paraventricular area: critical focus of a longitudinal neurocircuitry mediating food intake
TRANSCRIPT

Physiology & Behavior, Vol. 18, pp. 1111-1119. Pergamon Press and Brain Research Publ., 197"/. Printed in the U.S.A.
Paraventricular Area: Critical Focus of a Longitudinal Neurocircuitry
Mediating Food Intake
R I C H A R D M. GOLD, A L A N P. JONES AND P A U L E. SAWCHENKO
Psychology Department, University o f Massachusetts, Amherst, MA 01003
AND
G R E G O R Y K A P A T O S
Department o f Life Sciences, University o f Pittsburgh, Pittsburgh, PA 15260
(Received 17 December 1976)
GOLD, R. M., A. P. JONES, P. E. SAWCHENKO AND G. KAPATOS. Paraventricular area: critical focus of a longitudinal neurocircuitry mediating food intake. PHYSIOL. BEHAV. 18(6) 1111-1119, 1977. - Hyperphagia, obesity, and excessive linear growth, but not hyperdipsia, were produced by the asymmetrical combination of a parasagittal hypothalamic knife cut and a contralateral coronal knife cut. When the location of the coronal cut was varied systematically, it was found that cuts rostral to the coronal level of the paraventricular nucleus (PVN) neither produced nor prevented overeating, while cuts caudal to the PVN produced a robust hyperphagic response. Thus, the coronal level of the paraventricular nucleus was revealed as the rostral focus of a longitudinal satiety neurocircuitry.
Hyperphagia Obesity Paraventricular n Hypothalamus
FOCAL brain damage of the vent romedia l hypo tha lamus (VMH), tradit ionally produced by electrolyt ic lesions, can result in a wel l -documented syndrome of hyperphagia and obesity [23 ,32] . Early formula t ions of the syndrome postulated the existence of a discrete appet i te satiety center [6 ,58] . Recent data suggest that a broader brain circuitry subserves satiety.
The early reports of the VMH lesion syndrome carefully described the extensive areas of the hypo tha lamus damaged by these lesions [6, 29, 32, 33] . A recent anatomical analysis [23] , conf i rmed by several laborator ies [ 13,44] , reveals that lesions restricted to the VMN have no impact on eating or body weight. The effect ive locus of the VMH lesion lies, instead, rostral to the VMN. In the present report we conf i rm with coronal knife cuts the local izat ion of the anatomical substrate of hypo tha lamic hyperphagia to this more rostral plane. We also tenatively implicate the paraventr icular nucleus (PVN) which is located dorso- rostral to the VMN, and its project ions , as the most rostral neuroana tomica l focus within a broader , longi tudinal ly or iented satiety neurocircui t ry .
The evidence support ing these suggestions takes two main lines. First, parasagittal knife cuts extending to the
base of the brain alongside the medial ly located PVN produce hyperphagia and obesi ty [22 ,23] . The effect ive lesions and the effect ive parasagittal cuts are thus both at approximate ly the same rostral level. This suggests that satiety project ions enter or exi t the PVN or an adjacent midline structure en route from and /or to sites fur ther from the midline.
The second major line of evidence support ing a role of the PVN in appeti te regulation is provided by the demon- strat ion of the PVN as the site within the brain where norepinephr ine (NE) and epinephrine (E) infusions elicit eating at the lowest dose thresholds [42] . The PVN has been shown to be heavily innervated by NE and E terminals [35, 60, 61] . The paradoxical findings that NE st imulat ion or e lectrolyt ic or knife cut destruct ion in the PVN area both increase food intake can be accommoda ted [16 ,34] . The impor tan t fact is that lesion, knife cut, and chemical application studies all point towards one c o m m o n neuro- anatomical focus.
Overeating and weight gains have also been repor ted to result f rom the asymmetr ica l combina t ion of the above described parasagittal hypotha lamic knife cut on one side of the brain, and a contralateral mammil lary area lesion
1Supported by National Institutes of Mental Health Grant No. MH-26251 to R. M.G. We thank Jay Alexander and Patti Powers for technical assistance.
l l l l

1112 GOLD, JONES, SAWCHENKO AND KAPATOS
[ 2 8 ] , f rom cent ra l dep le t ions of the m o n o a m i n e neuro- t r ansmi t t e r s n o r e p i n e p h r i n e (NE) [ 1, 2, 4, 34, 38, 44] or se ro ton in ( 5 - H T ) [ 1 0 , 4 9 ] , and f rom bi la teral ly sym- metr ica l knife cuts in each of the three s t e reo tax ic planes [5, 21, 30, 46, 53, 56 ] . Collect ively, these data suggest the invo lvemen t of long i tud ina l (ascending a n d / o r descending) p ro jec t ions in the VMH lesion syndrome .
Some i n f o r m a t i o n on the pa th of these fibers is already available. As m e n t i o n e d previously, bi la teral parasagi t ta l knife cuts alongside the PVN ( ros t ro la te ra l to the VMN) are effect ive in p roduc ing h y p o t h a l a m i c obes i ty [ 2 1 , 2 2 ] . Such cuts, typica l ly 3 m m in height , and e x t e n d i n g to the base of the brain, are effect ive in parasagi t ta l planes 0.5 1.5 mm from the midt ine [ 2 2 , 5 5 ] .
Sclafani [53] used bi lateral ly symmet r i ca l coronal tran- sect ions e x t e n d i n g on each side of the midl ine at co- o rd ina tes an t e r io r and pos te r io r to the VMN. No hyper- phagia was observed fo l lowing any of Slafani 's cuts. In cont ras t , Grossman and Hennessey [30] repor t b o t h hype rphag ia and hyperd ips ia wi th more laterally extens ive coronal cuts pos te r ior to the VMN. It appears f rom these two studies tha t at coronal levels a round the VMN the fibers crit ical to obes i ty lie be tween 1.0 and 2.0 m m from the midl ine.
A more deta i led map of the route of these sat ie ty p ro jec t ions should foster more accurate ana tomica l , neuro- chemical , and behaviora l cha rac te r i za t ions of sat iety. The knife cut t e chn ique is well sui ted to such mapping . Whereas lesion t echn iques are relatively ind i sc r imina te in t ha t they tend to des t roy b o t h cellular e l ement s and axons at the lesion site, knife cuts pr imar i ly sever axons. Only sec- ondar i ly do knife cuts damage neu rona l cell bodies con- nec ted to the severed axons. Knife cuts can also reveal the d i rec t ion in which crit ical f ibers run, as they sever only those axons tha t cross the plane of the cut .
In previous work we have used asymmet r i ca l lesions to map the p ro jec t ions of h y p o t h a l a m i c aphagia and adipsia [ 2 0 ] , and, more recen t ly , of h y p o t h a l a m i c obes i ty [ 2 8 , 3 8 ] . In the first e x p e r i m e n t of the present repor t we combine the advantages of the two techn iques , knife cuts and a s y m m e t r y , to e x t e n d and ref ine the m app ing of the longi tud ina l sys tem damaged in the p r o d u c t i o n of hypo- tha lamic obesi ty .
EXPERIMENT 1
Rats were given a uni la tera l parasagi t ta l h y p o t h a l a m i c knife cut at the rostral locus at which bi la teral cuts rel iably p roduce hype rphag ia and obes i ty [ 2 2 ] . In add i t ion , each rat received a con t ra l a t e ra l h y p o t h a l a m i c or m idb ra in cu t in the coronal plane. By varying the loca t ion of the coronal cut we have t raced our p resen t cons t ruc t of the sat ie ty c i rcui t ry .
Method
Animals. Carwor th CFE and Charles River CD female a lb ino rats weighing 2 1 0 - 2 3 0 g were individual ly housed at 21 ± 2°C, u n d e r cycling light ( 1 2 h r on, 12 hr off) wi th unres t r i c ted access to tap water and to a high fat diet composed , by weight, of 2 parts Pur ina Lab Chow powder , 1 par t vegetable shor t en ing [ 1 2 , 2 1 ] . A m i n i m u m of 10 days a d a p t a t i o n to these cond i t i ons preceded surgery.
Surgery and Histology. All surgery was p e r f o r m e d u n d e r e the r anesthesia . A uni la tera l parasagi t ta l cut was made with a caudal ly d i rec ted 3 m m long re t rac t ing wire knife
[ 2 7 ] . With the incisor bar 3 m m below the in teraural line, the guide cannu la was lowered at 8 . 0 - 8 . 5 m m an te r io r to the earbars , 0.9 mm lateral to the midl ine . At this pos i t ion the wire knife was e x t e n d e d , lowered unt i l it con t ac t ed the base of the skull, and raised 3.0 ram. The wire knife was then re t r ac ted in to its guide cannula . In addi t ion , each rat received a con t ra la te ra l coronal cut with a 2.0 mm long medial ly d i rec ted wire which typical ly t ransec ted the b o t t o m 3 . 0 m m of the brain. The an te r io r -pos te r io r co- o rd ina te of the coronal cut was sys temat ica l ly varied.
Sham opera ted con t ro l animals were similarly t rea ted excep t tha t thei r brains were not invaded b e y o n d the pr icking of the dura.
Af ter behavioral data had been col lected for a m i n i m u m of 14 days pos topera t ive ly , animals were sacrificed. Frozen ho r i zon ta l sect ions were prepared th rough the knife cut areas and s ta ined with cresyl violet . The cuts were then r econs t ruc t ed based on the i r histological local izat ions. Data were only cons idered from those animals whose parasagit tal cut lay alongside the PVN 122], and which were 0.6 1.2 mm lateral to the midline.
Results and Diseuss#m
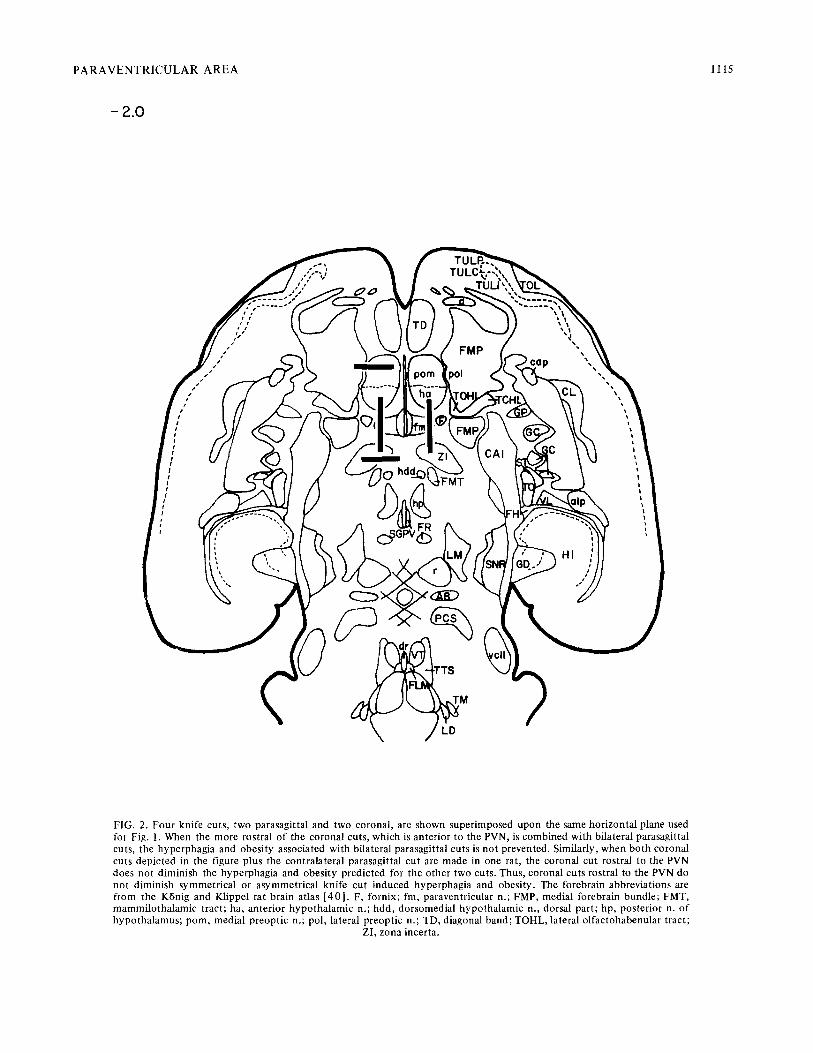
In Fig. 1 the coronal po r t i ons of these a symmet r i ca l cuts are supe r imposed on a ho r i zon ta l p lane passing t h r o u g h the PVN and parallel to the ho r i zon ta l zero plane of the K6nig and Klippel Atlas [401 (Jones, Gold, Simson and Zoll iker , hor izon ta l brain atlas, in p repa ra t ion ) .The n u m b e r next to each cut is the weight gain in g/day for the first two weeks af ter surgery. The water - food ra t io for the same interval is given in parentheses . Individual cuts appear on whichever side of Fig. 1 is least crowded. The cons t an t con t ra la te ra l parasagi t ta l cuts are no t represented . For a typical para- sagittal cut r e cons t ruc t i on see Fig. 2.
The data of Fig. I define a longi tudina l food sat ie ty system ex t end ing ros t ro-caudal ly to ( a n d / o r f rom) the precise coronal level of the PVN. As the cuts move rostrally past the PVN the rate of weight gain decreases dramat ica l ly over a dis tance of less than 0.4 mm. When the coronal cut is at or beh ind the caudal margin of the PVN the rats gain as m u c h as 10 13 g/day. Rapid weight gains persist as the cuts proceed caudal ly th rough the h y p o t h a l a m u s . Coronal cuts at or in f ron t of the rostral margin of the PVN produce weight gains comparab le to con t ro l v a l u e s ( 0 4 g / d a y ) . T h e caudal target (or source) of this p ro jec t ion has regre t tab ly not yet been de te rmined . Our data, plus extensive work in progress, and repor ts f rom o ther labora tor ies , suggest tha t there is no clearly de l ineated caudal focus.
In the medial- lateral d imens ion , effect ive coronal cuts had to e x t e n d more than 1 m m from t h e m i d l i n e . W i t h t h e excep t ion of one Y.gg /day (coronal coord ina te + 3.6) paradoxica l rat, coronal cuts l imi ted to the mos t medial one m m produced , at best. modes t (e.g., 4 g /day) weight gains. The high (2.91 water - food rat io of the paradoxica l 9.9 g/day weight gain rat wilh a medial cut suggests tha t the p i tu i ta ry , which underl ies the brain at this poin t , may possibly have been invaded by the cut . U n f o r t u n a t e l y , tha t rat ' s p i tu i ta ry was no t examined his tological ly.
Ten sham opera ted con t ro l rats gained 0.0--1.5 g/day, and ten rats wi th a single (uni la tera l ) parasagi t ta l cut gained 0.6 2.6 g/day. The best of the asymmet r ica l cuts p roduced fully as rapid gains as we have ever seen with bi lateral parasagit tal knife cuts. Ten rats with bi lateral parasagi t ta l knife cuts gained a mean of 10.1 g /day over the two weeks af ter surgery. On rare occasion, e lec t ro ly t ic lesions can

P A R A V E N T R I C U L A R A R E A 1113
1 3 -
1 2 -
I I - -
I 0 -
9 - -
8 - - m
m
7 - -
6 ~ -
S. 5 - -
n
m
m
4 - -
. 3 - - m
2 - -
o7
- I S- m
m
m
/ / 0
i1~ I I
' / C- t ! C'-'; " , , - ' J l ~- . . I / III1 W - - 2., (~ .s ) 'X '~
" . - 4 / f " ° L s " ~ . - " '!11 - ~ _ , . o ( 3 . , ) ~ ' X J . , ~ "~ f I .11 13.4.J . I - - ' l i i I , - - - . _ - . ~ , 1.4 (2 e ) ~ ~.~
0,3 t3.41 - - ~ - -
l f l t _ , . ~,,L,e tz.,) :_ ._. . . . ,.i I I ~ ' - r , ~ . ~ - _ e.e (,.,) ~ ' , , I 11 / k ~ ~ . ~ " , . o ( , . , ) , , . , ) .~ . .o , , ~ . . , . , ( . ~ . / \
r s t, . . . . . . ) a~ z ) f " I - - , . , s . . . . . - ~ _ , z s ( 4 4 ) ,O0(eO) ' • f m ' ' ' • • ~ 7 . 9 e.
/ / 1 ~ ' , , ~ 7 , . o ( , . , ) , / ~ o . - ~ . , , , _ . , , ~ : . , . , . - ~ , ~ . o ( , , ) v ,..7-~--~ L 1.31 t~-4s ~ | 4 . W , % ~ I ~1 / " I l l ( , .,.,<,.,, t
L/ l ~ ) I , t . o ~ , . 4 j ~ ~ l ~ ._ . .~ - - - ' i . s . , ( , . 3 ) / L - ) / / _ 3 . ' ~ , o)1 _ _ "" !' '~ ~---'~.<o c~ n ~ _ . ~ " ( ' ' ) \ ~m--", I f ~.,,,~"-- / II , L s ~ , . s . , ~ / J'~' " % t I ' , / I
• , ' r L _ . 3 f ; I , . , ( , . , ) . ~ _ v c-, / ~ - " ~', /~ r- ~ - ' , - - - ~ t / / ,,., <,.,) ~ - - - - - b l " t ' ~ '*'"., (,.,) \ \ , . , ( , . ; ) /
/ ,~_. . . - - -~I / ~ 1 /ll~----~-~ ,.,(,-,) ~ \ ~ . 4 .
, ( , ' "
~ . 3 . 9 (Ls)
~,~, ~ 6.4 ( I . r )
I I i , l l f i l l l J I I I l l l l l l l l l l i l l l l l ] l l l i l i l l l l l l l i l I l l i l , i
6 5 4 3 2 I 0 I 2 3 4
FIG. 1. Coronal knife cuts superimposed upon a horizontal section of the brain. Each cut was unilateral and was combined with a contralateral parasagittal knife cut (not shown) such as the one shown in Fig. 2. The postoperative weight gain (g/day/2 weeks) is shown next to each cut along with the postoperative water/food ratio, which is in parenthesis. The coronal cuts are drawn on whichever side of the figure is least crowded. The names of structures are deleted for clarity, but can be found in Fig. 2. The two parasagittal cuts shown in the midbrain alongside the raphe nuclei were also combined
with contralateral parasagittal hypothalamic knife cuts. fm = paraventricular nucleus.

1114 GOLD, JONES, SAWCHENKO AND KAPATOS
produce slightly greater rates of weight gain, but this requires extensive h y p o t h a l a m i c des t ruc t i on [23 ,241 .
The possibi l i ty tha t coronal cuts rostral to the PVN sever bo th sat ie ty and food- in take- in i t i a t ing f ibers and thus p reven t the express ion of h y p o t h a l a m i c obes i ty was ex- plored. Two rats received bi lateral parasagi t ta l cuts plus a uni la teral coronal cut rostral to the PVN (see Fig. 2). Bo th gained weight rapidly (10 and 11 g/day) . Two o the r rats received a uni la tera l parasagi t ta l cut c o m b i n e d wi th two con t ra la te ra l corona l cuts, one an ter ior , and one pos ter ior , to the PVN. These rats also gained weight rapidly (9 and 10 g/day) . Thus, a coronal cut rostral to the PVN ne i the r p roduces no r prevents h y p o t h a l a m i c obesi ty .
Bilateral parasagi t ta l cuts 0 . 5 - 1 . 5 m m f rom the midl ine will p roduce hype rphag ia and obes i ty provided the cuts sever the area alongside and slightly caudal to the PVN. A midl ine sagittal cut in one rat was w i t h o u t ef fec t (weight gain of 1.8 g/day) . This impl icates a s t ruc tu re near the midl ine such as the PVN, or f ibers tha t sweep close to the midl ine , but no t a midl ine crossing p ro jec t ion such as the supraopt ic commissure .
The p rec ip i tous d rop-of f in the rate of weight gain precisely as the corona l cuts pass the PVN indica tes t ha t the longi tudina l sat ie ty f ibers ascend to or descend f rom the PVN itself, or some s t ruc ture jus t vent ra l to the PVN. The PVN is so p r o m i n e n t l y staining, and the series of rats so large, tha t we are mos t con f iden t in this conclus ion . Indeed, the PVN served us as an ana tomica l l andmark long before it became evident tha t the sa t ie ty p ro jec t ions went to or f rom its coronal level. Relative to o the r bra in s t ruc tures , the PVN appears to be heavily c o m m i t t e d as a source, a long wi th the supraopt ic nucleus , of the pos te r ior p i tu i t a ry h o r m o n e s oxy toc in and vasopressin. These h o r m o n e s , involved, re- spect ively, in lac ta t ion and water r e t en t ion , have no t previously been impl ica ted in h y p o t h a l a m i c obesi ty .
With the bi lateral parasagi t ta l knife cuts or e lec t ro ly t ic lesions tha t p roduce obesi ty , w a t e r / f o o d rat ios are fre- quen t ly dramat ica l ly elevated [ 5 9 ] , o f t en going as high as 10.0 (unpub l i shed observat ions) . However, u p o n re- cons t ruc t i on of those symmet r i ca l cuts or lesions we have never been able to de t e rmine how the brain damage 9f the hyperdips ic plus hype rphag ic rats differed from tha t of the rats tha t were only hyperphagic . Hence, we have never been able to plan surgery so as to cons i s ten t ly include or exc lude overdr inking. However, a f te r a symmet r i ca l knife cuts the water in takes did sort in to dis t inct n e u r o a n a t o m i c a l groupings. When the coronal cut lay rostral to or im- media te ly beh ind the PVN, the w a t e r / f o o d rat ios were modera te ly elevated, wereas coronal cuts more t han 0.5 m m caudal to the PVN p roduced only elevated food intakes. For a very short dis tance, abou t 0.5 mm, just beh ind the PVN, b o t h wa te r / food rat ios and food in takes were excessive. We suspect tha t many of the elevated wa te r / food ratios, especially those caused by cuts close to the PVN, are a result of damage to the paravent r icu lo- sup raop t i co -hypophysea l t ract and result f rom an anti- d iuret ic h o r m o n e def ic iency. In Fig. 1 indiv idual post- operat ive wa te r / foo d rat ios are given in parentheses . Pre- operat ively , the range of wa te r / food ratios was 1.4 2.7 (mean = 2.0).
Note tha t the wa te r / food rat ios of the hype rphag ic rats with more caudal coronal cuts are cons i s ten t ly below 2.0. Absolu te water in takes in these animals were elevated as compared wi th cont ro l s , but the i r food in takes were elevated even more.
Cons i s ten t hype rphag ia and obesi ty w i t h o u t elevated w a t e r / f o o d rat ios can now con f iden t l y be p r o g r a m m e d by using asymmet r ica l knife cuts provided the coronal cu t is 1 - 3 m m caudal to the PVN. This conf i rms our p red ic t ion tha t a symmet r i ca l knife cuts should be more selective than symmet r ica l cuts.
EXPERIMENT 2
Having es tab l i shed the a symmet r i ca l knife cu t p rocedure as a reliable means of p roduc ing excessive weight gains, we u n d e r t o o k to direct ly compare aspects of the overeat ing s y n d r o m e in groups of animals bear ing bi la teral parasagi t ta l versus a symmet r i ca l knife cuts.
Diet appears to be cri t ical to the express ion of each of the overea t ing s y n d r o m e s he re to fo re descr ibed. While even rats with in tac t bra ins can be t e m p t e d in to obesi ty by highly pala table supe rmarke t diets [ 5 4 , 5 7 ] , VMH lesioned rats are even more responsive to diet pa la tabi l i ty . In general, b o t h VMH lesion and parasagi t ta l knife cut obesi t ies are mos t fully mani fes t wi th high-fat diets, wi th lesser ef fec ts observed using pel le ted lab chow and li t t le or no overea t ing seen on powdered lab chow [ 1 2 , 5 3 ] . H y p o t h a l a m i c NE deple t ions , b rough t abou t by e i the r midbra in or h y p o t h a l a m i c lesions, p roduce signif icant increases in food in take on high-fat , bu t no t pel leted diets [44 ] . However, a small but cons i s ten t hyperphag ia has been repor ted in ra ts on powdered diets af ter midbra in lesions of ascending NE p a t h w a y s [ 1,2]. In addi t ion , while relat ively specific 5 - H T dep le t ions resul t ing f rom lesions of the midbra in r aphe nuclei b lock the deve lopmen t of VMH lesion obes i ty in rats on a pel le ted diet , these animals do subsequen t ly overeat when offered high-fat diets [14 ] .
Linear g rowth and adipos i ty also appear to be dif- ferent ia l ly a f fec ted by the various brain incurs ions which p roduce overeat ing. While l inear g rowth rates are repor ted to range f rom norma l to s tun ted af ter VMH lesions [7, 9, 18] , excessive l inear g rowth fol lows bi la teral ly symmet r i ca l parasagit tal knife cuts [ 2 6 , 4 5 ] . Both man ipu la t ions , how- ever, p roduce increased adiposi ty (i.e., obesi ty) . In cont ras t , centra l 5 HT dep le t ions which p roduce overea t ing are repor ted to p roduce increased l inear g rowth w i t h o u t obes i ty [49 ] .
Method
Animals. Adul t female rats (Charles River C D ) w e i g h i n g 2 2 0 - 2 5 0 g were employed . All cond i t ions were as in E x p e r i m e n t 1 save tha t in lieu of the high-fa t diet , the rats were offered Purina lab chow pel le ts ad lib.
Surgery and Procedure. Genera l surgical p rocedures were as in Expe r imen t 1. Coord ina tes for the 3 mm tong × 3 mm high parasagi t ta l cuts were 8.0 m m an te r io r to earbars and 0.9 m m lateral to the midline. F rom this po in t , the cuts ex t ended caudally. The 2 m m long x 3 mm high coronal cut coord ina tes were an te r io r 5.5 mm and lateral 2.1 ram. F rom this po in t the cuts ex t ended medially. In this manne r , l 1 rats were given bilateral parasagi t ta l cuts, six were given asymmetr ica l cuts, as in Expe r imen t 1, and six were sham operated.
Naso-anal lengths, an index of l inear g rowth , were measured with calipers at surgery and 20 days later. Each animal ' s Lee Obesi ty lndex [ 4 1 ] , a measure highly cor- re lated with adiposi ty [ 8 ] , was ca lcula ted f rom naso-anal lengths and body weight by the fo rmula Lee Index = (104 ) (~ /Body Weight (g) ) /Naso-Anal Length (mm. ) Food in takes
P A R A V E N T R I C U L A R A R E A 1115
- 2 . 0
• i
,,;"V
!
/
\:,,, ,,;,' / " FMP "',
, , / / ~ ~ pol '
i )J/ ("C'~ / ~.L, )~I_LI~I'~,FMp)) \ g-~'~ \\/~ ', I
HI
#
FIG. 2. Four knife cuts, two parasagittal and two coronal, are shown superimposed upon the same horizontal plane used for Fig. 1. When the more rostral of the coronal cuts, which is anterior to the PVN, is combined with bilateral parasagittal cuts, the hyperphagia and obesity associated with bilateral parasagittal cuts is not prevented. Similarly, when both coronal cuts depicted in the figure plus the contralateral parasagittal cut are made in one rat, the coronal cut rostral to the PVN does not diminish the hyperphagia and obesity predicted for the other two cuts. Thus , coronal cuts rostral to the PVN do not diminish symmetr ical or asymmetr ical knife cut induced hyperphagia and obesity. The forebrain abbreviations are from the KBnig and Klippel rat brain atlas [ 40 ] . F, fornix; fm, paraventricular n.; FMP, medial forebrain bundle; FMT, mammilo tha lamic tract; ha, anterior hypotha lamic n.; hdd, dorsomedial hypotha lamic n., dorsal part; hp, posterior n. o f hypotha lamus ; pom, medial preoptic n.; pol, lateral preoptic n.; TD, diagonal band; TOHL, lateral olfactohabenular tract;
ZI, zona incerta.
1116 GOLD, JONES, SAWCHENKO AND KAPATOS
2 0 0 "
E 1 7 5 -
L~J 1 5 0 -
Z <Z T 125 - Q)
F - "1- I 0 0 -
L9
uJ 7 5 -
>- 50- Q 0
25-
~ \ ~ 0 ,_.
12
E I0
LIJ
Z
- - 6 ell O O 4 i,
. . . . Asymmet r i ca l Cuts (6)
= -= S y m m e t r i c a l Cuts GRI (6)
c ~ Symmet r i ca l Cuts GRrt (5)
........... Sham Operated (6)
/ / . . - / / / / l
" l J
I J
t
[ lllllr ////Z-
I I I I I I I I I I 1 I I I
2.25-, < 2 . 0 0 - (lC
C3 1 . 7 5 O O
LIJ }-- 1 .25
/
<[ I I 1 I l I 1 1
- 6 - 4 - 2 I' 2 4 6 8 I0 12 14 16 18 20 Surgery
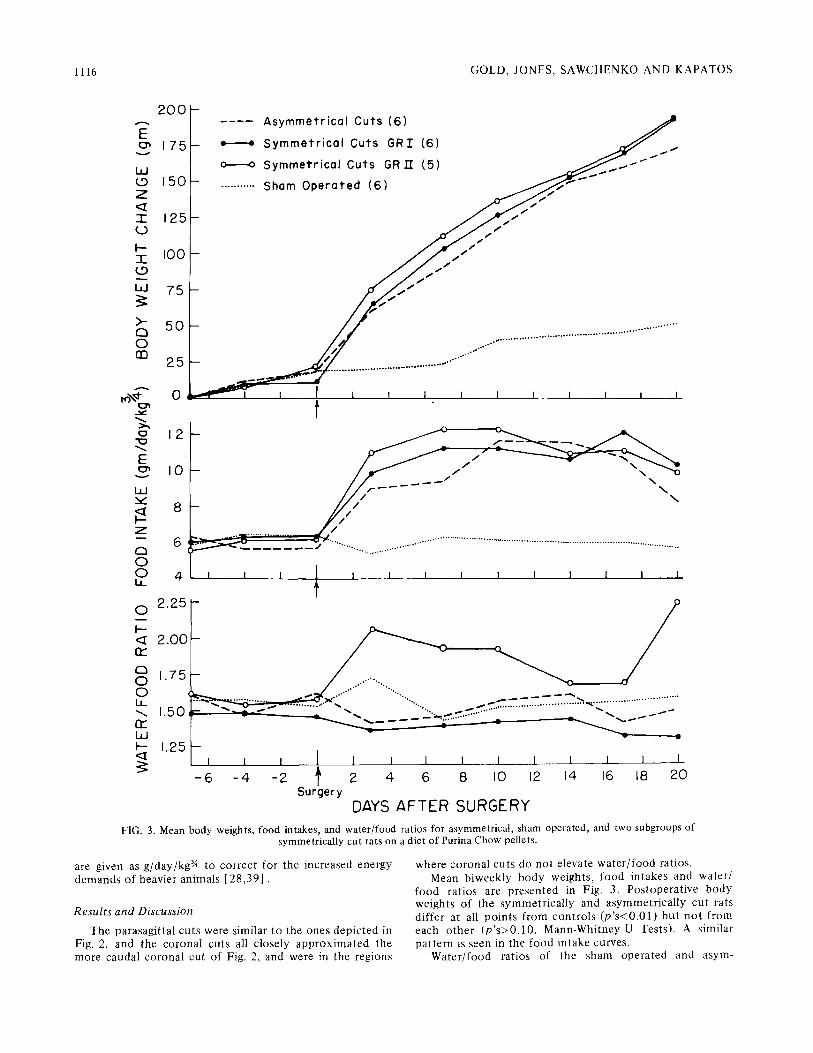
DAYS AFTER SURGERY FIG. 3. Mean body weights, food intakes, and water/food ratios for asymmetrical, sham operated, and two subgroups of
symmetrically cut rats on a diet of Purina Chow pellets.
are given as g /day /kg 3/" to cor rec t for the increased energy demands of heavier animals [ 2 8 , 3 9 ] .
Results and Discussion
The parasagi t ta l cuts were similar to the ones depic ted in Fig. 2, and the coronal cuts all closely a p p r o x i m a t e d the more caudal corona l cut of Fig. 2, and were in the regions
where coronal cuts do not elevate w a t e r / f o o d ratios. Mean b iweekly body weights, food in takes and water /
food rat ios are presen ted in Fig. 3. Pos topera t ive body weights of the symmet r ica l ly and asymmet r ica l ly cut rats dif fer at all po in t s f rom con t ro l s ( p ' s < 0 . 0 1 ) bu t n o t f rom each o t h e r ( p ' s > 0 . 1 0 , Mann-Whi tney U Tests). A similar pa t t e rn is seen in the food in take curves.
Wate r / food rat ios of the sham opera ted and asym-
PARAVENTRICULAR AREA 1117
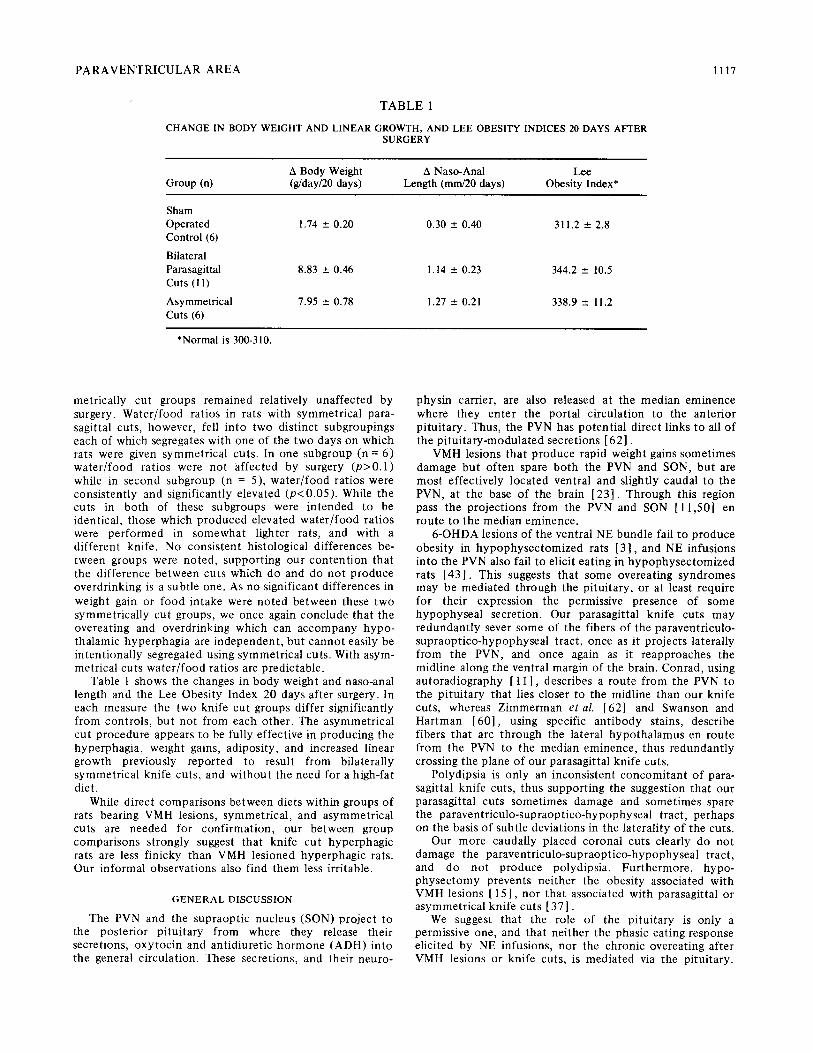
TABLE 1
CHANGE IN BODY WEIGHT AND LINEAR GROWTH, AND LEE OBESITY INDICES 20 DAYS AFTER SURGERY
A Body Weight A Naso-Anal Lee Group (n) (g/day/20 days) Length (mm/20 days) Obesity Index*
Sham Operated 1.74 ___ 0.20 0.30 _+ 0.40 311.2 ___ 2.8 Control (6)
Bilateral Parasagittal 8.83 _+ 0.46 1.14 +_ 0.23 344.2 _+ 10.5 Cuts (11)
Asymmetrical 7.95 _+ 0.78 1.27 --- 0.21 338.9 _+ 11.2 Cuts (6)
*Normal is 300-310.
metrically cut groups remained relatively unaffected by surgery. Water/food ratios in rats with symmetrical para- sagittal cuts, however, fell into two distinct subgroupings each of which segregates with one of the two days on which rats were given symmetrical cuts. In one subgroup (n = 6) water/food ratios were not affected by surgery (p>0.1) while in second subgroup (n = 5), water/food ratios were consistently and significantly elevated (p<0.05). While the cuts in both of these subgroups were intended to be identical, those which produced elevated water/food ratios were performed in somewhat lighter rats, and with a different knife. No consistent histological differences be- tween groups were noted, supporting our contention that the difference between cuts which do and do not produce overdrinking is a subtle one. As no significant differences in weight gain or food intake were noted between these two symmetrically cut groups, we once again conclude that the overeating and overdrinking which can accompany hypo- thalamic hyperphagia are independent, but cannot easily be intentionally segregated using symmetrical cuts. With asym- metrical cuts water/food ratios are predictable.
Table 1 shows the changes in body weight and naso-anal length and the Lee Obesity Index 20 days after surgery. In each measure the two knife cut groups differ significantly from controls, but not from each other. The asymmetrical cut procedure appears to be fully effective in producing the hyperphagia, weight gains, adiposity, and increased linear growth previously reported to result from bilaterally symmetrical knife cuts, and without the need for a high-fat diet.
While direct comparisons between diets within groups of rats bearing VMH lesions, symmetrical, and asymmetrical cuts are needed for confirmation, our between group comparisons strongly suggest that knife cut hyperphagic rats are less finicky than VMH lesioned hyperphagic rats. Our informal observations also find them less i r r i t ab le .
GENERAL DISCUSSION
The PVN and the supraoptic nucleus (SON) project to the posterior pituitary from where they release their secretions, oxytocin and antidiuretic hormone (ADH) into the general circulation. These secretions, and their neuro-
physin carrier, are also released at the median eminence where they enter the portal circulation to the anterior pituitary. Thus, the PVN has potential direct links to all of the pituitary-modulated secretions [62].
VMH lesions that produce rapid weight gains sometimes damage but often spare both the PVN and SON, but are most effectively located ventral and slightly caudal to the PVN, at the base of the brain [23]. Through this region pass the projections from the PVN and SON [11,50] en route to the median eminence.
6-OHDA lesions of the ventral NE bundle fail to produce obesity in hypophysectomized rats [3], and NE infusions into the PVN also fail to elicit eating in hypophysectomized rats [43]. This suggests that some overeating syndromes may be mediated through the pituitary, or at least require for their expression the permissive presence of some hypophyseal secretion. Our parasagittal knife cuts may redundantly sever some of the fibers of the paraventriculo- supraoptico-hypophyseal tract, once as it projects laterally from the PVN, and once again as it reapproaches the midline along the ventral margin of the brain. Conrad, using autoradiography [ 1 1 ], describes a route from the PVN to the pituitary that lies closer to the midline than our knife cuts, whereas Zimmerman e ta l . [62] and Swanson and Hartman [60], using specific antibody stains, describe fibers that arc through the lateral hypothalamus en route from the PVN to the median eminence, thus redundantly crossing the plane of our parasagittal knife cuts.
Polydipsia is only an inconsistent concomitant of para- sagittal knife cuts, thus supporting the suggestion that our parasagittal cuts sometimes damage and sometimes spare the paraventriculo-supraoptico-hypophyseal tract, perhaps on the basis of subtle deviations in the laterality of the cuts.
Our more caudally placed coronal cuts clearly do not damage the paraventriculo-supraoptico-hypophyseal tract, and do not produce polydipsia. Furthermore, hypo- physectomy prevents neither the obesity associated with VMH lesions [ 15], nor that associated with parasagittal or asymmetrical knife cuts [ 37 ].
We suggest that the role of the pituitary is only a permissive one, and that neither the phasic eating response elicited by NE infusions, nor the chronic overeating after VMH lesions or knife cuts, is mediated via the pituitary.

11 18 GOLD, JONES, SAWCHENKO AND KAPATOS
This suggest ion is f u r t he r suppor t ed by the recent repor t tha t sys temic co r t i cos t e rone re ins ta tes (bu t does no t by itself elicit) ea t ing in response to NE infus ions in to the PVN in sated h y p o p h y s e c t o m i z e d rats [ 4 3 ] .
The rou te of the sa t ie ty c i rcui t ry depic ted by our coronal cuts (Fig. 1) is cons i s t en t wi th cu r ren t maps of b o t h ascending NE [60 ,61] and 5 - H T [19 ,61] systems. Asymmet r i ca l cuts whose coronal m e m b e r lies near the m i d b r a i n - p o n t i n e origins of these sys tems are, however , general ly less effect ive in p roduc ing excessive weight gains than are more rostral cuts.
The magn i tude of the overea t ing observed af ter asym- metr ical knife cuts leads us to conc lude tha t damage to NE and 5 - H T sys tems may each c o n t r i b u t e to the syndrome , but ne i the r dep le t ion a lone can accoun t for the en t i re spec t rum of the syndrome . NE-deple t ion hype rphag ia is b locked by h y p o p h y s e c t o m y [ 3 ] , while tha t p roduced by e i the r symmet r i ca l or a symmet r i ca l knife cuts is no t b locked by h y p o p h y s e c t o m y [ 3 7 ] . Centra l 5-HT dep le t ions can p roduce overea t ing and e n h a n c e d weight gain, but this is en t i re ly accoun ted for by increased l inear growth , no t by increased ad ipos i ty [ 4 9 ] . Af ter knife cuts, increased adipos i ty and l inear g rowth are b o t h apparen t . It seems the re fore , f rom b o t h ana tomica l and behaviora l con- s iderat ions, t ha t i n t e r r u p t i o n by knife cuts of ascending NE a n d / o r ascending 5-HT sys tems can c o n t r i b u t e to hypo- tha lamic hyperphagia . It is likely, however , t ha t the i n t e r rup t i on of add i tona l neuroc i rcu i t ry , pe rhaps a de- scending p ro jec t ion , is also involved.
A descending sa t ie ty sys tem is suppor t ed by the recent f indings tha t subd iaph ragma t i c vago tomy reverses VMH lesion obes i ty [ 4 8 ] , and tha t subd iaphragmat i c v a g o t o m y (but no t a more selective gastric v a g o t o m y ) reverses knife cut induced obes i ty [ 5 1 ] . These data t en ta t ive ly suggest
tha t a p ro jec t ion f rom the PVN may inh ib i t the vagus. Severing this p ro jec t ion would, accord ing to this mode l , d i s inhib i t the hepa t ic a n d / o r coeliac b ranches of the subd iaphragmat i c vagus. This d i s inh ib i t ion of the vagus migh t be responsib le , for example , for the hype r in su l i nemia associated wi th , and perhaps c o n t r i b u t o r y to, VMH lesion and knife cut induced obes i t ies [7, 31, 3 6 ] . Direct ( m o n o s y n a p t i c ) c o n n e c t i o n s be tween per ikarya in the region of the PVN and the vagal nuclei of the h i n d b r a i n
have been descr ibed [ 5 0 ] . The me tabo l i c mechan i sm of h y p o t h a l a m i c obes i ty is
clearly still an open ques t ion , wi th many in te res t ing possibil i t ies. However this issue may f inal ly resolve, a centra l role for the PVN, or the region precisely vent ra l to and ros t ro-caudal ty coextens ive wi th the PVN, is revealed by our data.
We, and o thers te.g., [30, 47, 5 4 ] ) are cu r ren t ly explor ing each of the dys func t ions associated wi th hypo- tha lamic hyperphagia . Those tha t persist in a symmet r i ca l ly cut ra ts may be t ru ly indicat ive of the func t ion ( s ) of the c i rcui t ry tha t mus t be damaged in order to p roduce obesi ty. Those dys func t ions tha t drop out are likely due to damage to over lapping but func t iona l ly unre la t ed neuroc i rcu i t ry .
To date , the polydipsia , i r r i tabi l i ty , f inickiness ( this repor t ) , and sexual dys func t i ons (47,53.] , Gold and Gen t ry , u n p u b l i s h e d ) appear to drop out . The enhanced l inear g rowth tha t persists wi th b o t h symmet r ica l and asym- metr ica l knife cuts suggest a mechan i sm tha t b o t h elicits eat ing and dis inhibi ts the release of g rowth h o r m o n e , the la t ter bu t no t the fo rmer effect being b locked distal to the site of d i s inh ib i t ion by VMH lesions or h y p o p h y s e c t o m y . O the r aspects of the VMH lesion s y n d r o m e tha t endure in knife cut rats are hypoac t iv i t y [ 1 7 , 5 3 ] , hyper insu l inemia , and gastro in tes t ina l h y p o m o t i l i t y [ 52 ].
REFERENCES
1. Ahlskog, J. E. Food intake and amphetamine anorexia after selective forebrain norepinephrine loss. Brain Res. 82: 211-240, 1974.
2. Ahlskog, J. E. and B. G. Hoebel. Overeating and obesity from damage to a noradrenergic system in the brain. Science 182: 166-169, 1973.
3. Ahlskog, J. E., B. G. Hoebel and S. T. Breisch. Hyperphagia following lesions of the noradrenergic pathway is prevented by hypophysectomy. Fedn Proc. 33: 463, 1974.
4. Ahlskog, J. E., P. K. Randall and B. G. Hoebel. Hypothalamic hyperphagia: Dissociation from hyperphagia following de- struction of noradrenergic neurons. Science 190 : 399 401, 1975.
5. Albert, D. J. and L. H. Storlien. Hyperphagia in rats with cuts between the ventromedial and lateral hypothalamus. Science 165: 599-600, 1969.
6. Anand, B. K. and J. R. Brobeck. Hypothalamlc control of food intake in rats and cats. Yale J. Biol. Med. 24:123 140, 1951.
7. Bernardis, L. L. and L. A. Frohman. Effect of lesion size in the ventromedial hypothalamus on growth hormone and insulin levels in the weanling rat. Neuroendocrinology 6 : 3 1 9 328, 1970.
8. Bernardis, L. L. and B. D. Patterson. Correlation between "Lee Index" and carcass fat content in weanling and adult female rats with hypothalamic lesions. J. Endocrin. 4 0 : 5 2 7 528, 1968.
9. Bernardis, L. L. and F. R. Skelton. Growth and obesity following ventromedial hypothalamic lesions placed in female rats at four different ages. Neuroendocrinology 1: 265-275, 1965/1966.
10. Breisch, S. T., F. P. Zemlan and B. G. Hoebel. Hyperphagia and obesity following serotonin depletion by intraventricular p-chlorophenylalanine. Science 192: 382-385, 1976.
11. Conrad, L. C. A. and D. W. Pfaff. Efferents from medial basal forebrain and hypothalamus in the rat: An autoradiographic study of the anterior hypothalamus. J. eomp. Neurol. 169: 221-261, 1976.
12. Corbit, J. D. and E. Stellar. Palatability, food intake and obesity in normal and hyperphagic rats. Z comp. physiol. Psychol. 58:63 67, 1964.
13. Coscina, D. V., D. D. Godse and H. C. Stancer. Neurochemical correlates of hypothalamic obesity in rats. Behav. Biol. 16: 365-372, 1976.
14. Coscina, D. V. and H. C. Stancer. Selective blockage of hypothalamic hyperphagia and obesity in rats by serotonin- depleting midbrain lesions. Science 195:416-418 , 1977.
15. Cox, V. C., J. W. Kakolweski and E. J. Valenstein. Effect of ventromedial hypothalamic damage in hypophysectomized rats. J. comp. physiol. Psychol. 65:145 148, 1968.
16. Cruce, J. A. t:., N. B. Thoa and M. J. Jacobowitz. Catechol- amines in the brains of genetically obese rats. Brain Res. 101: 165--170, 1976.
17. Eng, R. The effect of vagotomy on hypothalanfic knife cut hypoactivity. Paper presented to the Eastern Psychological Association, Boston, 1977.
18. l.'rohman, L. A. and L. L. Bernardis. Growth hormone and insulin levels in weanling rats with ventromedial hypotbalamic lesions. Endocrin. 82:1125 1132, 1968.

P A R A V E N T R I C U L A R A R E A 1119
19. Fuxe, K. and G. Jonsson. Further mapping of central 5-hydro- xytryptamine neurons: Studies with the neurotoxic hydro- xytryptamines. Adv. Biochem. Psychopharmac. I0: 1-12, 1974.
20. Gold, R. M. Aphagia and adipsia following unilateral and bilaterally asymmetrical lesions in rats. Physiol. Behav. 2: 211-220, 1967.
21. Gold, R. M. Hypothalamic hyperphagia: Males get just as fat as females.J, comp. physioL PsychoL 71: 347-356, 1970.
22. Gold, R. M. Hypothalamic hyperphagia produced by para- sagittal knife cuts. Physiol. Behav. 5: 23-25, 1970.
23. Gold, R. M. Hypothalamic obesity: The myth of the ventro- medial nucleus. Science 182: 488-490, 1973.
24. Gold, R. M. Anodal electrolytic brain lesions: How current and electrode metal influence lesion size and hyperphagiosity. PhysioL Behav. 14: 625-632, 1975.
25. Gold, R. M., J. R. Ieni and E. L. Simson. Delayed or precocious hyperphagia after symmetrical or asymmetrical hypothalamic knife cut in male and female weanling rats. Physiol. Behav. 18: 275-281, 1977.
26. Gold, R. M. and G. Kapatos. Delayed hyperphagia and increased body length after hypothalamic knife cuts in weanling rats. J. comp. physiol. PsychoL 88: 202-209, 1975.
27. Gold, R. M., G. Kapatos and R. J. Carey. A retracting wire knife for stereotaxic brain surgery made from a microliter syringe. Physiol. Behav. 10 :813- 815, 1973.
28. Gold, R. M., P. M. Quackenbush and G. Kapatos. Obesity following combination of rostrolateral to VMH cut and contralateral mammillary area lesion. J. comp. physiol. Psychol. 79: 210-218, 1972.
29. Graff, H. and E. Stellar. Hyperphagia, obesity and finickiness. J. comp. physiol. Psychol. 55: 418-424, 1962.
30. Grossman, S. P. and J. W. Hennessy. Differential effects of cuts through the posterior hypothalamus on food intake and body weight in male and female rats. Physiol. Behav. 17: 89-102, 1976.
31. Hales, C. N. and G. C. Kennedy. Plasma glucose, non-esterified fatty acid and insulin concentrations in hypothalamic- hyperphagic rats. J. Biochem. 90: 620-624, 1964.
32. Hetherington, A. W. and S. W. Ranson. Hypothalamic lesions and adiposity in the rat. Anat. Rec. 78: 149-172, 1940.
33. Hoebel, B. G. Feeding and self stimulation. Ann. N. Y. Acad. ScL 157: 758-777, 1969.
34. Hoebel, B. G. Pharmacology of feeding. In: Handbook o f Psychopharmacology, edited by L. L. Iversen, S. D. Iversen and S. H. Snyder. In press.
35. H~;kfelt, T., K. Fuxe, M. Goldstein and O. Johansson. lmmuno-histochemical evidence for the existence of adrenaline neurons in the rat brain. Brain Res. 66: 235-251, 1974.
36. Hustvedt, B. E. and A. Lqvq. Correlations between hyper- insulinemia and hyperphagia in rats with ventromedial hypo- thalamic lesions. Acta physiol, scand. 84: 29-33, 1972.
37. leni, J. R. and R. M. Gold. Two satiety systems revealed by hypothalamic knife cuts in hypophysectomized rats. Physiol. Behav., 1977. In press.
38. Kapatos, G. and R. M. Gold. Evidence for ascending nor- adrenergic mediation of hypothalamic hyperphagia. Pharmacol. Biochem. Behav. 1: 81-87, 1973.
39. Kleiber, M. Body size and metabolic rate. Physiol. Rev. 27: 511 541, 1947.
40. K~Snig, J. F. R. and R. A. Klippel. The Rat Brain: A Stereo- taxic Atlas o f the Forebrain and Lower Parts o f the Brain Stem. Baltimore: Williams and Wilkins, 1963.
41. Lee, M. O. Determination of the surface area of the white rat with its application to expression of metabolic results. Am. J. Physiol. 89: 24-33, 1929.
42. Leibowitz, S. F. Paraventricutar nucleus: A primary site mediating adrenergic feeding-elicitation. Paper presented at the meeting of the Eastern Psychological Association, Philadelphia, 1974.
43. Leibowitz, S. F., K. Chang and R. L. Oppenheimer. Feeding elicited by noradrenergic stimulation of the paraventricular nucleus. Paper presented at the meeting of the Society for Neuroscience, Toronto, 1976.
44. Lorden, J., G. A. Oltmans and D. L. Margules. Central noradrenergic neurons: Differential effects on body weight of electrolytic and 6-hydroxydopamine lesions in rats. J. comp. physiol. Psychok 90: 144-155, 1976.
45. Mitchell, J. A., M. Hutchins, W. J. Schindler and V. Critchlow. Increases in plasma growth hormone concentration and naso- anal length following isolation of the medial basal hypo- thalamus. Neurendocrinology 12:161 - 173,1973.
46. Palka, Y., R. A. Liebelt and V. Critchlow. Obesity following partial or complete isolation of the ventromedial hypo- thalamus. Physiol. Behav. 7: 187-194, 1971.
47. Paxinos, G. The hypothalamus: Neural systems involved in feeding, irritability, aggression, and copulation in male rats. J. comp. physiol. Psychol. 87:110 119, 1974.
48. Powley, T. L. and C. A. Opsahl. Ventromedial hypothalamic obesity abolished by subdiaphragmatic vagotomy. Am. J. Physiol. 226:25 33, 1974.
49. Sailer, C. F. and E. M. Stricker. Hyperphagia and increased g ro w t h in ra ts a f t e r intraventricular injection of 5,7-dihydroxytryptamine. Science 192: 385-387, 1976.
50. Saper, C. B., A. D. Loewy, L. W. Swanson and W. M. Cowan. Direct hypothalamo-autonomic connections. Brain Res. 117: 305-312, 1976.
51. Sawchenko, P. E. Knife cut hypothalamic hyperphagia blocked by complete subdiaphragmatic vagotomy but not by selective gastric vagotomy. Presented to the Eastern Psycho- logical Association, Boston, 1977.
52. Sawchenko, P. E., T. L. Ralph and S. Balagura. Effects of lateral and ventromedial hypothalamic lesions on gastric retention and intestinal transit. Fedn Proc. 34: 397, 1975. (Abstract).
53. Sclafani, A. Neural pathways involved in the ventromedial hypothalamic lesion syndrome in the rat. J. comp. physiol. Psychol. 77: 70-96, 1971.
54. Sclafani, A. Appetite and hunger in experimental obesity syndromes. In: Hunger: Basic Mechanisms and Clinical Im- plications, edited by D. Novin, W. Wyrwicka and G. Bray. New York: Raven Press, 1976.
55. Sclafani, A., C. N. Berner and G. Maul. Feeding and drinking pathways between the medial and lateral hypothalamus in the rat. J. comp. physiol. Psychol. 85 :29 -51 , 1973.
56. Sclafani, A. and S. P. Grossman. Hyperphagia produced by knife cuts between the medial and lateral hypothalamus in the rat. Physiol. Behav. 4: 533-537, 1969.
57. Sclafani, A. and D. Springer. Dietary obesity in adult rats: Similarities to hypothalamic and human obesity syndromes. Physiol. Behav. 17: 461-471, 1976.
58. Stellar, E. The physiology of motivation. PsychoL Rev. 61: 5-22, 1954.
59. Stevenson, J. A. F. Neural control of food and water intake. In: The Hypothalamus, edited by W. Haymaker, E. Anderson and W. J. H. Nauta. Springfield: Charles C. Thomas, 1969.
60. Swanson, L. W. and B. K. Hartman. Central adrenergic s y s t e m - Immunofluorescence study of localization of cell bodies and their efferent connections in rat utilizing dop- amine-beta-hydroxylase as a marker. J. comp. Neurol. 163: 467-505, 1975.
61. Ungerstedt, U. Stereotaxic mapping of the monoamine path- ways in the rat. Acta physiol, scand., Suppl. 367 :1 -48 , 1971.
62. Zimmerman, E. A., P. W. Carmel, M. K. Husain, M. Ferin, M. Tannenbaum, A.G. Frantz and A.G. Robinson. Vasopressin and neurophysin: High concentrations in monkey hypo- physeal portal blood. Science 182:925 927, 1973.