las Heras por su ayuda desinteresada.
Pido disculpas si no he sido justa en los agradecimientos o si olvidé a alguien. Sin
embargo, creo que las palabras no alcanzan para expresar las infinitas gracias que
debo a todas las personas que me ayudaron.
Finalmente quiero agradecer a todas las personas que me regalaron una sonrisa
cuando la necesité pues eso me dio alegría y fuerza; también a todos los que me
dieron un motivo para estar triste, porque me permitieron ser más fuerte, luchar por
salir adelante y crecer.

Financiamiento
iv
ii. FINANCIAMIENTO
Esta tesis de doctorado se realizó en:
- Instituto de Investigaciones Materno Infantil, Facultad de Medicina (Campus
Centro), Universidad de Chile, Hospital Clínico San Borja Arriarán.
- Division of Basic Biomedical Sciences, School of Medicine, University of South
Dakota, USA.
Fue financiada por los siguientes Proyectos y Becas:
Proyectos:
- Proyecto FONDECYT 1010821/1050098
Investigador Responsable: Dra. Margarita Vega Blanco
- RE-ENTRY GRANT PLACIRH: PRE-070/2003
Investigador Responsable: Alejandra M. Villavicencio Galdeano
- NIH GRANT 2 P 20 RR016479
Investigador Responsable: Dr. Carlos Telleria
Becas:
- Beca Doctoral Mixta del Consejo Nacional de Investigaciones Científicas y
Tecnológicas (CONICET, Argentina).
- Beca Programa Regional de Capacitación en Salud Sexual y Reproductiva
para Latinoamérica y el Caribe (PROGRESAR, Chile).

Financiamiento
v
- Beca del Centro FONDAP de Estudios Moleculares de la Célula (Chile).
- Beca de Estadía Corta en el Extranjero del Proyecto MECESUP UCH 0306
(Chile).
- Beca de Arancel, Facultad de Medicina, Universidad de Chile.

Publicaciones y Presentaciones a Congresos originadas de esta Tesis
vi
iii. PUBLICACIONES Y PRESENTACIONES A CONGRESOS ORIGINADAS DE ESTA TESIS
PUBLICACIONES
A. Villavicencio, K. Bacallao, C. Avellaira, F. Gabler, A. Fuentes, M. Vega. Steroid
receptors and co-regulators levels in endometria from patients with polycystic ovarian
syndrome with and without endometrial hyperplasia. Gynecologic Oncology. (Enviado
y en segunda revisión para su publicación)
A. Villavicencio, K. Bacallao, F. Gabler, A. Fuentes, M. Vega. Deregulation of
endometrial homeostasis in patients with polycystic ovarian syndrome with and
without endometrial hyperplasia. European Journal of Cancer. (Enviado)
A. Villavicencio, A. Goyeneche, C. Telleria, K. Bacallao, F. Gabler, A. Fuentes, M.
Vega. Involvement of Akt, Ras and cell cycle regulators in the potential development
of endometrial hyperplasia in women with polycystic ovarian syndrome. Endocrine-
Related Cancer. (En preparación para su publicación)
PRESENTACIONES A CONGRESOS
Nacionales:
A. Villavicencio, F. Gabler, A. Estévez, V. Garcia, T. Sir-Petermann, E. Soto, M.
Vega. Expresión de receptores esteroidales y coactivadores en endometrios de
pacientes con Síndrome de Ovario Poliquístico con y sin hiperplasia endometrial. XIV
Reunión Anual de la Sociedad Chilena de Reproducción y Desarrollo, La Serena,
Chile. Agosto del 2003.

Publicaciones y Presentaciones a Congresos originadas de esta Tesis
vii
S. Quezada, M. Anido, A. Villavicencio, M.C. Johnson, V.Garcia, A. Palomino, F.
Gabler, M. Vega. Acción del hiperandrogensimo sobre la expresión de receptores
esteroidales en tejido endometrial humano. XIV Reunión Anual de la Sociedad
Chilena de Reproducción y Desarrollo, La Serena, Chile. Agosto del 2003.
A. Villavicencio, F. Gabler, A. Fuentes, M. Vega. Reguladores de la supervivencia
celular en endometrios de pacientes con Síndrome de Ovario Poliquístico. XV
Reunión Anual de la Sociedad Chilena de Reproducción y Desarrollo, El Quisco,
Chile. Agosto del 2004. PREMIO SCHRyD 2004
A. Villavicencio, A. Goyeneche, C. Telleria, K. Bacallao, F. Gabler, P. Wells, J.
Albornoz, M. Vega. Proteínas involucradas en el aumento de la supervivencia celular
en endometrios de pacientes con Síndrome de Ovario Poliquístico. XVI Reunión
Anual de la Sociedad Chilena de Reproducción y Desarrollo, Reñaca, Chile. Agosto
del 2005.
C. Avellaira, K. Bacallao, A. Villavicencio, F. Gabler, A. Fuentes, M. Anido, M. Vega.
Receptor de andrógenos y su regulación en endometrios de mujeres con Síndrome
de Ovario Poliquístico. XVI Reunión Anual de la Sociedad Chilena de Reproducción y
Desarrollo, Reñaca, Chile. Agosto del 2005.
K. Bacallao, F. Gabler, A. Villavicencio, T. Sir-Petermann, V. García, M. Anido, M.
Vega. Cambios en la expresión y actividad de reguladores del ciclo celular en
endometrios de pacientes con Síndrome de Ovario Poliquístico. XVI Reunión Anual
de la Sociedad Chilena de Reproducción y Desarrollo, Reñaca, Chile. Agosto del
2005.

Publicaciones y Presentaciones a Congresos originadas de esta Tesis
viii
Internacionales:
A. Villavicencio, F. Gabler, M. Anido, M. Vega. Expresión de RAS y PTEN en
endometrios de pacientes con Síndrome de Ovario Poliquístico con y sin hiperplasia
endometrial. XVIII Reunión Bienal de la Asociación Latinoamericana de
Investigadores en Reproducción Humana, Varadero, Cuba. Mayo del 2003.
A. Villavicencio, F. Gabler, A. Casals, A. Palomino, M. Vega. Receptores
esteroidales y coactivador AIB1 en endometrios de pacientes con Síndrome de
Ovario Poliquístico con y sin hiperplasia endometrial”. 12th Internacional Congress of
Endocrinology, Lisboa, Portugal. Septiembre del 2004.
A. Villavicencio, F. Gabler, A. Fuentes, M. Vega. Reguladores de la supervivencia
celular en endometrios de pacientes con Síndrome de Ovario Poliquístico”. 12th World
Congress on Human Reproduction, Venecia, Italia. Marzo del 2005.
K. Bacallao, C. Avellaira, A. Villavicencio, M. Anido, A. Fuentes, E. Soto, F. Gabler,
M. Vega. Determinación de la supervivencia celular en endometrios de pacientes con
Síndrome de Ovario Poliquístico sin y con hiperplasia endometrial. XIX Reunión
Bienal de la Asociación Latinoamericana de Investigadores en Reproducción
Humana, Cartagena de Indias, Colombia. Mayo del 2005.

ix

Resúmen
x
iv. RESUMEN
El endometrio es uno de los tejidos del organismo cuya función es regulada de
manera cíclica por los esteroides de origen ovárico. El estradiol producido en la fase
folicular del ciclo menstrual induce proliferación de las células endometriales, y la
progesterona producida en la fase lútea promueve la diferenciación de dichas
células. Para ello, las células endometriales expresan distintas isoformas de
receptores esteroidales. También, la acción esteroidal a través de los receptores
puede estar controlada por co-reguladores, tales como, co-activadores o co-
represores.
Se ha reportado que estradiol regularía la expresión de varias moléculas
involucradas en el control del ciclo celular, tanto en endometrio como en otros tejidos
y diversas líneas celulares. Además, es ampliamente conocida la acción mitogénica
de insulina en endometrio; no obstante, la acción proliferativa de los andrógenos no
está aún claramente definida. Lo anterior podría explicar, en parte, el mayor riesgo
relativo que presentan las pacientes con Síndrome de Ovario Poliquístico (SOP) de
desarrollar hiperplasia y carcinoma endometrial, ya que es una patología
caracterizada por alteraciones endocrino-metabólicas.
En consecuencia, el propósito de esta tesis fue evaluar en endometrios de mujeres
SOP si la expresión de receptores esteroidales y moléculas reguladoras del ciclo
celular, así como el grado de proliferación y apoptosis, se relaciona con el desarrollo
de hiperplasia endometrial. Además, determinar in vitro si la condición
hiperandrogénica y/o hiperestrogénica y/o hiperinsulínica, contribuyen a modificar la
expresión de marcadores del ciclo celular compatibles con hiperplasia endometrial.

Resúmen
xi
Para llevar a cabo el estudio se utilizaron 19 endometrios obtenidos de mujeres con
fertilidad probada en fase proliferativa del ciclo menstrual (EN), de pacientes SOP sin
y con hiperplasia endometrial (ESOP, n = 28; y ESOPH, n = 14, respectivamente), y
de mujeres con hiperplasia endometrial (HE, n = 12). En el estudio ex vivo, se
evaluó en estos tejidos la expresión génica (RT-PCR) y proteica (Inmunohistoquímica
y/o Western Blot) de los receptores de andrógenos (RA) y estrógenos (REα y REβ),
de co-reguladores, así como, de la enzima P450 aromatasa. Además, se estudió la
proliferación celular por Ki67, y la apoptosis a través de estudios de fragmentación
del DNA y de expresión proteica de Bcl-2, Bax, Pro-caspasa-3 y Caspasa-3.
También, se investigó la expresión de la onco-proteína Ras, de la proteína supresora
de tumores PTEN, y la expresión y actividad de la proteína Akt. Por otro lado, se
evaluó la expresión de reguladores del ciclo celular tales como: p27, p21, proteína
del retinoblastoma (Rb), Ciclina E y Cdk-2 (Western blot y/o Inmunohistoquímica), y
la actividad de Cdk-2. Además, se realizó un estudio in vitro, en el cual se evaluó la
respuesta del tejido endometrial (explantes) a hormonas esteroidales e insulina
adicionados a los cultivos, con el propósito de simular la condición SOP. Los
parámetros que se investigaron en el estudio in vitro fueron: el co-activador AIB1, y
reguladores del ciclo celular tales como: p27, Rb, Ciclina E y Cdk-2.
Los resultados muestran que el endometrio SOP presenta una mayor sensibilidad a
la acción esteroidal, lo cual fue evidenciado por la mayor expresión del RA en ESOP
y ESOPH; el incremento observado en la expresión del co-activador AIB1 y del REα
en ESOP; junto con la alta expresión del co-activador ARA70 y la localización
citoplasmática del co-represor NCoR en todos los grupos de endometrios estudiados.

Resúmen
xii
En los estudios in vitro también se observó un incremento en la expresión de AIB1,
en presencia de una alta concentración de andrógenos, estrógenos e insulina.
Por otro lado, no encontramos expresión de la enzima P450 aromatasa en los
endometrios estudiados, lo que nos hace pensar que será necesario realizar estudios
de actividad de la enzima a fin de conocer si aumenta la biodisponibilidad de
estrógenos en los endometrios SOP.
Concordantemente con nuestra proposición de que cambios en la expresión de
receptores esteroidales y co-reguladores, podrían desregular la expresión de
moléculas importantes en el control de la homeostasis tisular, encontramos un
aumento de la proliferación celular (por Ki67) en los endometrios SOP, en ausencia
de apoptosis en todos los grupos de endometrios estudiados. Por lo tanto, las
pacientes SOP presentan una desregulación en su homeostasis endometrial. Tal
desregulación fue evidenciada al estudiar a reguladores del ciclo celular como p27,
encontrando una disminución en su expresión nuclear en las células estromales de
pacientes SOP (específicamente, pacientes SOP insulino-resistentes) y de ESOPH.
Lo anterior, coincide con los hallazgos del estudio in vitro donde se observó que en
presencia de insulina existe una leve tendencia a la disminución de p27. Esto
sugiere una posible contribución de esta proteína para el progreso hacia la fase S del
ciclo celular. Por otro lado, se detectó un incremento en la expresión citoplasmática
de p27 en células epiteliales de ESOP, lo que podría indicar que en este tipo celular,
p27 favorecería el ensamblaje del complejo Ciclina E/Cdk-2 e inhibiría la apoptosis.
En cuanto a Ciclina E y Cdk-2, se encontró que ambas proteínas se expresaron
principalmente en las células epiteliales de los grupos de endometrios estudiados,

Resúmen
xiii
igual que para el caso de Ki67. Además, no se detectaron cambios en la expresión
y/o actividad de Ciclina E y Cdk-2 entre los cuatro grupos de endometrios, a
excepción de Cdk-2 donde se observó un aumento en su expresión nuclear desde
ESOP a ESOPH y EH. Estos resultados son importantes si tenemos en cuenta que
los endometrios de las mujeres normales expresan estas proteínas en fase
proliferativa, mientras que dicha expresión disminuye en fase secretora. En cambio,
los endometrios SOP expresan continuamente estas moléculas, dado que las
mujeres SOP son amenorreicas u oligo-menorreicas.
En el caso de la proteína supresora de tumores Rb, no se observaron cambios en su
expresión entre los endometrios estudiados. Sin embargo, en los ensayos in vitro, en
presencia de insulina se observó una tendencia a la disminución de Rb.
También se estudió a la proteína supresora de tumores PTEN, la cual no cambia su
expresión entre los endometrios de interés; no obstante, a través de estos resultados
no podemos concluir acerca de la funcionalidad de PTEN. Para ello, se analizó a la
proteína anti-apoptótica Akt, la que se encuentra río abajo de PTEN y que se activa
por fosforilación en ausencia o inactividad de PTEN. Encontramos un aumento en la
fosforilación de Akt en el residuo de Ser473 en ESOP y ESOPH al comparar con EN,
lo que sugiere que ésta sería una de las vías que conducen al aumento de la
supervivencia de las células endometriales de las pacientes SOP. Otra de las vías
que podría colaborar con lo anteriormente señalado es la de Ras, ya que
observamos sobreexpresión de esta proteína en los endometrios de pacientes SOP
sin y con hiperplasia, principalmente a nivel de las células epiteliales.

Resúmen
xiv
En lo que se refiere a los estudios in vitro, tal como fue descrito anteriormente, no
observamos cambios importantes en condiciones hiperandrogénicas,
hiperestrogénicas y/o hiperinsulínicas. Es factible pensar que ello puede deberse a
que en 18 h de cultivo es difícil reproducir un síndrome como es el SOP, y que
además, los cambios en la expresión de las proteínas que nos interesaron estudiar
pueden requerir más tiempo para ser visualizados.
Por lo expuesto anteriormente, podemos concluir que la condición endocrino-
metabólica inherente al SOP afecta la función normal del endometrio, a nivel de la
homeostasis tisular y de la supervivencia celular. De modo tal que los endometrios
sometidos a estos ambientes hormonales anormales, experimentan descoordinación
entre proliferación y apoptosis, constituyendo éste un evento importante que puede
conducir al crecimiento inadecuado o patológico del endometrio de mujeres con SOP,
y potencialmente al desarrollo de hiperplasia y/o carcinoma endometrial.

Abstract
xv
v. ABSTRACT
The endometrium is one of the tissues of the organism whose function is regulated in
an important way by ovarian steroids. The estradiol produced in the follicular phase
induces endometrial cell proliferation, and the progesterone produced in the luteal
phase promotes cell differentiation. To achieve this regulation, endometrial cells
express different isoforms of steroid receptors. Also, the steroid action through the
receptors can be controlled by co-regulators, such as co-activators or co-repressors.
It has been reported that estradiol would regulate the expression of various molecules
involved in the control of the cell cycle, not only in the endometrium but in other
tissues and cell lines, as well. Moreover, it is widely known the mitogenic action of
insulin in the endometrium, however, the proliferative action of androgens it is not yet
clearly defined. The latter could explain, in part, the higher relative risk in the
development of endometrial hyperplasia and carcinoma found in patients with
Polycystic Ovarian Syndrome (PCOS), which is a pathology characterized by
endocrine and metabolic alterations.
The aim of this thesis was to evaluate if the expression of steroid receptors,
regulators of the cell cycle, and the degree of proliferation and apoptosis is related to
the development of endometrial hyperplasia in PCOS-women. Also, to determine in
vitro if the hyperandrogenic or hyperestrogenic or hyperinsulinic condition, contributes
to modify the expression of cell-cycle markers compatible with endometrial
hyperplasia.

Abstract
xvi
For this purpose, endometria from four groups of women were obtained. The first
group corresponds to endometria from women with proven fertility in proliferative
phase (NE, n = 19); the second group is composed by endometrium of untreated-
PCOS women (PCOSE, n = 28); the third group corresponds to PCOS women with
endometrial hyperplasia (HPCOSE, n = 14); the last group is formed by endometrium
from women with endometrial hyperplasia (HE, n = 12).
In the ex-vivo study, the expression of androgen and estrogen receptors, co-
regulators, and the P450 aromatase enzyme in the tissues were evaluated by RT-PCR
and/or immunohistochemistry and/or Western blot. Also, cell proliferation was
evaluated by Ki67, and apoptosis by DNA fragmentation, Bcl-2, Bax, Pro-caspase-3
and Caspase-3 protein expression. In addition, the expression of Ras onco-protein,
PTEN tumor suppressor protein, and Akt protein expression and activity, was
investigated. Also, the expression of cell cycle regulators, such as, p27, p21,
retinoblastoma protein (Rb), Cyclin E, and Cdk-2 through Western blot and/or
immunohistochemistry, and Cdk-2 activity was determined. An in vitro study was
carried out for evaluating the answer of the endometrial tissue (explants) to steroid
hormones and insulin added to cultures, in order to simulate the PCOS condition.
The parameters investigated in vitro were: the AIB1 co-activator, and the cell cycle
regulators like: p27, Rb, Cyclin E and Cdk-2.
The results show that the PCOS endometrium has a greater sensitivity to the steroid
action, based on the higher expression of the androgen receptor in PCOSE and
HPCOSE, the increase of AIB1 and estrogen receptor-alpha in PCOSE, along with
the high expression of the ARA70 co-activator and the cytoplasmic localization of the

Abstract
xvii
NCoR co-repressor in all the endometria studied. In the in vitro studies, also was
observed an increase of the expression of AIB1 in the presence of a high
concentration of androgens, estrogens and insulin.
On the other hand, we did not detect expression of P450 aromatase enzyme in all
studied endometria, which suggests that it will be necessary to attain activity studies
to know if this enzyme enhances the bio-availability of estrogens in the PCOS
endometria.
Accordingly with our proposal that changes in the expression of steroid receptors and
co-regulators could deregulate the expression of important molecules involved in the
control of tissue homeostasis, we found an increase in cellular proliferation (by Ki67)
in the PCOS endometrium when compared to normal endometrium, in absence of
apoptosis in all the groups studied. Given the above, the PCOS-patients present
abnormal regulation in their endometrial homeostasis, based on the data obtained for
regulators of the cell cycle, like p27, finding a lowered expression level in the stromal
cells of HPCOSE, and in insulin-resistant PCOS patients. The latter are in agreement
with the results of the in vitro studies: a slight decrease of p27 in the presence of
insulin. This suggests a possible contribution of this protein to the initiation of S-
phase of the cell cycle. Moreover, it was found an increment in the cytoplasmic
expression of p27 in PCOSE epithelial cells, which could indicate that in this cell type,
p27 would promote the linkage of the Cyclin E/Cdk-2 complex and inhibit apoptosis.
The Cyclin E and Cdk-2 proteins were expressed mainly in the epithelial cells of the
endometria studied, same as Ki67. Also, no changes in the expression or activity of
Cyclin E and Cdk-2 were detected in the endometria studied, except for Cdk-2, where

Abstract
xviii
an increase of its nuclear expression from PCOSE to HPCOSE and HE was obtained.
These results are important if we take into account that the endometria of normal
women express Cyclin E and Cdk-2 in proliferative phase, but its expression
decreases in secretory phase. However, the PCOS endometria continuously express
those molecules, due to the fact that these women are amenorreic or oligo-menorreic.
In the case of the Rb tumor-suppressor protein, no noticeable changes in its
expression were detected between the studied endometria. However, in the in vitro
assays in the presence of insulin, a tendency to a decrease in the Rb level was
observed.
Also, the PTEN tumor suppressor protein was studied, but not changes in its
expression were found. We cannot conclude about the functionality of PTEN through
this study. To have a greater insight, the anti-apoptotic protein Akt was analyzed. Akt
is located downstream PTEN, and is activated by phosphorylation in the absence or
inactivity of PTEN. We found an increment in the phosphorylation of Akt in the
Ser473 residue in PCOSE and HPCOSE compared to NE, which suggests this would
be one of the reasons to explain the higher survival of the endometrial cells from
PCOS-patients. Another pathway that could contribute to cell survival is the over-
expression of Ras observed in PCOS-patients with and without hyperplasia, mainly in
epithelial cells.
As was described earlier, we did not find important changes in hyperandrogenic,
hyperestrogenic and/or hyperinsulinic conditions in vitro. It is feasible to think that this
is because it is not easy to reproduce a syndrome like PCOS in 18-hour cultures, and

Abstract
xix
moreover, the changes in the expression levels of proteins studied may need more
time to be detected.
Therefore, it is possible to conclude that the endocrine-metabolic condition inherent to
PCOS affects the normal endometrium functions at the level of the tissue
homeostasis and cell survival. Thus, endometria subjected to this type of abnormal
hormonal environments, experiment a lack of coordination between proliferation and
apoptosis, configuring an important event that can lead to inadequate or pathologic
growth of endometrium from PCOS women, and to the potential development of
endometrial hyperplasia and/or carcinoma.

Introducción
1
I. INTRODUCCION
I.1 EL ENDOMETRIO HUMANO
El endometrio es la capa que reviste al útero y se encuentra estructuralmente dividido
en dos capas: la funcionalis y la basalis. La capa funcionalis consta de un epitelio
superficial, el cual se invagina dando origen a las glándulas uterinas tubulares
(epitelio glandular) (1) y de un estroma constituido por fibroblastos, células
endoteliales, células musculares lisas y leucocitos. Además, otro importante
componente de todo este microambiente celular lo constituye la matriz extracelular
(MEC), la que en el endometrio humano presenta variaciones a través del ciclo
menstrual (2). La MEC está formada por la membrana basal en la que descansan las
células epiteliales, y por una matriz intersticial rica en colágeno en la que se localizan
los fibroblastos, además, rodea al endotelio de los vasos y a las células deciduales
(2). La morfología y la función de las células endometriales están fuertemente
influenciadas por los cambios cíclicos que ocurren en la síntesis de esteroides
sexuales por el ovario. Estos cambios producen características determinadas en la
arquitectura endometrial, lo que llevó a Noyes (3), en la década de los 50, a
establecer una serie de criterios que permiten determinar el día del ciclo en el cual se
encuentra el tejido endometrial. Basado en lo anterior, es factible definir dos etapas
principales a través del ciclo: la fase proliferativa y la fase secretora. Durante la fase
proliferativa en el endometrio (o fase folicular ovárica), el estradiol (E2) producido por
los folículos en desarrollo induce la proliferación celular en el endometrio. Esta activa
proliferación de todos los componentes del tejido, alcanza su máxima actividad en los

Introducción
2
días 8 a 10 de ciclo (4). Hacia el final de la fase proliferativa, el lumen de las
glándulas se ensancha y éstas adquieren un contorno ondulado. Al inicio de la fase
secretora se acumulan vesículas de glicógeno en la región basal de las células
glandulares (vesículas subnucleares), lo cual es el primer indicio de que la ovulación
ha ocurrido (5). En esta etapa, el endometrio aumenta su grosor debido a la
hipertrofia de las células glandulares y a edema en el estroma. Por acción de la
progesterona (P4) producida por el cuerpo lúteo, las células estromales son inducidas
a diferenciarse en células de tipo predeciduales (a partir del día 23 del ciclo),
aumentan de tamaño, se redondean y secretan algunos péptidos tales como proteína
ligante de factores de crecimiento insulino símil tipo 1 (IGFBP-1) y prolactina. Al final
de la fase secretora ocurre un dramático aumento de leucocitos polimorfonucleares y
de prostaglandinas (PGs) F2α y E2 (6, 7), que posiblemente estimulan la
vasoconstricción endometrial. Esto lleva a isquemia de la funcionalis y a una
destrucción parcial de la mucosa, lo que conduce a necrosis tisular durante la
menstruación (5).
Dado que el endometrio es uno de los tejidos cuya función es principalmente
regulada por los esteroides circulantes de origen ovárico, en situaciones endocrino-
metabólicas asociadas a anormalidades en la concentración sanguínea de algunas
hormonas y proteínas, como es el caso del Síndrome de Ovario Poliquístico (SOP),
los procesos que ocurren en el endometrio y la forma como las células responden a
los estímulos biológicos alterados, pueden verse afectados.

Introducción
3
I.2 SINDROME DE OVARIO POLIQUISTICO (SOP)
El SOP es una de las patologías más frecuentemente relacionadas con infertilidad y
aborto recurrente. Es un trastorno de alta prevalencia en las mujeres en edad fértil,
afectando entre el 5-10% de este grupo de mujeres (8). Se caracteriza por un
hiperandrogenismo funcional, amenorrea/oligomenorrea, e hiperinsulinemia asociada
a insulino-resistencia en el 50-60% de las pacientes con SOP (9). Además, en las
pacientes SOP no sólo se encuentran elevados los niveles séricos de andrógenos
sino que, además, ellas se caracterizan por presentar un hiperestrogenismo relativo
(dada la acción de los estrógenos no compensada por P4) (10). La hipersecreción de
insulina junto a la sobreexpresión de la enzima P450 c17 (responsable de la síntesis de
andrógenos en las células tecales del folículo ovárico), aumentan los andrógenos
intraováricos y se detiene el desarrollo folicular. En consecuencia, se produce la
acumulación de pequeños quistes foliculares subcorticales e incrementa el volumen
ovárico, lo que le da la imagen ultrasonográfica característica a este síndrome. A
pesar de que el SOP fue identificado hace más de 60 años (11), los conocimientos
actuales permiten identificarlo como un síndrome de gran heterogeneidad de
fenotipos y que presenta múltiples anormalidades. Es por ello que su etiogenia aún
no está establecida y podría incluir una combinación de factores genético-
ambientales. Además, este síndrome constituye un problema de salud mayor para la
mujer, con ramificaciones más allá de las anormalidades endocrinas y reproductivas
y está relacionado con enfermedades como diabetes tipo 2, hipertensión, función
suprarrenal alterada y enfermedades cardiovasculares (9, 10).

Introducción
4
Trabajos anteriores han reportado que el endometrio de mujeres con SOP se
comporta de manera diferente al endometrio de mujeres normales, lo cual se
evidencia con un aumento significativo de abortos recurrentes (1% en mujeres SOP)
comparado con la población normal (0,34%) (12, 13). Además, se ha asociado la
condición endocrina-metabólica inherente al SOP con una mayor probabilidad de
desarrollar hiperplasia endometrial (35,7%), como lo señalan estudios que establecen
una asociación entre mujeres nulíparas que padecen SOP e hiperplasia endometrial
(14, 15). De ellas, 60, 15 y 25% fueron clasificadas como hiperplasia simple,
compleja e hiperplasia con atipia citológica, respectivamente (15). Por otro lado, se
ha reportado que la condición SOP presenta un mayor riesgo relativo (3.1) de
desarrollar carcinoma endometrial (16, 17, 18) (Figura 1). Sin embargo, la naturaleza
multifactorial del SOP y su heterogeneidad hace difícil conocer los factores asociados
al SOP que son causa de riesgo para el desarrollo de hiperplasia y carcinoma
endometrial. Entre éstos se incluyen la obesidad, acción estrogénica no
compensada por progesterona, nuliparidad, hipertensión, diabetes mellitus tipo 2,
amenorrea/oligomenorrea, todas características que están asociadas al SOP (19).
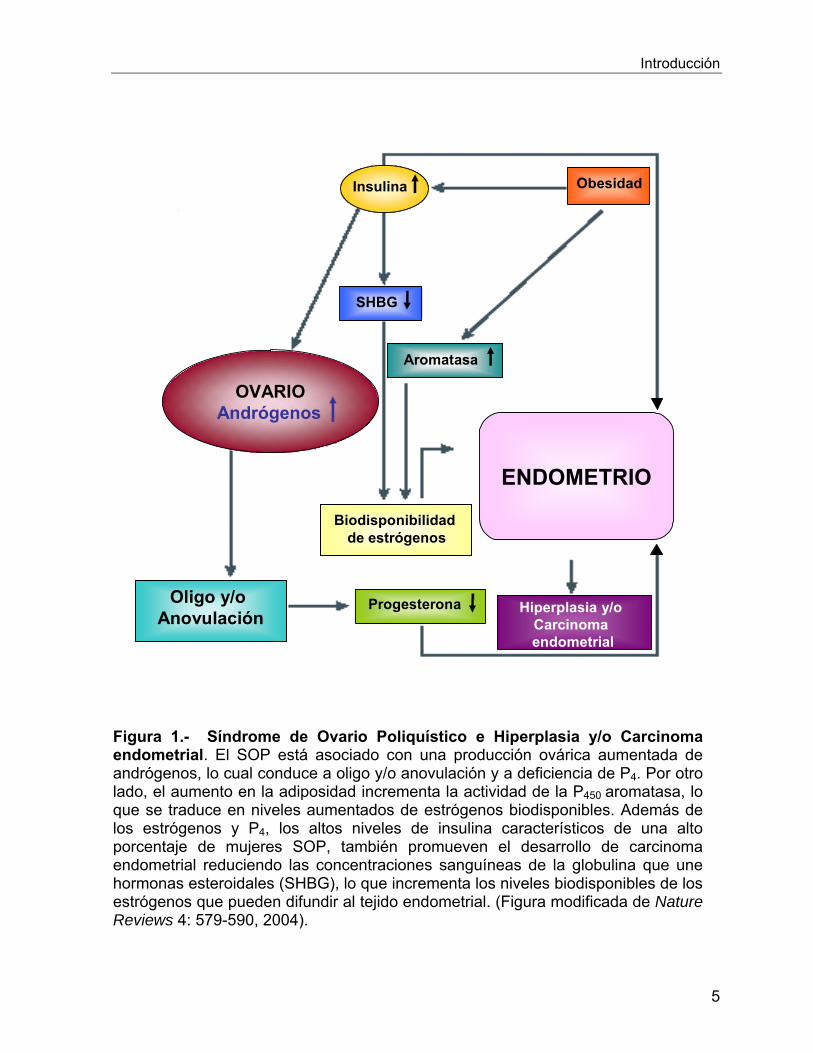
Introducción
5
Figura 1.- Síndrome de Ovario Poliquístico e Hiperplasia y/o Carcinoma endometrial. El SOP está asociado con una producción ovárica aumentada de andrógenos, lo cual conduce a oligo y/o anovulación y a deficiencia de P4. Por otro lado, el aumento en la adiposidad incrementa la actividad de la P450 aromatasa, lo que se traduce en niveles aumentados de estrógenos biodisponibles. Además de los estrógenos y P4, los altos niveles de insulina característicos de una alto porcentaje de mujeres SOP, también promueven el desarrollo de carcinoma endometrial reduciendo las concentraciones sanguíneas de la globulina que une hormonas esteroidales (SHBG), lo que incrementa los niveles biodisponibles de los estrógenos que pueden difundir al tejido endometrial. (Figura modificada de Nature Reviews 4: 579-590, 2004).
OVARIOAndrógenos
ENDOMETRIO
Oligo y/o Anovulación
Insulina Obesidad
Aromatasa
Progesterona
Biodisponibilidad de estrógenos
SHBG
Hiperplasia y/o Carcinoma endometrial
OVARIOAndrógenos
ENDOMETRIO
Oligo y/o Anovulación
Insulina Obesidad
Aromatasa
Progesterona
Biodisponibilidad de estrógenos
SHBG
Hiperplasia y/o Carcinoma endometrial
OVARIOAndrógenos
ENDOMETRIO
Oligo y/o Anovulación
Oligo y/o Anovulación
InsulinaInsulina Obesidad
AromatasaAromatasa
Progesterona
Biodisponibilidad de estrógenos
Biodisponibilidad de estrógenos
SHBG
Hiperplasia y/o Carcinoma endometrial

Introducción
6
I.3 HORMONAS OVARICAS
Tanto los estrógenos como la P4 son hormonas ováricas que controlan la ciclicidad
endometrial (20). En forma similar a la de otros miembros de la familia de hormonas
esteroidales, los andrógenos, promueven sus efectos genómicos vía receptores
nucleares. Progesterona y andrógenos se encuentran principalmente unidos a la
proteína ligante de esteroides sexuales (SHBG: sex-steroid-binding globulin), y solo
la fracción libre de estas hormonas es responsable de sus efectos hormonales.
Entonces, los efectos hormonales pueden estar regulados por un exceso o por una
disminución de SHBG. Así, concentraciones elevadas de andrógenos e insulina
inducen una disminución de la producción de SHBG (21). En este contexto, es
importante mencionar que la concentración circulante de SHBG está disminuida en
las pacientes con SOP (9).
Los efectos hormonales también pueden ser modulados localmente por factores de
crecimiento (22, 23).
I.4 RECEPTORES ESTEROIDALES
Es conocido que las acciones de los esteroides en los tejidos, incluido el endometrio,
están mediadas por receptores específicos, constituyendo factores de transcripción
importantes para numerosos genes, entre los que destacan aquellos relacionados
con la proliferación celular y el control del ciclo celular.
La expresión de los receptores está altamente controlada por los esteroides
circulantes, por lo que obviamente concentraciones plasmáticas alteradas de los
esteroides pueden conducir a una desregulación en la expresión endometrial de

Introducción
7
estos receptores, con consecuencias negativas sobre la implantación y/o viabilidad y
diferenciación de las células endometriales.
Por otro lado, en los últimos años se ha establecido que la transcripción mediada por
los receptores de estrógenos puede ser estimulada por mecanismos independientes
de los ligandos y que involucran vías de segundos mensajeros, como las proteínas
quinasas activadas por mitógenos (MAPK), que estimulan la fosforilación en residuos
de serina del receptor, las cuales pueden ser tejido-específicas (24).
I.4.1 Receptores endometriales de estrógenos
En condiciones fisiológicas, el efecto de los estrógenos sobre el tejido endometrial
puede realizarse a través de las distintas isoformas que presentan los receptores. Se
ha demostrado que los estrógenos (fundamentalmente E2) ejercen su acción a través
de dos isoformas del receptor, α yβ, codificadas por dos genes distintos ubicados en
diferentes cromosomas (cromosomas 6 y 14, respectivamente). Aunque la expresión
de ambos receptores aumenta por acción del estrógeno circulante durante la fase
proliferativa del ciclo menstrual normal, es la isoforma α la expresada de preferencia
en el endometrio humano normal, así como, en carcinoma endometrial (25, 26). El
predominio de cada una de estas isoformas es tejido-dependiente y, en ciertos
casos, se ha descrito una acción opuesta entre los subtipos de receptor al estar
unido a su ligando.
Además, resultan interesantes estudios realizados en ratones BERKO, los cuáles son
carentes de REβ, que sugieren un rol de esta isoforma del RE en la modulación de

Introducción
8
los efectos de estradiol sobre el REα, además de un efecto antiproliferativo en el
útero (27).
Considerando la capacidad de ambas isoformas para formar no sólo homodímeros
sino también heterodímeros, diferencias en la razón α/β, tanto en condiciones
normales como patológicas, podrían tener importantes implicaciones funcionales. Por
lo tanto, será interesante el análisis de lo anteriormente mencionado en endometrios
de pacientes SOP sin y con hiperplasia endometrial, ya que hasta el momento no hay
estudios al respecto.
I.4.2 Receptores de progesterona en endometrio
El receptor para P4 (RP) presenta dos formas, A y B, codificadas por el mismo gen,
pero utilizando distintos promotores (28). La isoforma A no posee el dominio de
transactivación constituido por 164 aminoácidos. La expresión de las formas A y B
tienen variaciones en el endometrio humano durante el ciclo menstrual, siendo la
isoforma A detectada de preferencia durante la fase secretora media, sugiriendo una
acción sobre la transcripción de genes involucrados en el proceso de implantación.
Los estrógenos inducen un aumento en la concentración tisular de la isoforma B,
siendo por lo tanto mayor su expresión en la fase proliferativa. Coincidentemente, en
cánceres endometriales dependientes de estrógenos se observa un predominio del
mRNA para la isoforma B (29). Por otro lado, en los endometrios SOP se ha
observado que la presencia del RP es similar a la de los endometrios normales (30).
Al igual que para los receptores de estrógenos, la unión de la P4 a las diferentes
formas de su receptor puede provocar respuestas antagónicas, lo que es tejido-

Introducción
9
dependiente. En este contexto, es importante considerar que la concentración
relativa de cada una de las isoformas puede ser importante para evaluar con mayor
precisión la acción esteroidal en estos tejidos.
I.4.3 Receptor de andrógenos endometrial
La concentración de andrógenos en el endometrio humano normal excede a la del
plasma (31, 32, 33) y al igual que los estrógenos y progesterona, los andrógenos
tienen una acción importante en la fisiología endometrial. Se ha demostrado que
tanto el mRNA como la proteína del receptor de andrógeno (RA), experimentan
cambios cíclicos durante el ciclo menstrual (34, 35). También, se ha observado un
aumento de dicho receptor en el núcleo de las células endometriales de fase
secretora inicial, lo que sugiere que el alza periovulatoria de andrógenos plasmáticos,
traería consigo un aumento en la unión del andrógeno al receptor. Se sabe que en la
fase proliferativa el RA es regulado positivamente por E2 (36, 37) y negativamente
por P4 en fase secretora (38). Además, se ha demostrado que los andrógenos
pueden interactuar con el RP (39) y con el RE, bloqueándolo (12) e induciendo
atrofia tisular en este último caso (40). Sin embargo, estudios ex vivo en endometrios
de pacientes SOP, publicados por nuestro grupo (30), sugieren que el
hiperestrogenismo es prevalente sobre los altos niveles de andrógenos en inducir la
expresión de los receptores esteroidales. Probablemente lo que tenga mayor
importancia en el SOP sea la condición hiperestrogénica relativa que presentan estas
mujeres, lo que genera en ellas un aumento de la acción de los estrógenos producto
de un estímulo más prolongado, y como es sabido los estrógenos modulan
positivamente la expresión de su propio receptor. Además, observamos que en los

Introducción
10
endometrios de mujeres SOP los RA se encuentran elevados, lo que en parte se
puede explicar por las concentraciones plasmáticas aumentadas de estrógenos y
andrógenos, lo que está de acuerdo con reportes previos (41). No obstante, no se
conoce acerca de la expresión de este receptor en endometrios de pacientes SOP
con hiperplasia.
I.5 CO-REGULADORES Y SU ACCION EN ENDOMETRIO HUMANO
Además de las isoformas de los receptores, la especificidad de la acción de los
esteroides está regulada por co-reguladores, los cuales pueden ser co-activadores o
co-represores de la transcripción génica mediada por los receptores esteroidales (42,
43, 44). Es probable que una sobreexpresión de los co-activadores o de los co-
represores en un determinado tejido, conduzca a un aumento o disminución de la
sensibilidad de ese tejido a la acción de los esteroides, respectivamente; por esto, es
importante considerar la concentración relativa entre estos co-reguladores.
La acción co-activadora se produce por estabilización del complejo de pre-iniciación
de la transcripción, o bien, por la capacidad de estos co-activadores de reclutar
histona-acetiltransferasas, las cuales por su acción desestabilizadora de las uniones
electrostáticas en el DNA, pueden aumentar el acceso de los factores de
transcripción al promotor favoreciendo la transcripción (45). Por otro lado, los co-
represores reclutan deacetilasas, las cuales inducen a la cromatina a adoptar una
estructura más compacta, la cual reprime la transcripción (45). En los últimos años,
se han identificado numerosos co-reguladores que modulan la acción de los
receptores esteroidales. Estos co-reguladores se expresan en diversos tejidos,

Introducción
11
incluyendo al endometrio humano. Entre los co-activadores más estudiados se
encuentran SRC-1/NCoA1; SRC-2/TIF2/GRIP1, SRC-3/RAC3/
ACTR/pCIP/AIB1/TRAM1 y ARA70. Cabe destacar que SRC-1 y SRC-3 (46, 47)
poseen actividad intrínseca de histona acetiltransferasa y entre los co-represores, el
NCoR y el SMRT, han sido los más investigados (45).
Se ha demostrado previamente que en el endometrio humano la expresión de co-
activadores está regulada durante el ciclo menstrual normal y más aún,
recientemente, se ha señalado que la expresión del co-activador SRC-3/AIB1
aumenta en endometrios de mujeres SOP tratadas con citrato de clomifeno (48). No
obstante, datos de nuestro laboratorio señalan que el citrato de clomifeno per se
induce una asincronía del desarrollo entre epitelio y estroma, alterando parámetros
de función endometrial. Lo anterior es de relevancia y debería ser considerado al
evaluar los modelos experimentales que utilizan este fármaco. A pesar de lo anterior,
se infiere que la mayor concentración plasmática de estrógenos y de andrógenos,
sumado a la elevada expresión endometrial de los RE, RA y de los co-activadores
encontrados en los endometrios de pacientes SOP, pudiesen favorecer la respuesta
exagerada a la acción de los esteroides en este tejido, conduciendo a alteraciones en
la fisiología endometrial. Por lo dicho anteriormente, será relevante estudiar la
expresión de algunos de estos co-reguladores en endometrios de pacientes SOP sin
tratamiento, a fin de elucidar si ellos aumentan la sensibilidad a la acción esteroidal
en estas pacientes, contribuyendo potencialmente al desarrollo de hiperplasia
endometrial.

Introducción
12
I.6 HOMEOSTASIS ENDOMETRIAL
Dado que el endometrio se encuentra bajo el control de esteroides sexuales, su
homeostasis está regulada por hormonas durante el ciclo menstrual. De esta forma,
el endometrio está sujeto a procesos de remodelación a través de la menstruación,
proliferación y diferenciación de las células endometriales, y durante dichos procesos
se produce un balance específico entre apoptosis y proliferación tanto en el epitelio
como en el estroma (49, 50).
I.6.1 Apoptosis
La apoptosis es un proceso activo que los organismos multicelulares han
desarrollado para eliminar las células no necesarias o potencialmente perjudiciales.
Por lo tanto, ocurre en situaciones fisiológicas y fisiopatológicas, y las células se
suicidan bajo la acción de controles internos y externos (51).
La activación de este proceso puede ser dependiente del ligando FAS y comenzar a
nivel de la membrana, que es lo que se conoce como vía extrínseca. También puede
ocurrir una activación independiente del ligando (vía mitocondrial o vía intrínseca), la
cual está dada por el desbalance entre proteínas pro- y anti-apoptóticas de la familia
Bcl-2. Ambas vías convergen en la activación de las caspasas-3, -6 y -7 siendo la
principal la caspasa-3. Sin embargo, en los últimos años se han identificado
procesos que si bien corresponden a apoptosis, son independientes de caspasas
(52).
Es importante mencionar que la apoptosis es un proceso que difiere de la muerte
celular por necrosis (53), ya que el primero afecta sólo a células aisladas y se

Introducción
13
manifiesta por cambios severos y característicos a nivel nuclear y citoplasmático. El
núcleo de las células apoptóticas presenta condensación de la cromatina y
desensamblaje de la envoltura nuclear. Uno de los marcadores más característicos
de apoptosis lo constituye la fragmentación del DNA en segmentos de 180- 200 pb
(54), aunque la ausencia de clivaje en el DNA no descarta la presencia de apoptosis.
A nivel citoplasmático, la característica morfológica más importante es la
fragmentación de la célula en los llamados cuerpos apoptóticos. La membrana
plasmática también presenta alteraciones, las que se manifiestan en la separación de
las células apoptóticas de sus vecinas (por pérdida de las uniones intercelulares),
exposición de marcadores de superficie y cambios en la composición de los lípidos
de membrana, entre ellos el fosfolípido fosfatidilserina se transloca desde la cara
citoplasmática de la membrana celular a la superficie externa de la célula. Este
desensamblaje apoptótico ordenado y secuencial es silente, y lleva a que estas
células pasen desapercibidas por el sistema de respuesta inflamatoria, y sean
solamente reconocidas por los macrófagos que fagocitan los cuerpos apoptóticos
(54). En relación a los agentes que inducen apoptosis, éstos son tejidos específicos.
En endometrio humano normal, la apoptosis ha sido observada a través de la fase
lútea, menstruación, en fase folicular temprana (50, 55, 56), y localmente en el sitio
de implantación (57, 58). Además, el proceso de apoptosis cíclico en el endometrio,
provee evidencias de la regulación hormonal de la apoptosis (59) y de la importancia
de este proceso en la remodelación endometrial.
Por otra parte, Gompel y cols. (60) describieron la presencia de la proteína anti-
apoptótica Bcl-2 en endometrio humano. Estos autores mostraron una variación en

Introducción
14
la expresión de Bcl-2 durante el ciclo endometrial, comenzando en la parte baja de la
capa funcional y extendiéndose por todas las glándulas durante la fase proliferativa y
desapareciendo cuando el epitelio se hace secretor. En tanto, en el estroma la
expresión de Bcl-2 se hace coincidente con la diferenciación de las células. Estos
resultados sugieren que la expresión de Bcl-2 en las glándulas es inhibida a medida
que aumenta la concentración de P4. Además, Castro y cols. (61), han entregado
evidencia adicional que avala la relación entre la disminución de la expresión de Bcl-
2 y del proto-oncogen c-myc con el aumento de células apoptóticas en endometrio en
fase secretora. Se ha descrito por medio de análisis de inmunohistoquímica, que
otras proteínas de la familia Bcl-2, tales como Bax, Bak, Bcl-Xs y Bcl-XL, tendrían
variaciones cíclicas en su expresión en el endometrio humano, siendo la proteína
pro-apoptótica Bax mayormente expresada en endometrio secretor que en
proliferativo (62, 63). Se conoce que Bax ejerce su efecto pro-apoptótico a través de
la liberación de citocromo c mitocondrial, lo cual activa a caspasas, enzimas
efectoras del proceso apoptótico. Del mismo modo, Bak, otra proteína pro-apotótica,
ha sido escasamente detectada en fase proliferativa, pero aumenta en fase secretora
donde estaría confinada casi exclusivamente al epitelio glandular (63). Por su parte,
en endometrio humano de fase proliferativa, es factible observar algunas células con
una fuerte tinción para Bax y una débil tinción de Bcl-2 sin ser apoptóticas, lo que
indica que existirían factores (que no usan la vía mitocondrial) involucrados también
en la regulación de la apoptosis en las células endometriales (62). Debido a ésto, se
ha sugerido que la acción de las proteínas de la familia Bcl-2 debe considerarse en
relación a la de otras proteínas reguladoras de la apoptosis durante el ciclo
menstrual.

Introducción
15
I.6.2 Proliferación
La proliferación cíclica es una característica de los órganos reproductivos femeninos
durante los años fértiles, y la hormona folículo estimulante (FSH) y E2 son las
principales hormonas estimuladoras de la proliferación, especialmente en ovario y
endometrio (64).
La proliferación del epitelio endometrial es máxima durante la fase folicular, y es
detenida después de la ovulación (65). Sin embargo, las células estromales pueden
proliferar durante la fase lútea (66) como ocurre en el proceso de implantación (57).
La capa basal (o basalis) del endometrio presenta una proliferación baja pero
constante a través del ciclo menstrual, y los cambios cíclicos son principalmente
marcados en la capa funcional (o funcionalis) del endometrio (67).
La proliferación de las células endometriales involucra una serie de eventos que
comienza con la unión de E2 a su receptor nuclear, con la consiguiente activación de
los elementos de respuesta a E2. Como consecuencia, se produce la activación de la
transcripción de algunos genes específicos para factores de crecimiento, entre otros,
que actúan de manera paracrina y/o autocrina en la expresión de proto-oncogenes,
tales como c-myc, c-fos, c-jun y c-ras (68). Es importante destacar que en
experimentos in vitro las células aisladas del epitelio endometrial presentan una baja
respuesta proliferativa a E2, por lo que se considera indispensable la presencia de las
células estromales en la respuesta mitogénica de las células glandulares en la fase
proliferativa. Por otra parte, P4 contrarresta el efecto de E2 ya que produce una
disminución en los receptores para estrógenos y un aumento en el metabolismo de
E2 a conjugados no activos (67).

Introducción
16
Con respecto al efecto de otros esteroides como los andrógenos, los cuales tienen un
efecto tanto proliferativo como apoptótico en otros tejidos reproductivos, poco se
sabe de su acción en la fisiología o fisiopatología del endometrio. En células
estromales se ha reportado un aumento en la expresión de receptores de factores de
crecimiento al ser estimuladas por andrógenos (69). Debido a estos antecedentes es
factible pensar que los andrógenos puedan tener alguna función en particular durante
el ciclo endometrial. Sin embargo, estudios realizados en células del epitelio
endometrial han mostrado que los andrógenos provocan una disminución en la
proliferación celular y en la secreción de glicodelina, proteína que sirve como
marcador de la función endometrial (70). Además, se ha propuesto que la mayor
concentración de andrógenos plasmáticos se correlacionaría con una disminución de
proteínas marcadoras de la función endometrial, así como, con un aumento de la
tasa de abortos recurrentes (12, 13).
La capacidad para medir proliferación celular y conocer los factores que la regulan,
es de gran interés ya que la proliferación está asociada con carcinogénesis
endometrial (71, 72).
Los eventos que tienen lugar en las células proliferantes son ilustrados en el ciclo
celular (Figura 2):
• La fase G1 representa el periodo entre la mitosis y la fase S. La mayoría de
las funciones especializadas de las células son realizadas durante esta fase y
en G0, y el contenido de DNA de las células diploides normales es equivalente
a un doble set de cromosomas. Las células están bajo la influencia de
factores de crecimiento, onco-genes, etc; que pueden hacer avanzar a las

Introducción
17
células a la fase S del ciclo celular o entrar en arresto en G0. Antes de entrar
a la fase S, hay un importante punto de restricción, bajo el control de un gen
supresor de tumores, p53, que es capaz de detener la proliferación de células
genéticamente defectuosas y conducirlas a G0 para ser reparadas o entrar en
apoptosis.
• La fase S es el periodo durante el cual el DNA es duplicado.
• En fase G2 las células presentan un contenido de DNA duplicado y se
preparan para la mitosis.
• Durante la Mitosis (M) se produce la división celular, con la consecuente
condensación de la cromatina y distribución de los cromosomas entre las
células hijas.
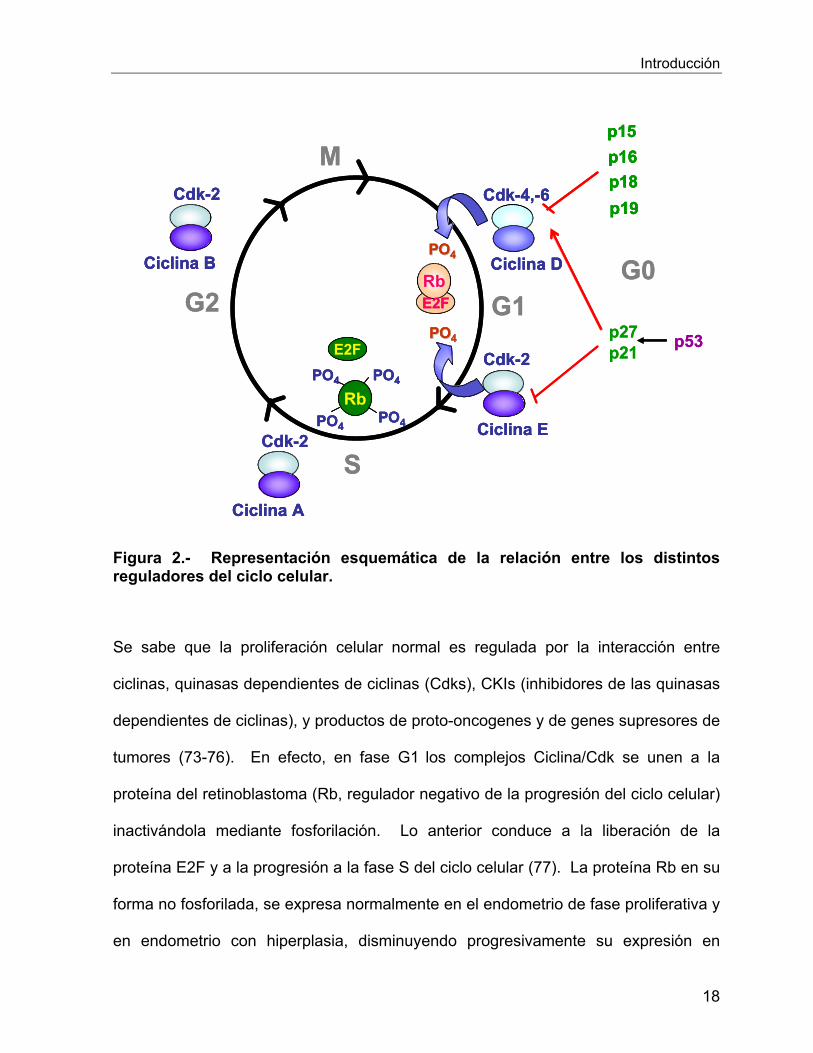
Introducción
18
Figura 2.- Representación esquemática de la relación entre los distintos reguladores del ciclo celular.
Se sabe que la proliferación celular normal es regulada por la interacción entre
ciclinas, quinasas dependientes de ciclinas (Cdks), CKIs (inhibidores de las quinasas
dependientes de ciclinas), y productos de proto-oncogenes y de genes supresores de
tumores (73-76). En efecto, en fase G1 los complejos Ciclina/Cdk se unen a la
proteína del retinoblastoma (Rb, regulador negativo de la progresión del ciclo celular)
inactivándola mediante fosforilación. Lo anterior conduce a la liberación de la
proteína E2F y a la progresión a la fase S del ciclo celular (77). La proteína Rb en su
forma no fosforilada, se expresa normalmente en el endometrio de fase proliferativa y
en endometrio con hiperplasia, disminuyendo progresivamente su expresión en
M
S
G2 G1
Cdk-4,-6
Ciclina D
Cdk-2
Ciclina E
Cdk-2
Ciclina BRbE2F
p16p18p19
p15
p27p21 p53
POPO44
POPO44
RbPO4
PO4PO4
PO4
E2F
Cdk-2
Ciclina A
G0
M
S
G2 G1
Cdk-4,-6
Ciclina D
Cdk-4,-6
Ciclina D
Cdk-2
Ciclina E
Cdk-2
Ciclina E
Cdk-2
Ciclina B
Cdk-2
Ciclina BRbE2FRbE2FRbE2F
p16p18p19
p15p16p18p19
p15p16p18p19
p15
p27p21 p53
POPO44
POPO44
POPO44
POPO44
RbPO4
PO4PO4
PO4
E2F
RbPO4
PO4PO4
PO4
E2F
Cdk-2
Ciclina A
Cdk-2
Ciclina A
G0

Introducción
19
carcinoma endometrial (78). Sin embargo, aún no se sabe si existe relación entre la
expresión de esta proteína y el potencial desarrollo de hiperplasia y/o carcinoma
endometrial en pacientes SOP.
Por otro lado, se sabe que la capacidad de los complejos Ciclina/Cdk para fosforilar
Rb es inhibida por inhibidores CKIs, entre los que se destacan las proteínas p15,
p16, p21 y p27, las cuales suprimen la actividad quinasa de los complejos
Ciclina/Cdk conduciendo al arresto del ciclo celular (73, 79). Por otro lado, la
proteína p53, estimula la síntesis de p21 y p27 regulando el ciclo celular y además
regula la apoptosis por activación de la transcripción del gen bax.
Reportes de la literatura indican que las Ciclinas muestran cambios en hiperplasia y
carcinoma endometrial respecto al endometrio normal; así se ha descrito, por
ejemplo, que la ciclina A se correlaciona positivamente con la expresión de Ki67,
marcador de proliferación celular (80). Sin embargo, no se conoce si existe una
desregulación a nivel de estos reguladores del ciclo celular en endometrios de
pacientes SOP con y sin hiperplasia.
Cambios en el balance entre apoptosis y proliferación pueden ser fisiológicos, como
son los cambios que ocurren durante el ciclo menstrual, donde durante las fases de
proliferación y diferenciación se produce un balance específico entre apoptosis y
proliferación tanto en el epitelio como en el estroma. Sin embargo, hasta la fecha no
se sabe si este balance existe a nivel del endometrio de las mujeres con SOP.

Introducción
20
I.7 REGULADORES DEL CICLO CELULAR
I.7.1 Regulación del ciclo celular por estrógenos
Son varios los mecanismos propuestos por los cuales el E2 regula de manera normal
las proteínas involucradas en el ciclo celular y en la proliferación celular. Entre éstos
se ha propuesto la inducción de la expresión de la ciclina D1; la regulación de la
distribución subcelular de los inhibidores de ciclinas (CKIs) p21 y p27; la unión de la
ciclina D1 al receptor de estrógenos aumentando su actividad transcripcional y la
fosforilación del receptor de estrógenos por la actividad de la Ciclina A/Cdk-2 (81).
En estudios animales, E2 reduce el tiempo en que las células están en fase G1 y
promueve la transición G1/S de las células epiteliales uterinas (82).
Los efectos hormonales sobre el ciclo celular son también modulados localmente por
factores de crecimiento tanto en endometrio normal (83, 84) como en células de
carcinoma endometrial (85). Sin embargo, en los endometrios de pacientes SOP no
se conoce el efecto de los estrógenos sobre reguladores del ciclo celular, ni tampoco
el efecto de los andrógenos e insulina.
I.7.2 Otros reguladores del ciclo celular
I.7.2.1. Bcl-2 y Bax
El proto-oncogén bcl-2 está implicado en controlar el ciclo celular junto con otros
miembros de la familia Bcl, tales como bax, bcl-X-long (bcl-XL), y bcl-X-short (bcl-Xs).
Bcl-2 funciona prolongando la supervivencia de las células normales y patológicas
bloqueando la apoptosis (86), y su acción es opuesta a la de Bax (62).

Introducción
21
La sobreexpresión de Bcl-2 se asociaría con transformación neoplásica (87, 88). En
este contexto, se ha postulado que E2 unido a su receptor regula la expresión
endometrial de la proteína anti-apoptótica Bcl-2 (89). Esto es especialmente
importante pues se ha descrito que durante y después de la transformación
neoplásica, la proliferación celular excede a la muerte celular, aunque es muy posible
que la apoptosis aumente temporalmente en un estadio temprano del proceso (90).
Así, se ha descrito que la expresión de Bcl-2 varía durante la transformación celular
en varios tumores humanos (91), siendo los niveles de Bcl-2 sostenidos en lesiones
pre-malignas y disminuidos luego del cambio a la malignidad. La regulación negativa
de Bcl-2 se asocia con disminución de p21 y aumento de la proteína Rb en su forma
fosforilada, siendo estos parámetros indicadores potenciales de los cambios
malignos.
Bcl-2 puede actuar como oncogén (92), pero la acción de Bcl-2 en endometrio
humano es compleja debido a la co-acción con otros miembros de la famila Bcl-2.
Por otro lado, Bax que funciona como una proteína supresora de tumores (93) se
expresa en hiperplasia endometrial mientras que está ausente en el 20,7% de los
carcinomas endometriales; esta pérdida en la expresión se asocia con mutación en el
gen bax (94).
Es importante recordar que los miembros de la familia Bcl-2 a la cual pertenecen
tanto Bcl-2 como Bax, requieren formar homo- o heterodímeros para cumplir con su
función reguladora de la viabilidad celular.

Introducción
22
I.7.2.2. Ras
La proteína Ras, producto del proto-oncogen ras, participa en la vía de señalización
de insulina y factores de crecimiento. Dichos factores al unirse a su receptor tirosina
quinasa, activan a diversas proteínas entre las que se encuentra Ras. La función de
Ras está regulada por la unión a GTP y su actividad GTPasa. Ras unido a GTP
transmite señales, pero al hidrolizar en GDP + Pi, se inactiva. Ras-GTP induce la
activación, finalmente, de MAPK que fosforilan a factores transcripcionales
aumentando la transcripción de ciertos genes que regulan la proliferación celular. Lo
interesante es que en 10 a 20% de los carcinomas endometriales dependientes de
estrógenos se han reportado mutaciones heterocigotas u homocigotas del proto-
oncogen ras convirtiéndolo en oncogen, el cual codifica para una proteína defectuosa
que conduce a una señal de proliferación continua en los tejidos. Dado que estas
mutaciones han sido descritas también en hiperplasia endometrial, se puede pensar
en que la mutación constituye un evento temprano en el desarrollo de carcinoma
endometrial (95).
I.7.2.3. Vía de la fosfatidil inositol-3-kinasa (PI3K)
Dentro de la vía PI3K, es importante la acción de la proteína supresora de tumores
PTEN (Phosphatase and Tensin homologue deleted on chromosome Ten) (96) por su
acción reguladora de la proliferación celular y de la adhesión intercelular. Sus
efectos descritos son disminuir los niveles de Ciclina D1, aumentar los del inhibidor
p27 (97), y activar moléculas pro-apoptóticas a través de vías dependientes e
independientes de Akt (75). La proteína PTEN posee un dominio de tirosina

Introducción
23
fosfatasa y también presenta características comunes con la tensina, péptido
relacionado con los filamentos de actina e implicado en mecanismos de adhesión
intercelular. Por su función fosfatasa, podría contrarrestar la acción de quinasas, y
regular la proliferación celular y por su homología con tensina, podría servir de
puente entre el citoesqueleto y proteínas de membranas como integrinas o
receptores para factores de crecimiento o insulina. De manera importante, esta
interacción está ayudando a que la célula reconozca la matriz extracelular del tejido
al cual pertenece. Las mutaciones homocigotas de este gen codifican una proteína
anormal con las funciones descritas alteradas. En hiperplasia endometrial se han
detectado mutaciones en el gen de PTEN (33,3%), lo mismo que en carcinoma
endometrial (45,8%) que pueden constituir eventos tempranos de la carcinogénesis
endometrial (98-100).

Hipótesis y Objetivos generales
24
II. HIPOTESIS
El ambiente hormonal presente en endometrios de mujeres SOP regula la expresión
de marcadores del ciclo celular, y altera factores que modulan la supervivencia de
células endometriales, creando un ambiente favorable para el desarrollo de
hiperplasia endometrial.
III. OBJETIVOS GENERALES
1- Evaluar en endometrios de mujeres SOP si la expresión de receptores
esteroidales y moléculas reguladoras del ciclo celular, así como el grado de
proliferación y apoptosis, se relaciona con el desarrollo de hiperplasia
endometrial.
2- Determinar si la condición in vitro hiperandrogénica y/o hiperestrogénica y/o
hiperinsulínica contribuyen a modificar la expresión de marcadores del ciclo
celular compatibles con hiperplasia endometrial.

Objetivos Específicos
25
IV. OBJETIVOS ESPECIFICOS
1- Determinar la expresión del mRNA de receptores esteroidales (REα, REβ y
RA) en endometrios obtenidos durante la fase proliferativa de mujeres
normales, endometrios de pacientes SOP, endometrios de mujeres SOP con
hiperplasia endometrial, y endometrios no SOP con hiperplasia (RT-PCR).
Comparar los resultados obtenidos con la expresión proteica de dichos
receptores en los cuatro grupos de pacientes estudiadas (Western Blot: WB,
y/o Inmunohistoquímica: IHQ).
2- Investigar si la expresión del co-activador AIB1 difiere en los tejidos
endometriales obtenidos de los distintos grupos en estudio (IHQ). Relacionar
dicha expresión con la de receptores esteroidales encontrada en esos mismos
tejidos.
3- Establecer el grado de apoptosis en base a la fragmentación del DNA
(TUNEL) y a la expresión de Bcl-2, Bax, Caspasa-3 activa (WB), así como el
grado de proliferación celular (IHQ de Ki67) en los cuatro grupos de pacientes
estudiadas.
4- Analizar la expresión proteica de Ras, PTEN, Akt total, Akt-P y de reguladores
del ciclo celular (Ciclina E, Cdk-2, Rb, p27 y p21), así como la actividad de
Cdk-2 en los grupos de pacientes mencionados anteriormente (WB y/o IHQ).
5- Relacionar los cambios en los niveles de expresión de reguladores del ciclo
celular, Ras, PTEN, Akt total y Akt-P, con la expresión de RE, RA, del co-
activador AIB1, índice de proliferación y apoptosis en los grupos en estudio.

Objetivos Específicos
26
6- Evaluar si en cultivos de explantes de endometrios de fase proliferativa
obtenidos de mujeres normales, la expresión de AIB1, y de reguladores del
ciclo celular, se modifica en presencia de una condición hiperandrogénica y/o
hiperestrogénica y/o hiperinsulínica in vitro (WB y/o IHQ).

Materiales y Métodos
27
V. MATERIALES Y METODOS
V.1 OBTENCION DE LAS MUESTRAS
Los endometrios fueron obtenidos de cuatro grupos de mujeres, previo
consentimiento escrito e informado. Este proyecto fue aprobado por los Comités de
Ética de la Facultad de Medicina de la Universidad de Chile, y del Hospital Clínico
San Borja Arriarán, Área Central de la Salud.
V.1.1 Controles Sanos (EN)
Se utilizaron como controles, voluntarias cíclicas normales en fase proliferativa del
ciclo menstrual (n = 19), con fertilidad probada, sin terapia con anticonceptivos
hormonales por lo menos tres meses antes de la obtención de la muestra, sometidas
a salpingoligadura, biopsia o histerectomía por causas no neoplásicas en el Servicio
de Ginecología y Obstetricia del Hospital Clínico San Borja-Arriarán, Campus Centro,
Facultad de Medicina, Universidad de Chile. La razón por la cual se utilizaron estos
endometrios como controles, se debió a que los endometrios obtenidos de pacientes
SOP, son comparables morfológicamente a endometrios normales de fase
proliferativa.
V.1.2 Pacientes SOP (ESOP)
Se incluyeron en total 28 pacientes entre 25-27 años con SOP (sin tratamiento
hormonal ni farmacológico) que asistieron al Consultorio de Infertilidad y Gineco-

Materiales y Métodos
28
Endocrinología del Hospital Clínico San Borja-Arriarán. Las biopsias de estas
pacientes fueron obtenidas mediante Pipelle.
Los criterios de inclusión fueron los que se mencionan a continuación, teniendo en
cuenta que se requieren al menos dos de estas tres características para establecer el
diagnóstico del síndrome: oligo y/o anovulación, signos clínicos (acné, hirsutismo,
seborrea, sin virilización) y/o bioquímicos (testosterona total >0,6 ng/ml, índice
andrógenos libres >4,5) de hiperandrogenismo y ovarios poliquísticos a la
ultrasonografía (9).
Algunas de las pacientes presentaron, además, signos clínicos de hiperinsulinemia
(obesidad androide: índice masa corporal >27, índice cintura/cadera >88 cm, con o
sin acantosis nigricans) e insulinemias indicativas de insulino- resistencia (insulinemia
2 h post sobrecarga de glucosa >60 UI/ml y glicemia/insulinemia en ayunas <4,5). Es
importante destacar que 38% de estas mujeres SOP presentaron insulino-resistencia.
Se excluyeron pacientes con PRL >35 ng/ml, testosterona total >2 ng/ml, DHEAS
>4000 µg/ml, 17-OH progesterona >2,5 ng/ml, TSH >5 mUI/l o con diabetes franca.
Las mediciones hormonales fueron determinadas por RIA o IRMA convencionales.
V.1.3 Pacientes SOP con Hiperplasia Endometrial (ESOPH)
Las muestras (n = 14) fueron obtenidas mediante biopsia endometrial de pacientes
SOP. El diagnóstico de hiperplasia endometrial fue confirmado por estudios
histológicos realizados por un anátomo-patólogo.

Materiales y Métodos
29
El 29% de estas mujeres SOP con hiperplasia endometrial fueron insulino-
resistentes.
V.1.4 Pacientes no SOP con Hiperplasia Endometrial (EH)
Las muestras de tejido endometrial (n = 12) fueron obtenidas luego del procedimiento
quirúrgico (legrado endometrial y/o pieza de histerectomía), según el criterio médico.
Los tejidos fueron clasificados según la fecha de la última regla (FUR),
concentraciones plasmáticas de estradiol y progesterona, y mediante estudio
histológico de acuerdo a los criterios de Noyes (3), el cual fue realizado por un
anátomo-patólogo con experiencia en este tipo de tejido. Posteriormente, se
procedió al desarrollo de los estudios ex vivo e in vitro, como se detalla a
continuación:
V.2 ESTUDIOS EX VIVO
Una vez obtenidos los tejidos endometriales de los cuatro grupos de pacientes, una
fracción (t = 0) fue congelada en N2 líquido y guardada a -80ºC (estudios de
expresión proteica y/o génica, y estudio de actividad de Cdk-2). Otra fracción fue
incluida en parafina para el análisis morfológico, y para los estudios
inmunohistoquímicos y de apoptosis (TUNEL).

Materiales y Métodos
30
V.3 ESTUDIOS IN VITRO
En el caso del grupo EN, el resto del tejido fue procesado inmediatamente para
cultivos de explantes de endometrio. Cada situación experimental se realizó en
duplicado o triplicado.
V.4 DISEÑO EXPERIMENTAL
V.5 CULTIVOS DE EXPLANTES DE TEJIDOS
Se realizaron cultivos de explantes de endometrio (de mujeres correspondientes al
grupo EN) de 20-30 mg (peso húmedo). Los mismos fueron cultivados durante 18 h
Biopsia de endometrio
Explantes T0 Cultivo de explantes
Análisis Morfológico
Expresión génica
Expresión proteica
Proliferación
Apoptosis
Actividad de Cdk-2
Dating histológico
Androstenediona Estradiol Insulina
x 18h
Expresión proteica

Materiales y Métodos
31
a 37ºC y 5% CO2/aire, en medio DMEM-F12 suplementado con albúmina bovina,
transferrina y selenio, antibióticos, glutamax, ácido ascórbico, sin rojo fenol y sin
insulina. Los explantes fueron no tratados (condición basal) o tratados con:
Androstenediona (A2: 10-6 - 10-5 mol/l), 17β-estradiol (E2:10-8 - 10-6 mol/l), Insulina
(10-7 - 10-6 mol/l) y combinaciones hormonales de A2 (10-6 mol/l) + E2 (10-6 mol/l) +
Insulina (10-6 mol/l).
El tiempo de incubación de 18 h fue establecido en estudios anteriores de nuestro
grupo en los cuales se determinó que se mantiene la arquitectura tisular. Las
concentraciones de hormonas utilizadas se encuentran dentro de rangos fisiológicos
y fueron establecidas en protocolos anteriores realizados por nuestro grupo (101) y
están basadas, además, en datos obtenidos por otros autores (41, 70, 102).
Una vez finalizada la incubación, los trozos de tejidos fueron congelados en N2
líquido y/o incluidos en parafina para los estudios de expresión proteica.
La realización del estudio in vitro es fundamental para establecer una relación causal
entre las hormonas involucradas en SOP (andrógenos, estrógenos e insulina) y la
modificación de los marcadores a estudiar.
V.6 ANALISIS MORFOLOGICO
En todas las muestras de tejido de las pacientes estudiadas (tanto para los estudios
ex vivo como in vitro) se realizó la tinción morfológica de rutina hematoxilina-eosina
para evaluar la arquitectura del tejido.

Materiales y Métodos
32
V.7 ESTUDIOS DE EXPRESION GENICA
V.7.1 Reacción de polimerización en cadena de DNA complementario (RT-PCR)
El RNA total se preparó utilizando TRIzol (Gibco BRL, Life Technology) y el DNA
complementario (cDNA) se sintetizó a partir de RNA tratado con DNAasa I, utilizando
la enzima SuperScript II Transcriptasa Reversa (Gibco BRL, Life Technology). De
100 a 300 ng de cDNA fueron amplificados utilizando los primers específicos de cada
gen a estudiar:
RA : ATG GCT GTC ATT CAG TAC TCC TGG A
AGA TGG GCT TGA CTT TCC CAG AAA G
REα : AGG CTG CGC GTT CGG C
AGC CAT ACT TCC CTT GTC AT
REβ : TTC CCA GCA ATG TCA CTA ACT
CTC TTT GAA CCT GGA CCA GTA
El cDNA fue amplificado en una mezcla que contenía:
RA: Buffer PCR, MgCl2 3,3 mM, dNTP 0,27 mM, 27 mU/ μl Taq polimerasa (Gibco
BRL, Life Technology), 0,87 μM de mezcla primer de RA.
REα: Buffer PCR, MgCl2 2,17 mM, desoxinucleótidos (dNTP) 0,16 mM, 65 mU/μl de
Taq polimerasa (Gibco BRL, Life Technology), 0,87 μM de mezcla primer de REα.

Materiales y Métodos
33
REβ: Buffer PCR, MgCl2 3,3 mM, dNTP 0,27 mM, 27 mU/μl Taq polimerasa (Gibco
BRL, Life Technology), 0,87 μM de mezcla primer de RE
Las condiciones de PCR fueron las siguientes:
RA: 30 ciclos de 1 min a 94°C de denaturación, 2 min a 55°C de anneling y 3 min a
72°C de extensión.
REα: 30 ciclos de 45 seg a 94°C de denaturación, 1min a 55°C de anneling y 1min
30 seg a 72°C de extensión.
REβ: 40 ciclos de 45 seg a 94°C de denaturación, 1min 30 seg a 55°C de anneling y
1min 30 seg a 72°C de extensión.
Los productos fueron separados en geles de agarosa y en todas las muestras se
amplificó un control interno de GAPDH (Glyceraldehyde-3-phosphate
dehydrogenase). Los resultados se calcularon como la razón entre el mRNA del gen/
mRNA de GAPDH y fueron expresados en unidades arbitrarias (U.A.).
V.8 ESTUDIOS DE EXPRESION PROTEICA
V.8.1 SDS-PAGE y Análisis de Western Blot
Trozos de tejido congelados de las pacientes estudiadas (estudios ex vivo e in vitro)
fueron homogenizados y lisados a 4°C usando un buffer de lisis conteniendo: Hepes

Materiales y Métodos
34
20 mM, EDTA 2 mM EGTA 2 mM, Triton 1%, PMSF 5 mM, Na3VO4 50 μM (Sigma),
además de un cocktail de inhibidores de proteasas (Roche, IN).
Luego de la centrifugación a 10,000 g por 20 min a 4°C, se determinó la
concentración de proteínas usando BCA protein assay kit (Pierce, IL). Los extractos
proteicos (50 µg) fueron sometidos a electroforesis en geles al 7,5% para el caso de
RA, REα, Akt total, Akt-P Ser473 y Akt-P Thr308; al 15% para Ras, PTEN, Bcl-2,
Bax, Pro-Caspasa-3, Caspasa-3, y además, se utilizaron geles en gradiente de
poliacrilamida (4-20 %) para el caso de Cdk-2, ciclina E, p27, p21 y Rb. Una vez
finalizada la corrida, los geles fueron electrotransferidos a membranas de PDVF.
Luego de la transferencia, las membranas fueron bloqueadas y a continuación
incubadas con el primer anticuerpo tal como se indica en la Tabla 1. Luego, las
membranas fueron lavadas en TTBS/ 0,1 % Tween 20 e incubadas con el segundo
anticuerpo correspondiente (Tabla 1) durante 1h a temperatura ambiente (TA).
Posteriormente, las membranas fueron lavadas nuevamente, y los anticuerpos
secundarios unidos a la membrana fueron detectados por quimioluminiscencia. Los
niveles proteicos fueron analizados por estudios densitométricos usando un
programa UN-SCAN-IT (Versión 5.1). Los resultados de Western Blot se expresan
como la razón entre la intensidad de la banda de la proteína en estudio y la de β-
Actina, como control interno.
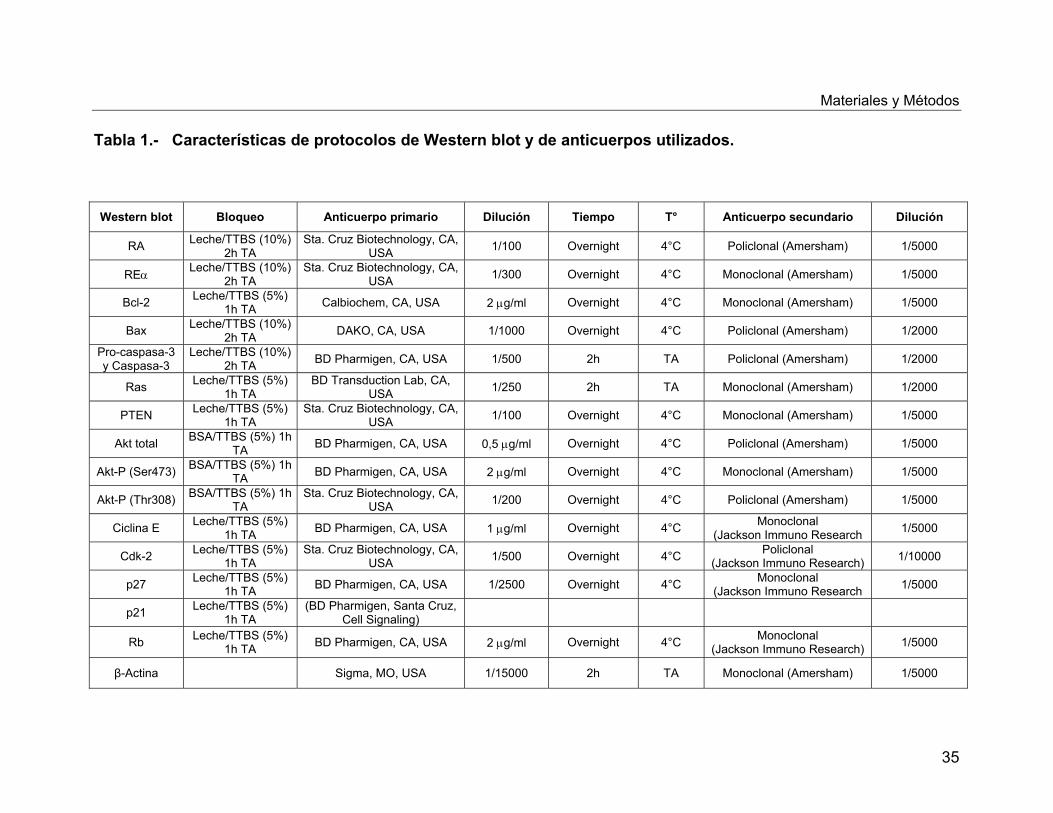
Materiales y Métodos
35
Tabla 1.- Características de protocolos de Western blot y de anticuerpos utilizados.
Western blot Bloqueo Anticuerpo primario Dilución Tiempo T° Anticuerpo secundario Dilución
RA Leche/TTBS (10%) 2h TA
Sta. Cruz Biotechnology, CA, USA 1/100 Overnight 4°C Policlonal (Amersham) 1/5000
REα Leche/TTBS (10%) 2h TA
Sta. Cruz Biotechnology, CA, USA 1/300 Overnight 4°C Monoclonal (Amersham) 1/5000
Bcl-2 Leche/TTBS (5%) 1h TA Calbiochem, CA, USA 2 μg/ml Overnight 4°C Monoclonal (Amersham) 1/5000
Bax Leche/TTBS (10%) 2h TA DAKO, CA, USA 1/1000 Overnight 4°C Policlonal (Amersham) 1/2000
Pro-caspasa-3 y Caspasa-3
Leche/TTBS (10%) 2h TA BD Pharmigen, CA, USA 1/500 2h TA Policlonal (Amersham) 1/2000
Ras Leche/TTBS (5%) 1h TA
BD Transduction Lab, CA, USA 1/250 2h TA Monoclonal (Amersham) 1/2000
PTEN Leche/TTBS (5%) 1h TA
Sta. Cruz Biotechnology, CA, USA 1/100 Overnight 4°C Monoclonal (Amersham) 1/5000
Akt total BSA/TTBS (5%) 1h TA BD Pharmigen, CA, USA 0,5 μg/ml Overnight 4°C Policlonal (Amersham) 1/5000
Akt-P (Ser473) BSA/TTBS (5%) 1h TA BD Pharmigen, CA, USA 2 μg/ml Overnight 4°C Monoclonal (Amersham) 1/5000
Akt-P (Thr308) BSA/TTBS (5%) 1h TA
Sta. Cruz Biotechnology, CA, USA 1/200 Overnight 4°C Policlonal (Amersham) 1/5000
Ciclina E Leche/TTBS (5%) 1h TA BD Pharmigen, CA, USA 1 μg/ml Overnight 4°C Monoclonal
(Jackson Immuno Research 1/5000
Cdk-2 Leche/TTBS (5%) 1h TA
Sta. Cruz Biotechnology, CA, USA 1/500 Overnight 4°C Policlonal
(Jackson Immuno Research) 1/10000
p27 Leche/TTBS (5%) 1h TA BD Pharmigen, CA, USA 1/2500 Overnight 4°C Monoclonal
(Jackson Immuno Research 1/5000
p21 Leche/TTBS (5%) 1h TA
(BD Pharmigen, Santa Cruz, Cell Signaling)
Rb Leche/TTBS (5%) 1h TA BD Pharmigen, CA, USA 2 μg/ml Overnight 4°C Monoclonal
(Jackson Immuno Research) 1/5000
β-Actina Sigma, MO, USA 1/15000 2h TA Monoclonal (Amersham) 1/5000

Materiales y Métodos
36
V.8.2 Estudio inmunohistoquímico
Se realizó en cortes incluidos en parafina de endometrios de los cuatro grupos de
pacientes estudiadas, y en los endometrios en fase proliferativa utilizados para los
ensayos in vitro. En todos ellos se determinó la expresión de las proteínas REα,
REβ, SRC-3/AIB1, P450 aromatasa, Ras, PTEN, Cdk-2, Ciclina E, y p27, y en el caso
de los estudios in vitro solo se realizó inmunodetección para SRC-3/AIB1.
Para ello, los tejidos fueron desparafinados y rehidratados. Para la recuperación de
antígenos, los cortes se incubaron 20 min en baño de agua entre 95-100º C en una
solución de citrato de sodio 10 mM, pH 6,0, excepto para REβ donde se utilizó un
buffer glicina 0,05 M, EDTA 0,01% pH 3,5, y en el caso de P450 aromatasa no se
necesitó realizar el procedimiento de recuperación antigénica. Para evitar la
actividad peroxidasa endógena, en la detección de REα, Ras y PTEN se realizó un
tratamiento con peróxido de hidrógeno al 3% por 5 min. Por otro lado, se realizó un
tratamiento durante 30 min con peróxido de hidrógeno al 3% en metanol para REβ y
P450 aromatasa, al 3% en agua para Ciclina E, al 0,5% en agua para p27 y Cdk-2, y
al 0,3% en etanol absoluto para SRC-3/AIB1. Posteriormente, los cortes se
incubaron en PBS-BSA al 4% durante 1 h para evitar uniones inespecíficas, y luego,
con los respectivos anticuerpos, de acuerdo a los protocolos que se muestran en la
Tabla 2.
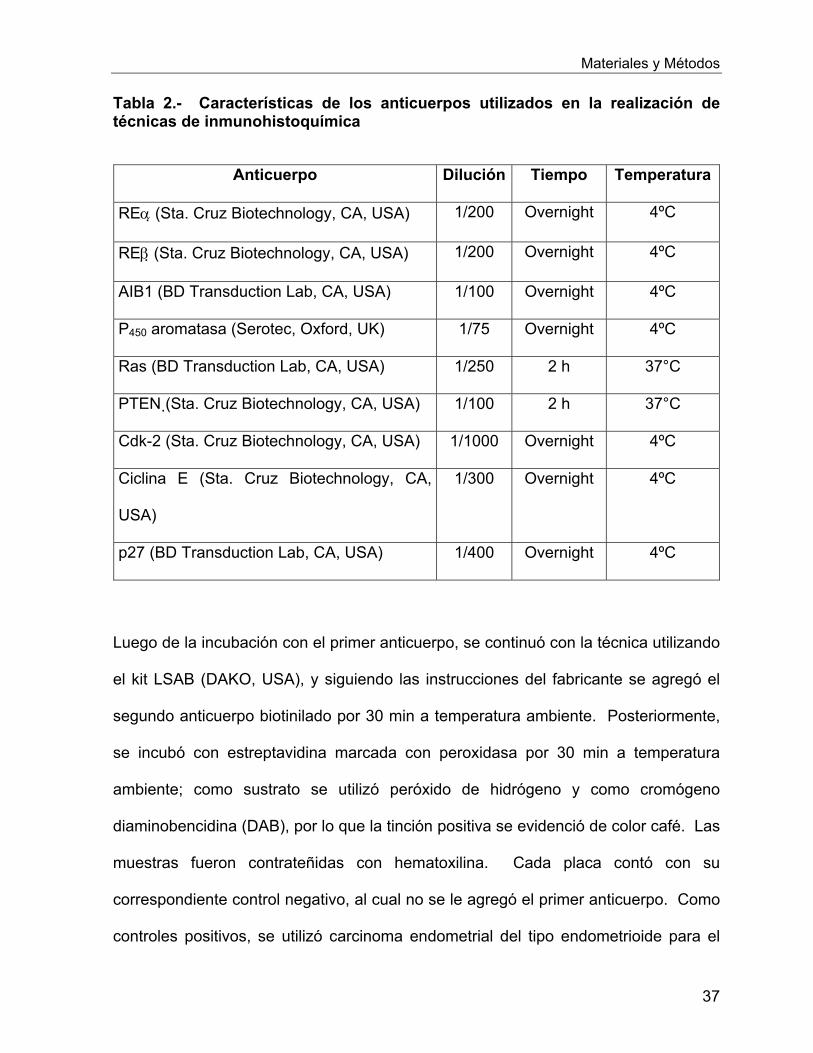
Materiales y Métodos
37
Tabla 2.- Características de los anticuerpos utilizados en la realización de técnicas de inmunohistoquímica
Anticuerpo Dilución Tiempo Temperatura
REα (Sta. Cruz Biotechnology, CA, USA) 1/200 Overnight 4ºC
REβ (Sta. Cruz Biotechnology, CA, USA) 1/200 Overnight 4ºC
AIB1 (BD Transduction Lab, CA, USA) 1/100 Overnight 4ºC
P450 aromatasa (Serotec, Oxford, UK) 1/75 Overnight 4ºC
Ras (BD Transduction Lab, CA, USA) 1/250 2 h 37°C
PTEN (Sta. Cruz Biotechnology, CA, USA) 1/100 2 h 37°C
Cdk-2 (Sta. Cruz Biotechnology, CA, USA) 1/1000 Overnight 4ºC
Ciclina E (Sta. Cruz Biotechnology, CA,
USA)
1/300 Overnight 4ºC
p27 (BD Transduction Lab, CA, USA) 1/400 Overnight 4ºC
Luego de la incubación con el primer anticuerpo, se continuó con la técnica utilizando
el kit LSAB (DAKO, USA), y siguiendo las instrucciones del fabricante se agregó el
segundo anticuerpo biotinilado por 30 min a temperatura ambiente. Posteriormente,
se incubó con estreptavidina marcada con peroxidasa por 30 min a temperatura
ambiente; como sustrato se utilizó peróxido de hidrógeno y como cromógeno
diaminobencidina (DAB), por lo que la tinción positiva se evidenció de color café. Las
muestras fueron contrateñidas con hematoxilina. Cada placa contó con su
correspondiente control negativo, al cual no se le agregó el primer anticuerpo. Como
controles positivos, se utilizó carcinoma endometrial del tipo endometrioide para el

Materiales y Métodos
38
caso del REα y SRC-3/AIB1, epididimo humano para REβ y cuerpo lúteo humano
para P450 aromatasa.
Las placas se evaluaron por tres observadores independientes mediante porcentaje
de células teñidas obtenidas del conteo de 1000 células e intensidad (i) de la tinción
evaluada como 1 (leve), 2 (moderada), o 3 (intensa). Para el cálculo se aplicó la
siguiente fórmula descrita por Lessey (103):
HS=Σ % células teñidas x (i +1)/100
V.9 ESTUDIO DE ACTIVIDAD DE CDK-2
Los extractos fueron preparados usando el siguiente buffer de lisis: Tris-HCl 50 mM
pH 7,5, NaCl 150 mM, EDTA 20 mM, NP40 0,5%, DTT 1mM, Aprotinina 2 µg/ml,
Leupeptina 2 µg/ml, Pepstatina 2 µg/ml, PMSF 1mM, NaF 1mM, β-glicerofosfato
1mM, Na3VO4 0,2 mM. Luego del proceso de homogenización, los extractos fueron
centrifugados en una microcentrífuga a 16,000 g a 4°C por 20 min. La concentración
de proteínas fue determinada usando BCA kit. A continuación, los lisados (100 µg de
proteína), fueron usados para inmunoprecipitar Cdk-2 con un anticuerpo policlonal.
Las perlas (beads) con las proteínas unidas fueron lavadas y luego resuspendidas en
30 µl del buffer quinasa (Tris-HCl 50 mM pH 7,5, MgCl2 15 mM, DTT 2 mM, 20 µM
ATP, 10 µCi [γ-32P] ATP 10µM, Histona H1 2 µg). La mezcla de la reacción fue
incubada a 30°C por 15 min. Luego, la reacción fue detenida adicionando el mismo
volumen de Buffer Laemmli 2 X. La histona fosforilada fue resuelta por SDS-PAGE y
detectada por autoradiografía usando Kodak film.

Materiales y Métodos
39
V.10 ESTUDIOS DE APOPTOSIS
V.10.1 Método fluorescente de detección de apoptosis (TUNEL)
Fue realizado mediante el sistema DeadEndTM Fluorometric TUNEL System
(Promega, WI, USA). Este consiste en la incorporación de 12-dUTP marcado con
FITC al extremo 3’ OH de la mólecula de DNA mediante la enzima deoxinucleotidil
transferasa (TdT), con lo cual se puede detectar la presencia de fragmentación del
DNA. Los cortes histológicos luego de ser desparafinados y rehidratados, fueron
tratados con proteinasa K 20 μg/ml por 10 min, lavados en PBS por 10 min e
incubados a temperatura ambiente por 1 h en cámara húmeda, con la mezcla de la
enzima TdT, los deoxinucleótidos marcados con FITC y el buffer de equilibro en las
proporciones indicadas por los fabricantes. Al control negativo no se le adicionó la
enzima TdT. Los cortes fueron contrateñidos con yoduro de propidio y analizados en
un microscopio de fluorescencia. Se evaluó el porcentaje de células con tinción
positiva en varios campos a 400x, contándose entre 800 a 1000 células.
V.11 ESTUDIOS DE PROLIFERACION
V.11.1 Expresión de la proteína Ki67
La proteína Ki67 está presente en componentes granulares del nucleolo durante las
fases G1 tardía, S, G2 y M; por lo tanto, su expresión se encuentra restringida a
células que están proliferando.
Las muestras de tejidos de las diferentes pacientes se sometieron a un análisis
inmunohistoquímico para Ki67. Para ello, las láminas fueron tratadas con Buffer Tris

Materiales y Métodos
40
100 mM pH 9,5 por 20 min a 100º C, peróxido de hidrógeno al 0,2% en metanol por
30 min y PBS-BSA al 4% por 60 min. Posteriormente, fueron incubadas con el
anticuerpo monoclonal anti-Ki67 0,05 μg/ml (Oncogene, CA, USA) durante toda la
noche a 4º C, y luego reveladas con el kit LSAB (DAKO, USA) de acuerdo al
protocolo descrito por el fabricante. Se evaluaron varios campos a 400x por tres
observadores en forma independiente, contándose 1000 células por lámina y los
resultados se expresaron como porcentaje de células positivas.
V.12 ANALISIS ESTADISTICO
Los resultados se expresan como promedio ± EEM en todos los estudios y se
muestran, en muchos casos, como porcentaje respecto del grupo de endometrios
normales en fase proliferativa (para los estudios ex vivo), o respecto del basal (para
los estudios in vitro).
Para el análisis estadístico se utilizó una versión standard del programa SPSS. De
esta forma, primero se realizó un test de normalidad (Kolmogorov-Smirnov), a fin de
conocer la distribución de los datos. A continuación, para las comparaciones entre
los distintos grupos en estudio se utilizó la prueba de ANOVA seguido de un post-
test (Tukey), para datos paramétricos. Para datos no paramétricos se utilizó el test
de Kruskal-Wallis H.

Resultados
41
VI. RESULTADOS
VI.1 CARACTERISTICAS DE LAS PACIENTES ESTUDIADAS
Los datos obtenidos de las pacientes de los cuatro grupos estudiados se muestran
en la Tabla 3. La edad de las mujeres voluntarias participantes en este estudio varía
de acuerdo al grupo al que pertenecen: en el grupo EN, la edad fue mayor que el de
las mujeres SOP sin y con hiperplasia endometrial, debido a que las mujeres
controles fueron pacientes quienes solicitaron salpingoligadura y que en forma
paralela aceptaron donar parte de su endometrio para este estudio. El grupo de
mujeres con hiperplasia endometrial (EH) presentaron una edad mayor que el de las
mujeres SOP (ESOP y ESOPH), debido a que las primeras corresponden a
pacientes peri-menopáusicas y que consultan fundamentalmente por metrorragia. El
incremento en el índice de masa corporal (IMC) de los grupos ESOP y ESOPH, en
comparación a los grupos EN y EH, es una condición inherente al síndrome. Por otro
lado, las pacientes de los grupos ESOP y ESOPH presentaron un hiperestrogenismo
relativo (E2/P4) mayor al comparar con el grupo EN (114% y 171%, respectivamente).
El elevado índice de andrógenos libres (IAL) de las mujeres SOP (ESOP y ESOPH)
se debe a la excesiva producción ovárica de andrógenos, principalmente
testosterona total (T), y a las reducidas concentraciones sanguíneas de la globulina
que une a esteroides sexuales (SHBG). Los coeficientes de variación intra- e inter-
ensayo de las mediciones hormonales fueron, respectivamente, 7,0% y 11% para T;
4,8% y 9,2% para P4; 3,8 y 7,9% para SHBG; 2,7% y 5% para E2; y 3,2% y 6,1% para
A4.
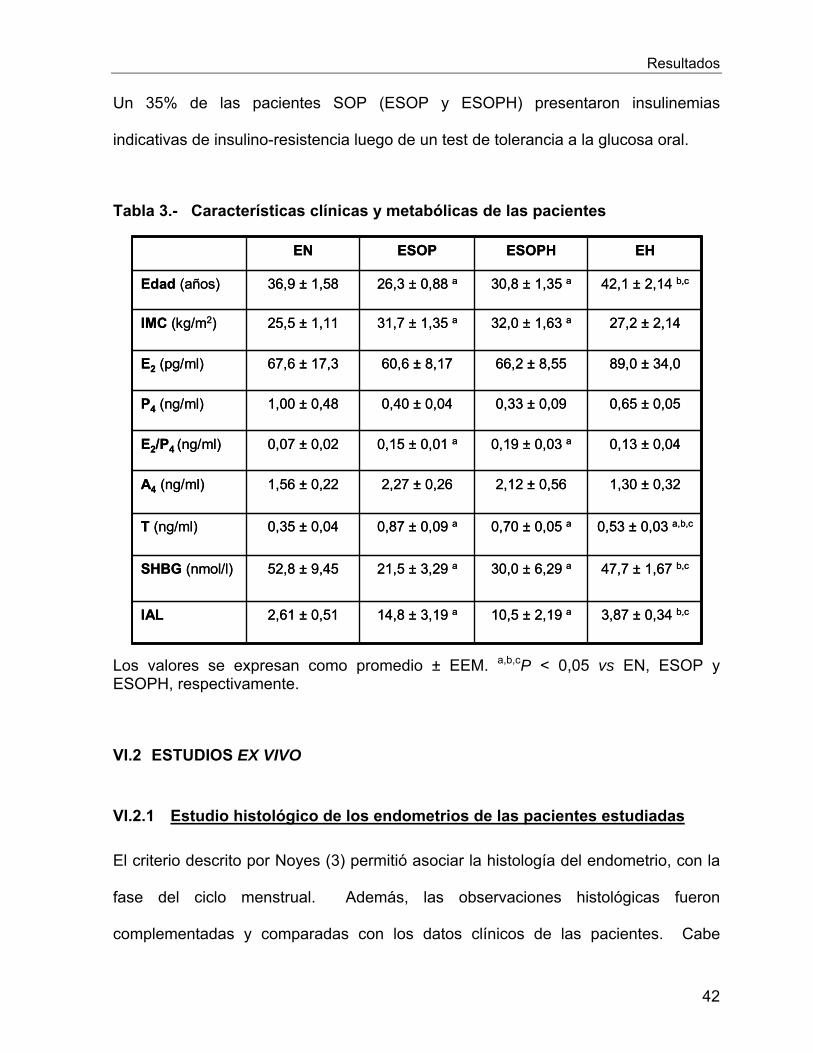
Resultados
42
Un 35% de las pacientes SOP (ESOP y ESOPH) presentaron insulinemias
indicativas de insulino-resistencia luego de un test de tolerancia a la glucosa oral.
Tabla 3.- Características clínicas y metabólicas de las pacientes
Los valores se expresan como promedio ± EEM. a,b,cP < 0,05 vs EN, ESOP y ESOPH, respectivamente.
VI.2 ESTUDIOS EX VIVO
VI.2.1 Estudio histológico de los endometrios de las pacientes estudiadas
El criterio descrito por Noyes (3) permitió asociar la histología del endometrio, con la
fase del ciclo menstrual. Además, las observaciones histológicas fueron
complementadas y comparadas con los datos clínicos de las pacientes. Cabe
0,13 ± 0,040,19 ± 0,03 a0,15 ± 0,01 a0,07 ± 0,02E2/P4 (ng/ml)
0,65 ± 0,050,33 ± 0,090,40 ± 0,041,00 ± 0,48P4 (ng/ml)
89,0 ± 34,066,2 ± 8,5560,6 ± 8,1767,6 ± 17,3E2 (pg/ml)
3,87 ± 0,34 b,c10,5 ± 2,19 a14,8 ± 3,19 a2,61 ± 0,51IAL
27,2 ± 2,1432,0 ± 1,63 a31,7 ± 1,35 a25,5 ± 1,11IMC (kg/m2)
1,30 ± 0,322,12 ± 0,562,27 ± 0,261,56 ± 0,22A4 (ng/ml)
0,53 ± 0,03 a,b,c0,70 ± 0,05 a0,87 ± 0,09 a0,35 ± 0,04T (ng/ml)
47,7 ± 1,67 b,c30,0 ± 6,29 a21,5 ± 3,29 a52,8 ± 9,45SHBG (nmol/l)
42,1 ± 2,14 b,c30,8 ± 1,35 a26,3 ± 0,88 a 36,9 ± 1,58Edad (años)
EHESOPHESOP EN
0,13 ± 0,040,19 ± 0,03 a0,15 ± 0,01 a0,07 ± 0,02E2/P4 (ng/ml)
0,65 ± 0,050,33 ± 0,090,40 ± 0,041,00 ± 0,48P4 (ng/ml)
89,0 ± 34,066,2 ± 8,5560,6 ± 8,1767,6 ± 17,3E2 (pg/ml)
3,87 ± 0,34 b,c10,5 ± 2,19 a14,8 ± 3,19 a2,61 ± 0,51IAL
27,2 ± 2,1432,0 ± 1,63 a31,7 ± 1,35 a25,5 ± 1,11IMC (kg/m2)
1,30 ± 0,322,12 ± 0,562,27 ± 0,261,56 ± 0,22A4 (ng/ml)
0,53 ± 0,03 a,b,c0,70 ± 0,05 a0,87 ± 0,09 a0,35 ± 0,04T (ng/ml)
47,7 ± 1,67 b,c30,0 ± 6,29 a21,5 ± 3,29 a52,8 ± 9,45SHBG (nmol/l)
42,1 ± 2,14 b,c30,8 ± 1,35 a26,3 ± 0,88 a 36,9 ± 1,58Edad (años)
EHESOPHESOP EN
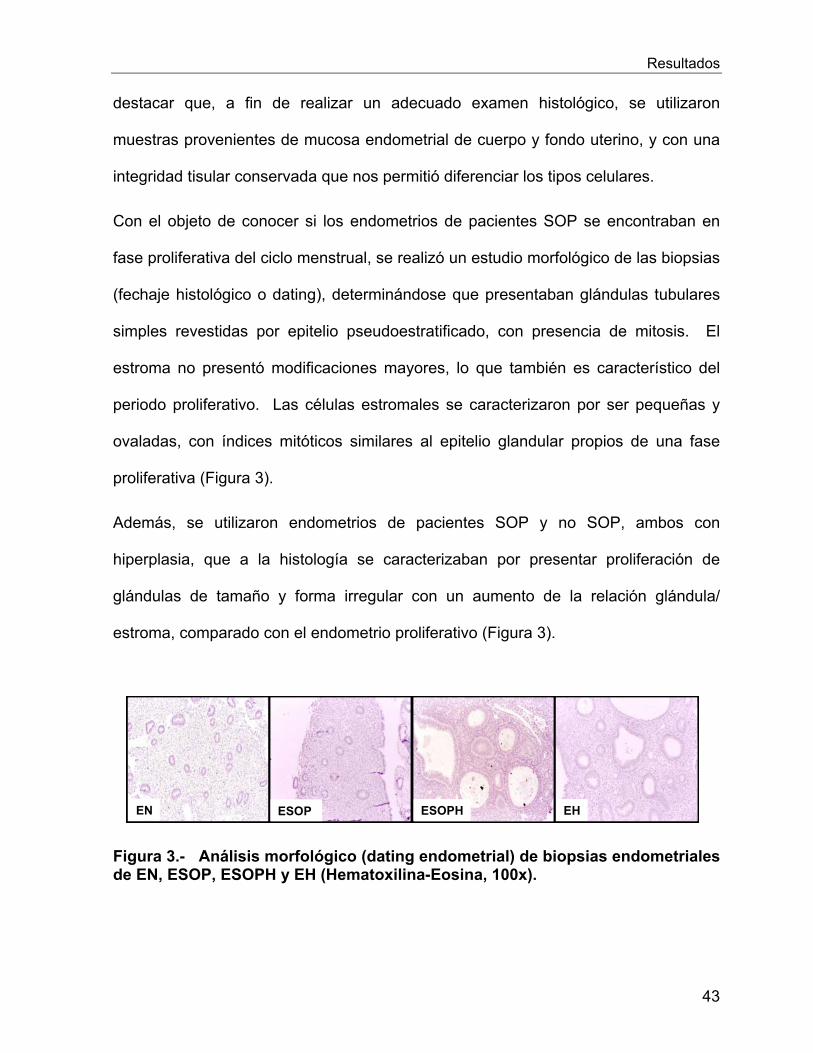
Resultados
43
destacar que, a fin de realizar un adecuado examen histológico, se utilizaron
muestras provenientes de mucosa endometrial de cuerpo y fondo uterino, y con una
integridad tisular conservada que nos permitió diferenciar los tipos celulares.
Con el objeto de conocer si los endometrios de pacientes SOP se encontraban en
fase proliferativa del ciclo menstrual, se realizó un estudio morfológico de las biopsias
(fechaje histológico o dating), determinándose que presentaban glándulas tubulares
simples revestidas por epitelio pseudoestratificado, con presencia de mitosis. El
estroma no presentó modificaciones mayores, lo que también es característico del
periodo proliferativo. Las células estromales se caracterizaron por ser pequeñas y
ovaladas, con índices mitóticos similares al epitelio glandular propios de una fase
proliferativa (Figura 3).
Además, se utilizaron endometrios de pacientes SOP y no SOP, ambos con
hiperplasia, que a la histología se caracterizaban por presentar proliferación de
glándulas de tamaño y forma irregular con un aumento de la relación glándula/
estroma, comparado con el endometrio proliferativo (Figura 3).
Figura 3.- Análisis morfológico (dating endometrial) de biopsias endometriales de EN, ESOP, ESOPH y EH (Hematoxilina-Eosina, 100x).
EHEN ESOPHESOP EHEHENEN ESOPHESOPESOP

Resultados
44
VI.2.2 Expresion génica y proteica de los receptores esteroidales en el
endometrio de las mujeres en estudio
VI.2.2.1. Expresión génica de RA, REα y REβ
Para evaluar la expresión génica de los receptores esteroidales, se realizó RT-PCR
convencional a las biopsias endometriales obtenidas de los cuatro grupos en estudio.
En dichas biopsias se amplificó un fragmento de 545-pb para RA, 200-pb para REα y
de 258-pb para REβ. Como se muestra en la Tabla 4, se observó un incremento
relativo en el mRNA para RA en ESOP (71%) y en EH (94%) comparado con
biopsias endometriales de mujeres con ciclos normales, siendo significativamente
alta su expresión en ESOPH al compararla con EN (P < 0.05). La expresión génica
para el REα no fue estadísticamente diferente entre los grupos estudiados, aunque
su expresión fue 40% mayor en ESOP vs EN. Por otro lado, se detectó un
incremento gradual del mRNA para REβ entre estos grupos (ESOP: 22%, ESOPH:
47%, y EH: 122%, respecto a EN).
Dado que las isoformas de los receptores de estrógeno pueden mediar acciones
contrapuestas del esteroide en los tejidos, se calculó la razón de la expresión génica
entre ambas isoformas. Se determinó que la razón REβ/REα fue mayor en ESOPH y
EH debido al incremento en la isoforma β, mientras que dicha razón fue menor en EN
y ESOP como consecuencia del aumento de la isoforma α (P < 0.05) (Tabla 4).
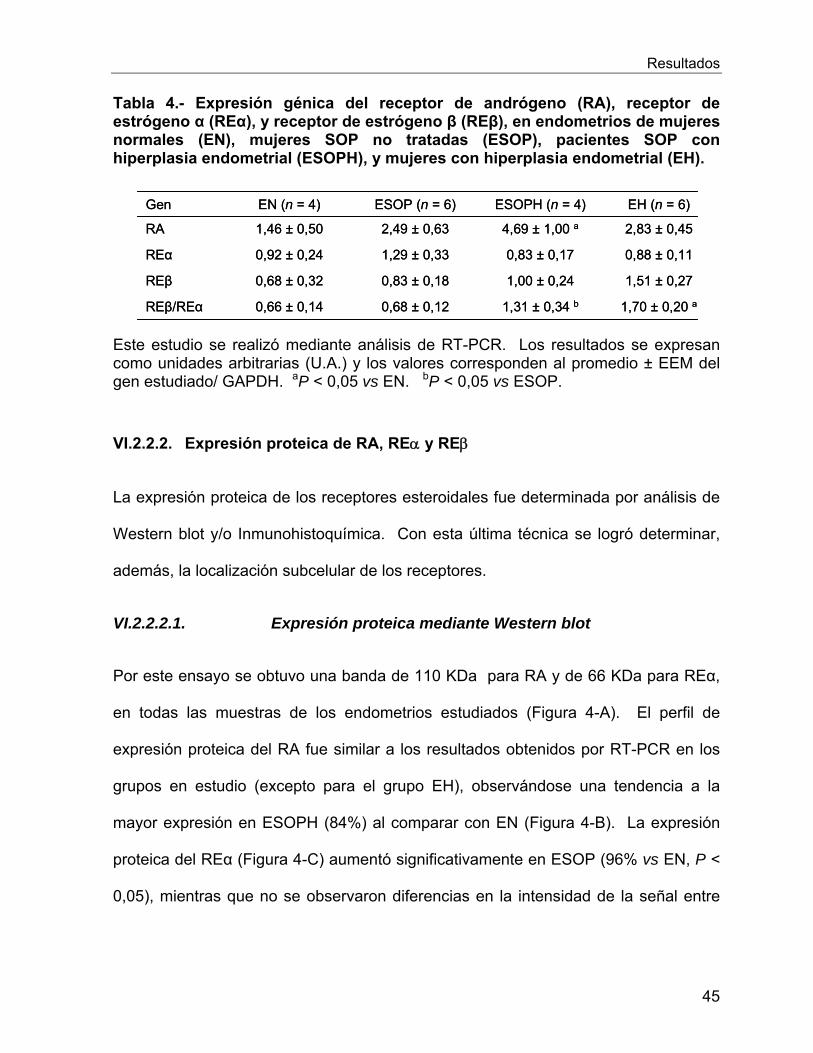
Resultados
45
Tabla 4.- Expresión génica del receptor de andrógeno (RA), receptor de estrógeno α (REα), y receptor de estrógeno β (REβ), en endometrios de mujeres normales (EN), mujeres SOP no tratadas (ESOP), pacientes SOP con hiperplasia endometrial (ESOPH), y mujeres con hiperplasia endometrial (EH).
Este estudio se realizó mediante análisis de RT-PCR. Los resultados se expresan como unidades arbitrarias (U.A.) y los valores corresponden al promedio ± EEM del gen estudiado/ GAPDH. aP < 0,05 vs EN. bP < 0,05 vs ESOP.
VI.2.2.2. Expresión proteica de RA, REα y REβ
La expresión proteica de los receptores esteroidales fue determinada por análisis de
Western blot y/o Inmunohistoquímica. Con esta última técnica se logró determinar,
además, la localización subcelular de los receptores.
VI.2.2.2.1. Expresión proteica mediante Western blot
Por este ensayo se obtuvo una banda de 110 KDa para RA y de 66 KDa para REα,
en todas las muestras de los endometrios estudiados (Figura 4-A). El perfil de
expresión proteica del RA fue similar a los resultados obtenidos por RT-PCR en los
grupos en estudio (excepto para el grupo EH), observándose una tendencia a la
mayor expresión en ESOPH (84%) al comparar con EN (Figura 4-B). La expresión
proteica del REα (Figura 4-C) aumentó significativamente en ESOP (96% vs EN, P <
0,05), mientras que no se observaron diferencias en la intensidad de la señal entre
1,51 ± 0,271,00 ± 0,240,83 ± 0,180,68 ± 0,32REβ
2,83 ± 0,454,69 ± 1,00 a2,49 ± 0,631,46 ± 0,50RA
0,88 ± 0,110,83 ± 0,171,29 ± 0,330,92 ± 0,24REα
1,70 ± 0,20 a1,31 ± 0,34 b0,68 ± 0,120,66 ± 0,14REβ/REα
EH (n = 6)ESOPH (n = 4)ESOP (n = 6)EN (n = 4)Gen
1,51 ± 0,271,00 ± 0,240,83 ± 0,180,68 ± 0,32REβ
2,83 ± 0,454,69 ± 1,00 a2,49 ± 0,631,46 ± 0,50RA
0,88 ± 0,110,83 ± 0,171,29 ± 0,330,92 ± 0,24REα
1,70 ± 0,20 a1,31 ± 0,34 b0,68 ± 0,120,66 ± 0,14REβ/REα
EH (n = 6)ESOPH (n = 4)ESOP (n = 6)EN (n = 4)Gen
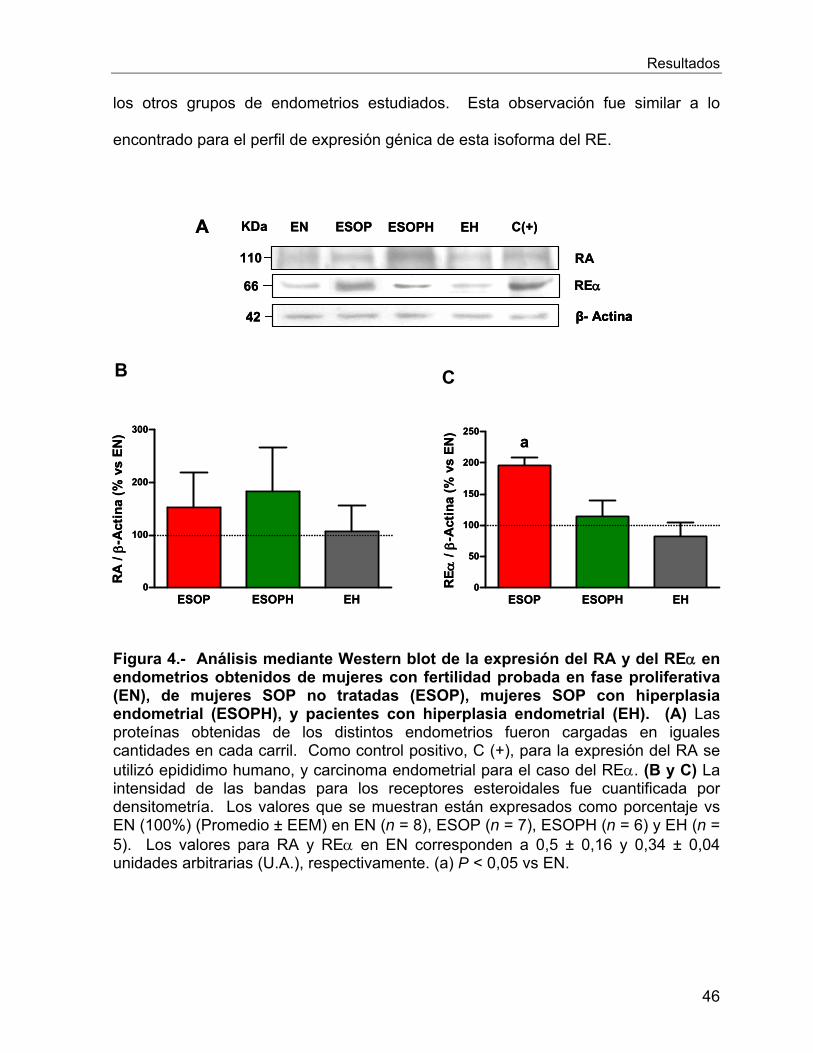
Resultados
46
los otros grupos de endometrios estudiados. Esta observación fue similar a lo
encontrado para el perfil de expresión génica de esta isoforma del RE.
Figura 4.- Análisis mediante Western blot de la expresión del RA y del REα en endometrios obtenidos de mujeres con fertilidad probada en fase proliferativa (EN), de mujeres SOP no tratadas (ESOP), mujeres SOP con hiperplasia endometrial (ESOPH), y pacientes con hiperplasia endometrial (EH). (A) Las proteínas obtenidas de los distintos endometrios fueron cargadas en iguales cantidades en cada carril. Como control positivo, C (+), para la expresión del RA se utilizó epididimo humano, y carcinoma endometrial para el caso del REα. (B y C) La intensidad de las bandas para los receptores esteroidales fue cuantificada por densitometría. Los valores que se muestran están expresados como porcentaje vs EN (100%) (Promedio ± EEM) en EN (n = 8), ESOP (n = 7), ESOPH (n = 6) y EH (n = 5). Los valores para RA y REα en EN corresponden a 0,5 ± 0,16 y 0,34 ± 0,04 unidades arbitrarias (U.A.), respectivamente. (a) P < 0,05 vs EN.
A EN ESOP ESOPH EH C(+)
REα
RA
66
110
KDa
β- Actina42
A EN ESOP ESOPH EH C(+)
REα
RA
66
110
KDa
β- Actina42 β- Actina42
B C
ESOP ESOPH EH0
100
200
300
RA
/β-
Act
ina
(% v
s EN
)
ESOP ESOPH EH0
100
200
300
RA
/β-
Act
ina
(% v
s EN
)
ESOP ESOPH EH0
50
100
150
200
250
RE α
/β-
Act
ina
(% v
s EN
) a
ESOP ESOPH EH0
50
100
150
200
250
RE α
/β-
Act
ina
(% v
s EN
) a
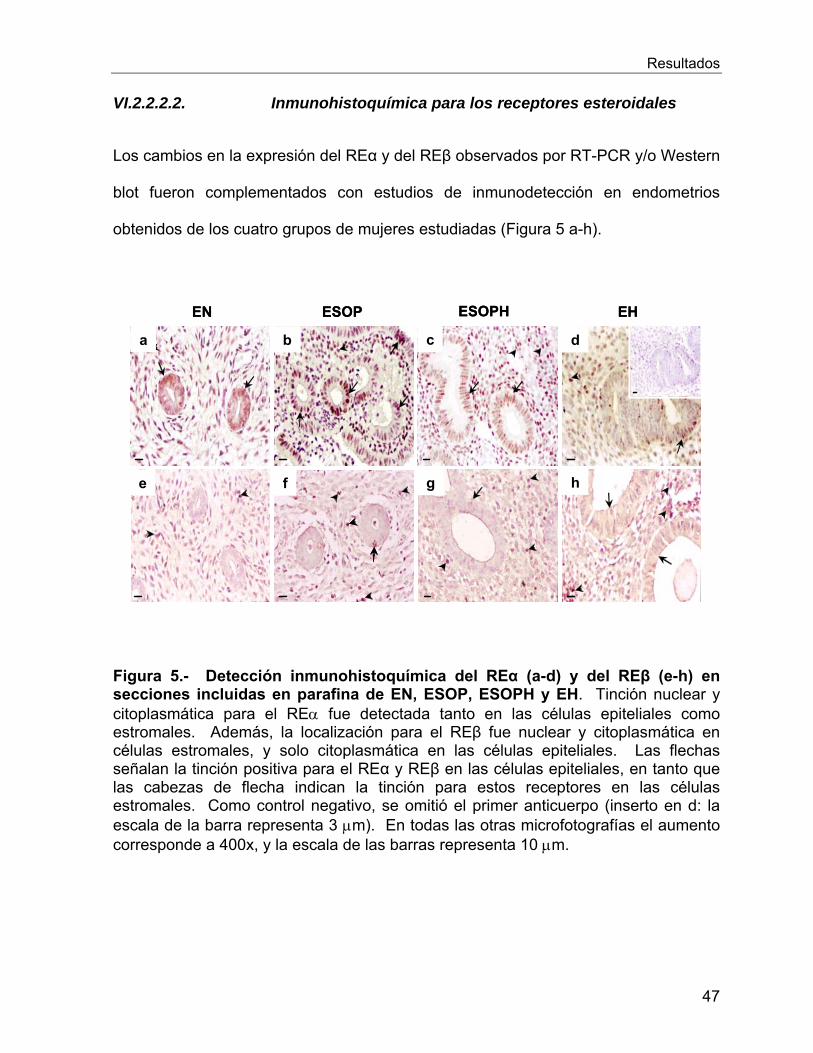
Resultados
47
VI.2.2.2.2. Inmunohistoquímica para los receptores esteroidales
Los cambios en la expresión del REα y del REβ observados por RT-PCR y/o Western
blot fueron complementados con estudios de inmunodetección en endometrios
obtenidos de los cuatro grupos de mujeres estudiadas (Figura 5 a-h).
Figura 5.- Detección inmunohistoquímica del REα (a-d) y del REβ (e-h) en secciones incluidas en parafina de EN, ESOP, ESOPH y EH. Tinción nuclear y citoplasmática para el REα fue detectada tanto en las células epiteliales como estromales. Además, la localización para el REβ fue nuclear y citoplasmática en células estromales, y solo citoplasmática en las células epiteliales. Las flechas señalan la tinción positiva para el REα y REβ en las células epiteliales, en tanto que las cabezas de flecha indican la tinción para estos receptores en las células estromales. Como control negativo, se omitió el primer anticuerpo (inserto en d: la escala de la barra representa 3 μm). En todas las otras microfotografías el aumento corresponde a 400x, y la escala de las barras representa 10 μm.
EN ESOP ESOPH EH
a b c d
e f g h
EN ESOP ESOPH EH
a b c d
e f g h

Resultados
48
La inmunotinción para el REα fue predominantemente localizada a nivel nuclear (HS:
EN = 2,78 ± 0,13; ESOP = 3,0 ± 0,10; ESOPH = 2,63 ± 0,23; EH = 2,44 ± 0,25) y
débilmente a nivel citoplasmático (HS: EN = 1,44 ± 0,16; ESOP = 1,73 ± 0,09;
ESOPH = 1,43 ± 0,18; EH = 1,71 ± 0,13) de las células endometriales de los grupos
estudiados (P < 0,05 núcleo vs citoplasma) (Figura 5 a-d). En cuanto a la isoforma β
del RE, se detectó tinción positiva principalmente en el núcleo y citoplasma de las
células estromales, y solo en el citoplasma de las células epiteliales (Figura 5 e-h).
Como se muestra en la Figura 6, mediante la evaluación semicuantitativa por
HScore, se observó que la tinción inmunohistoquímica para ambas isoformas del RE,
tuvieron un patrón similar de expresión en las células estromales del endometrio de
los cuatro grupos de pacientes (Figura 6-A); sin embargo, este patrón fue diferente
en las células epiteliales (Figura 6-B). Se observó un incremento gradual en la
expresión del REβ en el compartimiento epitelial, siendo este aumento significativo
en EH, en comparación a EN. No obstante, la expresión del REα no se modificó
significativamente a nivel epitelial entre los diferentes grupos estudiados (Figura 6-B).
Consecuentemente, la razón REβ/REα aumentó en forma significativa en el
compartimiento epitelial en ESOP (120%), ESOPH (270%) y EH (490%), al comparar
con EN. Por otro lado, la expresión del REβ fue mayor en las células estromales de
ESOP, ESOPH y EH al comparar con EN (P < 0,05) (Figura 6-A).
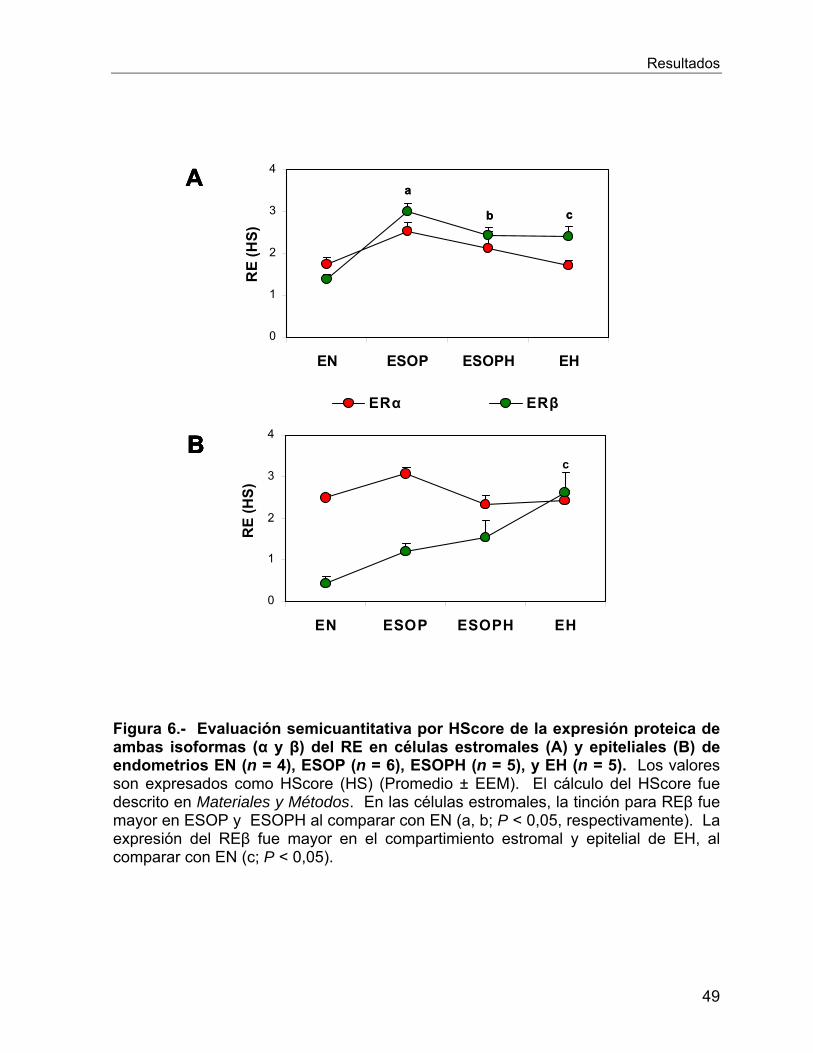
Resultados
49
Figura 6.- Evaluación semicuantitativa por HScore de la expresión proteica de ambas isoformas (α y β) del RE en células estromales (A) y epiteliales (B) de endometrios EN (n = 4), ESOP (n = 6), ESOPH (n = 5), y EH (n = 5). Los valores son expresados como HScore (HS) (Promedio ± EEM). El cálculo del HScore fue descrito en Materiales y Métodos. En las células estromales, la tinción para REβ fue mayor en ESOP y ESOPH al comparar con EN (a, b; P < 0,05, respectivamente). La expresión del REβ fue mayor en el compartimiento estromal y epitelial de EH, al comparar con EN (c; P < 0,05).
0
1
2
3
4
EN ESOP ESOPH EH
RE
(HS)
A
B
0
1
2
3
4
EN ESOP ESOPH EH
ER
(HS
)
ERα ERβ
c
a
b c
RE
(HS)
0
1
2
3
4
EN ESOP ESOPH EH
RE
(HS)
A
B
0
1
2
3
4
EN ESOP ESOPH EH
ER
(HS
)
ERα ERβ
c
a
b c
RE
(HS)
A
B
0
1
2
3
4
EN ESOP ESOPH EH
ER
(HS
)
ERα ERβ
cB
0
1
2
3
4
EN ESOP ESOPH EH
ER
(HS
)
ERα ERβ
B
0
1
2
3
4
EN ESOP ESOPH EH
ER
(HS
)
ERα ERβ
c
a
b c
RE
(HS)
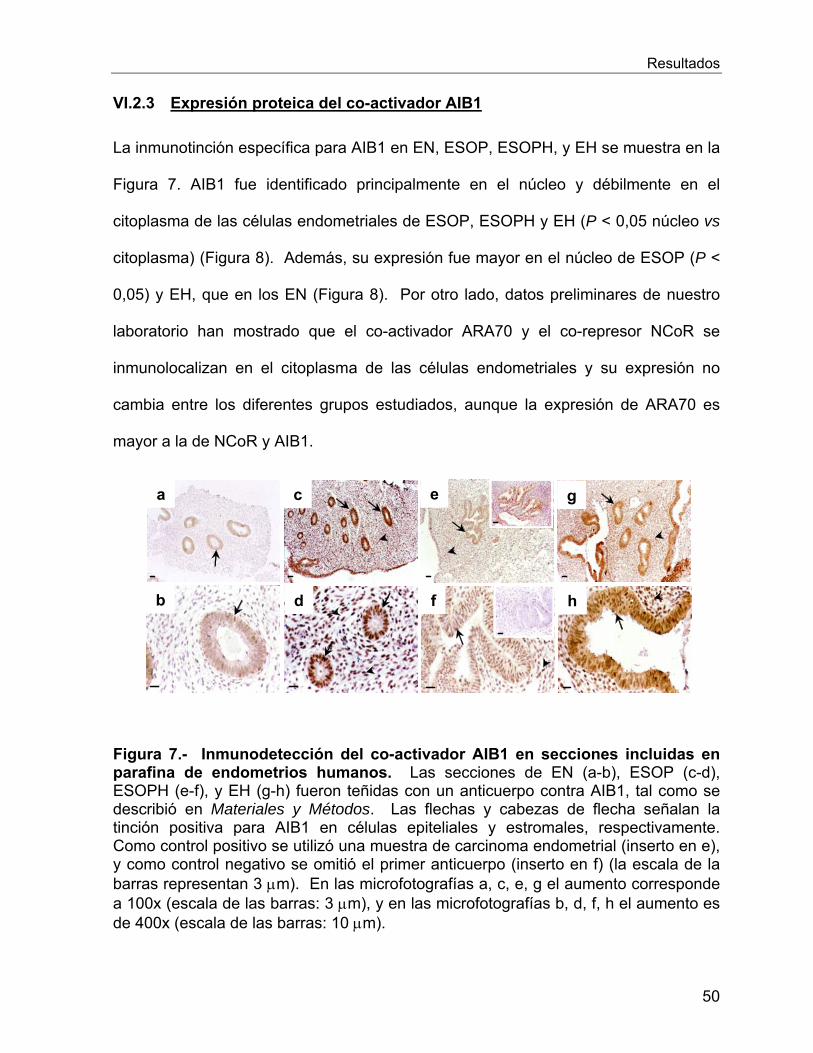
Resultados
50
VI.2.3 Expresión proteica del co-activador AIB1
La inmunotinción específica para AIB1 en EN, ESOP, ESOPH, y EH se muestra en la
Figura 7. AIB1 fue identificado principalmente en el núcleo y débilmente en el
citoplasma de las células endometriales de ESOP, ESOPH y EH (P < 0,05 núcleo vs
citoplasma) (Figura 8). Además, su expresión fue mayor en el núcleo de ESOP (P <
0,05) y EH, que en los EN (Figura 8). Por otro lado, datos preliminares de nuestro
laboratorio han mostrado que el co-activador ARA70 y el co-represor NCoR se
inmunolocalizan en el citoplasma de las células endometriales y su expresión no
cambia entre los diferentes grupos estudiados, aunque la expresión de ARA70 es
mayor a la de NCoR y AIB1.
Figura 7.- Inmunodetección del co-activador AIB1 en secciones incluidas en parafina de endometrios humanos. Las secciones de EN (a-b), ESOP (c-d), ESOPH (e-f), y EH (g-h) fueron teñidas con un anticuerpo contra AIB1, tal como se describió en Materiales y Métodos. Las flechas y cabezas de flecha señalan la tinción positiva para AIB1 en células epiteliales y estromales, respectivamente. Como control positivo se utilizó una muestra de carcinoma endometrial (inserto en e), y como control negativo se omitió el primer anticuerpo (inserto en f) (la escala de la barras representan 3 μm). En las microfotografías a, c, e, g el aumento corresponde a 100x (escala de las barras: 3 μm), y en las microfotografías b, d, f, h el aumento es de 400x (escala de las barras: 10 μm).
b d f h
a c e g
b d f h
a c e g
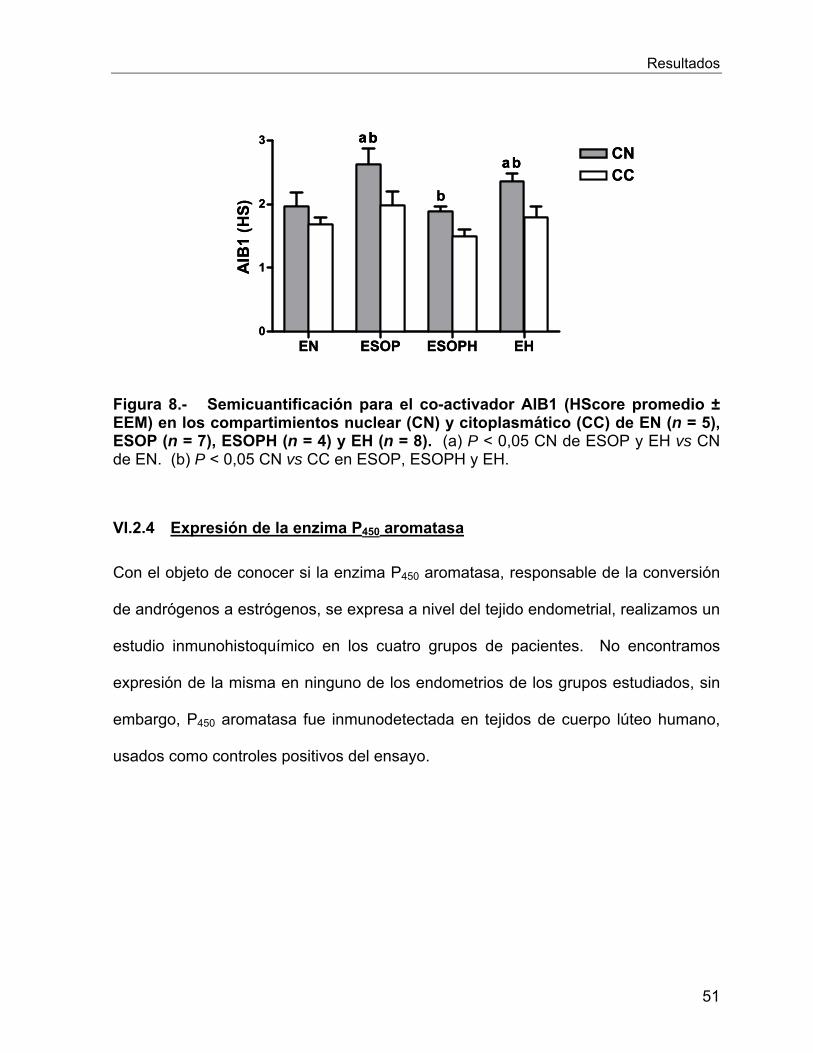
Resultados
51
Figura 8.- Semicuantificación para el co-activador AIB1 (HScore promedio ± EEM) en los compartimientos nuclear (CN) y citoplasmático (CC) de EN (n = 5), ESOP (n = 7), ESOPH (n = 4) y EH (n = 8). (a) P < 0,05 CN de ESOP y EH vs CN de EN. (b) P < 0,05 CN vs CC en ESOP, ESOPH y EH.
VI.2.4 Expresión de la enzima P450 aromatasa
Con el objeto de conocer si la enzima P450 aromatasa, responsable de la conversión
de andrógenos a estrógenos, se expresa a nivel del tejido endometrial, realizamos un
estudio inmunohistoquímico en los cuatro grupos de pacientes. No encontramos
expresión de la misma en ninguno de los endometrios de los grupos estudiados, sin
embargo, P450 aromatasa fue inmunodetectada en tejidos de cuerpo lúteo humano,
usados como controles positivos del ensayo.
b
ba
EN ESOP ESOPH EH0
1
2
3CNCC
AIB
1 (H
S)
ba
b
baba
EN ESOP ESOPH EH0
1
2
3CNCC
AIB
1 (H
S)
baba

Resultados
52
VI.2.5 Estudios de Proliferación
VI.2.5.1. Detección de Ki67 por inmunohistoquímica
La tinción positiva para Ki67, indicativa de proliferación celular, se localizó en el
núcleo de las células epiteliales y estromales de los cuatro grupos de endometrios
(Figura 9-A), lo que es compatible con la conocida expresión nuclear de este
antígeno en las células que se encuentran en las fases G1, S, G2 y M del ciclo
celular. Los resultados muestran mayor proliferación en las células epiteliales que en
las células estromales de todos los grupos estudiados, siendo este aumento
estadísticamente significativo en ESOP, ESOPH y EH (P < 0,05) (Figura 9-B).
Además, el porcentaje de células epiteliales con marca positiva para Ki67 fue mayor
en ESOP (45%, P < 0,05), ESOPH (23,5%) y EH (24%) al comparar con EN (Figura
9-B). Lo anterior es de especial importancia, dado que los endometrios controles, por
pertenecer a la fase proliferativa de un ciclo menstrual normal se encuentran en
activa proliferación. La expresión de Ki67 en estroma fue 31% mayor en ESOP
respecto a EN; 62% menor en ESOPH respecto a ESOP (P = 0.056); 66% y 74%
menor en EH vs EN y ESOP, respectivamente (P < 0,05).
Además, la expresión de este antígeno fue ligeramente mayor en las células
epiteliales (11%) y en las células estromales (5%) de endometrios de pacientes SOP
insulino-resistentes, al comparar con las pacientes SOP no insulino-resistentes. Sin
embargo, estos cambios no fueron estadísticamente significativos.
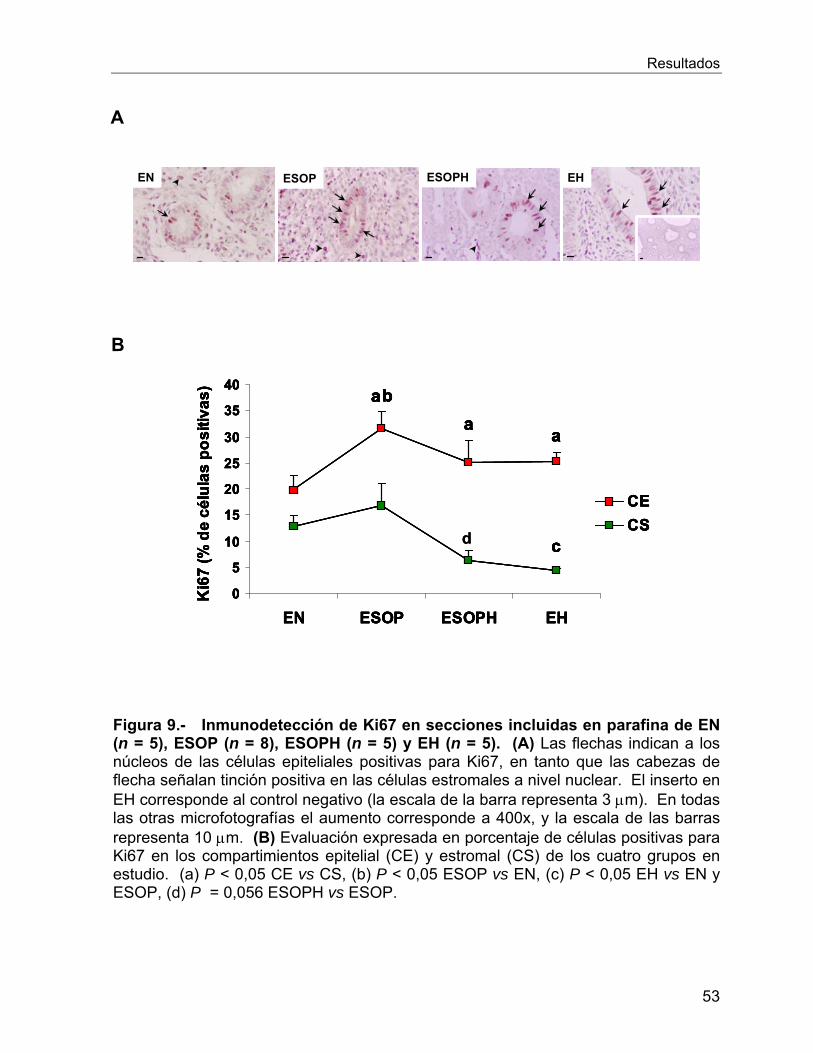
Resultados
53
Figura 9.- Inmunodetección de Ki67 en secciones incluidas en parafina de EN (n = 5), ESOP (n = 8), ESOPH (n = 5) y EH (n = 5). (A) Las flechas indican a los núcleos de las células epiteliales positivas para Ki67, en tanto que las cabezas de flecha señalan tinción positiva en las células estromales a nivel nuclear. El inserto en EH corresponde al control negativo (la escala de la barra representa 3 μm). En todas las otras microfotografías el aumento corresponde a 400x, y la escala de las barras representa 10 μm. (B) Evaluación expresada en porcentaje de células positivas para Ki67 en los compartimientos epitelial (CE) y estromal (CS) de los cuatro grupos en estudio. (a) P < 0,05 CE vs CS, (b) P < 0,05 ESOP vs EN, (c) P < 0,05 EH vs EN y ESOP, (d) P = 0,056 ESOPH vs ESOP.
B
EN ESOP ESOPH EHEN ESOP ESOPH EH
A
d
0
5
10
15
20
25
30
35
40
EN ESOP ESOPH EH
Ki6
7 (%
de
célu
las
posi
tivas
)
CECS
aaab
c
0
5
10
15
20
25
30
35
40
EN ESOP ESOPH EH
Ki6
7 (%
de
célu
las
posi
tivas
)
CECS
aaab
c
aaab
cd
0
5
10
15
20
25
30
35
40
EN ESOP ESOPH EH
Ki6
7 (%
de
célu
las
posi
tivas
)
CECS
aaab
c
0
5
10
15
20
25
30
35
40
EN ESOP ESOPH EH
Ki6
7 (%
de
célu
las
posi
tivas
)
CECS
aaab
c
aaab
c

Resultados
54
VI.2.6 Estudios de Apoptosis
La evaluación de apoptosis se realizó mediante la determinación de la expresión de
Bcl-2, Bax, Pro-caspasa-3, Caspasa-3, y de la fragmentación del DNA.
VI.2.6.1. Expresión de Bcl-2 y Bax por Western Blot
En los endometrios de todos los grupos de pacientes, la expresión de la proteína
anti-apoptótica Bcl-2 fue mayor que la de la proteína pro-apoptótica Bax (Figura 10-A
y -B). Además, se observó un incremento en la expresión proteica de Bcl-2 en
ambos grupos de pacientes SOP y este aumento fue estadísticamente significativo
en el grupo ESOPH (64%, P < 0,05 ESOPH vs EN). Una disminución significativa de
Bcl-2 fue observada en EH al comparar con ESOP y ESOPH. Para este ensayo,
usamos como control positivo endometrios de 2 pacientes con carcinoma
endometrial, y en ellos encontramos que Bcl-2 aumentó su expresión 1 vez al
comparar con EN. Por otro lado, la expresión de Bax presentó un perfil de expresión
similar al de Bcl-2 en todos los grupos en estudio, pero dichos cambios no fueron
estadísticamente significativos. En las muestras de carcinoma endometrial, usadas
como control positivo, se observó una menor expresión de Bax respecto a Bcl-2, y
además, la expresión de Bax disminuyó 0,4 veces respecto a EN.
Dado que Bcl-2 y Bax ejercen acciones contrapuestas para la supervivencia celular, y
a que ambas proteínas forman dímeros para ejercer su acción; es que la
concentración relativa de Bcl-2 y Bax es uno de los factores que determina el destino
final de la célula. Es por ello, que se calculó la razón entre Bcl-2 y Bax en todos los
grupos de endometrios estudiados, observándose un incremento de la misma
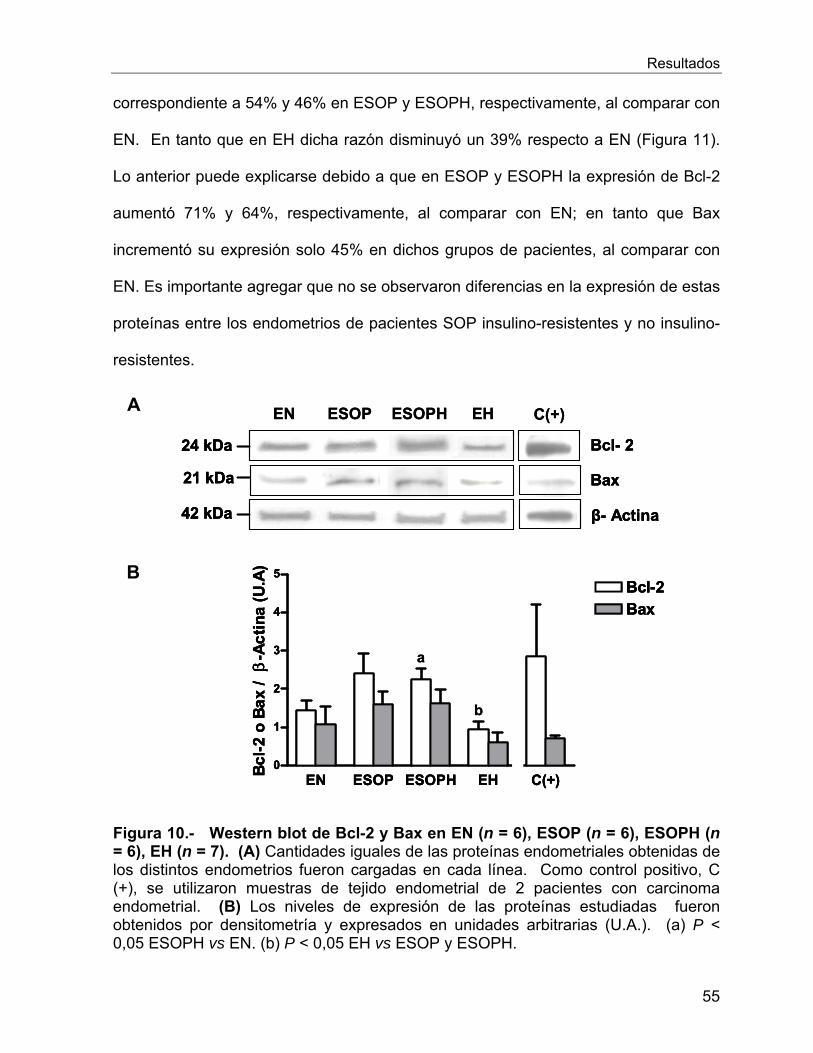
Resultados
55
correspondiente a 54% y 46% en ESOP y ESOPH, respectivamente, al comparar con
EN. En tanto que en EH dicha razón disminuyó un 39% respecto a EN (Figura 11).
Lo anterior puede explicarse debido a que en ESOP y ESOPH la expresión de Bcl-2
aumentó 71% y 64%, respectivamente, al comparar con EN; en tanto que Bax
incrementó su expresión solo 45% en dichos grupos de pacientes, al comparar con
EN. Es importante agregar que no se observaron diferencias en la expresión de estas
proteínas entre los endometrios de pacientes SOP insulino-resistentes y no insulino-
resistentes.
Figura 10.- Western blot de Bcl-2 y Bax en EN (n = 6), ESOP (n = 6), ESOPH (n = 6), EH (n = 7). (A) Cantidades iguales de las proteínas endometriales obtenidas de los distintos endometrios fueron cargadas en cada línea. Como control positivo, C (+), se utilizaron muestras de tejido endometrial de 2 pacientes con carcinoma endometrial. (B) Los niveles de expresión de las proteínas estudiadas fueron obtenidos por densitometría y expresados en unidades arbitrarias (U.A.). (a) P < 0,05 ESOPH vs EN. (b) P < 0,05 EH vs ESOP y ESOPH.
EN ESOP ESOPH EH C(+)
24 kDa Bcl- 2
Bax21 kDa
42 kDa β- Actina
EN ESOP ESOPH EH C(+)
24 kDa Bcl- 224 kDa Bcl- 2
Bax21 kDa Bax21 kDa
42 kDa β- Actina42 kDa β- Actina
A
B
EN ESOP ESOPH EH C(+)0
1
2
3
4
5Bcl-2Bax
Bcl-2
o B
ax /β
-Act
ina
(U.A
)
a
b
EN ESOP ESOPH EH C(+)0
1
2
3
4
5Bcl-2Bax
Bcl-2
o B
ax /β
-Act
ina
(U.A
)
EN ESOP ESOPH EH C(+)0
1
2
3
4
5Bcl-2Bax
Bcl-2
o B
ax /β
-Act
ina
(U.A
)
a
b
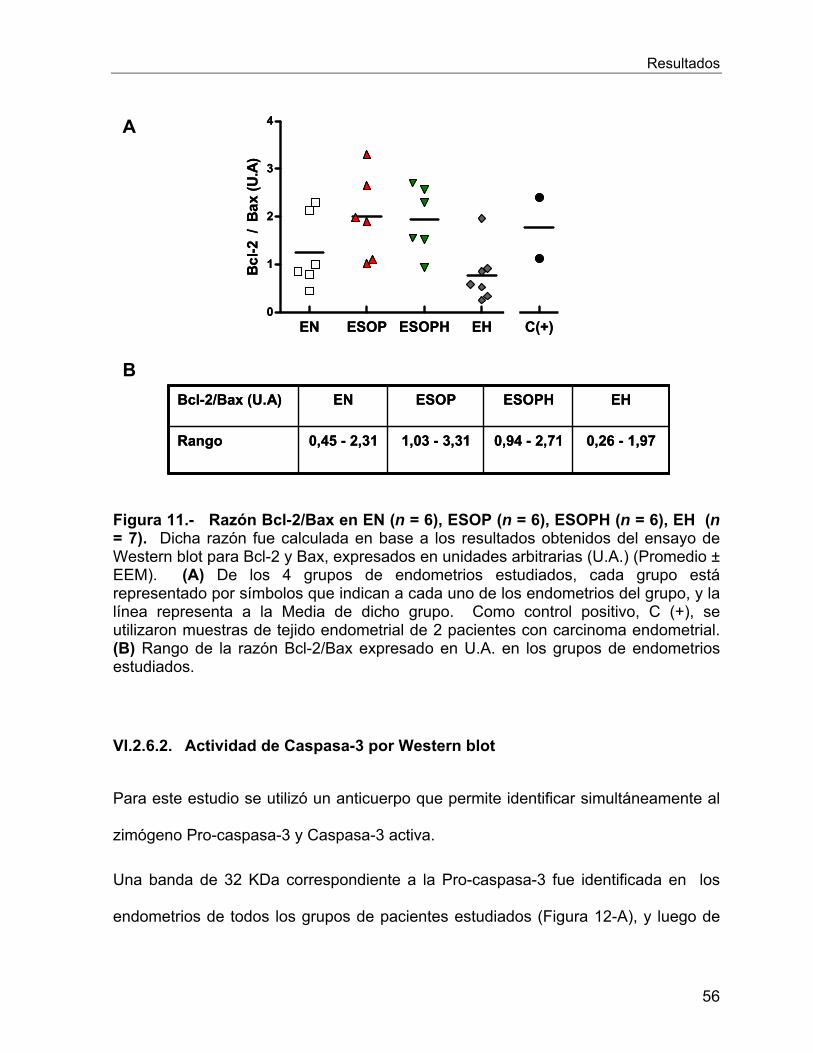
Resultados
56
Figura 11.- Razón Bcl-2/Bax en EN (n = 6), ESOP (n = 6), ESOPH (n = 6), EH (n = 7). Dicha razón fue calculada en base a los resultados obtenidos del ensayo de Western blot para Bcl-2 y Bax, expresados en unidades arbitrarias (U.A.) (Promedio ± EEM). (A) De los 4 grupos de endometrios estudiados, cada grupo está representado por símbolos que indican a cada uno de los endometrios del grupo, y la línea representa a la Media de dicho grupo. Como control positivo, C (+), se utilizaron muestras de tejido endometrial de 2 pacientes con carcinoma endometrial. (B) Rango de la razón Bcl-2/Bax expresado en U.A. en los grupos de endometrios estudiados.
VI.2.6.2. Actividad de Caspasa-3 por Western blot
Para este estudio se utilizó un anticuerpo que permite identificar simultáneamente al
zimógeno Pro-caspasa-3 y Caspasa-3 activa.
Una banda de 32 KDa correspondiente a la Pro-caspasa-3 fue identificada en los
endometrios de todos los grupos de pacientes estudiados (Figura 12-A), y luego de
B
0,26 - 1,970,94 - 2,711,03 - 3,310,45 - 2,31Rango
EHESOPHESOPENBcl-2/Bax (U.A)
0,26 - 1,970,94 - 2,711,03 - 3,310,45 - 2,31Rango
EHESOPHESOPENBcl-2/Bax (U.A)
EN ESOP ESOPH EH C(+)0
1
2
3
4
Bcl
-2 /
Bax
(U.A
)
EN ESOP ESOPH EH C(+)0
1
2
3
4
Bcl
-2 /
Bax
(U.A
)
A
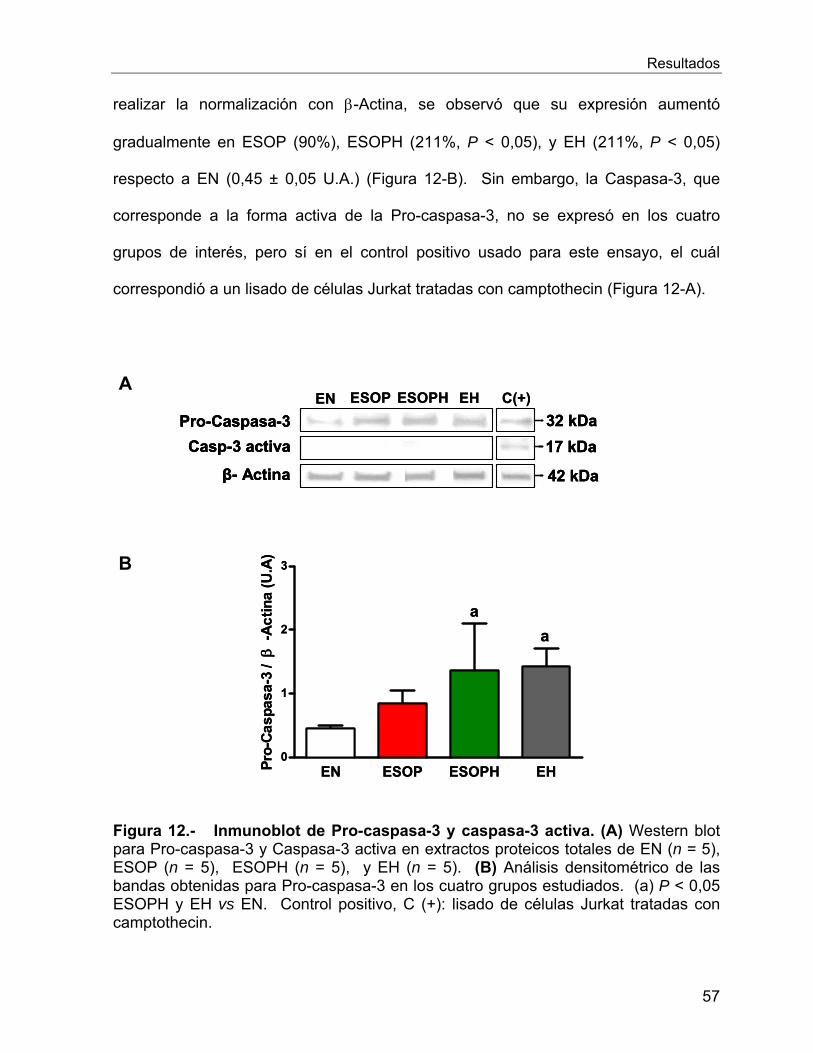
Resultados
57
realizar la normalización con β-Actina, se observó que su expresión aumentó
gradualmente en ESOP (90%), ESOPH (211%, P < 0,05), y EH (211%, P < 0,05)
respecto a EN (0,45 ± 0,05 U.A.) (Figura 12-B). Sin embargo, la Caspasa-3, que
corresponde a la forma activa de la Pro-caspasa-3, no se expresó en los cuatro
grupos de interés, pero sí en el control positivo usado para este ensayo, el cuál
correspondió a un lisado de células Jurkat tratadas con camptothecin (Figura 12-A).
Figura 12.- Inmunoblot de Pro-caspasa-3 y caspasa-3 activa. (A) Western blot para Pro-caspasa-3 y Caspasa-3 activa en extractos proteicos totales de EN (n = 5), ESOP (n = 5), ESOPH (n = 5), y EH (n = 5). (B) Análisis densitométrico de las bandas obtenidas para Pro-caspasa-3 en los cuatro grupos estudiados. (a) P < 0,05 ESOPH y EH vs EN. Control positivo, C (+): lisado de células Jurkat tratadas con camptothecin.
A
B
EN ESOP ESOPH EH C(+)Pro-Caspasa-3 32 kDa
β- Actina 42 kDa
Casp-3 activa 17 kDa
EN ESOP ESOPH EH C(+)Pro-Caspasa-3 32 kDaPro-Caspasa-3 32 kDa
β- Actina 42 kDaβ- Actina 42 kDa
Casp-3 activa 17 kDaCasp-3 activa 17 kDa
EN ESOP ESOPH EH0
1
2
3
Pro-
Casp
asa-
3 /β
-Act
ina
(U.A
)
aa
EN ESOP ESOPH EH0
1
2
3
Pro-
Casp
asa-
3 /β
-Act
ina
(U.A
)
aa

Resultados
58
VI.2.6.3. Determinación de la fragmentación del DNA por TUNEL
A través de este estudio, se observaron escasas figuras con signos positivos de
apoptosis en los endometrios los cuatro grupos en estudio, independientemente del
tipo celular analizado. Además, no se encontraron diferencias en el porcentaje de
células apoptóticas entre los distintos grupos de endometrios.
VI.2.7 Estudio de la onco-proteína Ras
VI.2.7.1. Expresión de la onco-proteína Ras por Western blot
La Figura 13 muestra que la expresión de Ras aumentó en ESOP (6 veces, P <
0,05), ESOPH (4,5 veces, P < 0,05), y en EH (1,5 veces) respecto a EN (0,2 ± 0,05
U.A.). Por otro lado, no de detectaron diferencias al comparar la expresión de Ras
entre endometrios de pacientes SOP insulino-resistentes y no insulino-resistentes.
Como control positivo, utilizamos muestras de tejido endometrial de pacientes con
carcinoma endometrial, donde se observó un incremento de 9,8 veces en la
expresión de Ras respecto a EN (P < 0,05).
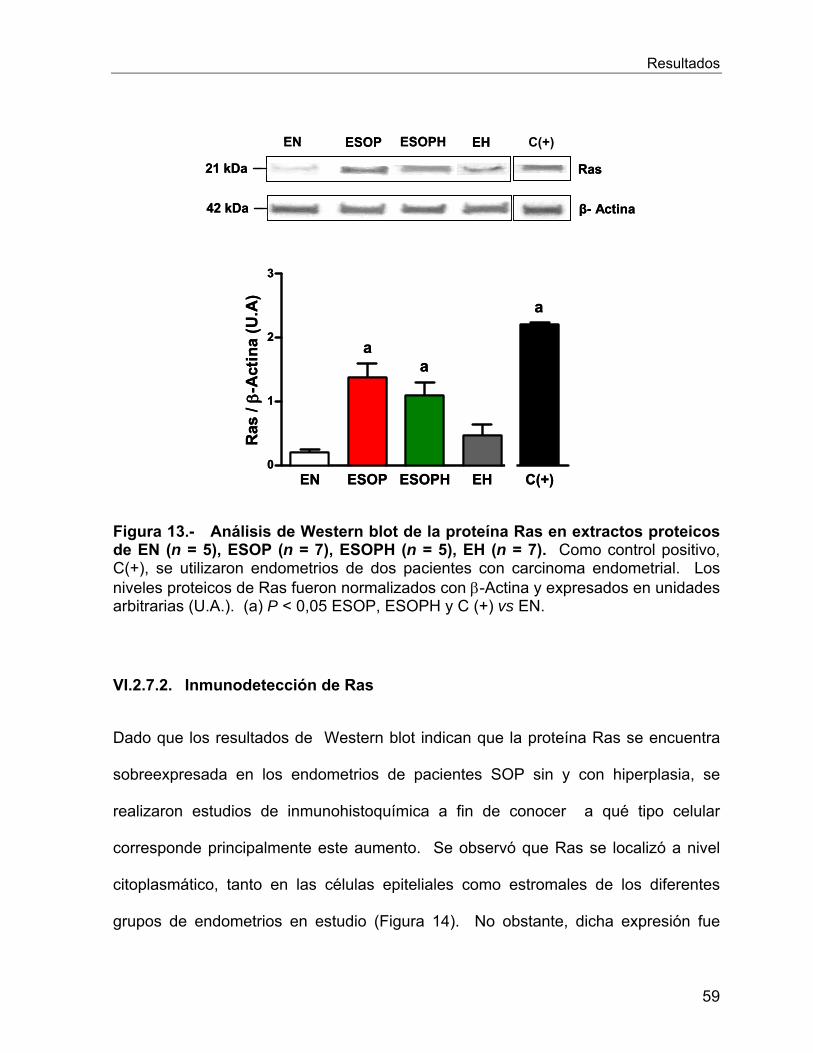
Resultados
59
Figura 13.- Análisis de Western blot de la proteína Ras en extractos proteicos de EN (n = 5), ESOP (n = 7), ESOPH (n = 5), EH (n = 7). Como control positivo, C(+), se utilizaron endometrios de dos pacientes con carcinoma endometrial. Los niveles proteicos de Ras fueron normalizados con β-Actina y expresados en unidades arbitrarias (U.A.). (a) P < 0,05 ESOP, ESOPH y C (+) vs EN.
VI.2.7.2. Inmunodetección de Ras
Dado que los resultados de Western blot indican que la proteína Ras se encuentra
sobreexpresada en los endometrios de pacientes SOP sin y con hiperplasia, se
realizaron estudios de inmunohistoquímica a fin de conocer a qué tipo celular
corresponde principalmente este aumento. Se observó que Ras se localizó a nivel
citoplasmático, tanto en las células epiteliales como estromales de los diferentes
grupos de endometrios en estudio (Figura 14). No obstante, dicha expresión fue
aa
a
EN ESOP ESOPH EH C(+)0
1
2
3R
as /β-
Act
ina
(U.A
)
aa
a
EN ESOP ESOPH EH C(+)0
1
2
3R
as /β-
Act
ina
(U.A
)
21 kDa
42 kDa
Ras
β- Actina
EN ESOP ESOPH EH C(+)
21 kDa
42 kDa
Ras
β- Actina
EN ESOP ESOPH EH C(+)
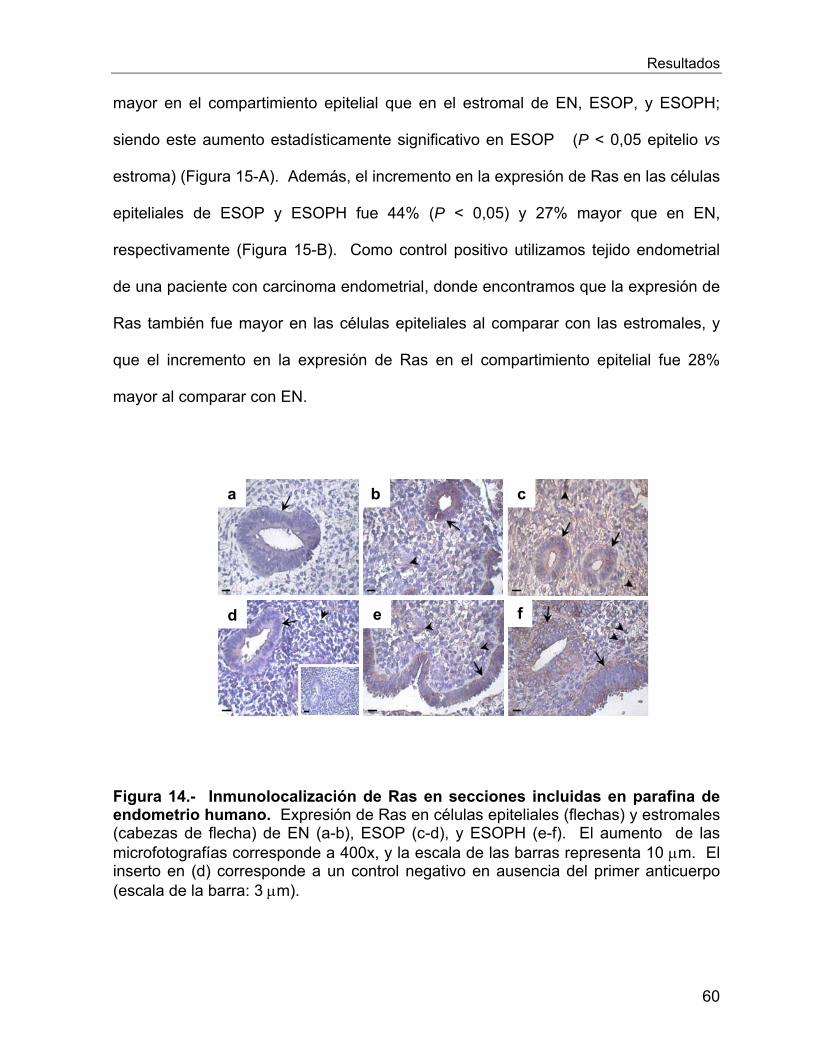
Resultados
60
mayor en el compartimiento epitelial que en el estromal de EN, ESOP, y ESOPH;
siendo este aumento estadísticamente significativo en ESOP (P < 0,05 epitelio vs
estroma) (Figura 15-A). Además, el incremento en la expresión de Ras en las células
epiteliales de ESOP y ESOPH fue 44% (P < 0,05) y 27% mayor que en EN,
respectivamente (Figura 15-B). Como control positivo utilizamos tejido endometrial
de una paciente con carcinoma endometrial, donde encontramos que la expresión de
Ras también fue mayor en las células epiteliales al comparar con las estromales, y
que el incremento en la expresión de Ras en el compartimiento epitelial fue 28%
mayor al comparar con EN.
Figura 14.- Inmunolocalización de Ras en secciones incluidas en parafina de endometrio humano. Expresión de Ras en células epiteliales (flechas) y estromales (cabezas de flecha) de EN (a-b), ESOP (c-d), y ESOPH (e-f). El aumento de las microfotografías corresponde a 400x, y la escala de las barras representa 10 μm. El inserto en (d) corresponde a un control negativo en ausencia del primer anticuerpo (escala de la barra: 3 μm).
a
d
b c
e f
a
d
b c
e f
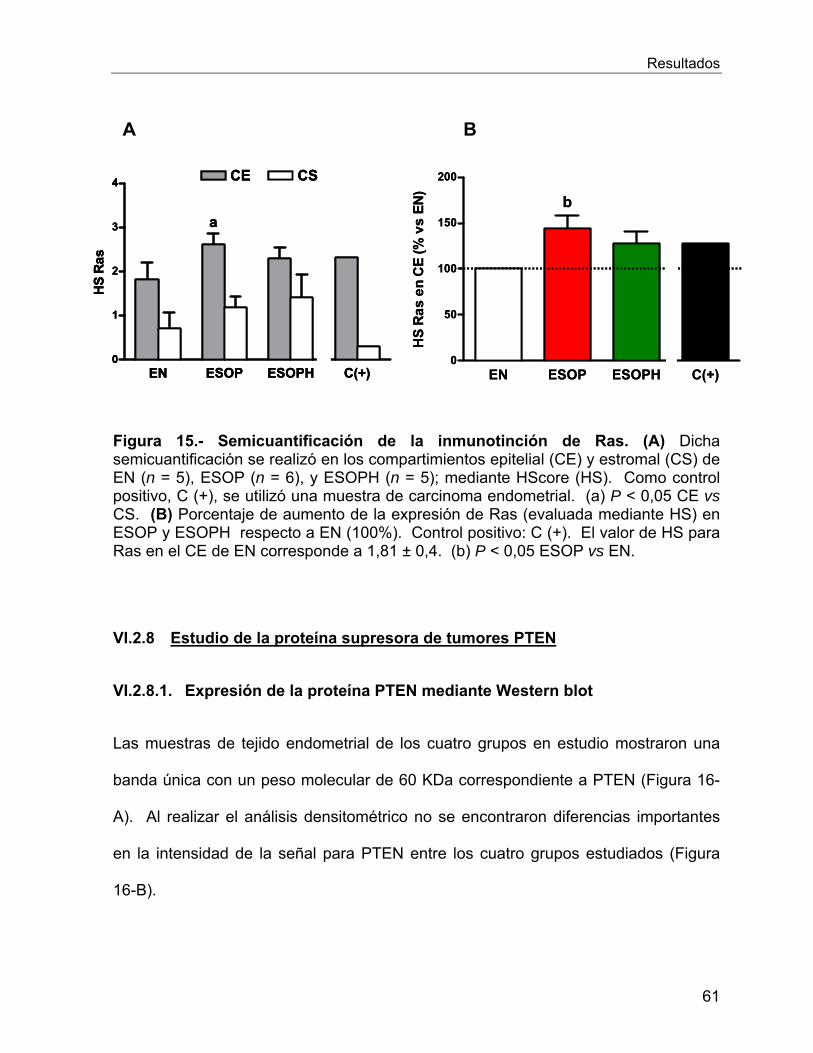
Resultados
61
Figura 15.- Semicuantificación de la inmunotinción de Ras. (A) Dicha semicuantificación se realizó en los compartimientos epitelial (CE) y estromal (CS) de EN (n = 5), ESOP (n = 6), y ESOPH (n = 5); mediante HScore (HS). Como control positivo, C (+), se utilizó una muestra de carcinoma endometrial. (a) P < 0,05 CE vs CS. (B) Porcentaje de aumento de la expresión de Ras (evaluada mediante HS) en ESOP y ESOPH respecto a EN (100%). Control positivo: C (+). El valor de HS para Ras en el CE de EN corresponde a 1,81 ± 0,4. (b) P < 0,05 ESOP vs EN.
VI.2.8 Estudio de la proteína supresora de tumores PTEN
VI.2.8.1. Expresión de la proteína PTEN mediante Western blot
Las muestras de tejido endometrial de los cuatro grupos en estudio mostraron una
banda única con un peso molecular de 60 KDa correspondiente a PTEN (Figura 16-
A). Al realizar el análisis densitométrico no se encontraron diferencias importantes
en la intensidad de la señal para PTEN entre los cuatro grupos estudiados (Figura
16-B).
EN ESOP ESOPH C(+)0
1
2
3
4 CSCE
HS
Ras
a
EN ESOP ESOPH C(+)0
1
2
3
4 CSCE
HS
Ras
a
EN ESOP ESOPH C(+)0
1
2
3
4 CSCE
HS
Ras
a
EN ESOP ESOPH C(+)0
50
100
150
200
HS
Ras
en
CE
(% v
s EN
)
b
EN ESOP ESOPH C(+)0
50
100
150
200
HS
Ras
en
CE
(% v
s EN
)
b
A B
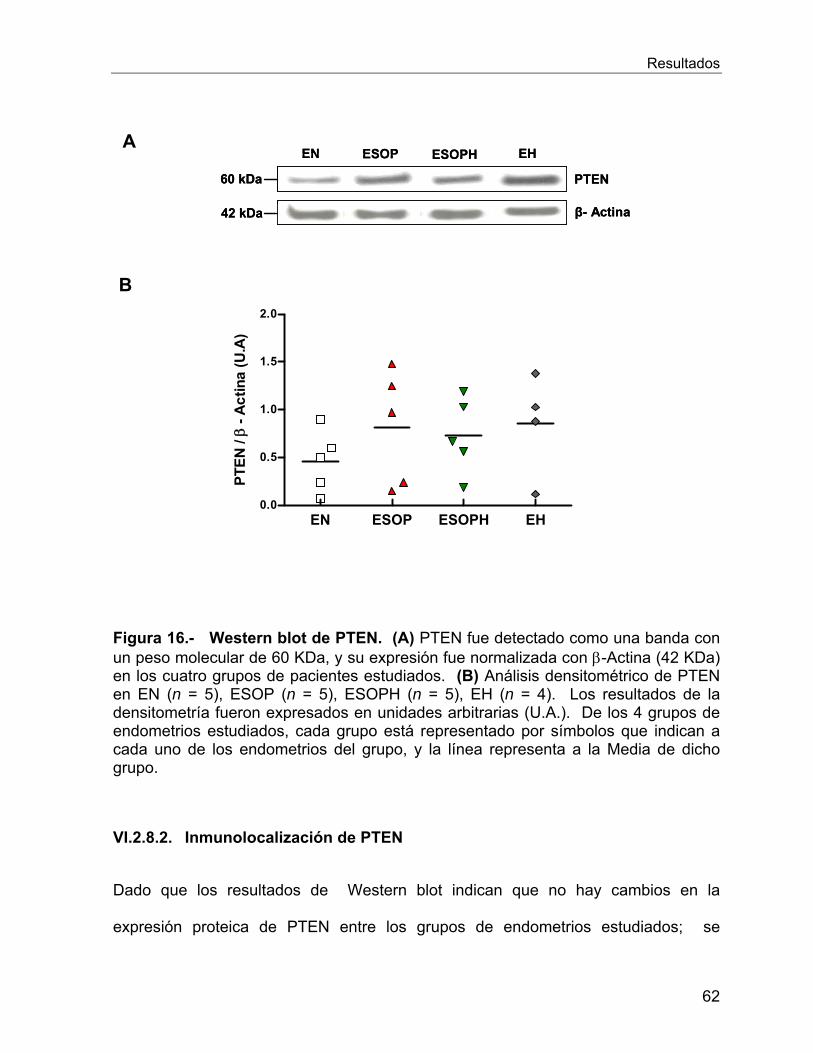
Resultados
62
Figura 16.- Western blot de PTEN. (A) PTEN fue detectado como una banda con un peso molecular de 60 KDa, y su expresión fue normalizada con β-Actina (42 KDa) en los cuatro grupos de pacientes estudiados. (B) Análisis densitométrico de PTEN en EN (n = 5), ESOP (n = 5), ESOPH (n = 5), EH (n = 4). Los resultados de la densitometría fueron expresados en unidades arbitrarias (U.A.). De los 4 grupos de endometrios estudiados, cada grupo está representado por símbolos que indican a cada uno de los endometrios del grupo, y la línea representa a la Media de dicho grupo.
VI.2.8.2. Inmunolocalización de PTEN
Dado que los resultados de Western blot indican que no hay cambios en la
expresión proteica de PTEN entre los grupos de endometrios estudiados; se
B
EN ESOP ESOPH EH0.0
0.5
1.0
1.5
2.0
PTEN
/β
- Act
ina
(U.A
)
PTEN
β- Actina42 kDa
60 kDa
EN ESOP ESOPH EH
PTEN
β- Actina42 kDa
60 kDa
EN ESOP ESOPH EHA
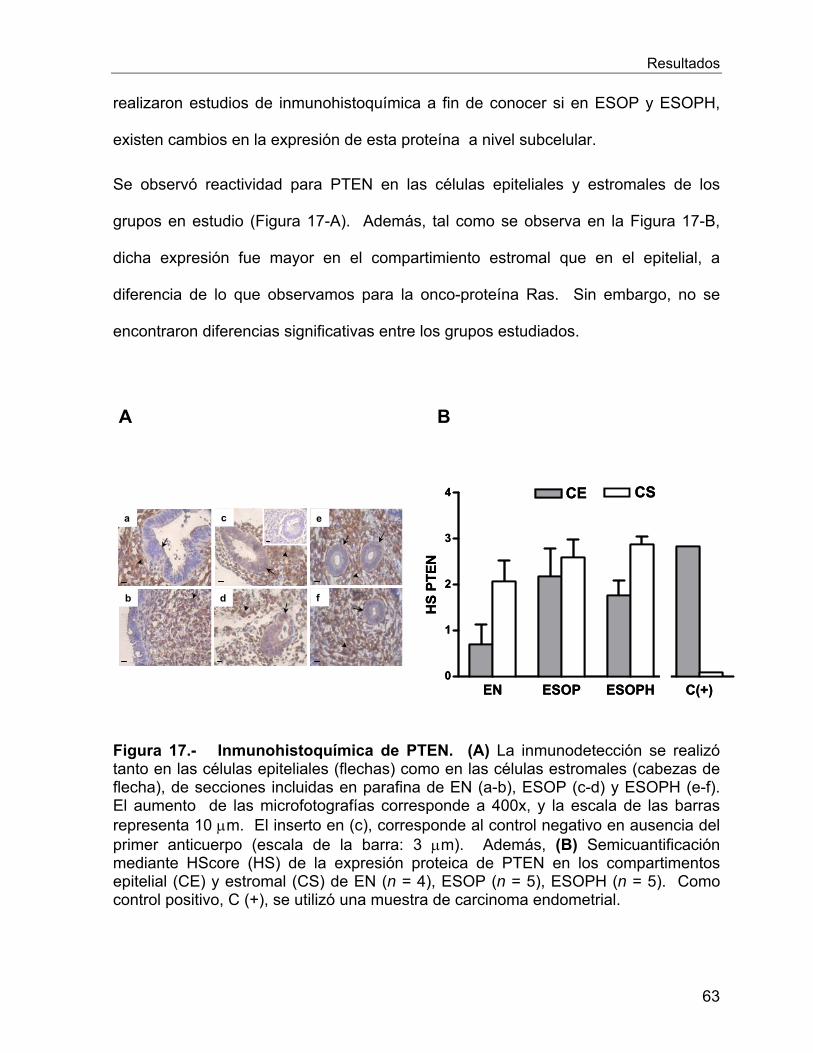
Resultados
63
realizaron estudios de inmunohistoquímica a fin de conocer si en ESOP y ESOPH,
existen cambios en la expresión de esta proteína a nivel subcelular.
Se observó reactividad para PTEN en las células epiteliales y estromales de los
grupos en estudio (Figura 17-A). Además, tal como se observa en la Figura 17-B,
dicha expresión fue mayor en el compartimiento estromal que en el epitelial, a
diferencia de lo que observamos para la onco-proteína Ras. Sin embargo, no se
encontraron diferencias significativas entre los grupos estudiados.
Figura 17.- Inmunohistoquímica de PTEN. (A) La inmunodetección se realizó tanto en las células epiteliales (flechas) como en las células estromales (cabezas de flecha), de secciones incluidas en parafina de EN (a-b), ESOP (c-d) y ESOPH (e-f). El aumento de las microfotografías corresponde a 400x, y la escala de las barras representa 10 μm. El inserto en (c), corresponde al control negativo en ausencia del primer anticuerpo (escala de la barra: 3 μm). Además, (B) Semicuantificación mediante HScore (HS) de la expresión proteica de PTEN en los compartimentos epitelial (CE) y estromal (CS) de EN (n = 4), ESOP (n = 5), ESOPH (n = 5). Como control positivo, C (+), se utilizó una muestra de carcinoma endometrial.
A B
EN ESOP ESOPH C(+)0
1
2
3
4 CSCE
HS
PTEN
EN ESOP ESOPH C(+)0
1
2
3
4 CSCE
HS
PTEN
a c e
b d f
a c e
b d f

Resultados
64
VI.2.9 Expresión y actividad de Akt
VI.2.9.1. Expresión de Akt total mediante Western Blot
Tal como se muestra en la Figura 18, los niveles de Akt total en endometrios ESOP y
ESOPH fueron 32% y 55% mayores, respectivamente, al comparar con los
endometrios controles. En tanto que el nivel de expresión de esta proteína en EH fue
similar a EN.
VI.2.9.2. Actividad de Akt
Cuando Akt es fosforilada en los residuos de Ser473 y/o Thr308 se activa y actúa
como un regulador de procesos fisiológicos como la apoptosis, la supervivencia
celular y el ciclo celular. Es por ello que mediante estudios de Western blot se evaluó
la fosforilación de esta proteína en ambos residuos. Se observó un incremento en
Akt-P en Ser473 de 131% en ESOP (P < 0,05) y de 97% en ESOPH (P < 0,05) al
comparar con EN, mientras que no hubo cambios para Akt-P en Thr308 entre los
grupos estudiados (Figura 19-A y -B). Tampoco se observaron diferencias en la
fosforilación de Akt entre los endometrios de pacientes SOP insulino-resistentes y no
insulino-resistentes.
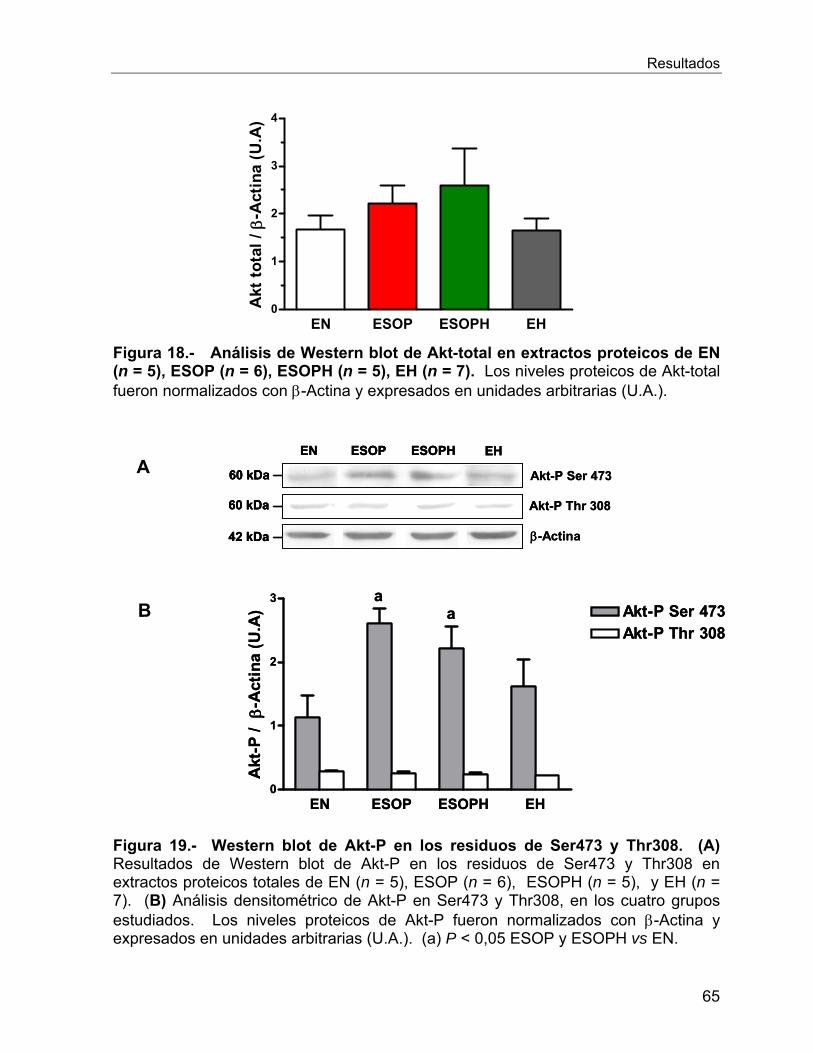
Resultados
65
Figura 18.- Análisis de Western blot de Akt-total en extractos proteicos de EN (n = 5), ESOP (n = 6), ESOPH (n = 5), EH (n = 7). Los niveles proteicos de Akt-total fueron normalizados con β-Actina y expresados en unidades arbitrarias (U.A.).
Figura 19.- Western blot de Akt-P en los residuos de Ser473 y Thr308. (A) Resultados de Western blot de Akt-P en los residuos de Ser473 y Thr308 en extractos proteicos totales de EN (n = 5), ESOP (n = 6), ESOPH (n = 5), y EH (n = 7). (B) Análisis densitométrico de Akt-P en Ser473 y Thr308, en los cuatro grupos estudiados. Los niveles proteicos de Akt-P fueron normalizados con β-Actina y expresados en unidades arbitrarias (U.A.). (a) P < 0,05 ESOP y ESOPH vs EN.
EN ESOP ESOPH EH0
1
2
3
4
Akt
tota
l /β-
Act
ina
(U.A
)
A
B
Akt-P Thr 308
β-Actina
EN ESOP ESOPH EH
Akt-P Ser 473
42 kDa
60 kDa
60 kDa
Akt-P Thr 308
β-Actina
EN ESOP ESOPH EHEN ESOP ESOPH EH
Akt-P Ser 473
42 kDa42 kDa
60 kDa60 kDa
60 kDa60 kDa
EN ESOP ESOPH EH0
1
2
3Akt-P Ser 473Akt-P Thr 308
Akt
-P /β-
Act
ina
(U.A
)
aa
EN ESOP ESOPH EH0
1
2
3Akt-P Ser 473Akt-P Thr 308
Akt
-P /β-
Act
ina
(U.A
)
aa

Resultados
66
VI.2.10 Reguladores del ciclo celular
VI.2.10.1. Expresión proteica de inhibidores del ciclo celular: p27 y p21
La función primordial de las proteínas p27 y p21, cuando se localizan a nivel nuclear,
es ejercer una regulación negativa sobre el ciclo celular, impidiendo que las células
pasen de la fase G1 a la fase S del ciclo celular. Basado en los resultados obtenidos
de Akt-P (Sección VI.2.9.) en los distintos grupos de endometrios, y sabiendo que
cuando Akt se encuentra activo es capaz de bloquear a p27 y p21, mediante
fosforilación, conduciendo a que estos inhibidores adopten una localización
citoplasmática; se realizó una evaluación de la expresión proteica de p27 y p21 por
Western blot y/o inmunohistoquímica.
VI.2.10.1.1. Expresión proteica de p27 y p21 mediante Western blot
La Figura 20 muestra que el inhibidor p27, conocido por asociarse al complejo Ciclina
E/Cdk-2 en fase G1 tardía para inhibir la fosforilación de Rb, no modificó su
expresión en ESOP, ESOPH y EH al comparar con endometrios normales.
La expresión del inhibidor p21 fue estudiada con 3 anticuerpos distintos, tal como se
indica en Materiales y Métodos. Sin embargo, su expresión no pudo ser visualizada
por los ensayos de Western Blot en ninguno de los grupos en estudio.
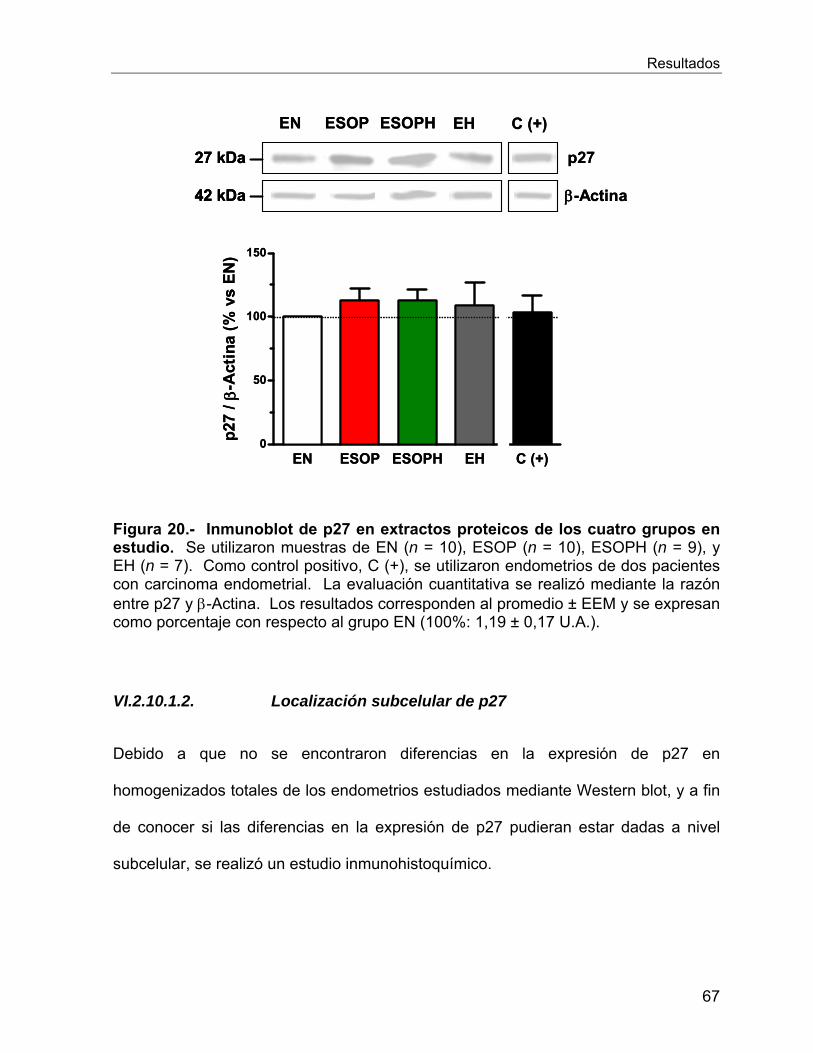
Resultados
67
Figura 20.- Inmunoblot de p27 en extractos proteicos de los cuatro grupos en estudio. Se utilizaron muestras de EN (n = 10), ESOP (n = 10), ESOPH (n = 9), y EH (n = 7). Como control positivo, C (+), se utilizaron endometrios de dos pacientes con carcinoma endometrial. La evaluación cuantitativa se realizó mediante la razón entre p27 y β-Actina. Los resultados corresponden al promedio ± EEM y se expresan como porcentaje con respecto al grupo EN (100%: 1,19 ± 0,17 U.A.).
VI.2.10.1.2. Localización subcelular de p27
Debido a que no se encontraron diferencias en la expresión de p27 en
homogenizados totales de los endometrios estudiados mediante Western blot, y a fin
de conocer si las diferencias en la expresión de p27 pudieran estar dadas a nivel
subcelular, se realizó un estudio inmunohistoquímico.
EN ESOP ESOPH EH C (+)0
50
100
150
p27
/ β-A
ctin
a (%
vs
EN)
EN ESOP ESOPH EH C (+)0
50
100
150
p27
/ β-A
ctin
a (%
vs
EN)
p27
β-Actina
EN ESOP ESOPH EH C (+)
42 kDa
27 kDa p27
β-Actina
EN ESOP ESOPH EH C (+)
42 kDa42 kDa
27 kDa27 kDa

Resultados
68
La inmunodetección para p27 se muestra en la Figura 21 donde se observa que la
tinción para p27 fue nuclear y citoplasmática, tanto a nivel estromal como epitelial en
todas las muestras analizadas.
Tal como se muestra en la Figura 22-A, en el compartimiento estromal, la expresión
nuclear de p27 disminuyó 64% en ESOP (P < 0,05 vs EN), 76% en ESOPH (P < 0,05
vs EN) y 45% en EH, al comparar con endometrios normales en fase proliferativa.
No se encontraron diferencias en la expresión de p27 en el citoplasma de las células
estromales.
Al estudiar el compartimiento epitelial (Figura 22-B), los resultados muestran mayor
expresión de p27 (47%) en el citoplasma de ESOP al comparar con EN, en tanto que
a nivel nuclear no se encontraron diferencias entre los grupos estudiados.
Dado que la insulino-resistencia es característica de un porcentaje importante de las
pacientes SOP, analizamos en forma separada la expresión proteica de p27 en el
endometrio de pacientes SOP no insulino-resistentes y de mujeres SOP insulino-
resistentes (Tabla 5). Encontramos que el grupo de mujeres SOP insulino-
resistentes expresó 78% y 57% menos p27 en el núcleo de las células estromales, al
comparar con las mujeres normales y las pacientes SOP no insulino-resistentes,
respectivamente (P < 0,05). En tanto que en las células epiteliales de endometrios
de mujeres SOP no insulino-resistentes, la expresión de p27 fue mayor en el
citoplasma (P < 0,05), al comparar con los controles normales (Tabla 5).
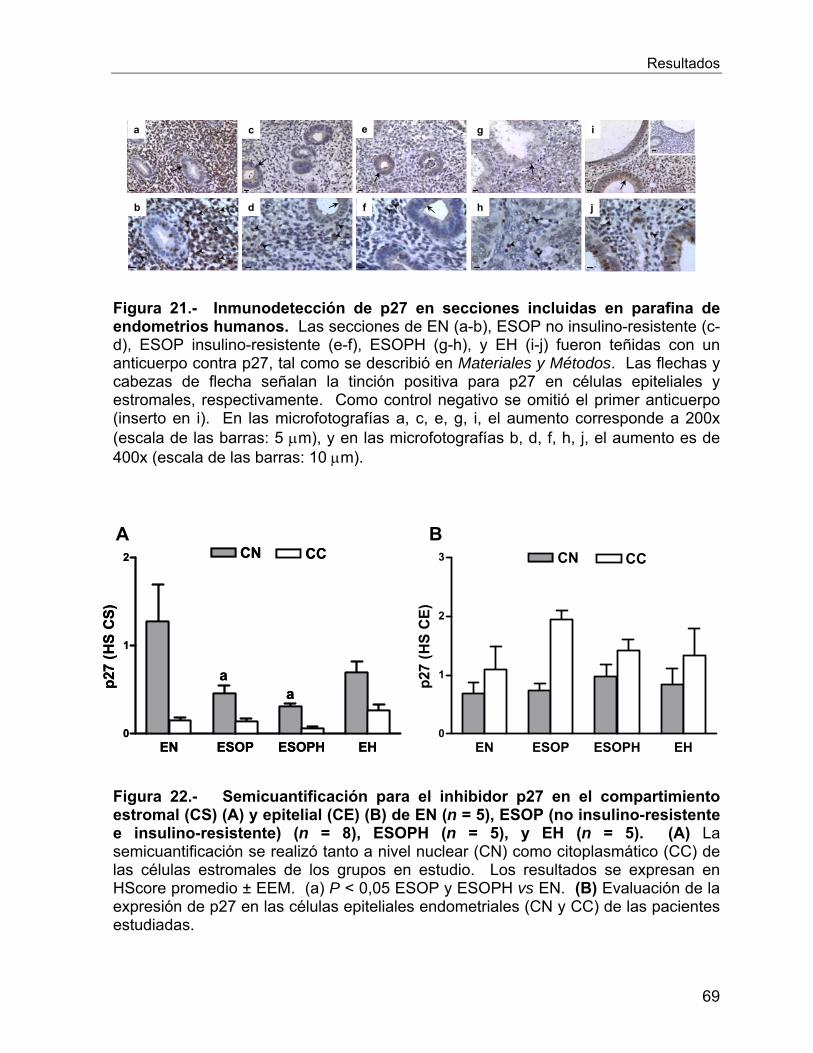
Resultados
69
Figura 21.- Inmunodetección de p27 en secciones incluidas en parafina de endometrios humanos. Las secciones de EN (a-b), ESOP no insulino-resistente (c-d), ESOP insulino-resistente (e-f), ESOPH (g-h), y EH (i-j) fueron teñidas con un anticuerpo contra p27, tal como se describió en Materiales y Métodos. Las flechas y cabezas de flecha señalan la tinción positiva para p27 en células epiteliales y estromales, respectivamente. Como control negativo se omitió el primer anticuerpo (inserto en i). En las microfotografías a, c, e, g, i, el aumento corresponde a 200x (escala de las barras: 5 μm), y en las microfotografías b, d, f, h, j, el aumento es de 400x (escala de las barras: 10 μm).
Figura 22.- Semicuantificación para el inhibidor p27 en el compartimiento estromal (CS) (A) y epitelial (CE) (B) de EN (n = 5), ESOP (no insulino-resistente e insulino-resistente) (n = 8), ESOPH (n = 5), y EH (n = 5). (A) La semicuantificación se realizó tanto a nivel nuclear (CN) como citoplasmático (CC) de las células estromales de los grupos en estudio. Los resultados se expresan en HScore promedio ± EEM. (a) P < 0,05 ESOP y ESOPH vs EN. (B) Evaluación de la expresión de p27 en las células epiteliales endometriales (CN y CC) de las pacientes estudiadas.
A B
EN ESOP ESOPH EH0
1
2
3 CN CC
p27
(HS
CE)
a
EN ESOP ESOPH EH0
1
2 CN CC
p27
(HS
CS)
aa
EN ESOP ESOPH EH0
1
2 CN CC
p27
(HS
CS)
a
a
b
c
d
e
f
g
h
i
j
a
b
c
d
e
f
g
h
i
j

Resultados
70
Tabla 5.- Semicuantificación mediante HScore de la expresión proteica de p27 a nivel nuclear del compartimiento estromal (CS), y a nivel citoplasmático del compartimiento epitelial (CE) de EN, ESOP no insulino-resistente y ESOP insulino-resistente.
Los valores se expresan en HScore promedio ± EEM. aP < 0,05 ESOP insulino-resistente y no insulino-resistente vs EN; bP < 0,05 ESOP insulino-resistente vs ESOP no insulino-resistente.
VI.2.10.2. Expresión y actividad de Cdk-2
La quinasa dependiente de ciclina Cdk-2 es importante en la progresión desde la
fase G1 a la fase S del ciclo celular, ya que fosforila a la proteína del retinoblastoma
(Rb), liberando al factor de transcripción E2F el cuál activa a genes necesarios para
la síntesis de DNA. En este estudio se evaluó tanto la expresión proteica de esta
quinasa (por Western blot e inmunohistoquímica), como su actividad.
Es importante adelantar, que a diferencia de lo encontrado para p27, en ninguno de
estos ensayos encontramos cambios en la expresión y actividad de esta quinasa
entre los endometrios de pacientes SOP insulino-resistentes y no insulino-
resistentes.
1,70 ± 0,160,28 ± 0,06 a,bESOP insulino-resistente
2,29 ± 0,06 a0,65 ± 0,10ESOP no insulino-resistente
1,10 ± 0,391,28 ± 0,42EN
CECS
1,70 ± 0,160,28 ± 0,06 a,bESOP insulino-resistente
2,29 ± 0,06 a0,65 ± 0,10ESOP no insulino-resistente
1,10 ± 0,391,28 ± 0,42EN
CECS
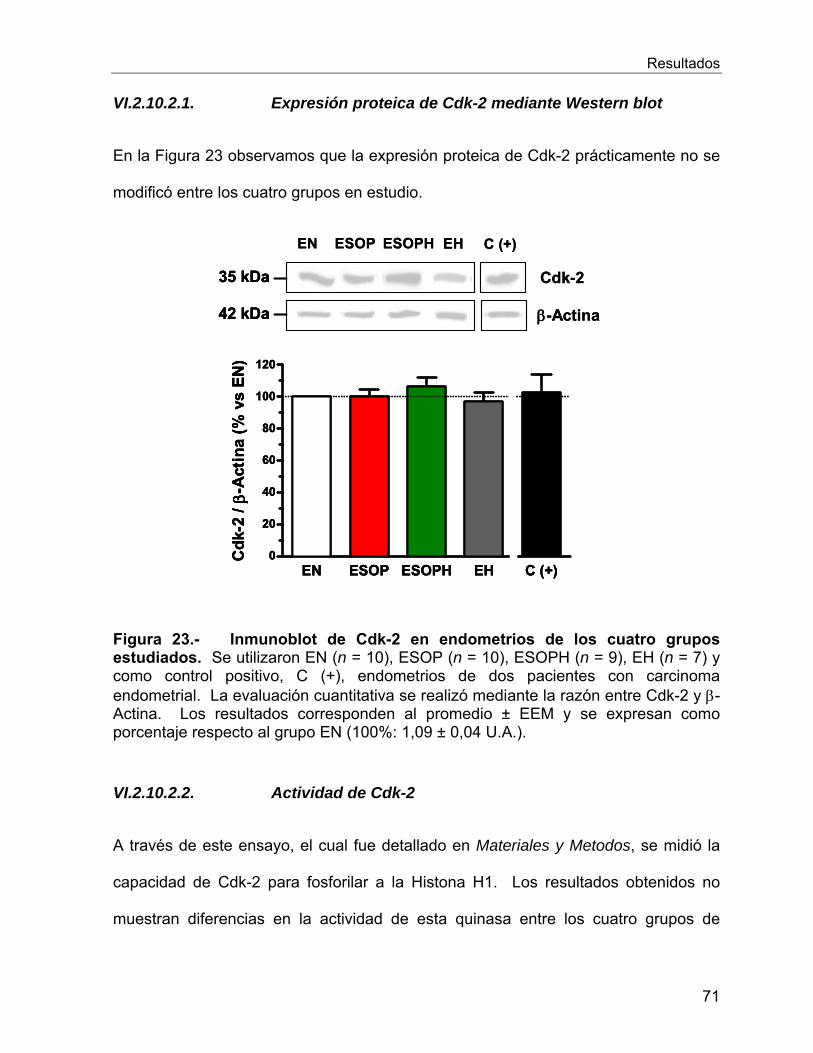
Resultados
71
VI.2.10.2.1. Expresión proteica de Cdk-2 mediante Western blot
En la Figura 23 observamos que la expresión proteica de Cdk-2 prácticamente no se
modificó entre los cuatro grupos en estudio.
Figura 23.- Inmunoblot de Cdk-2 en endometrios de los cuatro grupos estudiados. Se utilizaron EN (n = 10), ESOP (n = 10), ESOPH (n = 9), EH (n = 7) y como control positivo, C (+), endometrios de dos pacientes con carcinoma endometrial. La evaluación cuantitativa se realizó mediante la razón entre Cdk-2 y β-Actina. Los resultados corresponden al promedio ± EEM y se expresan como porcentaje respecto al grupo EN (100%: 1,09 ± 0,04 U.A.).
VI.2.10.2.2. Actividad de Cdk-2
A través de este ensayo, el cual fue detallado en Materiales y Metodos, se midió la
capacidad de Cdk-2 para fosforilar a la Histona H1. Los resultados obtenidos no
muestran diferencias en la actividad de esta quinasa entre los cuatro grupos de
EN ESOP ESOPH EH C (+)0
20
40
60
80
100
120
Cdk
-2 /β-
Act
ina
(% v
s EN
)
EN ESOP ESOPH EH C (+)0
20
40
60
80
100
120
Cdk
-2 /β-
Act
ina
(% v
s EN
)
EN ESOP ESOPH EH C (+)0
20
40
60
80
100
120
Cdk
-2 /β-
Act
ina
(% v
s EN
)
Cdk-2
β-Actina
EN ESOP ESOPH EH C (+)
42 kDa
35 kDa Cdk-2
β-Actina
EN ESOP ESOPH EH C (+)
42 kDa42 kDa
35 kDa35 kDa
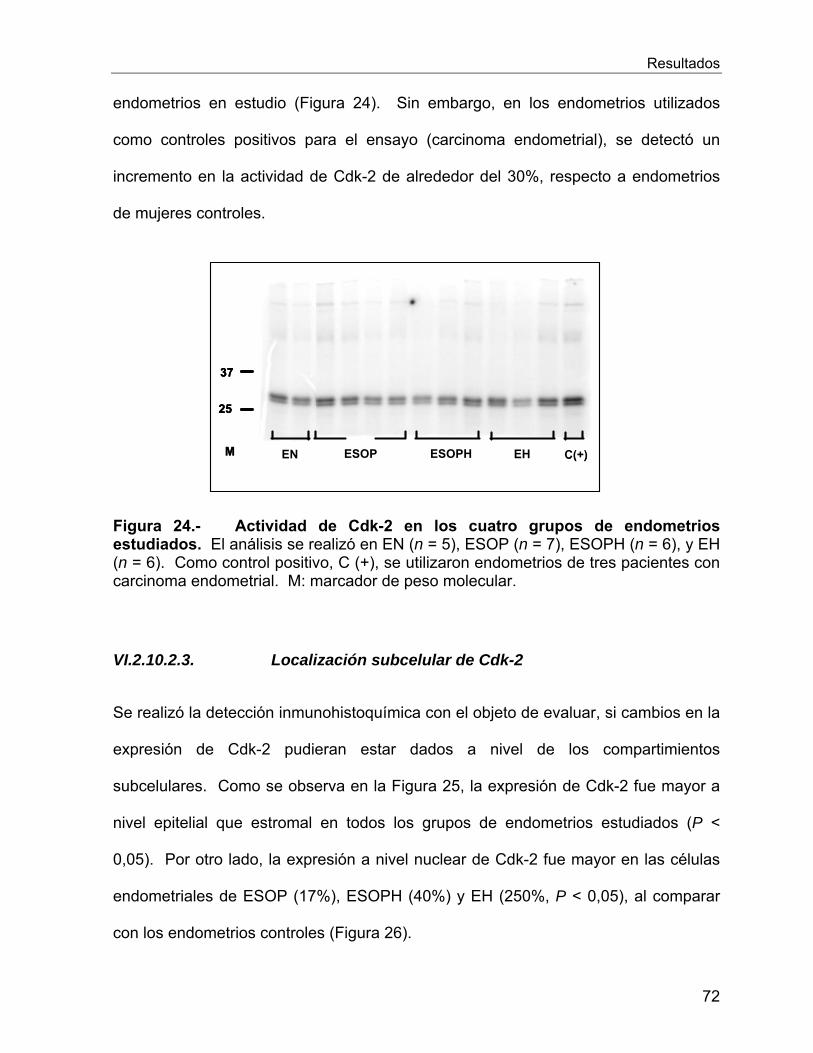
Resultados
72
endometrios en estudio (Figura 24). Sin embargo, en los endometrios utilizados
como controles positivos para el ensayo (carcinoma endometrial), se detectó un
incremento en la actividad de Cdk-2 de alrededor del 30%, respecto a endometrios
de mujeres controles.
Figura 24.- Actividad de Cdk-2 en los cuatro grupos de endometrios estudiados. El análisis se realizó en EN (n = 5), ESOP (n = 7), ESOPH (n = 6), y EH (n = 6). Como control positivo, C (+), se utilizaron endometrios de tres pacientes con carcinoma endometrial. M: marcador de peso molecular.
VI.2.10.2.3. Localización subcelular de Cdk-2
Se realizó la detección inmunohistoquímica con el objeto de evaluar, si cambios en la
expresión de Cdk-2 pudieran estar dados a nivel de los compartimientos
subcelulares. Como se observa en la Figura 25, la expresión de Cdk-2 fue mayor a
nivel epitelial que estromal en todos los grupos de endometrios estudiados (P <
0,05). Por otro lado, la expresión a nivel nuclear de Cdk-2 fue mayor en las células
endometriales de ESOP (17%), ESOPH (40%) y EH (250%, P < 0,05), al comparar
con los endometrios controles (Figura 26).
25
37
EN ESOP ESOPH EH C(+)M
25
37
25
37
EN ESOP ESOPH EH C(+)M
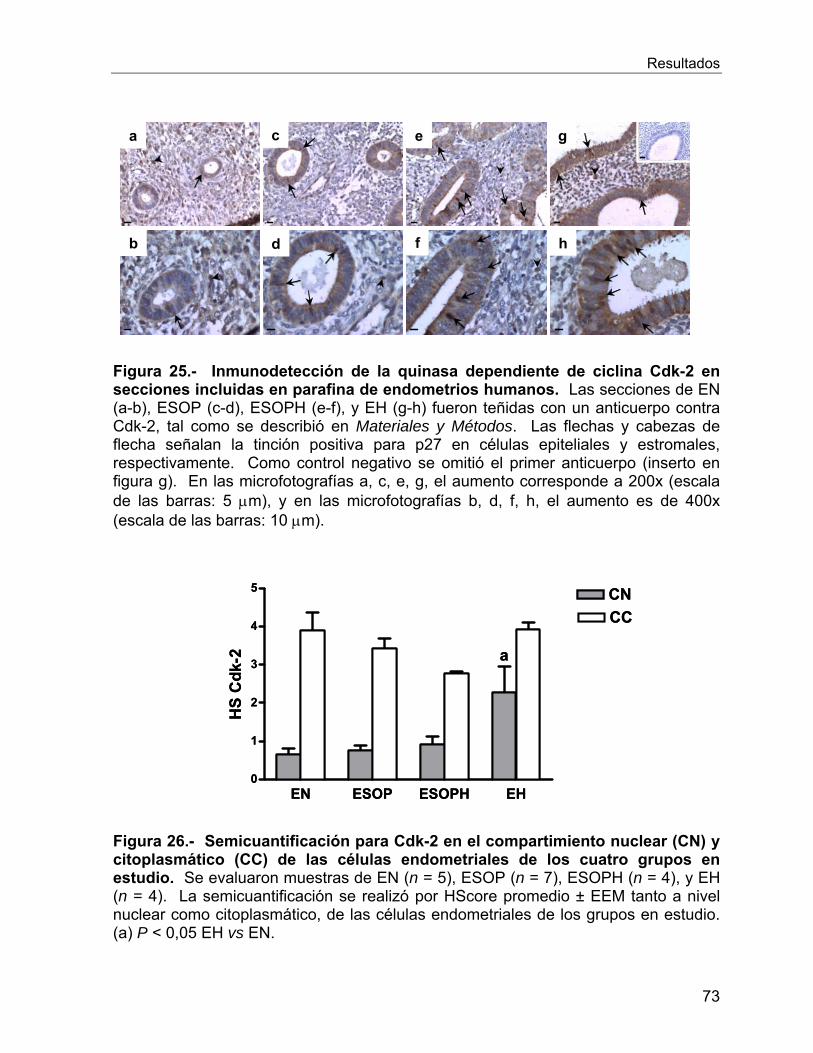
Resultados
73
Figura 25.- Inmunodetección de la quinasa dependiente de ciclina Cdk-2 en secciones incluidas en parafina de endometrios humanos. Las secciones de EN (a-b), ESOP (c-d), ESOPH (e-f), y EH (g-h) fueron teñidas con un anticuerpo contra Cdk-2, tal como se describió en Materiales y Métodos. Las flechas y cabezas de flecha señalan la tinción positiva para p27 en células epiteliales y estromales, respectivamente. Como control negativo se omitió el primer anticuerpo (inserto en figura g). En las microfotografías a, c, e, g, el aumento corresponde a 200x (escala de las barras: 5 μm), y en las microfotografías b, d, f, h, el aumento es de 400x (escala de las barras: 10 μm).
Figura 26.- Semicuantificación para Cdk-2 en el compartimiento nuclear (CN) y citoplasmático (CC) de las células endometriales de los cuatro grupos en estudio. Se evaluaron muestras de EN (n = 5), ESOP (n = 7), ESOPH (n = 4), y EH (n = 4). La semicuantificación se realizó por HScore promedio ± EEM tanto a nivel nuclear como citoplasmático, de las células endometriales de los grupos en estudio. (a) P < 0,05 EH vs EN.
EN ESOP ESOPH EH0
1
2
3
4
5 CNCC
HS
Cdk
-2 a
EN ESOP ESOPH EH0
1
2
3
4
5 CNCC
HS
Cdk
-2 a
a
b
c
d
e
f
g
h
a
b
c
d
e
f
g
h

Resultados
74
VI.2.10.3. Expresión de Ciclina E
La Ciclina E es importante porque forma complejo con Cdk-2 para permitir la
progresión hacia la fase S del ciclo celular. En este trabajo se estudió la expresión
proteica de esta Ciclina mediante Western blot e inmunohistoquímica.
VI.2.10.3.1. Expresión proteica de Ciclina E mediante Western blot
La Figura 27 muestra los niveles de expresión proteica de Ciclina E en endometrios
obtenidos de los cuatro grupos estudiados. Los resultados muestran que no existen
diferencias en la expresión de Ciclina E entre los distintos grupos de pacientes
comparados con el grupo de voluntarias normales. Por otro lado, existe una
tendencia a una mayor expresión de Ciclina E en ESOP (13% vs EN), sin
evidenciarse diferencias en la expresión de esta proteína entre pacientes SOP
insulino-resistentes y no insulino-resistentes.
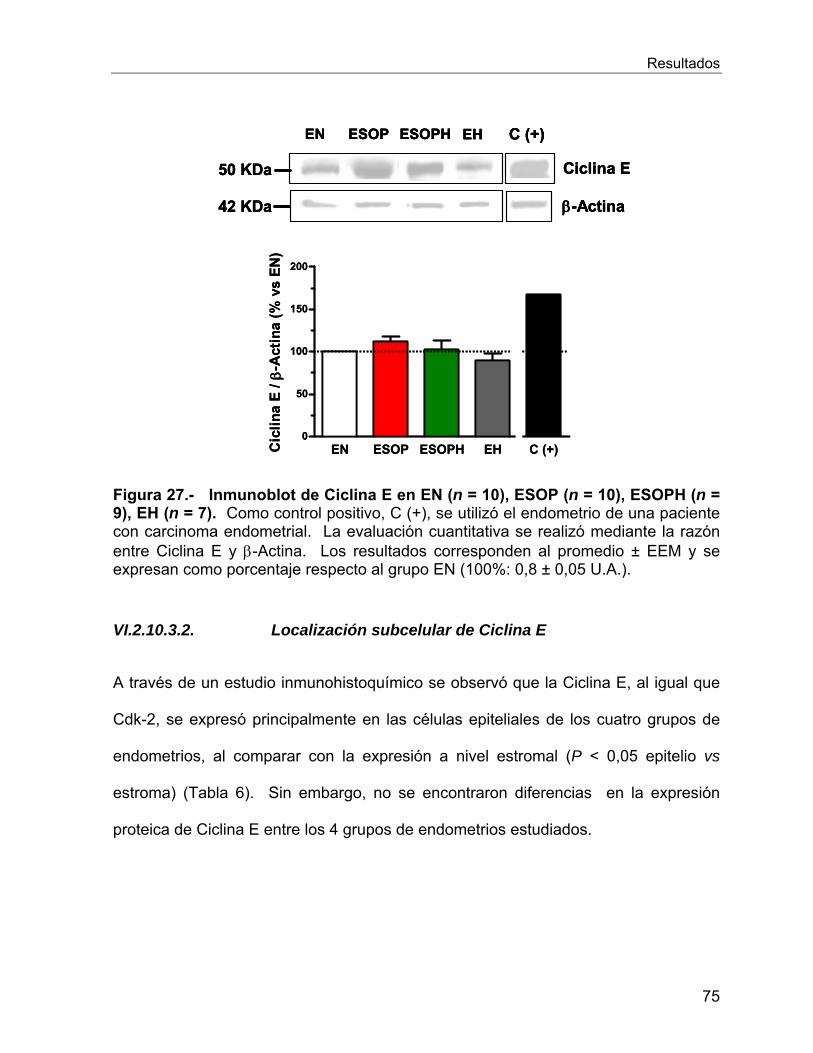
Resultados
75
Figura 27.- Inmunoblot de Ciclina E en EN (n = 10), ESOP (n = 10), ESOPH (n = 9), EH (n = 7). Como control positivo, C (+), se utilizó el endometrio de una paciente con carcinoma endometrial. La evaluación cuantitativa se realizó mediante la razón entre Ciclina E y β-Actina. Los resultados corresponden al promedio ± EEM y se expresan como porcentaje respecto al grupo EN (100%: 0,8 ± 0,05 U.A.).
VI.2.10.3.2. Localización subcelular de Ciclina E
A través de un estudio inmunohistoquímico se observó que la Ciclina E, al igual que
Cdk-2, se expresó principalmente en las células epiteliales de los cuatro grupos de
endometrios, al comparar con la expresión a nivel estromal (P < 0,05 epitelio vs
estroma) (Tabla 6). Sin embargo, no se encontraron diferencias en la expresión
proteica de Ciclina E entre los 4 grupos de endometrios estudiados.
Ciclina E
β-Actina
EN ESOP ESOPH EH C (+)
50 KDa
42 KDa
Ciclina E
β-Actina
EN ESOP ESOPH EH C (+)
50 KDa
42 KDa
Ciclina E
β-Actina
EN ESOP ESOPH EH C (+)
50 KDa
42 KDa
EN ESOP ESOPH EH C (+)0
50
100
150
200C
iclin
a E
/ β-A
ctin
a (%
vs
EN)
EN ESOP ESOPH EH C (+)0
50
100
150
200C
iclin
a E
/ β-A
ctin
a (%
vs
EN)

Resultados
76
Tabla 6.- Semicuantificación mediante HScore (HS) de la expresión de Ciclina E en el compartimiento epitelial (CE) y estromal (CS) de EN (n = 5), ESOP (n = 5), ESOPH (n = 4), y EH (n = 4). aP < 0,05 CE vs CS.
VI.2.10.4. Expresión de la proteína del Retinoblastoma (Rb)
VI.2.10.4.1. Expresión proteica de Rb por Western Blot
En la Figura 28 observamos que la expresión proteica de Rb total, prácticamente no
se modificó entre los cuatro grupos en estudio, aunque en el grupo ESOP, ESOPH y
EH dicha expresión se incrementó levemente (10, 15 y 7%, respectivamente)
respecto a los endometrios normales en fase proliferativa. Además, no se observaron
diferencias en la expresión de Rb entre pacientes SOP insuino-resistentes y no
insulino-resitentes. Como control positivo, se utilizó tejido endometrial de pacientes
con carcinoma endometrial, donde se observó un 14% de aumento en la expresión
de Rb, respecto a EN.
0,34 ± 0,171,64 ± 0,42 aEH
0,04 ± 0,020,82 ± 0,32 aESOPH
0,22 ± 0,071,06 ± 0,19 aESOP
0,33 ± 0,070,72 ± 0,20 aEN
CSCEHS
0,34 ± 0,171,64 ± 0,42 aEH
0,04 ± 0,020,82 ± 0,32 aESOPH
0,22 ± 0,071,06 ± 0,19 aESOP
0,33 ± 0,070,72 ± 0,20 aEN
CSCEHS
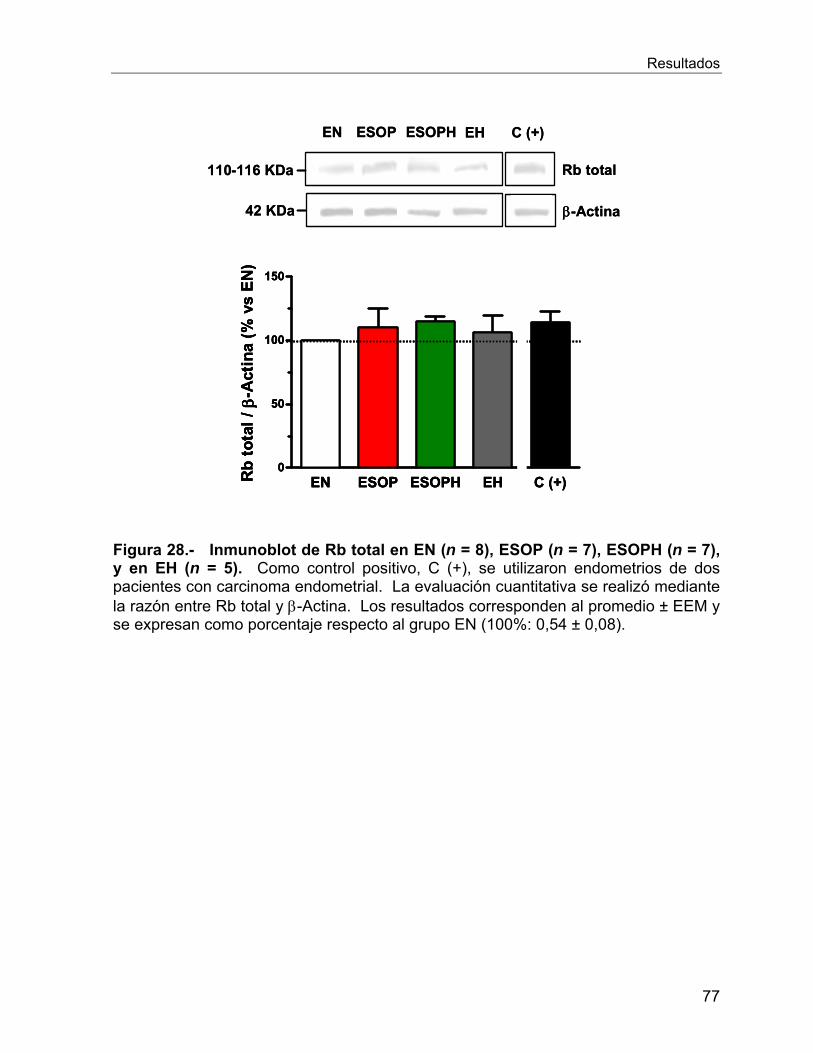
Resultados
77
Figura 28.- Inmunoblot de Rb total en EN (n = 8), ESOP (n = 7), ESOPH (n = 7), y en EH (n = 5). Como control positivo, C (+), se utilizaron endometrios de dos pacientes con carcinoma endometrial. La evaluación cuantitativa se realizó mediante la razón entre Rb total y β-Actina. Los resultados corresponden al promedio ± EEM y se expresan como porcentaje respecto al grupo EN (100%: 0,54 ± 0,08).
EN ESOP ESOPH EH C (+)0
50
100
150
Rb
tota
l /β-
Act
ina
(% v
s EN
)
EN ESOP ESOPH EH C (+)0
50
100
150
Rb
tota
l /β-
Act
ina
(% v
s EN
)
EN ESOP ESOPH EH C (+)0
50
100
150
Rb
tota
l /β-
Act
ina
(% v
s EN
)
Rb total
β-Actina
EN ESOP ESOPH EH C (+)
110-116 KDa
42 KDa
Rb total
β-Actina
EN ESOP ESOPH EH C (+)
110-116 KDa
42 KDa

Resultados
78
VI.3 ESTUDIOS IN VITRO
La alteración endocrina central de las pacientes SOP es el hiperandrogenismo; sin
embargo, el hiperestrogenismo relativo y el hiperinsulinismo son también
características de un porcentaje importante de estas mujeres. Por esta razón, se
estudió la acción de androstenediona, estradiol e insulina sobre la expresión proteica
del co-activador AIB1 y de marcadores del ciclo celular como: Ciclina E, Cdk-2, p27 y
Rb. Para ello, explantes de tejido endometrial en fase proliferativa provenientes de
mujeres controles, fueron cultivados por 18 h en presencia de estos factores.
VI.3.1 Evaluación morfológica del tejido endometrial posterior al cultivo
A fin de determinar si la integridad estructural de los explantes de endometrio se
modificaba luego de 18 h de cultivo, se realizó un estudio morfológico del tejido. Se
observó que la estructura histológica de los endometrios cultivados no se ve afectada
luego de 18 h de cultivo a 37ºC, ya que el tejido mantiene su arquitectura
característica a la de un endometrio en fase proliferativa del ciclo menstrual.
VI.3.2 Acción in vitro de concentraciones elevadas de androstenediona,
estradiol y/o insulina sobre la expresión del co-activador AIB1
VI.3.2.1. Expresión proteica del co-activador AIB1 por inmunohistoquímica
Luego de 18 h de cultivo con androstenediona (10-5 y 10-6 M) o estradiol (10-6 y 10-8
M), prácticamente no se modificó la expresión proteica de AIB1 al comparar con la
condición basal. Lo mismo se observó al incubar con insulina (10-6 M). Sin embargo,
en presencia de androstenediona, estradiol e insulina, se detectó un incremento de

Resultados
79
74% en la expresión proteica de AIB1; situación similar a la observada en
endometrios obtenidos de pacientes SOP en los estudios ex vivo (Tabla 7).
Tabla 7.- Efecto de androstenediona, estradiol y/o insulina sobre los niveles proteicos del co-activador AIB1
Los explantes de endometrio (n = 4) fueron cultivados por 18 h en presencia de distintas concentraciones de androstenediona, estradiol e insulina. Los resultados se expresan como porcentaje de células positivas para AIB1 (promedio ± EEM).
43,9 ± 18,3Androstenediona + Estradiol + Insulina (10-6M)
28,3 ± 13,9Insulina (10-6M)
18,6 ± 6,4Estradiol (10-8M)
27,3 ± 11,3Estradiol (10-6M)
23,0 ± 8,1Androstenediona (10-6M)
26,6 ± 8,3Androstenediona (10-5M)
25,3 ± 7,9Basal
% células epiteliales positivasAIB1
43,9 ± 18,3Androstenediona + Estradiol + Insulina (10-6M)
28,3 ± 13,9Insulina (10-6M)
18,6 ± 6,4Estradiol (10-8M)
27,3 ± 11,3Estradiol (10-6M)
23,0 ± 8,1Androstenediona (10-6M)
26,6 ± 8,3Androstenediona (10-5M)
25,3 ± 7,9Basal
% células epiteliales positivasAIB1

Resultados
80
VI.3.3 Acción in vitro de concentraciones elevadas de androstenediona,
estradiol y/o insulina sobre la expresión de reguladores del ciclo celular
VI.3.3.1. Expresión proteica de Ciclina E, Cdk-2, p27 y Rb por Western blot
Tal como se muestra en la Tabla 8, se observó que en presencia de androstenediona
y estradiol prácticamente no se modifica la expresión de las proteínas estudiadas.
Sin embargo, en presencia de insulina (10-7M), disminuye levemente la expresión de
p27, tal como se observó en los endometrios SOP de pacientes insulino-resistentes
en los estudios ex vivo. Además, a la menor concentración probada para insulina se
detectó un incremento del 15% en la expresión de Ciclina E y una disminución del
12% en la expresión de Rb. Por otro lado, la expresión proteica de Cdk-2 no
experimentó modificaciones en presencia de insulina.
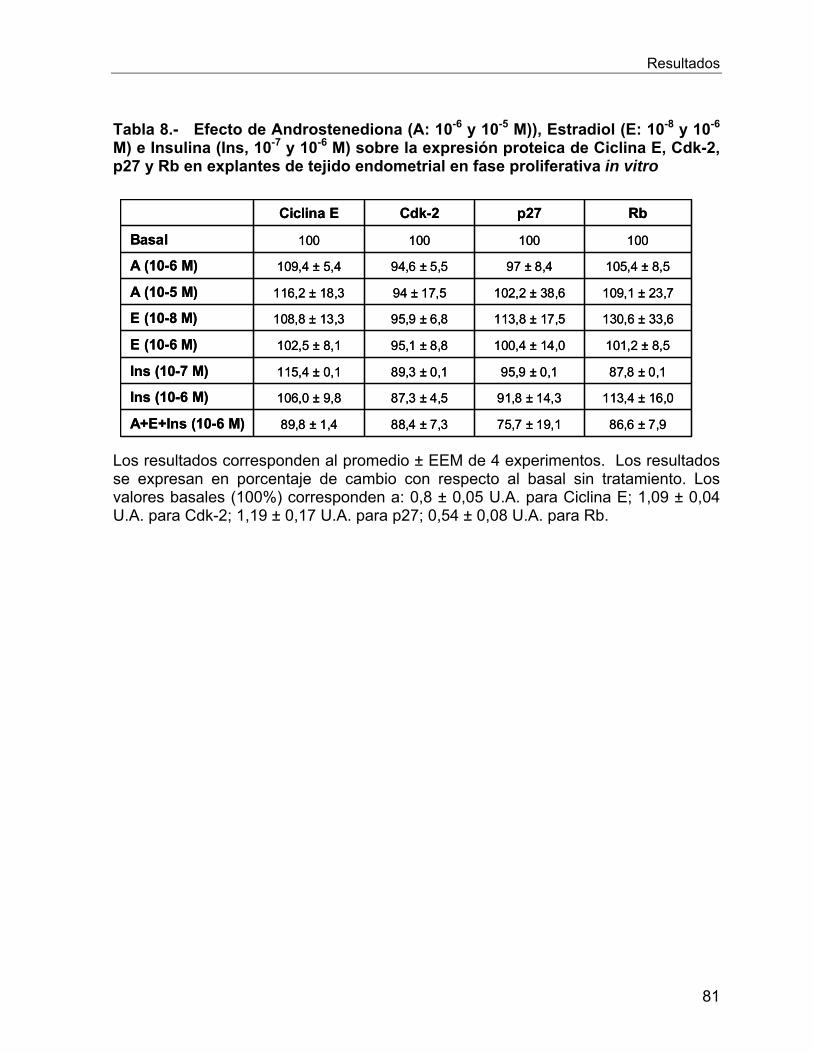
Resultados
81
Tabla 8.- Efecto de Androstenediona (A: 10-6 y 10-5 M)), Estradiol (E: 10-8 y 10-6 M) e Insulina (Ins, 10-7 y 10-6 M) sobre la expresión proteica de Ciclina E, Cdk-2, p27 y Rb en explantes de tejido endometrial en fase proliferativa in vitro
Los resultados corresponden al promedio ± EEM de 4 experimentos. Los resultados se expresan en porcentaje de cambio con respecto al basal sin tratamiento. Los valores basales (100%) corresponden a: 0,8 ± 0,05 U.A. para Ciclina E; 1,09 ± 0,04 U.A. para Cdk-2; 1,19 ± 0,17 U.A. para p27; 0,54 ± 0,08 U.A. para Rb.
86,6 ± 7,975,7 ± 19,188,4 ± 7,389,8 ± 1,4A+E+Ins (10-6 M)
113,4 ± 16,091,8 ± 14,387,3 ± 4,5106,0 ± 9,8Ins (10-6 M)
87,8 ± 0,195,9 ± 0,189,3 ± 0,1115,4 ± 0,1Ins (10-7 M)
101,2 ± 8,5100,4 ± 14,095,1 ± 8,8102,5 ± 8,1E (10-6 M)
130,6 ± 33,6113,8 ± 17,595,9 ± 6,8108,8 ± 13,3E (10-8 M)
109,1 ± 23,7102,2 ± 38,694 ± 17,5116,2 ± 18,3A (10-5 M)
105,4 ± 8,597 ± 8,494,6 ± 5,5109,4 ± 5,4A (10-6 M)
100100100100Basal
Rbp27Cdk-2Ciclina E
86,6 ± 7,975,7 ± 19,188,4 ± 7,389,8 ± 1,4A+E+Ins (10-6 M)
113,4 ± 16,091,8 ± 14,387,3 ± 4,5106,0 ± 9,8Ins (10-6 M)
87,8 ± 0,195,9 ± 0,189,3 ± 0,1115,4 ± 0,1Ins (10-7 M)
101,2 ± 8,5100,4 ± 14,095,1 ± 8,8102,5 ± 8,1E (10-6 M)
130,6 ± 33,6113,8 ± 17,595,9 ± 6,8108,8 ± 13,3E (10-8 M)
109,1 ± 23,7102,2 ± 38,694 ± 17,5116,2 ± 18,3A (10-5 M)
105,4 ± 8,597 ± 8,494,6 ± 5,5109,4 ± 5,4A (10-6 M)
100100100100Basal
Rbp27Cdk-2Ciclina E

Discusión
82
VII. DISCUSION
Los cambios cíclicos en el tracto genital femenino requieren remodelación del tejido
endometrial, lo cual está relacionado con las variaciones hormonales durante el ciclo
menstrual. Estos cambios llevan a una modificación de la arquitectura endometrial, a
fin de permitir el éxito del proceso reproductivo. Así, durante la fase folicular ovárica
(o fase proliferativa en el endometrio), el estradiol producido por los folículos en
desarrollo induce la proliferación celular en el endometrio, y luego de la ovulación, la
progesterona producida por el cuerpo lúteo durante la fase lútea, promueve la
diferenciación de las células endometriales (fase secretora).
Ha sido descrito por nuestro grupo y otros autores (104), que el endometrio en fase
secretora experimenta un aumento en el número de células que mueren, en parte
debido a apoptosis de las células endometriales. Este proceso de muerte celular
programada, así como el proceso de proliferación celular, son de gran relevancia
fisiológica pues son fundamentales para garantizar la homeostasis endometrial. En
todos estos procesos participan esteroides sexuales, receptores esteroidales, co-
reguladores, proteínas involucradas en la proliferación y apoptosis, y reguladores del
ciclo celular.
Considerando que el endometrio es un tejido cíclico, donde la acción regulatoria de
los esteroides sexuales es tan importante, es lógico pensar que en situaciones
endocrino-metabólicas asociadas a anormalidades en la concentración sanguínea de
algunas hormonas y proteínas, como es el caso del Síndrome de Ovario Poliquístico

Discusión
83
(SOP), los procesos que ocurren en el endometrio y la forma como las células
responden a los estímulos biológicos alterados, pueden verse afectados.
Como se mencionó anteriormente, el endometrio es un tejido cuya función es
regulada fundamentalmente por esteroides. La señalización en respuesta a la acción
esteroidal se realiza a través de receptores esteroidales, lo cual es un proceso
complejo que involucra la interacción de múltiples co-reguladores. De lo anterior se
desprende que la expresión alterada de estos receptores esteroidales y co-
reguladores puede llevar a cambios en la expresión génica, así como en la
progresión del ciclo celular (105), lo que puede traducirse en un descontrol del
crecimiento celular.
Estudios previos han examinado la expresión de receptores esteroidales y co-
reguladores en endometrios obtenidos de mujeres normales (45, 48, 106, 107),
pacientes SOP (48), y en carcinoma endometrial (45). Sin embargo, este es el
primer estudio que compara la expresión de receptores esteroidales y co-reguladores
entre endometrios normales y endometrios de mujeres SOP sin tratamiento (sin y con
hiperplasia endometrial). Los resultados del presente trabajo muestran que la
expresión génica y proteica del RA se encuentra relativamente elevada en
endometrios de mujeres SOP sin tratamiento, al compararlos con endometrios
normales de la fase proliferativa del ciclo menstrual. Resultados similares fueron
reportados por Apparao y cols. (41) en mujeres SOP tratadas con citrato de
clomifeno. Por otro lado, también se observa un incremento en la expresión del RA
en las pacientes SOP con hiperplasia endometrial al comparar con los endometrios
controles. Estos resultados pueden explicarse, en parte, por una acción

Discusión
84
hiperestrogénica no opuesta por progesterona a nivel endometrial; condición propia
del SOP. Se sabe que el RA es regulado en forma positiva por andrógenos y
estrógenos, y en forma negativa por progesterona (36, 37, 41), lo que sugiere un
efecto sinérgico de los andrógenos con los estrógenos en la regulación del RA.
Es ampliamente conocido que los genes para el RE son genes de respuesta a
estrógenos, los cuales aumentan su expresión luego del tratamiento con estrógenos,
tal como fue reportado en endometrio humano y de primate (36, 38, 103). En esta
tesis se observa un incremento significativo de la expresión proteica del REα en
endometrios de mujeres SOP sin hiperplasia, principalmente en el núcleo de las
células endometriales. En cuanto a la isoforma β del RE, a través de
inmunohistoquímica, observamos que su expresión proteica aumenta en forma
gradual desde ESOP a ESOPH y EH, principalmente en las células epiteliales. Al
respecto, Weihua y cols. (108) reportaron que la isoforma β del RE se encuentra
involucrada en la regulación del crecimiento epitelial, tal como fue demostrado en
ratones knockout para la isoforma β del RE (REβ-/-). Además, en la presente
investigación se evidenció un aumento de la expresión proteica del REβ en las
células estromales de ESOP, ESOPH, y EH. De manera interesante, se obtuvo un
incremento gradual en la razón REβ/REα, tanto a nivel génico como proteico, entre
los cuatro grupos de mujeres estudiadas. De los resultados expuestos
anteriormente, podemos inferir que el desbalance en la razón REβ/REα podría ser un
paso crítico en la progresión a hiperplasia y carcinoma endometrial dependiente de
estrógenos en las mujeres SOP, y que vías específicas para el REα y el REβ pueden
tener distintos roles en este proceso. Esta proposición está basada en resultados de

Discusión
85
varios estudios que muestran una expresión disminuida del REβ en algunos cánceres
(mama, ovario, colon, próstata), al comparar con tumores benignos o tejidos
normales, mientras que la expresión del REα persiste (109-115).
Recientemente, dos trabajos en cáncer de mama han sugerido que el REβ puede
disminuir la proliferación celular, a través de la inhibición de la expresión de Ciclina
D1 y Ciclina E (116, 117). Por otro lado, estudios in vitro en células HeLa corroboran
la hipótesis de Liu y cols. (118), referida a que estradiol activa la transcripción del gen
de ciclina D1 a través del REα, e inhibe dicha transcripción a través del REβ.
Además, se ha sugerido que el REα puede desempeñar un rol en la carcinogénesis
ovárica previniendo la apoptosis, mientras que la inhibición de la proliferación
inducida por el REβ podría explicarse por la inhibición de la expresión génica de bcl-2
(119). Estos estudios concuerdan con un reporte previo de nuestro laboratorio donde
observamos que la expresión elevada del REα en endometrios de mujeres SOP,
puede estar relacionada con la alta expresión proteica de Bcl-2 en las mismas
pacientes (30). Por lo tanto, el REα podría estar ejerciendo un efecto no protector,
en lo que respecta al desarrollo de hiperplasia en los endometrios SOP. También, es
importante considerar que otras proteínas, tales como onco-proteínas y proteínas
supresoras de tumores, podrían estar actuando en forma no protectora a nivel del
endometrio de estas pacientes.
La actividad transcripcional de receptores esteroidales, tales como RA y RE, se
encuentra influenciada por co-activadores y co-represores, los cuales activan o
reprimen la transcripción de genes en respuesta a estrógenos o andrógenos (120,
121). Estudios previos han mostrado que el co-activador AIB1 se encuentra

Discusión
86
amplificado y sobre-expresado en cáncer de ovario, de mama (122), hígado (123) y
estómago (124). Por otro lado, Torres-Arzayus y cols. (125) reportaron que la sobre-
expresión de AIB1 en ratones transgénicos, conduce a hipertrofia e hiperplasia
mamaria, y al desarrollo de tumores mamarios malignos. Dichos tumores también
aparecen en hipófisis y útero. Los resultados del presente trabajo muestran un
incremento en la expresión proteica de AIB1 a nivel nuclear en endometrios SOP al
comparar con los endometrios controles, situación que también se observa en los
estudios in vitro en presencia de una alta concentración de andrógenos, estrógenos e
insulina. Estos cambios en la expresión proteica de AIB1 van acompañados por una
alta expresión del co-activador ARA70 en los cuatro grupos de endometrios
estudiados. Cabe destacar que la alta expresión de ARA70 en endometrios de
pacientes SOP con hiperplasia, junto con el nivel de expresión normal de AIB1,
indicaría que ARA70 podría incrementar la sensibilidad a los esteroides en este
grupo de endometrios. Por otro lado, la localización citoplasmática de ARA70
sugiere que este co-activador actúa de una forma diferente a los co-activadores
clásicos, que se unen a los receptores esteroidales activados por el ligando y a los
factores de la maquinaria de transcripción basal (126), y por ende, ARA70 puede
representar una nueva clase de co-activador que actúa en el citoplasma (127). Al
respecto, se sabe que en el citoplasma ARA70 aumenta la estabilidad proteica del
RA unido al ligando, y que este co-activador permanece en el citoplasma una vez
que el complejo hormona-receptor se transloca al núcleo (120). No obstante, es
posible considerar que otros co-activadores puedan también contribuir a la acción de
AIB1 y ARA70 en los grupos de endometrios estudiados.

Discusión
87
La expresión nuclear de AIB1 encontrada en esta tesis concuerda con los resultados
de un reporte previo que muestra que este co-activador se localizada principalmente
en el núcleo de células mitóticas, y en el citoplasma de fibroblastos quiescentes
(128); lo que provee una importante correlación entre la localización subcelular y el
estado mitótico de la célula. En este contexto, resultados del presente trabajo que
serán discutidos más adelante, muestran un alto grado de proliferación en los
endometrios de mujeres SOP al comparar con los endometrios controles, lo cual
coincide con la localización nuclear del co-activador AIB1.
En cuanto al co-represor NCoR, su expresión fue similar entre los cuatro grupos de
pacientes, y estuvo principalmente localizado en el citoplasma de las células
endometriales. Esta localización subcelular sugiere que este NCoR no está
cumpliendo con su rol represor de la acción transcripcional de los receptores
esteroidales, en los endometrios de las pacientes en estudio, tal como fue reportado
previamente en neuronas (129).
En el presente estudio no se encontraron diferencias en la expresión de receptores
esteroidales y co-reguladores entre los endometrios de pacientes SOP con y sin
insulino-resistencia, lo que indica que probablemente la insulina no es un regulador
importante de la expresión endometrial de estos marcadores.
De los resultados discutidos hasta el momento, se puede postular que como el
balance entre co-activadores y co-represores es importante en la regulación final de
la transcripción, la expresión nuclear aumentada de AIB1 junto con el incremento en
la expresión del RE y RA en endometrios SOP, sumado a la alta expresión de ARA70
y a la localización citoplasmática de NCoR en todos los grupos de endometrios

Discusión
88
estudiados, podrían dar al endometrio de estas mujeres una mayor sensibilidad a la
acción esteroidal. Además, los cambios observados en los receptores esteroidales y
co-reguladores en endometrios SOP, podrían conducir a cambios en la expresión de
genes involucrados en la progresión del ciclo celular en endometrios de pacientes
SOP con hiperplasia. Entonces, pensamos que el ambiente hormonal anormal en las
pacientes SOP, podría des-regular la transcripción de genes involucrados en el
control del ciclo celular, y estas alteraciones podrían conducir al desarrollo de
hiperplasia endometrial en estas mujeres.
Además de los cambios mencionados anteriormente en la expresión de receptores
esteroidales y co-reguladores y sus potenciales consecuencias en el desarrollo de
hiperplasia endometrial en las mujeres SOP, también es importante considerar la
disponibilidad in situ de los andrógenos y estrógenos, lo cual depende en gran
medida de la expresión y actividad de numerosas enzimas. En esta tesis, se estudió
a la enzima P450 aromatasa, la cual convierte los andrógenos a estrógenos. Dicha
conversión ocurre principalmente en tejidos periféricos, incluyendo piel, músculo,
tejido adiposo y hueso. Así, la mayoría de la estrona formada a partir de andrógenos
por la actividad de la aromatasa en estos tejidos periféricos, es convertida a estrona
sulfato por la acción de la enzima sulfotransferasa, la que también está presente en
estos tejidos (130). Resultados de este trabajo indican que la proteína de la
aromatasa no se expresa en endometrio normal, lo que está de acuerdo con
antecedentes de otros autores, quienes reportaron que la actividad de esta enzima
no es detectable en endometrio normal (131). Además, este es el primer estudio que
analiza la expresión de esta enzima en endometrios de pacientes SOP sin y con

Discusión
89
hiperplasia, observándose ausencia de la misma en estos endometrios. También,
observamos su ausencia en endometrios con hiperplasia, tal como fue reportado
previamente (132). Sin embargo, la expresión génica y proteica de la enzima
aromatasa ha sido determinada en carcinoma endometrial (132). En relación a lo
anterior, existen reportes que muestran que en endometrio humano neoplásico, la
testosterona puede aromatizarse a estradiol, lo que conduce a un aumento in situ de
la concentración de estrógenos (131). Por lo tanto, será necesario realizar estudios
de actividad de esta enzima, a fin de elucidar si P450 aromatasa aumenta la
biodisponibilidad de estrógenos a nivel endometrial en las pacientes SOP. No
obstante, si bien existen estudios que señalan que los estrógenos producidos por
aromatización en el tejido, pueden funcionar como un factor mitogénico y de
crecimiento (133, 134), no se puede descartar que una exposición del tejido
endometrial a elevadas concentraciones de estrógenos y andrógenos por periodos
prolongados (como es el caso del SOP), pueden constituir factores de riesgo para
favorecer el desarrollo de hiperplasia y carcinoma endometrial.
Los fenómenos recientemente mencionados pueden desencadenarse, en parte, por
un desbalance entre proliferación y apoptosis, lo cual conduce a una alteración de la
homeostasis endometrial. En este contexto, es importante destacar que a través del
estudio de la proteína Ki67, en el presente trabajo se ha observado un aumento de la
proliferación celular en endometrios de pacientes SOP, en ausencia de apoptosis
(evidenciada a través de la ausencia de caspasa-3-activa y de fragmentación del
DNA por TUNEL). Por lo tanto, parece razonable postular que las pacientes SOP
presentan una desregulación de su homeostasis endometrial. Además, dado que no

Discusión
90
se encontraron diferencias en la proliferación y en la apoptosis entre endometrios de
pacientes SOP insulino-resistentes y no insulino-resistentes, es probable que el
hiperestrogenismo relativo y/o el hiperandrogenismo presente en las pacientes SOP,
puedan ser importantes en el aumento de la proliferación y en la ausencia de
apoptosis en estas mujeres.
Algunos autores han sugerido que la expresión de los receptores esteroidales estaría
relacionada con la regulación de la apoptosis en las células endometriales (89, 135,
136). Así, se ha descrito que estradiol unido a su receptor regula la expresión
endometrial de la proteína anti-apoptótica Bcl-2 (89), lo que está de acuerdo con los
resultados de esta tesis que indican un aumento de la expresión de Bcl-2 en los
endometrios de pacientes SOP (sin y con hiperplasia), quienes presentan un
hiperestrogenismo relativo, tal como se mencionó anteriormente. Por otro lado,
estudios in vitro mostraron que los andrógenos no inducen cambios en la expresión
génica de bax (pro-apoptótico) en células endometriales (101), lo cual coincide con el
perfil de expresión de Bax en endometrios de mujeres SOP sin y con hiperplasia, al
comparar con los endometrios controles. Por lo tanto, pareciera que el desbalance
Bcl2/Bax en los endometrios SOP sin y con hiperplasia, es uno de los
acontecimientos importantes que favorecen la supervivencia celular, previniendo la
apoptosis y aumentando la proliferación celular en estos grupos de endometrios.
Se sabe que la proliferación celular tanto de las células glandulares endometriales
normales como de las células de carcinoma endometrial que expresan receptores de
estrógeno, aumenta por la acción de estrógenos (137). Al respecto, se ha
investigado sobre los mecanismos del crecimiento dependiente de estrógenos de

Discusión
91
estas células, desde varios aspectos que incluyen: regulación positiva de receptores
esteroidales (103, 138), función de las enzimas que metabolizan esteroides (139),
efecto autocrino o paracrino de factores de crecimiento y citoquinas (140); sin
embargo, muy poco se conoce sobre el efecto directo de los estrógenos sobre el
ciclo celular. Recientes hallazgos demostraron que la proliferación inducida por
estradiol de las células epiteliales de endometrio normal, se inicia por activación
transcripcional de la Ciclina D1 a través de la unión de c-Jun a secuencias AP-1 en el
promotor de Ciclina D1 (141). Este proceso controlado, el cual tiene lugar en el
endometrio normal, se encuentra alterado en hiperplasia, pero principalmente en
carcinoma endometrial, lo cual se evidencia por sobreexpresión de Ciclina D1 (142),
Ciclina E, Ciclina B1 y Cdk-2 (143) y disminución o ausencia en la expresión de p27
(144). Sin embargo, no hay estudios que expliquen, a nivel del ciclo celular, la mayor
proliferación observada en los endometrios de las pacientes SOP. Cabe mencionar,
que dado que el SOP es una patología multifactorial que se caracteriza no solo por
un hiperestrogenismo relativo, sino también por hiperandrogenismo e
hiperinsulinismo, es importante considerar además de la posible acción estrogénica a
nivel endometrial, la acción de andrógenos e insulina. En el caso de insulina,
Nagamani y cols., encontraron sitios de unión para la misma en el estroma
endometrial de mujeres pre-menopáusicas y de mujeres con carcinoma endometrial.
Por otro lado, mediante estudios in vitro se demostró que insulina e IGF-1, aceleran
el crecimiento de las células de cáncer de endometrio (145). En lo que respecta a los
andrógenos, se ha demostrado un aumento en la expresión de receptores de
factores de crecimiento en células del estroma estimuladas con andrógenos (69). Sin
embargo, estudios realizados en células del epitelio endometrial han mostrado que

Discusión
92
los andrógenos provocan una disminución en la proliferación celular (70). Estos
antecedentes apoyan la proposición que indica que el rol de los andrógenos es
controversial a nivel endometrial.
Los resultados del presente estudio muestran que Ciclina E, la cual funciona en el
punto de chequeo entre las fases G1 y S del ciclo celular, se encuentra
principalmente localizada en las células epiteliales de los cuatro grupos de
endometrios en estudio, lo cual coincide con lo ya descrito por Milde-Langosch y cols.
(143) en carcinoma endometrial. Por otro lado, la expresión de Ciclina E no se
modifica entre los endometrios estudiados. Tampoco observamos cambios en la
expresión de Ciclina E en los estudios in vitro en presencia de andrógenos y/o
estrógenos, pero sí observamos un leve incremento en su expresión en presencia de
insulina. Sin embargo, dado que no encontramos diferencias en la expresión de
Ciclina E entre pacientes SOP insulino-resistentes y no insulino-resistentes, no se
puede concluir que insulina, a través de la Ciclina E, sea un factor que pueda
contribuir al desarrollo de hiperplasia y/o carcinoma endometrial en estas pacientes.
En conjunto, estos datos sugieren que en los endometrios de mujeres SOP (sin y con
hiperplasia), aún no existe una desregulación en la expresión de esta proteína.
También, es importante considerar que la alta expresión de Ciclina E en carcinoma
endometrial se correlaciona muy bien con el grado del tumor y el mal pronóstico
(146). Por lo tanto, es razonable pensar que dicha sobreexpresión pueda ser un
paso importante en la carcinogénesis endometrial.
Es importante destacar, además, que las células necesitan que Ciclina E se asocie a
Cdk-2, para así poder ingresar a la fase S del ciclo celular. De esta forma, la

Discusión
93
actividad quinasa generada por estos complejos, fosforila a la proteína
retinoblastoma (Rb), lo que resulta en la progresión del ciclo celular (74, 147).
Los datos obtenidos en este trabajo muestran que Cdk-2 se encuentra principalmente
localizada en las células epiteliales, lo que coincide con lo observado para el caso de
ciclina E y Ki67. Lo anterior, sugiere que el complejo Ciclina E/Cdk-2 está
involucrado en la regulación del crecimiento de las células endometriales epiteliales
de los grupos estudiados. Además, a nivel del núcleo de las células endometriales,
la expresión de Cdk-2 aumenta en forma gradual desde ESOP a ESOPH y EH, al
comparar con los controles normales; lo que también indicaría, que esta quinasa
puede estar ejerciendo una acción relacionada con la fosforilación de Rb en este
compartimiento subcelular. En nuestros estudios de actividad de Cdk-2 en
homogenizados, no se observan diferencias entre ESOP, ESOPH y EH al comparar
con los endometrios normales en fase proliferativa. Sin embargo, dicha actividad se
encuentra aumentada en las muestras de carcinoma endometrial, usadas como
control positivo en el presente estudio, lo cual coincide con la alta expresión
reportada para esta quinasa en carcinoma endometrial (143). Una explicación para
estos resultados puede estar basada en los antecedentes reportados por Shiozawa y
cols. (148), quienes mostraron una significativa regulación positiva de Ciclina E y
Cdk-2 en células epiteliales endometriales durante la fase proliferativa. Las muestras
utilizadas en el presente estudio como controles normales, corresponden a
endometrios en fase proliferativa de mujeres en edad fértil, cuyo grado de
proliferación es alto. Aún cuando la proliferación es mayor en las células epiteliales
de endometrios SOP al comparar con EN, es posible que el alto grado de

Discusión
94
proliferación sea una de las razones por las cuales no se evidencian cambios en la
expresión de Ciclina E, y en la expresión y actividad de Cdk-2 en los endometrios
SOP, al comparar con las voluntarias normales. Sin embargo, es relevante destacar
que mientras las mujeres normales expresan Ciclina E y Cdk-2 en sus endometrios
en fase proliferativa del ciclo menstrual, la expresión de estas proteínas disminuye en
fase secretora (149). En cambio, los endometrios de las pacientes SOP siempre
están expresando estos marcadores, dado que la mayoría de estas mujeres no
pueden ciclar por ser amenorreicas u oligomenorreicas.
Por otro lado, se sabe que los complejos Ciclina/Cdk pueden ser inactivados, lo cual
resulta en arresto del ciclo celular y en una disminución de la proliferación celular.
Dentro de estos inhibidores se encuentra p27, del cual se sabe que su expresión se
encuentra disminuida en carcinoma endometrial (150). Además, p27 se expresa
débilmente en endometrios de fase proliferativa y fuertemente en endometrios de
etapa secretora del ciclo menstrual, lo que indicaría que p27 está involucrado en la
supresión del crecimiento endometrial inducida por progesterona (149). Esta
posibilidad está apoyada por el hecho de que en endometrios con hiperplasia
tratados con medroxiprogesterona, se observa un incremento en la
inmunoreactividad para p27 y una disminución en la expresión de Ki67 (149).
Además, recientemente se ha reportado una doble función para p27 de acuerdo al
compartimiento subcelular donde se encuentre (151). En efecto, cuando está
localizado en el núcleo se comporta como un inhibidor de la proliferación celular,
inactivando principalmente al complejo Ciclina E/Cdk-2 (152) en cambio, cuando se
localiza en el citoplasma promueve el ensamblaje de los complejos Ciclina/Cdk,

Discusión
95
inhibe la apoptosis y regula la estructura del citoesqueleto y por ende, la migración
celular (151). En este contexto, observamos una importante reducción en la
expresión nuclear de p27 en las células estromales de pacientes SOP
(específicamente en las insulino-resistentes). Estos hallazgos nos permiten deducir
que la insulina estaría jugando un rol en la regulación de p27, lo cual también fue
evidenciado en los estudios in vitro, donde observamos una tendencia a una menor
expresión de p27 en presencia de insulina. Los datos de este estudio, también
muestran una menor expresión nuclear de p27 en las células estromales de mujeres
SOP con hiperplasia. Además, detectamos un incremento en la expresión
citoplasmática de p27 en las células epiteliales de pacientes SOP no insulino-
resistentes. Estos resultados, pudiesen indicar que a nivel epitelial, p27 favorecería
el ensamblaje del complejo Ciclina E/Cdk-2 e inhibiría la apoptosis en estos grupos
de endometrios, lo cual ha sido detallado anteriormente. Por otro lado, a nivel de las
células estromales, la menor expresión nuclear de p27 en ESOP sugiere una posible
contribución de esta proteína para el progreso hacia la fase S del ciclo celular. Sin
embargo, a nivel de las células estromales observamos una disminución en la
proliferación celular, a través del estudio de Ki67. Esta divergencia, podría estar
indicando que otros inhibidores distintos a p27 pudieran, en parte, ser los
responsables de la menor proliferación observada en las células estromales de los
endometrios de estos cuatro grupos de mujeres.
También, ha sido descrito que estradiol es capaz de regular la distribución subcelular
de p27 (81), por lo que podemos inferir que los cambios observados en la

Discusión
96
localización de p27 en los endometrios de pacientes SOP pueden, parcialmente, ser
el reflejo del hiperestrogenismo relativo que presentan estas mujeres.
De lo planteado anteriormente, es factible pensar que en respuesta a los mismos
estímulos, el mecanismo de crecimiento de las células epiteliales y estromales es
distinto, tal como fue reportado previamente (148,153, 154, 155). Por lo tanto, aún
cuando lo que realmente nos interesa es lo que sucede a nivel de las células
epiteliales, no se puede descartar la interacción entre epitelio y estroma, y la
contribución de lo anterior en la respuesta biológica final.
Mientras en algunos tumores humanos la proteína supresora de tumores Rb se
encuentra mutada, ha sido sugerido que esta inactivación no está involucrada en la
carcinogénesis endometrial (143). Lo anterior está de acuerdo con los resultados
obtenidos en el presente trabajo, donde no observamos cambios importantes en la
expresión proteica de Rb entre los grupos de endometrios estudiados, respecto a los
controles normales. Además, hallazgos previos mostraron una débil expresión de Rb
en células endometriales normales, pero una alta expresión en carcinoma
endometrial (143, 156). Por lo tanto, parece que en carcinoma endometrial, Rb se
encuentra desacoplada de su función inhibitoria del ciclo celular normal. No
obstante, es interesante destacar que en los estudios in vitro realizados en esta tesis,
se observa que en presencia de insulina existe una tendencia a la disminución de
esta proteína supresora de tumores. Sin embargo, no se puede sugerir que lo
anterior podría eventualmente contribuir al desarrollo de hiperplasia y/o carcinoma
endometrial, dado que en los ensayos ex vivo no se detectaron diferencias en la
expresión de Rb entre pacientes SOP insulino-resistentes y no insulino-resistentes.

Discusión
97
En estudios futuros será necesario estudiar en forma específica la fosforilación de Rb
en los grupos de endometrios de interés, a fin de confirmar lo concluido acerca de
estos resultados.
Otra proteína supresora de tumores es PTEN, quien promueve la inhibición de la
quinasa de adhesión focal (FAK) y del señalamiento río abajo de esta molécula (157,
158). Además, PTEN desfosforila al fosfatidil inositol 3, 4,5 trifosfato (PIP3) (159),
inhibiendo así la cascada de PI3K (160, 161). Estas dos vías convergen para
modular procesos específicos como son la organización del citoesqueleto y la
progresión del ciclo celular. En el presente trabajo, se encontró que en los grupos de
endometrios estudiados, no se observaron cambios en la expresión de esta proteína.
No obstante, a través de estos resultados no podemos concluir acerca de la
funcionalidad de PTEN. En este contexto, ha sido previamente reportado que PTEN
es capaz de inhibir la entrada hacia la fase S del ciclo celular, a través de la inhibición
de la fosforilación de la proteína Rb; y que este proceso está mediado por la
capacidad de PTEN para inhibir la vía de las PI3K (162).
La activación de la vía de las PI3K, la cual está mediada por factores de crecimiento
entre los que se encuentra insulina, gatilla múltiples vías de señalización que
conducen a la entrada al ciclo celular (163). Una de estas vías corresponde a la
activación de la serina/treonina kinasa Akt, por fosforilación en los residuos de
Ser473 y/o Thr308 (164). El incremento en la fosforilación de Akt en Ser473,
observado en esta tesis en los endometrios de mujeres SOP sin y con hiperplasia, al
comparar con las voluntarias normales, adquiere especial importancia dado que es
ampliamente conocido que Akt fosforilada es un regulador importante de procesos

Discusión
98
fisiológicos como la apoptosis y la supervivencia celular (165, 166). Además, Akt
activada se encuentra involucrada en el control de la transición G1/S, a través de la
fosforilación de los inhibidores p21 y p27, lo cuál conduce a que ellos adquieran una
localización citoplasmática (167, 168). Así, se ha visto que en cáncer de mama
existe una muy buena correlación entre la activación de Akt y la localización
citoplasmática de p27 (167). Estos reportes coinciden con lo observado por nosotros
en las células epiteliales de endometrios de pacientes SOP sin hiperplasia, donde
observamos una mayor expresión de p27 en el citoplasma. Es importante agregar,
que a través de un reciente estudio se sabe que la fosforilación de Akt en Ser473
cambia a través del ciclo menstrual a nivel endometrial. Además, los estrógenos
pueden jugar un rol en la regulación de estos cambios a través de una vía no
genómica que involucra a la vía de las PI3K (169). Por lo tanto, podemos inferir que
el hiperestrogenismo relativo presente en las pacientes SOP, podría ser uno de los
responsables de la activación de Akt en los endometrios de estas mujeres.
Hasta el momento, no sabemos por qué la fosforilación de Akt en el residuo Thr308
no experimenta cambios entre los cuatro grupos de endometrios estudiados. Sin
embargo, se sabe que PTEN inhibe la activación de Akt a través de la inhibición de la
fosforilación en Ser473 (170), lo que estaría indicando que en los endometrios de las
mujeres SOP sin y con hiperplasia, PTEN no está ejerciendo su acción, además, que
la vía de las PI3K es una de las rutas que conducen al aumento de la supervivencia
de las células endometriales de las pacientes SOP.
Por otro lado, la proteína Ras es capaz de activar a miembros de la familia de las
MAPK que conducen a la estimulación de numerosos factores transcripcionales.

Discusión
99
Como resultado de lo anterior, se expresan proteínas reguladoras del ciclo celular
que permiten la progresión a través del ciclo celular (171). Estos reportes se
relacionan muy bien con los resultados de este trabajo, que muestran una
sobreexpresión de Ras en los endometrios de pacientes SOP sin y con hiperplasia,
principalmente a nivel de las células epiteliales. Además, se sabe que Ras puede
interactuar con la vía de las PI3K (172), y activar a Akt (173); lo que sugiere que la
potencial función de Akt en los endometrios de estas pacientes, podría ser una parte
importante de la señal de supervivencia celular generada por Ras.
En lo que se refiere a los estudios in vitro, tal como fue mostrado anteriormente, no
observamos cambios importantes en condiciones hiperandrogénicas,
hiperestrogénicas y/o hiperinsulinémicas. Es factible pensar que ello puede deberse
a que en 18 h de cultivo es difícil reproducir un síndrome como es el SOP, y que
además, los cambios en la expresión de las proteínas que nos interesaron estudiar
pueden requerir más tiempo para ser visualizados.
Por lo expuesto anteriormente, es indudable que el SOP afecta la función normal del
endometrio, no sólo a nivel del proceso reproductivo, sino principalmente a nivel de la
homeostasis tisular. De modo tal que los endometrios sometidos a ambientes
hormonales anormales, experimentan descoordinación entre proliferación y
apoptosis, constituyendo éste un evento importante que puede conducir al
crecimiento inadecuado o patológico del endometrio de mujeres con SOP.
En consecuencia, la condición endocrina-metabólica inherente al SOP constituye un
excelente ejemplo de la acción anormal de los esteroides sobre la supervivencia
celular. Por ello, es necesario realizar más investigaciones que ayuden en la

Discusión
100
identificación de marcadores tempranos de hiperplasia y cáncer de endometrio, así
como en la localización de blancos terapéuticos en estas pacientes. Lo anterior,
adquiere especial relevancia cuando pensamos que se trata de mujeres en edad fértil
que, además de padecer trastornos endocrino-metabólicos y reproductivos, tienen
riesgo de desarrollar cáncer endometrial si no son tratadas a tiempo.

Conclusiones
101
VIII. CONCLUSIONES
El endometrio SOP presenta una mayor sensibilidad a la acción esteroidal,
que puede conducir a cambios en la expresión de genes involucrados en los
procesos de proliferación, apoptosis y progresión del ciclo celular.
El desbalance en la razón REβ/REα puede ser un paso crítico en la progresión
a hiperplasia endometrial dependiente de estrógenos en las mujeres SOP, y
vías específicas para ambas isoformas del RE pueden tener distintos roles en
este proceso.
El aumento en la proliferación y la escasa apoptosis, junto con las diferencias
encontradas en la expresión de proteínas relacionadas con estos procesos,
indica que en los endometrios SOP existe una desregulación de la
homeostasis tisular, principalmente ligada a una inadecuada proliferación.
La localización principal de Ciclina E, Cdk-2 y Ki67 en las células epiteliales,
sugiere que el complejo Ciclina E/Cdk-2 está involucrado en la regulación del
crecimiento de este tipo celular en los grupos de endometrios estudiados.
Aunque las voluntarias normales expresan Ciclina E y Cdk-2 en sus
endometrios en fase proliferativa, y se sabe que su expresión disminuye en
fase secretora, los endometrios de pacientes SOP están continuamente
expresando estos reguladores positivos del ciclo celular, dada su
característica de amenorreicas u oligomenorreicas.

Conclusiones
102
Las diferencias encontradas en la localización subcelular de p27 en las células
epiteliales de ESOP, podrían favorecer la desregulación en la homeostasis
endometrial de las mujeres SOP por inhibición de la apoptosis.
La activación de Akt y la sobreexpresión de Ras en ESOP y ESOPH sugiere
que estas proteínas son importantes reguladores de la supervivencia celular
en los endometrios de estas pacientes.
Las alteraciones observadas en la expresión de proteínas que regulan la
supervivencia celular endometrial de las pacientes SOP, pueden estar
favoreciendo el desarrollo de hiperplasia endometrial en estas pacientes.

Proyecciones
103
IX. PROYECCIONES
La presente investigación es el primer trabajo realizado que aborda el estudio de la
función endometrial en mujeres SOP sin y con hiperplasia endometrial. Lo
interesante es que los endometrios de estas pacientes experimentan cambios en la
expresión de algunos reguladores del ciclo celular. Por lo tanto, existe una clara
desregulación de la homeostasis endometrial en estas pacientes, que potencialmente
puede conducir al desarrollo de hiperplasia y/o carcinoma endometrial. Por lo tanto,
este trabajo podría ayudar en el diagnóstico y en los procedimientos terapéuticos de
las pacientes SOP, sugiriendo fuertemente para ello, solicitar biopsia endometrial
principalmente a las mujeres SOP obesas.

Anexos
104
X. ANEXOS
X.1 CONSENTIMIENTO INFORMADO PARA LA OBTENCION DE MUESTRAS
DE TEJIDO ENDOMETRIAL
Título del Proyecto: “Reguladores de la supervivencia celular en endometrios de
pacientes con Síndrome de Ovario Poliquístico. Potenciales consecuencias en el
desarrollo de hiperplasia endometrial”
Nombre del investigador principal: Alejandra Mariela Villavicencio Galdeano
Institución: Instituto de Investigaciones Materno Infantil (IDIMI), Facultad de Medicina,
Universidad de Chile
Teléfonos: 56-2-4248280/ 424828
Le estamos invitando a participar en el proyecto de investigación Reguladores de la
supervivencia celular en endometrios de pacientes con Síndrome de Ovario
Poliquístico. Potenciales consecuencias en el desarrollo de hiperplasia endometrial.
Esta investigación tiene por objetivo estudiar la regulación del ciclo celular en
endometrios normales, y la desregulación de los marcadores de su función en
endometrios de mujeres con el Síndrome de Ovario Poliquístico (SOP), ya que la
descordinación entre proliferación y apoptosis puede favorecer el desarrollo de
hiperplasia endometrial en estas pacientes.
Este estudio incluirá a un número total de 60 pacientes, de los Consultorios de
Infertilidad y de Gineco-Endocrinología del Hospital Clínico San Borja Arriarán,
Campus Centro, Facultad de Medicina, Universidad de Chile.
Si Ud. acepta participar en este proyecto, se le pedirá una muestra de tejido
endometrial obtenida por indicación médica de biopsia endometrial, histerectomía por
patología endometrial y/o esterilización tubaria, parte de la cual será utilizada en este
estudio. Dicho procedimiento no tendrá más riesgo para Ud. que el procedimiento
mismo.

Anexos
105
La participación en este estudio no le significará a Ud. ningún beneficio inmediato. Su
contribución será importante para la identificación de marcadores tempranos de
hiperplasia endometrial en las mujeres que padecen SOP.
Ud. es libre de participar o no en este estudio, de manera que si Ud. decide no
participar, no habrá consecuencias negativas para Ud. y recibirá la misma atención
previamente acordada con su médico tratante.
Toda la información será conservada con estricta confidencialidad, lo que incluye el
acceso de los investigadores o de agencias supervisoras de la investigación.
Además, la publicación de los resultados no incluirá su nombre e identidad.
Si Ud. necesita cualquier otra información sobre este proyecto o acerca de su
participación, puede llamar al Dr. Ariel Fuentes (Fono: 4248280) o dirigirse a la Dra.
Margarita Vega (Fono: 4248283).
Después de haber recibido y comprendido la información de este documento y de
haber podido aclarar todas mis dudas, otorgo mi consentimiento para participar en el
proyecto Reguladores de la supervivencia celular en endometrios de pacientes con
Síndrome de Ovario Poliquístico. Potenciales consecuencias en el desarrollo de
hiperplasia endometrial.
..................................... .......................... ...........................
Nombre de la paciente Firma Fecha
................................... ........................... ...........................
Nombre del médico Firma Fecha
....................................... ............................ ..........................
Nombre del investigador Firma Fecha

Bibliografía
106
XI. BIBLIOGRAFIA
1. Cormack DH. 1990 Endometrio. En Histología de HAM. Harper and Row Eds. 9º
edición. Harla. México. p 777-79.
2. Salamonsen LA and Nancarrow CD. 1994 Cell Biology of the Oviduct and
Endometrium. En: Molecular biology of the female reproductive system. Findlay JK
ed. Academic Press.
3. Noyes RW, Hertig DT and Rock J. 1950 Dating the endometrial biopsy. Fertil Steril
1: 3-25.
4. Ferenczy A and Guralnick. 1983 Endometrial microestructure: Structure-function
relationships throughout the menstrual cycle. Sem Reprod Endocrinol 1: 205-19.
5. Ferenczy A and Bergeron C. 1991 Histology of the human endometrium: From birth
to senescence. Ann N Y Acad Sci 622: 6-27.
6. Joshi SG. 1983 Progestin-regulated proteins of the human endometrium. Sem
Reprod Endocrinol 1: 221-36.
7. Neulen J, Zahradnik HP, Flecken U, et al. 1988 Effects of estradiol-17β and
progesterone on the synthesis of prostaglandin F2α, prostaglandin E2, and
prostaglandin I2 by fibroblast from human endometrium in vitro. Prostaglandins 36:
1511-19.
8. Legro RS, Spielman R, Urbanek M, et al. 1998 Phenotype and genotype in
polycystic ovary syndrome. Rec Prog Horm Res 53: 217-56.

Bibliografía
107
9. The Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group.
Revised 2003 consensus on diagnostic criteria and long-term health risks related to
polycystic ovary syndrome. 2004 Fertil Steril: 81: 19-25.
10. Homburg R. 1996 Polycystic ovary syndrome-from gynaecological curiosity to
multisystem endocrinopathy. Hum Reprod 11: 29-39.
11. Stein IF and Leventhal ML. 1935 Amenorrhea associated with bilateral polycystic
ovaries. Am J Obst Gynecol 29: 181-91.
12. Okon MA, Laird SM, Tuckerman EM, et al. 1998 Serum androgen levels in
women who have recurrent miscarriages and their correlation with markers of
endometrial function. Fertil Steril 69: 682-90.
13. Sagle M, Bishop K, Ridley N, et al. 1988 Recurrent early miscarriage and
polycystic ovaries. Br Med J 297: 1027-28.
14. Chamlain DL and Taylor HB. 1970 Endometrial hyperplasia in young women.
Obstet Gynecol 36: 659-66.
15. Cheung AP. 2001 Ultrasound and menstrual history in predicting endometrial
hyperplasia in polycystic ovary syndrome. Obst Gynecol 98: 325-31.
16. Balen A. 2001 Polycystic ovary syndrome and cancer. Hum Reprod Update 7:
522-25.
17. HendersonBE, Casagrande JT, Pike MC, et al. 1983 The epidemiology of
endometrial cancer in young women. Br J Cancer 47: 749-56.
18. Elwood JM, Cole P; Rothman KJ, et al. 1977 Epidemiology of endometrial cancer.
J Natl Cancer Inst 59: 1055-60.

Bibliografía
108
19. Coulam CB, Annegers JF and Kranz JS. 1983 Chronic anovulation syndrome and
associated neoplasia. Obstet Gynecol 61: 403-7.
20. Lebovic DI, Shifren JL, Ryan IP, et al. 2000 Ovarian steroid and cytokine
modulation of human endometrial angiogenesis. Hum Reprod 3: 67-77.
21. Petra P. 1999 The plasma sex steroid binding protein (SBP or SHBG): A critical
review of recent development on the structure, molecular biology and function. J
Steroid Biochem Mol Biol 64: 100-6.
22. Irwin JC, Utian WH and Eckert RL. 1991 Sex steroids and growth factors
differentially regulate the growth and differentiation of cultured human endometrial
stromal cells. Endocrinology 129: 2385-92.
23. Moller B, Rasmussen C, Lindblom B, et al. 2001 Expression of the angiogenic
growth factors VEGF, FGF-2, EGF and their receptors in normal human endometrium
during the menstrual cycle. Mol Hum Reprod 7: 65-72.
24. Lannigan DA. 2003 Estrogen receptor phosphorilation. Steroids 68: 1-9.
25. Enmark E, Pelto-Huikko M, Grandien K, et al. 1997 Human estrogen receptor
beta-gene structure, chromosomal localization and expression pattern. J Clin
Endocrinol Metab 82: 4258-65.
26. Fujimoto J, Sakaguchi H, Aoki I, et al. 2002 Clinical implications of the expression
of estrogen receptor-α and -β in primary and metastatic lesions of uterine endometrial
cancers. Oncology 62: 269-77.
27. Weihua Z, Saji S, Mäkinen S, et al. 2000 Estrogen receptor (ER) β, a modulator in
the uterus. Proc Natl Acad Sci USA 97: 5936-41.

Bibliografía
109
28. Kastner P, Krust A, Turcotte B, et al. 1990 Two distinct estrogen-regulated
promoters generate transcripts encoding the two functionally different human
progesterone receptor forms A and B. EMBO J 9: 1603-24.
29. Fujimoto J, Ichigo S, Hori M, et al. 1995 Expression of progesterone receptor form
A and B mRNAs in gynecologic malignant tumors. Tumor Biol 16: 254-60.
30. Maliqueo M, Clementi M, Gabler F, et al. 2003 Expression of steroid receptors
and proteins related to apoptosis in endometria of women with polycystic ovary
syndrome. Fertil Steril 80: 812-19.
31. Vermeulen-Meiners C, Poortman J, Nabuurs M, et al. 1988 The endogenous
concentration and subcellular distribution of androgens in normal human
premenopausal endometrium, myometrium and vagina. Gynecol Endocrinol 2: 121-
30.
32. Horie K, Takakura K, Imai K, et al. 1992 Immunohistochemical localization of
androgen receptor in the human endometrium, deciduas, placenta and pathological
conditions of the endometrium. Hum Reprod 7: 1461-66.
33. Suri NK. 1997 Steroid content in endometrial tissue. East Afr Med J 74: 242-48.
34. Kato J and Seto T. 1985 Correlation of androgen receptors with histological
differentiation in human endometrial carcinomas. Acta Obstet Gynecol Scand 64:
209-12.
35. Mertens HJMM, Heineman MJ, Koudstaal J, et al. 1996 Androgen receptor
content in human endometrium. Eur J Obstet Gynecol Reprod Biol 70: 11-13.

Bibliografía
110
36. Fujimoto J, NishigakiM, Hori M, et al. 1994 The effect of estrogen and androgen
on androgen receptors and mRNA levels in uterine leiomyoma, myometrium and
endometrium of human subjects. J Steroid Biochem Mol Biol 50: 137-43.
37. Lovely LP, Apparao KBC, Gui Y, et al. 2000 Characterization of the androgen
receptor in a well-differentiated endometrial adenocarcinoma cell line (Ishikawa). J
Steroid Biochem Mol Biol 74: 235-41.
38. Slayden OD, Nayak NR, Burton KA, et al. 2001 Progesterone antagonists
increase androgen receptor expression in the rhesus macaque and human
endometrium. J Clin Endocrinol Metab 86: 2668-79.
39. Vivat V, Gofflo D, Garcia T, et al. 1997 Sequences in the ligand-binding domains
of the human androgen and progesterone receptors which determine their distinct
ligand identities. J Mol Endocrinol 18: 147-60.
40. Loesser AA. 1938 The action of testosterone propionate on the uterus and breast.
Lancet 12: 373-74.
41. Apparao KBC, Lovely LP, Gui Y, et al. 2002 Elevated endometrial androgen
receptor expression in women with polycystic ovary syndrome. Biol Reprod 66: 297-
304.
42. Shibata H, Spencer TE, Onate SA, et al. 1997 Role of co-activators and co-
repressors in the mechanism of steroid/thyroid receptor action. Recent Prog Horm
Res 52: 141-64.
43. Torchia J, Glass C and Rosenfeld MG. 1998 Co-activators and co-repressors in
the integration of transcriptional responses. Curr Opin Cell Biol 10: 373-78.

Bibliografía
111
44. Parker MG. 1998 Transcriptional activation by oestrogen receptors. Biochem Soc
Symp 63: 45-50.
45. Kershah SM, Desouki MM, Koterba KL, et al. 2004 Expression of estrogen
receptor coregulators in normal and malignant human endometrium. Gynecol
Oncology 92: 304-13.
46. Spencer TE, Jenster G, Burcin MM, et al. 1997 Steroid receptor co-activator-1 is a
histone acetyltransferase. Nature 389: 194-98.
47. Chen H, Lin RJ, Schiltz RL, et al. 1997 Nuclear receptor coactivator ACTR is a
novel histone acetyltransferase and forms a multimeric activation complex with P/CAF
and CBP/p300. Cell 90: 569-80.
48. Gregory CW, Wilson EM, Apparao KBC, et al. 2002 Steroid receptor coactivator
expression throughout the menstrual cycle in normal and abnormal endometrium. J
Clin Endocrinol Metab 87: 2960-66.
49. Speroff L and Vande Wiele RL. 1971 Regulation of the human menstrual cycle.
Am J Obstet Gynecol 109: 234-47.
50. Tabibzadeh S. 1996 The signals and molecular pathways involved in human
menstruation, a unique process of tissue destruction and remodeling. Mol Hum
Reprod 2: 77-92.
51. Allen RT, Cluck MW and Agrawal DK. 1998 Mechanisms controlling cellular
suicide: role of Bcl-2 and caspases. Cell Mol Life Sci 54: 427-45.
52. Bröker LE, Kruyt FAE, and Giaccone G. 2005 Cell death independent of
caspasas: a review. Clin Cancer Res 11: 3155-62.

Bibliografía
112
53. Kroemer G, Petit P, Zamzami N, et al. 1995 The biochemistry of programmed cell
death. FASEB J 9: 1277-87.
54. Hale AJ, Smith CA, Sutherland LC, et al. 1996 Apoptosis: molecular regulation of
cell death. Eur J Biochem 236: 1-26.
55. Hopwood D and Levison DA. 1975 Atrophy and apoptosis in the cyclical human
endometrium. J Pathol 119: 159-66.
56. Verma V. 1983 Ultrastructural changes in human endometrium at different phases
of the menstrual cycle and their functional significance. Gynecol Obstet Invest 15:
193-212.
57. Parr MB and Parr EL. 1989 The implantation reaction. 2 ed. New York: Plenum
Press.
58. von Rango U, Classen-Linke I, Krusche CA, et al. 1998 The receptive
endometrium is characterized by apoptosis in the glands. Hum Reprod 13: 3177-89.
59. Tabidzadeh S, Kong QF, Satyaswaroop PG, et al. 1994 Distinct regional and
menstrual cycle dependent distribution of apoptosis in human endometrium. Potential
regulatory role of T cells and TNF-alfa. Endocr J 2: 87-95.
60. Gompel A, Sabourin JC, Martin A, et al. 1994 Bcl-2 expression in normal
endometrium during the menstrual cycle. Am J Pathol 144: 1195-202.
61. Castro A, Olguin P, Gabler F, et al. 1999 Apoptosis and expression of the proto-
oncogenes bcl-2 and c-myc in normal endometrium trough the human menstrual
cycle. 10th World Congress of Human Reproduction. Salvador, Bahía, Brazil. 4 al 8 de
Mayo, p 83.

Bibliografía
113
62. Tao XJ, Tilly KI, Maravei DV, et al. 1997 Differential expression of members of the
bcl-2 gene family in proliferative and secretory human endometrium: Galndular
epithelial cell apoptosis is associated with increased expression of bax. J Clin
Endocrinol Metab 82: 2738-46.
63. Tao XJ, Sayegh RA, Tilly JL, et al. 1998 Elevated expression of the proapoptotic
Bcl-2 family member, BAK, in the human endometrium coincident with apoptosis
during the secretory phase of the cycle. Fertil Steril 70: 338-43.
64. Speroff L, Glass R, and Kase N. 1994 Hormone biosyntesis, metabolism, and
mechanism of action. In: C. Mitchell (ed.) Clinical gynecologic endocrinology and
infertility. 5 ed. Pp. 31-92. Baltimore: Williams & Wilkins.
65. Kokeguchi S, Hayase R, and Sekiba K. 1992 Proliferative activity in normal
endometrium and endometrial carcinoma measured by immunohistochemistry using
Ki-67 and anti-DNA polymerase alfa antibody, and flow cytometry. Acta Med
Okayama 46: 113-21.
66. Clarke C and Sutherland R. 1990 Progestin regulation of cellular proliferation.
Endocr Rev 11: 266-72.
67. Ferenczy A, Bertrand G, and Gelfand MM. 1979 Proliferation kinetics of human
endometrium during the normal menstrual cycle. Am J Obstet Gynecol 133: 859-67.
68. Odom LD, Barrett JM, Pantazis CG, et al. 1989 Immunocytochemical study of ras
and myc proto-oncogene polypeptide expression in the human menstrual cycle. Am J
Obstet Gynecol 161: 1663-68.

Bibliografía
114
69. Watson H, Franks S and Bonney RC. 1997 Regulation of epidermal growth factor
receptor by androgens in human endometrial cells in culture. Hum Reprod 13: 2585-
91.
70. Tuckermann EM; Okon MA, Li TC, et al. 2000 Do androgen have a direct effect
on endometrial function? An in vitro study. Fertil Steril 74: 771-79.
71. Kurman RJ, Kaminski PF, and Norris HJ. 1985 The behavior of endometrial
hyperplasia. A long-term study of “untreated” hyperplasia in 170 patients. Cancer 56:
403-12.
72. Sherman ME. 2000 Theories of endometrial carcinogenesis: a multidisciplinary
approach. Mod Pathol 13: 295-308.
73. Sherr CJ and Roberts JM. 1995 Inhibitors of mammalian G1 cyclin-dependent
kinases. Genes Develop 9: 1149-63.
74. Nurse P. 1994 Ordering S phase and M phase in the cell cycle. Cell 79: 547-50.
75. Guzeloglu-Kayisli O, Kayisli UA, Al-Rejjal R, et al. 2003 Regulation of PTEN
(Phosphatase and tensin homolog deleted on chromosome 10) expression by
estradiol and progesterone in human endometrium. J Clin Endocrinol Metab 88: 5017-
26.
76. Tashiro H, Blazes MS, Wu R, et al. 1997 Mutations in PTEN are frequent in
endometrial carcinoma but rare in other common gynecological malignancies. Cancer
Res 57: 3935-40.
77. Weinberg RA. 1995 The retinoblastoma protein and cell cycle control. Cell 81:
323-30.

Bibliografía
115
78. Rubul Quddus M, Latkovich P, Castellano WJ, et al. 2002 Expression of cyclin D1
in normal, metaplasic, hyperplastic endometrium and endometrioid carcinoma
suggests a role in endometrial carcinogenesis. Arch Pathol Lab Med 126: 459-63.
79. Sherr CJ. 1996 Cancer cell cycles. Science 274: 1672-77.
80. Kyushima N. 2002 Expression of cyclin A in endometrial adenocarcinoma and its
correlation with proliferative activity and clinicopathological variables. J Cancer Res
Clin Oncol 128: 307-12.
81. Pestell RG, Albanese C, Reutens AT, et al. 1999 The cyclins and cyclin-
dependent kinase inhibitors in hormonal regulation of proliferation and differentiation.
Endocr Rev 20: 501-34.
82. Quarmby VE and Korach KS. 1984 The influence of 17 beta-estradiol on patterns
of cell division in the uterus. Endocrinology 114: 694-702.
83. Imai T, Kurachi H, Adachi K, et al. 1995 Changes in epidermal growth factor
receptor and the levels of its ligands during menstrual cycle in human endometrium.
Biol Reprod 52: 928-38.
84. Corleta H, Capp E, and Strowitzki T. 2000 Cycle modulation of insulin-like growth
factor-binding protein 1 in human endometrium. Braz J Med Biol Res 33: 1387-91.
85. Hata H, Hamano M, Watanabe J, et al. 1998 Role of estrogen and estrogen-
related growth factor in the mechanism of hormone dependency of endometrial
carcinoma cells. Oncology 55 Suppl 1: 35-44.
86. LeBrun DP, Warnke RA, and Cleary ML. 1993 Expression of bcl-2 in fetal tissues
suggests a role in morphogenesis. Am J Pathol 142: 743-53.

Bibliografía
116
87. Chan WK, Mole MM, Levison DA, et al. 1995 Nuclear and cytoplasmic Bcl-2
expression in endometrial hyperplasia and adenocarcinoma. J Pathol 177: 241-6.
88. Hockenbery DM. 1994 Bcl-2 in cancer, development and apoptosis. J Cell Sci 18:
51-5.
89. Otsuki Y, Ito I, Akao Y, et al. 1994 Cyclic Bcl-2 gene expression in human uterine
endometrium during the menstrual cycle. Lancet 344: 27-9.
90. Morsi HM, Leers MP, Radespiel-Troger M, et al. 2000 Apoptosis, bcl-2
expression, and proliferation in benign and malignant endometrial epithelium: An
approach using multiparameter flow cytometry. Gynecol Oncol 77: 11-7.
91. Crescenzi E and Palumbo G. 2001 Bcl-2 exerts a pRb-mediated cell cycle
inhibitory function in HEC1B endometrial carcinoma cells. Gynecol Oncol 81: 184-92.
92. Bozdogan O, Atasoy P, Erekul S, et al. 2002 Apoptosis-related proteins and
steroid hormone receptors in normal, hyperplastic, and neoplastic endometrium. Int J
Gynecol Pathol 21: 375-82.
93. Korsmeyer SJ. 1999 Bcl-2 gene family and the regulation of programmed cell
death. Cancer Res 59: 1693s-1700s.
94. Sakuragi N, Salah-Eldin AE, Watari H, et al. 2002 Bax, Bcl-2 and p53 expression
in endometrial cancer. Gynecol Oncol 86: 288-96.
95. Berchuck A. 1995 Biomarkers in the endometrium. J Cell Biochem Suppl 23: 174-
8.
96. Myers MP, Pass I, Batty IH, et al. 1998 The lipid phosphatase activity of PTEN is
critical for its tumor suppressor function. Proc Natl Acad Sci 95: 13513-18.

Bibliografía
117
97. Weng LP, Brown JL, and Eng C. 2001 PTEN coordinates G1 arrest by down-
regulating cyclin D1 via its protein phosphatase activity and up-regulating p27 via its
lipid phosphatase activity in a breast cancer model. Hum Mol Genetics 10: 599-604.
98. Tashiro H, Blazes MS, Wu R, et al. 1997 Mutations in PTEN are frequent in
endometrial carcinoma but rare in other common gynecological malignancies. Cancer
Res 57: 3935-40.
99. Yoshinaga K, Sasano H, Furukawa T, et al. 1998 The PTEN, BAX and IGFIIR
genes are mutated in endometrial atypical hyperplasia. Jpn J Cancer Res 89: 985-90.
100. Maxwell GL, Risinger JI, Gumbs C, et al. 1998 Mutation of the PTEN tumor
suppressor gene in endometrial hyperplasia. Cancer Res 58: 2500-03.
101. Maliqueo M, Quezada S, Clementi M, et al. 2004 Potential action of
androstenedione on the proliferation and apoptosis of stromal endometrial cells.
Reprod Biol Endocrinol 2: 1-10.
102. Greisen S, Flyvbjerg A, Ledet T, et al. 2002 Regulation of insulin-like growth
factor binding protein secretion by human granulose luteal cells in a polycystic ovary-
like environment. Fertil Steril 78: 162-68.
103. Lessey BA, Killam AP, Metzger DA, et al. 1988 Immunohistochemical analysis of
human uterine estrogen and progesterone receptors throughout the menstrual cycle.
J Clin Endocrinol Metab 67: 334-40.
104. Vaskivuo TE, Stenback F, Karhumaa P, et al. 2000 Apoptosis and apoptosis-
related proteins in human endometrium. Mol Cell Endocrinol 165: 75-83.

Bibliografía
118
105. Shang Y, Hu X, DiRenzo J, et al. 2000 Cofactor dynamics and sufficiency in
estrogen receptor-regulated transcription. Cell 103: 843-52.
106. Wieser F, Schneeberger C, Hudelist G, et al. 2002 Endometrial nuclear receptor
co-factors SRC-1 and N-CoR are increased in human endometrium during
menstruation. Mol Hum Reprod 8: 644-50.
107. Shiozawa T, Shih HC, Miyamoto T, et al. 2003 Cyclic changes in the expression
of steroid receptor coactivators and corepressors in the normal human endometrium.
J Clin Endocrinol Metab 88: 871-8.
108. Weihua Z, Makela S, Andersson LC, et al. 2001 A role for estrogen receptor β in
the regulation of growth of the ventral prostate. PNAS 98: 6330-35.
109. Brandenberger AW, Tee MK, and Jaffe RB. 1998 Estrogen receptor alpha (ER-
alpha) and beta (ER-beta) mRNAs in normal ovary, ovarian serous
cystadenocarcinoma and ovarian cancer cell lines: down-regulation of ER-beta in
neoplastic tissues. J Clin Endocrinol Metab 83: 1025-28.
110. Pujol P, Rey JM, Nirde P, et al. 1998 Differential expression of estrogen
receptor-alpha and -beta messenger RNAs as a potential marker of ovarian
carcinogenesis. Cancer Res 58: 5367-73.
111. Foley EF, Jazaeri AA, Shupnik MA, et al. 2000 Selective loss of estrogen
receptor beta in malignant human colon. Cancer Res 60: 245-8.
112. Rutherford T, Brown WD, Sapi E, et al. 2000 Absence of estrogen receptor-beta
expression in metastatic ovarian cancer. Obstet Gynecol 96: 417-21.

Bibliografía
119
113. Campbell-Thompson M, Lynch J, and Bhardwaj B. 2001 Expression of estrogen
receptor (ER) subtypes ER beta isoforms in colon cancer. Cancer Res 61: 632-40.
114. Roger P, Sahla ME, Makela S, et al. 2001 Decreased expression of receptor
beta protein in proliferative preinvasive mammary tumors. Cancer Res 61: 2537-41.
115. Fixemer T, Remberger K, and Bonkhoff H. 2003 Differential expression of the
estrogen receptor beta in human prostate tissue, premalignant changes, and in
primary, metastatic, and recurrent prostatic adenocarcinoma. Prostate 54: 79-87.
116. Paruthiyil S, Parmar H, Kerekatte V, et al. 2004 Estrogen receptor beta inhibits
human breast cancer cell proliferation and tumor formation by causing a G2 cell cycle
arrest. Cancer Res 64: 423-28.
117. Strom A, Hartman J, Foster JS, et al. 2004 Estrogen receptor β inhibits 17β-
estradiol-stimulated proliferation of the breast cancer cell line T47D. PNAS 101: 1566-
71.
118. Liu MM, Albanese C, Anderson CM, et al. 2002 Opposing action of estrogen
receptors alpha and beta on cyclin D1 gene expression. J Biol Chem 277: 24353-60.
119. Nilsen J, Mor G, and Naftolin F. 2000 Estrogen-regulated developmental
neuronal apoptosis is determined by estrogen receptor subtype and the Fas/Fas
ligand system. J Neurobiol 43: 64-78.
120. Heinlein CA and Chang C. 2002 Androgen receptor (AR) coregulators: an
overview. Endocr Rev 23: 175-200.
121. Klinge CM. 2000 Estrogen receptor interaction with co-activators and co-
repressors. Steroids 65: 227-51.

Bibliografía
120
122. Anzick SL, Kononem J, Walker RL, et al. 1997 AIB1, a steroid receptor
coactivator amplified in breast and ovarian cancer. Science 277: 965-8.
123. Wang Y, Wu MC, Sham JS, et al. 2002 Prognostic significance of c-myc and
AIB1 amplification in hepatocellular carcinoma. A broad survey using high-throughput
tissue microarray. Cancer 95: 2346-52.
124. Sakakura C, Hagiwara A, Yasuoka R, et al. 2000 Amplification and over-
expression of the AIB1 nuclear receptor co-activator gene in primary gastric cancers.
In J Cancer 89: 217-23.
125. Torres-Arzayus MI, Font de Mora J, Yuan J, et al. 2004 High tumor incidence
and activation of the PI3K/AKT pathway in transgenic mice define AIB1 as an
oncogene. Cancer Cell 6: 263-74.
126. Leo C and Don Chen J. 2000 The SRC family of nuclear receptor coactivators.
Gene 245: 1-11.
127. Kollara A, Kahn HJ, Marks A, et al. 2001 Loss of androgen receptor associated
protein 70 (ARA70) expression in a subset of HER2-positive breast cancers. Breast
Cancer Res Treat 67: 245-53.
128. Hermanson O, Glass CK, and Rosenfel M. 2002 Nuclear receptor coregulators:
multiple modes of modification. Trends Endocrinol Metab 13: 55-60.
129. Boutell JM, Thomas P, Neal JW, et al. 1999 Aberrant interactions of
transcriptional repressor proteins with the Huntington’s disease gene product,
huntingtin. Hum Mol Genet 8: 1647-55.

Bibliografía
121
130. Simpson ER, Meandro MS, Means GD, et al. 1994 Aromatase cytochrome
P450, the isoenzyme responsible for estrogen metabolism. Endocr Rev 15:342-55.
131. Sasano H and Harada N. 1998 Intratumoral aromatase in human breast,
endometrial and ovarian malignancies. Endocr Rev 19:593-607.
132. Watanabe K, Sasano H, Harada N, et al. 1995 Aromatase in human endometrial
carcinoma and hyperplasia. Immunohistochemical, in situ hybridization, and
biochemical studies. Am J Pathol 146: 491-500.
133. Bulun SE, Economos K, Miller D, et al. 1994 CYP19 (aromatase cytochrome
P450) gene expression in human malignant endometrial tumors. J Clin Endocrinol
Metab 79: 1831-1834.
134. Sasano H, Kaga K, Sato S, et al. 1996 Aromatase cytochrome P450 gene
expresión in endometrial carcinoma. Br J Cancer 74:1541-4.
135. Dahmoun M, Boman K, Cajander S, et al. 1999 Apoptosis, proliferation, and sex
hormone receptors in superficial parts of human endometrium at the end of the
secretory phase. J Clin Endocrinol Metab 84: 1737-43.
136. Critchley HOD, Tong S, Cameron ST, et al. 1999 Regulation of bcl-2 gene family
members in human endometrium by antiprogestin administration in vivo. J Reprod
Fertil 115: 389-95.
137. Strauss III and Coutifaris C. 1999 The Endometrium and Myometrium.
Regulation and Dysfunction: Reproductive Endocrinology, 4th edn Yen SSC, Jaffe RB
and Barbieri R (eds). WB Saunders Co.: Philadelphia, pp. 218-56.

Bibliografía
122
138. Gronroos M, Maenpaa J, Kangas L, et al. 1987 Steroid receptors and response
of endometrial cancer to hormones in vitro. Ann Chir Gynecol 202: 76-9.
139. Tseng L and Mazzella J. 1980 Cyclic Changes of Estradiol Metabolic Enzymes
in Human Endometrium During the Menstrual Cycle: The Endometrium Kimball FA
(ed). SP Medial and Scientific Books: New York, pp. 211-26.
140. Giudice LC. 1994 Growth factors and growth modulators in human uterine
endometrium: their potential relevance to reproductive medicine. Fertil Steril 61: 1-17.
141. Shiozawa T, Miyamoto T, Kashima H, et al. 2004 Estrogen-induced proliferation
of normal endometrial glandular cells is initiated by transcriptional activation of cyclin
D1 via binding of c-Jun to an AP-1 sequence. Oncogene 23: 8603-10.
142. Moreno-Bueno G, Rodríguez-Perales S, Sánchez-Estévez C. et al. 2004
Molecular alterations associated with cyclin D1 overexpression in endometrial cancer.
Int J Cancer 110: 194-200.
143. Milde-Langosch K, Bamberger AM, Goemann C, et al. 2001 Expression of cell-
cycle regulatory proteins in endometrial carcinomas: correlations with hormone
receptor status and clinicopathologic parameters. J Cancer Res Clin Oncol 127: 537-
44.
144. Corakci A and Kacar Özkara S. 2004 Significantly decreased p27 expression in
endometrial carcinoma compared to complex hyperplasia with atypia (correlation with
p53 expression). Pathol Oncol Res10: 89-97.
145. Nagamani M and Stuart CA. 1998 Specific binding and growth-promoting activity
of insulin in endometrial cancer cell lines in culture. Am J Obstet Gynecol 179: 6-12.

Bibliografía
123
146. Kurman RJ, Zaino RJ, and Norris HJ. 1995 Endometrial carcinoma. In: Kurman
RJ (ed) Blaustein’s pathology of the female genital tract. Springer, Berlin Heidelberg
New York, pp. 439-86.
147. Sherr JC, 1994 G1 phase progression: cycling on cue. Cell 79: 551-55
148. Shiozawa T, Li SF, Nakayama K. et al. 1996 Relationship between the
expression of cyclins/cyclin-dependent kinases and sex-steroid receptors/Ki67 in
normal human endometrial glands and stroma during the menstrual cycle. Mol Hum
Reprod 2: 745-52.
149. Shiozawa T, Nikaido T, Nakayama K, et al. 1998 Involvement of cyclin-
dependent kinase inhibitor p27Kip1 in growth inhibition of endometrium in the secretory
phase and of hyperplastic endometrium treated with progesterone. Mol Hum Reprod
4: 899-905.
150. Oshita T, Shigemasa K, Nagai N, et al. 2002 p27, cyclin E, and CDK-2
expression in normal and cancerous endometrium. Int J Oncol 21: 737-43.
151. Besson A, Assoian R, and Roberts JM. 2004 Regulation of the cytoskeleton: an
oncogenic function for CDK inhibitors? Nature Rev Cancer 4: 948-54.
152. Polyak K, Lee MH, Erdjument-Bromage H, et al. 1994 Cloning of p27Kip1, a
cyclin-dependent kinase inhibitor and a potential mediator of extracellular
antimitogenic signals. Cell 78: 59-66.
153. Amso NN, Crow J, and Shaw RW. 1994 Comparative immunohistochemical
study of oestrogen and progesterone receptors in the Fallopian tube and uterus at
different stages of the menstrual cycle and the menopause. Hum Reprod 9: 1027-37.

Bibliografía
124
154. Pickarts H, Beckmann R, Fleige B, et al. 1990 Steroid receptors and proliferative
activity in non-neoplastic and neoplastic endometria. Virchows Archiv A (Pathol Anat)
417: 163-71.
155. Li SF, Nakayama K, Masuzawa H, et al. 1993 The number of proliferating cell
nuclear antigen positive cells in endometriotic lesions differs from that in the
endometrium. Virchows Archiv A (Pathol Anat) 423: 257-63.
156. Yagimuna Y, Katayama H, Kawai K, et al. 1996 Analysis of the retinoblastoma
gene in human endometrial carcinoma. Obstet Gynecol 87: 755-959.
157. Gu J, Tamura M and Yamada KM. 1998 Tumor suppressor PTEN inhibits
integrin- and growth factor-mediated mitogen-activated protein (MAP) kinase
signaling pathways. J Cell Biol 143: 1375-83.
158. Tamura M, Gu J, Matsumoto K, et al. 1998 Inhibition of cell migration, spreading,
and focal adhesions by tumor suppressor PTEN. Science 280: 1614-17.
159. Maehama T and Dixon JE. 1998 The tumor suppressor, PTEN/MMAC1,
dephosphorylates the lipid second messenger, phosphatidylinositol 3,4,5-
trisphosphate. J Biol Chem 273: 13375-8.
160. Li D-M and Sun H. 1998 PTEN/MMAC1/TEP1 suppresses the tumorigenicity and
induces G1 cell cycle arrest in human glioblastoma cells. Proc Natl Acad Sci USA 95:
15406-11.
161. Wu X, Senechal K, Neshat MS, et al. 1998 The PTEN/MMAC1 tumor suppressor
phosphatase functions as a negative regulator of the phosphoinositide 3-kinase/Akt
pathway. Proc Natl Acad Sci USA 95: 15587-91.

Bibliografía
125
162. Paramio JM, Navarro M, Segrelles C, et al. 1999 PTEN tumour supresor is
linked to the cell cycle control through the retinoblastoma protein. Oncogene 18:
7462-68.
163. Klippel A, Escobedo MA, Wachowicz MS, et al. 1998 Activation of
phosphatidylinositol 3-kinase is sufficient for cell cycle entry and promotes cellular
changes characteristic of oncogenic transformation. Mol Cell Biol 18, 5699-711.
164. Vanhaesebroeck B and Alessi DR. 2000 The PI3K-PDK1 connection: more than
just a road to PKB. Biochem J 346: 561-76.
165. Scheid MP and Woodgett JR. 2001 PKB/AKT: functional insights from genetic
models. Nat Rev Mol Cell Biol 2: 760-8.
166. Nicholson KM and Anderson NG. 2002 The protein kinase B/Akt signaling
pathway in human malignancy. Cell Signal 14: 381-95.
167. Liang J, et al. 2002 PKB/Akt phosphorylates p27, impairs nuclear import of p27
and opposes p27-mediated G1 arrest. Nat Med 8: 1153-60.
168. Shin I, et al. 2002 PKB/Akt mediates cell-cycle progression by phosphorylation of
p27Kip1at threonine 157 and modulation of its cellular localization. Nat Med 8: 1145-
52.
169. Kayisli OG, Kayisli UA, Luleci G, et al. 2004 In vivo and in vitro regulation of Akt
activation in human endometrial cells is estrogen dependent. Biol Reprod 71: 714-21.
170. Brazil DP, Yang Z-Z, and Hemmings BA. 2004 Advances in protein kinase B
signaling: AKTion on multiple fronts. Trends Biochem Sci 29: 233-42.

Bibliografía
126
171. Pruitt K and Der CJ. 2001 Ras and Rho regulation of the cell cycle and
oncogenesis. Cancer Lett 171: 1-10.
172. Rodríguez-Viciana P, Warne PH, Vanhasebroeck B, et al. 1996 Activation of
phosphoinositide 3-kinase by interaction with Ras and by point mutation. EMBO J 15:
2442-51.
173. Downward J. 2003 Targeting Ras signalling pathways in cancer therapy. Nat.
Rev. Cancer 3: 11-22.