plant cyclins: a unified nomenclature for plant a-, b- and d-type cyclins based on sequence...
TRANSCRIPT

Plant Molecular Biology 32: 1003-1018, 1996. 1003 (~) 1996 Kluwer Academic Publishers. Printed in Belgium.
Invited Review
Plant cyclins: a unified nomenclature for plant A-, B- and D-type cyclins based on sequence organization
Jean-Pierre R e n a u d i n 1,*, John H. D o o n a n 2, D o n n a F r e e ma n 3, Junj i H a s h i m o t o 4, Her iber t Hir t 5,
Dirk Inz66, T h o m a s Jacobs 7, Hi roshi Kouch i 4, Pierre Rouz66, Margre t Sauter 8,
Ar nou l d Savour69, David A. Sorrell 3, Venkatesan Sundaresan 1° and James A.H. Mu r r a y 3,* 1Laboratory of Plant Biochemistry and Physiology, INRA/ENSAM/CNRS, 34060 Montpellier Cedex 1, France (*author for correspondence); 2Dept. of Cell Biology, John Innes Centre, Colney Lane, Norwich NR4 7UH, UK; 31nstitute of Biotechnology, University of Cambridge, Tennis Court Road, Cambridge CB2 1QT, UK (*author for correspondence); 4National Institute of AgrobiologicaI Resources, Tsukuba, Ibaraki, 305 Japan; 5Institute of Microbiology and Genetics, University of Vienna, Dr Borhgasse 9, 1030 Vienna, Austria; 6 Laboratoire Associ~ de I'INRA, University of Gent, K.L. Ledeganckstraat 35, 9000 Gent, Belgium; 7 University of lllinois, Urbana, IL 61801, USA; 8 Universitat Hamburg, Institut fiir Allgemeine Botanik, 22609 Hamburg, Germany; 9Molecular Biology Laboratory, Faculty of Sciences, 33 rue de St-Leu, 80039Amiens Cedex, France; l°Cold Spring Harbor Laboratory, Cold Spring Harbor, NY 11724, USA
Received 9 September 1996; accepted 16 September 1996
Key words: cyclins, nomenclature
Abstract
The comparative analysis of a large number of plant cyclins of the A/B family has recently revealed that plants possess two distinct B-type groups and three distinct A-type groups of cyclins [1]. Despite earlier uncertainties, this large-scale comparative analysis has allowed an unequivocal definition of plant cyclins into either A or B classes. We present here the most important results obtained in this study, and extend them to the case of plant D-type cyclins, in which three groups are identified. For each of the plant cyclin groups, consensus sequences have been established and a new, rational, plant-wide naming system is proposed in accordance with the guidelines of the Commission on Plant Gene Nomenclature. This nomenclature is based on the animal system indicating cyclin classes by an upper-case roman letter, and distinct groups within these classes by an arabic numeral suffix. The naming of plant cyclin classes is chosen to indicate homology to their closest animal class. The revised nomenclature of all described plant cyclins is presented, with their classification into groups CycA1, CycA2, CycA3, CycB 1, CycB2, CycD 1, CycD2 and CycD3.
Introduction
The cell division cycle is controlled in eukaryotes by cyclin-dependent kinases (CDKs) [2-4]. The kinase activity of CDKs is dependent on the binding of pos- itive regulators known as cyclins, first identified as proteins showing a cyclical pattern of accumulation and destruction during the synchronous divisions that characterise early embryonic development in marine invertebrates [5]. The two cyclins originally described
had different accumulation patterns and were termed A and B [5]. They were later shown to be the regulat- ory subunits of the CDK activity controlling the onset of mitosis [6]. Subsequent work has revealed that in mammals both the cyclin and CDK families display complex organization, with at least ten classes (A to J) of cyclin described to date [2, 7]. These associate with one or more members of the family of CDKs, of which there are at least eight types (CDKI-8). The presence of multiple cyclins and CDKs enables the

1004
fine regulation of cell cycle controls and checkpoints, since different kinase activities are involved at different points within the cell cycle. Cyclins of various classes associated with their cognate CDKs therefore regulate not only the G2-M transition (cyclin A and B, associ- ated with CDK1 and CDK2), but also GI exit (cyclins D and E with CDK2, CDK4 and CDK6) and S-phase progression (cyclin A with CDK2) [2, 8]. A similar theme on a simpler scale is seen in fungi, for example in budding yeast, where cell cycle control in S phase and mitosis is exercised by a number of B-type cyclins [9-11] and in G1 by the Cln cyclins [12, 13].
The archetypal cyclins A and B of animals possess a conserved 250-amino acid domain called the cyclin core [1, 14, 15]. The cyclin core of cyclin A is suffi- cient for binding and activation of CDK1 and CDK2 [16-18]. It displays a rigid tertiary structure organized in two folds of five helices (see Fig. 4). The first fold, composed of 100 amino acids, encompasses the CDK- binding site [19]. This domain is the defining feature of all cyclins, and is called the cyclin box. It is conserved to varying degrees in all cyclin classes [20], although the cyclin superfamily is rather diverse and the homo- logy between cyclins that regulate Gl and G2-M can be fairly low even within the cyclin box.
Cell cycle studies have been triggered in high- er plants by the importance of the proper control of cell division during plant development with respect to meristem activity, secondary organ formation, growth factor response, nodulation and in vitro regeneration [21, 22]. The conservation of the basic tools of the cell cycle machinery amongst eukaryotes has enabled the cloning of plant homologues to CDKs and cyclins by homology [23-25]. The majority of plant cyclins described to date display homology with cyclin classes A and B [26-28]. Recently, genetic complementation of yeast mutants enabled the cloning of a different class of plant cyclins, which show greater homology to animal cyclin D [29, 30].
Despite having homology with cyclin classes of other eukaryotes, all these plant cyclins also show specific structural characteristics that have prevented the number of structural groups of plant cyclins, their consensus sequences and their phylogenetic relation- ship with other eukaryotic cyclins from being assessed. This has led to an ad hoc nomenclature, which is often different for each species, and makes plant-wide ana- lysis or comparisons with animal cyclins very difficult. To avoid confusion, it is therefore essential to estab- lish a plant-wide nomenclature that will indicate the relationships between cyclins in different species.
The purpose of this review is to present an overview of homologies between the plant cyclins described to date and their relationship to established classes of animal and fungal cyclins. The definition of cyc- lin classes and groups which are conserved across plant species allows a plant-wide nomenclature to be developed. We believe that functional studies of plant cyclins will be greatly helped by the establishment of this new system of classification and naming.
Nomenclature of animal cyclins
The nomenclature of animal cyclins has developed from the initial naming of cyclin A and B [5], with different classes given sequential letters. In animals, several cyclin classes are further divided into com- ponent groups which are indicated by arabic numerals. Group membership are defined by sequence homology. Thus human cyclin D1 is closer to mouse cyclin D1 than to human cyclin D2. This inter-species conser- vation is indicative of distinct and conserved functions for the groups. The classification of animal cyclins was originally based solely on amino acid sequence simil- arity, but subsequent functional analysis has shown that cyclins within classes created on.sequence similarity grounds generally also carry out equivalent functions in different organisms.
Plant cyclins related to A-, B- and D-type animal cyclins are classified into eight clusters
In the past 5 years, 61 full-length or nearly full-length cyclin cDNAs have been isolated in plants (Table 1). Of these, 51 are related to A- and B-type cyclins and were found mostly by sequence amplification using the conservation of motifs in helices H 1 and H3 of the first cyclin fold of cyclins A and B (see Fig. 4). Ten D-type cyclins have been found either by complementation of G1-S cyclin-deficient yeast cells or by hybridization with probes derived from the clones obtained in this way. A detailed phylogenetic analysis of A- and B- type plant cyclins [ 1 ], to which we here add the analysis of plant D-type cyclins, shows that cyclin classes are conserved between animals and plants, and that each of the plant cyclins can be defined to the cyclin A, cyclin B or cyclin D classes (Fig. 1). We propose to name these major plant cyclin classes with the mnenomics CycA, CycB and CycD respectively, indicating their sequence relationship to the equivalent cyclin class (A,

1005
B and D) of other eukaryotes. The use of the three- letter Cyc prefix is consistent with the guidelines of the Commission on Plant Gene Nomenclature (CPGN) [31,321.
The analysis defines a total of eight groups within the major cyclin classes (Figs. 1-3). A numbering sys- tem is proposed to distinguish these as distinct gene families [1]. CycA plant cyclins form three groups, CycA1, CycA2 and CycA3 (Fig. 2). CycB plant cyc- lins form two groups, CycB1 and CycB2 (Fig. 3). The number of plant CycD cyclins is not yet sufficient to assess precisely the complexity of this class. Neverthe- less, the comparison often plant CycD cyclins suggests the occurrence of three groups amongst these cyclins, CycD 1, CycD2 and CycD3 (Fig. 1 ). This classification is robust in that it is obtained with various phylogenet- ic analysis methods with high confidence indices [1]. Cyclins from different species or families that lie with- in the same cyclin group are more closely related to each other than to cyclins from the same species in different groups.
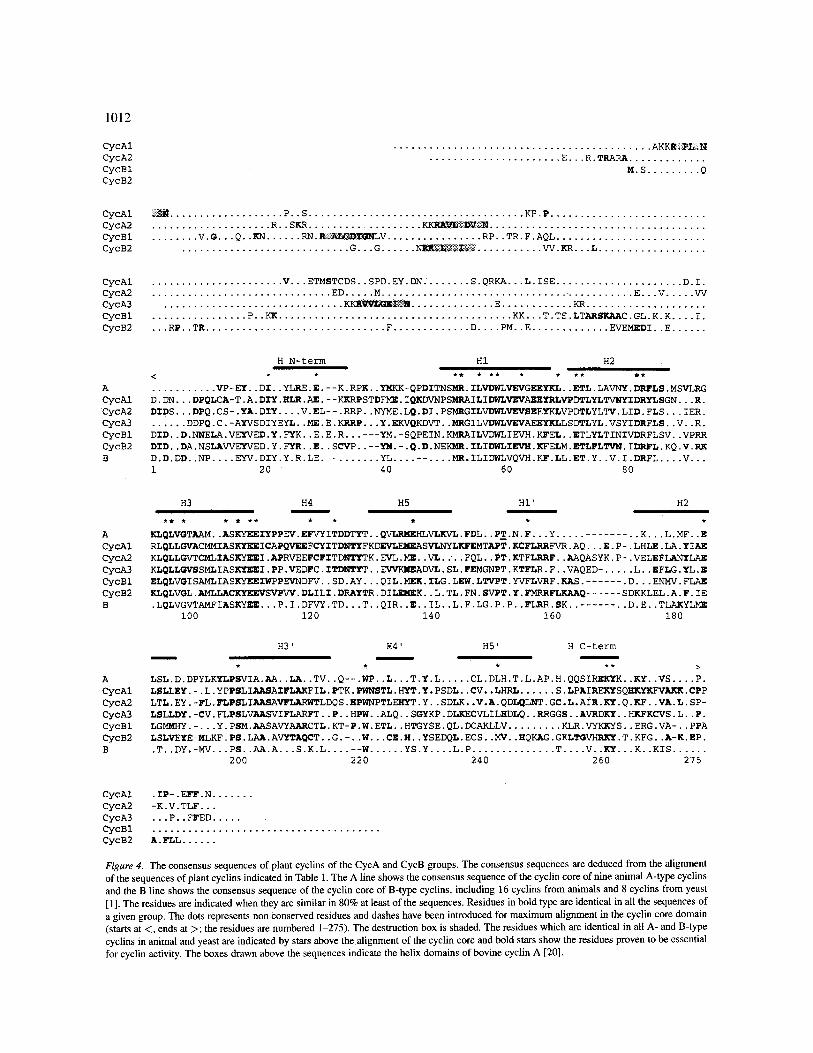
Figure 4 shows the consensus sequences of the 5 groups of plant CycA and CycB cyclins. The cyclin core of plant cyclins CycA and CycB is 75% to 84% conserved within each group, whereas this rate comes to 44% to 53% between the different CycA or CycB groups, and to 31 to 35% between CycA and CycB classes (Table 2).
It is apparent from Fig. 1 that plant cyclin groups arise within each plant cyclin class, and therefore can- not be related to the animal cyclin groups. Plant CycB 1 is therefore related to all animal B-type cyclins, but the nomenclature does not imply a greater similarity or similar function to mammalian cyclin B 1.
The plant cyclins represented in the analysis come from 14 species in seven Angiosperm families (Com- positae, Cruciferae, Gramineae, Leguminosae, Scro- phulariaceae, Solanaceae, Umbelliferae) and include examples from the monocots and dicots. A number of species have examples in many of the separate groups (Table 1). The occurrence of cyclins of classes CycA, CycB and CycD is therefore a general situation in higher plants, and it appears likely that the 8 groups described above are present in all higher plant spe- cies. CycB1 and CycB2 cyclins have been found in Arabidopsis, maize and tobacco (Table 1; Renaudin, unpublished), whereas members of the three CycA groups have been found in tobacco and in soybean (Table 1). The conservation of these groups suggests that their evolutionary origin arose before the appear-
ance of Angiosperms, and that they have distinct func- tions [27].
Sequence analysis of CycA and CycB plant cyclins
Members of the CycA and CycB classes of plant cyc- lins display the same typical structural organization as cyclins A and B from other eukaryotes (Fig. 4). They possess a conserved cyclin core with greatest simil- arity (26 to 37%, Table 2) to cyclin A and cyclin B. Within this domain, they share more than 90% of the 30 most strongly conserved residues of cyclin A and B, and notably all have the 5 residues which have been shown to be essential for cyclin activity by mutational analysis [16-18] (Fig. 4).
The phylogenetic relationship of CycA cyclins with animal cyclin A and of CycB cyclins with yeast and animal cyclin B (Fig. 1) [1] is confirmed only partly at the level of consensus sequence similarity. The cyclin core of CycA or of CycB cyclins is hardly, as a mean, significantly more similar to A-type or to B-type cyc- lins (Table 2). Several studies have emphasized that none of the plant cyclins has the complete consensus motifs previously defined as being typical of the A- and B-type cyclins from other eukaryotes. This is due to the fact that these consensus motifs were established using only metazoans for A-type cyclins or fungi and meta- zoans for B-type cyclins. Plants however represent a distinct kingdom, which may be less related to either metazoans or fungi than these groups are to each other [1, 33]. A detailed analysis retained only one zone in helix H1 of the cyclin box to separate unambiguously every eukaryotic A-type and B-type cyclin (Table 1) [ 1 ]. All A-type cyclins, including those of plants, have the typical LVEVxEEY (x = any amino acid) signa- ture, whereas all B-type cyclins have, instead, the typ- ical cyclin B signature (H/Q)x(K/R/Q)(F/L). It must also be noted that the typical signature of yeast and animal B-type cyclins, FLRRxSK, in helix HI' of the cyclin core, is found in all CycA and CycB cyclins in an altered form, lacking notably the serine residue. A second signature of yeast and animal B-type cyclins, AKYL in helix HU, is absent from plant CycA and CycB cyclins.
Plant CycA and CycB cyclins have a variable, approximately 180-amino-acid N-terminal domain, at the beginning of which is found a 9-amino-acid motif strongly resembling the destruction box of mitotic cyc- lins as defined in animals and fungi. The first residue of the destruction box, arginine, which has been shown

1006
Table 1. A list of plant cyclins. The sequences have been named according to the classification into eight structural groups of CycA, CycB and CycD plant cyclins proposed in [1] and here. Accession numbers in EMBL/GenBank are indicated.
New gene Old name(s) Species Accession Note Reference
name and MN
CycA1 group
Brana;CycA 1; 1
Glyma;CycA 1; 1
Nicta;CycA 1; 1
Nicta; CycA 1 ;2
Orysa;CycA 1; 1 *
Orysa;CycA1;2*
Zeama;CycA I; 1
Zeama;CycA 1 ;2
bncyc2 Brassica napus L25406
cyc3Gm Glycine max D50870
ntcyc25 Nicotiana tabacum D50735
ntCycA19 X92966
ntCycA30 Nicotiana tabacum X92967
cycOs 1 Oryza sativa -
cycOs3 Oryza sativa -
cycIIzm Zea mays U 10077
cycZm2w Zea mays U50064
[56]
[27]
[48]
[61]
[61] Hashimoto, unpublished.
Hashimoto, unpublished
[26] Hsieh and Wolniak, unpublished
CycA2 group
Arath;CycA2;1
Arath;CycA2;2
Arath;CycA2;3
Arath;CycA2;4
Brana;CycA2;1
Glyma;CycA2; 1
Medsa; CycA2; 1
Nicta;CycA2; 1
Pissa;CycA2;1 *
Pissa;CycA2;2*
cyc3aAt Arabidopsis thaliana Z31589 cyc3bAt Arabidopsis thaliana Z31402 3
cyc3c-at Arabidopsis thaliana U17890
atu17889 Arabidopsis thaliana U17889 4
bncyc 1 Brassica napus L25405
cyc2Gm Glycine max D50869 cycMs3 Medicago sativa X85783 5
ntcyc27 Nicotiana tabacum D50736
cycps 1 Pisum sativum -
cycps2 Pisum sativum -
[28]
[28]
Lu and Fed, unpublished
Lu and Fed, unpublished
[56]
[271 [49]
[481 Jacobs, unpublished Jacobs, unpublished
CycA3 group
Antma;CycA3; 1 *
Dauca;CycA3;1
Glyma;CycA3;l
Nicta; CycA3 ; 1
Nicta;CycA3;2
Nicta;CycA3;3
- Antirrhinum majus -
C 13-1 Daucus carota $49312
cyc 1Gin Glycine max D50868
ntCycA105 Nicotiana tabacum X92964
ntCycA59 Nicotiana tabacum X92965
ntCycA13 Nicotiana tabacum X93467
Gaudin et aL, unpublished
[41]
[27]
[61]
[61]
[61]
CycB1 group
Antma; CycB l ; 1
Antma;CycB 1 ;2 Arath;CycB1;1
Arath;CycB 1 ;2
Arath;CycB 1 ;3
Glyma;CycB1; 1
Glyma;CycB1;2
Glyma;CycB1;3
Luplu;CycB I; 1 *
Luplu;CycBl;2*
Luplu;CycB1;3*
Luplu;CycBl ;4* Nicta;CycB1; 1
Nicta;CycBl;2
amcyll Antirrhinum majus X76122 amcyl2 Antirrhinum majus X76123
cyclat Arabidopsis thaliana X62279 cyc lbAt Arabidopsis thaliana L27223
cycx3-ara Arabidopsis thaliana L27224
S 13-6;cyc5Gm Glycine max X62820
gmcyclin Glycine max Z26331
S 13-7 Glycine max X62303
cyc4Gm Glycine max D50871
CycB I a-ll Lupinus luteus -
CycB lb-ll Lupinus luteus -
CycB lc-ll Lupinus luteus -
CycB 1 d-ll Lupinus luteus -
Ntcyc I Nicotiana tabacum Z37978 Ntcyc29 Nicotiana tabacum D50737
[51] [51] [54]
[57]
[57]
[271 [41] Deckert and Gresshoff,
unpublished [41]
[27]
[58] Deckert et al., unpublished Deckert et al., unpublished Deckert et al., unpublished
[59] [481

Table 1. Continued.
New gene Old name(s) Species Accession Note Reference
name and MN
1007
Petcr;CycBl;l pummicy Petroselinum crispum L34207
Pissa;CycB1; 1 * cycps3 Pisum sativum -
Sesro CvcB I 1 * - Sesbania rostrata -
Zeama;CycB1;l cyclazm Zea mays U10079
Zeama;CycBl;2 cyclbzm Zea mays U10078
[60]
Jacobs, unpublished
Goormachtig et al.,
unpublished
[26]
[26]
CyeB2 group Arath;CycB2;l
Arath;CycB2;2
Medsa;CycB2; 1
Medsa;CycB2;2
Medsa;CycB2;3
Orysa; CycB2; 1
Orysa;CycB2;2
Zeama;CycB2; 1
cyc2aAt Arabidopsis thaliana Z31400 [28]
cyc2bAt Arabidopsis thaliana Z31401 II [28]
cycMs 1 Medicago sativa X82039 [49]
cycMsl Medicago sativa X68740 12 [50]
cycMs2 Medicago sativa X82040 [49]
cycMs2 Medicago sativa X68741 13 [50]
cyclIIMs Medicago sativa X78504 14 [53]
cycOs 1 Oryza sativa X82035 [52];
cycOs2 Oryza sativa X82036 15 [52];
cyclIIzm Zea mays U10076 [26]
Hashimoto, unpublished
Sauter, unpublished
CycD1 group
Antma;CycD1;1 * - Antirrhinum majus - Gaudin et aL, unpublished
Arath;CycD1;l cyclin ~1 Arabidopsis thaliana X83369 [29]
Heltu;CycD1; 1 * - Helianthus tuberosus - Freeman and Murray,
unpublished
CycD2 group Arath;CycD2;l cyclin o~2 Arabidopsis thaliana X83370 [29]
CycD3 group Antma;CycD3; 1. - Antirrhinum majus - Gaudin et al., unpublished
Antma;CycD3;2* - Antirrhinum majus - Gaudin et al., unpublished
Arath;CycD3;l cyclin ~3 Arabidopsis thaliana X83371 [29]
Heltu;CycD3;l * - Helianthus tuberosus - Freeman and Murray,
unpublished
Medsa;CycD3;1 cycms4 Medicago sativa X88864 [30]
Nicta;CycD3; 1 * - Nicotiana tabacum - Sorrell and Murray,
unpublished
* Provisional member number allocation, pending ratification by CPGN. Member numbers can only be formally allocated by the Commission for Plant Gene Nomenclature for sequences with an accession number. I Clone ntcyc25 (D50735) was isolated from plant tissue of Nicotiana tabacum cv. Samsun, and clone ntCycA 19 from BY-2 suspension culture cells originally derived from N. tabacum cv. Bright Yellow-2. The nucleotide sequences of the coding regions are 99.8% identical. Despite differences in 5 r- and 3r-untranslated regions, these are assumed here to represent the same gene defined as Nicta; CycA 1;1. 2 95% identity with Nicta; CycA1;l (X92966). 3 71.5% amino acid homology with Arath;CycA2;l in the complete protein. 4 99% identical to Arath;CycA2;2 except in C-terminus. 5 Complements an c~-pheromone-response yeast mutant. 6 81% amino acid identity with Nicta;CycA3;2 in the cyclin box (62% in the N-terminal region). 7 84% amino acid identity with Antma;CycB1;l in the C-terminal two thirds of the protein. 8 Incomplete cDNA, 100% identical to X62820, and assumed to be from same gene. 9 95% amino acid and nucleotide identity with Glyma;CycB1;1. 10 67% amino acid identity with Zeama;CycBl; 1 in the cyclin core. r171.4% amino acid homology with Arath;CycB2;1 in the complete protein. 12 Incomplete cDNA 100% identical to X82039 and assumed to be from same gene. 13 Incomplete cDNA 100% identical to X82040 and assumed to be from same gene. 14 97% identical to Medsa;CycB2;1. 15 77% nucleotide identity with Orysa;CycB2;1.

1008
Table 2. Homology of plant CycA and CycB cyclins. The table shows the homology (%) in the cyclin core (ca. 265 amino acids) amongst the cyclins of one given group or between the five groups of plant cyclins, animal A-type cyclins and yeast and animal B-type cyclins. The % homology within a group is calculated by counting the number of positions at which there is a consensus shown in Fig. 4. The % homology between groups is calculated by counting the positions at which the consensus for the two groups are identical.
CycA 1 CycA2 CycA3 CycB 1 CycB2
CycA1 84 CycA2 53 78 CycA3 48 50 76 CycB 1 34 35 32 CycB2 34 35 31 Cyclin A 37 36 37 Cyclin B 26 27 26
75 44 77 33 31 30 29
to be essential [34, 35], is systematically conserved. The leucine residue in position 4 in the motif is also strongly conserved. It is noteworthy that each group of CycA and CycB cyclins exhibits a different con- sensus sequence for its destruction box (Fig. 4) [1] as is also the case in A- and B-type animal and yeast cyclins. These structural differences probably reflect functional distinctions in the mode or timing of cyclin degradation.
Conserved motifs have been identified in the vari- able N-terminus of CycA and CycB plant cyclins, out- side the destruction box (Fig. 4) [ 1 ]. The most spec- tacular is in CycB 1 cyclins, which have a 12 amino acid long motif, located just upstream the cyclin core, and containing a continuous stretch of 6 conserved residues, TS(V/T)LTARSKAAC. Other motifs are found at the same or other positions in CycB2, CycA1 and CycA2 cyclins. None has homologues in molecu- lar databases. The function of these motifs remains unknown, but it is possible that they bind proteins involved in the compartmentation of the cyclin or the regulation of its activity. Conserved motifs upstream the cyclin core of animal cyclins B 1 and B2 act as cyto- plasmic retention signals responsible for the localiza- tion of these cyclins in the cytoplasm during interphase [36]. The CycA3 plant cyclins, which have almost no conserved motifs in their N-terminal domain, except for the destruction box, are characterized by a much reduced length of this domain: the sequence between their destruction box and the cyclin core is about 43 amino acid long, i.e. 2.5 to 4 times shorter than in other groups of cyclin A or B [ 1 ].
A special mention must be made of putative bipart- ite nuclear localization signals (NLS) which have been detected in nearly all CycB 1 sequences, in the N- terminal region upstream the LTARSKAAC motif, and in the three groups of CycA plant cyclins, in the region surrounding the destruction box [1 ]. Although they have not yet been proven to be functional, the presence of these motifs suggests that, at least during a part of the cell cycle, these cyclins are located in the nucleus. These motifs are absent from CycB2 plant cyclins. In contrast, A- and B-type cyclins from animals and fungi do not contain any NLS, although animal cyclin A is located in the nucleus, and animal cyclin B 1 relocates into the nucleus at the onset of mitosis [37-39].
The occurrence of modified consensus sequences in the cyclin core and the presence of putative NLS in plant cyclins further substantiate the separate evol- ution of plant cyclins in connection with the specific regulation of the plant cell cycle [21-25, 40]. Nev- ertheless, the conserved features that can be defined between plant and fungal/metazoan cyclin A and cyclin B classes respectively, strongly suggest that the distinc- tion between cyclin A and cyclin B was drawn before the division of the multicellular eukaryotes into separ- ate kingdoms. In support of this claim may be cited the limited degree of functional data on expression tim- ing of plant CycA and CycB cyclins, which indicates that CycA genes, like animal cyclin A, appear to be expressed during S phase, whereas CycB are activated later in G2 (see below). Moreover, plant CycA and CycB cyclins clearly belong functionally to the cyclin A/B family since they are effective inducers of meiotic maturation of immature Xenopus oocytes [26, 28, 41 ].

1009
CycB 1
[ CycB plants I
CycB2
B fungi ]
[ B animals [
CvcD3 I A animals [ -G
CycA2
I CycD plants I
CycA plants ] CycD2 ( ~ / / / / / CycAI
CycA3
CycD 1
I D animals I
Figure 1. A phylogenetic tree of eukaryotic cyclins. The tree was obtained with the Darwin system of the Computational Biochemistry Group (Zurich) from the alignment of 190 amino-acids in the most conserved part of the cyclin core from helix HI to helix H5 t (see Fig. 4) of the 61 plant cyclin sequences described in Table 1, and of representative examples of cyclins A, B, D, E, F and G from other eukaryotes. The length of the lines is proportional to the genetic distance between each node. Plant cyclins from the classes CycA and CycB are identified by different symbols indicating their groups (CycA1, =; CycA2, #; CycA3, A; CycB1, *; CycB2, o). Enlarged trees of the CycA and CycB classes are shown in Figs. 2 and 3. The three groups of CycD plant cyclins are identified as CycD1, CycD2 and CycD3. For clarity, species are indicated by the first letter of the generic and specific names. Member numbers are as indicated in Table 1. The six animal cyclins A are indicated as follows: A;a, Helobdella robusta (leech), U25177 (accession number); A;b, Drosophila melanogaster (insect), JC1390; A;c, Chlorohydra viridissima (hydra: coelenterate), X90983; A;d, Spisula solidissima (mollusc), A26328; A;e, Xenopus laevis, S 11678; A;f, Homo sapiens, S08277. The four yeast cyclins B are: B;a, S. cerevisiae CLB3, P24870; B;b, Saccharomyces cerevisiae CLB1, S14165; B;c, S. cerevisiae CLB5, $31290; B;d, Schizosaccharomyces pombe cig2 $44344. The five animal cyclins B are: B;e, Spisula solidissima, A30108; B;f, Gallus gallus B2 (chicken), $23596; B;g, Homo sapiens B1, A32992; B;h, Drosophila melanogaster, A35144; B3, Gallus gallus cyclin B3, $41708. The three human cyclins D are indicated by their names DI (P24385), D2 (P30279) and D3 (P30281). Human cyclin E (P24864) and cyclin F (A55501) and rat cyclin G (P39950) are also indicated.
Sequence analysis of CycD plant cyclins
Plant cyclins with homology to the cyclin D of ver- tebrates were isolated by their ability to rescue yeast mutants lacking endogenous G 1 cyclin activity. Three groups of D-type cyclins (originally named ~ cyclins) were isolated from Arabidopsis [29], and these groups have remained the basis for the definition of plant CycD cyclins, now termed CycDl , CycD2 and CycD3. Sub- sequently a CycD3 cyclin was also identified by yeast complementation from alfalfa [30], and further CycD homologues have been cloned f romAnt irrh inum, Heli-
anthus and Nicotiana by reduced-stringency hybridiz- ation (Table 1).
The overall homology between mammalian cyclin D and plant CycD is only 9-14% identity across the region corresponding to the cyclin core (Figs. 1 and 5; Table 3). However the plant cyclins share a specific fea- ture with their mammalian homologues, which is the presence of the retinoblastoma protein (Rb)-binding motif LxCxE. This sequence is absolutely conserved and is embedded within a somewhat longer stretch of homology at the N-terminus of the protein. In mam- malian cells, phosphorylation of Rb family proteins by

1010
[. CycA3 ]
Figure 2. A phylogenetic tree of plant cyclins of the CycA class. (See Fig. 1 for details). The three groups of CycA plant cyclins are identified as CycAl, CycA2 and CycA3. Species are indicated by the first letter of the generic and specific names. Member numbers are as indicated in Table 1. The numbers close to each line indicate the distances between nodes.
cyclin D-associated kinase activity is required for G1 exit [42]. The LxCxE motif of cyclin D is essential for cyclin D-associated kinases to bind to and phos- phorylate Rb [43]. The same LxCxE motif is also found in the transforming proteins of mammalian tumour vir- uses, and is required for their ability to inactivate pro- teins of the Rb family and provoke cellular transform- ation. Conservation of the Rb-binding motif between mammals and plants, and the recent discovery of a plant Rb homologue [62, 63], indicate that Rb proteins and cyclin D-kinase are likely to have evolved early in the evolution of multicellular eukaryotes.
Despite the relatively small number of CycD cyc- lins that have been sequenced, distinct features can be observed both in the Rb-binding region and within the cyclin core (Fig. 5). In the CycD1 class the leucine in the LxCxE motif is direcly preceded by an acid- ic residue, whereas in CycD2 and CycD3 there is an intervening residue. The consensus sequence of this whole region is different in CycD1 and CycD3. These
Table 3. Homology of plant CycD cyclins. The table shows the % homology between the positions corresponding approximately to the cyclin core (see Fig. 5) amongst the cyclins from one given group or between the three groups of plant and animal D-type cyclins. The % homology within a group is calcu- lated by counting the number of positions at which there is a consensus shown in Fig. 5. The % homology between groups is calcu- lated by counting the positions at which the consensus for the two groups are identical. The data for CycD2 is based on comparison of the Arath;CycD2 sequence with the other consensus sequences.
CycD 1 CycD2 CycD3
CycD1 45
CycD2 24 -
CycD3 20 26 47
Cyclin D 9.4 14 12

1011
Figure 3. A phylogenetic tree of plant cyclins of the CycB class. (See Fig. 1 for details). The two groups of CycB plant cyclins are identified as CycB 1 and CycB2. Species are indicated by the first letter of the generic and specific names. Member numbers are as indicated in Table 1. The numbers close to each line indicate the distances between nodes.
differences could be indicative of the existence of mul- tiple Rb-like proteins in plants, with different affinities for CycD groups.
Carboxy-terminal to the Rb-binding region, CycD proteins have a region of variable length and low homo- logy. Significantly higher conservation is observed over the cyclin core and the degree of conservation is also higher within CycD groups than between groups. Indeed the homology is 45-47% over the cyclin core within CycD groups, and 20-26% between groups (Table 3). All D-type cyclins, including plant CycD, share with cyclins A and B twelve conserved residues in the cyclin core, among which 11 are in the cyc- lin box and five are essential for cyclin activity [16- 18] (Fig. 5). One of these is the absolutely conserved tryptophan residue which in all CycD is part of a dis- tinctive W(I /M)LKV motif corresponding to helix H 1 in mitotic cyclins. The tyrosine of the highly conserved KYEE motif in helix H3 of mitotic cyclins is replaced by an uncharged residue (M,V) in CycD proteins. All
CycD3 sequences have a valine at this position, where- as CycD1 and CycD2 have a methionine at this pos- ition. A further common feature of plant CycD and animal cyclin D is the presence of at least one poten- tial PEST sequence believed to be responsible for the rapid proteolysis of proteins [44, 45]. This sequence is located in the N-terminal or in the C-terminal part of the cyclin or both [29, 30].
Plant cyclin groups show additional complexity
It has recently become clear that individual plant spe- cies have several distinct cyclins within a given group. Thus, for example, three distinct members of the CycA3 group have been found in tobacco and three members of the CycA2 group in Arabidopsis. Their interrelationships are relatively complex, as in some cases they are more similar to another group mem- ber in the same species, whereas in other cases there
1012
CycAl
CycA2
CycBI
CycB2
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . AKKI~ ~:~ N
...................... E...R.TRARA .............
M.S ......... Q
CycAl
CycA2
CycBl
CycB2
~ ................... P..S .................................... KP.P ..........................
.................... R..SKR ................... K K ~ ' ~ F ~ ....................................
........ V.G...Q..KN ...... RN.~!~G~ILV ................ RP..TR.F.AQL .........................
............................ G...G ...... ~ J ~ i ........... VV.KR...L ..................
CycAI
CycA2
CycA3
CycBI
CycB2
...................... V...ETMSTCDS..SPD.EY.DN ........ S.QRKA...L.ISE ..................... D.I.
.............................. ED ..... M ............................... . .......... E...V ..... VV
.............................. K K ~ ~ .............. E ............ KR ....................
................ P..KK ....................................... KK...T.TS.LTARSKAAC.GL.K.K .... I.
...RP..TR .............................. F ............. D .... PM..E ............. EVEM~DI..E ......
A
CycAI
CycA2
CycA3
CycBI
CycB2
B
H N-term HI H2 I
< * ** * ** * * ** **
........... vP-EY..DI..YLRE.E.--K.RPK..YMKK-QPDITNSMR.ILVDWLVK%rGEEYKL..ETL.LAVNY.D~FLS.MSvLRG
D.DN...DPQLCA-T.A.DIY.HLR.AE.--KKRPSTDFMj.IQKDvNPSMRAILIDWLvK~AEE~1~LvPDTLYLTVNYzDR~GN~..R.
DIDS...DPQ.CS-.YA.DIY .... V.EL--.RRP..NYME.LQ.DI.PSMRGILVDWLVEVSEEYKLVPDTLYLTV.LID.FLS...IER.
...... DD~Q.C.-AYvSDIYEYL..ME.E.KRRP...Y.EKvQKDVT..MRGILVDWLvK"dAEEYKLLSDTLYL.VSYIDRFLS..V..R.
DID..D.NNELA.VEYVED~Y.FYK..E.E.R...---YM.-SQPEIN.KMBAILVDWLIEvH.KFEL..ETLYLTINIVDRFLSV..VPRR
DID..DA.NSLAwEYVED.Y.FYR..E..SCvP..--YM.-.Q.D.NEKMR.ILIDWL~Ev~.KFELM.ETLFL~N.IDRFL.KQ.v.RK
D.D.DD..NP .... EYV.DIY.Y.R.LE.--. ...... YL .... --....MR.ILIDWLVQVH.KF.LL.ET.Y..V.I.DRFL .... V...
1 20 40 60 80
A
CycAI
CycA2
CycA3
CycBI
CycB2
B
H3 H4 H5 HI' H2'
** * * * ** * * * * *
KLQLVGTAA..ASKYEEIYPPEV.EFVYITDDTYT..QVLRMZHLVLKVL.FDL..PT.N.F...Y ..... - ...... ..K...L.MF..E
RLQLLGvACMMIASK~EI~APQvEEFCYITDNTYFKDK"~LEM~AS~L~EMTAPT.KCFLRRFvR.AQ...E.P-.LHLE.LA.YIAz KLQLL~VTCMLIASKYEEI.APRVEEFCFITDNT~TK.EVL.M~..VL .... FQL..PT.KTFLRRF..AAQASYK.P-.VELEFLANYLAE
KLQLI/gVSSMLIASI~IZEI.PP.VEDFC.ITDNTYT..~DVL.SL.FEMGNPT.KTFLR.F..VAQED-. .... L..EFLG.YL.E
ELQLVGISAMLIASKYEEIWPPEVNDFV..SD.AY...QIL.MEK.ILG.LEW.LTVPT.YVFLVRF.KAS.- ..... .D...ENMV.FLAE
I~SQLVGL.AMLLACK~/I~EVSVPVV.DLILI.DRA%"~R.DILEMZK..L.TL.FN.SVPT.Y.FMRRFLKAAQ ...... SDKKLEL.A.F.IE
.LQLVGVTAMFIASKYEE...P.I.DFVY.TD...T..QIR..E..IL..L.F.LG.P.P..FLRR.SK..- ..... ..D.E..TLAKYLME
100 120 140 160 180
A
CycAl
CycA2
CycA3
CycBl
CycB2
B
H3' H4' H5' H C-term
* * * ** >
LSL.D.DPYLKYLPSVIA.AA..LA..TV..Q--.WP..L...T.Y.L ..... CL.DLH.T.L.AP.H.QQSIREKYK..KY..VS .... P.
LSLLEY.-.L.YPPSLIAKSAXFLAKFIL.PTK.PWNSTL.HYT.Y.PSDL..CV..LHRL ...... S.LPAIREKYSQIIKYKFVAKK.CPP
LTL.EY.-FL.FLPSLIA••AvFLARWTLDQS.HPWNPTLEHYT.Y..SDLK..v.A.QDL•LNT.GC.L.AIR.KY.Q.KF..vA.L.S•-
L•LLDY.-•V.FLPSLVAASVIFLARFT..P..HPW..ALQ..SGYKP.DLKECVLILHDLQ..RRGG•..AVRDKY..HKFKCvS.L..P.
LGMMHY.-...Y.PSM.AASAVYAARCTL.KT-P.W.ETL..HTGYSE.QL.DCAKLLV ......... KLR.VYKKYS..ERG.VA-..PPA
LSLvEYE-MLKF.PS.LAA.AVYTA••T..G.-..W...CE.H..YSEDQL.ECS..MV..HQKAG.GKLTGVHRKY.T.KFG..A-K.EP.
.T..DY.-MV...PS..AA.A...S.K.L .... --W ...... YS.Y .... L.P .............. T .... V..KY...K..KIS ......
200 220 240 260 275
CycAl .IP-.EFF.N .......
CycA2 -K.V.TLF...
CycA3 ...P..FFED .....
CycBI ......................................
CycB2 A.FLL ......
Figure 4. The consensus sequences of plant cyclins of the CycA and CycB groups. The consensus sequences are deduced from the alignment of the sequences of plant cyclins indicated in Table 1. The A line shows the consensus sequence of the cyclin core of nine animal A-type cyclins and the B line shows the consensus sequence of the cyclin core of B-type cyclins, including 16 cyclins from animals and 8 cyclins from yeast [1]. The residues are indicated when they are similar in 80% at least of the sequences. Residues in bold type are identical in all the sequences of a given group. The dots represents non conserved residues and dashes have been introduced for maximum alignment in the cyclin core domain (starts at <, ends at >; the residues are numbered 1-275). The destruction box is shaded. The residues which are identical in all A- and B-type cyclins in animal and yeast are indicated by stars above the alignment of the cyclin core and bold stars show the residues proven to be essential for cyclin activity. The boxes drawn above the sequences indicate the helix domains of bovine cyclin A [20].

1013
CycDl
CyCD2
CycD3
CycD
a]] D
MS.S.S .... DL.C.EDS ........................... D ............. D. . .SIA .... E.ER .... G ....... Q .....
....... MAENLACGETSESWI I DNDDDD INYGGGFTNE IDYNHQLFAKDDNFGGNGS I PMMGS S S S SLSED. RIKEMLVREIEFCPGT.. D
..... S. . .DAL.C.EE ...................................... L. . .DL ......... E. EEL. .L. .KE.E ........
........... L.C.E ...................................................... E .....................
........... LLCCE ................................. RA. PD.. LL.. , RVL... L.. EE... P.. SYF. CVQK. I. P..
........... L.C.E ...................................................... E .....................
< 20
CycDI
CycD2
CycD3
CycD
m a/Am D
a]] D
. . . . . . . . . . D.S.R..S.AWILKVQ..Y.FQPL.AYL.V.Y.DRF . . . . . . P...GWP.QLL.VACLSLAAKMEE.L.PS..D.QV.G...
Y V K R L L S G D L D L ~ R N Q A L D W ~ L K ~ C A H Y H F G H L ~ I C L S ~ N Y L D R F L T S Y E L P K D K D W A A Q L L A V S ~ L S L A S K M E E T D V P H T V D L Q V E D P K F
.......... L...R.E.V.W.LKV...YGF..LTA.LA.NYLDRF ...... Q.DKPWM.QL.AV.CLS~ETQVPLLLD.QVE..Ky
.............. R ..... W.LKV...Y.F..L.A.L...Y.DRF ........... W..QL..V.CLSLAAK.EE..VP...D.QV...K.
............. MR...A.WMLEVCEEQ.CEEEVFPLAMNYLDR.L...p..K ..... LQLLGA.CM..ASK..ET..PLT.EKLCIYTD.
.............. R ..... W.L.V ............ L...Y.DR ............... QL .... C...A.K..E...p ............ * * * ** ** * * *
4(] 6[) 80 i00 ]20
CycDI
CycD2
CycD3
CycD
marmm D
all D
.FE... I.RME. LVL.VLDWRLRS, .PF .... FF..KID ....... G.L.S ..... ILS...E.S..EY.PS. IAAA..L..A..L...S..
VFEAKTIKRMELLVVTTLNWRLQALTPFSFIDYFVDKISGh~/SENLIYRSSR... FILNTTKAIEFLDFRPSEIAAAAAVSVSISGETECID
VFEAKTIQRMELL . LSTLKW. M. PVTP . SF. DHI . ILl{b . r. ....... F .... CE. . . LS . . . D. RFV. . . PSV. A. ATM. . VI .... P ....
.FE. K. I.RM~LL .... L.W ..... TP..F ........................... L ............ PS..A.A ...............
• . .P. .L. . .E ..... KLKW.LAA. , .HDF .... L .............. KHAQTF.ALCAT.D. .F. . .PPSM.A.GS . .AA. .GL ......
.......... E ...... L.W ......... F ........................... L ............ PS..A .................
140 160 180 200 220
CycDl
CycD~
CycD3
CycD
mamm D
all D
• . .H. . .E.WCDGL.K. .I. .C. .L .................. PK ...... RV ............... S ..... S .......... S.S. .K.
EEKALSSLIY... ~q4QERVKRCLNLMRSLTGEENVRGTSLSQEQARVAVRA.. VPASPVGVLEATCL. SYR. SEERTVESCTNSSQSSPDNN
. . .Y...L...L...K.K...C..LI .................. KRK ....... P. SP °GVID ..... S...SN.SW .......... S ....
............... K ..... C..L ................... K ....................... S...S .............. S ....
....... LT. . L ........ DCL. ACQE . QTEA. L . . SL .... Q ...................... TPTDV .....................
..................... C ......................................................................
240 260 274>
CycDI
CycD2
CycD3
CycD
man~ D
all D
R K L . . . S . . . . . . . . . . . . . . . . . . . . . . . . .
NNNNNSKQEEEKTMRENKRVIHCLLQPKTTST
• .P..KK .........................
Figure 5. The consensus sequences of plant cyclins of the CycD groups• (See Fig. 4 for details). The CycD2 line shows the complete sequence ofArabidopsis CycD2. The CycD and mammD lines show positions at which at least 80% or 100% (bold type) CycD sequences in the CycD class and in vertebrate cyclins D, respectively, have identical residues• The sequences for human, mouse and rat DI, D2 and D3, and chicken D2 were used. The all D line shows positions at which cyclin D and CycD sequences have the same conserved residues. The stars indicate the residues which are identical in all eukaryotic cyclin A, B and D, and bold stars shows the residues essential for cyclin activity• The cyclin core (numbered) is delimited between < and >.
is a closer relative in a different species, suggesting that these group members may have distinct functions. Thus, for example, of the three CycA2 group members found in Arabidopsis, two are most closely related to each other, but the third is closer to Brassica napus, pea and alfalfa CycA2 cyclins (Fig. 2). These data reflect the great complexity of plant genome evolution, and presumably indicate the occurence of both recent and more ancient gene duplication events in plant cyclins, as well as genome fusions in the creation of certain species. This has resulted in the presence of large num- bers of cyclin genes in many plants, the significance of which has not yet been evaluated functionally and which leads one to expect that the number of plant cyc- lins in the groups defined above will continue to grow in the future. It also suggests that there may be difficulty
in assessing the specific roles of each of these cyclins in the plant cell cycle and during plant development.
The complexity of relationships and the relatively small number of cyclins currently within each group leads us to conclude that further sub-divisions within cyclin groups cannot be defined at present. However we leave open the possibility for defining sub-groups in the future should a meaningful plant-wide basis for further sub-divisions become apparent (see below).
C P G N gene nomenclature for plant cyclins
The nomenclature rules of the CPGN define the formal structure of plant gene names as consisting of a spe- cies acronym (as used in SwissProt), the gene family name, and a member number (MN) [31, 32]. Mem-

1014
ber numbers identify the individual coding sequence of a member of a multigene family within a species. We therefore suggest that each cyclin group should be treated as a multigene family, with the genes encoding different members of each family being indicated by individual MN.
The full formal names of the three Ara- bidopsis (abbreviation Arath) CycA2 genes under CPGN nomenclature are therefore: Arath;CycA2;1, Arath;CycA2;2, Arath;CycA2;3. In Table 1 we present the revised nomenclature for all plant cyclins known to us. It should be noted that MN are allocated only by the CPGN when the sequences of cloned genes acquire an EMBL/Genbank accession number. Table 1 there- fore shows provisional MN for genes currently without accession numbers.
MN are allocated independently and sequentially for each plant species, and there is therefore no rela- tionship between member numbers of a gene family in different species. This is unimportant if all members of a group have very similar or identical functions, since in this case referring to 'CycA2' describes a consist- ent pattern of expression and function across species for all member genes. However, the greater conserva- tion of some group members between different species is indicative of functional distinctions. Furthermore, it has recently become apparent that group members can have dramatically different expression patterns as recently described for three CycA3 homologues of Nicotiana tabacum [61] and two CycD3 homologues of Antirrhinum (Gaudin et al., unpublished).
We therefore suggest the future possibility of cre- ating further sub-divisions within cyclin groups. Such divisions may be useful to indicate either a further degree of sequence conservation (subgroups) or a con- served and independent function of certain group mem- bers across different plant species. The CPGN rules permit gene names up to 8 characters, so we pro- pose that if subgroups are created these are indicated by a lower-case letter appended to the gene family name. This would therefore indicate distinct subgroups with plant-wide significance. However the gene fam- ily will continue to be regarded at the group level and MN will therefore remain unaltered. Thus genes encoding two CycD3 cyclins have been identified in Antirrhinum (Antma; Cyc D 3 ;1 and Antma; CycD 3 ;2 ), which have distinct expression patterns. If this beha- viour is conserved in other species, the creation of two CycD3 subgroups, called CycD3a and CycD3b, could be justified. The formal gene names would there- fore become Antma;CycD3a; 1 and Antma;CycD3b;2.
A similar proposal has been adopted for naming his- tone variants that differ in expression patterns [46].
The use of such a suffix would thus indicate sim- ilar expression patterns and roles or highly conserved sequences in different species. We believe that this proposal allows maximum flexibility for future devel- opments.
Functional significance of cyclin classes
The naming of cyclins in all species has been on the basis of sequence similarity to homologues known to have distinct functions. We have adopted the sequence relationships principle as the basis for naming plant cyclins, resulting in their assembly into A, B and D classes. However, this naming by no means signifies that the plant cyclins are necessarily true functional homologues of yeast or animal A-, B- or D-type cyc- lins. The little data available to date indicate however that CycA, CycB and CycD genes exhibit different reg- ulation during the cell cycle. The steady-state levels of CycA RNAs increase at or after the onset of S phase till G2 and occasionally M phases [21, 27, 28, 47-49]. The CycB RNAs examined to date are only present in G2 and M phases [21, 27, 47-54]. The general tim- ing of CycA and CycB expression is consistent with the expression patterns of their homologues in mammalian cells, in which cyclin A is expressed during S phase, accumulates further during G2 and is destroyed dur- ing mitosis, and cyclin B is expressed somewhat later during G2 and is also destroyed during mitosis.
Interestingly, the cyclins of different CycA groups [27] and of different CycB groups [28] have differ- ent characteristics of expression when assayed in the same biological system, although these data remain too scarce to assess their validity as defining the behaviour of the whole group. In two recent cases however the expression of CycA3 genes has been found to occur earlier in S phase and to be much more restricted to this phase than that of the two other CycA groups [27, 61]. One CycA2 cyclin has been found to be potentially involved in the reentry of quiescent cells into division in alfalfa [49], although this is probably not a general feature of the CycA2 group [28, 47, 61 ].
CycD groups show clear differential expression patterns of mRNAs between tissues [29], and show dif- ferential cell cycle regulation and different responses to sucrose and plant hormones in culture systems [29, 55]. The regulation of CycD3 in synchronized Ara- bidopsis suspension cultures cells is reminiscent of the

behaviour of mammalian cyclin D [55], suggesting a potential analogous role to their mammalian homo- logues. Recent data also suggest that two CycD3 genes in Antirrhinum have distinct meristematic expression patterns (Gaudin et al., unpublished), indicating that functional differences may also be present between different CycD3 group members.
Summary: The nomenclature of plantcyclins
Plant cyclins are defined according to class and group. Within a group, individual genes from a particular spe- cies are identified by a MN. The formal gene name is therefore made up of species identifier, cyclin group name and MN.
Class. Cyclin A, B or D. New cyclins with great- er homology to other vertebrate cyclin classes can be named accordingly.
Group. Currently recognised groups have gene names CycA1, CycA2, CycA3, CycB1, CycB2, CycD1, CycD2 and CycD3.
Member number. MN are allocated by the CPGN when an EMBL/GenBank accession number is provided. They are allocated sequentially for genes in each cyclin group in any particular species.
Subgroup. Subgroups are not currently defined but may be designated in the future as functional ana- lysis and increased numbers of sequences demon- strate sequence conservation between species or dis- tinct functions for different members of a defined cyc- lin group. The use of a given subgroup suffix should indicate highly homologous sequences or equivalent functional roles between several species. They will be indicated by a lower-case letter suffix to the gene name, but this will not affect the MN, which is allocated at the group level.
The formal gene name for a plant cyclin of classes A, B or D will therefore be of the form shown in Fig. 6, although the species abbreviation and MN may be omitted after the first citation.
Gene products. CPGN nomenclature guides only the naming of genes, but we propose that the pro- tein product is conveniently named in a similar man- ner. When the plant species involved is evident from the context, the protein product may be called, for example, CycA1 in the case where only a single gene is known in that cyclin group. The form 'cyclin A I ' is also acceptable. When it is necessary to refer to a particular MN of a group, the form CycA1;1 is pre- ferred. If subgroups are subsequently defined, the pro-
1015
M a t h ; CycA1; 1 , member number, / / ~ distinguishing multiple
genes in the same group
species abbreviation as in Swissprot ] cyclin ~oup
cyclin class, / three letter designation indicating closest for cyclin homology to
animal cyclin class
Figure 6. Gene nomenclature for plant cyclins.
tein product of members of distinct subgroups can be referred to, for example, as CycD3a and CycD3b, or cyclin D3a and cyclin D3b.
Allocation of names to new cyclins and cyclin-like proteins
The class and group of a new cyclin related to CycA, CycB or CycD can be assigned using the guidelines in this paper. A file containing the acces- sion numbers of all plant cyclins sequences and sample metazoans and fungi cyclins currently present in sequence databases can be obtained from the Mendel WWW server (http://probe.nalusda.gov:8300/cgi- bin/browse/mendel). This can be used to run sequence comparisons as presented here on the Darwin system of the Computational Biochemistry Group at Zurich (program PhyloTree: send message containing 'help' in the body to [email protected]). Proposed gene names should then be submitted with the relevant data to the CPGN Registry (either at John Innes Centre, Nor- wich, UK: [email protected] or at Waksman Institute, Rutgers, USA: [email protected]) as decribed in [32]. CPGN will then issue a MN for the gene sequence. According to normal CPGN procedure, decisions are normally returned within two weeks and new gene designations posted on Mendel within one month.
A cyclin clearly belonging to an existing class, but not falling within current groups may necessitate the creation of a new group. Proposals for new groups should be made to CPGN. Cyclins with clear relation- ships to other mammalian cyclin classes will be named accordingly, eg CycC for a cyclin C homologue. Cyc- lins of unknown class or with no homology will be designated Cyc0 (zero).

1016
Conclusion: A rational classification and nomenclature for plant cyclins
Whilst there is no specific logic to the naming of animal cyclin classes, the nomenclature is now widely accep- ted and understood. Recent analysis has shown that A and B-like plant cyclins can be unambiguously defined as being related to one of these two animal cyclin classes [ 1 ], indicating that the cyclin classes are con- served between multicellular eukaryotes. We there- fore propose that the same nomenclature be adopted for plant cyclin classes, replacing a number of cur- rent nomenclatures [26, 27, 29], of which the most prevalent is the independent numbering of cyclins in each plant species. The relationship of one plant cyclin to another is impossible to deduce from their current names (see Table 1, old names), nor does the current naming indicate sequence relationships to the cyclins of other eukaryotes. Here we develop the nomenclature proposed in [1] and define plant cyclins of A, B and D classes, but the same system is equally applicable to further plant cyclins with homology to an existing animal class.
Each class of plant cyclins contains groups which can be clearly defined by features located in various positions in their protein sequences. These groups are identified by numbers 1, 2 and 3. In certain cases, mul- tiple cyclins from a single species are found in a group. These are regarded as members of a multigene family, and assigned a MN by the CPGN. We have left open the possibility of creating subgroups, if conserved struc- tural or functional differences are found between group members, but in this case the MN remain unaltered.
We believe our proposal has a number of important advantages. 1. It indicates plant-wide homologies. 2. It indicates homologies with animal and fungal cyc- lins. 3. The proposed system will absorb the large increase in the number of plant cyclins that we anticipate to be described over the next few years, whilst still main- taining their interrelationships. 4. It will facilitate communication between research- ers working with different plant species by providing a common nomenclature. 5. It will stimulate functional analysis by allowing easy comparison between species. 6. It will stimulate communication and interaction among cell cycle researchers in animal, plant and fungai systems.
We therefore believe that the plant-wide classific- ation of plant cyclins will provide a valuable tool to understand their various functions during the plant cell cycle.
Acknowledgements
We thank all the authors who have communicated data prior to publication, and notably J. Deckert, J. Jelenska, V. Gaudin, S. Goormachtig, W.L. Hsieh and S. Wolniak. We also thank T. Hunt, J. Pines, C. Price and D. Lonsdale for invaluable comments on the manuscript and guidance on CPGN nomenclature.
References
1. Renaudin JP, Savour6 A, Philippe H, Van montagu M, Inz6 D, Rouz6 P, Characterization and classification of plant cyclins sequences related to A and B-type cyclins. In: Francis D, Dudits D, Inz6 D (eds) Plant Cell Division, Portland Press, London, in press.
2. Pines J: Cyclins and cyclin-dependent kinases: a biochemical view. Biochem J 308:697-711 (1995).
3. Nigg EA: Cyclin-dependent protein kinases: key regulators of the eukaryotic cell cycle. Bioessays 17:471-480 (1995).
4. Norbury C, Nurse P: Animal cell cycles and their control. Annu Rev Biochem 61:441-470 (1992).
5. Evans T, Rosenthal ET, Youngblom J, Distel D, Hunt T: Cyclin: A protein specified by maternal mRNA in sea urchin eggs that is destroyed at each cleavage division. Cell 33:389-396 (1983).
6. Meijer L, Arion D, Golsteyn R, Pines J, Brizuela L, Hunt T, Beach D: Cyclin is a component of the sea urchin egg M-phase specific histone HI kinase. EMBO J 8:2275-2282 (1989).
7. Nakamura T, Sanokawa R, Sasaki YE Ayusawa D, Oishi M, Mori N: Cyclin I: a new cyclin encoded by a gene isolated from human brain. Exp Cell Res 221:534-542 (1995).
8. Pines J: Cyclins and cyclin-dependent kinases: Theme and variations. Adv Cancer Res 66:181-212 (1995).
9. Fitch I, Dahmann C, Surana U, Amon A, Nasmyth K, Goetsch L, Byers B, Futcher B: Characterization of four B-type cyclin genes of the budding yeast Saccharomyces cerevisiae. Mol Biol Cell 3:805-818 (1992).
10. Fisher D, Nurse P: Cyclins of the fission yeast Schizosacchar- omyces pombe. Semin Cell B iol 6:73-78 (1995).
11. Schwob E, Nasmyth K: CLB5 and CLB6, a new pair of B cyc- lins involved in DNA replication in Saccharomyces cerevisiae. Genes Devel. 7:1160-1175 (1993).
12. Cross FR: Starting the cell cycle: What's the point? Curr Opin Cell Biol 7:790-797 (1995).
13. Dirick L, Bohm T, Nasmyth K: Roles and regulation of Cln- Cdc28 kinases at the start of the cell cycle of Saccharomyces cerevisiae. EMBO J 14:4803-4813 (1995).
14. Nugent JHA, Alfa CE, Young T, Hyams JS: Conserved struc- tural motifs in cyclins identified by sequence analysis. J Cell Sci 99:669-674 (1991).

15. O'Farrell P, Leopold P: A consensus of cyclin sequences reveals homology with the ras oncogene. Cold Spring Harbor Symp Quant Biol 56:83-92 (1991).
16. Zheng XF, Ruderman JV: Functional analysis of the P Box, a domain in cyclin B required for the activation of Cdc25. Cell 75:155-164 (1993).
17. Kobayashi H, Stewart E, Poon R, Adamczewski JP, Gannon J, Hunt T: Identification of the domains in cyclin A required for the binding to, and activation of, p34 cdc2 and p32 cdk2 protein kinase subunits. Mol Biol Cell 3:1279-1294 (1992).
18. Lees EM, Harlow E: Sequences within the conserved cyclin box of human cyclin A are sufficient for binding to and activ- ation of cdc2 kinase. Mol Cell Biol 13:1194-1201 (1993).
19. Jeffrey PD, Russo AA, Polyak K, Gibbs E, Hurwitz J, Mas- sagud J, Pavletich NP: Mechanism of cdk activation revealed by the structure of a cyclinA-CDK2 complex. Nature 376: 313-320 (1995).
20. Brown NR, Noble MEM, Endicott JA, Garman EF, Wakat- suki S, Mitchell E, Rasmussen B, Hunt T, Johnson LN: The crystal structure of cyclin A. Structure 3:1235-1247 (1995).
21. Shaul O, Van montagu M, Inzd D: Regulation of cell division in Arabidopsis. Crit Rev Plant Sci 15:97-112 (1996).
22. Jacobs T: Control of the cell cycle. Devel Biol 153:1-15 (1992).
23. Jacobs TW: Cell cycle control. Annu Rev Plant Physiol Plant Mol Biol 46:317-339 (1995).
24. Ferreira P, Hemerly A, Van Montagu M, Inzd D: Control of cell proliferation during plant development. Plant Mol Biol 26:1289-1303 (1994).
25. Francis D, Halford NG: The plant cell cycle. Physiol Plant 93: 365-374 (1995).
26. Renaudin JP, Colasanti J, Rime H, Yuan Z, Sundaresan V: Cloning of four cyclins from maize indicates that higher plants have three structurally distinct groups of mitotic cyclins. Proc Natl Acad Sci USA 91:7375-7379 (1994).
27. Kouchi H, Sekine M, Hata S: Distinct classes of mitotic cyclins are differentially expressed in the soybean shoot apex during the cell cycle. Plant Cell 7:1143-1155 (1995).
28. Ferreira P, Hemerly A, Engler JD, Bergounioux C, Burssens S, Van Montagu M, Engler G, Inz6 D: Three discrete classes of Arabidopsis cyclins are expressed during different intervals of the cell cycle. Proc Natl Acad Sci USA 91:11313-11317 (1994).
29. Soni R, Carmichael JP, Shah ZH, Murray JAH: A family of cyclin D homologs from plants differentially controlled by growth regulators and containing the conserved retinoblastoma protein interaction motif. Plant Cell 7:85-103 (1995).
30. Dahl M, Meskiene I, Bogre L, Ha DTC, Swoboda I, Hub- mann R, Hirt H, Heberlebors E: The D-type alfalfa cyclin gene cycMs4 complements G(I) cyclin-deficient yeast and is induced in the G( 1 ) phase of the cell cycle. Plant Cell 7:1847- 1857 (1995).
31. Lonsdale DM, Price CA: Eukaryotic gene nomenclature: a resolvable problem? Trends Biochem Sci, in press (1996).
32. Price CA, Reardon EM, Lonsdale DM: A guide to naming sequenced plant genes. Plant Mol B iol 30:225-227 (1996).
33. Herendeen PS, Crane PR, The fossil history of the monocoty- ledons. In: Ruddall et aL (ed) Monocotyledons: Systemat- ics and Evolution, pp. 1-21. Royal Botanical Gardens, Kew (1995).
34. Glotzer M, Murray AW, Kirschner MW: Cyclin is degraded by the ubiquitin pathway. Nature 349:132-138 (1991).
35. Lorca T, Devault A, Colas P, VanLoon A, Fesquet D, Lazaro JB, Dorde M: Cyclin A-Cys41 does not undergo cell cycle-
1017
dependent degradation in Xenopus extracts. FEBS Lett 306: 90-93 (1992).
36. Pines J, Hunter T: The differential localization of human cyc- lins A and B is due to a cytoplasmic retention signal in cyclin B. EMBO J 13:3772-3781 (1994).
37. Pines J, Hunter T: Human cyclins A and B1 are differentially located in the cells and undergo cell cycle dependent nuclear transport. J. Cell Biol 115:1-17 (199l).
38. Ookata K, Hisanaga S, Bulinski JC, Murofushi H, Aizawa H, Itoh TJ, Hotani H, Okumura E, Tachibana K, Kishimoto T: Cyclin B interaction with microtubule-associated protein 4 (MAP4) targets p34(cdc2) kinase to microtubules and is a potential regulator of M-phase microtubule dynamics. J Cell Biol 128:849-862 (1995).
39. Jackman M, Firth M, Pines J: Human cyclins B1 and B2 are localized to strikingly different structures: B 1 to microtubules, B2 primarily to the Golgi apparatus. EMBO J 14:1646-1654 (1995).
40. Furuya M: Cell division patterns in multicellular plants. Annu Rev Plant Physiol 35:349-373 (1984).
41. Hata S, Kouchi H, Suzuka I, Ishii T: Isolation and characteriza- tion of cDNA clones for plant cyclins. EMBO J 10:2681-2688 (1991).
42. Weinberg RA: The retinoblastoma protein and cell cycle con- trol. Cell 81:323-330 (1995).
43. Dowdy SF, Hinds PW, Louie K, Reed SI, Arnold A, Weinberg RA: Physical interaction of the retinoblastoma protein with human D cyclins. Cell 73:499-511 (1993).
44. Rechsteiner M: Regulation of enzyme levels by proteolysis: the role of PEST regions. Adv Enzyme Regul 27:135-151 (1988).
45. Rogers S, Wells R, Rechsteiner M: Amino acid sequences common to rapidly degraded proteins: the PEST hypothesis. Science 234:364-368 (1986).
46. Gigot C, Spiker S: Nomenclature of genes encoding histones. Plant Mol Biol Rep 12:$39-$40 (1994).
47. Shaul O, Mironov V, Burssens S, Van montagu M, Inzd M: Two Arabidopsis cyclin promoters mediate distinctive tran- scriptional oscillation in synchronized tobacco BY-2 cells. Proc Natl Acad Sci USA 93:4868-4872 (1996).
48. Setiady YY, Sekine M, Hariguchi N, Yamamoto T, Kouchi H, Shinmyo A: Tobacco mitotic cyclins: cloning, characteriza- tion, gene expression and functional assay. Plant J 8:949-957 (1995).
49. Meskiene I, Bogre L, Dahl M, Pirck M, Ha DTC, Swoboda I, Heberlebors E, Ammerer G, Hirt H: cycMs3, a novel B-Type alfalfa cyclin gene, is induced in the G(0)-to-G(1) transition of the cell cycle. Plant Cell 7:759-771 (1995).
50. Hirt H, Mink M, B6gre L, Gy6rgyey J, Jonak C, Gartner A, Dudits D, Heberle-Bors E: Alfalfa cyclins: differential expres- sion during the cell cycle and in plant organs. Plant Cell 4: 1531-1538 (1992).
51. Fobert PR, Coen ES, Murphy GJP, Doonan JH: Patterns of cell division revealed by transcriptional regulation of genes during the cell cycle in plants. EMBO J 13:616-624 (1994).
52. Sauter M, Mekhedov SL, Kende H: Gibberellin promotes his- tone H 1 kinase activity and the expression of cdc2 and cyclin genes during the induction of rapid growth in deepwater rice internodes. Plant J 7:623-632 (1995).
53. Savourd A, Feher A, Kalo P, Petrovics G, Csanadi G, Szecsi J, Kiss G, Brown S, Kondorosi A, Kondorosi E: Isolation of a full- length mitotic cyclin cDNA clone CyclIIMs from Medicago sativa: chromosomal mapping and expression. Plant Mol Biol 27:1059-1070 (1995).

1018
54. Hemerly A, Bergounioux C, Van Montagu M, Inz6 D, Fer- reira P: Genes regulating the plant cell cycle: Isolation of a mitotic-like cyclin from Arabidopsis thaliana. Proc Natl Acad Sci USA 89:3295-3299 (1992).
55. Fuerst RAUL, Soni R, Murray JAH, Lindsey K: Cyclin tran- scripts in synchronized Arabidopsis cells. Plant Physiol, in press (1996).
56. Szarka S, Fitch M, Schaerer S, Moloney M: Classification and expression of a family of cyclin gene homologues in Brassica napus. Plant Mol Biol 27:263-275 (1995).
57. Day IS, Reddy ASN, Golovkin M: Isolation of a new mitotic- like cyclin from Arabidopsis: complementation of a yeast cyc- lin mutant with a plant cyclin. Plant Mol Biol 30:565-575 (1996).
58. Deckert J, Jelenska J, Gwozdz EA, Legocki AB: The isolation of a lupine cDNA clone coding for a putative cyclin protein. Biochimie 78:90-94 (1996).
59. Qin LX, Richard L, Perennes C, Gadal P, Bergounioux C: Identification of a cell cycle-related gene, cyclin, in Nicotiana tabacum (L). Plant Physiol 108:425--426 (1995).
60. Logemann E, Wu SC, Schroder J, Schmelzer E, Somssich IE, Hahlbrock K: Gene activation by UV light, fungal elicit- or or fungal infection in Petroselinum crispum is correlated with repression of cell cycle-related genes. Plant J 8:865-876 (1995).
61. Reichheld JP, Chaubet N, Shen WH, Renaudin JP, Gigot C: Multiple A-type cyclins express sequentially during the cell cycle in Nicotiana tabacum BY2 cells. Proc Natl Acad Sci USA, in press (1996).
62. Murray JAH, Freeman D, Greenwood J, Huntley R, Makkerh J, Riou-Khamlichi C, Sorrell DA, Cockcroft C, Carmichael JP, Soni R, Shah ZH: Plant D cyclins and retinoblastoma (Rb) protein homologues. In: Francis D, Dudits D, Inz6 D, (eds.), Plant Cell Division. Portland Press, London, in press (1996).
63. Xie Q, Sanz-Burgos AP, Hannon GJ, GutiErrez C: Plant cells contain a novel member of the retinoblastoma family of growth regulatory proteins. EMBO J, 15:4900---4908 (1996).