pathway analysis report - files.hsls.pitt.edu redirect page
TRANSCRIPT

Pathway Analysis Report
Gene Name
This report contains the pathway analysis results for the submitted sample 'Gene Name'. Analysis was performed against Reactome version 66 on 04/12/2018. The web link to these results is:
https://reactome.org/PathwayBrowser/#/ANALYSIS=MjAxODEyMDQwMjI0MjhfNDA4MA%3D%3D
Please keep in mind that analysis results are temporarily stored on our server. The storage period depends on usage of the service but is at least 7 days. As a result, please note that this URL is only
valid for a limited time period and it might have expired.

Table of Contents
Introduction1.
Properties2.
Genome-wide overview3.
Most significant pathways4.
Pathways details5.
Identifiers found6.
Identifiers not found7.

https://reactome.org Page 2
1. Introduction
Reactome is a curated database of pathways and reactions in human biology. Reactions can be con-sidered as pathway 'steps'. Reactome defines a 'reaction' as any event in biology that changes the state of a biological molecule. Binding, activation, translocation, degradation and classical bio-chemical events involving a catalyst are all reactions. Information in the database is authored by expert biologists, entered and maintained by Reactome’s team of curators and editorial staff. Re-actome content frequently cross-references other resources e.g. NCBI, Ensembl, UniProt, KEGG (Gene and Compound), ChEBI, PubMed and GO. Orthologous reactions inferred from annotation for Homo sapiens are available for 17 non-human species including mouse, rat, chicken, puffer fish, worm, fly, yeast, rice, and Arabidopsis. Pathways are represented by simple diagrams follow-ing an SBGN-like format.
Reactome's annotated data describe reactions possible if all annotated proteins and small mo-lecules were present and active simultaneously in a cell. By overlaying an experimental dataset on these annotations, a user can perform a pathway over-representation analysis. By overlaying quantitative expression data or time series, a user can visualize the extent of change in affected pathways and its progression. A binomial test is used to calculate the probability shown for each result, and the p-values are corrected for the multiple testing (Benjamini–Hochberg procedure) that arises from evaluating the submitted list of identifiers against every pathway.
To learn more about our Pathway Analysis, please have a look at our relevant publications:
Fabregat A, Sidiropoulos K, Garapati P, Gillespie M, Hausmann K, Haw R, … D’Eustachio P (2016). The reactome pathway knowledgebase. Nucleic Acids Research, 44(D1), D481–D487.
https://doi.org/10.1093/nar/gkv1351.
Fabregat A, Sidiropoulos K, Viteri G, Forner O, Marin-Garcia P, Arnau V, … Hermjakob H (2017). Reactome pathway analysis: a high-performance in-memory approach. BMC Bioinformatics, 18.

https://reactome.org Page 3
2. Properties
This is an expression analysis: The numbers are used to produce a scaled coloured overlay over Reactome pathway diagrams, as a means to visualize relative expression levels. Note that the numeric values do not have to be expression data, for instance by using gene association
scores the same analysis can be used to visualize genotyping results.
•
703 out of 1159 identifiers in the sample were found in Reactome, where 1337 pathways were hit by at least one of them.
•
All non-human identifiers have been converted to their human equivalent. •
This report is filtered to show only results for species 'Homo sapiens' and resource 'all re-sources'.
•
The unique ID for this analysis (token) is MjAxODEyMDQwMjI0MjhfNDA4MA%3D%3D. This ID is valid for at least 7 days in Reactome’s server. Use it to access Reactome services with your data.
•

https://reactome.org Page 4
3. Genome-wide overview
Cell CycleDNA Replication
Musclecontraction
Disease
Circadian ClockDNA Repair
Geneexpression (Transcription)
Organelle biogenesisand maintenance
Vesicle-mediatedtransport
DevelopmentalBiology
Mitophagy
SignalTransduction
Reproduction
Extracellularmatrix organization
Immune SystemMetabolism
of RNA
Cellular responsesto external stimuli
Proteinlocalization
Hemostasis
Neuronal System
ProgrammedCell Death
Digestionand absorption
Metabolism Transport ofsmall molecules
Chromatinorganization
Cell-Cellcommunication
Metabolismof proteins
1.4E2
-1.96E1
This figure shows a genome-wide overview of the results of your pathway analysis. Reactome path-ways are arranged in a hierarchy. The center of each of the circular "bursts" is the root of one top-
level pathway, for example "DNA Repair". Each step away from the center represents the next level lower in the pathway hierarchy. The color code denotes over-representation of that pathway in your input dataset. Light grey signifies pathways which are not significantly over-represented.
https://reactome.org Page 5
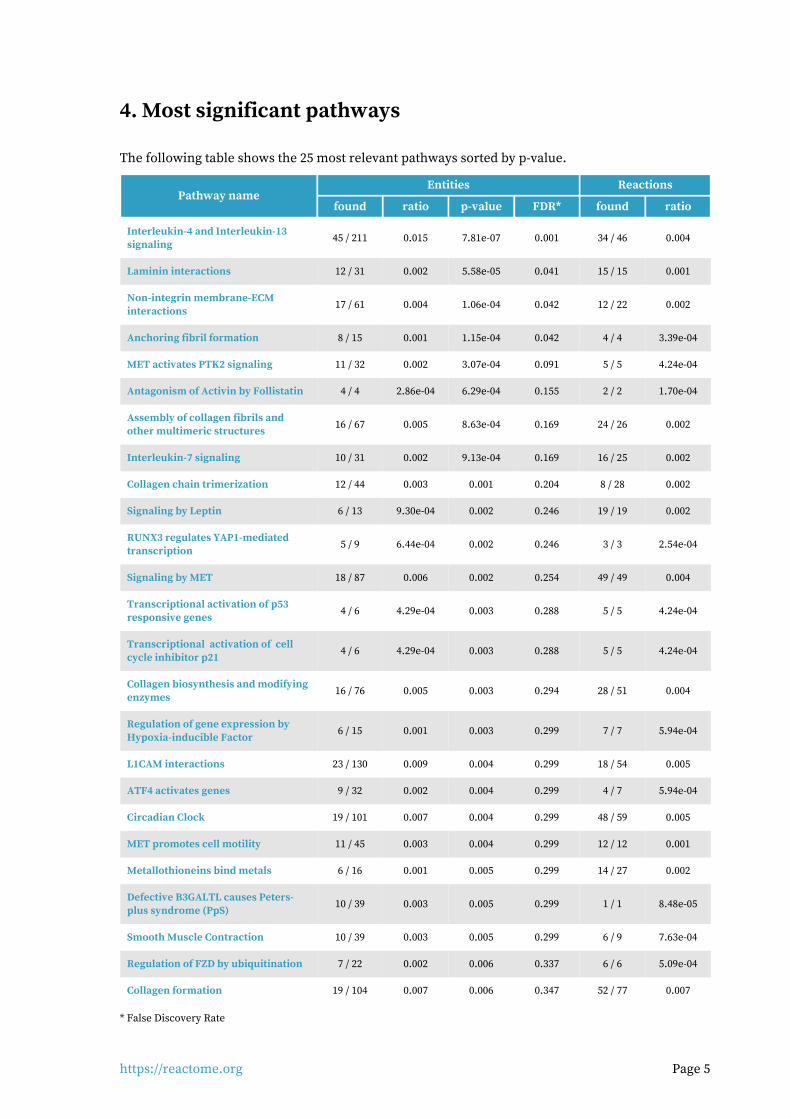
4. Most significant pathways
The following table shows the 25 most relevant pathways sorted by p-value.
Entities ReactionsPathway name
found ratio p-value FDR* found ratio
Interleukin-4 and Interleukin-13 signaling
45 / 211 0.015 7.81e-07 0.001 34 / 46 0.004
Laminin interactions 12 / 31 0.002 5.58e-05 0.041 15 / 15 0.001
Non-integrin membrane-ECM interactions
17 / 61 0.004 1.06e-04 0.042 12 / 22 0.002
Anchoring fibril formation 8 / 15 0.001 1.15e-04 0.042 4 / 4 3.39e-04
MET activates PTK2 signaling 11 / 32 0.002 3.07e-04 0.091 5 / 5 4.24e-04
Antagonism of Activin by Follistatin 4 / 4 2.86e-04 6.29e-04 0.155 2 / 2 1.70e-04
Assembly of collagen fibrils and other multimeric structures
16 / 67 0.005 8.63e-04 0.169 24 / 26 0.002
Interleukin-7 signaling 10 / 31 0.002 9.13e-04 0.169 16 / 25 0.002
Collagen chain trimerization 12 / 44 0.003 0.001 0.204 8 / 28 0.002
Signaling by Leptin 6 / 13 9.30e-04 0.002 0.246 19 / 19 0.002
RUNX3 regulates YAP1-mediated transcription
5 / 9 6.44e-04 0.002 0.246 3 / 3 2.54e-04
Signaling by MET 18 / 87 0.006 0.002 0.254 49 / 49 0.004
Transcriptional activation of p53 responsive genes
4 / 6 4.29e-04 0.003 0.288 5 / 5 4.24e-04
Transcriptional activation of cell cycle inhibitor p21
4 / 6 4.29e-04 0.003 0.288 5 / 5 4.24e-04
Collagen biosynthesis and modifying enzymes
16 / 76 0.005 0.003 0.294 28 / 51 0.004
Regulation of gene expression by Hypoxia-inducible Factor
6 / 15 0.001 0.003 0.299 7 / 7 5.94e-04
L1CAM interactions 23 / 130 0.009 0.004 0.299 18 / 54 0.005
ATF4 activates genes 9 / 32 0.002 0.004 0.299 4 / 7 5.94e-04
Circadian Clock 19 / 101 0.007 0.004 0.299 48 / 59 0.005
MET promotes cell motility 11 / 45 0.003 0.004 0.299 12 / 12 0.001
Metallothioneins bind metals 6 / 16 0.001 0.005 0.299 14 / 27 0.002
Defective B3GALTL causes Peters-plus syndrome (PpS)
10 / 39 0.003 0.005 0.299 1 / 1 8.48e-05
Smooth Muscle Contraction 10 / 39 0.003 0.005 0.299 6 / 9 7.63e-04
Regulation of FZD by ubiquitination 7 / 22 0.002 0.006 0.337 6 / 6 5.09e-04
Collagen formation 19 / 104 0.007 0.006 0.347 52 / 77 0.007
* False Discovery Rate

https://reactome.org Page 6
5. Pathways details
For every pathway of the most significant pathways, we present its diagram, as well as a short sum-mary, its bibliography and the list of inputs found in it.
Interleukin-4 and Interleukin-13 signaling (R-HSA-6785807)1.
cytosol
nucleoplasm
mitochondrial matrix
endoplasmic reticulum lumen
IL4R
JAK2
IL4R:JAK2IL4R:JAK2IL4R:JAK2 IL2RG
JAK3
IL2RG:JAK3
\\
STAT3-upregulated genesSTAT3-upregulated genesSTAT3-upregulated genesfor extracellular proteinsfor extracellular proteinsfor extracellular proteins
IL4:IL4R:JAK2IL4:IL4R:JAK2IL4:IL4R:JAK2
STAT3-upregulated extracellularSTAT3-upregulated extracellularSTAT3-upregulated extracellularproteinsproteinsproteins
IL4:IL4R:JAK2:IL2RG:JAK3IL4:IL4R:JAK2:IL2RG:JAK3IL4:IL4R:JAK2:IL2RG:JAK3
p-Y705-STAT3 dimerp-Y705-STAT3 dimerp-Y705-STAT3 dimer
\\
4 4
\\
ATP
IL13-bound tyrosine-IL13-bound tyrosine-IL13-bound tyrosine-phosphorylated IL13R type IIphosphorylated IL13R type IIphosphorylated IL13R type II
IL4
ADP
IL13RA1:TYK2
IL4:IL4R:JAK2:IL13RA1:TYK2IL4:IL4R:JAK2:IL13RA1:TYK2IL4:IL4R:JAK2:IL13RA1:TYK2
STAT1,STAT3,STAT6STAT1,STAT3,STAT6STAT1,STAT3,STAT6
TYK2
IL13-bound tyrosine-IL13-bound tyrosine-IL13-bound tyrosine-phosphorylated IL13 receptor typephosphorylated IL13 receptor typephosphorylated IL13 receptor type
II with STAT1,STAT3,STAT6II with STAT1,STAT3,STAT6II with STAT1,STAT3,STAT6
IL13RA1
3 3
JAK1
ATP
IL13-bound tyrosine-IL13-bound tyrosine-IL13-bound tyrosine-phosphorylated IL13 receptor typephosphorylated IL13 receptor typephosphorylated IL13 receptor type
II with phosphorylated STAT1,II with phosphorylated STAT1,II with phosphorylated STAT1,STAT3,STAT6STAT3,STAT6STAT3,STAT6
IL4:IL4R:JAK2:IL2RG:JAK3:IL4:IL4R:JAK2:IL2RG:JAK3:IL4:IL4R:JAK2:IL2RG:JAK3:JAK1JAK1JAK1
ADP
\\
3 3
ATP
p-Y701-STAT1,p-Y705-STAT3,p-p-Y701-STAT1,p-Y705-STAT3,p-p-Y701-STAT1,p-Y705-STAT3,p-Y641-STAT6Y641-STAT6Y641-STAT6
ADP
IL4:p-Y-IL4R:JAK2:p-Y-IL4:p-Y-IL4R:JAK2:p-Y-IL4:p-Y-IL4R:JAK2:p-Y-IL2RG:JAK3:p-Y-JAK1IL2RG:JAK3:p-Y-JAK1IL2RG:JAK3:p-Y-JAK1
\\ 2
\\
p-Y701-STAT1 dimer,p-Y705-STAT3p-Y701-STAT1 dimer,p-Y705-STAT3p-Y701-STAT1 dimer,p-Y705-STAT3dimer,p-Y641-STAT6 dimerdimer,p-Y641-STAT6 dimerdimer,p-Y641-STAT6 dimer
STAT3-upregulatedSTAT3-upregulatedSTAT3-upregulatedgenes for plasmagenes for plasmagenes for plasma
membrane proteinsmembrane proteinsmembrane proteins
\\
IL4:p-Y-IL4R:JAK2:p-Y-IL2RG:IL4:p-Y-IL4R:JAK2:p-Y-IL2RG:IL4:p-Y-IL4R:JAK2:p-Y-IL2RG:JAK3:p-Y-JAK1:STAT3,STAT6JAK3:p-Y-JAK1:STAT3,STAT6JAK3:p-Y-JAK1:STAT3,STAT6
IL4,IL13-upregulatedextracellular genes
STAT3-upregulatedSTAT3-upregulatedSTAT3-upregulatedplasma membraneplasma membraneplasma membrane
proteinsproteinsproteins
\\
IL4,IL13-upregulated extracellularproteins
ATP ADP
GATA3 gene
IL4, IL13
IL4:p-Y-IL4R:JAK2:p-Y-IL2RG:IL4:p-Y-IL4R:JAK2:p-Y-IL2RG:IL4:p-Y-IL4R:JAK2:p-Y-IL2RG:JAK3:p-Y-JAK1:p-Y705-JAK3:p-Y-JAK1:p-Y705-JAK3:p-Y-JAK1:p-Y705-
STAT3,p-Y641-STAT6STAT3,p-Y641-STAT6STAT3,p-Y641-STAT6
GATA3
\\
HSPA8 gene, ALOX15gene
HSPA8, ALOX15
\\
p-Y641-STAT6 dimer
\\
\\ \\
STAT3-upregulated genes forSTAT3-upregulated genes forSTAT3-upregulated genes forcytosolic proteinscytosolic proteinscytosolic proteins
p-Y701-STAT1 dimer,p-Y705-STAT3p-Y701-STAT1 dimer,p-Y705-STAT3p-Y701-STAT1 dimer,p-Y705-STAT3dimer,p-Y641-STAT6 dimerdimer,p-Y641-STAT6 dimerdimer,p-Y641-STAT6 dimer
STAT3-upregulated cytosolicSTAT3-upregulated cytosolicSTAT3-upregulated cytosolicproteinsproteinsproteins
STAT3,STAT6STAT3,STAT6STAT3,STAT6
\\
IL13-upregulated genes forIL13-upregulated genes forIL13-upregulated genes forplasma membrane proteinsplasma membrane proteinsplasma membrane proteins
IL13-upregulated proteinsIL13-upregulated proteinsIL13-upregulated proteins
\\
IL13-downregulated genes forIL13-downregulated genes forIL13-downregulated genes forextracellular proteinsextracellular proteinsextracellular proteins
p-Y705-STAT3 dimer, p-Y614-p-Y705-STAT3 dimer, p-Y614-p-Y705-STAT3 dimer, p-Y614-STAT6 dimerSTAT6 dimerSTAT6 dimer
IL13-downregulated proteinsIL13-downregulated proteinsIL13-downregulated proteins
\\
p-Y705-STAT3 dimer, p-Y641-STAT6p-Y705-STAT3 dimer, p-Y641-STAT6p-Y705-STAT3 dimer, p-Y641-STAT6dimerdimerdimer
HMOX1 gene
HMOX1
\\
MAOA gene
MAOA
STAT6 upregulated plasmamembrane proteins
STAT6 upregulatedplasma membrane protein
genes
SOCS5,(SOCS1)SOCS5,(SOCS1)SOCS5,(SOCS1)
IL4:IL4R:JAK2:IL13RA:TYK2:IL4:IL4R:JAK2:IL13RA:TYK2:IL4:IL4R:JAK2:IL13RA:TYK2:SOCS5,(SOCS1)SOCS5,(SOCS1)SOCS5,(SOCS1)
\\
IL4,IL13-downregulatedgenes for extracellular
proteins
\\
NDN gene, TP53gene
IL4,IL13-downregulatedextracellular proteins
NDN, TP53
\\
FASLG gene
FASLG(1-281)
\\ 2 p-Y705-STAT3,p-Y641-p-Y705-STAT3,p-Y641-p-Y705-STAT3,p-Y641-STAT6STAT6STAT6
\\
IL18 gene, ALOX5gene
IL18, ALOX5
IL13
\\
STAT3-upregulated genes forSTAT3-upregulated genes forSTAT3-upregulated genes fornuclear proteinsnuclear proteinsnuclear proteins
STAT3-upregulated nuclearSTAT3-upregulated nuclearSTAT3-upregulated nuclearproteinsproteinsproteins
?
CD36 gene
4xPalmC-CD36Pa
PaPa Pa
\\
BCL2 gene, BCL2L1 gene
Bcl-2/Bcl-X(L)
IL13
IL13RA2 IL13:IL13RA2
\\
STAT6 upregulatedextracellular protein genes
IL13:IL13RA:TYK2
STAT6 upregulated extracellularproteins
\\
HSP90B1
\\
PTGS2 gene
IL13:IL13RA:TYK2:IL4R:IL13:IL13RA:TYK2:IL4R:IL13:IL13RA:TYK2:IL4R:JAK2JAK2JAK2
PTGS2
IL13:IL13RA:TYK2:IL4R:IL13:IL13RA:TYK2:IL4R:IL13:IL13RA:TYK2:IL4R:JAK2:JAK1JAK2:JAK1JAK2:JAK1
SOCS1 gene
SOCS1
\\
1.4E2
-1.96E1
Interleukin-4 (IL4) is a principal regulatory cytokine during the immune response, crucially im-portant in allergy and asthma (Nelms et al. 1999). When resting T cells are antigen-activated and ex-pand in response to Interleukin-2 (IL2), they can differentiate as Type 1 (Th1) or Type 2 (Th2) T helper cells. The outcome is influenced by IL4. Th2 cells secrete IL4, which both stimulates Th2 in an autocrine fashion and acts as a potent B cell growth factor to promote humoral immunity (Nelms et al. 1999).

https://reactome.org Page 7
Interleukin-13 (IL13) is an immunoregulatory cytokine secreted predominantly by activated Th2 cells. It is a key mediator in the pathogenesis of allergic inflammation. IL13 shares many functional properties with IL4, stemming from the fact that they share a common receptor subunit. IL13 re-ceptors are expressed on human B cells, basophils, eosinophils, mast cells, endothelial cells, fibro-blasts, monocytes, macrophages, respiratory epithelial cells, and smooth muscle cells, but unlike IL4, not T cells. Thus IL13 does not appear to be important in the initial differentiation of CD4 T cells into Th2 cells, rather it is important in the effector phase of allergic inflammation (Hershey et al. 2003). IL4 and IL13 induce “alternative activation” of macrophages, inducing an anti-inflammatory phen-otype by signaling through IL4R alpha in a STAT6 dependent manner. This signaling plays an im-portant role in the Th2 response, mediating anti-parasitic effects and aiding wound healing (Go-rdon & Martinez 2010, Loke et al. 2002) There are two types of IL4 receptor complex (Andrews et al. 2006). Type I IL4R (IL4R1) is predomin-antly expressed on the surface of hematopoietic cells and consists of IL4R and IL2RG, the common gamma chain. Type II IL4R (IL4R2) is predominantly expressed on the surface of nonhematopoietic cells, it consists of IL4R and IL13RA1 and is also the type II receptor for IL13. (Obiri et al. 1995, Aman et al. 1996, Hilton et al. 1996, Miloux et al. 1997, Zhang et al. 1997). The second receptor for IL13 consists of IL4R and Interleukin-13 receptor alpha 2 (IL13RA2), sometimes called Interleukin-13 binding protein (IL13BP). It has a high affinity receptor for IL13 (Kd = 250 pmol/L) but is not suf-ficient to render cells responsive to IL13, even in the presence of IL4R (Donaldson et al. 1998). It is reported to exist in soluble form (Zhang et al. 1997) and when overexpressed reduces JAK-STAT sig-naling (Kawakami et al. 2001). It's function may be to prevent IL13 signalling via the functional IL4R:IL13RA1 receptor. IL13RA2 is overexpressed and enhances cell invasion in some human can-cers (Joshi & Puri 2012).
The first step in the formation of IL4R1 (IL4:IL4R:IL2RB) is the binding of IL4 with IL4R (Hoffman et al. 1995, Shen et al. 1996, Hage et al. 1999). This is also the first step in formation of IL4R2 (IL4:IL4R:IL13RA1). After the initial binding of IL4 and IL4R, IL2RB binds (LaPorte et al. 2008), to form IL4R1. Alternatively, IL13RA1 binds, forming IL4R2. In contrast, the type II IL13 complex (IL13R2) forms with IL13 first binding to IL13RA1 followed by recruitment of IL4R (Wang et al. 2009).
Crystal structures of the IL4:IL4R:IL2RG, IL4:IL4R:IL13RA1 and IL13:IL4R:IL13RA1 complexes have been determined (LaPorte et al. 2008). Consistent with these structures, in monocytes IL4R is tyr-osine phosphorylated in response to both IL4 and IL13 (Roy et al. 2002, Gordon & Martinez 2010) while IL13RA1 phosphorylation is induced only by IL13 (Roy et al. 2002, LaPorte et al. 2008) and IL2RG phosphorylation is induced only by IL4 (Roy et al. 2002).
Both IL4 receptor complexes signal through Jak/STAT cascades. IL4R is constitutively-associated with JAK2 (Roy et al. 2002) and associates with JAK1 following binding of IL4 (Yin et al. 1994) or IL13 (Roy et al. 2002). IL2RG constitutively associates with JAK3 (Boussiotis et al. 1994, Russell et al. 1994). IL13RA1 constitutively associates with TYK2 (Umeshita-Suyama et al. 2000, Roy et al. 2002, LaPorte et al. 2008, Bhattacharjee et al. 2013).
IL4 binding to IL4R1 leads to phosphorylation of JAK1 (but not JAK2) and STAT6 activation (Takeda et al. 1994, Ratthe et al. 2007, Bhattacharjee et al. 2013).
https://reactome.org Page 8
IL13 binding increases activating tyrosine-99 phosphorylation of IL13RA1 but not that of IL2RG. IL4 binding to IL2RG leads to its tyrosine phosphorylation (Roy et al. 2002). IL13 binding to IL4R2 leads to TYK2 and JAK2 (but not JAK1) phosphorylation (Roy & Cathcart 1998, Roy et al. 2002).
Phosphorylated TYK2 binds and phosphorylates STAT6 and possibly STAT1 (Bhattacharjee et al. 2013).
A second mechanism of signal transduction activated by IL4 and IL13 leads to the insulin receptor substrate (IRS) family (Kelly-Welch et al. 2003). IL4R1 associates with insulin receptor substrate 2 and activates the PI3K/Akt and Ras/MEK/Erk pathways involved in cell proliferation, survival and translational control. IL4R2 does not associate with insulin receptor substrate 2 and consequently the PI3K/Akt and Ras/MEK/Erk pathways are not activated (Busch-Dienstfertig & González-Rodríguez 2013).
References
Nelms K, Keegan AD, Zamorano J, Ryan JJ & Paul WE (1999). The IL-4 receptor: signaling mechan-
isms and biologic functions. Annu. Rev. Immunol., 17, 701-38.
Hershey GK (2003). IL-13 receptors and signaling pathways: an evolving web. J. Allergy Clin. Im-
munol., 111, 677-90; quiz 691.
Edit history
Date Action Author
2015-07-01 Authored Jupe S
2015-07-01 Created Jupe S
2016-09-02 Edited Jupe S
2016-09-02 Reviewed Leibovich SJ
2018-08-30 Modified Croft D
Entities found in this pathway (24)
Input UniProt Id #Fold cha...
IL6R P08887 1.55
PIM1 P11309 -1.80e+00
VCAM1 P19320 -1.21e+01
IL12A P29459 -4.10e+00
MYC P01106 1.8
MCL1 Q07820 1.5
MUC1 P15941 1.8
HGF P14210 -1.98e+00
JAK2 O60674 1.75
HPS5 P0DJI8 3
LIF P15018 -5.73e+00
CEBPD P49716 2.8
FOXO1 Q12778 4.91
TNFRSF1B P20333 1.77
CDKN1A P38936 -2.01e+00
FOXO3 O43524 2.41
PIK3R1 P27986 3.08

https://reactome.org Page 9
Input UniProt Id #Fold cha...
MAOA P21397 9.7
SOCS1 O15524 -2.28e+00
S1PR1 P21453 -2.85e+00
HMOX1 P09601 -2.54e+00
BCL6 P41182 1.7
STAT3 P40763 1.53
VEGFA P15692 -2.33e+00
Input Ensembl Id #Fold cha...
IL6R ENSG00000160712 1.55
PIM1 ENSG00000137193 -1.80e+00
VCAM1 ENSG00000162692 -1.21e+01
IL12A ENSG00000168811 -4.10e+00
MYC ENSG00000136997 1.8
MCL1 ENSG00000143384 1.5
MUC1 ENSG00000185499 1.8
HGF ENSG00000019991 -1.98e+00
LIF ENSG00000128342 -5.73e+00
CEBPD ENSG00000221869 2.8
FOXO1 ENSG00000150907 4.91
TNFRSF1B ENSG00000028137 1.77
CDKN1A ENSG00000124762 -2.01e+00
FOXO3 ENSG00000118689 2.41
PIK3R1 ENSG00000145675 3.08
MAOA ENSG00000189221 9.7
SOCS1 ENSG00000185338 -2.28e+00
S1PR1 ENSG00000170989 -2.85e+00
HMOX1 ENSG00000100292 -2.54e+00
BCL6 ENSG00000113916 1.7
VEGFA ENSG00000112715 -2.33e+00

https://reactome.org Page 10
Laminin interactions (R-HSA-3000157)2.
cytosol
IBSP:Collagen type I fibrilIBSP:Collagen type I fibrilIBSP:Collagen type I fibril
Degradation of the extracellularmatrixCollagen formation Elastic fibre formation
FN1 dimer
\\
10
Integrin alpha5beta1:Fibronectinmatrix
FN1(32-2386)
Integrin alphaVbeta3
Nidogens 1, 2Nidogens 1, 2Nidogens 1, 2
Laminins with gamma-1, gamma-3Laminins with gamma-1, gamma-3Laminins with gamma-1, gamma-3
DMP1
DMP1:Integrin alphVbeta3
Laminins with gamma-1,Laminins with gamma-1,Laminins with gamma-1,gamma-3:Nidogens 1,2gamma-3:Nidogens 1,2gamma-3:Nidogens 1,2
Laminins with gamma-1,Laminins with gamma-1,Laminins with gamma-1,gamma-3:Nidogens:HSPG2gamma-3:Nidogens:HSPG2gamma-3:Nidogens:HSPG2
DMD
Dystroglycan:Dystrophin:Dystroglycan:Dystrophin:Dystroglycan:Dystrophin:LamininsLamininsLaminins
Tenascin-C hexamer
Integrin alphaVbeta1
Laminins with gamma-1,Laminins with gamma-1,Laminins with gamma-1,gamma-3:Nidogens:Collagengamma-3:Nidogens:Collagengamma-3:Nidogens:Collagen
type IV networktype IV networktype IV network
DSPP(463-1301)
LamininsLamininsLaminins
DSPP(463-1301):IntegrinalphaVbeta1
Integrin alpha6beta1,Integrin alpha6beta1,Integrin alpha6beta1,alpha7beta1, alpha1beta1,alpha7beta1, alpha1beta1,alpha7beta1, alpha1beta1,alpha2beta1, alphaVbeta1alpha2beta1, alphaVbeta1alpha2beta1, alphaVbeta1
Dystroglycan
Integrin alpha6beta1,Integrin alpha6beta1,Integrin alpha6beta1,alpha7beta1, alpha1beta1,alpha7beta1, alpha1beta1,alpha7beta1, alpha1beta1,alpha2beta1, alphaVbeta1:alpha2beta1, alphaVbeta1:alpha2beta1, alphaVbeta1:
Laminin-111Laminin-111Laminin-111
AGRN, HSPG2
Dystroglycan:AGRN:HSPG2
Dystroglycan:NRXN1
Laminins withLaminins withLaminins withgamma-1gamma-1gamma-1
Laminin-111Laminin-111Laminin-111
Collagen type IV network:Collagen type IV network:Collagen type IV network:Laminin-1Laminin-1Laminin-1
AGRN(30-2045)
AGRN:Laminins with gamma-AGRN:Laminins with gamma-AGRN:Laminins with gamma-111
Laminin-332Laminin-332Laminin-332
NTN4
NTN4:Laminins with gamma-1,NTN4:Laminins with gamma-1,NTN4:Laminins with gamma-1,gamma-3gamma-3gamma-3
Collagen type VII fibrilSulfatide
DAG1(30-653)
Laminins:SulfatideLaminins:SulfatideLaminins:Sulfatide
Integrin alpha3beta1,alpha6beta4
AGRN:Alpha-dystroglycan
Integrin alpha3beta1,Integrin alpha3beta1,Integrin alpha3beta1,alpha6beta4:Laminins-332, 511,alpha6beta4:Laminins-332, 511,alpha6beta4:Laminins-332, 511,
521, (211, 221)521, (211, 221)521, (211, 221)
LRP4:MUSK
AGRN:LRP4:MUSK
Beta amyloid fibril
AGRN:Beta amyloid fibril
NCAM1, PTPRS
AGRN:NCAM1, PTPRS
DDR1 dimerCollagen type I, II, III, IV, V, XICollagen type I, II, III, IV, V, XICollagen type I, II, III, IV, V, XIfibrilsfibrilsfibrils
Collagen types I-V, VIICollagen types I-V, VIICollagen types I-V, VII
DDR1 dimer:Collagen type I, II, III, IV, V, XIDDR1 dimer:Collagen type I, II, III, IV, V, XIDDR1 dimer:Collagen type I, II, III, IV, V, XIfibrilsfibrilsfibrils
Integrin alpha7beta1:Integrin alpha7beta1:Integrin alpha7beta1:Laminin-211, 221, 411, 512,Laminin-211, 221, 411, 512,Laminin-211, 221, 411, 512,
521521521Integrin alpha2beta1Integrin alpha2beta1Integrin alpha2beta1
Collagen type IVCollagen type IVCollagen type IVnetworksnetworksnetworks
Integrin alpha2beta1:Laminin-332Integrin alpha2beta1:Laminin-332Integrin alpha2beta1:Laminin-332
DDR2 dimer
BGN:Collagen types I, VI, (IX)
Laminin-332Laminin-332Laminin-332
Collagen type I, II, III, V, Xfibrils
Hydroxylapatite
CEACAMheterodimer
Ca2+
NRXN1
Integrin cell surface interactions
Collagen type VII fibril:Collagen type VII fibril:Collagen type VII fibril:Laminin-332Laminin-332Laminin-332
SPARC Collagen type I fibril
Mn2+
Integrin alpha5beta1
DDR2 dimer:Collagen type I, II, III, V, Xfibrils
ADAM12,ADAM15,ADAM19ADAM12,ADAM15,ADAM19ADAM12,ADAM15,ADAM19
SH3PXD2A:PI(3,4)P2
VTN
SH3PXD2A:PI(3,4)P2:SH3PXD2A:PI(3,4)P2:SH3PXD2A:PI(3,4)P2:ADAM12,ADAM15,ADAM19ADAM12,ADAM15,ADAM19ADAM12,ADAM15,ADAM19
Collagen types II, III, VCollagen types II, III, VCollagen types II, III, V
Laminin-211, 221, 332, 411, 512, 521Laminin-211, 221, 332, 411, 512, 521Laminin-211, 221, 332, 411, 512, 521
Collagen type I fibril:SPARC:Collagen type I fibril:SPARC:Collagen type I fibril:SPARC:Hydroxylapatitie:Ca2+Hydroxylapatitie:Ca2+Hydroxylapatitie:Ca2+
VTN:Collagen types II,III,V
Collagen type I,IV,VICollagen type I,IV,VICollagen type I,IV,VI
Integrinalpha6beta1
Integrin alpha6beta1:Integrin alpha6beta1:Integrin alpha6beta1:Laminin-211, 221, 332, 411,Laminin-211, 221, 332, 411,Laminin-211, 221, 332, 411,
512, 521512, 521512, 521
2
VTN:Collagen type I,IV,VI
Integrins alphaVbeta1, alphaVbeta3, alphaVbeta5,alpha2bbeta3
FN1(32-2386):Collagen types I-V, VIIFN1(32-2386):Collagen types I-V, VIIFN1(32-2386):Collagen types I-V, VII
VTN:Integrins alphaVbeta1, alphaVbeta3,alphaVbeta5, alpha2bbeta3
SERPINE1
Vitronectin:Plasminogenactivator inhibitor 1
Collagen type I, II, III,VI fibrils
Integrinalpha7beta1
Laminin-211, 221, 411, 512, 521Laminin-211, 221, 411, 512, 521Laminin-211, 221, 411, 512, 521
DCN
DCN:Collagen type I, II, III, VIfibrils
Integrin alpha5beta1:FN1 dimer
Collagen types II, IIICollagen types II, IIICollagen types II, III
COMP pentamer
COMP interactors
COMP pentamer:COMPinteractors
BGN
Laminin-111Laminin-111Laminin-111
BGN:Collagen types II, IIIBGN:Collagen types II, IIIBGN:Collagen types II, III
Laminins with alpha-1, -2 or -5Laminins with alpha-1, -2 or -5Laminins with alpha-1, -2 or -5
Collagen types I, VI, (IX)
Syndecaninteractions
TGF betaTGF betaTGF beta
Tenascins C, R, (X, N)
SLRPs
Lecticans
SLRPs:TGF betaSLRPs:TGF betaSLRPs:TGF beta
Tenascins C, R, (X, N):Lecticans
Fibronectin matrix
Laminins with alpha-1, -2 or -5:HSPG2(22-Laminins with alpha-1, -2 or -5:HSPG2(22-Laminins with alpha-1, -2 or -5:HSPG2(22-4391)4391)4391)
Tenascins C, R, (X, N):Fibronectinmatrix
Integrin alphaVbeta3,Integrin alphaVbeta3,Integrin alphaVbeta3,alphaVbeta6,alphaVbeta6,alphaVbeta6,alpha2beta1,alpha2beta1,alpha2beta1,alpha7beta1,alpha7beta1,alpha7beta1,alpha8beta1,alpha8beta1,alpha8beta1,alpha9beta1,alpha9beta1,alpha9beta1,alphaXbeta1alphaXbeta1alphaXbeta1
TNC:Integrin alphaVbeta3,TNC:Integrin alphaVbeta3,TNC:Integrin alphaVbeta3,alphaVbeta6, alpha2beta1,alphaVbeta6, alpha2beta1,alphaVbeta6, alpha2beta1,alpha7beta1, alpha8beta1,alpha7beta1, alpha8beta1,alpha7beta1, alpha8beta1,alpha9beta1, alphaXbeta1alpha9beta1, alphaXbeta1alpha9beta1, alphaXbeta1
HSPG2(22-4391)
HSPG2:Dystroglycan
Collagen type IV networks:Collagen type VII fibrilCollagen type IV networks:Collagen type VII fibrilCollagen type IV networks:Collagen type VII fibril
FGF2(10-155), Fibronectn matrix,Transthyretin tetramer, PDGFA
homodimer, PDGFB homodimer
HSPG2:FGF2(10-155), Fibronectn matrix,Transthyretin tetramer, PDGFA
homodimer, PDGFB homodimer
HAPLN1
Aggrecan
HA
Aggrecan:HA:HAPLN1
Endostatin dimer
Laminin-111:EndostatinLaminin-111:EndostatinLaminin-111:Endostatindimerdimerdimer
LamininsLamininsLaminins Laminin network
Laminins-332, 511, 521,Laminins-332, 511, 521,Laminins-332, 511, 521,(211, 221)(211, 221)(211, 221)
IBSP
1.4E2
-1.96E1
Laminins are a large family of conserved, multidomain trimeric basement membrane proteins. There are many theoretical trimer combinations but only 18 have been described (Domogatskaya et al. 2012, Miner 2008, Macdonald et al. 2010) and the existence of isoforms laminin-212 and/or laminin-222 (Durbeej et al. 2010) awaits further confirmation. The chains assemble through coiled-coil domains at their C-terminal end. Alpha chains additionally have a large C-terminal globular do-main containing five LG subdomains (LG1-5). The N termini are often referred to as the short arms. These have varying numbers of laminin-type epidermal growth factor-like (LE) repeats. Trimer as-sembly is controlled by highly specific coiled-coil interactions (Domogatskaya et al. 2012). Some laminin isoforms are modified extracellularly by proteolytic processing at the N- or C-terminal ends prior to their binding to cellular receptors or other matrix molecules (Tzu & Marinkovitch 2008).
The cell adhesion properties of laminins are mediated primarily through the alpha chain G domain to integrins, dystroglycan, Lutheran glycoprotein, or sulfated glycolipids. The N-terminal globular domains of the alpha-1 (Colognato-Pyke et al. 1995) and alpha-2 chains (Colognato et al. 1997) and globular domains VI (Nielsen & Yamada 2001) and IVa (Sasaki & Timpl 2001) of the alpha-5 chain can bind to several integrin isoforms (alpha1beta1, alpha2beta1, alpha3beta1, and alphaVbeta3), which enables cell binding at both ends of laminins with these alpha chains.
References
Domogatskaya A, Rodin S & Tryggvason K (2012). Functional diversity of laminins. Annu. Rev. Cell
Dev. Biol., 28, 523-53.
Edit history
Date Action Author
2008-05-07 Reviewed Hynes R, Humphries MJ, Yamada KM
2012-08-08 Authored Jupe S
2013-01-24 Created Jupe S
2013-08-13 Edited Jupe S
https://reactome.org Page 11
Date Action Author
2013-08-13 Reviewed Ricard-Blum S
2018-08-23 Modified Schmidt EE
Entities found in this pathway (12)
Input UniProt Id #Fold cha...
LAMC1 P11047 1.73
LAMB1 P07942 1.69
COL4A5 P29400 -1.87e+00
COL4A3 Q01955 4.74
LAMA3 Q16787 1.82
COL4A4 P53420 3.93
COL7A1 Q02388 2.34
COL4A1 P02462 2.52
LAMA2 P24043 4.19
COL4A2 P08572 1.85
ITGA2 P17301 -2.06e+00
NID1 P14543 2.91

https://reactome.org Page 12
Non-integrin membrane-ECM interactions (R-HSA-3000171)3.
cytosol
IBSP:Collagen type I fibrilIBSP:Collagen type I fibrilIBSP:Collagen type I fibril
Degradation of the extracellularmatrixCollagen formation Elastic fibre formation
FN1 dimer
\\
10
Integrin alpha5beta1:Fibronectinmatrix
FN1(32-2386)
Integrin alphaVbeta3
Nidogens 1, 2Nidogens 1, 2Nidogens 1, 2
Laminins with gamma-1, gamma-3Laminins with gamma-1, gamma-3Laminins with gamma-1, gamma-3
DMP1
DMP1:Integrin alphVbeta3
Laminins with gamma-1,Laminins with gamma-1,Laminins with gamma-1,gamma-3:Nidogens 1,2gamma-3:Nidogens 1,2gamma-3:Nidogens 1,2
Laminins with gamma-1,Laminins with gamma-1,Laminins with gamma-1,gamma-3:Nidogens:HSPG2gamma-3:Nidogens:HSPG2gamma-3:Nidogens:HSPG2
DMD
Dystroglycan:Dystrophin:Dystroglycan:Dystrophin:Dystroglycan:Dystrophin:LamininsLamininsLaminins
Tenascin-C hexamer
Integrin alphaVbeta1
Laminins with gamma-1,Laminins with gamma-1,Laminins with gamma-1,gamma-3:Nidogens:Collagengamma-3:Nidogens:Collagengamma-3:Nidogens:Collagen
type IV networktype IV networktype IV network
DSPP(463-1301)
LamininsLamininsLaminins
DSPP(463-1301):IntegrinalphaVbeta1
Integrin alpha6beta1,Integrin alpha6beta1,Integrin alpha6beta1,alpha7beta1, alpha1beta1,alpha7beta1, alpha1beta1,alpha7beta1, alpha1beta1,alpha2beta1, alphaVbeta1alpha2beta1, alphaVbeta1alpha2beta1, alphaVbeta1
Dystroglycan
Integrin alpha6beta1,Integrin alpha6beta1,Integrin alpha6beta1,alpha7beta1, alpha1beta1,alpha7beta1, alpha1beta1,alpha7beta1, alpha1beta1,alpha2beta1, alphaVbeta1:alpha2beta1, alphaVbeta1:alpha2beta1, alphaVbeta1:
Laminin-111Laminin-111Laminin-111
AGRN, HSPG2
Dystroglycan:AGRN:HSPG2
Dystroglycan:NRXN1
Laminins withLaminins withLaminins withgamma-1gamma-1gamma-1
Laminin-111Laminin-111Laminin-111
Collagen type IV network:Collagen type IV network:Collagen type IV network:Laminin-1Laminin-1Laminin-1
AGRN(30-2045)
AGRN:Laminins with gamma-AGRN:Laminins with gamma-AGRN:Laminins with gamma-111
Laminin-332Laminin-332Laminin-332
NTN4
NTN4:Laminins with gamma-1,NTN4:Laminins with gamma-1,NTN4:Laminins with gamma-1,gamma-3gamma-3gamma-3
Collagen type VII fibrilSulfatide
DAG1(30-653)
Laminins:SulfatideLaminins:SulfatideLaminins:Sulfatide
Integrin alpha3beta1,alpha6beta4
AGRN:Alpha-dystroglycan
Integrin alpha3beta1,Integrin alpha3beta1,Integrin alpha3beta1,alpha6beta4:Laminins-332, 511,alpha6beta4:Laminins-332, 511,alpha6beta4:Laminins-332, 511,
521, (211, 221)521, (211, 221)521, (211, 221)
LRP4:MUSK
AGRN:LRP4:MUSK
Beta amyloid fibril
AGRN:Beta amyloid fibril
NCAM1, PTPRS
AGRN:NCAM1, PTPRS
DDR1 dimerCollagen type I, II, III, IV, V, XICollagen type I, II, III, IV, V, XICollagen type I, II, III, IV, V, XIfibrilsfibrilsfibrils
Collagen types I-V, VIICollagen types I-V, VIICollagen types I-V, VII
DDR1 dimer:Collagen type I, II, III, IV, V, XIDDR1 dimer:Collagen type I, II, III, IV, V, XIDDR1 dimer:Collagen type I, II, III, IV, V, XIfibrilsfibrilsfibrils
Integrin alpha7beta1:Integrin alpha7beta1:Integrin alpha7beta1:Laminin-211, 221, 411, 512,Laminin-211, 221, 411, 512,Laminin-211, 221, 411, 512,
521521521Integrin alpha2beta1Integrin alpha2beta1Integrin alpha2beta1
Collagen type IVCollagen type IVCollagen type IVnetworksnetworksnetworks
Integrin alpha2beta1:Laminin-332Integrin alpha2beta1:Laminin-332Integrin alpha2beta1:Laminin-332
DDR2 dimer
BGN:Collagen types I, VI, (IX)
Laminin-332Laminin-332Laminin-332
Collagen type I, II, III, V, Xfibrils
Hydroxylapatite
CEACAMheterodimer
Ca2+
NRXN1
Integrin cell surface interactions
Collagen type VII fibril:Collagen type VII fibril:Collagen type VII fibril:Laminin-332Laminin-332Laminin-332
SPARC Collagen type I fibril
Mn2+
Integrin alpha5beta1
DDR2 dimer:Collagen type I, II, III, V, Xfibrils
ADAM12,ADAM15,ADAM19ADAM12,ADAM15,ADAM19ADAM12,ADAM15,ADAM19
SH3PXD2A:PI(3,4)P2
VTN
SH3PXD2A:PI(3,4)P2:SH3PXD2A:PI(3,4)P2:SH3PXD2A:PI(3,4)P2:ADAM12,ADAM15,ADAM19ADAM12,ADAM15,ADAM19ADAM12,ADAM15,ADAM19
Collagen types II, III, VCollagen types II, III, VCollagen types II, III, V
Laminin-211, 221, 332, 411, 512, 521Laminin-211, 221, 332, 411, 512, 521Laminin-211, 221, 332, 411, 512, 521
Collagen type I fibril:SPARC:Collagen type I fibril:SPARC:Collagen type I fibril:SPARC:Hydroxylapatitie:Ca2+Hydroxylapatitie:Ca2+Hydroxylapatitie:Ca2+
VTN:Collagen types II,III,V
Collagen type I,IV,VICollagen type I,IV,VICollagen type I,IV,VI
Integrinalpha6beta1
Integrin alpha6beta1:Integrin alpha6beta1:Integrin alpha6beta1:Laminin-211, 221, 332, 411,Laminin-211, 221, 332, 411,Laminin-211, 221, 332, 411,
512, 521512, 521512, 521
2
VTN:Collagen type I,IV,VI
Integrins alphaVbeta1, alphaVbeta3, alphaVbeta5,alpha2bbeta3
FN1(32-2386):Collagen types I-V, VIIFN1(32-2386):Collagen types I-V, VIIFN1(32-2386):Collagen types I-V, VII
VTN:Integrins alphaVbeta1, alphaVbeta3,alphaVbeta5, alpha2bbeta3
SERPINE1
Vitronectin:Plasminogenactivator inhibitor 1
Collagen type I, II, III,VI fibrils
Integrinalpha7beta1
Laminin-211, 221, 411, 512, 521Laminin-211, 221, 411, 512, 521Laminin-211, 221, 411, 512, 521
DCN
DCN:Collagen type I, II, III, VIfibrils
Integrin alpha5beta1:FN1 dimer
Collagen types II, IIICollagen types II, IIICollagen types II, III
COMP pentamer
COMP interactors
COMP pentamer:COMPinteractors
BGN
Laminin-111Laminin-111Laminin-111
BGN:Collagen types II, IIIBGN:Collagen types II, IIIBGN:Collagen types II, III
Laminins with alpha-1, -2 or -5Laminins with alpha-1, -2 or -5Laminins with alpha-1, -2 or -5
Collagen types I, VI, (IX)
Syndecaninteractions
TGF betaTGF betaTGF beta
Tenascins C, R, (X, N)
SLRPs
Lecticans
SLRPs:TGF betaSLRPs:TGF betaSLRPs:TGF beta
Tenascins C, R, (X, N):Lecticans
Fibronectin matrix
Laminins with alpha-1, -2 or -5:HSPG2(22-Laminins with alpha-1, -2 or -5:HSPG2(22-Laminins with alpha-1, -2 or -5:HSPG2(22-4391)4391)4391)
Tenascins C, R, (X, N):Fibronectinmatrix
Integrin alphaVbeta3,Integrin alphaVbeta3,Integrin alphaVbeta3,alphaVbeta6,alphaVbeta6,alphaVbeta6,alpha2beta1,alpha2beta1,alpha2beta1,alpha7beta1,alpha7beta1,alpha7beta1,alpha8beta1,alpha8beta1,alpha8beta1,alpha9beta1,alpha9beta1,alpha9beta1,alphaXbeta1alphaXbeta1alphaXbeta1
TNC:Integrin alphaVbeta3,TNC:Integrin alphaVbeta3,TNC:Integrin alphaVbeta3,alphaVbeta6, alpha2beta1,alphaVbeta6, alpha2beta1,alphaVbeta6, alpha2beta1,alpha7beta1, alpha8beta1,alpha7beta1, alpha8beta1,alpha7beta1, alpha8beta1,alpha9beta1, alphaXbeta1alpha9beta1, alphaXbeta1alpha9beta1, alphaXbeta1
HSPG2(22-4391)
HSPG2:Dystroglycan
Collagen type IV networks:Collagen type VII fibrilCollagen type IV networks:Collagen type VII fibrilCollagen type IV networks:Collagen type VII fibril
FGF2(10-155), Fibronectn matrix,Transthyretin tetramer, PDGFA
homodimer, PDGFB homodimer
HSPG2:FGF2(10-155), Fibronectn matrix,Transthyretin tetramer, PDGFA
homodimer, PDGFB homodimer
HAPLN1
Aggrecan
HA
Aggrecan:HA:HAPLN1
Endostatin dimer
Laminin-111:EndostatinLaminin-111:EndostatinLaminin-111:Endostatindimerdimerdimer
LamininsLamininsLaminins Laminin network
Laminins-332, 511, 521,Laminins-332, 511, 521,Laminins-332, 511, 521,(211, 221)(211, 221)(211, 221)
IBSP
1.4E2
-1.96E1
Several non-integrin membrane proteins interact with extracellular matrix proteins. Transmem-brane proteoglycans may associate with integrins and growth factor receptors to influence their function, or they can signal independently, often influencing the actin cytoskeleton.
References
Rosso F, Giordano A, Barbarisi M & Barbarisi A (2004). From cell-ECM interactions to tissue engin-
eering. J. Cell. Physiol., 199, 174-80.
Couchman JR (2010). Transmembrane signaling proteoglycans. Annu. Rev. Cell Dev. Biol., 26, 89-
114.
Edit history
Date Action Author
2012-07-31 Authored Jupe S
2013-01-24 Created Jupe S
2013-04-26 Edited Jupe S
2013-05-22 Reviewed Ricard-Blum S
2018-08-23 Modified Schmidt EE
Entities found in this pathway (17)
Input UniProt Id #Fold cha...
LAMC1 P11047 1.73
SDC1 P18827 -1.80e+00
LAMB1 P07942 1.69
DMD P11532 1.97
COL4A5 P29400 -1.87e+00
COL4A3 Q01955 4.74
LAMA3 Q16787 1.82
https://reactome.org Page 13
Input UniProt Id #Fold cha...
COL4A4 P53420 3.93
COL5A3 P25940 1.9
COL4A1 P02462 2.52
COL11A1 P12107 5.14
LAMA2 P24043 4.19
COL4A2 P08572 1.85
ITGA2 P17301 -2.06e+00
COL1A1 P02452 -2.48e+00
COL3A1 P02461 -1.81e+00
THBS1 P07996 3.85

https://reactome.org Page 14
Anchoring fibril formation (R-HSA-2214320)4.
cytosol
Collagen fibrils
PXDN:Br-
Network formingtropocollagens
Collagen networks
2
Collagen alpha-1(VII)Collagen alpha-1(VII)Collagen alpha-1(VII)trimertrimertrimer
Collagen type VII hexamerCollagen type VII hexamerCollagen type VII hexamer
Collagen type I fibrils with histidino-hydroxylysinoleucine cross-links
2 Collagen type IV networks withCollagen type IV networks withCollagen type IV networks withsulfilimine cross-linkssulfilimine cross-linkssulfilimine cross-links
Collagen type XVII fibril:Integrin alpha6beta4
BPAG1e:PlectinBPAG1e:PlectinBPAG1e:Plectin
Collagen type VII fibril:Collagen type VII fibril:Collagen type VII fibril:Laminin-332Laminin-332Laminin-332
CD151 Type I hemidesmosomeType I hemidesmosomeType I hemidesmosomecomplexcomplexcomplex
Collagen type I fibrils with hydroxylysino-5-Collagen type I fibrils with hydroxylysino-5-Collagen type I fibrils with hydroxylysino-5-ketonorleucine crosslinksketonorleucine crosslinksketonorleucine crosslinks
Collagen type 1 fibrils cross-linked bydehydro-lysinonorleucine crosslinks
Endostatin releasingEndostatin releasingEndostatin releasingproteasesproteasesproteases
Collagen type XI fibril:Collagen type II fibril
Collagen alpha-1(VII) NC2 regionCollagen alpha-1(VII) NC2 regionCollagen alpha-1(VII) NC2 region
Tropocollagens
Collagen type I fibrils withCollagen type I fibrils withCollagen type I fibrils withhydroxylysyl-pyrrole cross-linkshydroxylysyl-pyrrole cross-linkshydroxylysyl-pyrrole cross-links
Collagen type VII -NC2Collagen type VII -NC2Collagen type VII -NC2hexamerhexamerhexamer
Collagen fibres
Laminin-332Laminin-332Laminin-332
Collagen type VII fibril
Collagen type IV networksCollagen type IV networksCollagen type IV networks
18
18
18
18
Collagen type XVIII
Collagen type XVCollagen type XVCollagen type XV
COL15A1(1212-1388)
COL18A1(1572-11754)
COL18A1(?-1571)
COL15A1(?-1211)
2
Collagen type I fibril
Collagen type IX
Anchoring fibril complexAnchoring fibril complexAnchoring fibril complex
Collagen type I fibrils with lysyl-pyridinolineCollagen type I fibrils with lysyl-pyridinolineCollagen type I fibrils with lysyl-pyridinolinecross-linkscross-linkscross-links
H2O2
Collagen type I fibril with freehydroxylysines
H2O
Collagen type I fibrils with deH-HLNL cross-links
6
Collagen type VII NC2proteinases
Prolysyl oxidases
Lysyl oxidasepropeptides
Lysyl oxidases
Procollagen C-Procollagen C-Procollagen C-proteinasesproteinasesproteinases
2
Collagen type XI fibril
O2
Collagen type I fibrils with hydroxylysyl-Collagen type I fibrils with hydroxylysyl-Collagen type I fibrils with hydroxylysyl-pyridinoline cross-linkspyridinoline cross-linkspyridinoline cross-links
H2O2
Collagen type II fibril
NH3
Lysyl oxidases:Cu2+
Collagen type I fibril withhydroxyallysines
Collagen type II fibril:Collagentype IX
H2O
Collagen type XII, XIV fibrils
Collagen type I fibril withallysines
Collagen type I fibrils with lysyl-pyrroleCollagen type I fibrils with lysyl-pyrroleCollagen type I fibrils with lysyl-pyrrolecross-linkscross-linkscross-links
Collagen type I, II fibrils
Collagen type I,II:XII,XIV fibrils
Collagen type I fibrils with lysino-5-Collagen type I fibrils with lysino-5-Collagen type I fibrils with lysino-5-ketonorleucine cross-linksketonorleucine cross-linksketonorleucine cross-links
Collagen type X network
Collagen type X:type II fibrils
1.4E2
-1.96E1
Cellular compartments: extracellular region.
Collagen VII forms anchoring fibrils, composed of antiparallel dimers that connect the dermis to the epidermis (Bruckner-Tuderman 2009, Has & Kern 2010). During fibrillogenesis, the nascent type VII procollagen molecules dimerize in an antiparallel manner. The C-propeptide is then re-moved by Bone morphogenetic protein 1 (Rattenholl et al. 2002) and the processed antiparallel di-mers laterally aggregate (Villone et al. 2008, Gordon & Hahn 2010).
References
Chung HJ & Uitto J (2010). Type VII collagen: the anchoring fibril protein at fault in dystrophic epi-
dermolysis bullosa. Dermatol Clin, 28, 93-105.
Edit history
Date Action Author
2012-04-30 Authored Jupe S
2012-04-30 Created Jupe S
2012-10-08 Reviewed Kalamajski S, Raleigh S
2012-11-12 Edited Jupe S
2012-11-19 Reviewed Ricard-Blum S
2018-08-23 Modified Schmidt EE
Entities found in this pathway (8)
Input UniProt Id #Fold cha...
COL4A5 P29400 -1.87e+00

https://reactome.org Page 15
Input UniProt Id #Fold cha...
COL4A3 Q01955 4.74
LAMA3 Q16787 1.82
COL4A4 P53420 3.93
COL7A1 Q02388 2.34
COL4A1 P02462 2.52
COL4A2 P08572 1.85
COL1A1 P02452 -2.48e+00

https://reactome.org Page 16
MET activates PTK2 signaling (R-HSA-8874081)5.
cytosol
nucleoplasm
recycling endosome membrane
ATP
HGF:p-4Y-MET dimer:p-Y317-SHC1-HGF:p-4Y-MET dimer:p-Y317-SHC1-HGF:p-4Y-MET dimer:p-Y317-SHC1-222
ADP
SPINT1,2SPINT1,2SPINT1,2
HGF:p-4Y-MET dimer:p-Y317-HGF:p-4Y-MET dimer:p-Y317-HGF:p-4Y-MET dimer:p-Y317-SHC1-2:GRB2:SOS1SHC1-2:GRB2:SOS1SHC1-2:GRB2:SOS1
SPINT1
CIN85:endophilinCIN85:endophilinCIN85:endophilin
HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):
CIN85:endophillinCIN85:endophillinCIN85:endophillin
pro-HGF
H2O
GAB1
HGF dimerHGF dimerHGF dimer
HGF:p-4Y-MET dimer:HGF:p-4Y-MET dimer:HGF:p-4Y-MET dimer:GAB1GAB1GAB1
HGFAC dimer
METHGF dimer:METHGF dimer:METHGF dimer:MET
GRB2-1 HGF:p-4Y-MET dimer:GRB2-1:HGF:p-4Y-MET dimer:GRB2-1:HGF:p-4Y-MET dimer:GRB2-1:GAB1GAB1GAB1
HGF:p-4Y-MET dimer:GAB1,HGF:p-4Y-MET dimer:GAB1,HGF:p-4Y-MET dimer:GAB1,HGF:p-4Y-MET dimer:GRB2:HGF:p-4Y-MET dimer:GRB2:HGF:p-4Y-MET dimer:GRB2:
GAB1GAB1GAB1
ATP
ADP
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1GRB2-1:p-5Y-GAB1GRB2-1:p-5Y-GAB1
STAT3
HGF:p-4Y-MET dimer:STAT3HGF:p-4Y-MET dimer:STAT3HGF:p-4Y-MET dimer:STAT3
ATP
ADP
p-Y705-STAT3P
2
p-Y705-STAT3 dimerp-Y705-STAT3 dimerp-Y705-STAT3 dimer
PIK3CA:PIK3R1PIK3CA:PIK3R1PIK3CA:PIK3R1
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:PI3KGRB2-1:p-5Y-GAB1:PI3KGRB2-1:p-5Y-GAB1:PI3K
\\
2
p-Y705-STAT3 dimerp-Y705-STAT3 dimerp-Y705-STAT3 dimer
HGF:METHGF:METHGF:METdimerdimerdimer
PI(4,5)P2
ATP
PI(3,4,5)P3
HGF:MonoUb-K,p-4Y-METHGF:MonoUb-K,p-4Y-METHGF:MonoUb-K,p-4Y-METdimer:GRB2:p-Y-CBL,(HGF:dimer:GRB2:p-Y-CBL,(HGF:dimer:GRB2:p-Y-CBL,(HGF:
MonoUb-K,p-Y1003,4Y-MonoUb-K,p-Y1003,4Y-MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):CIN85:METdimer:p-Y-CBL):CIN85:METdimer:p-Y-CBL):CIN85:endophillin,(MET:Ub-LRIG1)endophillin,(MET:Ub-LRIG1)endophillin,(MET:Ub-LRIG1)
ADP
EPS15:HGS:STAM
HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):
CIN85:endophillin:EPS15:HGS:STAM,(MET:Ub-LRIG1:CIN85:endophillin:EPS15:HGS:STAM,(MET:Ub-LRIG1:CIN85:endophillin:EPS15:HGS:STAM,(MET:Ub-LRIG1:EPS15:HGS:STAM)EPS15:HGS:STAM)EPS15:HGS:STAM)
PIP3 activates AKT signaling
TNS3HGF:p-4Y-MET dimer:TNS3HGF:p-4Y-MET dimer:TNS3HGF:p-4Y-MET dimer:TNS3
TNS4:ITGB1
HGF:p-4Y-MET dimer:TNS4:ITGB1HGF:p-4Y-MET dimer:TNS4:ITGB1HGF:p-4Y-MET dimer:TNS4:ITGB1
8 8
RAF/MAP kinasecascade
ATP
HGF:p-Y1234,Y1235,Y1349,Y1356-MET dimerHGF:p-Y1234,Y1235,Y1349,Y1356-MET dimerHGF:p-Y1234,Y1235,Y1349,Y1356-MET dimer
CRK,CRKL
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:CRK,CRKLGRB2-1:p-5Y-GAB1:CRK,CRKLGRB2-1:p-5Y-GAB1:CRK,CRKL
PTPN11
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:PTPN11p-5Y-GAB1:PTPN11p-5Y-GAB1:PTPN11
RAPGEF1HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:CRK,CRKL:RAPGEF1GRB2-1:p-5Y-GAB1:CRK,CRKL:RAPGEF1GRB2-1:p-5Y-GAB1:CRK,CRKL:RAPGEF1
2 2
ADP
H2O
CBL:GRB2,CBL
Pi
PTK2
HGF:p-4Y-MET dimer:PTK2HGF:p-4Y-MET dimer:PTK2HGF:p-4Y-MET dimer:PTK2
HGF:p-Y1234,Y1235,Y1356-METHGF:p-Y1234,Y1235,Y1356-METHGF:p-Y1234,Y1235,Y1356-METdimerdimerdimer
PTPRJ
HGF:p-4Y-MET dimer:GRB2:HGF:p-4Y-MET dimer:GRB2:HGF:p-4Y-MET dimer:GRB2:CBL,(HGF:p-Y1003,4Y-METCBL,(HGF:p-Y1003,4Y-METCBL,(HGF:p-Y1003,4Y-MET
dimer:CBL)dimer:CBL)dimer:CBL)
ATP
ADP
HGF:p-4Y-MET dimer:GRB2:p-Y-HGF:p-4Y-MET dimer:GRB2:p-Y-HGF:p-4Y-MET dimer:GRB2:p-Y-CBL,(HGF:p-Y1003,4Y-MET dimer:p-CBL,(HGF:p-Y1003,4Y-MET dimer:p-CBL,(HGF:p-Y1003,4Y-MET dimer:p-
Y-CBL)Y-CBL)Y-CBL)
ATP
HGF:p-4Y-MET dimer:p-Y397-PTK2HGF:p-4Y-MET dimer:p-Y397-PTK2HGF:p-4Y-MET dimer:p-Y397-PTK2
ADP
RAP1:GDP
HGF:MonoUb-K,p-4Y-MET dimer:HGF:MonoUb-K,p-4Y-MET dimer:HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,(HGF:MonoUb-K,p-GRB2:p-Y-CBL,(HGF:MonoUb-K,p-GRB2:p-Y-CBL,(HGF:MonoUb-K,p-
Y1003,4Y-MET dimer:p-Y-CBL)Y1003,4Y-MET dimer:p-Y-CBL)Y1003,4Y-MET dimer:p-Y-CBL)
RAP1:GTP
GTP
Integrin alpha2beta1,alpha3:beta1:(collagen,laminin,Integrin alpha2beta1,alpha3:beta1:(collagen,laminin,Integrin alpha2beta1,alpha3:beta1:(collagen,laminin,fibronectin)fibronectin)fibronectin)
GDP
MyrG-p-Y419-SRC
P
4 4
HGF:p-4Y-MET dimer:p-Y397-PTK2:HGF:p-4Y-MET dimer:p-Y397-PTK2:HGF:p-4Y-MET dimer:p-Y397-PTK2:MyrG-p-Y419-SRCMyrG-p-Y419-SRCMyrG-p-Y419-SRC
HGF:p-Y,1349,Y1356-MET dimerHGF:p-Y,1349,Y1356-MET dimerHGF:p-Y,1349,Y1356-MET dimer
PTPN1,PTPN2
HPN heterodimer
ATP
ADP
HGF:p-4Y-MET dimer:p-Y194,Y397-HGF:p-4Y-MET dimer:p-Y194,Y397-HGF:p-4Y-MET dimer:p-Y194,Y397-PTK2:MyrG-p-Y419-SRCPTK2:MyrG-p-Y419-SRCPTK2:MyrG-p-Y419-SRC
GRB2-1:SOS1
HGF:p-4Y-MET dimer:GRB2-1:HGF:p-4Y-MET dimer:GRB2-1:HGF:p-4Y-MET dimer:GRB2-1:SOS1SOS1SOS1
?(DOCK7)
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:CRK,CRKL:(DOCK7)p-5Y-GAB1:CRK,CRKL:(DOCK7)p-5Y-GAB1:CRK,CRKL:(DOCK7)
2
2
p21 RAS:GDP
ATP
GTP
ADP
HGF:p-4Y-MET dimer:p-Y194,Y397,HGF:p-4Y-MET dimer:p-Y194,Y397,HGF:p-4Y-MET dimer:p-Y194,Y397,Y576,Y577-PTK2:MyrG-p-Y419-SRCY576,Y577-PTK2:MyrG-p-Y419-SRCY576,Y577-PTK2:MyrG-p-Y419-SRC
p21 RAS:GTP
GDP
?RAC1:GDP
HGF:p-4Y-MET dimer,(HGF:p-Y1003,4Y-HGF:p-4Y-MET dimer,(HGF:p-Y1003,4Y-HGF:p-4Y-MET dimer,(HGF:p-Y1003,4Y-METdimer)METdimer)METdimer)
RAC1:GTP
GDPGTP
MUC20 HGF:p-4Y MET dimer:MUC20HGF:p-4Y MET dimer:MUC20HGF:p-4Y MET dimer:MUC20
Ub
LRIG1
MET:MET:MET:LRIG1LRIG1LRIG1
SOS1
RANBP9
HGF:p-4Y-MET dimer:RANBP9:HGF:p-4Y-MET dimer:RANBP9:HGF:p-4Y-MET dimer:RANBP9:SOS1SOS1SOS1
HGF:p-4Y-MET dimer:RANBP10HGF:p-4Y-MET dimer:RANBP10HGF:p-4Y-MET dimer:RANBP10
?
ARF6:GTP
MET:Ub-MET:Ub-MET:Ub-LRIG1LRIG1LRIG1
GGA3
unknown ubiquitin ligase
HGF:p-4Y-MET:p-5Y-GAB1,HGF:p-4Y-MET:p-5Y-GAB1,HGF:p-4Y-MET:p-5Y-GAB1,HGF:p-4Y-MET:GRB2-1:p-5Y-HGF:p-4Y-MET:GRB2-1:p-5Y-HGF:p-4Y-MET:GRB2-1:p-5Y-
GAB1:CRK,CRKL:GGA3:GAB1:CRK,CRKL:GGA3:GAB1:CRK,CRKL:GGA3:ARF6:GTPARF6:GTPARF6:GTP
Ub
RANBP10\\
GGC-RAB4:GTP
H2O
USP8
SHC1-2
HGF:p-4Y-MET dimer:SHC1-2HGF:p-4Y-MET dimer:SHC1-2HGF:p-4Y-MET dimer:SHC1-2
1.4E2
-1.96E1
MET receptor activates the focal adhesion kinase PTK2 (FAK1) in a process that depends on the simultaneous interaction of PTK2 with integrins and with MET. SRC is needed for PTK2 to become fully active. Activation of PTK2 is needed for HGF-induced cell motility (Beviglia et al. 1999, Parr et al. 2001, Chen and Chen 2006, Lietha et al. 2007, Chen et al. 2011, Brami-Cherrier et al. 2014).
References
Beviglia L & Kramer RH (1999). HGF induces FAK activation and integrin-mediated adhesion in
MTLn3 breast carcinoma cells. Int. J. Cancer, 83, 640-9.
Chen SY & Chen HC (2006). Direct interaction of focal adhesion kinase (FAK) with Met is required for FAK to promote hepatocyte growth factor-induced cell invasion. Mol. Cell. Biol., 26, 5155-67.
Parr C, Davies G, Nakamura T, Matsumoto K, Mason MD & Jiang WG (2001). The HGF/SF-induced phosphorylation of paxillin, matrix adhesion, and invasion of prostate cancer cells were sup-
pressed by NK4, an HGF/SF variant. Biochem. Biophys. Res. Commun., 285, 1330-7.
Brami-Cherrier K, Gervasi N, Arsenieva D, Walkiewicz K, Boutterin MC, Ortega A, ... Arold ST (2014). FAK dimerization controls its kinase-dependent functions at focal adhesions. EMBO J.,
33, 356-70.
Lietha D, Cai X, Ceccarelli DF, Li Y, Schaller MD & Eck MJ (2007). Structural basis for the autoinhib-
ition of focal adhesion kinase. Cell, 129, 1177-87.
Edit history
Date Action Author
2016-05-20 Created Orlic-Milacic M

https://reactome.org Page 17
Date Action Author
2016-06-14 Edited Orlic-Milacic M
2016-06-14 Authored Orlic-Milacic M
2016-07-11 Reviewed Heynen G, Birchmeier W
2018-08-30 Modified Croft D
Entities found in this pathway (11)
Input UniProt Id #Fold cha...
LAMC1 P11047 1.73
LAMB1 P07942 1.69
LAMA3 Q16787 1.82
COL5A3 P25940 1.9
HGF P14210 -1.98e+00
COL11A1 P12107 5.14
LAMA2 P24043 4.19
COL1A1 P02452 -2.48e+00
ITGA2 P17301 -2.06e+00
COL3A1 P02461 -1.81e+00
COL27A1 Q8IZC6 -1.98e+00
https://reactome.org Page 18
Antagonism of Activin by Follistatin (R-HSA-2473224)6.
cytosol
nucleoplasm
22
12 12
ACVR2A,BACVR1B
Activin A,AB,B:ACVR2A,B:ACVR1BActivin A,AB,B:ACVR2A,B:ACVR1BActivin A,AB,B:ACVR2A,B:ACVR1B
ATP
Activin A,AB,B:ACVR2A,B:p-ACVR1BActivin A,AB,B:ACVR2A,B:p-ACVR1BActivin A,AB,B:ACVR2A,B:p-ACVR1B
ADP
Activin A,AB,BActivin A,AB,BActivin A,AB,B
2 2
Activin AB,BActivin AB,BActivin AB,B
ACVR1C
Activin AB,B:ACVR2A,B:ACVR1CActivin AB,B:ACVR2A,B:ACVR1CActivin AB,B:ACVR2A,B:ACVR1C
1010
ATP
Activin AB,B:ACVR2A,B:p-ACVR1CActivin AB,B:ACVR2A,B:p-ACVR1CActivin AB,B:ACVR2A,B:p-ACVR1C
ADP
22
SMAD2/3
ATP
p-2S-SMAD2/3
ADP
Activin:ACVR2A,B:p-ACVR1B,CActivin:ACVR2A,B:p-ACVR1B,CActivin:ACVR2A,B:p-ACVR1B,C
2SMAD4
p-2S-SMAD2/3:SMAD4 p-2S-SMAD2/3:SMAD4
FOXH1
Activin Response Element
SMAD2,3:SMAD4:FOXH1:Activin Response Element
FOXH1:DRAP1
2
FST
Activin A,AB,B:FSTActivin A,AB,B:FSTActivin A,AB,B:FST
2FSTL3 Activin A,AB,B:FSTL3Activin A,AB,B:FSTL3Activin A,AB,B:FSTL3
1.4E2
-1.96E1
Cellular compartments: extracellular region.
Both Follistatin (FST) and Follistatin-like-3 (FSTL3) irreversibly bind Activin dimers and prevent Activin from interacting with its receptor (reviewed in Schneyer et al. 2004, Xia and Schneyer 2009). Though functionally similar in vitro, FST and FSTL3 do not function identically in vivo. Mice lack-ing FST die shortly after birth due to defects in muscle and bone (Matzuk et al. 1995); mice lacking FSTL3 are viable but have altered glucose metabolism (Mukherjee et al. 2007).
References
Schneyer A, Sidis Y, Xia Y, Saito S, del Re E, Lin HY & Keutmann H (2004). Differential actions of
follistatin and follistatin-like 3. Mol. Cell. Endocrinol., 225, 25-8.
Xia Y & Schneyer AL (2009). The biology of activin: recent advances in structure, regulation and
function. J. Endocrinol., 202, 1-12.
Matzuk MM, Lu N, Vogel H, Sellheyer K, Roop DR & Bradley A (1995). Multiple defects and perinat-
al death in mice deficient in follistatin. Nature, 374, 360-3.
Mukherjee A, Sidis Y, Mahan A, Raher MJ, Xia Y, Rosen ED, ... Schneyer AL (2007). FSTL3 deletion reveals roles for TGF-beta family ligands in glucose and fat homeostasis in adults. Proc. Natl.
Acad. Sci. U.S.A., 104, 1348-53.
Edit history
Date Action Author
2012-09-21 Edited May B
2012-09-21 Authored May B

https://reactome.org Page 19
Date Action Author
2012-09-22 Created May B
2012-11-14 Reviewed Chen YG
2018-08-23 Modified Schmidt EE
Entities found in this pathway (4)
Input UniProt Id #Fold cha...
INHBB P09529 12.69
INHBA P08476 2.11
FST P19883 -3.46e+00
FSTL3 O95633 3.58

https://reactome.org Page 20
Assembly of collagen fibrils and other multimeric structures (R-HSA-2022090)7.
cytosol
Collagen fibrils
PXDN:Br-
Network formingtropocollagens
Collagen networks
2
Collagen alpha-1(VII)Collagen alpha-1(VII)Collagen alpha-1(VII)trimertrimertrimer
Collagen type VII hexamerCollagen type VII hexamerCollagen type VII hexamer
Collagen type I fibrils with histidino-hydroxylysinoleucine cross-links
2 Collagen type IV networks withCollagen type IV networks withCollagen type IV networks withsulfilimine cross-linkssulfilimine cross-linkssulfilimine cross-links
Collagen type XVII fibril:Integrin alpha6beta4
BPAG1e:PlectinBPAG1e:PlectinBPAG1e:Plectin
Collagen type VII fibril:Collagen type VII fibril:Collagen type VII fibril:Laminin-332Laminin-332Laminin-332
CD151 Type I hemidesmosomeType I hemidesmosomeType I hemidesmosomecomplexcomplexcomplex
Collagen type I fibrils with hydroxylysino-5-Collagen type I fibrils with hydroxylysino-5-Collagen type I fibrils with hydroxylysino-5-ketonorleucine crosslinksketonorleucine crosslinksketonorleucine crosslinks
Collagen type 1 fibrils cross-linked bydehydro-lysinonorleucine crosslinks
Endostatin releasingEndostatin releasingEndostatin releasingproteasesproteasesproteases
Collagen type XI fibril:Collagen type II fibril
Collagen alpha-1(VII) NC2 regionCollagen alpha-1(VII) NC2 regionCollagen alpha-1(VII) NC2 region
Tropocollagens
Collagen type I fibrils withCollagen type I fibrils withCollagen type I fibrils withhydroxylysyl-pyrrole cross-linkshydroxylysyl-pyrrole cross-linkshydroxylysyl-pyrrole cross-links
Collagen type VII -NC2Collagen type VII -NC2Collagen type VII -NC2hexamerhexamerhexamer
Collagen fibres
Laminin-332Laminin-332Laminin-332
Collagen type VII fibril
Collagen type IV networksCollagen type IV networksCollagen type IV networks
18
18
18
18
Collagen type XVIII
Collagen type XVCollagen type XVCollagen type XV
COL15A1(1212-1388)
COL18A1(1572-11754)
COL18A1(?-1571)
COL15A1(?-1211)
2
Collagen type I fibril
Collagen type IX
Anchoring fibril complexAnchoring fibril complexAnchoring fibril complex
Collagen type I fibrils with lysyl-pyridinolineCollagen type I fibrils with lysyl-pyridinolineCollagen type I fibrils with lysyl-pyridinolinecross-linkscross-linkscross-links
H2O2
Collagen type I fibril with freehydroxylysines
H2O
Collagen type I fibrils with deH-HLNL cross-links
6
Collagen type VII NC2proteinases
Prolysyl oxidases
Lysyl oxidasepropeptides
Lysyl oxidases
Procollagen C-Procollagen C-Procollagen C-proteinasesproteinasesproteinases
2
Collagen type XI fibril
O2
Collagen type I fibrils with hydroxylysyl-Collagen type I fibrils with hydroxylysyl-Collagen type I fibrils with hydroxylysyl-pyridinoline cross-linkspyridinoline cross-linkspyridinoline cross-links
H2O2
Collagen type II fibril
NH3
Lysyl oxidases:Cu2+
Collagen type I fibril withhydroxyallysines
Collagen type II fibril:Collagentype IX
H2O
Collagen type XII, XIV fibrils
Collagen type I fibril withallysines
Collagen type I fibrils with lysyl-pyrroleCollagen type I fibrils with lysyl-pyrroleCollagen type I fibrils with lysyl-pyrrolecross-linkscross-linkscross-links
Collagen type I, II fibrils
Collagen type I,II:XII,XIV fibrils
Collagen type I fibrils with lysino-5-Collagen type I fibrils with lysino-5-Collagen type I fibrils with lysino-5-ketonorleucine cross-linksketonorleucine cross-linksketonorleucine cross-links
Collagen type X network
Collagen type X:type II fibrils
1.4E2
-1.96E1
Collagen trimers in triple-helical form, referred to as procollagen or collagen molecules, are expor-ted from the ER and trafficked through the Golgi network before secretion into the extracellular space. For fibrillar collagens namely types I, II, III, V, XI, XXIV and XXVII (Gordon & Hahn 2010, Ricard-Blum 2011) secretion is concomitant with processing of the N and C terminal collagen propeptides. These processed molecules are known as tropocollagens, considered to be the units of higher order collagen structures. They form within the extracellular space via a process that can proceed spontaneously, but in the cellular environment is regulated by many collagen binding pro-teins such as the FACIT (Fibril Associated Collagens with Interrupted Triple helices) family colla-gens and Small Leucine-Rich Proteoglycans (SLRPs). The architecture formed ultimately depends on the collagen subtype and the cellular conditions. Structures include the well-known fibrils and fibres formed by the major structural collagens type I and II plus several different types of supra-molecular assembly (Bruckner 2010). The mechanical and physical properties of tissues depend on the spatial arrangement and composition of these collagen-containing structures (Kadler et al. 1996, Shoulders & Raines 2009, Birk & Bruckner 2011).
Fibrillar collagen structures are frequently heterotypic, composed of a major collagen type in asso-ciation with smaller amounts of other types, e.g. type I collagen fibrils are associated with types III and V, while type II fibrils frequently contain types IX and XI (Wess 2005). Fibres composed exclus-ively of a single collagen type probably do not exist, as type I and II fibrils require collagens V and XI respectively as nucleators (Kadler et al. 2008, Wenstrup et al. 2011). Much of the structural un-derstanding of collagen fibrils has been obtained with fibril-forming collagens, particularly type I, but some central features are believed to apply to at least the other fibrillar collagen subtypes (Wess 2005). Fibril diameter and length varies considerably, depending on the tissue and collagen types (Fang et al. 2012). The reasons for this are poorly understood (Wess 2005).

https://reactome.org Page 21
Some tissues such as skin have fibres that are approximately the same diameter while others such as tendon or cartilage have a bimodal distribution of thick and thin fibrils. Mature type I collagen fibrils in tendon are up to 1 cm in length, with a diameter of approx. 500 nm. An individual fibrillar collagen triple helix is less than 1.5 nm in diameter and around 300 nm long; collagen molecules must assemble to give rise to the higher-order fibril structure, a process known as fibrillogenesis, prevented by the presence of C-terminal propeptides (Kadler et al. 1987). In electron micrographs, fibrils have a banded appearance, due to regular gaps where fewer collagen molecules overlap, which occur because the fibrils are aligned in a quarter-stagger arrangement (Hodge & Petruska 1963). Collagen microfibrils are believed to have a quasi-hexagonal unit cell, with tropocollagen ar-ranged to form supertwisted, right-handed microfibrils that interdigitate with neighbouring mi-crofibrils, leading to a spiral-like structure for the mature collagen fibril (Orgel et al. 2006, Holmes & Kadler 2006).
Neighbouring tropocollagen monomers interact with each other and are cross-linked covalently by lysyl oxidase (Orgel et al. 2000, Maki 2006). Mature collagen fibrils are stabilized by lysyl oxidase-mediated cross-links. Hydroxylysyl pyridinoline and lysyl pyridinoline cross-links form between (hydroxy) lysine and hydroxylysine residues in bone and cartilage (Eyre et al. 1984). Arginoline cross-links can form in cartilage (Eyre et al. 2010); mature bovine articular cartilage contains roughly equimolar amounts of arginoline and hydroxylysyl pyridinoline based on peptide yields. Mature collagen fibrils in skin are stabilized by the lysyl oxidase-mediated cross-link histidino-hydroxylysinonorleucine (Yamauch et al. 1987). Due to the quarter-staggered arrangement of colla-gen molecules in a fibril, telopeptides most often interact with the triple helix of a neighbouring collagen molecule in the fibril, except for collagen molecules in register staggered by 4D from an-other collagen molecule. Fibril aggregation in vitro can be unipolar or bipolar, influenced by tem-perature and levels of C-proteinase, suggesting a role for the N- and C- propeptides in regulation of the aggregation process (Kadler et al. 1996). In vivo, collagen molecules at the fibril surface may re-tain their N-propeptides, suggesting that this may limit further accretion, or alternatively repres-ents a transient stage in a model whereby fibrils grow in diameter through a cycle of deposition, cleavage and further deposition (Chapman 1989).
In vivo, fibrils are often composed from more than one type of collagen. Type III collagen is found associated with type I collagen in dermal fibrils, with the collagen III on the periphery, suggesting a regulatory role (Fleischmajer et al. 1990). Type V collagen associates with type I collagen fibrils, where it may limit fibril diameter (Birk et al. 1990, White et al. 1997). Type IX associates with the surface of narrow diameter collagen II fibrils in cartilage and the cornea (Wu et al. 1992, Eyre et al. 2004). Highly specific patterns of crosslinking sites suggest that collagen IX functions in interfibril-lar networking (Wess 2005). Type XII and XIV collagens are localized near the surface of banded collagen I fibrils (Nishiyama et al. 1994). Certain fibril-associated collagens with interrupted triple helices (FACITs) associate with the surface of collagen fibrils, where they may serve to limit fibril fusion and thereby regulate fibril diameter (Gordon & Hahn 2010). Collagen XV, a member of the multiplexin family, is almost exclusively associated with the fibrillar collagen network, in very close proximity to the basement membrane. In human tissues collagen XV is seen linking banded collagen fibers subjacent to the basement membrane (Amenta et al. 2005). Type XIV collagen, SLRPs and discoidin domain receptors also regulate fibrillogenesis (Ansorge et al. 2009, Kalamajski et al. 2010, Flynn et al. 2010).

https://reactome.org Page 22
Collagen IX is cross-linked to the surface of collagen type II fibrils (Eyre et al. 1987). Type XII and XIV collagens are found in association with type I (Walchli et al. 1994) and type II (Watt et al. 1992, Eyre 2002) fibrils in cartilage. They are thought to associate non-covalently via their COL1/NC1 do-mains (Watt et al. 1992, Eyre 2002).
Some non-fibrillar collagens form supramolecular assemblies that are distinct from typical fibrils. Collagen VII forms anchoring fibrils, composed of antiparallel dimers that connect the dermis to the epidermis (Bruckner-Tuderman 2009). During fibrillogenesis, the nascent type VII procollagen molecules dimerize in an antiparallel manner. The C-propeptides are then removed by Bone morphogenetic protein 1 (Rattenholl et al. 2002) and the processed antiparallel dimers aggregate laterally. Collagens VIII and X form hexagonal networks and collagen VI forms beaded filament (Gordon & Hahn 2010, Ricard-Blum et al. 2011).
References
Kadler KE, Holmes DF, Trotter JA & Chapman JA (1996). Collagen fibril formation. Biochem J, 316,
1-11.
Orgel JP, San Antonio JD & Antipova O (2011). Molecular and structural mapping of collagen fibril
interactions. Connect. Tissue Res., 52, 2-17.
Edit history
Date Action Author
2011-08-05 Authored Jupe S
2011-11-25 Created Jupe S
2012-10-08 Reviewed Kalamajski S, Raleigh S
2012-11-12 Edited Jupe S
2012-11-19 Reviewed Ricard-Blum S
2018-08-23 Modified Schmidt EE
Entities found in this pathway (16)
Input UniProt Id #Fold cha...
COL4A5 P29400 -1.87e+00
PCOLCE Q15113 -1.55e+00
COL4A3 Q01955 4.74
LAMA3 Q16787 1.82
COL4A4 P53420 3.93
COL7A1 Q02388 2.34
COL5A3 P25940 1.9
COL4A1 P02462 2.52
COL11A1 P12107 5.14
COL4A2 P08572 1.85
PLEC Q15149 1.79
COL1A1 P02452 -2.48e+00
COL3A1 P02461 -1.81e+00
COL27A1 Q8IZC6 -1.98e+00
COL15A1 P39059 -1.90e+00
MMP7 P09237 28.84


https://reactome.org Page 24
Interleukin-7 signaling (R-HSA-1266695)8.
cytosol
nucleoplasm
IL7:IL7R:JAK1
AcK(9,14,18,79)-p(S10,T11)-histone H3
IL2RG:JAK3 IL7:IL7R:JAK1:IL2RG:JAK3
?
STAT5A,STAT5BSTAT5A,STAT5BSTAT5A,STAT5B
\\
CRLF2:IL7R
TSLP
IL7R:TSLP:CRLF2IL7R:TSLP:CRLF2IL7R:TSLP:CRLF2
IL7
HGF(495-728)
HGF(495-728):IL7HGF(495-728):IL7HGF(495-728):IL7
IL7R
JAK1
IL7R:JAK1
JAK3
IL2RG
CISH,SOCS1,SOCS2CISH,SOCS1,SOCS2CISH,SOCS1,SOCS2
ATPADP
IL7:p-Y449-IL7R:JAK1:IL2RG:p-JAK3
IL7:IL7R:JAK1:IL2RG:p-JAK3
ADPATP
?
BRWD1 gene
2x(p-STAT5A,p-STAT5B):BRWD12x(p-STAT5A,p-STAT5B):BRWD12x(p-STAT5A,p-STAT5B):BRWD1genegenegene
\\
\\
CISH gene, SOCS1 gene,CISH gene, SOCS1 gene,CISH gene, SOCS1 gene,SOCS2 gene:p-STAT5 dimerSOCS2 gene:p-STAT5 dimerSOCS2 gene:p-STAT5 dimer
\\
IL7R:TSLP:CRLF2:IL7R:TSLP:CRLF2:IL7R:TSLP:CRLF2:STAT3STAT3STAT3
\\ BRWD1
ATP
PI3K regulatory subunitsPI3K regulatory subunitsPI3K regulatory subunits
IL7:p-Y449-IL7R:JAK1:p-FYN:IL2RG:IL7:p-Y449-IL7R:JAK1:p-FYN:IL2RG:IL7:p-Y449-IL7R:JAK1:p-FYN:IL2RG:JAK3:PI3K-regulatory subunitsJAK3:PI3K-regulatory subunitsJAK3:PI3K-regulatory subunits
IL7R:TSLP:CRLF2:p-IL7R:TSLP:CRLF2:p-IL7R:TSLP:CRLF2:p-STAT3STAT3STAT3
ADP
p-STAT5A, p-STAT5Bp-STAT5A, p-STAT5Bp-STAT5A, p-STAT5B
p-STAT5 dimerp-STAT5 dimerp-STAT5 dimer
\\
p-STAT3
IRS1,IRS2IRS1,IRS2IRS1,IRS2
p-STAT5 dimerp-STAT5 dimerp-STAT5 dimer
?
?
Immunoglobulin kappalocus
RAG1:RAG2recombinase
RAG1:RAG2recombinase:
Immunoglobulin kappalocus
BRWD1:AcK(9,14,18,79)-p(S10,T11)-histone H3
SMARCA4
BRWD1:SMARCA4
\\
ATP
STAT3
IL7:p-Y449-IL7R:JAK1:IL2RG:IL7:p-Y449-IL7R:JAK1:IL2RG:IL7:p-Y449-IL7R:JAK1:IL2RG:p-JAK3:p-STAT5A,p-STAT5Bp-JAK3:p-STAT5A,p-STAT5Bp-JAK3:p-STAT5A,p-STAT5B
ADP
IL7:p-Y449-IL7R:JAK1:IL7:p-Y449-IL7R:JAK1:IL7:p-Y449-IL7R:JAK1:IL2RG:p-JAK3:STAT5A,IL2RG:p-JAK3:STAT5A,IL2RG:p-JAK3:STAT5A,
STAT5BSTAT5BSTAT5B
BRWD1:AcK9,14-pS10-histone H3
?
2
IL7:p-Y449-IL7R:JAK1:IL2RG:JAK3:PI3K-IL7:p-Y449-IL7R:JAK1:IL2RG:JAK3:PI3K-IL7:p-Y449-IL7R:JAK1:IL2RG:JAK3:PI3K-regulatory subunits:IRS1,IRS2regulatory subunits:IRS1,IRS2regulatory subunits:IRS1,IRS2
?
CISH gene, SOCS1 gene,CISH gene, SOCS1 gene,CISH gene, SOCS1 gene,SOCS2 geneSOCS2 geneSOCS2 gene
1.4E2
-1.96E1
Interleukin-7 (IL7) is produced primarily by T zone fibroblastic reticular cells found in lymphoid organs, and also expressed by non-hematopoietic stromal cells present in other tissues including the skin, intestine and liver. It is an essential survival factor for lymphocytes, playing a key anti-ap-optotic role in T-cell development, as well as mediating peripheral T-cell maintenance and prolifer-ation. This dual function is reflected in a dose-response relationship that distinguishes the survival function from the proliferative activity; low doses of IL7 (<1 ng/ml) sustain only survival, higher doses (>1 ng/ml) promote survival and cell cycling (Kittipatarin et al. 2006, Swainson et al. 2007).
The IL7 receptor is a heterodimeric complex of the the common cytokine-receptor gamma chain (IL2RG, CD132, or Gc) and the IL7-receptor alpha chain (IL7R, IL7RA, CD127). Both chains are members of the type 1 cytokine family. Neither chain is unique to the IL7 receptor as IL7R is util-ized by the receptor for thymic stromal lymphopoietin (TSLP) while IL2RG is shared with the re-ceptors for IL2, IL4, IL9, IL15 and IL21. IL2RG consists of a single transmembrane region and a 240aa extracellular region that includes a fibronectin type III (FNIII) domain thought to be involved in receptor complex formation. It is expressed on most lymphocyte populations. Null mutations of IL2RG in humans cause X-linked severe combined immunodeficiency (X-SCID), which has a pheno-type of severely reduced T-cell and natural killer (NK) cell populations, but normal numbers of B cells. In addition to reduced T- and NK-cell numbers, Il2rg knockout mice also have dramatically reduced B-cell populations suggesting that Il2rg is more critical for B-cell development in mice than in humans. Patients with severe combined immunodeficiency (SCID) phenotype due to IL7R mutations (see Puel & Leonard 2000), or a partial deficiency of IL7R (Roifman et al. 2000) have markedly reduced circulating T cells, but normal levels of peripheral blood B cells and NK cells, similar to the phenotype of IL2RG mutations, highlighting a requirement for IL7 in T cell lymph-opoiesis. It has been suggested that IL7 is essential for murine, but not human B cell development, but recent studies indicate that IL7 is essential for human B cell production from adult bone mar-row and that IL7-induced expansion of the progenitor B cell compartment is increasingly critical for human B cell production during later stages of development (Parrish et al. 2009).

https://reactome.org Page 25
IL7 has been shown to induce rapid and dose-dependent tyrosine phosphorylation of JAKs 1 and 3, and concomitantly tyrosine phosphorylation and DNA-binding activity of STAT5a/b (Foxwell et al. 1995). IL7R was shown to directly induce the activation of JAKs and STATs by van der Plas et al. (1996). Jak1 and Jak3 knockout mice displayed severely impaired thymic development, further sup-porting their importance in IL7 signaling (Rodig et al. 1998, Nosaka et al. 1995).
The role of STAT5 in IL7 signaling has been studied largely in mouse models. Tyr449 in the cyto-plasmic domain of IL7RA is required for T-cell development in vivo and activation of JAK/STAT5 and PI3k/Akt pathways (Jiang et al. 2004, Pallard et al. 1999). T-cells from an IL7R Y449F knock-in mouse did not activate STAT5 (Osbourne et al. 2007), indicating that IL7 regulates STAT5 activity via this key tyrosine residue. STAT5 seems to enhance proliferation of multiple cell lineages in mouse models but it remains unclear whether STAT5 is required solely for survival signaling or also for the induction of proliferative activity (Kittipatarin & Khaled, 2007).
The model for IL7 receptor signaling is believed to resemble that of other Gc family cytokines, based on detailed studies of the IL2 receptor, where IL2RB binds constitutively to JAK1 while JAK3 is pre-associated uniquely with the IL2RG chain. Extending this model to IL7 suggests a similar series of events: IL7R constitutively associated with JAK1 binds IL7, the resulting trimer recruits IL2RG:JAK3, bringing JAK1 and JAK3 into proximity. The association of both chains of the IL7 re-ceptor orients the cytoplasmic domains of the receptor chains so that their associated kinases (Janus and phosphatidylinositol 3-kinases) can phosphorylate sequence elements on the cytoplas-mic domains (Jiang et al. 2005). JAKs have low intrinsic enzymatic activity, but after mutual phos-phorylation acquire much higher activity, leading to phosphorylation of the critical Y449 site on IL7R. This site binds STAT5 and possibly other signaling adapters, they in turn become phos-phorylated by JAK1 and/or JAK3. Phosphorylated STATs translocate to the nucleus and trigger the transcriptional events of their target genes.
The role of the PI3K/AKT pathway in IL7 signaling is controversial. It is a potential T-cell survival pathway because in many cell types PI3K signaling regulates diverse cellular functions such as cell cycle progression, transcription, and metabolism. The ERK/MAPK pathway does not appear to be involved in IL7 signaling (Crawley et al. 1996).

https://reactome.org Page 26
It is not clear how IL7 influences cell proliferation. In the absence of a proliferative signal such as IL7 or IL3, dependent lymphocytes arrest in the G0/G1 phase of the cell cycle. To exit this phase, cells typically activate specific G1 Cyclin-dependent kinases/cyclins and down regulate cell cycle in-hibitors such as Cyclin-dependent kinase inhibitor 1B (Cdkn1b or p27kip1). There is indirect evid-ence suggesting a possible role for IL7 stimulated activation of PI3K/AKT signaling, obtained from transformed cell lines and thymocytes, but not confirmed by observations using primary T-cells (Kittipatarin & Khaled, 2007). IL7 withdrawal results in G1/S cell cycle arrest and is correlated with loss of cdk2 activity (Geiselhart et al. 2001), both events which are known to be regulated by the de-phosphorylating activity of Cdc25A. Expression of a p38 MAPK-resistant Cdc25A mutant in an IL-7-dependent T-cell line as well as in peripheral, primary T-cells was sufficient to sustain cell survival and promote cell cycling for several days in the absence of IL7 (Khaled et al. 2005). Cdkn1b is a member of the CIP/KIP family of cyclin-dependent cell cycle inhibitors (CKIs) that negatively regu-lates the G1/S transition. In IL7 dependent T-cells, the expression of Cdkn1b was sufficient to cause G1 arrest in the presence of IL7. Withdrawal of IL7 induced the upregulation of Cdkn1b and arres-ted cells in G1 while siRNA knockout of Cdkn1b enhanced cell cycle progression. However, adopt-ive transfer of Cdkn1b-deficient lymphocytes into IL7 deficient mice indicated that loss of Cdkn1b could only partially compensate for the IL7 signal needed by T-cells to expand in a lymphopenic en-vironment (Li et al. 2006), so though Cdkn1b may be involved in negative regulation of the cell cycle through an effect on cdk2 activity, its absence is not sufficient to fully induce cell cycling under lymphopenic conditions.
References
Edit history
Date Action Author
2010-05-17 Authored Ray KP
2011-05-06 Edited Jupe S
2011-05-06 Created Jupe S
2011-11-03 Reviewed Puck J
2016-03-23 Edited Orlic-Milacic M
2016-05-11 Revised Mandal M
2017-07-26 Reviewed Kumar U
2017-08-21 Reviewed Goronzy JJ
2018-08-24 Modified Schmidt EE
Entities found in this pathway (9)
Input UniProt Id #Fold cha...
PIK3R3 Q92569 -3.21e+00
PIK3R1 P27986 3.08
TSLP Q969D9 -6.97e+00
IRS2 Q9Y4H2 4.46
IRS1 P35568 1.66
HGF P14210 -1.98e+00
STAT3 P40763 1.53
SOCS1 O15524 -2.28e+00

https://reactome.org Page 27
Input UniProt Id #Fold cha...
STAT5A P42229 1.5
Input Ensembl Id #Fold cha...
SOCS1 ENSG00000185338 -2.28e+00

https://reactome.org Page 28
Collagen chain trimerization (R-HSA-8948216)9.
endoplasmic reticulum lumen
cytosol
2
Alpha-1(I) propeptidesAlpha-1(I) propeptidesAlpha-1(I) propeptides
Alpha-2(I)propeptides
C-linked procollagen type IC-linked procollagen type IC-linked procollagen type Itrimertrimertrimer
3Alpha-1(II) propeptides C-linked procollagen type IItrimer
3Alpha-1(III) propeptidesAlpha-1(III) propeptidesAlpha-1(III) propeptidesC-linked procollagen type IIIC-linked procollagen type IIIC-linked procollagen type III
trimertrimertrimer
3
2
Alpha-3(IV) chainsAlpha-3(IV) chainsAlpha-3(IV) chains
Alpha-5(IV) chainsAlpha-5(IV) chainsAlpha-5(IV) chains
Alpha-1(IV) chainsAlpha-1(IV) chainsAlpha-1(IV) chains
Alpha-2(IV) chainsAlpha-2(IV) chainsAlpha-2(IV) chains
Alpha-4(IV) chainsAlpha-4(IV) chainsAlpha-4(IV) chains
Alpha-6(IV) chains
C-linked procollagen type IVC-linked procollagen type IVC-linked procollagen type IVtrimerstrimerstrimers
3
4
2
Alpha-1(V)propeptides
Alpha-3(V)Alpha-3(V)Alpha-3(V)propeptidespropeptidespropeptides
Alpha-2(V)propeptides C-linked procollagen type V
trimers
Alpha-3-6(VI)propeptides
Alpha-2(VI) chains
Alpha-1(VI) chains
C-linked procollagen type VItrimer
3Alpha-1(VII) chainsAlpha-1(VII) chainsAlpha-1(VII) chains C-linked procollagen type VIIC-linked procollagen type VIIC-linked procollagen type VIItrimertrimertrimer
2
Alpha-2(VIII) chains
Alpha-1(VIII) chains
C-linked procollagen type VIIItrimer
Alpha-2(IX) chains
Alpha-1(IX) chains
Alpha-3(IX) chains
C-linked procollagen type IXtrimer
3Alpha-1(X) chains C-linked procollagen type Xtrimer
Alpha-1(XI)Alpha-1(XI)Alpha-1(XI)propeptidespropeptidespropeptides
Alpha-2(XI)propeptides
C-linked procollagen type XItrimer
3Alpha-1(XII) chainsC-linked procollagen type XII
trimer
3Alpha-1(XIII) chains C-linked procollagen type XIIItrimer
3Alpha-1(XIV) chains C-linked procollagen type XIVtrimer
3Alpha-1(XIX) chainsC-linked procollagen type XIX
trimer
3Alpha-1(XV) chainsAlpha-1(XV) chainsAlpha-1(XV) chains C-linked procollagen type XVC-linked procollagen type XVC-linked procollagen type XVtrimertrimertrimer
3Alpha-1(XVI) chainsC-linked procollagen type XVI
trimer 3Alpha-1(XVII) chains C-linked procollagen type XVIItrimer
3Alpha-1(XXVIII) chainsC-linked procollagen type XXVIII
trimer
3Collagen alpha-1(XVIII)chains
C-linked procollagen type XVIIItrimer
3Alpha-1(XX) chainsC-linked procollagen type XX
trimer 3Alpha-1(XXI) chains C-linked procollagen type XXItrimer
3Alpha-1(XXII) chainsC-linked procollagen type XXII
trimer
3Alpha-1(XXIII) chains C-linked procollagen type XXIIItrimer
3Alpha-1(XXIV)propeptides
C-linked procollagen type XXIVtrimer
3Alpha-1(XXV) chainsC-linked procollagen type XXV
trimer
3Alpha-1(XXVI) chains C-linked procollagen type XXVItrimer
3Alpha-1(XXVII)Alpha-1(XXVII)Alpha-1(XXVII)propeptidespropeptidespropeptides
C-linked procollagen type XXVIIC-linked procollagen type XXVIIC-linked procollagen type XXVIItrimertrimertrimer
1.4E2
-1.96E1
The C-propeptides of collagen propeptide chains are essential for the association of three peptide chains into a trimeric but non-helical procollagen. This initial binding event determines the com-position of the trimer, brings the individual chains into the correct register and initiates formation of the triple helix at the C-terminus, which then proceeds towards the N-terminus in a zipper-like fashion (Engel & Prockop 1991). Most early refolding studies were performed with collagen type III, which contains a disulfide linkage at the C-terminus of its triple helix (Bächinger et al. 1978, Bruck-ner et al. 1978) that acts as a permanent linker even after removal of the non-collagenous domains.
Mutations within the C-propeptides further suggest that they are crucial for the correct interaction of the three polypeptide chains and for subsequent correct folding (refs. in Boudko et al. 2011).
References
Byers PH, Click EM, Harper E & Bornstein P (1975). Interchain disulfide bonds in procollagen are located in a large nontriple-helical COOH-terminal domain. Proc Natl Acad Sci U S A, 72, 3009-13
.
Bächinger HP, Brückner P, Timpl R & Engel J (1978). The role of cis-trans isomerization of peptide bonds in the coil leads to and comes from triple helix conversion of collagen. Eur J Biochem, 90,
605-13.
Edit history
Date Action Author
2012-04-11 Authored Jupe S
2012-05-24 Reviewed Canty-Laird EG
2016-11-03 Edited Jupe S
2016-11-11 Created Jupe S

https://reactome.org Page 29
Date Action Author
2018-08-23 Modified Schmidt EE
Entities found in this pathway (12)
Input UniProt Id #Fold cha...
COL4A5 P29400 -1.87e+00
COL4A3 Q01955 4.74
COL4A4 P53420 3.93
COL7A1 Q02388 2.34
COL5A3 P25940 1.9
COL4A1 P02462 2.52
COL11A1 P12107 5.14
COL4A2 P08572 1.85
COL1A1 P02452 -2.48e+00
COL3A1 P02461 -1.81e+00
COL27A1 Q8IZC6 -1.98e+00
COL15A1 P39059 -1.90e+00

https://reactome.org Page 30
Signaling by Leptin (R-HSA-2586552)10.
cytosol
nucleoplasm
2
LEP
LEPR:JAK2LEPR:JAK2LEPR:JAK2
LEP:LEPR:JAK2LEP:LEPR:JAK2LEP:LEPR:JAK2
24
24
LEP:LEPR:p-JAK2LEP:LEPR:p-JAK2LEP:LEPR:p-JAK2
6 6
ATP
LEP:p-LEPR:p-JAK2LEP:p-LEPR:p-JAK2LEP:p-LEPR:p-JAK2
ADP
2
PTPN11
LEP:p-LEPR:p-JAK2:SHP2LEP:p-LEPR:p-JAK2:SHP2LEP:p-LEPR:p-JAK2:SHP2
4 4
ATP
LEP:p-LEPR:p-JAK2:p-SHP2LEP:p-LEPR:p-JAK2:p-SHP2LEP:p-LEPR:p-JAK2:p-SHP2
ADP
2
STAT5A,STAT5A,STAT5A,STAT5BSTAT5BSTAT5B
LEP:p-LEPR:p-JAK2:STAT5LEP:p-LEPR:p-JAK2:STAT5LEP:p-LEPR:p-JAK2:STAT5
2 2
ATP
LEP:p-LEPR:p-JAK2:p-STAT5LEP:p-LEPR:p-JAK2:p-STAT5LEP:p-LEPR:p-JAK2:p-STAT5
ADP
2
p-STAT5A, p-STAT5Bp-STAT5A, p-STAT5Bp-STAT5A, p-STAT5B
2STAT3
LEP:p-LEPR:p-JAK2:STAT3LEP:p-LEPR:p-JAK2:STAT3LEP:p-LEPR:p-JAK2:STAT3
RAF/MAP kinase cascade
ATPADP
2
2
ATP
LEP:p-LEPR:p-JAK2:p-LEP:p-LEPR:p-JAK2:p-LEP:p-LEPR:p-JAK2:p-STAT3STAT3STAT3
ADP
2p-Y705-STAT3P
2SH2B1-2
LEP:p-LEPR:p-JAK2:SH2B1LEP:p-LEPR:p-JAK2:SH2B1LEP:p-LEPR:p-JAK2:SH2B12
IRS1,2IRS1,2IRS1,2
LEP:p-LEPR:p-JAK2:SH2B1:IRS1,2LEP:p-LEPR:p-JAK2:SH2B1:IRS1,2LEP:p-LEPR:p-JAK2:SH2B1:IRS1,2
22
ATPADP
LEP:p-LEPR:p-JAK2:SH2B1:p-IRS1,2LEP:p-LEPR:p-JAK2:SH2B1:p-IRS1,2LEP:p-LEPR:p-JAK2:SH2B1:p-IRS1,2
2
SOCS3
LEP:p-LEPR:p-LEP:p-LEPR:p-LEP:p-LEPR:p-JAK2:SOCS3JAK2:SOCS3JAK2:SOCS3
2
2p-Y705-STAT3 dimerp-Y705-STAT3 dimerp-Y705-STAT3 dimer
\\p-Y705-STAT3 dimerp-Y705-STAT3 dimerp-Y705-STAT3 dimer
p-STAT5 dimerp-STAT5 dimerp-STAT5 dimer
p-STAT5 dimerp-STAT5 dimerp-STAT5 dimer
1.4E2
-1.96E1
Cellular compartments: plasma membrane, cytosol.
Leptin (LEP, OB, OBS), a circulating adipokine, and its receptor LEPR (DB, OBR) control food intake and energy balance and are implicated in obesity-related diseases (recently reviewed in Amitani et al. 2013, Dunmore and Brown 2013, Cottrell and Mercer 2012, La Cava 2012, Marroqui et al. 2012, Paz-Filho et al. 2012, Denver et al. 2011, Lee 2011, Marino et al. 2011, Morton and Schwartz 2011, Scherer and Buettner 2011, Shan and Yeo 2011, Wauman and Tavernier 2011, Dardeno et al. 2010, Bjorbaek 2009, Morris and Rui 2009, Myers et al. 2008), including cancer (Guo et al. 2012), inflam-mation (Newman and Gonzalez-Perez 2013, Iikuni et al. 2008), and angiogenesis (Gonzalez-Perez et al. 2013).
The identification of spontaneous mutations in the leptin gene (ob or LEP) and the leptin receptor gene (Ob-R, db or LEPR) genes in mice opened up a new field in obesity research. Leptin was dis-covered as the product of the gene affected by the ob (obesity) mutation, which causes obesity in mice. Likewise LEPR is the product of the gene affected by the db (diabetic) mutation. Leptin bind-ing to LEPR induces canonical (JAK2/STATs; MAPK/ERK 1/2, PI-3K/AKT) and non-canonical signal-ing pathways (PKC, JNK, p38 MAPK and AMPK) in diverse cell types. The binding of leptin to the long isoform of LEPR (OB-Rl) initiates a phosphorylation cascade that results in transcriptional ac-tivation of target genes by STAT5 and STAT3 and activation of the PI3K pathway(not shown here), the MAPK/ERK pathway, and the mTOR/S6K pathway. Shorter LEPR isoforms with truncated intra-cellular domains are unable to activate the STAT pathway, but can transduce signals by way of ac-tivation of JAK2, IRS-1 or ERKs, including MAPKs.

https://reactome.org Page 31
LEPR is constitutively bound to the JAK2 kinase. Binding of LEP to LEPR causes a conformational change in LEPR that activates JAK2 autophosphorylation followed by phosphorylation of LEPR by JAK2. Phosphorylated LEPR binds STAT3, STAT5, and SHP2 which are then phosphorylated by JAK2. Phosphorylated JAK2 binds SH2B1 which then binds IRS1/2, resulting in phosphorylation of IRS1/2 by JAK2. Phosphorylated STAT3 and STAT5 dimerize and translocate to the nucleus where they activate transcription of target genes (Jovanovic et al. 2010). SHP2 activates the MAPK path-way. IRS1/2 activate the PI3K/AKT pathway which may be the activator of mTOR/S6K.
Several isoforms of LEPR have been identified (reviewed in Gorska et al. 2010). The long isoform (LEPRb, OBRb) is expressed in the hypothalamus and all types of immune cells. It is the only iso-form known to fully activate signaling pathways in response to leptin. Shorter isoforms (LEPRa, LEPRc, LEPRd, and a soluble isoform LEPRe) are able to interact with JAK kinases and activate oth-er pathways, however their roles in energy homeostasis are not fully characterized.
References
Wauman J & Tavernier J (2011). Leptin receptor signaling: pathways to leptin resistance. Front.
Biosci., 16, 2771-93.
Dardeno TA, Chou SH, Moon HS, Chamberland JP, Fiorenza CG & Mantzoros CS (2010). Leptin in
human physiology and therapeutics. Front Neuroendocrinol, 31, 377-93.
Bjørbaek C (2009). Central leptin receptor action and resistance in obesity. J. Investig. Med., 57,
789-94.
Myers MG, Cowley MA & Münzberg H (2008). Mechanisms of leptin action and leptin resistance.
Annu. Rev. Physiol., 70, 537-56.
Morris DL & Rui L (2009). Recent advances in understanding leptin signaling and leptin resistance.
Am. J. Physiol. Endocrinol. Metab., 297, E1247-59.
Edit history
Date Action Author
2012-11-15 Authored May B
2012-11-15 Created May B
2012-11-24 Edited May B
2013-08-31 Reviewed Scherer T
2013-10-26 Reviewed Gonzalez-Perez RR
2018-08-30 Modified Croft D
Entities found in this pathway (6)
Input UniProt Id #Fold cha...
IRS2 Q9Y4H2 4.46
IRS1 P35568 1.66
LEP P41159 9.44
STAT3 P40763 1.53
STAT5A P42229 1.5
JAK2 O60674 1.75

https://reactome.org Page 32
RUNX3 regulates YAP1-mediated transcription (R-HSA-8951671)11.
cytosol
nucleoplasm
\\
RUNX3
BCL2L11
\\
CBFB
HES1
RUNX3:CBFB
\\ZFHX3
\\
6
Ub
RUNX3:ZFHX3
Dimeric TGFB1
ITGAL gene,(ITGA4gene)
26S proteasome
\\
YAP1:TEAD1,TEAD4,(TEAD2,YAP1:TEAD1,TEAD4,(TEAD2,YAP1:TEAD1,TEAD4,(TEAD2,TEAD3)TEAD3)TEAD3)
CDKN1A gene
RUNX3:YAP1:TEAD1,TEAD4,RUNX3:YAP1:TEAD1,TEAD4,RUNX3:YAP1:TEAD1,TEAD4,(TEAD2,TEAD3)(TEAD2,TEAD3)(TEAD2,TEAD3)
CDKN1A
p-2S-SMAD3:p-2S-SMAD3:SMAD4
RUNX3:p-2S-SMAD3:p-2S-SMAD3:SMAD4
Signaling by TGF-beta ReceptorComplex
ITGAL,(ITGA4)
FOXO3
RUNX1:CBFB,(Ac-K94,K171-RUNX3:CBFB:EP300:BRD2)
CDKN2A gene
RUNX3:p-2S-SMAD3:p-2S-RUNX3:p-2S-SMAD3:p-2S-RUNX3:p-2S-SMAD3:p-2S-SMAD3:SMAD4:CDKN1A geneSMAD3:SMAD4:CDKN1A geneSMAD3:SMAD4:CDKN1A gene
RUNX1:CBFB,(Ac-K94,K171-RUNX3:CBFB:EP300:BRD2):
CDKN2A gene
\\
BRD2homodimer
\\
CTNNB1:TCF7L2,(LEF1,TCF7L1,TCF7)
RUNX3:CTNNB1:TCF7L2,(LEF1,TCF7L1,TCF7)
Signaling by NOTCH1
\\
EP300
TEADs:YAP1TEADs:YAP1TEADs:YAP1
CTGF gene
RUNX3:CBFB:EP300
TEADs:YAP1:CTGFTEADs:YAP1:CTGFTEADs:YAP1:CTGFgenegenegene
3
PolyUb-RUNX3
\\
MyrG-p-Y419-SRC
P
p14-ARF mRNA
SMURF
RUNX3
MyrG-p-Y419-SRC:RUNX3
EP300
\\
CTGF
TEAD:WWTR1(TAZ)TEAD:WWTR1(TAZ)TEAD:WWTR1(TAZ)
JAG1 gene
RUNX3:JAG1 geneRUNX3:JAG1 geneRUNX3:JAG1 gene\\ RORC-2
TP53 Tetramer
CCND1 gene
CCND1
RUNX3:TP53 tetramer
CTNNB1:TCF7L2,LEF1:CCND1 Gene
ATP
p-Y-RUNX3P
CCND1
Ac-K94,K171-RUNX3:CBFB:BRD2:CCND1
\\
ADP
2
2
Ac-CoA
Ac-K94,K171-RUNX3:CBFB:EP300
CTNNB1:TCF7L2,LEF1
CoA-SH
p-S166,S188-MDM2p-S166,S188-MDM2p-S166,S188-MDM2dimerdimerdimer
\\
RUNX3:p-S166,S188-MDM2RUNX3:p-S166,S188-MDM2RUNX3:p-S166,S188-MDM2dimerdimerdimer
JAG1
6Ub
PolyUb-K94,K148-RUNX3
p14-ARF
\\
SPP1 gene
SPP1
MYC
RUNX3:CTNNB1:TCF7L2,(LEF1)
MYC gene
TCF7L1/TCF7L2/LEF1:CTNNB1
TCF7L1/TCF7L2/LEF1:TCF7L1/TCF7L2/LEF1:TCF7L1/TCF7L2/LEF1:CTNNB1:MYC geneCTNNB1:MYC geneCTNNB1:MYC gene
NOTCH1NOTCH1NOTCH1CoactivatorCoactivatorCoactivator
ComplexComplexComplex
RUNX3:NOTCH1RUNX3:NOTCH1RUNX3:NOTCH1Coactivator ComplexCoactivator ComplexCoactivator Complex
RORC gene
RUNX1 gene
RUNX3:CBFB:RORCgene
RUNX3:CBFB:RUNX1 gene
HDAC4
Ac-K94,K171-RUNX3:CBFB:BRD2:CCND1:HDAC4
RUNX3:CBFB:ITGAL gene,(ITGA4
gene)
\\
RUNX1 mRNA
\\PolyUb-K94,K148-RUNX3
2
2
RUNX3:CBFB:CCND1:HDAC4
H2O
CH3COO-
\\
HES1 gene
RUNX3:NOTCH1 coactivatorRUNX3:NOTCH1 coactivatorRUNX3:NOTCH1 coactivatorcomplex:HES1 genecomplex:HES1 genecomplex:HES1 gene
RUNX3:TCF7L2,(LEF1,TCF7L1)
BCL2L11 gene
RUNX3:p-2S-SMAD3:p-2S-RUNX3:p-2S-SMAD3:p-2S-RUNX3:p-2S-SMAD3:p-2S-SMAD3:SMAD4:FOXO3:BCL2L11SMAD3:SMAD4:FOXO3:BCL2L11SMAD3:SMAD4:FOXO3:BCL2L11
genegenegene
Ac-K94,K171-RUNX3:CBFB:EP300:BRD2
NOTCH1 CoactivatorNOTCH1 CoactivatorNOTCH1 CoactivatorComplex:HES1 GeneComplex:HES1 GeneComplex:HES1 Gene
KRAS:GTP
1.4E2
-1.96E1
Association of RUNX3 with the TEADs:YAP1 complex inhibits loading of the TEADs:YAP1 to the CT-GF promoter, thus negatively regulating transcription of the CTGF gene which encodes the Con-nective tissue growth factor (Yagi et al. 1999, Zhao et al. 2008, Qiao et al. 2016).
References
Yagi R, Chen LF, Shigesada K, Murakami Y & Ito Y (1999). A WW domain-containing yes-associated
protein (YAP) is a novel transcriptional co-activator. EMBO J., 18, 2551-62.
Qiao Y, Lin SJ, Chen Y, Voon DC, Zhu F, Chuang LS, ... Ito Y (2016). RUNX3 is a novel negative regu-
lator of oncogenic TEAD-YAP complex in gastric cancer. Oncogene, 35, 2664-74.
Zhao B, Ye X, Yu J, Li L, Li W, Li S, ... Guan KL (2008). TEAD mediates YAP-dependent gene induc-
tion and growth control. Genes Dev., 22, 1962-71.
Edit history
Date Action Author
2016-12-08 Created Orlic-Milacic M
2016-12-13 Authored Orlic-Milacic M
2017-01-31 Edited Orlic-Milacic M
2017-01-31 Reviewed Ito Y, Chuang LS
2018-08-23 Modified Schmidt EE
Entities found in this pathway (4)
Input UniProt Id #Fold cha...
CTGF P29279 2.56
TEAD1 P28347 1.64
TEAD2 Q15562 -1.66e+00

https://reactome.org Page 33
Input UniProt Id #Fold cha...
TEAD3 Q99594 1.53
Input Ensembl Id #Fold cha...
CTGF ENSG00000118523 2.56

https://reactome.org Page 34
Signaling by MET (R-HSA-6806834)12.
cytosol
nucleoplasm
recycling endosome membrane
ATP
HGF:p-4Y-MET dimer:p-Y317-SHC1-HGF:p-4Y-MET dimer:p-Y317-SHC1-HGF:p-4Y-MET dimer:p-Y317-SHC1-222
ADP
SPINT1,2SPINT1,2SPINT1,2
HGF:p-4Y-MET dimer:p-Y317-HGF:p-4Y-MET dimer:p-Y317-HGF:p-4Y-MET dimer:p-Y317-SHC1-2:GRB2:SOS1SHC1-2:GRB2:SOS1SHC1-2:GRB2:SOS1
SPINT1
CIN85:endophilinCIN85:endophilinCIN85:endophilin
HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):
CIN85:endophillinCIN85:endophillinCIN85:endophillin
pro-HGF
H2O
GAB1
HGF dimerHGF dimerHGF dimer
HGF:p-4Y-MET dimer:HGF:p-4Y-MET dimer:HGF:p-4Y-MET dimer:GAB1GAB1GAB1
HGFAC dimer
METHGF dimer:METHGF dimer:METHGF dimer:MET
GRB2-1 HGF:p-4Y-MET dimer:GRB2-1:HGF:p-4Y-MET dimer:GRB2-1:HGF:p-4Y-MET dimer:GRB2-1:GAB1GAB1GAB1
HGF:p-4Y-MET dimer:GAB1,HGF:p-4Y-MET dimer:GAB1,HGF:p-4Y-MET dimer:GAB1,HGF:p-4Y-MET dimer:GRB2:HGF:p-4Y-MET dimer:GRB2:HGF:p-4Y-MET dimer:GRB2:
GAB1GAB1GAB1
ATP
ADP
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1GRB2-1:p-5Y-GAB1GRB2-1:p-5Y-GAB1
STAT3
HGF:p-4Y-MET dimer:STAT3HGF:p-4Y-MET dimer:STAT3HGF:p-4Y-MET dimer:STAT3
ATP
ADP
p-Y705-STAT3P
2
p-Y705-STAT3 dimerp-Y705-STAT3 dimerp-Y705-STAT3 dimer
PIK3CA:PIK3R1PIK3CA:PIK3R1PIK3CA:PIK3R1
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:PI3KGRB2-1:p-5Y-GAB1:PI3KGRB2-1:p-5Y-GAB1:PI3K
\\
2
p-Y705-STAT3 dimerp-Y705-STAT3 dimerp-Y705-STAT3 dimer
HGF:METHGF:METHGF:METdimerdimerdimer
PI(4,5)P2
ATP
PI(3,4,5)P3
HGF:MonoUb-K,p-4Y-METHGF:MonoUb-K,p-4Y-METHGF:MonoUb-K,p-4Y-METdimer:GRB2:p-Y-CBL,(HGF:dimer:GRB2:p-Y-CBL,(HGF:dimer:GRB2:p-Y-CBL,(HGF:
MonoUb-K,p-Y1003,4Y-MonoUb-K,p-Y1003,4Y-MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):CIN85:METdimer:p-Y-CBL):CIN85:METdimer:p-Y-CBL):CIN85:endophillin,(MET:Ub-LRIG1)endophillin,(MET:Ub-LRIG1)endophillin,(MET:Ub-LRIG1)
ADP
EPS15:HGS:STAM
HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):
CIN85:endophillin:EPS15:HGS:STAM,(MET:Ub-LRIG1:CIN85:endophillin:EPS15:HGS:STAM,(MET:Ub-LRIG1:CIN85:endophillin:EPS15:HGS:STAM,(MET:Ub-LRIG1:EPS15:HGS:STAM)EPS15:HGS:STAM)EPS15:HGS:STAM)
PIP3 activates AKT signaling
TNS3HGF:p-4Y-MET dimer:TNS3HGF:p-4Y-MET dimer:TNS3HGF:p-4Y-MET dimer:TNS3
TNS4:ITGB1
HGF:p-4Y-MET dimer:TNS4:ITGB1HGF:p-4Y-MET dimer:TNS4:ITGB1HGF:p-4Y-MET dimer:TNS4:ITGB1
8 8
RAF/MAP kinasecascade
ATP
HGF:p-Y1234,Y1235,Y1349,Y1356-MET dimerHGF:p-Y1234,Y1235,Y1349,Y1356-MET dimerHGF:p-Y1234,Y1235,Y1349,Y1356-MET dimer
CRK,CRKL
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:CRK,CRKLGRB2-1:p-5Y-GAB1:CRK,CRKLGRB2-1:p-5Y-GAB1:CRK,CRKL
PTPN11
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:PTPN11p-5Y-GAB1:PTPN11p-5Y-GAB1:PTPN11
RAPGEF1HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:CRK,CRKL:RAPGEF1GRB2-1:p-5Y-GAB1:CRK,CRKL:RAPGEF1GRB2-1:p-5Y-GAB1:CRK,CRKL:RAPGEF1
2 2
ADP
H2O
CBL:GRB2,CBL
Pi
PTK2
HGF:p-4Y-MET dimer:PTK2HGF:p-4Y-MET dimer:PTK2HGF:p-4Y-MET dimer:PTK2
HGF:p-Y1234,Y1235,Y1356-METHGF:p-Y1234,Y1235,Y1356-METHGF:p-Y1234,Y1235,Y1356-METdimerdimerdimer
PTPRJ
HGF:p-4Y-MET dimer:GRB2:HGF:p-4Y-MET dimer:GRB2:HGF:p-4Y-MET dimer:GRB2:CBL,(HGF:p-Y1003,4Y-METCBL,(HGF:p-Y1003,4Y-METCBL,(HGF:p-Y1003,4Y-MET
dimer:CBL)dimer:CBL)dimer:CBL)
ATP
ADP
HGF:p-4Y-MET dimer:GRB2:p-Y-HGF:p-4Y-MET dimer:GRB2:p-Y-HGF:p-4Y-MET dimer:GRB2:p-Y-CBL,(HGF:p-Y1003,4Y-MET dimer:p-CBL,(HGF:p-Y1003,4Y-MET dimer:p-CBL,(HGF:p-Y1003,4Y-MET dimer:p-
Y-CBL)Y-CBL)Y-CBL)
ATP
HGF:p-4Y-MET dimer:p-Y397-PTK2HGF:p-4Y-MET dimer:p-Y397-PTK2HGF:p-4Y-MET dimer:p-Y397-PTK2
ADP
RAP1:GDP
HGF:MonoUb-K,p-4Y-MET dimer:HGF:MonoUb-K,p-4Y-MET dimer:HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,(HGF:MonoUb-K,p-GRB2:p-Y-CBL,(HGF:MonoUb-K,p-GRB2:p-Y-CBL,(HGF:MonoUb-K,p-
Y1003,4Y-MET dimer:p-Y-CBL)Y1003,4Y-MET dimer:p-Y-CBL)Y1003,4Y-MET dimer:p-Y-CBL)
RAP1:GTP
GTP
Integrin alpha2beta1,alpha3:beta1:(collagen,laminin,Integrin alpha2beta1,alpha3:beta1:(collagen,laminin,Integrin alpha2beta1,alpha3:beta1:(collagen,laminin,fibronectin)fibronectin)fibronectin)
GDP
MyrG-p-Y419-SRC
P
4 4
HGF:p-4Y-MET dimer:p-Y397-PTK2:HGF:p-4Y-MET dimer:p-Y397-PTK2:HGF:p-4Y-MET dimer:p-Y397-PTK2:MyrG-p-Y419-SRCMyrG-p-Y419-SRCMyrG-p-Y419-SRC
HGF:p-Y,1349,Y1356-MET dimerHGF:p-Y,1349,Y1356-MET dimerHGF:p-Y,1349,Y1356-MET dimer
PTPN1,PTPN2
HPN heterodimer
ATP
ADP
HGF:p-4Y-MET dimer:p-Y194,Y397-HGF:p-4Y-MET dimer:p-Y194,Y397-HGF:p-4Y-MET dimer:p-Y194,Y397-PTK2:MyrG-p-Y419-SRCPTK2:MyrG-p-Y419-SRCPTK2:MyrG-p-Y419-SRC
GRB2-1:SOS1
HGF:p-4Y-MET dimer:GRB2-1:HGF:p-4Y-MET dimer:GRB2-1:HGF:p-4Y-MET dimer:GRB2-1:SOS1SOS1SOS1
?(DOCK7)
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:CRK,CRKL:(DOCK7)p-5Y-GAB1:CRK,CRKL:(DOCK7)p-5Y-GAB1:CRK,CRKL:(DOCK7)
2
2
p21 RAS:GDP
ATP
GTP
ADP
HGF:p-4Y-MET dimer:p-Y194,Y397,HGF:p-4Y-MET dimer:p-Y194,Y397,HGF:p-4Y-MET dimer:p-Y194,Y397,Y576,Y577-PTK2:MyrG-p-Y419-SRCY576,Y577-PTK2:MyrG-p-Y419-SRCY576,Y577-PTK2:MyrG-p-Y419-SRC
p21 RAS:GTP
GDP
?RAC1:GDP
HGF:p-4Y-MET dimer,(HGF:p-Y1003,4Y-HGF:p-4Y-MET dimer,(HGF:p-Y1003,4Y-HGF:p-4Y-MET dimer,(HGF:p-Y1003,4Y-METdimer)METdimer)METdimer)
RAC1:GTP
GDPGTP
MUC20 HGF:p-4Y MET dimer:MUC20HGF:p-4Y MET dimer:MUC20HGF:p-4Y MET dimer:MUC20
Ub
LRIG1
MET:MET:MET:LRIG1LRIG1LRIG1
SOS1
RANBP9
HGF:p-4Y-MET dimer:RANBP9:HGF:p-4Y-MET dimer:RANBP9:HGF:p-4Y-MET dimer:RANBP9:SOS1SOS1SOS1
HGF:p-4Y-MET dimer:RANBP10HGF:p-4Y-MET dimer:RANBP10HGF:p-4Y-MET dimer:RANBP10
?
ARF6:GTP
MET:Ub-MET:Ub-MET:Ub-LRIG1LRIG1LRIG1
GGA3
unknown ubiquitin ligase
HGF:p-4Y-MET:p-5Y-GAB1,HGF:p-4Y-MET:p-5Y-GAB1,HGF:p-4Y-MET:p-5Y-GAB1,HGF:p-4Y-MET:GRB2-1:p-5Y-HGF:p-4Y-MET:GRB2-1:p-5Y-HGF:p-4Y-MET:GRB2-1:p-5Y-
GAB1:CRK,CRKL:GGA3:GAB1:CRK,CRKL:GGA3:GAB1:CRK,CRKL:GGA3:ARF6:GTPARF6:GTPARF6:GTP
Ub
RANBP10\\
GGC-RAB4:GTP
H2O
USP8
SHC1-2
HGF:p-4Y-MET dimer:SHC1-2HGF:p-4Y-MET dimer:SHC1-2HGF:p-4Y-MET dimer:SHC1-2
1.4E2
-1.96E1
MET is a receptor tyrosine kinase (RTK) (Cooper et al. 1984, Park et al. 1984) activated by binding to its ligand, Hepatocyte growth factor/Scatter factor (HGF/SF) (Bottaro et al. 1991, Naldini et al. 1991). Similar to other related RTKs, such as EGFR, ligand binding induces MET dimerization and trans-autophosphorylation, resulting in the active MET receptor complex (Ferracini et al. 1991, Longati et al. 1994, Rodrigues and Park 1994, Kirchhofer et al. 2004, Stamos et al. 2004, Hays and Watowich 2004). Phosphorylated tyrosines in the cytoplasmic tail of MET serve as docking sites for binding of adapter proteins, such as GRB2, SHC1 and GAB1, which trigger signal transduction cascades that activate PI3K/AKT, RAS, STAT3, PTK2, RAC1 and RAP1 signaling (Ponzetto et al. 1994, Pelicci et al. 1995, Weidner et al. 1995, Besser et al. 1997, Shen and Novak 1997, Beviglia and Kramer 1999, Rodrigues et al. 2000, Sakkab et al. 2000, Schaeper et al. 2000, Lamorte et al. 2002, Wang et al. 2002, Chen and Chen 2006, Palamidessi et al. 2008, Chen et al. 2011, Murray et al. 2014).
Activation of PLC gamma 1 (PLCG1) signaling by MET remains unclear. It has been reported that PLCG1 can bind to MET directly (Ponzetto et al. 1994) or be recruited by phosphorylated GAB1 (Gual et al. 2000). Tyrosine residue Y307 of GAB1 that serves as docking sites for PLCG1 may be phosphorylated either by activated MET (Watanabe et al. 2006) or SRC (Chan et al. 2010). Another PCLG1 docking site on GAB1, tyrosine residue Y373, was reported as the SRC target, while the kinase for the main PLCG1 docking site, Y407 of GAB1, is not known (Chan et al. 2010).
Signaling by MET promotes cell growth, cell survival and motility, which are essential for embryon-ic development (Weidner et al. 1993, Schmidt et al. 1995, Uehara et al. 1995, Bladt et al. 1995, Maina et al. 1997, Maina et al. 2001, Helmbacher et al. 2003) and tissue regeneration (Huh et al. 2004, Borowiak et al. 2004, Liu 2004, Chmielowiec et al. 2007). MET signaling is frequently aberrantly ac-tivated in cancer, through MET overexpression or activating MET mutations (Schmidt et al. 1997, Pennacchietti et al. 2003, Smolen et al. 2006, Bertotti et al. 2009).

https://reactome.org Page 35
Considerable progress has recently been made in the development of HGF-MET inhibitors in can-cer therapy. These include inhibitors of HGF activators, HGF inhibitors and MET antagonists, which are protein therapeutics that act outside the cell. Kinase inhibitors function inside the cell and have constituted the largest effort towards MET-based therapeutics (Gherardi et al. 2012).
Pathogenic bacteria of the species Listeria monocytogenes, exploit MET receptor as an entryway to host cells (Shen et al. 2000, Veiga and Cossart 2005, Neimann et al. 2007).
For review of MET signaling, please refer to Birchmeier et al. 2003, Trusolino et al. 2010, Gherardi et al. 2012, Petrini 2015.
References
Ponzetto C, Bardelli A, Zhen Z, Maina F, dalla Zonca P, Giordano S, ... Comoglio PM (1994). A multi-functional docking site mediates signaling and transformation by the hepatocyte growth
factor/scatter factor receptor family. Cell, 77, 261-71.
Gual P, Giordano S, Williams TA, Rocchi S, Van Obberghen E & Comoglio PM (2000). Sustained re-cruitment of phospholipase C-gamma to Gab1 is required for HGF-induced branching tubulo-
genesis. Oncogene, 19, 1509-18.
Chan PC, Sudhakar JN, Lai CC & Chen HC (2010). Differential phosphorylation of the docking pro-tein Gab1 by c-Src and the hepatocyte growth factor receptor regulates different aspects of cell
functions. Oncogene, 29, 698-710.
Ferracini R, Longati P, Naldini L, Vigna E & Comoglio PM (1991). Identification of the major auto-phosphorylation site of the Met/hepatocyte growth factor receptor tyrosine kinase. J. Biol.
Chem., 266, 19558-64.
Longati P, Bardelli A, Ponzetto C, Naldini L & Comoglio PM (1994). Tyrosines1234-1235 are critical for activation of the tyrosine kinase encoded by the MET proto-oncogene (HGF receptor). Onco-
gene, 9, 49-57.
Edit history
Date Action Author
2015-10-27 Created Orlic-Milacic M
2016-06-14 Edited Orlic-Milacic M
2016-06-14 Authored Orlic-Milacic M
2016-07-11 Reviewed Heynen G, Birchmeier W
2018-08-23 Modified Schmidt EE
Entities found in this pathway (18)
Input UniProt Id #Fold cha...
LAMC1 P11047 1.73
LAMB1 P07942 1.69
PIK3R1 P27986 3.08
PIK3CA P42336 1.51
LAMA3 Q16787 1.82
COL5A3 P25940 1.9
COL11A1 P12107 5.14
LAMA2 P24043 4.19

https://reactome.org Page 36
Input UniProt Id #Fold cha...
ITGA2 P17301 -2.06e+00
SH3GL3 Q99963 -5.36e+00
RANBP9 Q96S59 1.66
HGF P14210 -1.98e+00
STAT3 P40763 1.53
COL1A1 P02452 -2.48e+00
COL3A1 P02461 -1.81e+00
COL27A1 Q8IZC6 -1.98e+00
LIG1 Q96JA1 -2.02e+00
SPINT2 O43291 3.45

https://reactome.org Page 37
Transcriptional activation of p53 responsive genes (R-HSA-69560)13.
cytosol
nucleoplasm
ATP
CDKN1A,CDKN1A,CDKN1A,CDKN1BCDKN1BCDKN1B
ADP
CCNE:CDK2
Cyclin E:CDK2:CDKN1A,Cyclin E:CDK2:CDKN1A,Cyclin E:CDK2:CDKN1A,CDKN1BCDKN1BCDKN1B
CDKN1A
TP53 Tetramer
ADP
ATP
6Ub
PolyUb,p-S166,S188-MDM2:PolyUb,p-S166,S188-MDM2:PolyUb,p-S166,S188-MDM2:PolyUb,p-S342,S367,S403-MDM4PolyUb,p-S342,S367,S403-MDM4PolyUb,p-S342,S367,S403-MDM4
p-S15-TP53Tetramer
p-S387-RFWD2P
p-S387-RFWD2
P
26S proteasome
Ub
CDKN1A gene
RFWD2
ubiquitinated phospho-COP1(ser-387)
p-S387-RFWD2:p-S15-TP53
PKMYT1
14-3-3 dimer
\\
4
DNA DNA DSBs:p-MRN:p-S1981,Ac-K3016-ATM:KAT5:K63PolyUb-K14,K16,p-S139-H2AFX,DNA DNA DSBs:p-MRN:p-S1981,Ac-K3016-ATM:KAT5:K63PolyUb-K14,K16,p-S139-H2AFX,DNA DNA DSBs:p-MRN:p-S1981,Ac-K3016-ATM:KAT5:K63PolyUb-K14,K16,p-S139-H2AFX,Me2K21-HIST1H4A-Nucleosome:p-5T-MDC1:p-S102-WHSC1:RNF8:Zn2+:SUMO1:p-T4827-Me2K21-HIST1H4A-Nucleosome:p-5T-MDC1:p-S102-WHSC1:RNF8:Zn2+:SUMO1:p-T4827-Me2K21-HIST1H4A-Nucleosome:p-5T-MDC1:p-S102-WHSC1:RNF8:Zn2+:SUMO1:p-T4827-HERC2:UBE2N:UBE2V2:RNF168:PIAS4:p-S25,S1778-TP53BP1:p-4S,2T-BRCA1-A complex:HERC2:UBE2N:UBE2V2:RNF168:PIAS4:p-S25,S1778-TP53BP1:p-4S,2T-BRCA1-A complex:HERC2:UBE2N:UBE2V2:RNF168:PIAS4:p-S25,S1778-TP53BP1:p-4S,2T-BRCA1-A complex:
CHEK2CHEK2CHEK2
ATP ADP
DNA DNA DSBs:p-MRN:p-S1981,Ac-K3016-ATM:KAT5:K63PolyUb-K14,K16,p-DNA DNA DSBs:p-MRN:p-S1981,Ac-K3016-ATM:KAT5:K63PolyUb-K14,K16,p-DNA DNA DSBs:p-MRN:p-S1981,Ac-K3016-ATM:KAT5:K63PolyUb-K14,K16,p-S139-H2AFX,Me2K21-HIST1H4A-Nucleosome:p-5T-MDC1:p-S102-WHSC1:S139-H2AFX,Me2K21-HIST1H4A-Nucleosome:p-5T-MDC1:p-S102-WHSC1:S139-H2AFX,Me2K21-HIST1H4A-Nucleosome:p-5T-MDC1:p-S102-WHSC1:
RNF8:Zn2+:SUMO1:p-T4827-HERC2:UBE2N:UBE2V2:RNF168:PIAS4:p-S25,RNF8:Zn2+:SUMO1:p-T4827-HERC2:UBE2N:UBE2V2:RNF168:PIAS4:p-S25,RNF8:Zn2+:SUMO1:p-T4827-HERC2:UBE2N:UBE2V2:RNF168:PIAS4:p-S25,S1778-TP53BP1:p-4S,2T-BRCA1-A complexS1778-TP53BP1:p-4S,2T-BRCA1-A complexS1778-TP53BP1:p-4S,2T-BRCA1-A complex
4
\\
Ub-p-S123-CDC25A
p-S123-CDC25AP
Ubiquitinligase
2
p-S166,S188-MDM2p-S166,S188-MDM2p-S166,S188-MDM2dimer, p-S166,S188-dimer, p-S166,S188-dimer, p-S166,S188-MDM2,MDM4:TP53MDM2,MDM4:TP53MDM2,MDM4:TP53
p-T68-CHEK2 dimer
33
2
PolyUb-TP53Tetramer
ATP
ADP
Ub
p-S,3T-CHEK2
P P P P
CDC25A p-S123-CDC25AP
DNA Double Strand Break Response
p-S1981,Ac-K3016-ATM
P Ac
44
ATP
ADP
p-S166,S188-MDM2:p-S166,S188-MDM2:p-S166,S188-MDM2:MDM4MDM4MDM4
ATP
ADP
p-S166,S188-MDM2:p-p-S166,S188-MDM2:p-p-S166,S188-MDM2:p-S403-MDM4S403-MDM4S403-MDM4
PolyUb-TP53Tetramer
4
4
ATP
ADP
p-S15,S20-TP53 Tetramer
ZNF385A
p-S15,S20-TP53 Tetramer:p-S15,S20-TP53 Tetramer:p-S15,S20-TP53 Tetramer:ZNF385AZNF385AZNF385A
p-S15,S20-TP53 Tetramer:p-S15,S20-TP53 Tetramer:p-S15,S20-TP53 Tetramer:ZNF385A:CDKN1A GeneZNF385A:CDKN1A GeneZNF385A:CDKN1A Gene
p-S435-GTSE1P
\\
p-S435-GTSE1:PolyUb-TP53 Tetramer
CDKN1A mRNA
p-S435-GTSE1:PolyUb-TP53 Tetramer
ATP
\\
PCBP4:CDKN1A mRNAPCBP4:CDKN1A mRNAPCBP4:CDKN1A mRNA
CHEK1
\\
p-S317,S345-CHEK1
P P
PCBP4
ADP
p-S435-GTSE1
P
ATR:ATRIP
12
CHEK2
p-T68-CHEK2P
PHF20
p14-ARF:p-S166,S188-MDM2p14-ARF:p-S166,S188-MDM2p14-ARF:p-S166,S188-MDM2dimer,p-S166,S188-MDM2:dimer,p-S166,S188-MDM2:dimer,p-S166,S188-MDM2:
MDM4MDM4MDM4
ATP
CDC25C p-S216-CDC25C
P
ADP
ADP
p-S216-CDC25C
P
ATPp-S216-CDC25C:14-3-3protein complex
ATP
WEE1
p-WEE1
P
ADP
CCNA:CCNA:CCNA:CDK2CDK2CDK2
Cyclin A:Cdk2:p21/p27Cyclin A:Cdk2:p21/p27Cyclin A:Cdk2:p21/p27complexcomplexcomplex
ATP
CCNB:CDK1
ATP
CCNB:p-T14-CDK1
ADP
RPA heterotrimer
Persistent single-strandedDNA
RPA complexed to ssDNA
ATR:ATRIP:RPA:ssDNAsignaling complex
RAD17:RFC
Rad17-RFC complex boundto DNA
RAD9:HUS1:RAD1RAD9:HUS1:RAD1RAD9:HUS1:RAD1
Rad9-Hus1-Rad1 bound toRad9-Hus1-Rad1 bound toRad9-Hus1-Rad1 bound toDNADNADNA
CLSPNCDK:DDK:MCM10:active pre-
replicative complex:CDC45
Cdc45:CDK:DDK:Mcm10:claspin:pre-replicative complex
2
2
4 4
ATP
Cdc45:CDK:DDK:Mcm10:Activated claspin:pre-
replicative complex
ADP
ATR:ATRIP:RPA:3' overhangingATR:ATRIP:RPA:3' overhangingATR:ATRIP:RPA:3' overhangingssDNA-DSBs:p-MRN:p-S1981,Ac-ssDNA-DSBs:p-MRN:p-S1981,Ac-ssDNA-DSBs:p-MRN:p-S1981,Ac-
K3016-ATM:KAT5:BRCA1-CK3016-ATM:KAT5:BRCA1-CK3016-ATM:KAT5:BRCA1-Ccomplex:EXO1,DNA2:BLM,WRN:complex:EXO1,DNA2:BLM,WRN:complex:EXO1,DNA2:BLM,WRN:p-S990,Ac-K1249-BRIP1:RAD17:p-S990,Ac-K1249-BRIP1:RAD17:p-S990,Ac-K1249-BRIP1:RAD17:
RFC:RAD9:HUS1:RAD1:RHNO1:RFC:RAD9:HUS1:RAD1:RHNO1:RFC:RAD9:HUS1:RAD1:RHNO1:TOPBP1TOPBP1TOPBP1
4
4
ATP
Ub
ADP
Kinetochore
ATP
CCNB1:p-T14,T161-CDK1
CCNB1:p-T14,Y15,T161-CDK1
ADP
4
4
p-S166,S188-MDM2 PP
ATP
ADP
ADP
p-5S,T-MDM2
P PP
P
P
P
2
2
ATP
ADP
p-S166,S188-MDM2:p-S346,p-S166,S188-MDM2:p-S346,p-S166,S188-MDM2:p-S346,S367,S403-MDM4S367,S403-MDM4S367,S403-MDM4
MAD1L1
Mad1:kinetochore complex
MAD2L1
Kinetochore:Mad1:MAD2Complex
Kinetochore:Mad1:MAD2*Complex
ActivatedMAD2L1
CDC20
MAD2*CDC20 complex
BUB3
BUB1B
hBUBR1:hBUB3:MAD2*:CDC20 complex
2
p-S166,S188-MDM2,p-S166,S188-MDM2,p-S166,S188-MDM2,MDM4MDM4MDM4
p-S166,S188-MDM2 dimer, p-S166,p-S166,S188-MDM2 dimer, p-S166,p-S166,S188-MDM2 dimer, p-S166,S188-MDM2:MDM4S188-MDM2:MDM4S188-MDM2:MDM4
phosphorylated anaphasepromoting complex (APC/C)
MCC:APC/Ccomplex
\\
1.4E2
-1.96E1
p53 causes G1 arrest by inducing the expression of a cell cycle inhibitor, p21 (El-Deiry et al, 1993; Harper et al, 1993; Xiong et al, 1993). P21 binds and inactivates Cyclin-Cdk complexes that mediate G1/S progression, resulting in lack of phosphorylation of Rb, E2F sequestration and cell cycle arrest at the G1/S transition. Mice with a homozygous deletion of p21 gene are deficient in their ability to undergo a G1/S arrest in response to DNA damage (Deng et al, 1995).
References
el-Deiry WS, Tokino T, Velculescu VE, Levy DB, Parsons R, Trent JM, ... Vogelstein B (1993). WAF1,
a potential mediator of p53 tumor suppression. Cell, 75, 817-25.
Harper JW, Adami GR, Wei N, Keyomarsi K & Elledge SJ (1993). The p21 Cdk-interacting protein
Cip1 is a potent inhibitor of G1 cyclin-dependent kinases. Cell, 75, 805-16.
Xiong Y, Hannon GJ, Zhang H, Casso D, Kobayashi R & Beach D (1994). p21 is a universal inhibitor
of cyclin kinases. Nature, 366, 701-4.
Deng C, Zhang P, Harper JW, Elledge SJ & Leder P (1995). Mice lacking p21CIP1/WAF1 undergo nor-
mal development, but are defective in G1 checkpoint control. Cell, 82, 675-84.
Edit history
Date Action Author
2003-06-05 Created Khanna KK
2018-08-23 Modified Schmidt EE
Entities found in this pathway (2)
Input UniProt Id #Fold cha...
CDKN1A P38936 -2.01e+00
ZNF385A Q96PM9 -1.76e+00
Input Ensembl Id #Fold cha...
CDKN1A ENSG00000124762, ENST00000244741 -2.01e+00

https://reactome.org Page 38
Transcriptional activation of cell cycle inhibitor p21 (R-HSA-69895)14.
cytosol
nucleoplasm
ATP
CDKN1A,CDKN1A,CDKN1A,CDKN1BCDKN1BCDKN1B
ADP
CCNE:CDK2
Cyclin E:CDK2:CDKN1A,Cyclin E:CDK2:CDKN1A,Cyclin E:CDK2:CDKN1A,CDKN1BCDKN1BCDKN1B
CDKN1A
TP53 Tetramer
ADP
ATP
6Ub
PolyUb,p-S166,S188-MDM2:PolyUb,p-S166,S188-MDM2:PolyUb,p-S166,S188-MDM2:PolyUb,p-S342,S367,S403-MDM4PolyUb,p-S342,S367,S403-MDM4PolyUb,p-S342,S367,S403-MDM4
p-S15-TP53Tetramer
p-S387-RFWD2P
p-S387-RFWD2
P
26S proteasome
Ub
CDKN1A gene
RFWD2
ubiquitinated phospho-COP1(ser-387)
p-S387-RFWD2:p-S15-TP53
PKMYT1
14-3-3 dimer
\\
4
DNA DNA DSBs:p-MRN:p-S1981,Ac-K3016-ATM:KAT5:K63PolyUb-K14,K16,p-S139-H2AFX,DNA DNA DSBs:p-MRN:p-S1981,Ac-K3016-ATM:KAT5:K63PolyUb-K14,K16,p-S139-H2AFX,DNA DNA DSBs:p-MRN:p-S1981,Ac-K3016-ATM:KAT5:K63PolyUb-K14,K16,p-S139-H2AFX,Me2K21-HIST1H4A-Nucleosome:p-5T-MDC1:p-S102-WHSC1:RNF8:Zn2+:SUMO1:p-T4827-Me2K21-HIST1H4A-Nucleosome:p-5T-MDC1:p-S102-WHSC1:RNF8:Zn2+:SUMO1:p-T4827-Me2K21-HIST1H4A-Nucleosome:p-5T-MDC1:p-S102-WHSC1:RNF8:Zn2+:SUMO1:p-T4827-HERC2:UBE2N:UBE2V2:RNF168:PIAS4:p-S25,S1778-TP53BP1:p-4S,2T-BRCA1-A complex:HERC2:UBE2N:UBE2V2:RNF168:PIAS4:p-S25,S1778-TP53BP1:p-4S,2T-BRCA1-A complex:HERC2:UBE2N:UBE2V2:RNF168:PIAS4:p-S25,S1778-TP53BP1:p-4S,2T-BRCA1-A complex:
CHEK2CHEK2CHEK2
ATP ADP
DNA DNA DSBs:p-MRN:p-S1981,Ac-K3016-ATM:KAT5:K63PolyUb-K14,K16,p-DNA DNA DSBs:p-MRN:p-S1981,Ac-K3016-ATM:KAT5:K63PolyUb-K14,K16,p-DNA DNA DSBs:p-MRN:p-S1981,Ac-K3016-ATM:KAT5:K63PolyUb-K14,K16,p-S139-H2AFX,Me2K21-HIST1H4A-Nucleosome:p-5T-MDC1:p-S102-WHSC1:S139-H2AFX,Me2K21-HIST1H4A-Nucleosome:p-5T-MDC1:p-S102-WHSC1:S139-H2AFX,Me2K21-HIST1H4A-Nucleosome:p-5T-MDC1:p-S102-WHSC1:
RNF8:Zn2+:SUMO1:p-T4827-HERC2:UBE2N:UBE2V2:RNF168:PIAS4:p-S25,RNF8:Zn2+:SUMO1:p-T4827-HERC2:UBE2N:UBE2V2:RNF168:PIAS4:p-S25,RNF8:Zn2+:SUMO1:p-T4827-HERC2:UBE2N:UBE2V2:RNF168:PIAS4:p-S25,S1778-TP53BP1:p-4S,2T-BRCA1-A complexS1778-TP53BP1:p-4S,2T-BRCA1-A complexS1778-TP53BP1:p-4S,2T-BRCA1-A complex
4
\\
Ub-p-S123-CDC25A
p-S123-CDC25AP
Ubiquitinligase
2
p-S166,S188-MDM2p-S166,S188-MDM2p-S166,S188-MDM2dimer, p-S166,S188-dimer, p-S166,S188-dimer, p-S166,S188-MDM2,MDM4:TP53MDM2,MDM4:TP53MDM2,MDM4:TP53
p-T68-CHEK2 dimer
33
2
PolyUb-TP53Tetramer
ATP
ADP
Ub
p-S,3T-CHEK2
P P P P
CDC25A p-S123-CDC25AP
DNA Double Strand Break Response
p-S1981,Ac-K3016-ATM
P Ac
44
ATP
ADP
p-S166,S188-MDM2:p-S166,S188-MDM2:p-S166,S188-MDM2:MDM4MDM4MDM4
ATP
ADP
p-S166,S188-MDM2:p-p-S166,S188-MDM2:p-p-S166,S188-MDM2:p-S403-MDM4S403-MDM4S403-MDM4
PolyUb-TP53Tetramer
4
4
ATP
ADP
p-S15,S20-TP53 Tetramer
ZNF385A
p-S15,S20-TP53 Tetramer:p-S15,S20-TP53 Tetramer:p-S15,S20-TP53 Tetramer:ZNF385AZNF385AZNF385A
p-S15,S20-TP53 Tetramer:p-S15,S20-TP53 Tetramer:p-S15,S20-TP53 Tetramer:ZNF385A:CDKN1A GeneZNF385A:CDKN1A GeneZNF385A:CDKN1A Gene
p-S435-GTSE1P
\\
p-S435-GTSE1:PolyUb-TP53 Tetramer
CDKN1A mRNA
p-S435-GTSE1:PolyUb-TP53 Tetramer
ATP
\\
PCBP4:CDKN1A mRNAPCBP4:CDKN1A mRNAPCBP4:CDKN1A mRNA
CHEK1
\\
p-S317,S345-CHEK1
P P
PCBP4
ADP
p-S435-GTSE1
P
ATR:ATRIP
12
CHEK2
p-T68-CHEK2P
PHF20
p14-ARF:p-S166,S188-MDM2p14-ARF:p-S166,S188-MDM2p14-ARF:p-S166,S188-MDM2dimer,p-S166,S188-MDM2:dimer,p-S166,S188-MDM2:dimer,p-S166,S188-MDM2:
MDM4MDM4MDM4
ATP
CDC25C p-S216-CDC25C
P
ADP
ADP
p-S216-CDC25C
P
ATPp-S216-CDC25C:14-3-3protein complex
ATP
WEE1
p-WEE1
P
ADP
CCNA:CCNA:CCNA:CDK2CDK2CDK2
Cyclin A:Cdk2:p21/p27Cyclin A:Cdk2:p21/p27Cyclin A:Cdk2:p21/p27complexcomplexcomplex
ATP
CCNB:CDK1
ATP
CCNB:p-T14-CDK1
ADP
RPA heterotrimer
Persistent single-strandedDNA
RPA complexed to ssDNA
ATR:ATRIP:RPA:ssDNAsignaling complex
RAD17:RFC
Rad17-RFC complex boundto DNA
RAD9:HUS1:RAD1RAD9:HUS1:RAD1RAD9:HUS1:RAD1
Rad9-Hus1-Rad1 bound toRad9-Hus1-Rad1 bound toRad9-Hus1-Rad1 bound toDNADNADNA
CLSPNCDK:DDK:MCM10:active pre-
replicative complex:CDC45
Cdc45:CDK:DDK:Mcm10:claspin:pre-replicative complex
2
2
4 4
ATP
Cdc45:CDK:DDK:Mcm10:Activated claspin:pre-
replicative complex
ADP
ATR:ATRIP:RPA:3' overhangingATR:ATRIP:RPA:3' overhangingATR:ATRIP:RPA:3' overhangingssDNA-DSBs:p-MRN:p-S1981,Ac-ssDNA-DSBs:p-MRN:p-S1981,Ac-ssDNA-DSBs:p-MRN:p-S1981,Ac-
K3016-ATM:KAT5:BRCA1-CK3016-ATM:KAT5:BRCA1-CK3016-ATM:KAT5:BRCA1-Ccomplex:EXO1,DNA2:BLM,WRN:complex:EXO1,DNA2:BLM,WRN:complex:EXO1,DNA2:BLM,WRN:p-S990,Ac-K1249-BRIP1:RAD17:p-S990,Ac-K1249-BRIP1:RAD17:p-S990,Ac-K1249-BRIP1:RAD17:
RFC:RAD9:HUS1:RAD1:RHNO1:RFC:RAD9:HUS1:RAD1:RHNO1:RFC:RAD9:HUS1:RAD1:RHNO1:TOPBP1TOPBP1TOPBP1
4
4
ATP
Ub
ADP
Kinetochore
ATP
CCNB1:p-T14,T161-CDK1
CCNB1:p-T14,Y15,T161-CDK1
ADP
4
4
p-S166,S188-MDM2 PP
ATP
ADP
ADP
p-5S,T-MDM2
P PP
P
P
P
2
2
ATP
ADP
p-S166,S188-MDM2:p-S346,p-S166,S188-MDM2:p-S346,p-S166,S188-MDM2:p-S346,S367,S403-MDM4S367,S403-MDM4S367,S403-MDM4
MAD1L1
Mad1:kinetochore complex
MAD2L1
Kinetochore:Mad1:MAD2Complex
Kinetochore:Mad1:MAD2*Complex
ActivatedMAD2L1
CDC20
MAD2*CDC20 complex
BUB3
BUB1B
hBUBR1:hBUB3:MAD2*:CDC20 complex
2
p-S166,S188-MDM2,p-S166,S188-MDM2,p-S166,S188-MDM2,MDM4MDM4MDM4
p-S166,S188-MDM2 dimer, p-S166,p-S166,S188-MDM2 dimer, p-S166,p-S166,S188-MDM2 dimer, p-S166,S188-MDM2:MDM4S188-MDM2:MDM4S188-MDM2:MDM4
phosphorylated anaphasepromoting complex (APC/C)
MCC:APC/Ccomplex
\\
1.4E2
-1.96E1
Both p53-independent and p53-dependent mechanisms of induction of p21 mRNA have been demonstrated. p21 is transcriptionally activated by p53 after DNA damage (el-Deiry et al., 1993).
References
el-Deiry WS, Tokino T, Velculescu VE, Levy DB, Parsons R, Trent JM, ... Vogelstein B (1993). WAF1,
a potential mediator of p53 tumor suppression. Cell, 75, 817-25.
Edit history
Date Action Author
2003-06-05 Created Khanna KK
2018-08-30 Modified Croft D
Entities found in this pathway (2)
Input UniProt Id #Fold cha...
CDKN1A P38936 -2.01e+00
ZNF385A Q96PM9 -1.76e+00
Input Ensembl Id #Fold cha...
CDKN1A ENSG00000124762, ENST00000244741 -2.01e+00

https://reactome.org Page 39
Collagen biosynthesis and modifying enzymes (R-HSA-1650814)15.
cytosol
endoplasmic reticulum lumen
UDP
P4HB
Collagen propeptidesCollagen propeptidesCollagen propeptides
P4HB:CollagenP4HB:CollagenP4HB:Collagenpropeptidespropeptidespropeptides
Procollagen C-linkedtrimers
\\
2OG
3,4-Hyp collagen propeptides3,4-Hyp collagen propeptides3,4-Hyp collagen propeptides
O2
Collagen and procollagentriple helices:Serpin H1
CO2
4-Hyp collagen4-Hyp collagen4-Hyp collagenpropeptides:P4HBpropeptides:P4HBpropeptides:P4HB
LEPRE1:PPIB:CRTAP
SUCCA
Prolyl 4-hydroxylases
Prolyl 3-hydroxylases:Fe2+:3,4-HypProlyl 3-hydroxylases:Fe2+:3,4-HypProlyl 3-hydroxylases:Fe2+:3,4-Hypcollagen propeptidescollagen propeptidescollagen propeptides
Lysyl hydroxylases:LysylLysyl hydroxylases:LysylLysyl hydroxylases:Lysylhydroxylase procollagenhydroxylase procollagenhydroxylase procollagen
substratessubstratessubstrates
Lysyl hydroxylases:LysylLysyl hydroxylases:LysylLysyl hydroxylases:Lysylhydroxylated collagenhydroxylated collagenhydroxylated collagen
propeptidespropeptidespropeptides
Lysyl hydroxylases
2OG
CO2
SUCCA
O2
COLGALT1,COLGALT2:LysylCOLGALT1,COLGALT2:LysylCOLGALT1,COLGALT2:Lysylhydroxylated collagenhydroxylated collagenhydroxylated collagen
propeptidespropeptidespropeptides
Procollagen N-Procollagen N-Procollagen N-proteinasesproteinasesproteinases
2OGSUCCA
Procollagen C-Procollagen C-Procollagen C-proteinasesproteinasesproteinases
2 Collagen type VI dimer
CO2
2 Collagen type VI tetramer
O2
\\Collagens and
tropocollagens:SerpinH1
Collagens andtropocollagens
UDP-Gal UDP
COLGALT1,COLGALT2
COLGALT1,COLGALT2:Galactosyl-COLGALT1,COLGALT2:Galactosyl-COLGALT1,COLGALT2:Galactosyl-hydroxylysyl collagen propeptideshydroxylysyl collagen propeptideshydroxylysyl collagen propeptides
\\Transmembrane collagens Transmembrane collagens
Prolyl 3-hydroxylases:Fe2+:4-Prolyl 3-hydroxylases:Fe2+:4-Prolyl 3-hydroxylases:Fe2+:4-Hyp collagen propeptidesHyp collagen propeptidesHyp collagen propeptides
SERPINH1
Fibrillar procollagensFibrillar procollagensFibrillar procollagens
Fibrillar procollagens -NFibrillar procollagens -NFibrillar procollagens -N
PLOD3:Fe2+ dimer
Tropocollagens
PCOLCEsPCOLCEsPCOLCEs
Collagen type VI
\\
4-Hyp collagen4-Hyp collagen4-Hyp collagenpropeptidespropeptidespropeptides
\\
Lysyl hydroxylated collagenLysyl hydroxylated collagenLysyl hydroxylated collagenpropeptidespropeptidespropeptides
\\
Galactosyl-hydroxylysyl collagen propeptidesGalactosyl-hydroxylysyl collagen propeptidesGalactosyl-hydroxylysyl collagen propeptides
\\
Prolyl 3-hydroxylases:Fe2+
\\
Glucosyl-galactosyl-hydroxylysylGlucosyl-galactosyl-hydroxylysylGlucosyl-galactosyl-hydroxylysylcollagen propeptidescollagen propeptidescollagen propeptides
Collagen chaintrimerization
UDP-Glc
PLOD3:Fe2+ dimer:Glucosyl-PLOD3:Fe2+ dimer:Glucosyl-PLOD3:Fe2+ dimer:Glucosyl-galactosyl-hydroxylysyl collagengalactosyl-hydroxylysyl collagengalactosyl-hydroxylysyl collagen
propeptidespropeptidespropeptides
PLOD3:Fe2+ dimer:LysylPLOD3:Fe2+ dimer:LysylPLOD3:Fe2+ dimer:Lysylhydroxylated collagenhydroxylated collagenhydroxylated collagen
propeptidespropeptidespropeptides PLOD3:Fe2+ dimer:Galactosyl-PLOD3:Fe2+ dimer:Galactosyl-PLOD3:Fe2+ dimer:Galactosyl-hydroxylysyl collagen propeptideshydroxylysyl collagen propeptideshydroxylysyl collagen propeptides
1.4E2
-1.96E1
The biosynthesis of collagen is a multistep process. Collagen propeptides are cotranslationally translocated into the ER lumen. Propeptides undergo a number of post-translational modifications. Proline and lysine residues may be hydroxylated by prolyl 3-, prolyl 4- and lysyl hydroxylases. 4-hy-droxyproline is essential for intramolecular hydrogen bonding and stability of the triple helical col-lagenous domain. In fibril forming collagens approximately 50% of prolines are 4-hydroxylated; the extent of this and of 3-hydroxyproline and lysine hydroxylation varies between tissues and collagen types (Kivirikko et al. 1972, 1992). Hydroxylysine molecules can form cross-links between collagen molecules in fibrils, and are sites for glycosyl- and galactosylation. Collagen peptides all have non-collagenous domains; collagens within the subclasses have common chain structures. These non-collagenous domains have regulatory functions; some are biologically active when cleaved from the main peptide chain. Fibrillar collagens all have a large triple helical domain (COL1) bordered by N and C terminal extensions, called the N and C propeptides, which are cleaved prior to forma-tion of the collagen fibril. The C propeptide, also called the NC1 domain, is highly conserved. It dir-ects chain association during intracellular assembly of the procollagen molecule from three colla-gen propeptide alpha chains (Hulmes 2002). The N-propeptide has a short linker (NC2) connecting the main triple helix to a short minor one (COL2) and a globular N-terminal region NC3. NC3 do-mains are variable both in size and the domains they contain.

https://reactome.org Page 40
Collagen propeptides typically undergo a number of post-translational modifications. Proline and lysine residues are hydroxylated by prolyl 3-, prolyl 4- and lysyl hydroxylases. 4-hydroxyproline is essential for intramolecular hydrogen bonding and stability of the triple helical collagenous do-main. Prolyl 4-hydroxylase may also have a role in alpha chain association as no association of the C-propeptides of type XII collagen was seen in the presence of prolyl 4-hydroxylase inhibitors (Mazzorana et al. 1993, 1996). In fibril forming collagens approximately 50% of prolines are 4-hy-droxylated; the extent of this is species dependent, lower hydroxylation correlating with lower am-bient temperature and thermal stability (Cohen-Solal et al. 1986, Notbohm et al. 1992). Similarly the extent of 3-hydroxyproline and lysine hydroxylation varies between tissues and collagen types (Kivirikko et al. 1992). Hydroxylysine molecules can form cross-links between collagen molecules in fibrils, and are sites for glycosyl- and galactosylation.
Collagen molecules fold and assemble through a series of distinct intermediates (Bulleid 1996). In-dividual collagen polypeptide chains are translocated co-translationally across the membrane of the endoplasmic reticulum (ER). Intra-chain disulfide bonds are formed within the N-propeptide, and hydroxylation of proline and lysine residues occurs within the triple helical domain (Kivirikko et al. 1992). When the peptide chain is fully translocated into the ER lumen the C-propeptide folds, the conformation being stabilized by intra-chain disulfide bonds (Doege and Fessler 1986). Pro al-pha-chains associate via the C-propeptides (Byers et al. 1975, Bachinger et al. 1978), or NC2 do-mains for FACIT family collagens (Boudko et al. 2008) to form an initial trimer which can be stabil-ized by the formation of inter-chain disulfide bonds (Schofield et al. 1974, Olsen et al. 1976), though these are not a prerequisite for further folding (Bulleid et al. 1996). The triple helix then nucleates and folds in a C- to N- direction. The association of the individual chains and subsequent triple helix formation are distinct steps (Bachinger et al. 1980). The N-propeptides associate and in some cases form inter-chain disulfide bonds (Bruckner et al., 1978). Procollagen is released via carriers into the exracellular space (Canty & Kadler 2005). Fibrillar procollagens undergo removal of the C- and N-propeptides by procollagen C and N proteinases respectively, both Zn2+ dependent metallopro-teinases. Propeptide processing is a required step for normal collagen I and III fibril formation, but collagens can retain some or all of their non-collagenous propeptides. Retained collagen type V and XI N-propeptides contribute to the control of fibril growth by sterically limiting lateral molecule ad-dition (Fichard et al. 1995). Processed fibrillar procollagen is termed tropocollagen, which is con-sidered to be the unit of higher order fibrils and fibres. Tropocollagens of the fibril forming colla-gens I, II, III, V and XI sponteneously aggregate in vitro in a manner that has been compared with crystallization, commencing with a nucleation event followed by subsequent organized aggregation (Silver et al. 1992, Prockop & Fertala 1998). Fibril formation is stabilized by lysyl oxidase catalyzed crosslinks between adjacent molecules (Siegel & Fu 1976).
References
Kadler KE, Baldock C, Bella J & Boot-Handford RP (2007). Collagens at a glance. J Cell Sci, 120, 1955-
8.
Myllyharju J & Kivirikko KI (2004). Collagens, modifying enzymes and their mutations in humans,
flies and worms. Trends Genet, 20, 33-43.
Edit history
Date Action Author
2010-07-20 Authored Jupe S

https://reactome.org Page 41
Date Action Author
2011-10-12 Created Jupe S
2012-05-14 Edited Jupe S
2012-05-24 Reviewed Canty-Laird EG
2018-08-23 Modified Schmidt EE
Entities found in this pathway (16)
Input UniProt Id #Fold cha...
SERPINH1 P50454 -1.61e+00
COL4A5 P29400 -1.87e+00
PCOLCE Q15113 -1.55e+00
COL4A3 Q01955 4.74
COL4A4 P53420 3.93
COL7A1 Q02388 2.34
COL5A3 P25940 1.9
COL4A1 P02462 2.52
COL11A1 P12107 5.14
COL4A2 P08572 1.85
ADAMTS2 O95450 1.63
ADAMTS14 Q8WXS8 -5.36e+00
COL1A1 P02452 -2.48e+00
COL3A1 P02461 -1.81e+00
COL27A1 Q8IZC6 -1.98e+00
COL15A1 P39059 -1.90e+00

https://reactome.org Page 42
Regulation of gene expression by Hypoxia-inducible Factor (R-HSA-1234158)16.
cytosol
nucleoplasmmitochondrial inner membrane
HIF1A,HIF2A
2OG
SUCCACO2
hydroxyAsn-HIF1A,HIF2A
2
2
2
2
EPAS1
2xHP-EPAS1
2
2
2
2
HIF1A
2xHP-HIF1A
HIF3A
HP-HIF3A
hydroxyPro-HIF-alphahydroxyPro-HIF-alphahydroxyPro-HIF-alpha
hydroxyPro-HIF-alpha:VHL:EloB,C:hydroxyPro-HIF-alpha:VHL:EloB,C:hydroxyPro-HIF-alpha:VHL:EloB,C:CUL2:RBX1:LIMD1,AJUBA,WTIP:PHD2,3CUL2:RBX1:LIMD1,AJUBA,WTIP:PHD2,3CUL2:RBX1:LIMD1,AJUBA,WTIP:PHD2,3
UBE2D1,2,3:Ubiquitin
UBE2D1,2,3
ub-hydroxyPro-HIF-alpha:VHL:EloB:EloC:ub-hydroxyPro-HIF-alpha:VHL:EloB:EloC:ub-hydroxyPro-HIF-alpha:VHL:EloB:EloC:CUL2:RBX1CUL2:RBX1CUL2:RBX1
\\
O2
\\
HIF-alphaHIF-alphaHIF-alpha
HIF-alphaHIF-alphaHIF-alpha ARNT
HIFHIFHIF
\\
EP300
CREBBP
DNA
HIF:CBP:p300HIF:CBP:p300HIF:CBP:p300
O2
2OG
CO2
SUCCA
2
2
2
2
EPAS1
O2
2OG
SUCCA
CO2
2xHP-EPAS1
PHD1,3
2
2
2
2
HIF1A
2xHP-HIF1A
HIF3A
HP-HIF3A
VHL:EloB,C:CUL2:RBX1hydroxyPro-HIF-alphahydroxyPro-HIF-alphahydroxyPro-HIF-alpha
hydroxyPro-HIF-alpha:VHL:EloB,C:CUL2:RBX1hydroxyPro-HIF-alpha:VHL:EloB,C:CUL2:RBX1hydroxyPro-HIF-alpha:VHL:EloB,C:CUL2:RBX1
UBE2D1,2,3:Ubiquitin
UBE2D1,2,3
ub-HIF-alpha:VHL:EloB,C:CUL2:RBX1ub-HIF-alpha:VHL:EloB,C:CUL2:RBX1ub-HIF-alpha:VHL:EloB,C:CUL2:RBX1
\\
FIH1:FIH1
\\
CA9
\\
\\
EPO
Ub
VEGFA
CA9 geneVEGFA gene
EPO gene
CITED2
\\
HIGD1A
HIGD1A gene
PHD2,3:LIMD1,AJUBA,WTIP:VHL:EloB:EloC:CUL2:RBX1
ub-hydroxyPro-HIF-alpha:VHL:EloB:EloC:ub-hydroxyPro-HIF-alpha:VHL:EloB:EloC:ub-hydroxyPro-HIF-alpha:VHL:EloB:EloC:CUL2:RBX1:LIMD1,AJUBA,WTIF:PHD2,3CUL2:RBX1:LIMD1,AJUBA,WTIF:PHD2,3CUL2:RBX1:LIMD1,AJUBA,WTIF:PHD2,3
ub-hydroxyPro-HIF-alpha:VCB (with or withoutub-hydroxyPro-HIF-alpha:VCB (with or withoutub-hydroxyPro-HIF-alpha:VCB (with or withoutLIMD1)LIMD1)LIMD1)
VBC complex (with or withoutLIMD1)
26S proteasome
1.4E2
-1.96E1
Cellular compartments: nucleoplasm.
HIF-alpha (HIF1A, HIF2A (EPAS1), HIF3A) is translocated to the nucleus, possibly by two pathways: importin 4/7 (Chachami et al. 2009) and importin alpha/beta (Depping et al. 2008). Once in the nuc-leus HIF-alpha heterodimerizes with HIF-beta (ARNT) (Wang et al. 1995, Jiang et al. 1996, Tian et al. 1997, Gu et al. 1998, Erbel et al. 2003) and recruits CBP and p300 to promoters of target genes (Ebert and Bunn 1998, Kallio et al. 1998, Ema et al. 1999, Gu et al. 2001, Dames et al. 2002, Freedman et al. 2002).
References
Chachami G, Paraskeva E, Mingot JM, Braliou GG, Görlich D & Simos G (2009). Transport of hypox-ia-inducible factor HIF-1alpha into the nucleus involves importins 4 and 7. Biochem Biophys
Res Commun, 390, 235-40.
Kallio PJ, Okamoto K, O'Brien S, Carrero P, Makino Y, Tanaka H & Poellinger L (1998). Signal trans-duction in hypoxic cells: inducible nuclear translocation and recruitment of the CBP/p300 co-
activator by the hypoxia-inducible factor-1alpha. EMBO J, 17, 6573-86.
Depping R, Steinhoff A, Schindler SG, Friedrich B, Fagerlund R, Metzen E, ... Köhler M (2008). Nuc-lear translocation of hypoxia-inducible factors (HIFs): involvement of the classical importin al-
pha/beta pathway. Biochim Biophys Acta, 1783, 394-404.
Tian H, McKnight SL & Russell DW (1997). Endothelial PAS domain protein 1 (EPAS1), a transcrip-
tion factor selectively expressed in endothelial cells. Genes Dev, 11, 72-82.
Jiang BH, Rue E, Wang GL, Roe R & Semenza GL (1996). Dimerization, DNA binding, and transactiv-
ation properties of hypoxia-inducible factor 1. J Biol Chem, 271, 17771-8.
Edit history
https://reactome.org Page 43
Date Action Author
2011-03-09 Edited May B
2011-03-09 Authored May B
2011-03-18 Created May B
2012-05-19 Reviewed Rantanen K
2018-08-30 Modified Croft D
Entities found in this pathway (4)
Input UniProt Id #Fold cha...
HIGD1A Q9Y241 2.26
HIF3A Q9Y2N7 6.03
CITED2 Q99967 3.04
VEGFA P15692 -2.33e+00
Input Ensembl Id #Fold cha...
HIGD1A ENSG00000181061 2.26
VEGFA ENSG00000112715 -2.33e+00

https://reactome.org Page 44
L1CAM interactions (R-HSA-373760)17.
cytosol
cytosol
cytosol
clathrin-coated endocytic vesicle
nucleoplasm
endosome
L1CAM
NCAN
L1:Neurocan
p-2T-MAP2K1
NRP1
ADP
HSPA8
ATP
p-Y1176-L1CAM
P
ADP
SRC-1
ERM:PIP2pL1:EzrinL1 trans-homodimer
L1CAM
F-actin
pL1:ERM:F-actinpL1:ERM:F-actinpL1:ERM:F-actin
KIAA1598
pL1:Shootin-1:F-actinpL1:Shootin-1:F-actinpL1:Shootin-1:F-actin
F-actin
ANK1
2Pi
Clathrin:AP-2 complex CRMP-2:NUMB:alpha adaptin
L1:NUMB:CRMP-2:alpha-adaptin
NRP1
L1CAML1:clathrin-coated vesicle
NFASC:NRCAM
NFASC
ANK1
NFASC
GTP
SH3GL2
Pi
L1:AP-2 Clathrin complex
GDP
Dynamin-1/2/3Dynamin-1/2/3Dynamin-1/2/3
L1:AP-2 Clathrin complex
p90rskp90rskp90rsk
ATP
ADP
pL1 (S1152):p90rsk:clathrin-pL1 (S1152):p90rsk:clathrin-pL1 (S1152):p90rsk:clathrin-dynamin complexdynamin complexdynamin complex
p-T185,Y187-MAPK1
P P
ATPADP
pL1 (S1204, 1248):ERK2:clathrin-dynamin complex
KIF4 dimer
AP2 clathrin:L1:KIF4:AP2 clathrin:L1:KIF4:AP2 clathrin:L1:KIF4:microtubulemicrotubulemicrotubule
22
PALM-C3,4-GAP43PaPa
H2O
GAP43
PALM
LYPLA2
MicrotubuleMicrotubuleMicrotubule
L1 homodimer
ANK1-3ANK1-3ANK1-3
L1 trans-homodimer:AnkyrinL1 trans-homodimer:AnkyrinL1 trans-homodimer:AnkyrinL1:HNK-1
Laminin-111Laminin-111Laminin-111
L1:LamininL1:LamininL1:Laminin
SPTA:SPTB:F-actinSPTA:SPTB:F-actinSPTA:SPTB:F-actin
L1 dimer:Ankyrin:Spectrin:L1 dimer:Ankyrin:Spectrin:L1 dimer:Ankyrin:Spectrin:F-actinF-actinF-actin
KCNQ2,3, SCNAs:SCNBsKCNQ2,3, SCNAs:SCNBsKCNQ2,3, SCNAs:SCNBs
NFASC:NRCAM:ANK1-3:NFASC:NRCAM:ANK1-3:NFASC:NRCAM:ANK1-3:SPTA:SPTB:F-actinSPTA:SPTB:F-actinSPTA:SPTB:F-actin
NFASC:NRCAM:ANK1-3:SPTA:NFASC:NRCAM:ANK1-3:SPTA:NFASC:NRCAM:ANK1-3:SPTA:SPTB:F-actin:KCNQ2,3, SCNAs:SPTB:F-actin:KCNQ2,3, SCNAs:SPTB:F-actin:KCNQ2,3, SCNAs:
SCNBsSCNBsSCNBs
ATP
ADP
pL1 (Y1229):L1CAM
unidentified protein tyrosinekinase
NCAM1
L1:NCAM1 complex
L1CAM
FGFR1c L1:FGFR1
EGFR L1-EGFR trans-heterodimer
L1:NRP1Integrins L1:Integrincomplex
\\
ATP
VAV2p-Y172-VAV2P
ADP
unidentified protein tyrosinekinase
RAC1:GDP
RAC1:GTP
GDP
GTP
22
ATP
PAK1 dimer
PAK1 ADP
pPAK1:Rac1-GTP
22
ATP
p-2S-MAP2K1:MAPK3p-T,Y-MAPK3:p-2S-MAP2K1
ADP
22
ATP
p-S,T-MAP2K2:MAPK1p-S,T-MAP2K2:p-T,Y-MAPK1
ADP
ATP
Casein kinase II
pL1:CK-II
ADP
unknown protein
CNTN2
L1:Axonin-1
CNTN1
L1:CNTN1
ALCAML1:ALCAML1:ALCAML1:ALCAM RANBP9L1:RanBPML1:RanBPML1:RanBPM
5
CD24(29-534)
L1:HSA
ATP
EPHB2
ADP
p-Y-L1:EPHB2p-Y-L1:EPHB2p-Y-L1:EPHB2
ANK1
CHL1CHL1:Ankyrin-GCHL1:HSP7CCHL1:NRP1
Integrin alpha1beta1, alpha2beta1,Integrin alpha1beta1, alpha2beta1,Integrin alpha1beta1, alpha2beta1,alpha10beta1alpha10beta1alpha10beta1
CHL1:alpha1beta1/alpha2beta1CHL1:alpha1beta1/alpha2beta1CHL1:alpha1beta1/alpha2beta1integrinsintegrinsintegrins
CNTN6CHL1:CNTN6
NRCAM
NrCAM:Ankyrin-G
NrCAM:Axonin-1
CNTN2
PSD-95 familyPSD-95 familyPSD-95 family
NrCAM:SAPNrCAM:SAPNrCAM:SAPmembersmembersmembers
NRP2
NRP2:NrCAMNRP2:NrCAMNRP2:NrCAM
Neurofascin:Ankyrin-Gcomplex
Trans neurofascin dimer:ankyrin-G
ATP
NFASC:pNFASC
ADP
unidentified protein tyrosinekinase
DCXpNFASC:
Doublecortin
SDCBP
Neurofascin:Syntenin-1 complex
Contactin1:CASPR complexNeurofascin:CNTN1:
CASPR complex
1.4E2
-1.96E1
The L1 family of cell adhesion molecules (L1CAMs) are a subfamily of the immunoglobulin super-family of transmembrane receptors, comprised of four structurally related proteins: L1, Close Homolog of L1 (CHL1), NrCAM, and Neurofascin. These CAMs contain six Ig like domains, five or six fibronectin like repeats, a transmembrane region and a cytoplasmic domain. The L1CAM family has been implicated in processes integral to nervous system development, including neurite out-growth, neurite fasciculation and inter neuronal adhesion.
L1CAM members are predominately expressed by neuronal, as well as some nonneuronal cells, during development. Except CHL1 all the other members of L1 family contain an alternatively spliced 12-nclueotide exon, encoding the amino acid residues RSLE in the neuronal splice forms but missing in the non-neuronal cells. The extracellular regions of L1CAM members are divergent and differ in their abilities to interact with extracellular, heterophilic ligands. The L1 ligands in-clude other Ig-domain CAMs (such as NCAM, TAG-1/axonin and F11), proteoglycans type molecules (neurocan), beta1 integrins, and extra cellular matrix protein laminin, Neuropilin-1, FGF and EGF receptors. Some of these L1-interacting proteins also bind to other L1CAM members. For example TAG-1/axonin interact with L1 and NrCAM; L1, neurofascin and CHL1 binds to contactin family members. The cytoplasmic domains of L1CAM members are most highly conserved. Nevertheless, they have different cytoplasmic binding partners, and even those with similar binding partners may be involved in different signaling complexes and mechanisms. The most conserved feature of L1CAMs is their ability to interact with the actin cytoskeletal adapter protein ankyrin. The cytoplas-mic ankyrin-binding domain, exhibits the highest degree of amino acid conservation throughout the L1 family.
References
Kamiguchi H (2003). The mechanism of axon growth: what we have learned from the cell adhesion
molecule L1. Mol Neurobiol, 28, 219-28.
Kamiguchi H & Lemmon V (1997). Neural cell adhesion molecule L1: signaling pathways and
growth cone motility. J Neurosci Res, 49, 1-8.
Herron LR, Hill M, Davey F & Gunn-Moore FJ (2009). The intracellular interactions of the L1 family
of cell adhesion molecules. Biochem J, 419, 519-31.
Schmid RS & Maness PF (2008). L1 and NCAM adhesion molecules as signaling coreceptors in neur-
onal migration and process outgrowth. Curr Opin Neurobiol, 18, 245-50.
Maness PF & Schachner M (2007). Neural recognition molecules of the immunoglobulin superfam-
ily: signaling transducers of axon guidance and neuronal migration. Nat Neurosci, 10, 19-26.
https://reactome.org Page 45
Edit history
Date Action Author
2008-07-16 Created Garapati P V
2008-07-30 Edited Garapati P V
2008-07-30 Authored Garapati P V
2010-02-16 Reviewed Maness PF
2018-08-23 Modified Schmidt EE
Entities found in this pathway (22)
Input UniProt Id #Fold cha...
LAMC1 P11047 1.73
SCN7A Q01118 2.69
LAMB1 P07942 1.69
KCNQ3 O43525 5.4
ANK2 Q01484 -2.00e+00
DLG3 Q92796 -1.57e+00
ITGA10 O75578 13.87
SPTBN4 Q9H254 -2.11e+00
ITGA2 P17301 -2.06e+00
ALCAM Q13740 1.79
SCN1B Q07699 -2.44e+00
EPHB2 P29323 -3.38e+00
TUBA4A P68363, P68366 -2.76e+00
RANBP9 Q96S59 1.66
TUBB3 Q13509 -2.82e+00
TUBA1A Q71U36 -2.42e+00
RPS6KA2 Q15349 1.89
TUBA1B P68363, Q9H853 -1.72e+00
RPS6KA5 O75582 -5.16e+00
NRP2 O60462 2.07
DNM1 Q05193 -3.37e+00
ACTB P60709 1.87

https://reactome.org Page 46
ATF4 activates genes (R-HSA-380994)18.
cytosol
nucleoplasm
endoplasmic reticulum lumen
DDIT3 gene
\\
ATF3
ATF4
\\
ASNS
\\
CCL2
\\
DDIT3
ATF6(1-380)
NF-YNF-YNF-Y
HERPUD1 gene \\
HERPUD1
\\
IGFBP1IGFPB1 gene
\\
IL8
KSRP:mRNA Degradation Complex
IL8 geneATF3 geneASNS gene
CCL2 gene
1.4E2
-1.96E1
ATF4 is a transcription factor and activates expression of IL-8, MCP1, IGFBP-1, CHOP, HERP1 and ATF3.
References
Yamaguchi Y, Larkin D, Lara-Lemus R, Ramos-Castañeda J, Liu M & Arvan P (2008). Endoplasmic reticulum (ER) chaperone regulation and survival of cells compensating for deficiency in the ER
stress response kinase, PERK. J Biol Chem, 283, 17020-9.
Averous J, Bruhat A, Jousse C, Carraro V, Thiel G & Fafournoux P (2004). Induction of CHOP expres-sion by amino acid limitation requires both ATF4 expression and ATF2 phosphorylation. J Biol
Chem, 279, 5288-97.
Armstrong JL, Flockhart R, Veal GJ, Lovat PE & Redfern CP (2010). Regulation of Endoplasmic Re-ticulum Stress-induced Cell Death by ATF4 in Neuroectodermal Tumor Cells. J Biol Chem, 285,
6091-100.
Gargalovic PS, Imura M, Zhang B, Gharavi NM, Clark MJ, Pagnon J, ... Lusis AJ (2006). Identification of inflammatory gene modules based on variations of human endothelial cell responses to oxid-
ized lipids. Proc Natl Acad Sci U S A, 103, 12741-6.
Gargalovic PS, Gharavi NM, Clark MJ, Pagnon J, Yang WP, He A, ... Lusis AJ (2006). The unfolded protein response is an important regulator of inflammatory genes in endothelial cells. Arterio-
scler Thromb Vasc Biol, 26, 2490-6.
Edit history
Date Action Author
2008-11-19 Created May B
2008-12-02 Reviewed Matthews L, D'Eustachio P, Gillespie ME
https://reactome.org Page 47
Date Action Author
2009-06-02 Edited May B
2009-06-02 Authored May B
2010-04-30 Reviewed Urano F
2014-02-07 Modified May B
Entities found in this pathway (5)
Input UniProt Id #Fold cha...
ASNS P08243 -2.00e+00
HERPUD1 Q15011 1.67
ATF3 P18847 3.93
NFYB P25208 2.02
DDIT3 P35638 -2.07e+00
Input Ensembl Id #Fold cha...
ASNS ENSG00000070669 -2.00e+00
HERPUD1 ENSG00000051108 1.67
ATF3 ENSG00000162772 3.93
DDIT3 ENSG00000175197 -2.07e+00

https://reactome.org Page 48
Circadian Clock (R-HSA-400253)19.
cytosol
nucleoplasm
ARNTL
NR1D1
ferriheme b
CLOCK,NPAS2CLOCK,NPAS2CLOCK,NPAS2
BMAL1:CLOCK,NPAS2BMAL1:CLOCK,NPAS2BMAL1:CLOCK,NPAS2
\\
RORA activates geneexpression
p-BMAL1:p-CLOCK,NPAS2p-BMAL1:p-CLOCK,NPAS2p-BMAL1:p-CLOCK,NPAS2
NR1D1 (REV-ERBA) represses gene expression
NRIP1
p-BMAL1:p-CLOCK,NPAS2:p-BMAL1:p-CLOCK,NPAS2:p-BMAL1:p-CLOCK,NPAS2:RORA geneRORA geneRORA gene
CRY1
CRY2
PER1
PER2
CREM:PER1 geneCREM:PER1 geneCREM:PER1 gene
NFIL3:BHLHE41:PER2NFIL3:BHLHE41:PER2NFIL3:BHLHE41:PER2genegenegene
RBM4:PER1 mRNARBM4:PER1 mRNARBM4:PER1 mRNA
p-S133-CREB:CRTC1,2,3:PPARGC1A genep-S133-CREB:CRTC1,2,3:PPARGC1A genep-S133-CREB:CRTC1,2,3:PPARGC1A gene
CRY1,2
BMAL1:CLOCK,NPAS2:CRYBMAL1:CLOCK,NPAS2:CRYBMAL1:CLOCK,NPAS2:CRY
2
2
CRY:PER:KinaseCRY:PER:KinaseCRY:PER:Kinase
p-CRY:p-PER:Kinasep-CRY:p-PER:Kinasep-CRY:p-PER:Kinase
CSNK1E,CSNK1D
SIRT1
\\
p-CRY:p-PER:Kinasep-CRY:p-PER:Kinasep-CRY:p-PER:Kinase
BMAL1:CLOCK,NPAS2:CRY:PERBMAL1:CLOCK,NPAS2:CRY:PERBMAL1:CLOCK,NPAS2:CRY:PER
BTRC
p-PER1,p-PER2p-PER1,p-PER2p-PER1,p-PER2
Beta-TrCP1:PERBeta-TrCP1:PERBeta-TrCP1:PER
FBXL3
p-CRY1,p-CRY2
FBXL3:CRY
\\
ub-p-CRY1,ub-p-CRY2
\\
Ub
ub-p-PER1,ub-p-PER2ub-p-PER1,ub-p-PER2ub-p-PER1,ub-p-PER2
BTRC:CUL1:SKP1
Ub
RORA
PPARGC1A
PPARGC1AATP
ADP
PER1,PER2PER1,PER2PER1,PER2
NPAS2
HDAC3
NCOR1
EP300
PPARGC1A
CLOCK
\\
p-T69,T71-ATF2P P
MEF2C,D:PPARGC1A
p-PPARGC1A
\\
HIF1A
PPARGC1A gene
RORA gene
PP1 catalytic subunitPP1 catalytic subunitPP1 catalytic subunit
\\
ARNTL gene
SIRT1
\\
CLOCK gene
RAI1
\\
NPAS2 gene
PPARA:RXRACoactivator complex
BMAL1:CLOCK,NPAS2 activates circadian geneexpression
CRY1 gene
p-BMAL1:p-CLOCK,p-BMAL1:p-CLOCK,p-BMAL1:p-CLOCK,NPAS2:CRY1 geneNPAS2:CRY1 geneNPAS2:CRY1 gene
\\
\\
CRY2 gene
PER1 gene
p-BMAL1:p-CLOCK,p-BMAL1:p-CLOCK,p-BMAL1:p-CLOCK,NPAS2:PER1 geneNPAS2:PER1 geneNPAS2:PER1 gene
\\
p-CREB:CRTC1:PER1p-CREB:CRTC1:PER1p-CREB:CRTC1:PER1genegenegene
SIK1
PER2 gene
p-BMAL1:p-CLOCK,p-BMAL1:p-CLOCK,p-BMAL1:p-CLOCK,NPAS2:PER2 geneNPAS2:PER2 geneNPAS2:PER2 gene
\\
NR3C1:DexamethasoneNR3C1:DexamethasoneNR3C1:Dexamethasone
NR1D1 gene
p-BMAL1:p-CLOCK,NPAS2:NR1D1 genep-BMAL1:p-CLOCK,NPAS2:NR1D1 genep-BMAL1:p-CLOCK,NPAS2:NR1D1 gene\\
RORA:Coactivator:ARNTL gene
NR1D1:heme:Corepressors:ARNTL
gene
NR1D1:heme:NR1D1:heme:NR1D1:heme:Corepressors:PPARGC1ACorepressors:PPARGC1ACorepressors:PPARGC1A
genegenegene
NR1D1:heme:Corepressors:CLOCK
gene
RORA:EP300:NPAS2RORA:EP300:NPAS2RORA:EP300:NPAS2genegenegeneNR1D1:heme:NR1D1:heme:NR1D1:heme:
Corepressors:NPAS2Corepressors:NPAS2Corepressors:NPAS2genegenegene
RORA:Coactivator:NR1D1geneNR1D1:heme:
Corepressors:NR1D1gene
1.4E2
-1.96E1
Cellular compartments: nucleoplasm, cytosol.
At the center of the mammalian circadian clock is a negative transcription/translation-based feed-back loop: The BMAL1:CLOCK/NPAS2 (ARNTL:CLOCK/NPAS2) heterodimer transactivates CRY and PER genes by binding E-box elements in their promoters; the CRY and PER proteins then inhibit transactivation by BMAL1:CLOCK/NPAS2. BMAL1:CLOCK/NPAS2 activates transcription of CRY, PER, and several other genes in the morning. Levels of PER and CRY proteins rise during the day and inhibit expression of CRY, PER, and other BMAL1:CLOCK/NPAS2-activated genes in the after-noon and evening. During the night CRY and PER proteins are targeted for degradation by phos-phorylation and polyubiquitination, allowing the cycle to commence again in the morning.
Transcription of the BMAL1 (ARNTL) gene is controlled by ROR-alpha and REV-ERBA (NR1D1), both of which are targets of BMAL1:CLOCK/NPAS2 in mice and both of which compete for the same element (RORE) in the BMAL1 promoter. ROR-alpha (RORA) activates transcription of BMAL1; REV-ERBA represses transcription of BMAL1. This mutual control forms a secondary, reinforcing loop of the circadian clock. REV-ERBA shows strong circadian rhythmicity and confers circadian expression on BMAL1.

https://reactome.org Page 49
BMAL1 can form heterodimers with either CLOCK or NPAS2, which act redundantly but show dif-ferent tissue specificity. The BMAL1:CLOCK and BMAL1:NPAS2 heterodimers activate a set of genes that possess E-box elements (consensus CACGTG) in their promoters. This confers circadian expression on the genes. The PER genes (PER1, PER2, PER3) and CRY genes (CRY1, CRY2) are among those activated by BMAL1:CLOCK and BMAL1:NPAS2. PER and CRY mRNA accumulates during the morning and the proteins accumulate during the afternoon. PER and CRY proteins form complexes in the cytosol and these are bound by either CSNK1D or CSNK1E kinases which phos-phorylate PER and CRY. The phosphorylated PER:CRY:kinase complex is translocated into the nuc-leus due to the nuclear localization signal of PER and CRY. Within the nucleus the PER:CRY com-plexes bind BMAL1:CLOCK and BMAL1:NPAS2, inhibiting their transactivation activity and their phosphorylation. This reduces expression of the target genes of BMAL1:CLOCK and BMAL1:NPAS2 during the afternoon and evening.
PER:CRY complexes also traffic out of the nucleus into the cytosol due to the nuclear export signal of PER. During the night PER:CRY complexes are polyubiquitinated and degraded, allowing the cycle to begin again. Phosphorylated PER is bound by Beta-TrCP1, a cytosolic F-box type compon-ent of some SCF E3 ubiquitin ligases. CRY is bound by FBXL3, a nucleoplasmic F-box type compon-ent of some SCF E3 ubiquitin ligases. Phosphorylation of CRY1 by Adenosine monophosphate-activ-ated kinase (AMPK) enhances degradation of CRY1. PER and CRY are subsequently polyubiquitin-ated and proteolyzed by the 26S proteasome.
The circadian clock is cell-autonomous and some, but not all cells of the body exhibit circadian rhythms in metabolism, cell division, and gene transcription. The suprachiasmatic nucleus (SCN) in the hypothalamus is the major clock in the body and receives its major input from light (via ret-inal neurons) and a minor input from nutrient intake. The SCN and other brain tissues determine waking and feeding cycles and influence the clocks in other tissues by hormone secretion and nervous stimulation. Independently of the SCN, other tissues such as liver receive inputs from sig-nals from the brain and from nutrients.
References
Hastings MH, Maywood ES & O'Neill JS (2008). Cellular circadian pacemaking and the role of
cytosolic rhythms. Curr Biol, 18, R805-R815.
Ko CH & Takahashi JS (2006). Molecular components of the mammalian circadian clock. Hum Mol
Genet, 15, R271-7.
Takahashi JS, Hong HK, Ko CH & McDearmon EL (2008). The genetics of mammalian circadian or-
der and disorder: implications for physiology and disease. Nat Rev Genet, 9, 764-75.
Green CB, Takahashi JS & Bass J (2008). The meter of metabolism. Cell, 134, 728-42.
Edit history
Date Action Author
2009-03-24 Created May B
2009-05-18 Edited May B
2009-05-18 Authored May B
2009-05-27 Reviewed D'Eustachio P
2010-06-23 Reviewed Hirota T, Delaunay F, Kay SA, Albrecht U

https://reactome.org Page 50
Date Action Author
2017-12-16 Modified May B
Entities found in this pathway (11)
Input UniProt Id #Fold cha...
NAMPT P43490 2.03
CPT1A P50416 1.64
SREBF1 P36956 -1.68e+00
PER1 O15534 9.02
NPAS2 Q99743 2.18
NFIL3 Q16649 2.2
PPP1CB P62140 2.08
KLF15 Q9UIH9 24.7
DBP Q10586 -3.15e+00
NR3C1 P04150 -2.01e+00
Input Ensembl Id #Fold cha...
USP46 ENSG00000109189 1.68
NAMPT ENSG00000105835 2.03
CPT1A ENSG00000110090 1.64
SREBF1 ENSG00000072310 -1.68e+00
PER1 ENSG00000179094, ENST00000317276 9.02
NPAS2 ENSG00000170485 2.18
KLF15 ENSG00000163884 24.7
DBP ENSG00000105516 -3.15e+00

https://reactome.org Page 51
MET promotes cell motility (R-HSA-8875878)20.
cytosol
nucleoplasm
recycling endosome membrane
ATP
HGF:p-4Y-MET dimer:p-Y317-SHC1-HGF:p-4Y-MET dimer:p-Y317-SHC1-HGF:p-4Y-MET dimer:p-Y317-SHC1-222
ADP
SPINT1,2SPINT1,2SPINT1,2
HGF:p-4Y-MET dimer:p-Y317-HGF:p-4Y-MET dimer:p-Y317-HGF:p-4Y-MET dimer:p-Y317-SHC1-2:GRB2:SOS1SHC1-2:GRB2:SOS1SHC1-2:GRB2:SOS1
SPINT1
CIN85:endophilinCIN85:endophilinCIN85:endophilin
HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):
CIN85:endophillinCIN85:endophillinCIN85:endophillin
pro-HGF
H2O
GAB1
HGF dimerHGF dimerHGF dimer
HGF:p-4Y-MET dimer:HGF:p-4Y-MET dimer:HGF:p-4Y-MET dimer:GAB1GAB1GAB1
HGFAC dimer
METHGF dimer:METHGF dimer:METHGF dimer:MET
GRB2-1 HGF:p-4Y-MET dimer:GRB2-1:HGF:p-4Y-MET dimer:GRB2-1:HGF:p-4Y-MET dimer:GRB2-1:GAB1GAB1GAB1
HGF:p-4Y-MET dimer:GAB1,HGF:p-4Y-MET dimer:GAB1,HGF:p-4Y-MET dimer:GAB1,HGF:p-4Y-MET dimer:GRB2:HGF:p-4Y-MET dimer:GRB2:HGF:p-4Y-MET dimer:GRB2:
GAB1GAB1GAB1
ATP
ADP
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1GRB2-1:p-5Y-GAB1GRB2-1:p-5Y-GAB1
STAT3
HGF:p-4Y-MET dimer:STAT3HGF:p-4Y-MET dimer:STAT3HGF:p-4Y-MET dimer:STAT3
ATP
ADP
p-Y705-STAT3P
2
p-Y705-STAT3 dimerp-Y705-STAT3 dimerp-Y705-STAT3 dimer
PIK3CA:PIK3R1PIK3CA:PIK3R1PIK3CA:PIK3R1
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:PI3KGRB2-1:p-5Y-GAB1:PI3KGRB2-1:p-5Y-GAB1:PI3K
\\
2
p-Y705-STAT3 dimerp-Y705-STAT3 dimerp-Y705-STAT3 dimer
HGF:METHGF:METHGF:METdimerdimerdimer
PI(4,5)P2
ATP
PI(3,4,5)P3
HGF:MonoUb-K,p-4Y-METHGF:MonoUb-K,p-4Y-METHGF:MonoUb-K,p-4Y-METdimer:GRB2:p-Y-CBL,(HGF:dimer:GRB2:p-Y-CBL,(HGF:dimer:GRB2:p-Y-CBL,(HGF:
MonoUb-K,p-Y1003,4Y-MonoUb-K,p-Y1003,4Y-MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):CIN85:METdimer:p-Y-CBL):CIN85:METdimer:p-Y-CBL):CIN85:endophillin,(MET:Ub-LRIG1)endophillin,(MET:Ub-LRIG1)endophillin,(MET:Ub-LRIG1)
ADP
EPS15:HGS:STAM
HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):(HGF:MonoUb-K,p-Y1003,4Y-METdimer:p-Y-CBL):
CIN85:endophillin:EPS15:HGS:STAM,(MET:Ub-LRIG1:CIN85:endophillin:EPS15:HGS:STAM,(MET:Ub-LRIG1:CIN85:endophillin:EPS15:HGS:STAM,(MET:Ub-LRIG1:EPS15:HGS:STAM)EPS15:HGS:STAM)EPS15:HGS:STAM)
PIP3 activates AKT signaling
TNS3HGF:p-4Y-MET dimer:TNS3HGF:p-4Y-MET dimer:TNS3HGF:p-4Y-MET dimer:TNS3
TNS4:ITGB1
HGF:p-4Y-MET dimer:TNS4:ITGB1HGF:p-4Y-MET dimer:TNS4:ITGB1HGF:p-4Y-MET dimer:TNS4:ITGB1
8 8
RAF/MAP kinasecascade
ATP
HGF:p-Y1234,Y1235,Y1349,Y1356-MET dimerHGF:p-Y1234,Y1235,Y1349,Y1356-MET dimerHGF:p-Y1234,Y1235,Y1349,Y1356-MET dimer
CRK,CRKL
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:CRK,CRKLGRB2-1:p-5Y-GAB1:CRK,CRKLGRB2-1:p-5Y-GAB1:CRK,CRKL
PTPN11
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:PTPN11p-5Y-GAB1:PTPN11p-5Y-GAB1:PTPN11
RAPGEF1HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:CRK,CRKL:RAPGEF1GRB2-1:p-5Y-GAB1:CRK,CRKL:RAPGEF1GRB2-1:p-5Y-GAB1:CRK,CRKL:RAPGEF1
2 2
ADP
H2O
CBL:GRB2,CBL
Pi
PTK2
HGF:p-4Y-MET dimer:PTK2HGF:p-4Y-MET dimer:PTK2HGF:p-4Y-MET dimer:PTK2
HGF:p-Y1234,Y1235,Y1356-METHGF:p-Y1234,Y1235,Y1356-METHGF:p-Y1234,Y1235,Y1356-METdimerdimerdimer
PTPRJ
HGF:p-4Y-MET dimer:GRB2:HGF:p-4Y-MET dimer:GRB2:HGF:p-4Y-MET dimer:GRB2:CBL,(HGF:p-Y1003,4Y-METCBL,(HGF:p-Y1003,4Y-METCBL,(HGF:p-Y1003,4Y-MET
dimer:CBL)dimer:CBL)dimer:CBL)
ATP
ADP
HGF:p-4Y-MET dimer:GRB2:p-Y-HGF:p-4Y-MET dimer:GRB2:p-Y-HGF:p-4Y-MET dimer:GRB2:p-Y-CBL,(HGF:p-Y1003,4Y-MET dimer:p-CBL,(HGF:p-Y1003,4Y-MET dimer:p-CBL,(HGF:p-Y1003,4Y-MET dimer:p-
Y-CBL)Y-CBL)Y-CBL)
ATP
HGF:p-4Y-MET dimer:p-Y397-PTK2HGF:p-4Y-MET dimer:p-Y397-PTK2HGF:p-4Y-MET dimer:p-Y397-PTK2
ADP
RAP1:GDP
HGF:MonoUb-K,p-4Y-MET dimer:HGF:MonoUb-K,p-4Y-MET dimer:HGF:MonoUb-K,p-4Y-MET dimer:GRB2:p-Y-CBL,(HGF:MonoUb-K,p-GRB2:p-Y-CBL,(HGF:MonoUb-K,p-GRB2:p-Y-CBL,(HGF:MonoUb-K,p-
Y1003,4Y-MET dimer:p-Y-CBL)Y1003,4Y-MET dimer:p-Y-CBL)Y1003,4Y-MET dimer:p-Y-CBL)
RAP1:GTP
GTP
Integrin alpha2beta1,alpha3:beta1:(collagen,laminin,Integrin alpha2beta1,alpha3:beta1:(collagen,laminin,Integrin alpha2beta1,alpha3:beta1:(collagen,laminin,fibronectin)fibronectin)fibronectin)
GDP
MyrG-p-Y419-SRC
P
4 4
HGF:p-4Y-MET dimer:p-Y397-PTK2:HGF:p-4Y-MET dimer:p-Y397-PTK2:HGF:p-4Y-MET dimer:p-Y397-PTK2:MyrG-p-Y419-SRCMyrG-p-Y419-SRCMyrG-p-Y419-SRC
HGF:p-Y,1349,Y1356-MET dimerHGF:p-Y,1349,Y1356-MET dimerHGF:p-Y,1349,Y1356-MET dimer
PTPN1,PTPN2
HPN heterodimer
ATP
ADP
HGF:p-4Y-MET dimer:p-Y194,Y397-HGF:p-4Y-MET dimer:p-Y194,Y397-HGF:p-4Y-MET dimer:p-Y194,Y397-PTK2:MyrG-p-Y419-SRCPTK2:MyrG-p-Y419-SRCPTK2:MyrG-p-Y419-SRC
GRB2-1:SOS1
HGF:p-4Y-MET dimer:GRB2-1:HGF:p-4Y-MET dimer:GRB2-1:HGF:p-4Y-MET dimer:GRB2-1:SOS1SOS1SOS1
?(DOCK7)
HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:HGF:p-4Y-MET:p-5Y-GAB1, HGF:p-4Y-MET:GRB2-1:p-5Y-GAB1:CRK,CRKL:(DOCK7)p-5Y-GAB1:CRK,CRKL:(DOCK7)p-5Y-GAB1:CRK,CRKL:(DOCK7)
2
2
p21 RAS:GDP
ATP
GTP
ADP
HGF:p-4Y-MET dimer:p-Y194,Y397,HGF:p-4Y-MET dimer:p-Y194,Y397,HGF:p-4Y-MET dimer:p-Y194,Y397,Y576,Y577-PTK2:MyrG-p-Y419-SRCY576,Y577-PTK2:MyrG-p-Y419-SRCY576,Y577-PTK2:MyrG-p-Y419-SRC
p21 RAS:GTP
GDP
?RAC1:GDP
HGF:p-4Y-MET dimer,(HGF:p-Y1003,4Y-HGF:p-4Y-MET dimer,(HGF:p-Y1003,4Y-HGF:p-4Y-MET dimer,(HGF:p-Y1003,4Y-METdimer)METdimer)METdimer)
RAC1:GTP
GDPGTP
MUC20 HGF:p-4Y MET dimer:MUC20HGF:p-4Y MET dimer:MUC20HGF:p-4Y MET dimer:MUC20
Ub
LRIG1
MET:MET:MET:LRIG1LRIG1LRIG1
SOS1
RANBP9
HGF:p-4Y-MET dimer:RANBP9:HGF:p-4Y-MET dimer:RANBP9:HGF:p-4Y-MET dimer:RANBP9:SOS1SOS1SOS1
HGF:p-4Y-MET dimer:RANBP10HGF:p-4Y-MET dimer:RANBP10HGF:p-4Y-MET dimer:RANBP10
?
ARF6:GTP
MET:Ub-MET:Ub-MET:Ub-LRIG1LRIG1LRIG1
GGA3
unknown ubiquitin ligase
HGF:p-4Y-MET:p-5Y-GAB1,HGF:p-4Y-MET:p-5Y-GAB1,HGF:p-4Y-MET:p-5Y-GAB1,HGF:p-4Y-MET:GRB2-1:p-5Y-HGF:p-4Y-MET:GRB2-1:p-5Y-HGF:p-4Y-MET:GRB2-1:p-5Y-
GAB1:CRK,CRKL:GGA3:GAB1:CRK,CRKL:GGA3:GAB1:CRK,CRKL:GGA3:ARF6:GTPARF6:GTPARF6:GTP
Ub
RANBP10\\
GGC-RAB4:GTP
H2O
USP8
SHC1-2
HGF:p-4Y-MET dimer:SHC1-2HGF:p-4Y-MET dimer:SHC1-2HGF:p-4Y-MET dimer:SHC1-2
1.4E2
-1.96E1
Direct and indirect interactions of MET with integrins, focal adhesion kinase PTK2 (FAK1), tensin-4 (TNS4) and GTPases RAP1 and RAC1, induce morphological changes that promote cell motility and play an important role in HGF-induced invasiveness of cancer cells (Weidner et al. 1993, Beviglia et al. 1999, Sakkab et al. 2000, Parr et al. 2001, Trusolino et al. 2001, Lamorte et al. 2002, Chen and Chen 2006, Watanabe et al. 2006, Muharram et al. 2014, Murray et al. 2014).
References
Beviglia L & Kramer RH (1999). HGF induces FAK activation and integrin-mediated adhesion in
MTLn3 breast carcinoma cells. Int. J. Cancer, 83, 640-9.
Sakkab D, Lewitzky M, Posern G, Schaeper U, Sachs M, Birchmeier W & Feller SM (2000). Signaling of hepatocyte growth factor/scatter factor (HGF) to the small GTPase Rap1 via the large docking
protein Gab1 and the adapter protein CRKL. J. Biol. Chem., 275, 10772-8.
Parr C, Davies G, Nakamura T, Matsumoto K, Mason MD & Jiang WG (2001). The HGF/SF-induced phosphorylation of paxillin, matrix adhesion, and invasion of prostate cancer cells were sup-
pressed by NK4, an HGF/SF variant. Biochem. Biophys. Res. Commun., 285, 1330-7.
Lamorte L, Royal I, Naujokas M & Park M (2002). Crk adapter proteins promote an epithelial-mes-enchymal-like transition and are required for HGF-mediated cell spreading and breakdown of
epithelial adherens junctions. Mol. Biol. Cell, 13, 1449-61.
Chen SY & Chen HC (2006). Direct interaction of focal adhesion kinase (FAK) with Met is required for FAK to promote hepatocyte growth factor-induced cell invasion. Mol. Cell. Biol., 26, 5155-67.
Edit history

https://reactome.org Page 52
Date Action Author
2016-06-09 Created Orlic-Milacic M
2016-06-14 Edited Orlic-Milacic M
2016-06-14 Authored Orlic-Milacic M
2016-07-11 Reviewed Heynen G, Birchmeier W
2018-08-23 Modified Schmidt EE
Entities found in this pathway (11)
Input UniProt Id #Fold cha...
LAMC1 P11047 1.73
LAMB1 P07942 1.69
LAMA3 Q16787 1.82
COL5A3 P25940 1.9
HGF P14210 -1.98e+00
COL11A1 P12107 5.14
LAMA2 P24043 4.19
COL1A1 P02452 -2.48e+00
ITGA2 P17301 -2.06e+00
COL3A1 P02461 -1.81e+00
COL27A1 Q8IZC6 -1.98e+00

https://reactome.org Page 53
Metallothioneins bind metals (R-HSA-5661231)21.
cytosol
7
7
MT4
MT1A
Zn2+
MT4:7Zn2+
MT1A:7Zn2+MT1A:7Zn2+MT1A:7Zn2+
7
Cd2+
MT1A:7Cd2+MT1A:7Cd2+MT1A:7Cd2+
6
As3+
Zn2+
MT1A:6As3+MT1A:6As3+MT1A:6As3+
7
MT4:7Cd2+
7
MT1B
MT1B:7Zn2+
7
MT1B:7Cd2+
Cd2+
10
MT4:10Cu1+
7
MT1E
Cu1+
MT1E:7Zn2+MT1E:7Zn2+MT1E:7Zn2+
Zn2+
Cd2+
Zn2+
7
MT1E:7Cd2+MT1E:7Cd2+MT1E:7Cd2+
Cd2+
7
MT1F
MT1F:7Zn2+
Zn2+
7
MT1F:7Cd2+
Cd2+
7
MT1G
MT1G:7Zn2+MT1G:7Zn2+MT1G:7Zn2+
Zn2+7
MT1G:7Cd2+MT1G:7Cd2+MT1G:7Cd2+
Cd2+
7
MT1H
MT1H:7Zn2+
Zn2+
7
MT1H:7Cd2+
Cd2+
7
MT1M
MT1M:7Zn2+MT1M:7Zn2+MT1M:7Zn2+
Zn2+
7
MT1M:7Cd2+MT1M:7Cd2+MT1M:7Cd2+
Cd2+
7
MT1X
MT1X:7Zn2+MT1X:7Zn2+MT1X:7Zn2+
Zn2+
7
MT1X:7Cd2+MT1X:7Cd2+MT1X:7Cd2+
Cd2+7
MT2A
MT2A:7Zn2+MT2A:7Zn2+MT2A:7Zn2+
Zn2+
7
MT2A:7Cd2+MT2A:7Cd2+MT2A:7Cd2+
Cd2+10
Cu1+
MT2A:10Cu1+MT2A:10Cu1+MT2A:10Cu1+
7
MT3
MT3:7Zn2+
Zn2+
7
MT3:7Cd2+
Cd2+12
MT3:12Cu1+
Cu1+
\\
3
4
Cu2+
MT3:4Zn2+:4Cu+
1.4E2
-1.96E1
Metallothioneins are highly conserved, cysteine-rich proteins that bind metals via thiolate bonds (recent general reviews in Capdevila et al. 2012, Blindauer et al. 2014, reviews of mammalian metal-lothioneins in Miles et al. 2000, Maret 2011, Vasak and Meloni 2011, Thirumoorthy et al. 2001, Bab-ula et al. 2012). Mammals contain 4 general metallothionein isoforms (MT1,2,3,4). The MT1 isoform has radiated in primates to 8 or 9 functional proteins (depending on classification of MT1L). Each mammalian metallothionein binds a total of 7 divalent metal ions in two clusters, the alpha and beta clusters. Though the functions of metallothioneins have not been fully elucidated, they appear to participate in detoxifying heavy metals (reviewed in Sharma et al. 2013), storing and transporting zinc, and redox biochemistry. Metallothioneins interact with many other cellular proteins, with most interactions involving proteins of the central nervous system (reviewed in Atrian and Capdev-ila 2013).
References
Miles AT, Hawksworth GM, Beattie JH & Rodilla V (2000). Induction, regulation, degradation, and biological significance of mammalian metallothioneins. Crit. Rev. Biochem. Mol. Biol., 35, 35-70
.
Babula P, Masarik M, Adam V, Eckschlager T, Stiborova M, Trnkova L, ... Kizek R (2012). Mammali-
an metallothioneins: properties and functions. Metallomics, 4, 739-50.
Vašák M & Meloni G (2011). Chemistry and biology of mammalian metallothioneins. J. Biol. Inorg.
Chem., 16, 1067-78.
Thirumoorthy N, Shyam Sunder A, Manisenthil Kumar K, Senthil Kumar M, Ganesh G & Chatterjee M (2011). A review of metallothionein isoforms and their role in pathophysiology. World J Surg
Oncol, 9, 54.
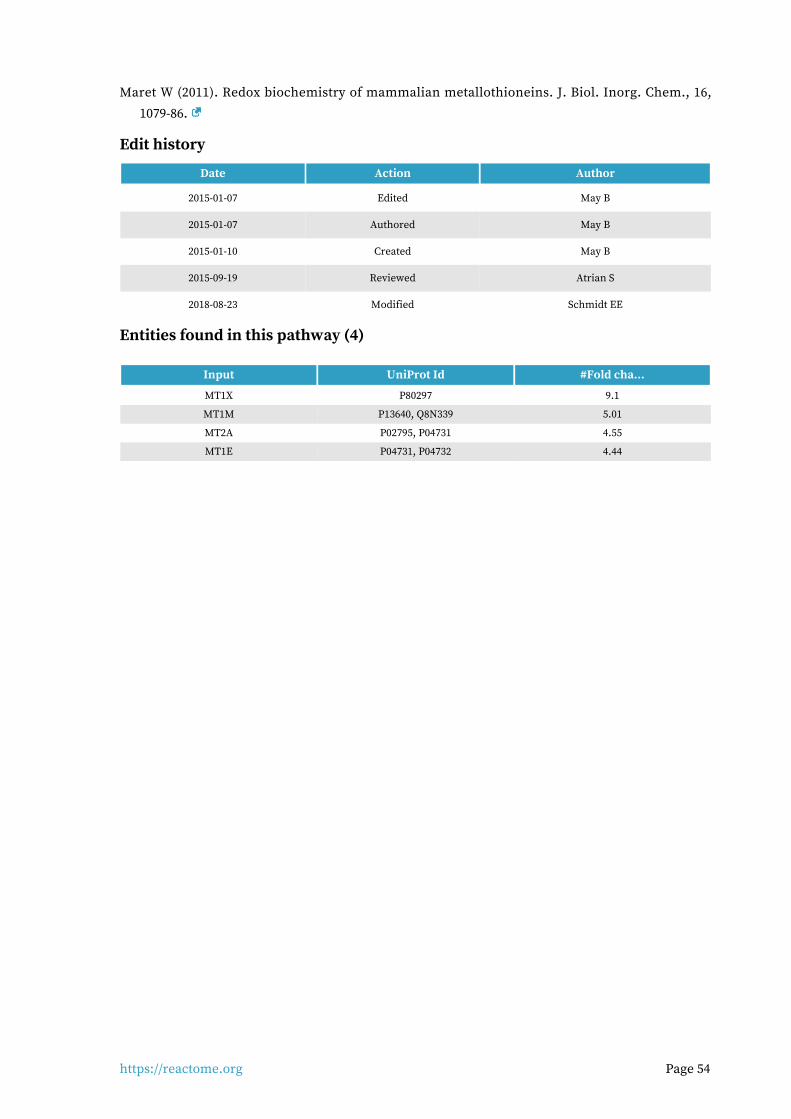
https://reactome.org Page 54
Maret W (2011). Redox biochemistry of mammalian metallothioneins. J. Biol. Inorg. Chem., 16,
1079-86.
Edit history
Date Action Author
2015-01-07 Edited May B
2015-01-07 Authored May B
2015-01-10 Created May B
2015-09-19 Reviewed Atrian S
2018-08-23 Modified Schmidt EE
Entities found in this pathway (4)
Input UniProt Id #Fold cha...
MT1X P80297 9.1
MT1M P13640, Q8N339 5.01
MT2A P02795, P04731 4.55
MT1E P04731, P04732 4.44

https://reactome.org Page 55
Defective B3GALTL causes Peters-plus syndrome (PpS) (R-HSA-5083635)22.
Golgi lumen
cytosol
endoplasmic reticulum lumen
Mucins
UDP-GalNAc
Tn antigens
UDP
Man-DAG1
POMT1:POMT2
UDP-Gal
Core 4 mucins
GCNTs
UDP
UDP
GlcNAc-Man-DAG1
UDP-GlcNAc
POMGNT2
UDP
GlcNAc,1,2-Man-DAG1
B3GNTs
UDP
POMGNT1
A4GNT
Sialyl-2,3 T antigens
UDP-GlcNAc
DOLP-Man
GalNAc-GlcNAc-Man-DAG1
GCNT3
B3GALNT2
DOLP
UDP
UDP
Core 2a mucins
Core 5 mucins
\\
GalNAc-GlcNAc-Man6P-DAG1
\\
POMK
Core 6 mucins
UDP
Core 7 mucins
Core 8 mucins
Sialyl Tn antigens
Disialyl T antigens
\\
C1GALT1:C1GALT1C1
UDP-GalNAc\\
UDP-Gal
CMP-Neu5Ac
CMP
ST6GAL1
B4GAT1:LARGE,GYLTL1B
CMP
PAPS
PAP
Core 2S mucins
ST6GALNAC2
CHST4
GALNTs
CMP-Neu5Ac
CMP
Xyl-GlcA
ST3GAL1-4 ST6GALNAC3/4
CMP-Neu5Ac
CMP
UDP-GlcNAc
DAG1(30-653)
UDP-Xyl
Xyl-GlcA-Xyl-GlcA
Core 1 mucins
UDP-GlcA
UDP-GlcNAc
UDP-Gal
UDP-GalNAc
ATP
ADP
UDP
CMP-Neu5Ac
UDP
CMP-Neu5Ac
Core 2 mucins
Core 3 mucins
UDP
UDP-GlcNAc
Man-DAG1
GlcA-Xyl-GlcA
Sialyl-2,6 T antigens
extended Core 6 mucins
B4GALT5
POFUT2 substrates
POFUT2
glc-fuc-proteins
B3GALTL
UDP-Glc
B3GALTL
O-fuc-proteinsO-fuc-proteinsO-fuc-proteins
1.4E2
-1.96E1
Diseases: eye disease, orofacial cleft.
Human beta-1,3-glucosyltransferase like protein (B3GALTL, HGNC Approved Gene Symbol: B3GLCT; MIM:610308; CAZy family GT31), localised on the ER membrane, glucosylates O-fucosylated proteins. The resultant glc-beta-1,3-fuc disaccharide modification on thrombospondin type 1 repeat (TSR1) domain-containing proteins is thought to assist in the secretion of many of these proteins from the ER lumen, and mediate an ER quality-control mechanism of folded TSRs (Vasudevan et al. 2015). Defects in B3GALTL can cause Peters plus syndrome (PpS; MIM:261540), an autosomal recessive disorder characterised by anterior eye chamber defects, short stature, delay in growth and mental developmental and cleft lip and/or palate (Heinonen & Maki 2009).
References
Heinonen TY & Maki M (2009). Peters'-plus syndrome is a congenital disorder of glycosylation caused by a defect in the beta1,3-glucosyltransferase that modifies thrombospondin type 1 re-
peats. Ann. Med., 41, 2-10.
Vasudevan D, Takeuchi H, Johar SS, Majerus E & Haltiwanger RS (2015). Peters plus syndrome
mutations disrupt a noncanonical ER quality-control mechanism. Curr. Biol., 25, 286-95.
Edit history
Date Action Author
2013-11-07 Edited Jassal B
2013-11-07 Authored Jassal B
2013-11-07 Created Jassal B
2015-12-18 Modified Jassal B
2015-12-18 Reviewed Hansen L, Joshi HJ
Entities found in this pathway (10)
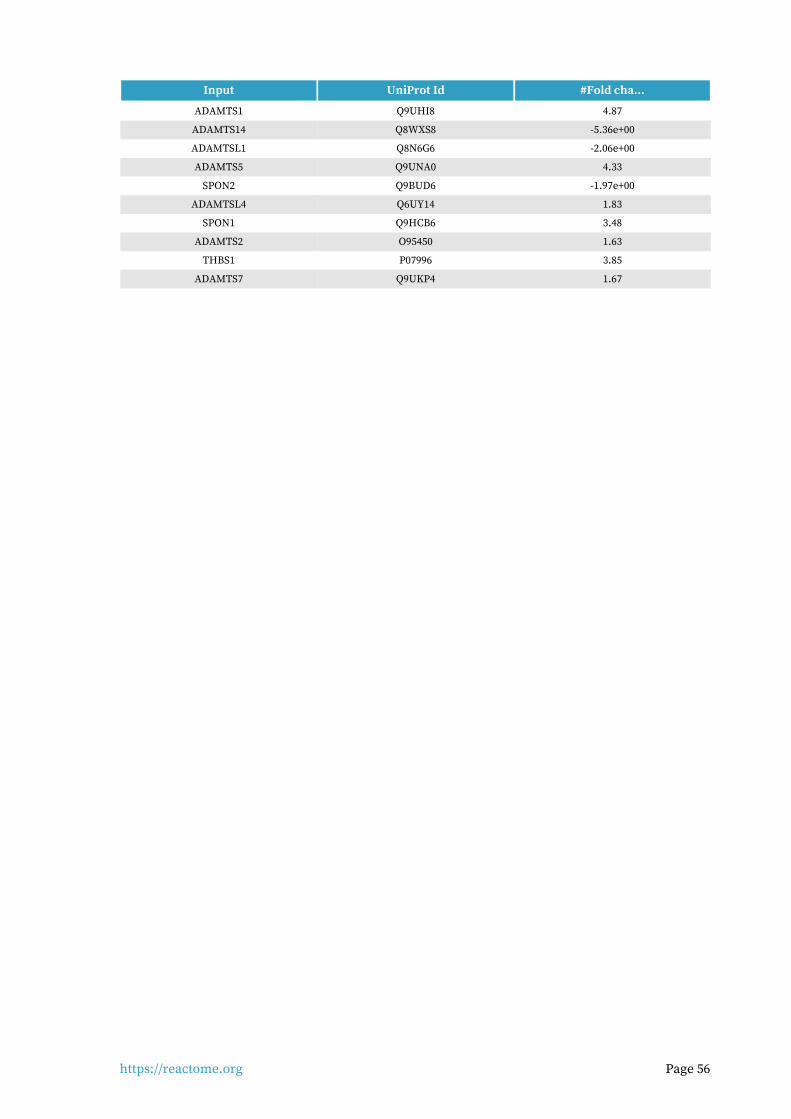
https://reactome.org Page 56
Input UniProt Id #Fold cha...
ADAMTS1 Q9UHI8 4.87
ADAMTS14 Q8WXS8 -5.36e+00
ADAMTSL1 Q8N6G6 -2.06e+00
ADAMTS5 Q9UNA0 4.33
SPON2 Q9BUD6 -1.97e+00
ADAMTSL4 Q6UY14 1.83
SPON1 Q9HCB6 3.48
ADAMTS2 O95450 1.63
THBS1 P07996 3.85
ADAMTS7 Q9UKP4 1.67

https://reactome.org Page 57
Smooth Muscle Contraction (R-HSA-445355)23.
cytosol
cytoplasmic vesicle lumen
Ca2+
ANXA2
ANXA6
ANXA1
CAV3:TRIM72:DYSF:ANXA6:CAV3:TRIM72:DYSF:ANXA6:CAV3:TRIM72:DYSF:ANXA6:ANXA1:ANXA2ANXA1:ANXA2ANXA1:ANXA2
p-S,T-PAK1,2
CALM1
4CALM1:4xCa2+
Ca2+
MYLK(1-1914)
MYLK:Ca2+:CALM1
ATP
Smooth Muscle MyosinSmooth Muscle MyosinSmooth Muscle MyosinLight ChainLight ChainLight Chain Phosphorylated SmoothPhosphorylated SmoothPhosphorylated Smooth
Muscle Myosin Light ChainMuscle Myosin Light ChainMuscle Myosin Light Chain
ADP
Inactive Myosin ActinInactive Myosin ActinInactive Myosin ActinContractile ComplexContractile ComplexContractile Complex
Calcium BoundCalcium BoundCalcium BoundMyosin Actin ComplexMyosin Actin ComplexMyosin Actin Complex
2
ADP:Calcium Bound MyosinADP:Calcium Bound MyosinADP:Calcium Bound MyosinActin ComplexActin ComplexActin Complex
ADP
2
ATP
ATP:Calcium Bound MyosinATP:Calcium Bound MyosinATP:Calcium Bound MyosinActin ComplexActin ComplexActin Complex
2
Pi
ATP Bound Smooth MuscleATP Bound Smooth MuscleATP Bound Smooth MuscleMyosin ComplexMyosin ComplexMyosin Complex
DYSF
CAV3
TRIM72
CAV3:TRIM72:DYSF
1.4E2
-1.96E1
Cellular compartments: cytosol, plasma membrane.
Layers of smooth muscle cells can be found in the walls of numerous organs and tissues within the body. Smooth muscle tissue lacks the striated banding pattern characteristic of skeletal and cardiac muscle. Smooth muscle is triggered to contract by the autonomic nervous system, hormones, auto-crine/paracrine agents, local chemical signals, and changes in load or length.
Actin:myosin cross bridging is used to develop force with the influx of calcium ions (Ca2+) initiat-ing contraction. Two separate protein pathways, both triggered by calcium influx contribute to con-traction, a calmodulin driven kinase pathway, and a caldesmon driven pathway.
Recent evidence suggests that actin, myosin, and intermediate filaments may be far more volatile then previously suspected, and that changes in these cytoskeletal elements along with alterations of the focal adhesions that anchor these proteins may contribute to the contractile cycle.
Contraction in smooth muscle generally uses a variant of the same sliding filament model found in striated muscle, except in smooth muscle the actin and myosin filaments are anchored to focal ad-hesions, and dense bodies, spread over the surface of the smooth muscle cell. When actin and my-osin move across one another focal adhesions are drawn towards dense bodies, effectively squeez-ing the cell into a smaller conformation. The sliding is triggered by calcium:caldesmon binding, caldesmon acting in an analogous fashion to troponin in striated muscle. Phosphorylation of myos-in light chains also is involved in the initiation of an effective contraction.
References
Webb RC (2003). Smooth muscle contraction and relaxation. Adv Physiol Educ, 27, 201-6.
Edit history
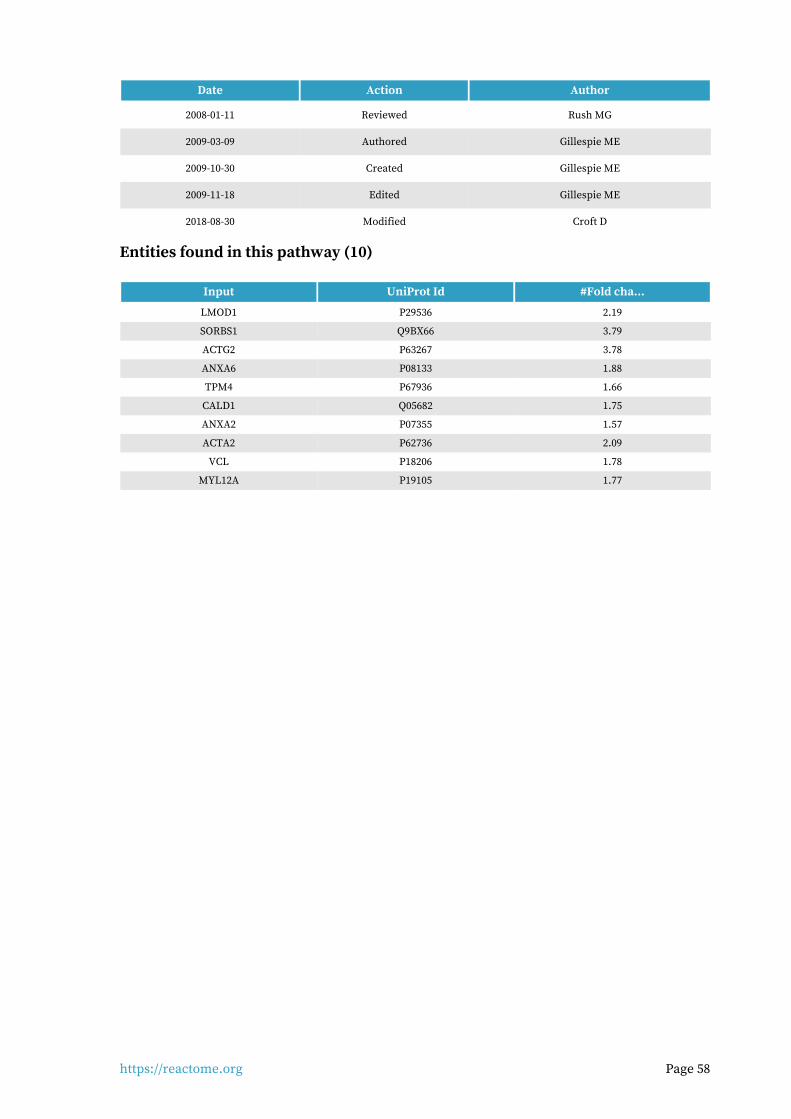
https://reactome.org Page 58
Date Action Author
2008-01-11 Reviewed Rush MG
2009-03-09 Authored Gillespie ME
2009-10-30 Created Gillespie ME
2009-11-18 Edited Gillespie ME
2018-08-30 Modified Croft D
Entities found in this pathway (10)
Input UniProt Id #Fold cha...
LMOD1 P29536 2.19
SORBS1 Q9BX66 3.79
ACTG2 P63267 3.78
ANXA6 P08133 1.88
TPM4 P67936 1.66
CALD1 Q05682 1.75
ANXA2 P07355 1.57
ACTA2 P62736 2.09
VCL P18206 1.78
MYL12A P19105 1.77

https://reactome.org Page 59
Regulation of FZD by ubiquitination (R-HSA-4641263)24.
cytosol
nucleoplasm
early endosome lumen
early endosome lumen
Ub
p-DVL
ATP ADP
\\ 26S proteasome
GSK3B:AXIN1
PIP5K1B
canonical FZDreceptors
ppDVL:PIP5K1B
PI4P
DVL:CUL3:KLHL12:RBX1
ATP
DVL
ADP
PI(4,5)P2
WNT:FZD:LRP5/6
LRP5/6
\\
ATP
DVL
WNT:FZD:LRP5/6:DVL
GSK3BFRAT1,2
ADP
FRAT1,2:GSK3beta
Casein kinase II
Ub
WNT:FZD:LRP5/6:DVL:AXIN:GSK3B
canonical WNTs
LEO1
CUL3:KLHL12:RBX1
CDC73
TCF/LEF:CTNNB1:PAF1-like complex
\\
DACT1
\\
DVL2
XPO1:YWHAZ:p-S20-CBY1:CTNNB1
DVL2:DACT1
5 5
ATP ADP
WNT:FZD:p5S/T-LRP5/6:DVL:AXIN:GSK3B
SRY,SOX2,SOX3,SOX4,SOX6,SRY,SOX2,SOX3,SOX4,SOX6,SRY,SOX2,SOX3,SOX4,SOX6,SOX7,SOX9,SOX17SOX7,SOX9,SOX17SOX7,SOX9,SOX17
5 5
ATP
CTNNB1:SRY,SOX2,SOX3,CTNNB1:SRY,SOX2,SOX3,CTNNB1:SRY,SOX2,SOX3,SOX4,SOX6,SOX7,SOX9,SOX17SOX4,SOX6,SOX7,SOX9,SOX17SOX4,SOX6,SOX7,SOX9,SOX17
ADP
WNT:FZD:p10S/T-LRP5/6:DVL:AXIN:GSK3B
SOX4,SOX13,SOX17SOX4,SOX13,SOX17SOX4,SOX13,SOX17
CSNK1G2
TCF/LEF:CTNNB1:SOX4,TCF/LEF:CTNNB1:SOX4,TCF/LEF:CTNNB1:SOX4,SOX13,SOX17SOX13,SOX17SOX13,SOX17
TNKS1/2
AXINAXINAXIN AXIN:TNKSAXIN:TNKSAXIN:TNKS
NAD+NAM
RUNX3:TCF7L2,(LEF1,TCF7L1)
p-S33,S37,T41,S45 CTNNB1:p-AXIN:CK1alpha:GSK3B:phospho-
ub-APC (20 aa repeat region):PP2A:AMER1 complex
SMURF2
Degradation of beta-catenin by thedestruction complex
AXIN:SMURF2AXIN:SMURF2AXIN:SMURF2
MYC
Ub
ub-AXIN:SMURF2ub-AXIN:SMURF2ub-AXIN:SMURF2
CCDC88C
DVL:CCDC88C
Ub
\\
CTNNBIP1
CTNNBIP1:CTNNB1
\\
WNT ligand biogenesis and trafficking
RibC-AXIN:TNKS:RNF146RibC-AXIN:TNKS:RNF146RibC-AXIN:TNKS:RNF146
SMARCA4
TERT
TCF/LEF:CTNNB1:SMARCA4:TERT
RibC-AXIN:TNKSRibC-AXIN:TNKSRibC-AXIN:TNKS
H2O
WIF1
USP34
WIF1-binding WNTs
CSNK1E
WIF1:WNT
TCF/LEF:CTNNB1:BCL9:PYGO
MEN1ASH2LKMT2D
ZNRF3,RNF43
RBBP5
TCF/LEF:CTNNB1:BCL9:PYGO:SET1-
like complex
MYC gene
TCF7L1/TCF7L2/LEF1:CTNNB1
DKK
TCF7L1/TCF7L2/LEF1:CTNNB1:MYCTCF7L1/TCF7L2/LEF1:CTNNB1:MYCTCF7L1/TCF7L2/LEF1:CTNNB1:MYCgenegenegene
KRM1/2
KRM:DKK:LRP5/6
\\
CTBP1
APC
TCF/LEF:CTNNB1:APC:CTBP:BTRC
\\
TCF/LEF:TLE:TCF/LEF:TLE:TCF/LEF:TLE:HDAC1HDAC1HDAC1 TCF/LEF:CTNNB1
CBY1
CBY1:CTNNB1
BCL9PYGO
TLE1:HDAC1TLE1:HDAC1TLE1:HDAC1
APC:CTBP:CTNNB1:BTRC
pS20-CBY1:CTNNB1
ATP
HDAC1
ADP
TLE tetramerTLE tetramerTLE tetramer
p-AKT1/2
AXIN2 gene
TCF/LEF:CTNNB1:AXIN2 geneTCF/LEF:CTNNB1:AXIN2 geneTCF/LEF:CTNNB1:AXIN2 gene
YWHAZ
TCF/LEF:CTNNB1
YWHAZ:p-S20-CBY:CTNNB1
\\
N4GlycoAsn-PalmS-WNT3A:N4GlycoAsn-PalmS-WNT3A:N4GlycoAsn-PalmS-WNT3A:FZD:LRP5/6FZD:LRP5/6FZD:LRP5/6
AXIN2
N4GlycoAsn-PalmS-WNT3A:N4GlycoAsn-PalmS-WNT3A:N4GlycoAsn-PalmS-WNT3A:FZD:LRP5/6:ZNRF3/RNF43FZD:LRP5/6:ZNRF3/RNF43FZD:LRP5/6:ZNRF3/RNF43
N4GlycoAsn-PalmS-WNT3A:N4GlycoAsn-PalmS-WNT3A:N4GlycoAsn-PalmS-WNT3A:ub-FZD:LRP5/6:ZNRF3/ub-FZD:LRP5/6:ZNRF3/ub-FZD:LRP5/6:ZNRF3/
RNF43RNF43RNF43
Ub
H2O
N4GlycoAsn-PalmS-WNT3A:N4GlycoAsn-PalmS-WNT3A:N4GlycoAsn-PalmS-WNT3A:ub-FZD:LRP5/6ub-FZD:LRP5/6ub-FZD:LRP5/6
XIAP
USP8
TLE:XIAPTLE:XIAPTLE:XIAP
XPO1
XPO1:YWHAZ:p-S20-CBY1:CTNNB1
Ub
ub-TLE:XIAPub-TLE:XIAPub-TLE:XIAP
CTNNB1:AXIN:GSK3:CK1alpha:ub-APC:PP2A:AMER1 complex
ub-TLEub-TLEub-TLE
SOST
SOST:LRP5/6
6 6MYC gene: H2BMYC gene: H2BMYC gene: H2B
K121ub nucleosomeK121ub nucleosomeK121ub nucleosome
AdoMet
MYC gene:H2B K121ubMYC gene:H2B K121ubMYC gene:H2B K121ubH3K4me3 nucleosomeH3K4me3 nucleosomeH3K4me3 nucleosome
AdoHcy
BTRC
TRRAP
RUVBL1
KAT5 TCF/LEF:CTNNB1:RUVBL1:TRRAP:KAT5
2 2
TCF4 gene:nucleosome
Ac-CoA
TCF4 gene:acetyl H4nucleosome
CoA-SH
CTNNB1
CTNNB1
sFRP
N4GlycoAsn-PalmS WNT3A
WNT3A:sFRP
LGRLGRLGRRSPORSPORSPO
RSPO:LGRRSPO:LGRRSPO:LGR RSPO:LGR:ZNRF3,RSPO:LGR:ZNRF3,RSPO:LGR:ZNRF3,RNF43RNF43RNF43
RSPO:LGR:ub-RSPO:LGR:ub-RSPO:LGR:ub-ZNRF3,RNF43ZNRF3,RNF43ZNRF3,RNF43
Ub
K63polyUb-APC
\\
WNT3A:FZD5:p-LRP6:WNT3A:FZD5:p-LRP6:WNT3A:FZD5:p-LRP6:GSK3B:AXIN:CAV1GSK3B:AXIN:CAV1GSK3B:AXIN:CAV1
WNT3A:FZD5:p-LRP6:WNT3A:FZD5:p-LRP6:WNT3A:FZD5:p-LRP6:GSK3B:AXIN:CAV1GSK3B:AXIN:CAV1GSK3B:AXIN:CAV1
CSNK1A1
PP2A
AMER1
\\
KRM:DKK:LRP5/6
\\
CREBBP, EP300
TCF/LEF:CTNNB1:CBP/p300
ub-DVL:CUL3:KLHL12:RBX1
RYKRYK-binding WNTs
RYK:WNTs
pp-DVL
CHD8
CTNNB1:CHD8
DVL1
HECW1
DVL1:HECW1
CXXC4
RNF146
DVL:CXXC4
Ub-RibC-AXIN:TNKS:RNF146Ub-RibC-AXIN:TNKS:RNF146Ub-RibC-AXIN:TNKS:RNF146
ub-DVL1:HECW1
\\
1.4E2
-1.96E1
WNT responsiveness is influenced by expression levels of FZD and LRP proteins. Levels of these receptors at the cell surface are regulated in part by endocytosis, but the mechanisms are not fully elucidated (Garliardi et al, 2008). A number of recent studies have identified a role for ubiquitina-tion in the localization and turnover of WNT receptors at the plasma membrane. ZNRF3 and RNF43 are E3 ligases that have been shown to ubiquitinate FZD proteins and promote their lyso-somal degradation, while the deubiquitinating enzyme USP8 promotes recycling of the receptor back to the plasma membrane (Hao et al, 2012; Mukai et al, 2010). This balance of ubiquitination and deubiquitination is in turn regulated by the R-spondin (RSPO) proteins, agonists of WNT signal-ing which appear to act by downregulating ZNRF3 and RNF43, thus potentiating both canonical and non-canonical pathways (Hao et al, 2012; reviewed in Abo and Clevers, 2012; Fearon and Spence, 2012, Papartriantafyllou, 2012).
References
Hao HX, Xie Y, Zhang Y, Charlat O, Oster E, Avello M, ... Cong F (2012). ZNRF3 promotes Wnt re-
ceptor turnover in an R-spondin-sensitive manner. Nature, 485, 195-200.
Mukai A, Yamamoto-Hino M, Awano W, Watanabe W, Komada M & Goto S (2010). Balanced ubi-quitylation and deubiquitylation of Frizzled regulate cellular responsiveness to Wg/Wnt. EMBO
J., 29, 2114-25.
Gagliardi M, Piddini E & Vincent JP (2008). Endocytosis: a positive or a negative influence on Wnt
signalling?. Traffic, 9, 1-9.
Abo A & Clevers HC (2012). Modulating WNT receptor turnover for tissue repair. Nat. Biotechnol.,
30, 835-6.
Fearon ER & Spence JR (2012). Cancer biology: a new RING to Wnt signaling. Curr. Biol., 22, R849-
51.
Edit history
Date Action Author
2007-09-04 Edited Matthews L
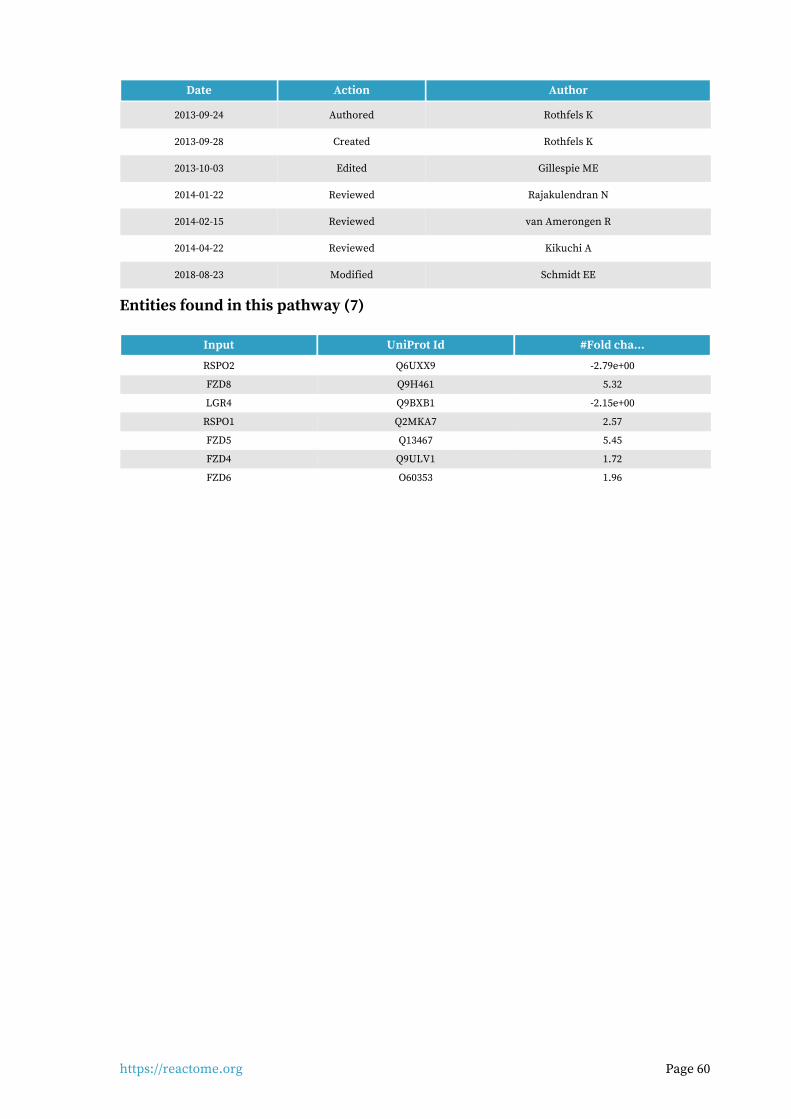
https://reactome.org Page 60
Date Action Author
2013-09-24 Authored Rothfels K
2013-09-28 Created Rothfels K
2013-10-03 Edited Gillespie ME
2014-01-22 Reviewed Rajakulendran N
2014-02-15 Reviewed van Amerongen R
2014-04-22 Reviewed Kikuchi A
2018-08-23 Modified Schmidt EE
Entities found in this pathway (7)
Input UniProt Id #Fold cha...
RSPO2 Q6UXX9 -2.79e+00
FZD8 Q9H461 5.32
LGR4 Q9BXB1 -2.15e+00
RSPO1 Q2MKA7 2.57
FZD5 Q13467 5.45
FZD4 Q9ULV1 1.72
FZD6 O60353 1.96

https://reactome.org Page 61
Collagen formation (R-HSA-1474290)25.

https://reactome.org Page 62
Collagen is a family of at least 29 structural proteins derived from over 40 human genes (Myllyharju & Kivirikko 2004). It is the main component of connective tissue, and the most abundant protein in mammals making up about 25% to 35% of whole-body protein content. A defining feature of colla-gens is the formation of trimeric left-handed polyproline II-type helical collagenous regions. The packing within these regions is made possible by the presence of the smallest amino acid, glycine, at every third residue, resulting in a repeating motif Gly-X-Y where X is often proline (Pro) and Y of-ten 4-hydroxyproline (4Hyp). Gly-Pro-Hyp is the most common triplet in collagen (Ramshaw et al. 1998). Collagen peptide chains also have non-collagenous domains, with collagen subclasses having common chain structures. Collagen fibrils are mostly found in fibrous tissues such as tendon, liga-ment and skin. Other forms of collagen are abundant in cornea, cartilage, bone, blood vessels, the gut, and intervertebral disc. In muscle tissue, collagen is a major component of the endomysium, constituting up to 6% of muscle mass. Gelatin, used in food and industry, is collagen that has been irreversibly hydrolyzed. On the basis of their fibre architecture in tissues, the genetically distinct collagens have been di-vided into subgroups. Group 1 collagens have uninterrupted triple-helical domains of about 300 nm, forming large extracellular fibrils. They are referred to as the fibril-forming collagens, consist-ing of collagens types I, II, III, V, XI, XXIV and XXVII. Group 2 collagens are types IV and VII, which have extended triple helices (>350 nm) with imperfections in the Gly-X-Y repeat sequences. Group 3 are the short-chain collagens. These have two subgroups. Group 3A have continuous triple-helical domains (type VI, VIII and X). Group 3B have interrupted triple-helical domains, referred to as the fibril-associated collagens with interrupted triple helices (FACIT collagens, Shaw & Olsen 1991). FACITs include collagen IX, XII, XIV, XVI, XIX, XX, XXI, XXII and XXVI plus the transmem-brane collagens (XIII, XVII, XXIII and XXV) and the multiple triple helix domains and interruptions (Multiplexin) collagens XV and XVIII (Myllyharju & Kivirikko 2004). The non-collagenous domains of collagens have regulatory functions; several are biologically active when cleaved from the main peptide chain. Fibrillar collagen peptides all have a large triple helical domain (COL1) bordered by N and C terminal extensions, called the N- and C-propeptides, which are cleaved prior to formation of the collagen fibril. The intact form is referred to as a collagen propeptide, not procollagen, which is used to refer to the trimeric triple-helical precursor of collagen before the propeptides are removed. The C-propeptide, also called the NC1 domain, directs chain association during assembly of the procollagen molecule from its three constituent alpha chains (Hulmes 2002).
Fibril forming collagens are the most familiar and best studied subgroup. Collagen fibres are ag-gregates or bundles of collagen fibrils, which are themselves polymers of tropocollagen complexes, each consisting of three polypeptide chains known as alpha chains. Tropocollagens are considered the subunit of larger collagen structures. They are approximately 300 nm long and 1.5 nm in dia-meter, with a left-handed triple-helical structure, which becomes twisted into a right-handed coiled-coil 'super helix' in the collagen fibril. Tropocollagens in the extracellular space polymerize spontaneously with regularly staggered ends (Hulmes 2002). In fibrillar collagens the molecules are staggered by about 67 nm, a unit known as D that changes depending upon the hydration state. Each D-period contains slightly more than four collagen molecules so that every D-period repeat of the microfibril has a region containing five molecules in cross-section, called the 'overlap', and a re-gion containing only four molecules, called the 'gap'. The triple-helices are arranged in a hexagonal or quasi-hexagonal array in cross-section, in both the gap and overlap regions (Orgel et al. 2006). Collagen molecules cross-link covalently to each other via lysine and hydroxylysine side chains. These cross-links are unusual, occuring only in collagen and elastin, a related protein.

https://reactome.org Page 63
The macromolecular structures of collagen are diverse. Several group 3 collagens associate with larger collagen fibers, serving as molecular bridges which stabilize the organization of the extracel-lular matrix. Type IV collagen is arranged in an interlacing network within the dermal-epidermal junction and vascular basement membranes. Type VI collagen forms distinct microfibrils called beaded filaments. Type VII collagen forms anchoring fibrils. Type VIII and X collagens form hexagonal networks. Type XVII collagen is a component of hemidesmosomes where it is com-plexed wtih alpha6Beta4 integrin, plectin, and laminin-332 (de Pereda et al. 2009). Type XXIX colla-gen has been recently reported to be a putative epidermal collagen with highest expression in suprabasal layers (Soderhall et al. 2007). Collagen fibrils/aggregates arranged in varying combina-tions and concentrations in different tissues provide specific tissue properties. In bone, collagen triple helices lie in a parallel, staggered array with 40 nm gaps between the ends of the tropocolla-gen subunits, which probably serve as nucleation sites for the deposition of crystals of the mineral component, hydroxyapatite (Ca10(PO4)6(OH)2) with some phosphate. Collagen structure affects cell-cell and cell-matrix communication, tissue construction in growth and repair, and is changed in development and disease (Sweeney et al. 2006, Twardowski et al. 2007). A single collagen fibril can be heterogeneous along its axis, with significantly different mechanical properties in the gap and overlap regions, correlating with the different molecular organizations in these regions (Mi-nary-Jolandan & Yu 2009).
References
Prockop DJ & Kivirikko KI (1995). Collagens: molecular biology, diseases, and potentials for therapy
. Annu Rev Biochem, 64, 403-34.
Gordon MK & Hahn RA (2010). Collagens. Cell Tissue Res, 339, 247-57.
Edit history
Date Action Author
2011-08-05 Authored Jupe S
2011-08-05 Created Jupe S
2012-04-11 Edited Jupe S
2012-05-24 Reviewed Canty-Laird EG
2018-08-23 Modified Schmidt EE
Entities found in this pathway (19)
Input UniProt Id #Fold cha...
SERPINH1 P50454 -1.61e+00
COL4A5 P29400 -1.87e+00
PCOLCE Q15113 -1.55e+00
COL4A3 Q01955 4.74
LAMA3 Q16787 1.82
COL4A4 P53420 3.93
COL7A1 Q02388 2.34
COL5A3 P25940 1.9
COL4A1 P02462 2.52
COL11A1 P12107 5.14
COL4A2 P08572 1.85

https://reactome.org Page 64
Input UniProt Id #Fold cha...
ADAMTS2 O95450 1.63
PLEC Q15149 1.79
ADAMTS14 Q8WXS8 -5.36e+00
COL1A1 P02452 -2.48e+00
COL3A1 P02461 -1.81e+00
COL27A1 Q8IZC6 -1.98e+00
COL15A1 P39059 -1.90e+00
MMP7 P09237 28.84
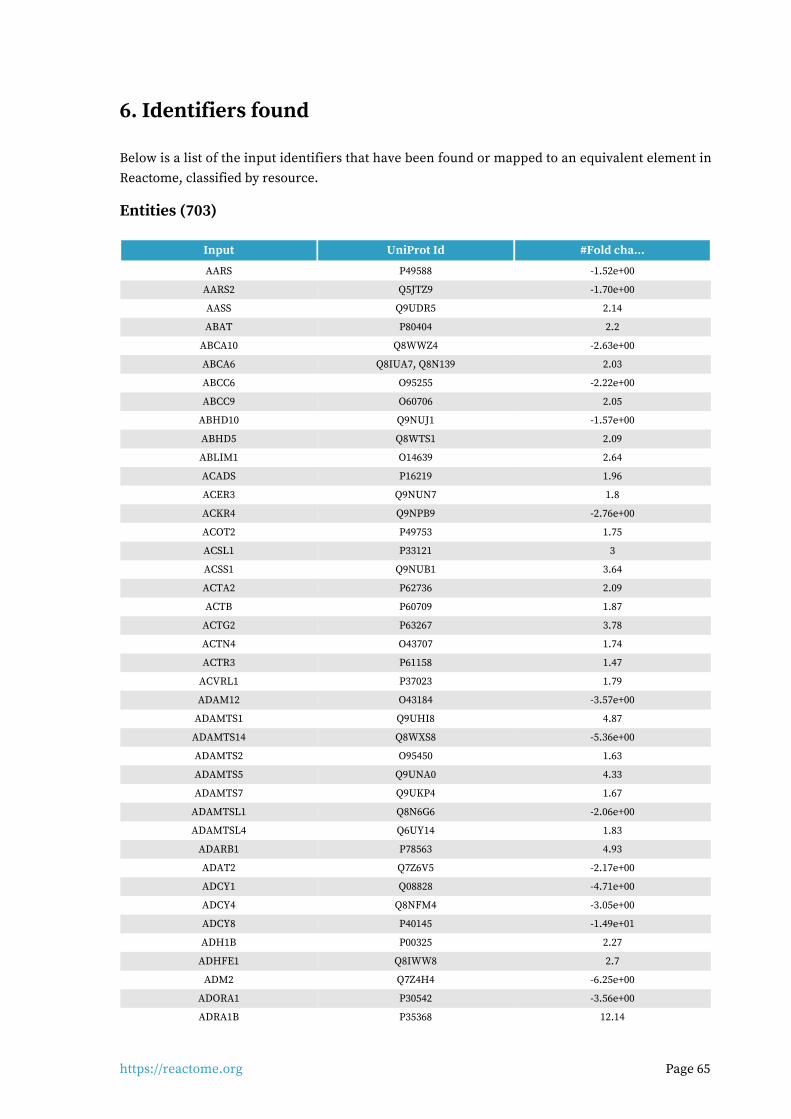
https://reactome.org Page 65
6. Identifiers found
Below is a list of the input identifiers that have been found or mapped to an equivalent element in Reactome, classified by resource.
Entities (703)
Input UniProt Id #Fold cha...
AARS P49588 -1.52e+00
AARS2 Q5JTZ9 -1.70e+00
AASS Q9UDR5 2.14
ABAT P80404 2.2
ABCA10 Q8WWZ4 -2.63e+00
ABCA6 Q8IUA7, Q8N139 2.03
ABCC6 O95255 -2.22e+00
ABCC9 O60706 2.05
ABHD10 Q9NUJ1 -1.57e+00
ABHD5 Q8WTS1 2.09
ABLIM1 O14639 2.64
ACADS P16219 1.96
ACER3 Q9NUN7 1.8
ACKR4 Q9NPB9 -2.76e+00
ACOT2 P49753 1.75
ACSL1 P33121 3
ACSS1 Q9NUB1 3.64
ACTA2 P62736 2.09
ACTB P60709 1.87
ACTG2 P63267 3.78
ACTN4 O43707 1.74
ACTR3 P61158 1.47
ACVRL1 P37023 1.79
ADAM12 O43184 -3.57e+00
ADAMTS1 Q9UHI8 4.87
ADAMTS14 Q8WXS8 -5.36e+00
ADAMTS2 O95450 1.63
ADAMTS5 Q9UNA0 4.33
ADAMTS7 Q9UKP4 1.67
ADAMTSL1 Q8N6G6 -2.06e+00
ADAMTSL4 Q6UY14 1.83
ADARB1 P78563 4.93
ADAT2 Q7Z6V5 -2.17e+00
ADCY1 Q08828 -4.71e+00
ADCY4 Q8NFM4 -3.05e+00
ADCY8 P40145 -1.49e+01
ADH1B P00325 2.27
ADHFE1 Q8IWW8 2.7
ADM2 Q7Z4H4 -6.25e+00
ADORA1 P30542 -3.56e+00
ADRA1B P35368 12.14
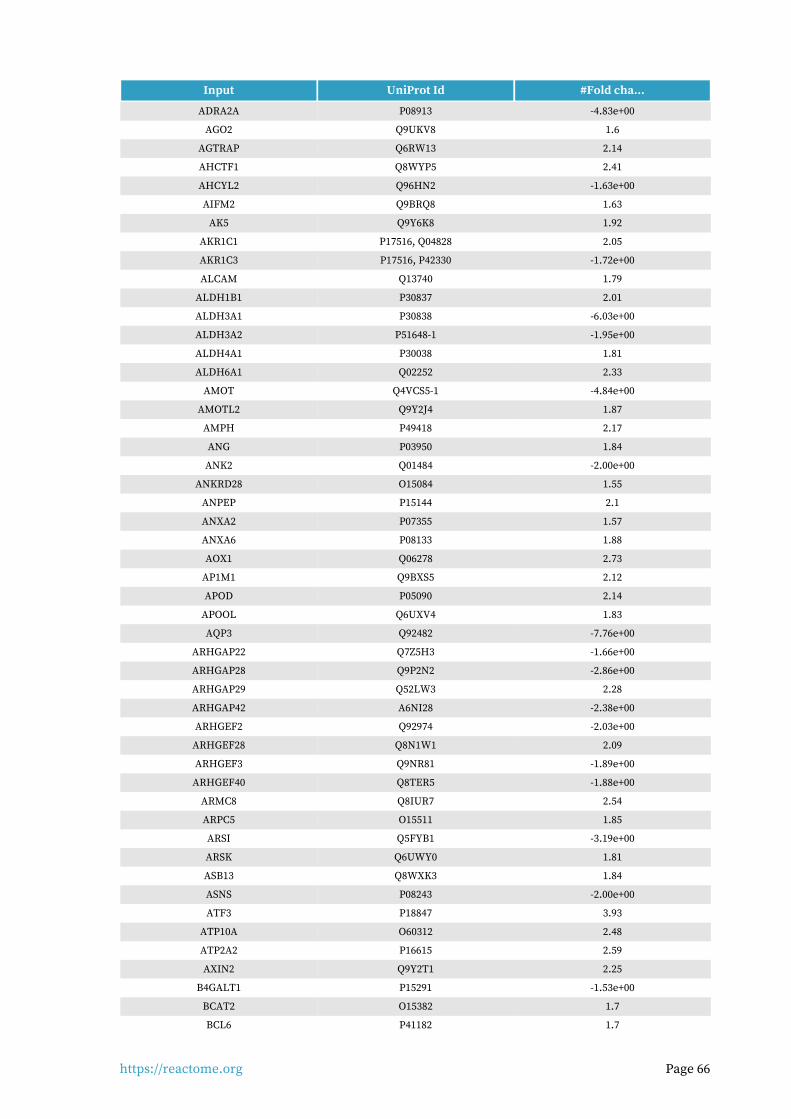
https://reactome.org Page 66
Input UniProt Id #Fold cha...
ADRA2A P08913 -4.83e+00
AGO2 Q9UKV8 1.6
AGTRAP Q6RW13 2.14
AHCTF1 Q8WYP5 2.41
AHCYL2 Q96HN2 -1.63e+00
AIFM2 Q9BRQ8 1.63
AK5 Q9Y6K8 1.92
AKR1C1 P17516, Q04828 2.05
AKR1C3 P17516, P42330 -1.72e+00
ALCAM Q13740 1.79
ALDH1B1 P30837 2.01
ALDH3A1 P30838 -6.03e+00
ALDH3A2 P51648-1 -1.95e+00
ALDH4A1 P30038 1.81
ALDH6A1 Q02252 2.33
AMOT Q4VCS5-1 -4.84e+00
AMOTL2 Q9Y2J4 1.87
AMPH P49418 2.17
ANG P03950 1.84
ANK2 Q01484 -2.00e+00
ANKRD28 O15084 1.55
ANPEP P15144 2.1
ANXA2 P07355 1.57
ANXA6 P08133 1.88
AOX1 Q06278 2.73
AP1M1 Q9BXS5 2.12
APOD P05090 2.14
APOOL Q6UXV4 1.83
AQP3 Q92482 -7.76e+00
ARHGAP22 Q7Z5H3 -1.66e+00
ARHGAP28 Q9P2N2 -2.86e+00
ARHGAP29 Q52LW3 2.28
ARHGAP42 A6NI28 -2.38e+00
ARHGEF2 Q92974 -2.03e+00
ARHGEF28 Q8N1W1 2.09
ARHGEF3 Q9NR81 -1.89e+00
ARHGEF40 Q8TER5 -1.88e+00
ARMC8 Q8IUR7 2.54
ARPC5 O15511 1.85
ARSI Q5FYB1 -3.19e+00
ARSK Q6UWY0 1.81
ASB13 Q8WXK3 1.84
ASNS P08243 -2.00e+00
ATF3 P18847 3.93
ATP10A O60312 2.48
ATP2A2 P16615 2.59
AXIN2 Q9Y2T1 2.25
B4GALT1 P15291 -1.53e+00
BCAT2 O15382 1.7
BCL6 P41182 1.7
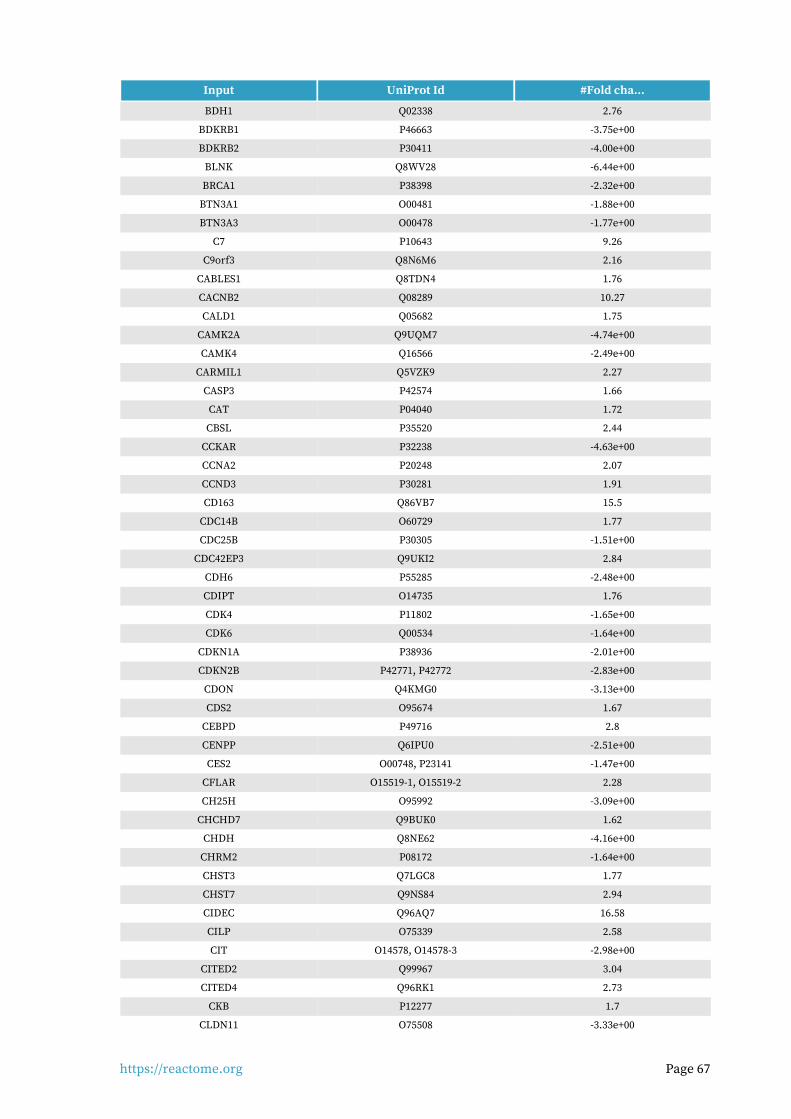
https://reactome.org Page 67
Input UniProt Id #Fold cha...
BDH1 Q02338 2.76
BDKRB1 P46663 -3.75e+00
BDKRB2 P30411 -4.00e+00
BLNK Q8WV28 -6.44e+00
BRCA1 P38398 -2.32e+00
BTN3A1 O00481 -1.88e+00
BTN3A3 O00478 -1.77e+00
C7 P10643 9.26
C9orf3 Q8N6M6 2.16
CABLES1 Q8TDN4 1.76
CACNB2 Q08289 10.27
CALD1 Q05682 1.75
CAMK2A Q9UQM7 -4.74e+00
CAMK4 Q16566 -2.49e+00
CARMIL1 Q5VZK9 2.27
CASP3 P42574 1.66
CAT P04040 1.72
CBSL P35520 2.44
CCKAR P32238 -4.63e+00
CCNA2 P20248 2.07
CCND3 P30281 1.91
CD163 Q86VB7 15.5
CDC14B O60729 1.77
CDC25B P30305 -1.51e+00
CDC42EP3 Q9UKI2 2.84
CDH6 P55285 -2.48e+00
CDIPT O14735 1.76
CDK4 P11802 -1.65e+00
CDK6 Q00534 -1.64e+00
CDKN1A P38936 -2.01e+00
CDKN2B P42771, P42772 -2.83e+00
CDON Q4KMG0 -3.13e+00
CDS2 O95674 1.67
CEBPD P49716 2.8
CENPP Q6IPU0 -2.51e+00
CES2 O00748, P23141 -1.47e+00
CFLAR O15519-1, O15519-2 2.28
CH25H O95992 -3.09e+00
CHCHD7 Q9BUK0 1.62
CHDH Q8NE62 -4.16e+00
CHRM2 P08172 -1.64e+00
CHST3 Q7LGC8 1.77
CHST7 Q9NS84 2.94
CIDEC Q96AQ7 16.58
CILP O75339 2.58
CIT O14578, O14578-3 -2.98e+00
CITED2 Q99967 3.04
CITED4 Q96RK1 2.73
CKB P12277 1.7
CLDN11 O75508 -3.33e+00
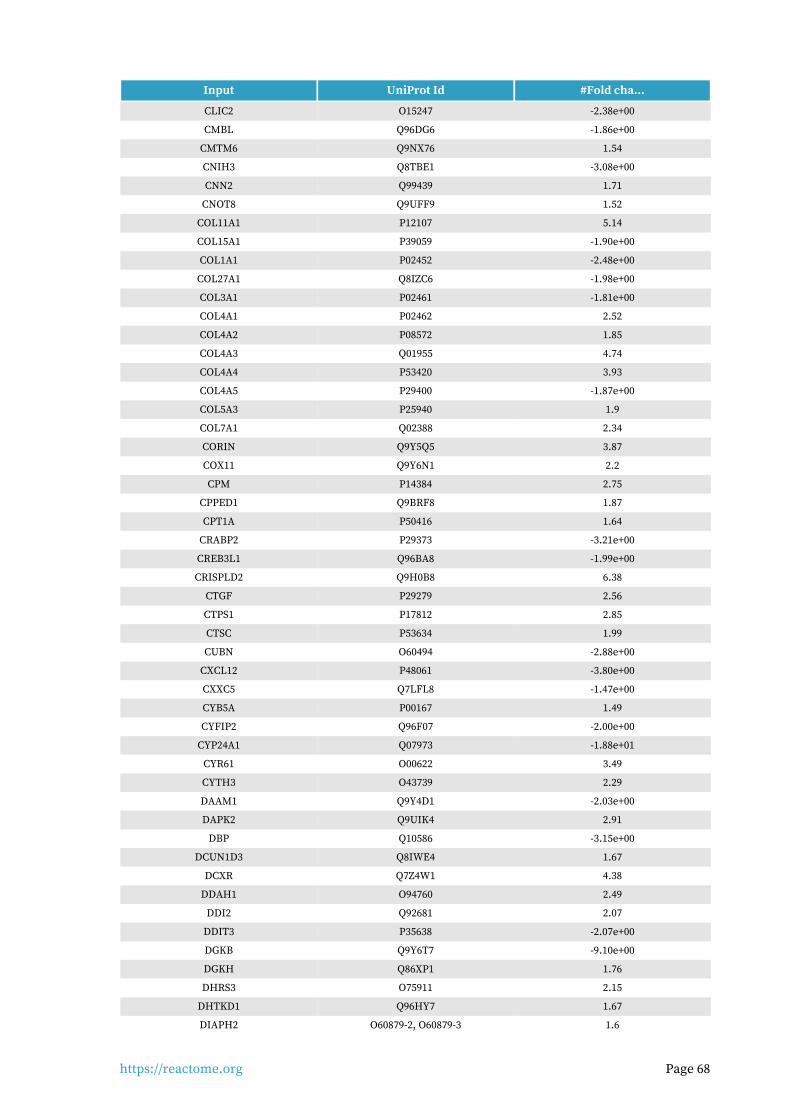
https://reactome.org Page 68
Input UniProt Id #Fold cha...
CLIC2 O15247 -2.38e+00
CMBL Q96DG6 -1.86e+00
CMTM6 Q9NX76 1.54
CNIH3 Q8TBE1 -3.08e+00
CNN2 Q99439 1.71
CNOT8 Q9UFF9 1.52
COL11A1 P12107 5.14
COL15A1 P39059 -1.90e+00
COL1A1 P02452 -2.48e+00
COL27A1 Q8IZC6 -1.98e+00
COL3A1 P02461 -1.81e+00
COL4A1 P02462 2.52
COL4A2 P08572 1.85
COL4A3 Q01955 4.74
COL4A4 P53420 3.93
COL4A5 P29400 -1.87e+00
COL5A3 P25940 1.9
COL7A1 Q02388 2.34
CORIN Q9Y5Q5 3.87
COX11 Q9Y6N1 2.2
CPM P14384 2.75
CPPED1 Q9BRF8 1.87
CPT1A P50416 1.64
CRABP2 P29373 -3.21e+00
CREB3L1 Q96BA8 -1.99e+00
CRISPLD2 Q9H0B8 6.38
CTGF P29279 2.56
CTPS1 P17812 2.85
CTSC P53634 1.99
CUBN O60494 -2.88e+00
CXCL12 P48061 -3.80e+00
CXXC5 Q7LFL8 -1.47e+00
CYB5A P00167 1.49
CYFIP2 Q96F07 -2.00e+00
CYP24A1 Q07973 -1.88e+01
CYR61 O00622 3.49
CYTH3 O43739 2.29
DAAM1 Q9Y4D1 -2.03e+00
DAPK2 Q9UIK4 2.91
DBP Q10586 -3.15e+00
DCUN1D3 Q8IWE4 1.67
DCXR Q7Z4W1 4.38
DDAH1 O94760 2.49
DDI2 Q92681 2.07
DDIT3 P35638 -2.07e+00
DGKB Q9Y6T7 -9.10e+00
DGKH Q86XP1 1.76
DHRS3 O75911 2.15
DHTKD1 Q96HY7 1.67
DIAPH2 O60879-2, O60879-3 1.6

https://reactome.org Page 69
Input UniProt Id #Fold cha...
DIO2 Q92813 2.89
DLG3 Q92796 -1.57e+00
DLL4 Q9NR61 12.92
DMC1 Q14565 3.54
DMD P11532 1.97
DMPK Q09013 1.81
DNAJC6 O75061 4.06
DNM1 Q05193 -3.37e+00
DOCK5 Q9H7D0 1.69
DOK5 Q9P104 -1.97e+00
DOK6 Q6PKX4 -2.21e+00
DPYSL3 Q14195 -1.53e+00
DUSP1 P28562 7.64
DUSP10 Q9Y6W6 -2.29e+00
DUSP5 Q16690 2.44
EBF1 Q9UH73 2.17
EDA Q92838, Q92838-3 2.84
EDA2R Q9HAV5 -1.95e+00
EGR2 P11161 -8.56e+00
EHMT2 Q96KQ7 -1.44e+00
ELF1 O43921 1.88
ENDOD1 O94919 3.86
ENPP1 P22413 2.52
ENTPD4 Q9Y227 1.5
EPB41 P11171 -2.05e+00
EPHA4 P54764 2.55
EPHB2 P29323 -3.38e+00
EPHB3 P54753 -2.72e+00
EPHB4 P54760 -1.92e+00
EPHB6 O15197 2.5
EXOSC10 Q01780 1.56
EXT1 Q16394 -1.81e+00
FABP4 P15090 9.39
FAM20A Q96MK3 2.14
FAM20C Q8IXL6 1.93
FBN2 P35556 4.2
FBXO32 Q969P5 2.14
FDXR P22570 -1.96e+00
FERMT2 Q96AC1 1.53
FGD4 Q96M96 4.77
FGF14 Q92915 2
FGF22 Q9HCT0 6.6
FGFR2 P21802-1, P21802-18, P21802-3, P21802-5 -4.53e+00
FKBP5 Q13451 15.25
FLRT3 Q9NZU0 -5.17e+00
FNBP1 Q96RU3 1.78
FOXO1 Q12778 4.91
FOXO3 O43524 2.41
FPGS Q05932-1, Q05932-2 1.54
FRK P42685 -2.24e+00
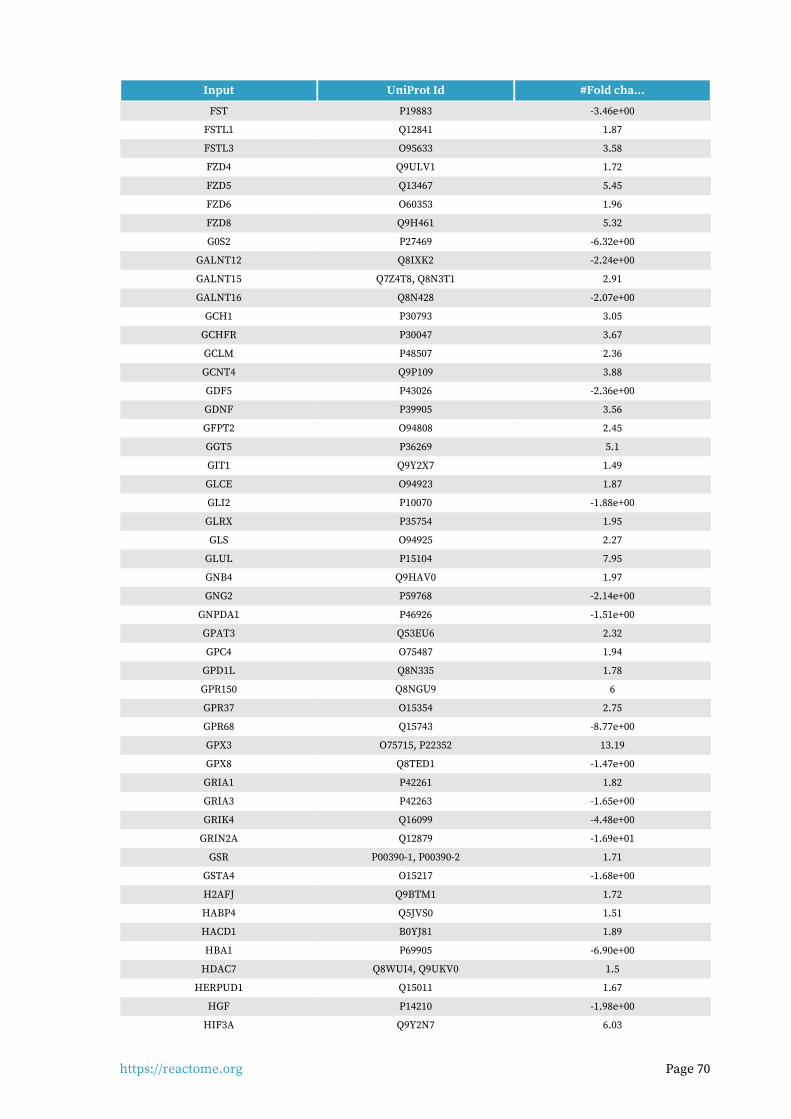
https://reactome.org Page 70
Input UniProt Id #Fold cha...
FST P19883 -3.46e+00
FSTL1 Q12841 1.87
FSTL3 O95633 3.58
FZD4 Q9ULV1 1.72
FZD5 Q13467 5.45
FZD6 O60353 1.96
FZD8 Q9H461 5.32
G0S2 P27469 -6.32e+00
GALNT12 Q8IXK2 -2.24e+00
GALNT15 Q7Z4T8, Q8N3T1 2.91
GALNT16 Q8N428 -2.07e+00
GCH1 P30793 3.05
GCHFR P30047 3.67
GCLM P48507 2.36
GCNT4 Q9P109 3.88
GDF5 P43026 -2.36e+00
GDNF P39905 3.56
GFPT2 O94808 2.45
GGT5 P36269 5.1
GIT1 Q9Y2X7 1.49
GLCE O94923 1.87
GLI2 P10070 -1.88e+00
GLRX P35754 1.95
GLS O94925 2.27
GLUL P15104 7.95
GNB4 Q9HAV0 1.97
GNG2 P59768 -2.14e+00
GNPDA1 P46926 -1.51e+00
GPAT3 Q53EU6 2.32
GPC4 O75487 1.94
GPD1L Q8N335 1.78
GPR150 Q8NGU9 6
GPR37 O15354 2.75
GPR68 Q15743 -8.77e+00
GPX3 O75715, P22352 13.19
GPX8 Q8TED1 -1.47e+00
GRIA1 P42261 1.82
GRIA3 P42263 -1.65e+00
GRIK4 Q16099 -4.48e+00
GRIN2A Q12879 -1.69e+01
GSR P00390-1, P00390-2 1.71
GSTA4 O15217 -1.68e+00
H2AFJ Q9BTM1 1.72
HABP4 Q5JVS0 1.51
HACD1 B0YJ81 1.89
HBA1 P69905 -6.90e+00
HDAC7 Q8WUI4, Q9UKV0 1.5
HERPUD1 Q15011 1.67
HGF P14210 -1.98e+00
HIF3A Q9Y2N7 6.03

https://reactome.org Page 71
Input UniProt Id #Fold cha...
HIGD1A Q9Y241 2.26
HIPK2 Q9H2X6 1.51
HMGB2 P26583 2.36
HMOX1 P09601 -2.54e+00
HPS5 P0DJI8 3
HRH1 P35367 -1.61e+00
HS2ST1 Q7LGA3 1.79
HS3ST3B1 Q9Y662 1.95
HSD17B11 Q14515 1.69
HSD17B6 O14756 -4.53e+00
HSPA12B Q96MM6 -5.32e+00
HSPA2 P54652 5.86
HSPA4L O95757 -2.45e+00
HSPA6 P17066 -5.92e+00
IARS P41252 -1.57e+00
IER3 P46695 -2.24e+00
IFIT1 P09914 -3.04e+00
IGF2BP2 Q9Y6M1 1.71
IGFBP2 P18065 4.19
IL12A P29459 -4.10e+00
IL16 Q14005 2
IL34 Q6ZMJ4 -4.19e+00
IL6R P08887 1.55
IMPA2 O14732 4.97
ING2 Q9H160 2.45
INHBA P08476 2.11
INHBB P09529 12.69
INHBE P58166 -5.11e+00
INPP5A Q14642 2.61
INPP5D Q92835 -4.35e+00
IRAK3 Q9Y616 1.94
IRS1 P35568 1.66
IRS2 Q9Y4H2 4.46
ISCA1 Q9BUE6 1.43
ITGA10 O75578 13.87
ITGA11 Q9UKX5 -3.05e+00
ITGA2 P17301 -2.06e+00
ITGA8 P53708 2.1
ITGBL1 O95965 2.1
ITPKB P27987 -1.82e+00
JADE1 Q6IE81 2.31
JADE3 Q92613 -1.88e+00
JAG1 P78504 -1.47e+00
JAK2 O60674 1.75
JUN P05412 -2.12e+00
KALRN O60229 -3.01e+00
KANK1 Q14678 2.35
KCND3 Q9UK17 2.79
KCNE3 Q9Y6H6 -2.51e+00
KCNK6 Q9Y257 7.4
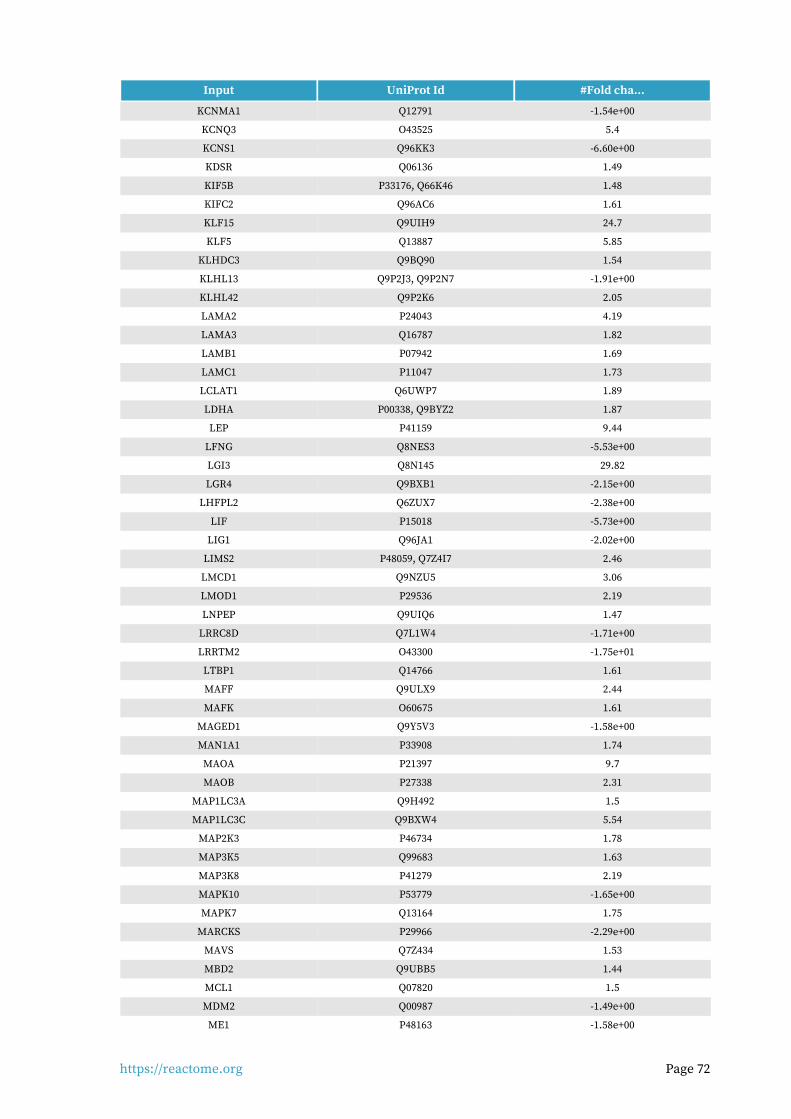
https://reactome.org Page 72
Input UniProt Id #Fold cha...
KCNMA1 Q12791 -1.54e+00
KCNQ3 O43525 5.4
KCNS1 Q96KK3 -6.60e+00
KDSR Q06136 1.49
KIF5B P33176, Q66K46 1.48
KIFC2 Q96AC6 1.61
KLF15 Q9UIH9 24.7
KLF5 Q13887 5.85
KLHDC3 Q9BQ90 1.54
KLHL13 Q9P2J3, Q9P2N7 -1.91e+00
KLHL42 Q9P2K6 2.05
LAMA2 P24043 4.19
LAMA3 Q16787 1.82
LAMB1 P07942 1.69
LAMC1 P11047 1.73
LCLAT1 Q6UWP7 1.89
LDHA P00338, Q9BYZ2 1.87
LEP P41159 9.44
LFNG Q8NES3 -5.53e+00
LGI3 Q8N145 29.82
LGR4 Q9BXB1 -2.15e+00
LHFPL2 Q6ZUX7 -2.38e+00
LIF P15018 -5.73e+00
LIG1 Q96JA1 -2.02e+00
LIMS2 P48059, Q7Z4I7 2.46
LMCD1 Q9NZU5 3.06
LMOD1 P29536 2.19
LNPEP Q9UIQ6 1.47
LRRC8D Q7L1W4 -1.71e+00
LRRTM2 O43300 -1.75e+01
LTBP1 Q14766 1.61
MAFF Q9ULX9 2.44
MAFK O60675 1.61
MAGED1 Q9Y5V3 -1.58e+00
MAN1A1 P33908 1.74
MAOA P21397 9.7
MAOB P27338 2.31
MAP1LC3A Q9H492 1.5
MAP1LC3C Q9BXW4 5.54
MAP2K3 P46734 1.78
MAP3K5 Q99683 1.63
MAP3K8 P41279 2.19
MAPK10 P53779 -1.65e+00
MAPK7 Q13164 1.75
MARCKS P29966 -2.29e+00
MAVS Q7Z434 1.53
MBD2 Q9UBB5 1.44
MCL1 Q07820 1.5
MDM2 Q00987 -1.49e+00
ME1 P48163 -1.58e+00

https://reactome.org Page 73
Input UniProt Id #Fold cha...
METTL7A Q9H8H3 4.96
MFAP2 P55001 -1.58e+00
MFAP4 P55083 -1.95e+00
MFGE8 Q08431 2.1
MGAT1 P26572 1.81
MMP15 P51511 2.36
MMP7 P09237 28.84
MORF4L2 Q15014 2
MOSPD2 Q8NHP6 1.61
MPST P25325 1.99
MRPL49 Q13405 -1.46e+00
MSRB1 Q9NZV6 1.59
MSRB3 Q8IXL7 1.66
MT1E P04731, P04732 4.44
MT1M P13640, Q8N339 5.01
MT1X P80297 9.1
MT2A P02795, P04731 4.55
MTMR10 Q9NXD2 1.62
MTOR P42345 1.58
MUC1 P15941 1.8
MX1 P20591 -1.98e+00
MYC P01106 1.8
MYL12A P19105 1.77
NADK O95544 1.72
NAMPT P43490 2.03
NCOA3 Q9Y6Q9 2.65
NEDD4L Q96PU5 -2.09e+00
NEGR1 Q7Z3B1 2.75
NF2 P35240 2.04
NFE2 Q16621 -7.48e+00
NFIL3 Q16649 2.2
NFKBIA P25963 1.74
NFYB P25208 2.02
NID1 P14543 2.91
NLN Q9BYT8 1.73
NNMT P40261 4.45
NOV Q9UIW2 -5.98e+00
NPAS2 Q99743 2.18
NPC1 O15118 1.82
NPC1L1 Q9UHC9-2 -5.94e+00
NQO1 P15559 -1.68e+00
NR0B1 P51843-1, P51843-2 -9.56e+00
NR1D2 Q14995 -1.54e+00
NR2F2 P24468 1.73
NR3C1 P04150 -2.01e+00
NR4A1 P22736 2.5
NR4A3 Q92570-1, Q92570-2 4.33
NRP2 O60462 2.07
NT5E P21589 -1.83e+00
NUAK1 O60285 2.57
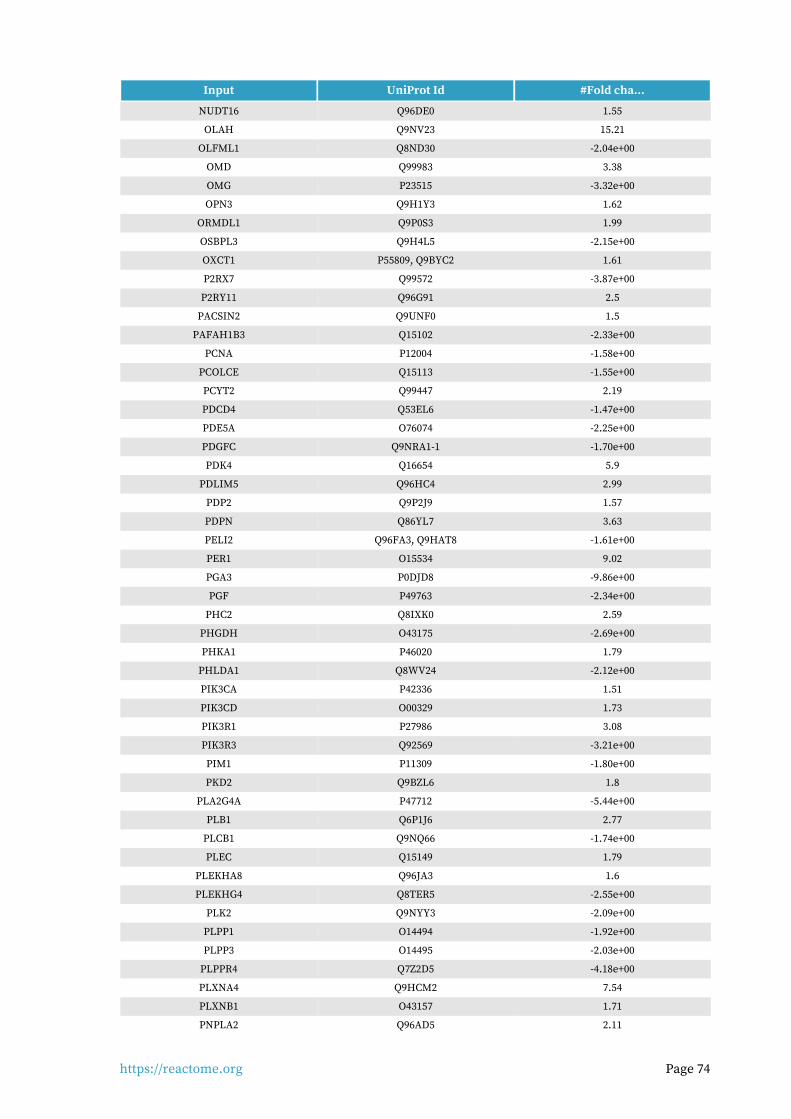
https://reactome.org Page 74
Input UniProt Id #Fold cha...
NUDT16 Q96DE0 1.55
OLAH Q9NV23 15.21
OLFML1 Q8ND30 -2.04e+00
OMD Q99983 3.38
OMG P23515 -3.32e+00
OPN3 Q9H1Y3 1.62
ORMDL1 Q9P0S3 1.99
OSBPL3 Q9H4L5 -2.15e+00
OXCT1 P55809, Q9BYC2 1.61
P2RX7 Q99572 -3.87e+00
P2RY11 Q96G91 2.5
PACSIN2 Q9UNF0 1.5
PAFAH1B3 Q15102 -2.33e+00
PCNA P12004 -1.58e+00
PCOLCE Q15113 -1.55e+00
PCYT2 Q99447 2.19
PDCD4 Q53EL6 -1.47e+00
PDE5A O76074 -2.25e+00
PDGFC Q9NRA1-1 -1.70e+00
PDK4 Q16654 5.9
PDLIM5 Q96HC4 2.99
PDP2 Q9P2J9 1.57
PDPN Q86YL7 3.63
PELI2 Q96FA3, Q9HAT8 -1.61e+00
PER1 O15534 9.02
PGA3 P0DJD8 -9.86e+00
PGF P49763 -2.34e+00
PHC2 Q8IXK0 2.59
PHGDH O43175 -2.69e+00
PHKA1 P46020 1.79
PHLDA1 Q8WV24 -2.12e+00
PIK3CA P42336 1.51
PIK3CD O00329 1.73
PIK3R1 P27986 3.08
PIK3R3 Q92569 -3.21e+00
PIM1 P11309 -1.80e+00
PKD2 Q9BZL6 1.8
PLA2G4A P47712 -5.44e+00
PLB1 Q6P1J6 2.77
PLCB1 Q9NQ66 -1.74e+00
PLEC Q15149 1.79
PLEKHA8 Q96JA3 1.6
PLEKHG4 Q8TER5 -2.55e+00
PLK2 Q9NYY3 -2.09e+00
PLPP1 O14494 -1.92e+00
PLPP3 O14495 -2.03e+00
PLPPR4 Q7Z2D5 -4.18e+00
PLXNA4 Q9HCM2 7.54
PLXNB1 O43157 1.71
PNPLA2 Q96AD5 2.11

https://reactome.org Page 75
Input UniProt Id #Fold cha...
POLD4 Q9HCU8 1.77
POU2F2 P09086 -3.97e+00
PPARG P37231 2.05
PPIP5K1 Q6PFW1 -1.81e+00
PPM1B O75688 1.72
PPP1CB P62140 2.08
PPP1R14A Q96A00 7.52
PPP1R1B Q9UD71 -8.56e+00
PRDM1 O75626 2.39
PREB Q9HCU5 2.17
PRKAA1 Q13131 1.82
PRKACB P22694 -1.51e+00
PRKAG2 P54619, Q9UGJ0 2.97
PRKCE Q02156 -2.29e+00
PRKD1 Q15139 1.62
PRKG2 Q13237 -3.17e+00
PRODH O43272 32.79
PSAT1 Q9Y617 -2.43e+00
PTGES O14684 -1.86e+00
PTK2B Q14289 2.17
PTX3 P26022 4.61
PYCR1 P32322, Q96C36 -1.65e+00
PYGB P11216 1.49
QPCT Q16769 2.22
QPRT Q15274 -2.31e+00
RAB31 Q13636 1.94
RAB43 Q86YS6 2.87
RAB7B Q96AH8 -3.54e+00
RAC3 Q9Y6Q9 2.86
RAD1 O60671 1.8
RAET1G Q6H3X3 -2.22e+00
RAF1 P04049 1.72
RALA P11233 -1.51e+00
RAMP3 O60896 -1.87e+01
RANBP9 Q96S59 1.66
RAP2B P61225 -2.45e+00
RASGRP2 Q7LDG7 3.58
RBM12 Q99829 1.55
RBPJ Q06330 -1.60e+00
REV3L O60673 2.17
RGCC Q9H4X1 9.02
RGL1 Q9NZL6 -2.12e+00
RGMB Q6NW40 -1.99e+00
RHEB Q15382 1.93
RHNO1 Q9BSD3 -1.52e+00
RHOB P62745 2.28
RHOBTB3 O94955 2.06
RHOJ Q9H4E5 -2.28e+00
RIOK3 O14730 1.45
RNF144B Q7Z419 4.45

https://reactome.org Page 76
Input UniProt Id #Fold cha...
RNF217 Q8TC41 2.1
RNF41 Q9H4P4 1.42
ROBO1 Q9Y6N7 -2.21e+00
ROCK1 Q13464 1.48
ROCK2 O75116 1.73
RPS6KA2 Q15349 1.89
RPS6KA5 O75582 -5.16e+00
RRP9 O43818 1.88
RSPO1 Q2MKA7 2.57
RSPO2 Q6UXX9 -2.79e+00
RXFP1 Q9HBX9 -3.98e+00
S1PR1 P21453 -2.85e+00
SAMD8 Q96LT4 1.5
SAMHD1 Q9Y3Z3 14.26
SAP30 O75446 2.35
SAT1 Q9H2H9 2.49
SCD O00767 -3.97e+00
SCN1B Q07699 -2.44e+00
SCN7A Q01118 2.69
SDC1 P18827 -1.80e+00
SEH1L Q96EE3-2 1.63
SEMA3A Q14563 -2.36e+00
SEMA6D Q8NFY4 -2.53e+00
SERPINH1 P50454 -1.61e+00
SESN2 P58004 -1.92e+00
SH3D19 Q5HYK7 1.65
SH3GL3 Q99963 -5.36e+00
SHB Q15464 -1.94e+00
SHMT2 P34897 -1.65e+00
SKP2 Q13309 1.93
SLC14A1 Q13336 -1.32e+01
SLC19A2 O60779, Q9BZV2 2.1
SLC25A32 Q9H2D1 1.82
SLC26A6 Q9BXS9 1.88
SLC2A12 Q8TD20 -1.89e+00
SLC35C1 Q96A29 1.52
SLC35D1 Q9NTN3 1.82
SLC36A1 Q7Z2H8 1.66
SLC38A1 Q9H2H9 1.76
SLC38A2 Q96QD8 2.35
SLC38A5 Q8WUX1 -1.97e+00
SLC39A10 Q9ULF5 -2.35e+00
SLC41A2 Q96JW4 -1.92e+00
SLC4A4 Q9Y6R1 -1.71e+00
SLC4A7 Q9Y6M7 -1.90e+00
SLC5A6 Q9Y289 2.26
SLC6A6 P31641 -4.26e+00
SLC6A9 P48067 -4.54e+00
SLC7A5 Q01650 -2.24e+00
SLC7A6 Q92536 3.09
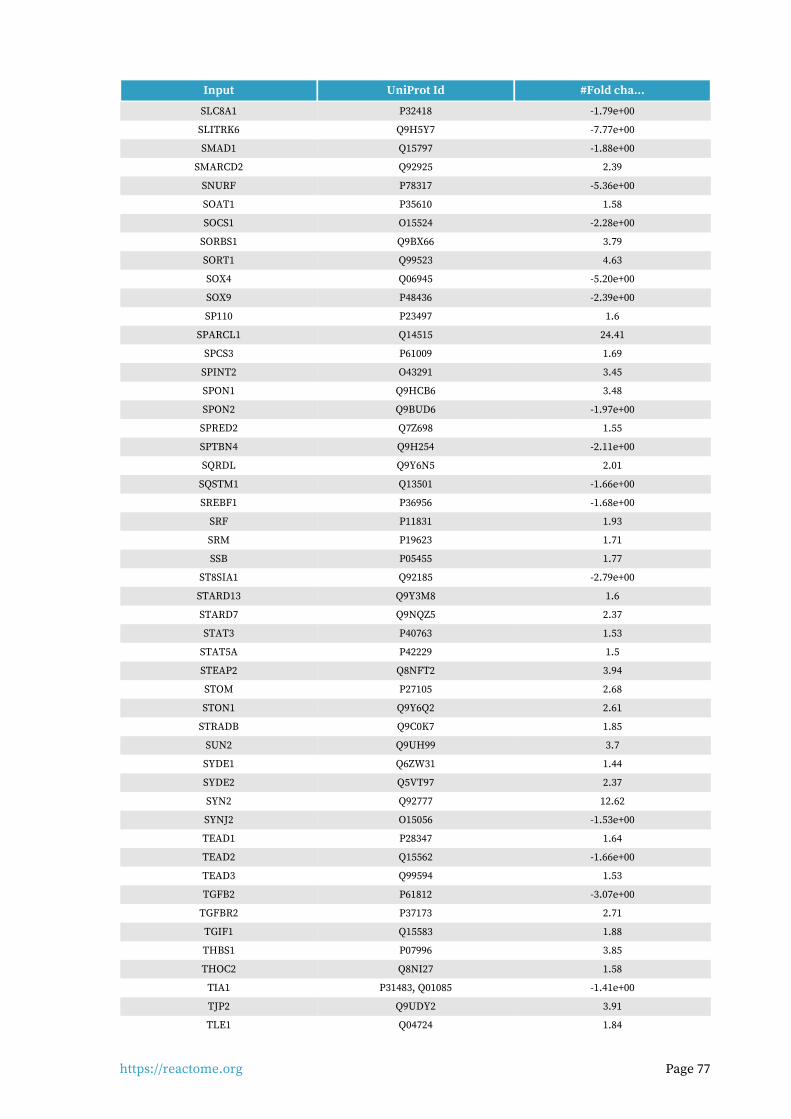
https://reactome.org Page 77
Input UniProt Id #Fold cha...
SLC8A1 P32418 -1.79e+00
SLITRK6 Q9H5Y7 -7.77e+00
SMAD1 Q15797 -1.88e+00
SMARCD2 Q92925 2.39
SNURF P78317 -5.36e+00
SOAT1 P35610 1.58
SOCS1 O15524 -2.28e+00
SORBS1 Q9BX66 3.79
SORT1 Q99523 4.63
SOX4 Q06945 -5.20e+00
SOX9 P48436 -2.39e+00
SP110 P23497 1.6
SPARCL1 Q14515 24.41
SPCS3 P61009 1.69
SPINT2 O43291 3.45
SPON1 Q9HCB6 3.48
SPON2 Q9BUD6 -1.97e+00
SPRED2 Q7Z698 1.55
SPTBN4 Q9H254 -2.11e+00
SQRDL Q9Y6N5 2.01
SQSTM1 Q13501 -1.66e+00
SREBF1 P36956 -1.68e+00
SRF P11831 1.93
SRM P19623 1.71
SSB P05455 1.77
ST8SIA1 Q92185 -2.79e+00
STARD13 Q9Y3M8 1.6
STARD7 Q9NQZ5 2.37
STAT3 P40763 1.53
STAT5A P42229 1.5
STEAP2 Q8NFT2 3.94
STOM P27105 2.68
STON1 Q9Y6Q2 2.61
STRADB Q9C0K7 1.85
SUN2 Q9UH99 3.7
SYDE1 Q6ZW31 1.44
SYDE2 Q5VT97 2.37
SYN2 Q92777 12.62
SYNJ2 O15056 -1.53e+00
TEAD1 P28347 1.64
TEAD2 Q15562 -1.66e+00
TEAD3 Q99594 1.53
TGFB2 P61812 -3.07e+00
TGFBR2 P37173 2.71
TGIF1 Q15583 1.88
THBS1 P07996 3.85
THOC2 Q8NI27 1.58
TIA1 P31483, Q01085 -1.41e+00
TJP2 Q9UDY2 3.91
TLE1 Q04724 1.84
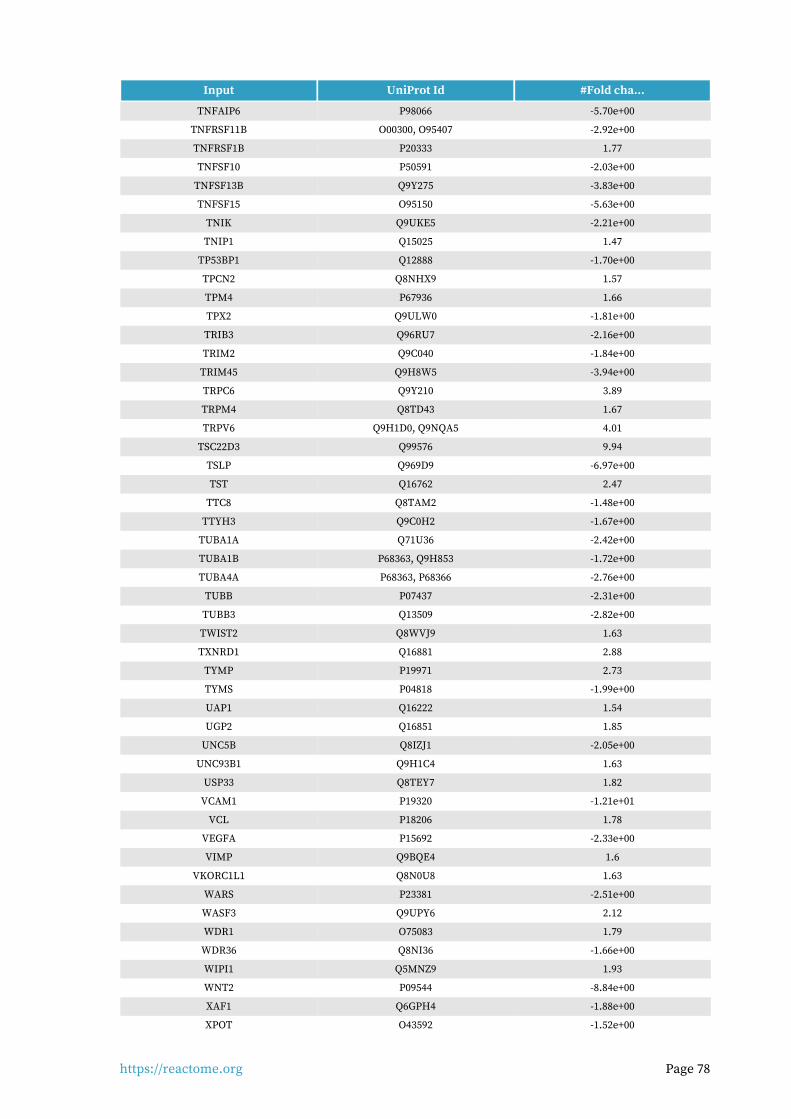
https://reactome.org Page 78
Input UniProt Id #Fold cha...
TNFAIP6 P98066 -5.70e+00
TNFRSF11B O00300, O95407 -2.92e+00
TNFRSF1B P20333 1.77
TNFSF10 P50591 -2.03e+00
TNFSF13B Q9Y275 -3.83e+00
TNFSF15 O95150 -5.63e+00
TNIK Q9UKE5 -2.21e+00
TNIP1 Q15025 1.47
TP53BP1 Q12888 -1.70e+00
TPCN2 Q8NHX9 1.57
TPM4 P67936 1.66
TPX2 Q9ULW0 -1.81e+00
TRIB3 Q96RU7 -2.16e+00
TRIM2 Q9C040 -1.84e+00
TRIM45 Q9H8W5 -3.94e+00
TRPC6 Q9Y210 3.89
TRPM4 Q8TD43 1.67
TRPV6 Q9H1D0, Q9NQA5 4.01
TSC22D3 Q99576 9.94
TSLP Q969D9 -6.97e+00
TST Q16762 2.47
TTC8 Q8TAM2 -1.48e+00
TTYH3 Q9C0H2 -1.67e+00
TUBA1A Q71U36 -2.42e+00
TUBA1B P68363, Q9H853 -1.72e+00
TUBA4A P68363, P68366 -2.76e+00
TUBB P07437 -2.31e+00
TUBB3 Q13509 -2.82e+00
TWIST2 Q8WVJ9 1.63
TXNRD1 Q16881 2.88
TYMP P19971 2.73
TYMS P04818 -1.99e+00
UAP1 Q16222 1.54
UGP2 Q16851 1.85
UNC5B Q8IZJ1 -2.05e+00
UNC93B1 Q9H1C4 1.63
USP33 Q8TEY7 1.82
VCAM1 P19320 -1.21e+01
VCL P18206 1.78
VEGFA P15692 -2.33e+00
VIMP Q9BQE4 1.6
VKORC1L1 Q8N0U8 1.63
WARS P23381 -2.51e+00
WASF3 Q9UPY6 2.12
WDR1 O75083 1.79
WDR36 Q8NI36 -1.66e+00
WIPI1 Q5MNZ9 1.93
WNT2 P09544 -8.84e+00
XAF1 Q6GPH4 -1.88e+00
XPOT O43592 -1.52e+00
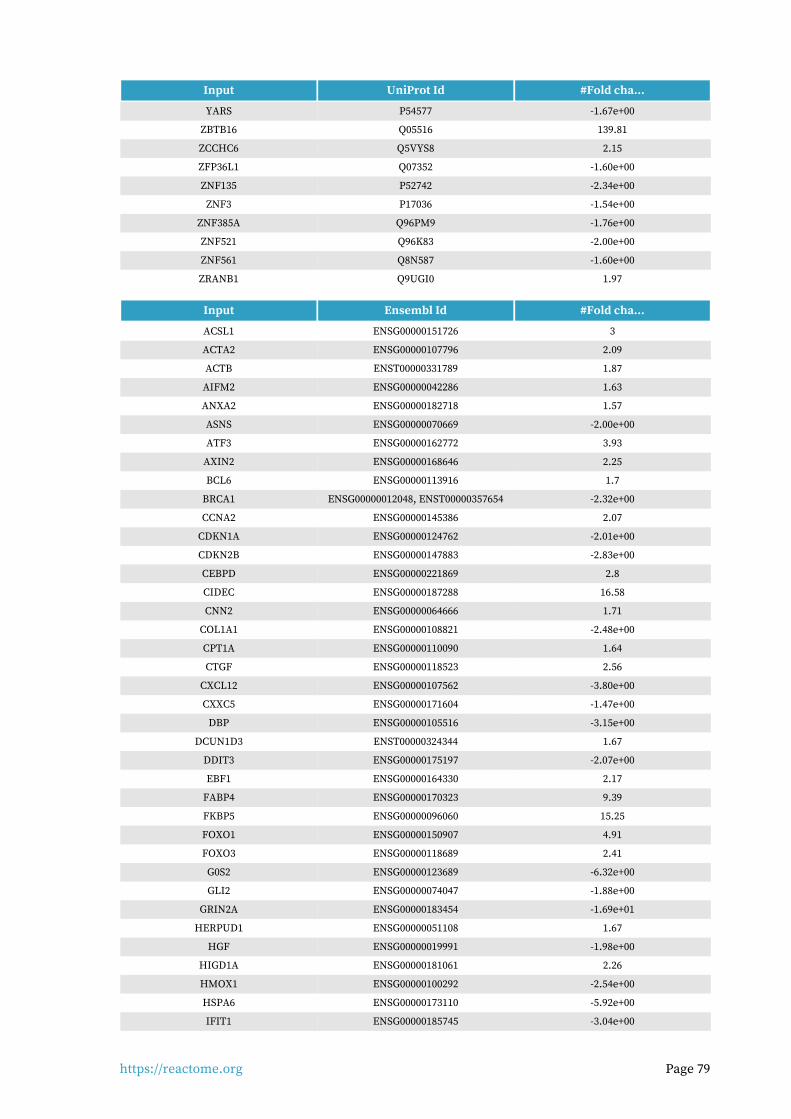
https://reactome.org Page 79
Input UniProt Id #Fold cha...
YARS P54577 -1.67e+00
ZBTB16 Q05516 139.81
ZCCHC6 Q5VYS8 2.15
ZFP36L1 Q07352 -1.60e+00
ZNF135 P52742 -2.34e+00
ZNF3 P17036 -1.54e+00
ZNF385A Q96PM9 -1.76e+00
ZNF521 Q96K83 -2.00e+00
ZNF561 Q8N587 -1.60e+00
ZRANB1 Q9UGI0 1.97
Input Ensembl Id #Fold cha...
ACSL1 ENSG00000151726 3
ACTA2 ENSG00000107796 2.09
ACTB ENST00000331789 1.87
AIFM2 ENSG00000042286 1.63
ANXA2 ENSG00000182718 1.57
ASNS ENSG00000070669 -2.00e+00
ATF3 ENSG00000162772 3.93
AXIN2 ENSG00000168646 2.25
BCL6 ENSG00000113916 1.7
BRCA1 ENSG00000012048, ENST00000357654 -2.32e+00
CCNA2 ENSG00000145386 2.07
CDKN1A ENSG00000124762 -2.01e+00
CDKN2B ENSG00000147883 -2.83e+00
CEBPD ENSG00000221869 2.8
CIDEC ENSG00000187288 16.58
CNN2 ENSG00000064666 1.71
COL1A1 ENSG00000108821 -2.48e+00
CPT1A ENSG00000110090 1.64
CTGF ENSG00000118523 2.56
CXCL12 ENSG00000107562 -3.80e+00
CXXC5 ENSG00000171604 -1.47e+00
DBP ENSG00000105516 -3.15e+00
DCUN1D3 ENST00000324344 1.67
DDIT3 ENSG00000175197 -2.07e+00
EBF1 ENSG00000164330 2.17
FABP4 ENSG00000170323 9.39
FKBP5 ENSG00000096060 15.25
FOXO1 ENSG00000150907 4.91
FOXO3 ENSG00000118689 2.41
G0S2 ENSG00000123689 -6.32e+00
GLI2 ENSG00000074047 -1.88e+00
GRIN2A ENSG00000183454 -1.69e+01
HERPUD1 ENSG00000051108 1.67
HGF ENSG00000019991 -1.98e+00
HIGD1A ENSG00000181061 2.26
HMOX1 ENSG00000100292 -2.54e+00
HSPA6 ENSG00000173110 -5.92e+00
IFIT1 ENSG00000185745 -3.04e+00

https://reactome.org Page 80
Input Ensembl Id #Fold cha...
IL12A ENSG00000168811 -4.10e+00
IL6R ENSG00000160712 1.55
ITGBL1 ENSG00000198542 2.1
JAG1 ENSG00000101384 -1.47e+00
KLF15 ENSG00000163884 24.7
KLF5 ENSG00000102554 5.85
KLHDC3 ENSG00000124702 1.54
LEP ENSG00000174697 9.44
LIF ENSG00000128342 -5.73e+00
MAOA ENSG00000189221 9.7
MCL1 ENSG00000143384 1.5
MDM2 ENSG00000135679 -1.49e+00
ME1 ENSG00000065833 -1.58e+00
MT2A ENSG00000125148 4.55
MUC1 ENSG00000185499 1.8
MX1 ENSG00000157601 -1.98e+00
MYC ENSG00000136997 1.8
NAMPT ENSG00000105835 2.03
NFE2 ENSG00000123405 -7.48e+00
NPAS2 ENSG00000170485 2.18
NR4A3 ENSG00000119508 4.33
PCNA ENSG00000132646 -1.58e+00
PDCD4 ENSG00000150593 -1.47e+00
PER1 ENSG00000179094, ENST00000317276 9.02
PIK3R1 ENSG00000145675 3.08
PIM1 ENSG00000137193 -1.80e+00
PLK2 ENSG00000145632 -2.09e+00
PPARG ENSG00000132170 2.05
PRDM1 ENSG00000057657 2.39
PREB ENSG00000138073 2.17
RALA ENSG00000006451 -1.51e+00
RGCC ENSG00000102760 9.02
RGL1 ENSG00000143344 -2.12e+00
ROBO1 ENSG00000169855 -2.21e+00
S1PR1 ENSG00000170989 -2.85e+00
SAMHD1 ENSG00000101347 14.26
SCD ENSG00000099194 -3.97e+00
SERPINH1 ENSG00000149257 -1.61e+00
SESN2 ENSG00000130766 -1.92e+00
SOCS1 ENSG00000185338 -2.28e+00
SREBF1 ENSG00000072310 -1.68e+00
STAT3 ENSG00000168610 1.53
THBS1 ENSG00000137801 3.85
TNFRSF1B ENSG00000028137 1.77
TNIK ENSG00000154310 -2.21e+00
TRIB3 ENSG00000101255 -2.16e+00
TRIM2 ENSG00000109654 -1.84e+00
TRIM45 ENSG00000134253 -3.94e+00
TSC22D1 ENSG00000102804 2.28
TXNRD1 ENSG00000198431 2.88

https://reactome.org Page 81
Input Ensembl Id #Fold cha...
TYMS ENSG00000176890 -1.99e+00
USP46 ENSG00000109189 1.68
VCAM1 ENSG00000162692 -1.21e+01
VEGFA ENSG00000112715 -2.33e+00
WIPI1 ENSG00000070540 1.93
XAF1 ENSG00000132530 -1.88e+00
ZNF385A ENSG00000161642 -1.76e+00

https://reactome.org Page 82
7. Identifiers not found
These 456 identifiers were not found neither mapped to any entity in Reactome.
10-Mar 4-Sep AAED1 AC007879.7 AC008522.1 AC159540.1 ACKR2 ADAP1
ADGRA3 ADGRB2 AFAP1 AFAP1L1 AFAP1L2 AIF1L AKAP12 AKAP2
ALPK3 ANGPTL1 ANGPTL2 ANGPTL7 ANKMY2 ANKRD13A ANXA4 AP5B1
APBB2 APCDD1 ARID5A ARL4C ARMCX6 ARRDC2 ARRDC4 ATL3
B3GALT2 BAIAP2-AS1 BATF3 BMPER BNC2 BRIX1 C10orf10 C14orf132
C14orf159 C16orf52 C1QTNF1 C20orf96 C2CD2 C4orf46 CACUL1 CADPS2
CALCOCO2 CAMK1D CAMK2N1 CASC15 CAV2 CBX1 CCDC107 CCDC34
CCDC54 CCDC69 CCDC71L CCL8 CD302 CD82 CDK2AP2 CDV3
CELF2CH17-
472G23.4CHRDL2 CLIC4 CLIC6 CORO2B CORO6 CPA4
CPED1 CPZ CREB5 CRIM1 CRIP2 CRYBG3 CTB-92J24.3 CTC-441N14.2
CTTNBP2 DAAM2 DANCR DBNDD2 DCHS1 DGCR6 DNAJB4 DNASE1
DNM1P47 DSTN DTNA DTWD1 DUSP23 ECHDC1 EFR3B EGR3
EIF4EBP2 EMP1 EMP2 EMP3 ENC1 ENOSF1 EOGT ERGIC1
ERRFI1 ETV5 EVA1C EVI2A FADS3 FAIM2 FAM101B FAM102A
FAM105A FAM107A FAM107B FAM129B FAM171A2 FAM171B FAM180A FAM196A
FAM198B FAM43A FAM46A FAM46B FAM46C FAM49A FAM63B FER1L6
FHOD1 FIBIN FKRP FNDC3B FOXF2 FOXN2 FRMD5 FRZB
GADD45B GALM GDF15 GDF7 GK5 GOLGA2P7 GPM6B GPR153
GPRASP1 GPRASP2 GPRC5B GRAMD3 GRAMD4 GRIK1-AS1 GXYLT2 HAPLN3
HEG1 HIPK3 HSPB3 IFI44L IGDCC4 IGFLR1 IGSF10 IVNS1ABP
KANK4 KAZALD1 KCTD12 KCTD9 KIAA0408 KIAA0895L KIAA0922 KIAA1755
KLF13 KLF6 KLF9 KLHL23 LARP4 LARP4B LBH LDB2
LHFP LINC00702 LINC00890 LINC00906 LINC00968 LMO3 LRCH2 LRRC15
LRRC17 LRRC25 LRRFIP2 LRRN1 LSP1 MAGED4 MAMDC2 MAP1A
MAP4K3 MAPRE2 MAPRE3 MARCKSL1 MAST4 MBNL1 MCTP1 MEIS2
MEIS3 MEST METRNL MEX3A MEX3B MFSD6 MIAT MICAL2
MIR22HG MIR99AHG MMD MOB3B MPLKIP MPP3 MSANTD3 MTSS1
MTSS1L MXRA5 MYADM MYH13 MYO1E MYOZ2 N4BP3 NAV3
NBL1 NBPF1 NCAM2 NCKAP5 NEDD9 NEK10 NEK11 NEXN
NKD1 NKD2 NPR3 NSMF NT5DC1 NXPH3 NYNRIN OLFML2A
OSR2 OTOGL PALLD PAPLN PAWR PCDH18 PDLIM1 PDZD2
PEAK1 PEG10 PER3 PGBD5 PGPEP1 PHOSPHO2 PID1 PIEZO2
PIFO PKD1L2 PKDCC PKIA PKNOX2 PLCE1-AS1 PLCXD3 PLXDC1
PLXDC2 POM121L9P PPP1R12C PPTC7 PREPL PRR5L PRRX1 PRSS35
PRUNE2 PTGFRN PTPDC1 PTPRG PXDC1 PXYLP1 RAB11FIP1 RAB11FIP4
RAPGEF5 RASD1 RASD2 RASL10B RASL11A RASL11B RASL12 RASSF2
RASSF8 RBM43 RBPMS RCAN1 RCAN3 RELL1 RFTN2 RGMB-AS1
RND2 RND3 RNPEPL1RP11-
1035H13.3RP11-
124N14.3RP11-
1415C14.4RP11-175K6.1 RP11-17A4.2
RP11-184M15.1
RP11-20J15.3 RP11-253M7.1RP11-
357D18.1RP11-366L20.2 RP11-434D9.1
RP11-448G15.3
RP11-493E12.3
RP11-496D24.2
RP11-548K23.11
RP11-54O7.3 RP11-703I16.1 RP11-770J1.5RP11-
824M15.3RP11-
890B15.2RP11-
903H12.5
RP4-621F18.2 RP4-756G23.5 RP5-864K19.6RPL36A-
HNRNPH2RPLP0P2 RRAD RRAS2 RTKN2
RWDD4 SACS SALL2 SAMD12 SAMD4A SAMD5 SCARA3 SEC1P

https://reactome.org Page 83
SECTM1 SELENBP1 SEMA3B SENP7 SERPINA3_1 SFT2D2 SGCD SGTB
SH3BP2 SH3BP5 SH3RF3 SIDT2 SIPA1L2 SLC16A12 SLC16A4 SLC22A23
SLC35F5 SLC35G2 SLC39A13 SLC45A1 SLC7A14 SMIM3 SMTN SMTNL2
SNX21 SOBP SPATS2 SPEN SPOCK1 SRPX SSBP2 SSBP3
SSH2 STC1 STEAP1 STEAP4 STIM2 STK17B STK38L STUM
STX11 STX12 STX2 STXBP4 SUSD6 SWAP70 SYNPO2 TACC1
TAGLN TANC2 TBX18 TCEAL1 TCEAL3 TCEAL4 TCEAL9 TENM4
TEX2 TIMP4 TLCD2 TLDC2 TM2D1 TM4SF1 TM7SF3 TMCC2
TMEM119 TMEM150A TMEM19 TMEM2 TMEM200A TMEM217 TMEM26 TMEM35A
TMEM47 TMEM50A TMEM87B TNFAIP2 TRAK2 TRERF1 TRNP1 TRO
TSKU TSPAN11 TSPAN8 TTC28 TTC39C UBALD2 UBASH3B UHMK1
ULK2 USP53 VASH2 VGLL3 VPS13D VSTM4 VWCE WBP1L
WDR63 WFDC1XXbac-
BPG246D15.9YIF1B YPEL4 ZBTB22 ZBTB47 ZC3H12C
ZCCHC5 ZDHHC12 ZDHHC23 ZDHHC9 ZFAND5 ZFP36L2 ZHX2 ZHX3
ZMYM3 ZNF395 ZNF438 ZNF488 ZNF618 ZNF821 ZSCAN31 ZYX