part 3 from individual behaviour to social systems 鄭先祐 (ayo) 國立台南大學...
TRANSCRIPT

Part 3 From individual behaviour to social systems
鄭先祐 (Ayo)
國立台南大學 環境與生態學院

evolution of animal signals 2
Part 3 Social systems
Chap. 7 The evolution of animal signals Chap. 8 Sexual selection and mate choice Chap. 9 Sociality and kin selection in insects Chap.10 Predicting family dynamics in social
vertebrates Chap. 11 The ecology of relationships Chap. 12 The social gene

The Evolution of Animal signals (動物訊號的演
化 )
鄭先祐 (Ayo)
國立台南大學 環境與生態學院

evolution of animal signals 4
The evolution of animal signals (動物訊號的演化 )
7.1 Introduction 7.2 The signaller’s ( 發出信息者 ) perspective
7.2.1 Getting the message across 7.2.2 The influence of the physical environment 7.2.3 The influence of the audience 7.2.4 The influence of other signallers
7.3 The receiver’s ( 接收者 ) perspective 7.4 conclusions and prospects

evolution of animal signals 5
7.2 The signaller’s perspective
It serves to influence the receiver’s behaviour in a way that benefits the signaller. 但有些情況,亦可能是對接收者有利。
Signaller, 發出訊息,是有風險和能量的成本。 演化會傾向 Greater effectiveness and lower fit
ness cost. ( 增加效力,且下降成本 )

evolution of animal signals 6
7.2.1 getting the message across 訊息要能傳達,特別於長距離,吵雜的環境。 譬如: male anoles’ head-bobbing display. 1. the display begins with a series of head mo
vements that are of high acceleration, velocity and amplitude compared with the rest of the display. This movements are conspicuous. (Fig. 7.1)
2. the anoles’ assertion display is repetitive. 確實辨識。
3. the anoles’ signal is stereotyped.

evolution of animal signals 7
Fig. 7.1 A schematic diagram of how the assertion display of A. auratus is created, together with two typical examples of the display.

evolution of animal signals 8
Fig. 7.1 the assertion display of A. auratus.

evolution of animal signals 9
7.2.2 The influence of the physical environment
Marchetti (1993), the plumage patterns of eight species of warblers of the genus Phylloscopus, which breed in the forests of Kashmir, India.
All of the species are small and greenish in color, but they possess varying numbers of pale color patches on their wings, crown, rump and tail.

evolution of animal signals 10
物理環境影響訊息的展現 Experimental manipulation of male coloration
in P. inornatus directly affected territory size, with males that were tendered more conspicuous obtaining larger territories than control individuals.
Species with more patches tended to occupy dark, dense habitats, while those with fewer patches tended to breed in open area.

evolution of animal signals 11
Wiley (1991) male territorial songs of eastern North American oscine birds
This study considered 120 species of oscines. Three of properties concerned the temporal s
tructure of the song, and were indicative of rapid modulation: the minimal period of repeated elements, the presence or absence of one or more buzzes and the presence or absence of ‘side-bands’ on a song spectrogram.
棲地區分為六種,三種是樹林型,另三種是 open habitats.

evolution of animal signals 12
Short repetition periods, buzzes and side-bands less common among birds of forested habitats.
這三種特質,於樹林內都較容易被干擾。

evolution of animal signals 13
7.2.3 The influence of the audience
Endler (1992), the coloration of male guppies provide evidence for the matching of display design to receiver sensory capacities.
Wild populations of puppies have complex and polymorphic colour patterns, comprising patches of eight major types: red-orange, yellow, bronze-green, cream-white, blue, silver, brown, black and body colour.
Those colours, restricted to adult males, play a role in mate choice.

evolution of animal signals 14
Female preferences vary among populations, but typically favour males with more carotenoid (red-orange and yellow) patches, more structural colour (blue, green and silver) patches, and patterns that contrast in colour or patch size with the visual background.
Conspicuous and highly colourful fish suffer greater risk of predation.
Selection had favored those colour patterns that were most conspicuous to guppies while being least conspicuous to predator.

evolution of animal signals 15
Proctor (1991) water mite
運用 female’s pre-existing sensory properties. ( 原本就存在的感應能力 ), which evolved for reasons unrelated to communication.
Mite 是 ambush predators. 於水上感應獵物活動時的水波。
雄 mite 運用其腳製造水的波動,讓雌 mite 誤以為是獵物。 愈飢餓的雌 mite , 愈會受到吸引。

evolution of animal signals 16
7.2.4 The influence of other signallers
於許多訊息傳送的過程,訊息的傳達需要能獨特出色。
Loftus-Hills & Littlejohn (1992), two species of narrow-mouthed toad, widely distributed in the southern US.
Males rely on an advertisement call to attract females.
sympatric or allopatric 分布情況,可有四種。 Sympatric 傾向可以區分開。

evolution of animal signals 17
Greenfield and Roizen (1993) chorusing in the neotropical katydid (美洲大螽斯 )
An isolated male will usually maintain a regular chirp rhythm for several minutes.
倘若兩隻 male 相距 10 公尺內,會有 synchronization 的現象。
這種 synchronization 是透過 an inhibitory resetting 的機制。
聽到聲音,會抑制發聲,而在下一個預期出聲時,再發聲。搶先發聲。
Females show a preference for the leading call in a closely synchronized sequence.

evolution of animal signals 18
7.3 The receiver’s perspective
7.3.1 cooperation and conflict ( 合作與衝突 ) in signal evolution
7.3.2 physical constraints on deceit ( 欺騙 ) 7.3.3 strategic constraints on deceit 7.3.4 The possibility of rare deception 7.3.5 strategic explanations for signal diversit
y

evolution of animal signals 19
cooperation and conflict in signal evolution
One of the best-known instances of cooperative signalling is the use of a dance ‘language’ by honey bees.
Many kinds of communication involve a conflict of interest between signaller and receiver. Males are typically under stronger selection to ac
quire many mates. Females would benefit from an accurate assessm
ent of male mate quality.

evolution of animal signals 20
Threat displays
遇到 predator 出現,發出警告的訊息。 倘若有 a deceitful’ mutant 出現,它將會獲利。
族群會朝向欺騙的方向演化。 什麼機制可以維持 ‘ honesty’ ?
7.3.2 physical constraints on deceit 7.3.3 strategic constraints on deceit

evolution of animal signals 21
7.3.2 physical constraints on deceit
維持雄性優越訊息的 honesty ,有些可以用 physical constraint 。
譬如:動物通常無法自體合成 carotenoid colour ,需要從食物獲取。
雄性展現 carotenoid colour ,必要有充分食物作為基礎。

evolution of animal signals 22
7.3.3 Strategic constraints on deceit (欺騙 )
有許多訊息無法靠 physical constraint ,需要有 strategic constraint 維護 honesty
Handicap principle (Zahavi, 1977) , sexual display 信息的產生需要很高的 cost 。 倘若身體不夠好,勉強的付出高 cost 的 sexual di
splay ,會下降其生存的維護。 但是相關這方面,欠缺驗證。 最好的例子是 tail ornaments of male barn swallo
ws (Moller, 1994)

evolution of animal signals 23
7.3.4 The possibility of rare deception (欺騙 ) Redondo (1993), begging by magpie and great
spotted cuckoo chicks. The chicks that begged most intensely were found
to be those that had received smaller quantities of food from parents, indicating that the begging display provides reliable information about hunger.
The above reliable system of parent-offspring communication, however, is exploited by the chicks of great spotted cuckoos, obligate brood parasites that are raised alongside host chicks, and severely depress the breeding success of magpies.

evolution of animal signals 24
7.3.5 Strategic explanations of signal diversity
動物通常有可靠的資訊顯現出發出訊息者的狀態,而且有些可有避免欺騙的機制 (Handicap principle) 。 但是鮮少研究驗證是否有 reliability 的天擇?
Zahavi (1987) 認為產出訊息的代價,可以維持honesty ,假若是對依靠訊息產出者的本身狀態而產出的信息。 但對發出逃走的訊息,則要靠掠食者掠食的壓力。 這方面的驗證,仍未有研究。

evolution of animal signals 25
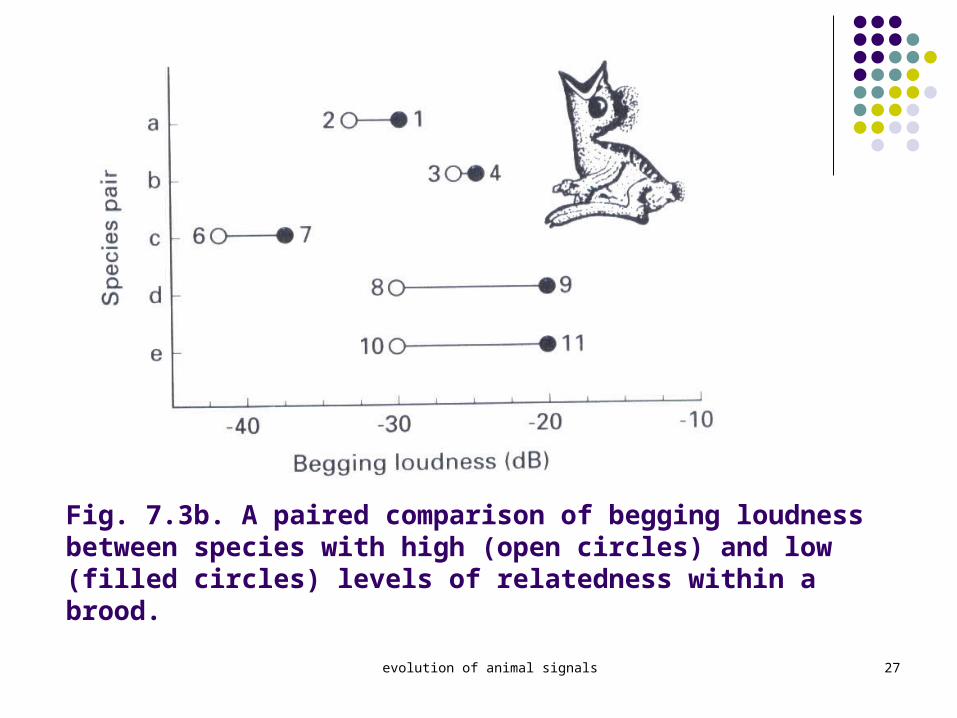
Briskie et al. (1994) 11 spp. Of passerine birds
這些鳥有不同程度的混合育幼的現象。 倘若是親緣關係相差較大,幼鳥的 beg 叫聲
和動作會較誇大。 如: cuckoo chick begs more loudly than a host c
hick because it has nothing to lose by depriving its nestmates of food)
Revealed that the loudness of nestling begging calls among the sample species was positively correlated with the percentage of extrapair young, illustrated in Fig. 7.3.

evolution of animal signals 26
Fig. 7.3 a phylogeny of the bird species (Briskie et al., 1994)
evolution of animal signals 27
Fig. 7.3b. A paired comparison of begging loudness between species with high (open circles) and low (filled circles) levels of relatedness within a brood.

evolution of animal signals 28
7.4 conclusions and prospects
兩種不同天擇的力量:有效率 vs. 可靠性 站在訊息產出者的立場:訊息的產出,需要有效率,低成本,高獲利。
站在接受者的立場:維持訊息的真實性 (honesty) ,訊息的產出需要提高成本。

evolution of animal signals 29
7.4.1 unanswered questions
Multiple signals Why are animal displays so complex? Models of signalling have typically assumed that o
nly a single form of display is abailable to signallers.
It is only recently that they have begun to address the possibility of multicomponent displays (Johnstone, 1996)

evolution of animal signals 30
Multiple signallers
The influence of competition among signallers on information transfer during communication. Receivers can acquire reliable information about
signallers by focusing on costly displays that only superior or more needy individuals can afford to use. (7.3)
However, where several signallers vie for attention, competition has the potential to interfere with receiver assessment of signaller condition.

evolution of animal signals 31
http://mail.nutn.edu.tw/~hycheng
問題與討論