physical mapofthe myxococcus xanthus chromosome
TRANSCRIPT
JOURNAL OF BACTERIOLOGY, Mar. 1991, p. 2109-2115 Vol. 173, No. 60021-9193/91/062109-07$02.00/0Copyright C 1991, American Society for Microbiology
Physical Map of the Myxococcus xanthus ChromosomeHONGWU CHEN,' ADAM KUSPA,2 INGRID M. KESELER,lt AND LAWRENCE J. SHIMKETS1*
Department of Microbiology, University of Georgia, Athens, Georgia 30602,1 and Department of Biology, University ofCalifornia, San Diego, La Jolla, California 920932
Received 21 August 1990/Accepted 7 January 1991
The genome of Myxococcus xanthus, which is 9,454 kbp, is one of the largest bacterial genomes. Theorganization of the DNA and the distribution of genes encoding social and developmental behaviors wereexamined by using pulsed field gel electrophoresis. Intact genomic DNA was digested with AseI into 16restriction fragments, which were separated by contour-clamped homogeneous electric field electrophoresis,purified, and radiolabeled. Each AseI fragment was hybridized to SpeI-digested DNA and to an M. xanthusgenomic library contained in yeast artificial chromosomes. Some SpeI restriction fragments and yeast artificialchromosome clones contained AseI sites and hybridized with two different AseI restriction fragments, providingevidence for the juxtaposition of these AseI restriction fragments in the chromosome. The deduced AseIphysical map is circular, suggesting that this bacterium contains a single, circular chromosome. Transposableelements shown by transduction to be in or near genes of interest were located on specific AseI restrictionfragments by restriction analysis and Southern hybridization. Most AseI restriction fragments contained genesinvolved in social and developmental behaviors.
The myxobacterium Myxococcus xanthus grows in thesoil, where it feeds on organic material and preys on othermicroorganisms (for reviews, see references 31 to 33). Uponstarvation on a solid surface, thousands of bacteria aggregateby gliding and form a fruiting body containing dormantmyxospores. This process requires social interactions be-tween cells, and much of the Myxococcus research focuseson the molecular basis of these social and developmentalbehaviors.The genome size of M. xanthus is 9,454 kbp, which is
unusually large for a bacterium (4). Since an efficient conju-gation system for mapping chromosomal genes has not beendevised, the physical organization of the DNA and the orderof genes on the DNA are not known. Currently, geneticmapping is performed by transposon tagging and generalizedtransduction, which is limited to a range of about 60 kbp (12).We examined the organization of the genome with pulsedfield gel electrophoresis and found it to be composed of asingle, circular chromosome. Genes involved in the socialand developmental behavior of this organism were found inmany regions of the chromosome.
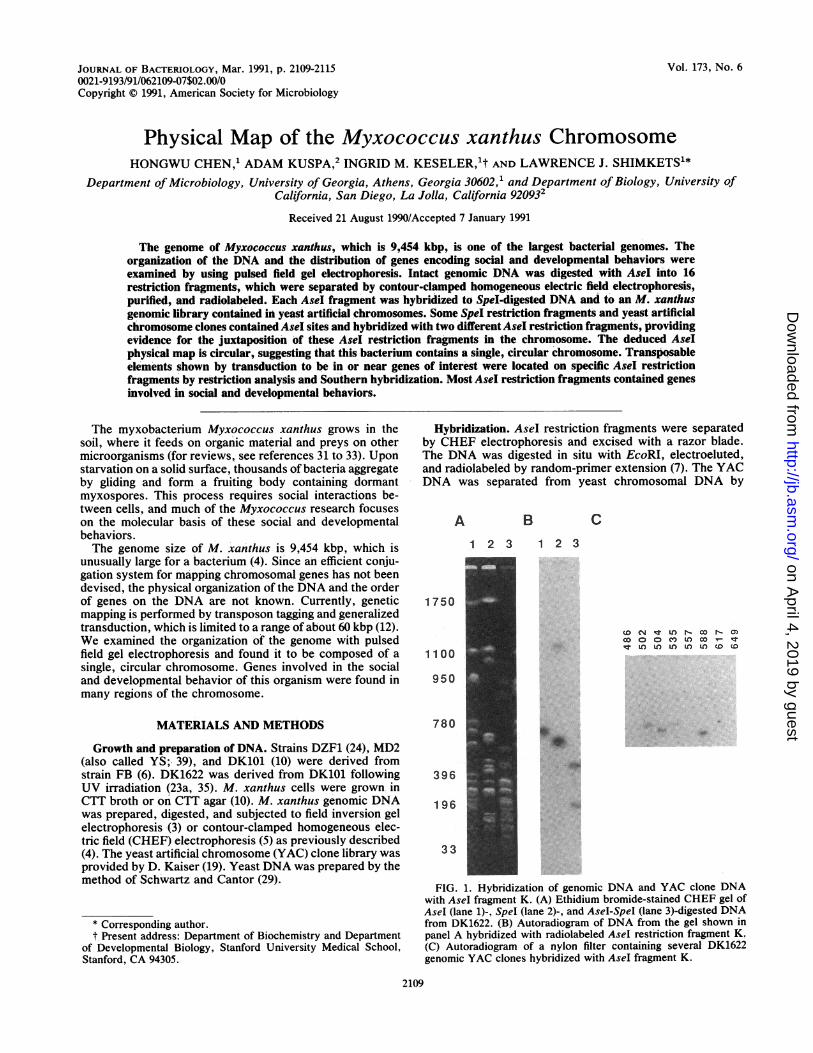
Hybridization. AseI restriction fragments were separatedby CHEF electrophoresis and excised with a razor blade.The DNA was digested in situ with EcoRI, electroeluted,and radiolabeled by random-primer extension (7). The YACDNA was separated from yeast chromosomal DNA by
A B C1 2 3 1 2 3
1 750
1100
950
CD C' Ri I r~- cc N1- 0Co 0 0 C' In COw
MATERIALS AND METHODS
Growth and preparation of DNA. Strains DZF1 (24), MD2(also called YS; 39), and DK101 (10) were derived fromstrain FB (6). DK1622 was derived from DK101 followingUV irradiation (23a, 35). M. xanthus cells were grown inCTT broth or on CTT agar (10). M. xanthus genomic DNAwas prepared, digested, and subjected to field inversion gelelectrophoresis (3) or contour-clamped homogeneous elec-tric field (CHEF) electrophoresis (5) as previously described(4). The yeast artificial chromosome (YAC) clone library wasprovided by D. Kaiser (19). Yeast DNA was prepared by themethod of Schwartz and Cantor (29).
* Corresponding author.t Present address: Department of Biochemistry and Department
of Developmental Biology, Stanford University Medical School,Stanford, CA 94305.
780
396
196
33
FIG. 1. Hybridization of genomic DNA and YAC clone DNAwith AseI fragment K. (A) Ethidium bromide-stained CHEF gel ofAseI (lane 1)-, Spel (lane 2)-, and AseI-SpeI (lane 3)-digested DNAfrom DK1622. (B) Autoradiogram of DNA from the gel shown inpanel A hybridized with radiolabeled Asel restriction fragment K.(C) Autoradiogram of a nylon filter containing several DK1622genomic YAC clones hybridized with AseI fragment K.
2109
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
2110 CHEN ET AL.
TABLE 1. Juxtaposition of AseI restriction fragments inthe M. xanthus chromosome
Linkage of Linking SpeI YAC linkingAseI fragments fragment clone(s)
A-B H 119, 890, 1464B-G N 1387, 1512G-J D 1751J-O R NoneO-K T NoneK-D Q 535, 1400D-L 0 818L-I F 1546I-P P 1305P-E J 799, 1242E-F U 948F-H U 1451H-C U 706, 735C-M U 984M-N U 174, 883, 1410, 1881N-A H 1730
CHEF electrophoresis. DNA was transferred to nylon filtersand baked (21). Prehybridization was performed at 42°C for2 h in a buffer containing 6x SSPE, 1Ox Denhardt solution,0.5% sodium dodecyl sulfate (SDS), and 100 ,ug of sonicatedsalmon sperm DNA per ml. Hybridization was carried outovernight at 68°C in the same buffer except for the absenceof the Denhardt solution. Membranes were washed twice in1x SSPE-1% SDS at 37°C for 15 minutes each and then in0.lx SSPE-1% SDS for 2 h at 69°C.
RESULTS
The structure of a genome can be determined by construc-tion of a physical map. The genome of M. xanthus DK1622contains 16 AseI restriction fragments and 21 SpeI restric-tion fragments (4), and the objective of this work was todetermine the order of these fragments in the intact genome.Each of the AseI restriction fragments was purified, radio-labeled, and probed against Spel-digested DNA, AseI-SpeI-double-digested DNA, and an M. xanthus DK1622 genomiclibrary contained in 145 YAC clones. Many of the SpeIrestriction fragments, referred to as linking fragments, con-tain an AseI site and consequently hybridize with twodifferent AseI fragments, providing evidence that these AseIfragments are adjacent to each other in the native genome.Similarly, some of the YAC clones will contain an AseI siteand hybridize with two different Asel restriction fragments;these clones are referred to as linking clones.An example of the hybridization results with AseI frag-
ment K used as the probe is shown in Fig. 1. Figure 1A is anethidium bromide-stained agarose gel of AseI (lane 1)-, SpeI(lane 2)-, and Asel-Spel (lane 3)-digested DNA; Fig. 1B is anautoradiogram obtained by hybridization of these DNAsamples with radiolabeled AseI fragment K. The probehybridizes with only AseI fragment K (lane 1), indicatingthat it is a specific probe. It also hybridizes, with Spelrestriction fragments Q (637 kbp) and T (1,118 kbp; lane 2),indicating that these fragments are adjacent to each other inthe intact genome. Fragment K also hybridized with AseI-SpeI-double-digested fragments of 490 and 228 kbp (lane 3).DNA from an M. xanthus library contained in 145 YACclones was separated by CHEF electrophoresis, transferredto a nylon membrane, and probed with AseI fragment K.Figure 1C shows the three types of hybridization results that
TABLE 2. Hybridization of AseI restriction fragments withSpeI-digested and AseI-SpeI-digested DNA from DK1622
AseI fragment Hybridization detected with fragments:probea SpeI-digesteda AseI-SpeI-digested'
A (91) H (137) 91B (196) H (137) 6
N (430) 190C (204) U (2,294) 204D (247) 0 (450) 100
Q (637) 147E (275) B (33) 33
C (53) 53J (279) 49U (2,294) 140
F (298) U (2,294) 298G (302) D (77) 62
N (430) 240H (357) U (2,294) 3571 (396) F (127) 63
L (301) 301P (497) 32
J (615) D (77) 15R (682) 600
K (718) Q (637) 490T (1,118) 228
L (780) A (11) 11F (127) 64M (355) 3550 (450) 350
M (955) U (2,294) 955N (948) E (105) 105
H (137) 401 (162) 162K (301) 301U (2,294) 340
0 (1,100) G (128) 128R (682) 82T (1,118) 890
P (1,750) J (279) 230P (497) 465S (1,055) 1,055
a The AseI and SpeI restriction fragments have been assigned alphabeticalnames as described by Chen et al. (4). Numbers refer to the sizes of therestriction fragments in kilobase pairs.
b Numbers refer to the sizes of the restriction products in kilobase pairs.The AseI-SpeI fragment listed in a given line is derived from the SpeI fragmentpreceding it in the same line. For example, the 6-kbp AseI-SpeI fragmnent withhomology to AseI fragment B is derived from SpeI fragment H rather than N.
were obtained. Many YAC clones, such as 486, 557, 617, and649, did not hybridize with the probe, while other YACclones, such as 502, 504, and 588, showed strong hybridiza-tion. A few YAC clones, such as 535, hybridized weaklywith the probe, suggesting that they contain only partialhomology with fragment K. Clone 535 contains an AseIrestriction site and is therefore a linking clone; it contains alarge amount of DNA that is derived from the restrictionfragment that joins K and would be expected to hybridizewith that fragment. AseI fragment D hybridized with YACclone 535 as well as SpeI restriction fragment Q (Table 1).These results suggest that AseI fragments D and K are joinedto each other in the intact genome.From the linking fragment data, it was possible to deter-
mine the order of all of the AseI restriction fragments exceptfor C, H, F, and M, which are internal to the 2,294-kbp SpeIrestriction fragment U (Table 1). The order of C, H, F, andM was unambiguously determined by identification of link-
J. BACTERIOL.
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
PHYSICAL MAP OF M. XANTHUS 2111
A B1 2 3 4 1 2 3 4
1100 -oM+N
780 L
K
615
396 IH
F+G
9 __B+C
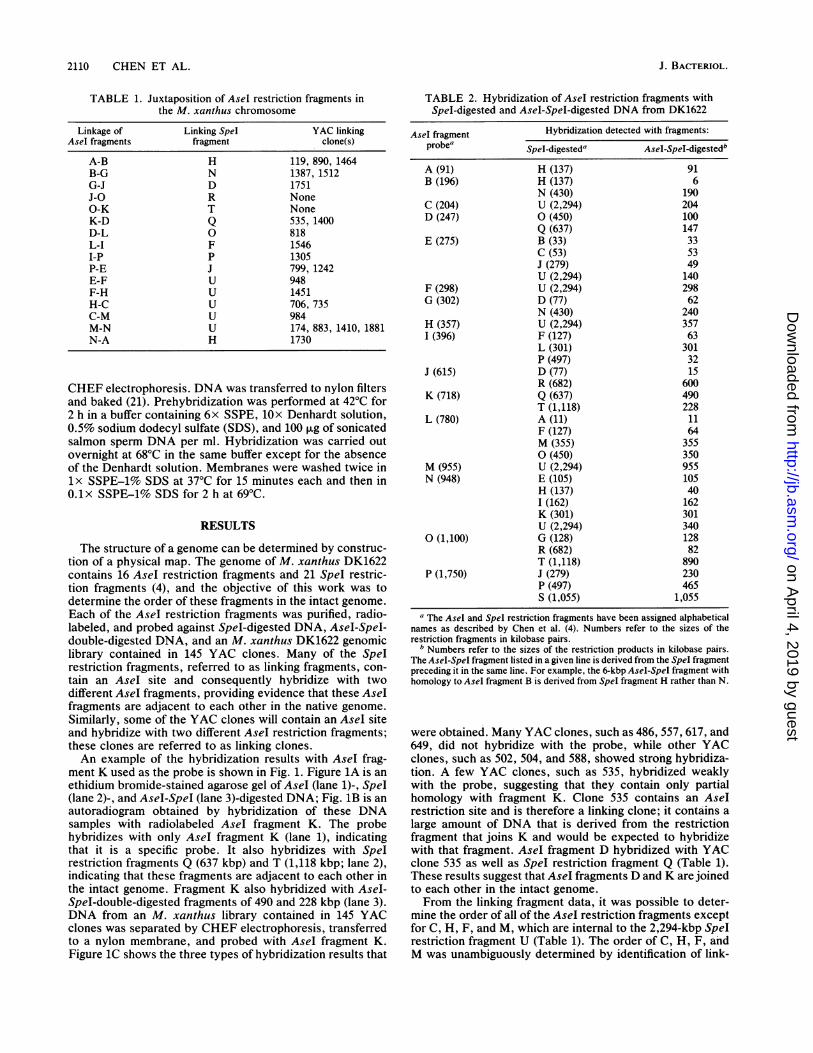
FIG. 2. AseI and SpeI restriction map of M. xanthus DK1622.Numbers refer to length in kilobase pairs. Letters refer to the namesof the restriction fragments as assigned by Chen et al. (4).
ing clones. Conversely, the YAC linking clone data sug-
gested an order for all but fragment 0, for which no linkingclones were identified. The linking fragment data clearlyidentify the neighbors of 0. Where overlapping data sets areavailable, the AseI fragment linkages determined from YAClinking clone data are in complete agreement with the linkingfragment results. The only possible interpretation of theseexperiments is that all of the Asel fragments are containedon a single, circular DNA molecule.
5000
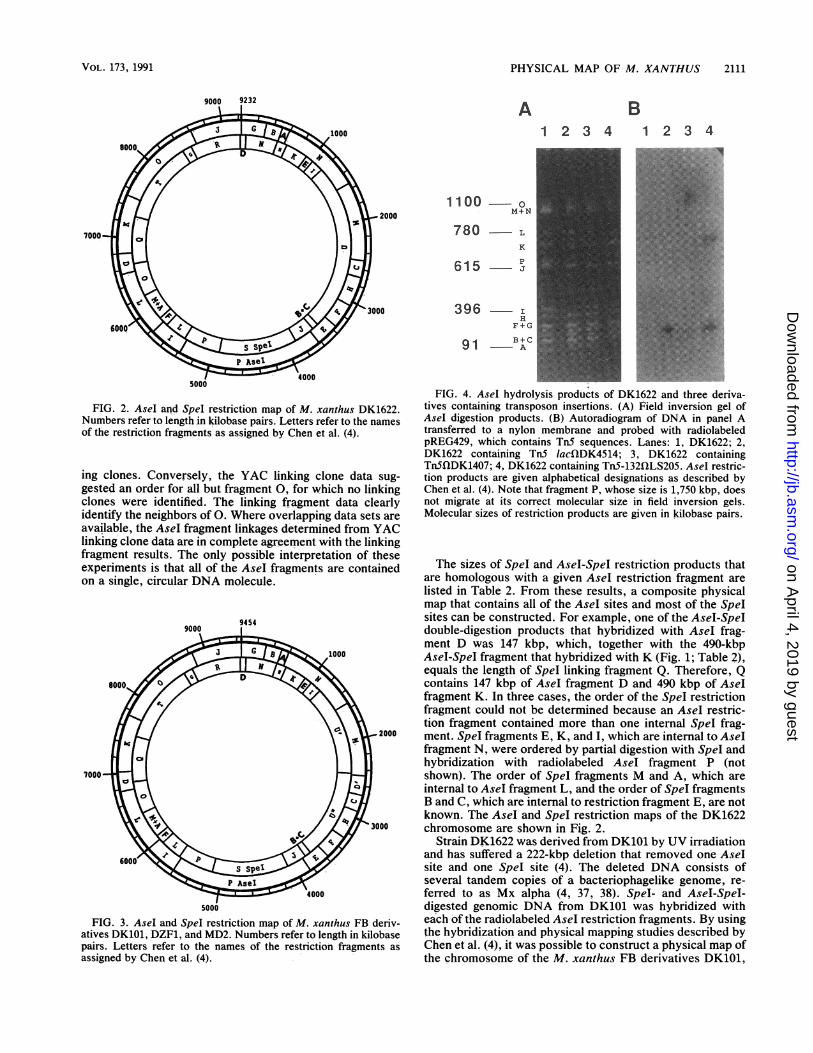
FIG. 3. AseI and SpeI restriction map of M. xanthus FB deriv-atives DK101, DZF1, and MD2. Numbers refer to length in kilobasepairs. Letters refer to the names of the restriction fragments asassigned by Chen et al. (4).
FIG. 4. AseI hydrolysis products of DK1622 and three deriva-tives containing transposon insertions. (A) Field inversion gel ofAseI digestion products. (B) Autoradiogram of DNA in panel Atransferred to a nylon membrane and probed with radiolabeledpREG429, which contains TnS sequences. Lanes: 1, DK1622; 2,DK1622 containing TnS lacQDK4514; 3, DK1622 containingTnSflDK1407; 4, DK1622 containing TnS-132fQLS205. AseI restric-tion products are given alphabetical designations as described byChen et al. (4). Note that fragment P, whose size is 1,750 kbp, doesnot migrate at its correct molecular size in field inversion gels.Molecular sizes of restriction products are given in kilobase pairs.
The sizes of SpeI and AseI-SpeI restriction products thatare homologous with a given AseI restriction fragment arelisted in Table 2. From these results, a composite physicalmap that contains all of the AseI sites and most of the SpeIsites can be constructed. For example, one of the AseI-Speldouble-digestion products that hybridized with AseI frag-ment D was 147 kbp, which, together with the 490-kbpAseI-SpeI fragment that hybridized with K (Fig. 1; Table 2),equals the length of SpeI linking fragment Q. Therefore, Qcontains 147 kbp of AseI fragment D and 490 kbp of AseIfragment K. In three cases, the order of the Spel restrictionfragment could not be determined because an AseI restric-tion fragment contained more than one internal SpeI frag-ment. SpeI fragments E, K, and I, which are internal to AseIfragment N, were ordered by partial digestion with SpeI andhybridization with radiolabeled AseI fragment P (notshown). The order of SpeI fragments M and A, which areinternal to AseI fragment L, and the order of SpeI fragmentsB and C, which are internal to restriction fragment E, are notknown. The AseI and SpeI restriction maps of the DK1622chromosome are shown in Fig. 2.
Strain DK1622 was derived from DK101 by UV irradiationand has suffered a 222-kbp deletion that removed one AseIsite and one -SpeI site (4). The deleted DNA consists ofseveral tandem copies of a bacteriophagelike genome, re-ferred to as Mx alpha (4, 37, 38). SpeI- and AseI-SpeI-digested genomic DNA from DK101 was hybridized witheach of the radiolabeled AseI restriction fragments. By usingthe hybridization and physical mapping studies described byChen et al. (4), it was possible to construct a physical map ofthe chromosome of the M. xanthus FB derivatives DK101,
.2000
VOL. 173, 1991
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
2112 CHEN ET AL.
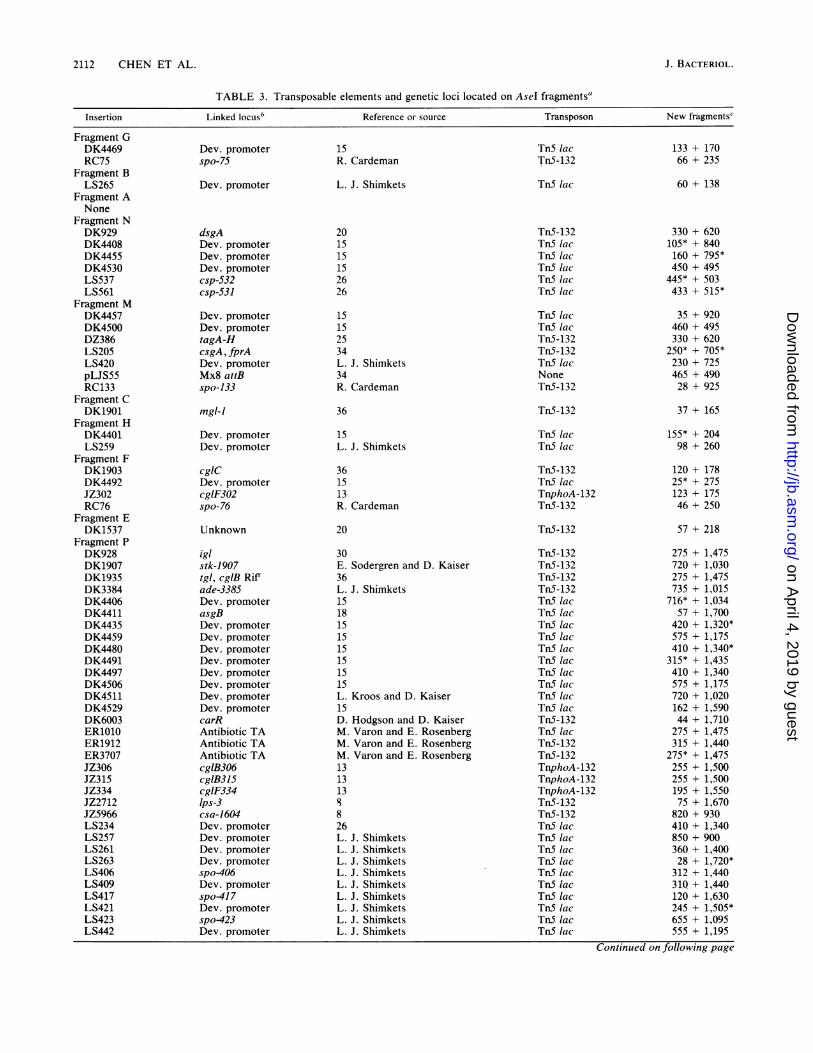
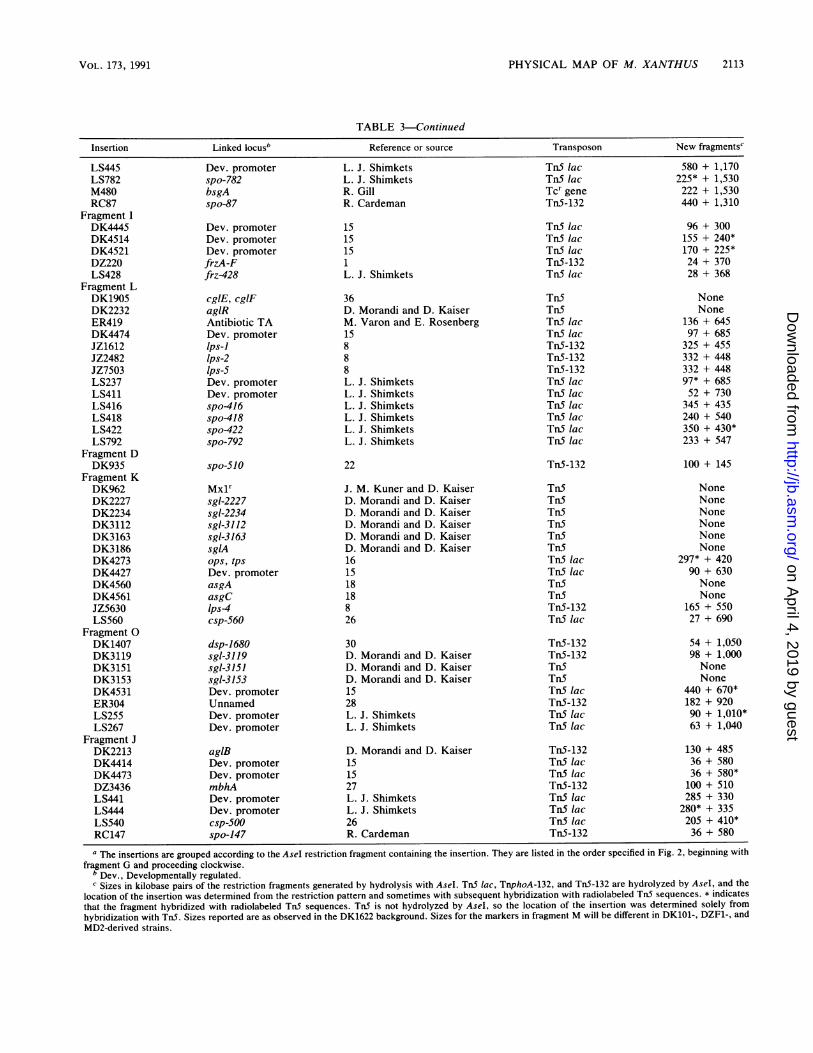
TABLE 3. Transposable elements and genetic loci located on Asel fragments'
Insertion Linked locusb Reference or source Transposon New fragments'
15R. Cardeman
L. J. Shimkets
201515152626
15152534L. J. Shimkets34R. Cardeman
36
15L. J. Shimkets
361513R. Cardeman
20
30E. Sodergren and D. Kaiser36L. J. Shimkets1518151515151515L. Kroos and D. Kaiser15D. Hodgson and D. KaiserM. Varon and E. RosenbergM. Varon and E. RosenbergM. Varon and E. Rosenberg131313s
826L. J. ShimketsL. J. ShimketsL. J. ShimketsL. J. ShimketsL. J. ShimketsL. J. ShimketsL. J. ShimketsL. J. ShimketsL. J. Shimkets
133 + 17066 + 235
Tn5 lacTn5-132
Tn5 lac
TnS-132TnS lacTn5 lacTnS lacTn5 lacTn5 lac
Tn5 lacTnS lacTnS-132TnS-132TnS lacNoneTnS-132
Tn5-132
Tn5 lacTnS lac
TnS-132TnS lacTnphoA-132Tn5-132
TnS-132
Tn5-132Tn5-132TnS-132TnS-132Tn5 lacTn5 lacTnS lacTn5 lacTn5 lacTn5 lacTn5 lacTn5 lacTn5 lacTn5 lacTn5-132TnS lacTn5-132Tn5-132TnphoA-132TnphoA-132TnphoA-132TnS-132Tn5-132TnS lacTn5 lacTnS lacTnS lacTn5 lacTnS lacTnS lacTn5 lacTn5 lacTnS lac
60 + 138
330 + 620105* + 840160 + 795*450 + 495
445* + 503433 + 515*
35 + 920460 + 495330 + 620
250* + 705*230 + 725465 + 49028 + 925
37 + 165
155* + 20498 + 260
120 + 17825* + 275123 + 17546 + 250
57 + 218
275 + 1,475720 + 1,030275 + 1,475735 + 1,015
716* + 1,03457 + 1,700
420 + 1,320*575 + 1,175410 + 1,340*315* + 1,435410 + 1,340575 + 1,175720 + 1,020162 + 1,59044 + 1,710
275 + 1,475315 + 1,440
275* + 1,475255 + 1,500255 + 1,500195 + 1,55075 + 1,670
820 + 930410 + 1,340850 + 900360 + 1,40028 + 1,720*
312 + 1,440310 + 1,440120 + 1,630245 + 1,505*655 + 1,095555 + 1,195
Continued on following page
Fragment GDK4469RC75
Fragment BLS265
Fragment ANone
Fragment NDK929DK4408DK4455DK4530LS537LS561
Fragment MDK4457DK4500DZ386LS205LS420pLJS55RC133
Fragment CDK1901
Fragment HDK4401LS259
Fragment FDK1903DK4492JZ302RC76
Fragment EDK1537
Fragment PDK928DK1907DK1935DK3384DK4406DK4411DK4435DK4459DK4480DK4491DK4497DK4506DK4511DK4529DK6003ER1010ER1912ER3707JZ306JZ315JZ334JZ2712JZ5966LS234LS257LS261LS263LS406LS409LS417LS421LS423LS442
Dev. promoterspo-75
Dev. promoter
dsgADev. promoterDev. promoterDev. promotercsp-532csp-531
Dev. promoterDev. promotertagA-HcsgA, fprADev. promoterMx8 attBspo-133
mgl-l
Dev. promoterDev. promoter
cglCDev. promotercgIF302spo-76
Unknown
iglstk-1907tgl, cglB Rif'ade-3385Dev. promoterasgBDev. promoterDev. promoterDev. promoterDev. promoterDev. promoterDev. promoterDev. promoterDev. promotercarRAntibiotic TAAntibiotic TAAntibiotic TAcglB306cglB315cglF334lps-3csa-1604Dev. promoterDev. promoterDev. promoterDev. promoterspo406Dev. promoterspo-417Dev. promoterspo423Dev. promoter
J. BACTERIOL.
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
PHYSICAL MAP OF M. XANTHUS 2113
TABLE 3-Continued
Insertion Linked locusb Reference or source Transposon New fragmentsc
Dev. promoterspo-782bsgAspo-87
Dev. promoterDev. promoterDev. promoterfrzA-Ffrz-428
cglE, cglFagiRAntibiotic TADev. promoterlps-llps-2lps-5Dev. promoterDev. promoterspo416spo418spo422spo-792
spo-510
Mxlrsgl-2227sgl-2234sgl-3112sgl-3163sglAops, tpsDev. promoterasgAasgCIps4csp-560
dsp-1680sgl-3119sgl-3151sgl-3153Dev. promoterUnnamedDev. promoterDev. promoter
aglBDev. promoterDev. promotermbhADev. promoterDev. promotercsp-500spo-147
L. J. ShimketsL. J. ShimketsR. GillR. Cardeman
1515151L. J. Shimkets
36D. Morandi and D. KaiserM. Varon and E. Rosenberg15888L. J. ShimketsL. J. ShimketsL. J. ShimketsL. J. ShimketsL. J. ShimketsL. J. Shimkets
22
J. M. Kuner and D. KaiserD. Morandi and D. KaiserD. Morandi and D. KaiserD. Morandi and D. KaiserD. Morandi and D. KaiserD. Morandi and D. Kaiser16151818826
30D. Morandi and D. KaiserD. Morandi and D. KaiserD. Morandi and D. Kaiser1528L. J. ShimketsL. J. Shimkets
D. Morandi and D. Kaiser151527L. J. ShimketsL. J. Shimkets26R. Cardeman
a The insertions are grouped according to the AseI restriction fragment containing the insertion. They are listed in the order specified in Fig. 2, beginning with
fragment G and proceeding clockwise.b Dev., Developmentally regulated.' Sizes in kilobase pairs of the restriction fragments generated by hydrolysis with AseI. TnS lac, TnphoA-132, and TnS-132 are hydrolyzed by AseI, and the
location of the insertion was determined from the restriction pattern and sometimes with subsequent hybridization with radiolabeled TnS sequences. * indicates
that the fragment hybridized with radiolabeled TnS sequences. TnS is not hydrolyzed by AseI, so the location of the insertion was determined solely from
hybridization with TnS. Sizes reported are as observed in the DK1622 background. Sizes for the markers in fragment M will be different in DK101-, DZF1-, and
MD2-derived strains.
Tn5 lacTn5 lacTcr geneTn5-132
580 + 1,170225* + 1,530222 + 1,530440 + 1,310
LS445LS782M480RC87
Fragment IDK4445DK4514DK4521DZ220LS428
Fragment LDK1905DK2232ER419DK4474JZ1612JZ2482JZ7503LS237LS411LS416LS418LS422LS792
Fragment DDK935
Fragment KDK962DK2227DK2234DK3112DK3163DK3186DK4273DK4427DK4560DK4561JZ5630LS560
Fragment 0DK1407DK3119DK3151DK3153DK4531ER304LS255LS267
Fragment JDK2213DK4414DK4473DZ3436LS441LS444LS540RC147
Tn5 lacTnS lacTn5 lacTn5-132Tn5 lac
Tn5Tn5Tn5 lacTn5 lacTn5-132Tn5-132Tn5-132Tn5 lacTn5 lacTn5 lacTn5 lacTn5 lacTn5 lac
Tn5-132
Tn5Tn5Tn5Tn5Tn5Tn5Tn5 lacTn5 lacTnSTnSTn5-132Tn5 lac
Tn5-132Tn5-132Tn5Tn5TnS lacTn5-132TnS lacTn5 lac
Tn5-132TnS lacTn5 lacTn5-132TnS lacTnS lacTnS lacTn5-132
96 + 300155 + 240*170 + 225*24 + 37028 + 368
NoneNone
136 + 64597 + 685
325 + 455332 + 448332 + 44897* + 68552 + 730
345 + 435240 + 540350 + 430*233 + 547
100 + 145
NoneNoneNoneNoneNoneNone
297* + 42090 + 630NoneNone
165 + 55027 + 690
54 + 1,05098 + 1,000NoneNone
440 + 670*182 + 92090 + 1,010*63 + 1,040
130 + 48536 + 58036 + 580*100 + 510285 + 330
280* + 335205 + 410*36 + 580
VOL. 173, 1991
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

2114 CHEN ET AL.
DZF1, and MD2, which contain the Mx alpha region (Fig. 3).The restriction map is identical to that of DK1622 except forthe region in which the deletion occurred.
Location of genetic markers. Through the use of transpo-son tagging (17) and generalized transduction (2, 23), trans-poson insertions that are in or near specific developmentalgenes have been discovered. Maps of genes that are sepa-rated by less than 60 kbp are available from generalizedtransduction studies (12). Unfortunately, the locations ofthese genes on the chromosome have not been determinedsince long-range mapping techniques were unavailable. It isnow possible to determine which AseI fragment contains atransposon by using pulsed field gel electrophoresis andDNA hybridization. Since Tn5-132 (11) and Tn.5 lac (14)contain AseI restriction sites, the AseI fragment harboringthe transposon may be identified by pulsed field gel electro-phoresis of Asel-digested DNA (4). Tn5 (11) does notcontain an Asel site, but the AseI fragment containing Tn5can be identified by hybridization with radiolabeled TnSsequences.An example of this approach is shown in Fig. 4. Figure 4A
shows Asel-digested genomic DNA from DK1622 and threerelated strains which was subjected to field inversion gelelectrophoresis. The alphabetical designations refer to thenames of the AseI restriction fragments as assigned by Chenet al. (4). The autoradiogram in Fig. 4B shows the results ofhybridization of radiolabeled plasmid pREG429 with theDNA observed in Fig. 4A. This plasmid contains the 2.5-kbpHpaI-SalI fragment from Tn5 which includes most of IS5OLas well as the gene encoding the neomycin phosphotransfer-ase (9). Lane 1 contains DK1622 DNA which does notcontain Tn5 and does not hybridize with pREG429. Lane 2contains DNA from a derivative of DK1622 containing asingle Tn5 lac insertion, fQDK4514. The 396-kbp AseI re-striction fragment I was hydrolyzed into restriction frag-ments of about 160 and 240 kbp. A 4-kbp AseI restrictionproduct internal to Tn5 lac was also produced but was toosmall to be seen on this gel. The 240-kbp restriction fragmenthybridizes with TnS sequences (Fig. 4B), but because anAseI site is located near one end of Tn5 lac, the 160-kbprestriction fragment has no homology with the probe andhybridization was not observed. Lane 3 contains DNA froma strain with Tn5 insertion fQDK1407. Since TnS does notcontain an Asel restriction site and is too small to signifi-cantly change the mobility of a large AseI fragment, thelocation of the transposon could not be determined fromexamination of an ethidium bromide-stained gel. However,hybridization with radiolabeled TnS sequences revealed thatthis transposon was located in Asel fragment 0. Lane 4contains DNA from a strain carrying TnS-132 insertionQILS205, which contains an AseI restriction fragment nearthe center of the transposon. Restriction fragment M washydrolyzed into 240- and 710-kbp fragments, both of whichhybridized with the Tn5 probe.
Using this approach, we determined the locations of over100 transposons that had been shown by generalized trans-duction to be in or adjacent to specific developmental genes(Table 3). The fact that few transposons map to the samelocation argues that many more genes for development andmotility remain to be identified and mapped. Clustering oflarge numbers of genes of similar function was observed onlyin the case of the sgl and dsp genes for social motility whichare located in the adjacent Asel fragments K and 0.
DISCUSSION
The genome of M. xanthus is one of the largest bacterialgenomes known (4, 33). Our results indicate that the DNA isarranged in a single, circular chromosome. The reasons forthe large size of the chromosome are not clear. Excludinggenes that are essential for cell viability, less than 1% of thegenome is absolutely essential for development under labo-ratory conditions (16). However, there may be an obviousselective pressure for the maintenance of the large genomesize. Since genes involved in the social and developmentalbiology of this organism are found in most regions of thechromosome, periodic selection for developmental compe-tence under conditions of nutritional stress may purge thepopulation of variants suffering large deletions.Transposable elements have been used extensively in the
genetic analysis of M. xanthus, and more than 100 differentinsertions were available for this study (12). These trans-posons provide an ideal venue for merging the geneticinformation with the physical map. The assignment of genesto specific Asel fragments provides an important first step inthe construction of a fine-structure genetic map. In thisstudy, it was not possible to distinguish between the twopossible locations of a transposon. For example, whilefQDK4469 is clearly in AseI fragment G, we cannot tellwhether it is 130 kbp from the B-G junction or the G-Jjunction (Table 3). However, recent advances in this tech-nology will make mapping much simpler and straightforwardin the future. An EcoRI physical map has been constructedfor each of the YAC clones (19). We are currently aligningthe YAC clones with the Asel-Spel physical map, and thecomposite physical map will have much more detail sinceEcoRI sites are relatively common. In the future, one will beable to map a cloned gene by hybridization with this YACclone library. More fine-structure placement will be possibleif one can then determine either the size of the EcoRIfragment containing the gene or the distance of the gene froma defined marker such as an AseI site. In the future, it maynot even be necessary to clone a gene in order to map it. Ifthe gene is near a transposable element, an inverse polymer-ase chain reaction may be used to generate homologousprobes that can be used for hybridization with the YAClibrary. These advances will increase the simplicity andaccuracy of gene mapping and make this technology avail-able to those who do not have access to pulsed field gelelectrophoresis. This should help streamline genetic analysisof M. xanthus.
ACKNOWLEDGMENTS
This work was supported by grants DCB8710705 andDCB9001755 from the National Science Foundation and a grantfrom the Georgia Power Co.We gratefully acknowledge the individuals who contributed M.
xanthus strains for this study: Richard Cardeman, Yvonne Cheng,Ronald Gill, David Hodgson, Dale Kaiser, Michael Kalos, LeeKroos, Mark McBride, Kathleen O'Connor, Jorge Rodriguez, Jo-seph Romeo, Eugene Rosenberg, Amy Rosenbluh, Mazal Varon,Robin Weinberg, James Zissler, and David Zusman. We also thankMary Case and Norman Giles for the use of their CHEF electro-phoresis unit, Bert Ely for help with the computer graphics, andJames Dana for excellent technical assistance.
REFERENCES1. Blackhart, B. D., and D. R. Zusman. 1985. Cloning and com-
plementation analysis of the frizzy genes of Myxococcus xan-thus. Mol. Gen. Genet. 198:243-254.
2. Campos, J. M., J. Geisselsoder, and D. R. Zusman. 1978.
J. BACTERIOL.
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

PHYSICAL MAP OF M. XANTHUS 2115
Isolation of bacteriophage Mx4, a generalized transducingphage for Myxococcus xanthus. J. Mol. Biol. 119:167-178.
3. Carle, G. F., M. Frank, and M. V. Olson. 1986. Electrophoreticseparations of large DNA molecules by periodic inversion of theelectric field. Science 232:65-68.
4. Chen, H.-W., I. M. Keseler, and L. J. Shimkets. 1990. Genomesize of Myxococcus xanthus determined by pulsed-field gelelectrophoresis. J. Bacteriol. 172:4206-4213.
5. Chu, G., D. Vollrath, and R. W. Davis. 1986. Separation of largeDNA molecules by contour-clamped homogeneous electricfields. Science 234:1582-1585.
6. Dworkin, M. 1962. Nutritional requirements for the vegetativegrowth of Myxococcus xanthus. J. Bacteriol. 84:250-257.
7. Feinberg, A. P., and B. Vogelstein. 1983. A technique forradiolabelling DNA restriction endonuclease fragments to highspecific activity. Anal. Biochem. 132:6-13.
8. Fink, J. M., M. Kalos, and J. F. Zissler. 1989. Isolation of cellsurface antigen mutants of Myxococcus xanthus by use ofmonoclonal antibodies. J. Bacteriol. 171:2033-2041.
9. Gill, R. E., M. G. Cull, and S. Fly. 1988. Genetic identificationand cloning of a gene required for developmental cell interac-tions in Myxococcus xanthus. J. Bacteriol. 170:5279-5288.
10. Hodgkin, J., and D. Kaiser. 1977. Cell-to-cell stimulation ofmovement in nonmotile mutants of Myxococcus xanthus. Proc.Natl. Acad. Sci. USA 74:2938-2942.
11. Jorgensen, R. A., S. J. Rothstein, and W. S. Reznikoff. 1979. Arestriction enzyme cleavage map of TnS and location of a regionencoding neomycin resistance. Mol. Gen. Genet. 177:65-72.
12. Kaiser, D. 1984. Genetics of myxobacteria, p. 163-184. In E.Rosenberg (ed.), Myxobacteria: development and cell interac-tions. Springer-Verlag, New York.
13. Kalos, M., and J. Zissler. 1990. Defects in contact-stimulatedgliding during aggregation by Myxococcus xanthus. J. Bacteriol.172:6476-6493.
14. Kroos, L., and D. Kaiser. 1984. Construction of TnS lac, atransposon that fuses lacZ expression to exogenous promoters,and its introduction into Myxococcus xanthus. Proc. Natl.Acad. Sci. USA 81:5816-5820.
15. Kroos, L., and D. Kaiser. 1987. Expression of many develop-mentally regulated genes in Myxococcus depends on a sequenceof cell interactions. Genes Dev. 1:840-854.
16. Kroos, L., A. Kuspa, and D. Kaiser. 1986. A global analysis ofdevelopmentally regulated genes in Myxococcus xanthus. Dev.Biol. 117:252-266.
17. Kuner, J. M., and D. Kaiser. 1981. Introduction of transposonTnS into Myxococcus for analysis of developmental and othernonselectable mutants. Proc. Natl. Acad. Sci. USA 78:425-429.
18. Kuspa, A., and D. Kaiser. 1989. Genes required for develop-mental signaling in Myxococcus xanthus: three asg loci. J.Bacteriol. 171:2762-2772.
19. Kuspa, A., D. Vollrath, Y. Cheng, and D. Kaiser. 1989. Physicalmapping of the Myxococcus xanthus genome by random cloningin yeast artificial chromosomes. Proc. Natl. Acad. Sci. USA86:8917-8921.
20. LaRossa, R., J. Kuner, D. Hagen, C. Manoil, and D. Kaiser.1983. Developmental cell interaction of Myxococcus xanthus:analysis of mutants. J. Bacteriol. 153:1394-1404.
21. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular
cloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
22. Manoil, C., and D. Kaiser. 1980. Accumulation of guanosinetetraphosphate and guanosine pentaphosphate in Myxococcusxanthus during starvation and myxospore formation. J. Bacte-riol. 141:297-304.
23. Martin, S., E. Sodergren, T. Masuda, and D. Kaiser. 1978.Systematic isolation of transducing phages for Myxococcusxanthus. Virology 88:44-53.
23a.Morandi, D., and D. Kaiser. Unpublished data.24. Morrison, C. E., and D. R. Zusman. 1979. Myxococcus xanthus
mutants with temperature-sensitive, stage-specific defects: evi-dence for independent pathways in development. J. Bacteriol.140:1036-1042.
25. O'Connor, K. A., and D. R. Zusman. 1990. Genetic analysis ofthe tag mutants of Myxococcus xanthus provides evidence fortwo developmental aggregation systems. J. Bacteriol. 172:3868-3878.
26. Rhie, H.-G., and L. J. Shimkets. 1989. Developmental bypasssuppression of Myxococcus xanthus csgA mutations. J. Bacte-riol. 171:3268-3276.
27. Romeo, J. M., and D. R. Zusman. 1987. Cloning of the gene formyxobacterial hemagglutinin and isolation and analysis of struc-tural gene mutations. J. Bacteriol. 169:3801-3808.
28. Rosenbluh, A., and E. Rosenberg. 1990. Role of autocide AMI indevelopment of Myxococcus xanthus. J. Bacteriol. 172:4307-4314.
29. Schwartz, D. C., and C. R. Cantor. 1984. Separation of yeastchromosome-sized DNAs by pulsed field gradient gel electro-phoresis. Cell 37:67-75.
30. Shimkets, L. J. 1986. Role of cell cohesion in Myxococcusxanthus fruiting body formation. J. Bacteriol. 166:842-848.
31. Shimkets, L. J. 1987. Control of morphogenesis in myxobacte-ria. Crit. Rev. Microbiol. 14:195-227.
32. Shimkets, L. J. 1989. The role of the cell surface in the socialand adventurous behavior of myxobacteria. Mol. Microbiol.3:1295-1298.
33. Shimkets, L. J. 1990. Social and developmental biology of themyxobacteria. Microbiol. Rev. 54:473-501.
34. Shimkets, L. J., and S. J. Asher. 1988. Use of recombinationtechniques to examine the structure of the csg locus of Myxo-coccus xanthus. Mol. Gen. Genet. 211:63-71.
35. Shimkets, L. J., and D. Kaiser. 1982. Induction of coordinatedmovement of Myxococcus xanthus cells. J. Bacteriol. 152:451-461.
36. Sodergren, E., and D. Kaiser. 1983. Insertions of TnS near genesthat govern stimulatable cell motility in Myxococcus. J. Mol.Biol. 167:295-310.
37. Starich, T., P. Cordes, and J. Zissler. 1985. Transposon taggingto detect a latent virus in Myxococcus xanthus. Science 230:541-543.
38. Starich, T., and J. Zissler. 1989. Movement of multiple DNAunits between Myxococcus xanthus cells. J. Bacteriol. 171:2323-2336.
39. Wireman, J. W., and M. Dworkin. 1975. Morphogenesis anddevelopmental interactions in myxobacteria. Science 189:516-522.
VOL. 173, 1991
on April 4, 2019 by guest
http://jb.asm.org/
Dow
nloaded from