the role of the hypothalamic paraventricular nucleus in
TRANSCRIPT

Loyola University Chicago Loyola University Chicago
Loyola eCommons Loyola eCommons
Dissertations Theses and Dissertations
1989
The Role of the Hypothalamic Paraventricular Nucleus in Stress-The Role of the Hypothalamic Paraventricular Nucleus in Stress-
Induced Renin and Corticosterone Secretion Induced Renin and Corticosterone Secretion
Kathy Richardson Morton Loyola University Chicago
Follow this and additional works at: https://ecommons.luc.edu/luc_diss
Part of the Neuroscience and Neurobiology Commons
Recommended Citation Recommended Citation Morton, Kathy Richardson, "The Role of the Hypothalamic Paraventricular Nucleus in Stress-Induced Renin and Corticosterone Secretion" (1989). Dissertations. 2653. https://ecommons.luc.edu/luc_diss/2653
This Dissertation is brought to you for free and open access by the Theses and Dissertations at Loyola eCommons. It has been accepted for inclusion in Dissertations by an authorized administrator of Loyola eCommons. For more information, please contact [email protected].
This work is licensed under a Creative Commons Attribution-Noncommercial-No Derivative Works 3.0 License. Copyright © 1989 Kathy Richardson Morton

,. , ... ·--:-i···,1Ty UBRJ.\i·:;y .. LG'(()L!\ l::·~ \ \i L.:n,::;:.
. MEDICAL CE:NTEH .: ... l'.:··
THE ROLE OF THE HYPOTHALAMIC PARAVENTRICULAR NUCLEUS
IN STRESS-INDUCED RENIN AND CORTICOSTERONE SECRETION
by
Kathy Richardson Morton
A Dissertation Submitted to the Faculty of the Graduate School
of Loyola University of Chicago in Partial Fulfillment
of the Requirements for the Degree of
Doctor of Philosophy
May
1989

ACKNOWLEDGEMENTS
I express my thanks to Dr. Louis Van de Kar, for providing the
support, guidance and friendship that has made this dissertation a
productive and enjoyable experience. Appreciation is also extended to my
committee members for their suggestions and insightful discussions during
the course of this dissertation work. Special thanks is given to Dr. Mark
Brownfield for his exhaustive help with the irnrnunocytochemistry and
histological analyses; to Dr. Stanley Lorens for use of the stress
chambers and assistance with the sotalol injections; to Dr. Celeste Napier
for experimental design suggestions and help with the microinj ection
techniques; to Dr. Stephen Jones for help with the fluorometric
catecholamine assay; and to Dr. Alexander Karczmar and Dr. Israel Hanin
for their guidance as Departmental Chairmen.
I am grateful to my laboratory co-workers, Ms. Kayoko Kunimoto and
Dr. Janice Urban, for their assistance on many occasions. I also thank
Ms. Heather Read for the preparation of the brain pathways diagram. I
express my gratitude to Ms. Diane Pikowski and her parents, Mr. and Mrs.
Carl Villafan, for their concern and support for both myself and my
daughter Lindsay. I also wish to thank Gerry and Dorothy Hunchak, Toni
Price, Lynn Rohrmann, Loek Van de Kar, Susan Schmitt, Celeste Napier, Ron
Price, Peter Rittenhouse and Denise Lokhorst for their kindness during my
trips to Chicago.
Finally, I gratefully acknowledge the support of the faculty, staff
and students of the Department of Pharmacology; the Graduate School; Dee
Miller; Mary Ann Jurgus; and the financial backing supplied by Loyola
University and the Arthur J. Schmitt Foundation.
ii

DEDICATION
This dissertation is dedicated to my parents,
Virgil and Audrey Bullington, for their constant
support and encouragement, and to my daughter, Lindsay.
iii

VITA
The author, Kathy Richardson Morton, was born on January 26, 1957,
in Knoxville, Tennessee. She completed her secondary education at West
High School in Knoxville, Tennessee in June, 1975. In September, 1975,
Ms. Morton entered the University of Tennessee, where she graduated magna
cum laude with a Bachelor of Arts degree in Biology in June, 1978.
Following graduation, she was employed as an Associate Chemist for the
Bristol-Myers Company in Cincinnati, Ohio. In 1982, Ms. Morton did a
summer rotation in the laboratory of Dr. Robert Ten Eick at Northwestern
University.
In January, 1983, Ms. Morton was accepted as a graduate student in
the Department of Pharmacology at Loyola University of Chicago Stritch
School of Medicine, where she was awarded a Loyola University Basic
Science Fellowship. In July, 1986, she received a Sigma Xi Grant-in-Aid
of Research, and in March, 1987, was awarded an Arthur J. Schmitt
Dissertation Fellowship. In May, 1988, Ms. Morton received an award for
Best Neuroscience Dissertation from Loyola University for the Chicago
Chapter of Neuroscience. The author also served as a Representative to
the Graduate Student Council (1983-84) and the Pharmacology Faculty
Meetings (1987). She is a student member of the Society for Neuroscience.
Ms. Morton has accepted a post-doctoral position in the laboratory
of Dr. Neil MacLusky in the.Department of Reproductive Science at the
University of Toronto and Toronto General Hospital. She has been awarded
a post-doctoral fellowship from the Medical Research Council of Canada.
iv

TABLE OF CONTENTS
ACKNOWLEDGEMENTS. ii
DEDICATION. iii
VITA .. .iv
LIST OF TABLES. .viii
LIST OF FIGURES .. ix
Chapter I . INTRODUCTION. . . . . . . . . . . . . . . . . . . . . . . . . . l
II. LITERATURE REVIEW . A. RENIN ..... .
1. Introduction. 2. Description of Renin. 3. Physiological Aspects of the Angiotensins
a. Angiotensinogen. b. Angiotensin I. . . .. . c. Angiotensin II . . .. .
4. Regulation of Renin Secretion a. Introduction ...... . b. Role of the Sympathetic Nervous System
in Renin Secretion . . . . . . . . c. Role of Extra-Renal Beta Receptors
in Renin Secretion . . . . . . . d. Role of Brain Serotonin Neurons
5 5 5 5 6 6 7 7 9 9
.10
.11
in Renin Secretion . . . . . . . .12 1. Role of Beta Receptors in the Serotonergic
Stimulation of Renin Secretion . . . . .13 e. Role of the Hypothalamic Paraventricular Nucleus
(PVN) in Renin Secretion . . . . . .15 f. Other Hypothalamic Areas Involved
in Renin Secretion . . . . . . . . 16 g. Extrahypothalamic Pathways Involved
in Renin Secretion . . . . . . . . . .17 h. Renin and Angiotensin in the Brain . .17 i. Role of Brain Catecholamines in Renin Secretion. .19
B. CORTICOSTERONE . . . . . . . . . . .21 1. Introduction. . . . . . . . . . .21 2. Corticotropin Releasing Factor (CRF). .22 3. Other Factors Affecting ACTH Release. .25 4. Role of Brain Catecholamines in
Corticosterone Secretion. .26 a. Introduction . . . . .26 b. Anatomical Evidence. .26
v

c. Role of the Ventral Noradrenergic Bundle in Corticosterone Secretion . . . . . . . . . .
d. Stimulation of CRF/Corticosterone Secretion by Injections of Epinephrine and Norepinephrine.
e. Role of Dopamine Neurons in Corticosterone Secretion . . . . . . . . .
5. Role of 5-HT Neurons in Corticosterone Secretion. C. STRESS .... .
1. Introduction .. . a. Definition .. b. Cannon's Alarm Reaction. c. Selye's General Adaptation Syndrome. d. Sympathetic and Endocrine Stress Responses e. Physiological Changes in Response to Stress.
2. The Conditioned Fear Paradigm ..... a. Description. . . . . . . . . . . . . b. Pavlov's Dog: Classical Conditioning c. Definition of Fear d. Measures of Fear . . . . . . . . . e. Emotional Factors ........ .
3. Cardiovascular Changes in Response to Classical Conditioning ................ .
4. Neuroendocrine Changes in Response to Stress. a. Stress-Induced Renin Secretion ..... b. Stress-Induced Corticosterone Secretion.
1. Introduction ....... . 2. Stress-Induced Secretion of CRF .. . 3. Stress-Induced Secretion of ACTH .. . 4. Role of Vasopressin in Stress-Induced
Corticosterone Secretion. . . . . . . 5. Role of Catecholamines in Stress-Induced
Corticosterone Secretion. . . i. Evidence for an Inhibitory Role.
ii. Evidence for a Stimulatory Role. iii. Role of Epinephrine ....
6. Role of Serotonin in Stress-Induced Corticosterone Secretion. . . . . .
c. Effect of Peripheral Sympathectomy on StressInduced Renin and Corticosterone Secretion .
5. Central Nervous System Pathways Involved in the Stress Response . . . a. Sensory Afferents ..... . b. The Limbic System ..... .
1. Septo-Hippocampal System. 2. Amygdala ........ . 3. Hypothalamic Ventromedial Nucleus
c. Role of Brain Catecholamines in Stress D. HYPOTHALAMIC PARAVENTRICULAR NUCLEUS (PVN)
1. Introduction ....... . 2. Morphology ........ . 3. Role of the PVN in Behavior 4. Afferent Connections to the PVN
vi
.29
.30
.31
.32
.35
.35
.35
.35
.36
.37
.38
.38
.38
.39
.40
.40
.41
.42
.45
.45
.47
.47
.47
.48
.so
.51
.51
.53
.57
.. 58
.61
.62
.62
.63
.63
.63
.65
.66
.68
.68
.69
.70
.71

5. Efferent Projections of the PVN ..... 6. Catecholaminergic Innervation of the PVN. 7. Neuroactive Substances in the PVN
a. Neuropeptide Y ........... .
III. METHODS . .
A. Animals B. Surgery C. Electrolytic Destruction of the PVN D. Electrolytic Destruction of the Nucleus Reuniens. E. Microinjection of Ibotenic Acid into the PVN F. Microinjections of 6-0HDA into the PVN. G. Microinjections of Sotalol into the PVN H. Histology ...... . I. Immunocytochemistry ... . J. Stress (CER) Procedure .. . K. Biochemical Determinations.
1. Plasma Renin Activity .. 2. Plasma Renin Concentration 3. Plasma Angiotensinogen 4. Plasma Corticosterone.
L. Statistics . . . . . . . 1. Analysis of Variance . 2. Student-Newman-Keuls' Test
IV. RESULTS ....
A. Electrolytic Lesions in the PVN. B. Electrolytic Lesions in the Nucleus Reuniens c. Injection of Ibotenic Acid into the PVN. D. Injection of 6-0HDA into the PVN E. Injection of Sotalol into the PVN. F. Comparison of Normal Plasma Pools to Experimental
V. DISCUSSION ...
Controls
.73
.74
.74
.75
. 77
. 77
. 77
.78
.78
.79
.80
.81
.82
.82
.83
.85
.85
.87
.88
.88
.89
.90
.91
.93
.93
.98 103 109 116 121
..... 126
A. Overview. . . . . . . . . . . . . . . . . 126 B. Justification of the Stress Paradigm. . . 126
1. Classical Conditioning of Fear Stimuli 127 C. Electrolytic and Ibotenic Acid Lesions in the PVN 129 D. Injection of 6-0HDA into the PVN. . . . . 132 E. Injection of Sotalol into the PVN . . . . 134 F. Hypothetical Organization of Brain Pathways Mediating
Stress-Induced Renin and Corticosterone Secretion . . 139
VI. LITERATURE CITED
vii

LIST OF TABLES
TABLE 1 Comparison of Mean Values: Normal PRA Pool versus Experimental Sham- or Vehicle-Treated Groups . . . . . . 122
TABLE 2 Newman-Keuls' Multiple Range Test for PRA ..... 123
TABLE 3 Comparison of Mean Values: Normal Corticosterone Pool versus Experimental Sham- or Vehicle-Treated Groups ... 124
TABLE 4 Newman-Keuls' Multiple Range Test for Corticosterone . . ....... 125
viii

LIST OF FIGURES
FIGURE 1 Coronal section of rat brain with an intact paraventricular nucleus ....... . . .. 94
FIGURE 2 Coronal section of rat brain with an electrolytic lesion in the paraventricular nucleus. . .. 94
FIGURE 3 The effect of stress on plasma renin activity in rats with electrolytic PVN lesions . . . . . . . . . .95
FIGURE 4 The effect of stress on plasma renin concentration in rats with electrolytic PVN lesions. . . . . . . .96
FIGURE 5 The effect of stress on plasma corticosterone levels in rats with electrolytic PVN lesions. . . . . . . .97
FIGURE 6 Coronal section of rat brain with an electrolytic lesion in the thalamic nucleus reuniens. . . . . . . .. 99
FIGURE 7 The effect of stress on plasma renin activity in rats with electrolytic lesions in the nucleus reuniens . 100
FIGURE 8 The effect of stress on plasma renin concentration in rats with electrolytic lesions in the nucleus reuniens .101
FIGURE 9 The effect of stress on plasma corticosterone levels in rats with electrolytic lesions in the nucleus reuniens. 102
FIGURE 10 Intact paraventricular nucleus .. ..... 104
FIGURE 11 Paraventricular nucleus following bilateral injection of ibotenic acid. . . . . . . . . . . . . . . . . .105
FIGURE 12 The effect of stress on plasma renin activity in rats with ibotenic acid lesions in the PVN. . . . . . .106
FIGURE 13 The effect of stress on plasma renin concentration in rats with ibotenic acid lesions in the PVN ......... 107
FIGURE 14 The effect of stress on plasma corticosterone levels in rats with ibotenic acid lesions in the PVN .108
FIGURE 15A Coronal section from rat injected with vehicle and stained with DBH antiserum. . . . . . . . . . . .111
ix

FIGURE lSB Coronal section from rat injected with 6-0HDA and stained with DBH antiserum, showing loss of DBH immunoreactivity. . . . . . . . . .. 111
FIGURE 16 The effect of stress on plasma renin activity in rats with 6-0HDA lesions . . . . . . . . . . . . . . . . .112
FIGURE 17 The effect of stress on plasma renin concentration in rats with 6-0HDA lesions . . . . . . . . . . . . .113
FIGURE 18 The effect of stress on plasma corticosterone levels in rats with 6-0HDA lesions ................ 114
FIGURE 19 The effect of stress on plasma angiotensinogen levels in rats with 6-0HDA lesions ................ 115
FIGURE 20 Coronal section of the paraventricular nucleus following bilateral injection of sotalol. . . . . . . .. 117
FIGURE 21 The effect of stress on plasma renin activity in rats injected with either vehicle or sotalol, or in non-inj ected rats . . . . . . . . . . . . . . . . . . . . 118
FIGURE 22 The effect of stress on plasma renin concentration in rats injected with either vehicle or sotalol, or in non-injected rats. . . . . . . . . . . . . . . . .119
FIGURE 23 The effect of stress on plasma corticosterone levels in rats injected with either vehicle or sotalol, or in non-injected rats .................. 120
FIGURE 24 Hypothetical brain pathways mediating stress-induced renin and corticosterone secretion ............. 146
x

CHAPTER I
INTRODUCTION
Stress has profound effects on neuroendocrine function,
increasing plasma corticosterone and prolactin levels, and plasma renin
activity (PRA). Neurons in the hypothalamic paraventricular nucleus (PVN)
play an important role as integrators of endocrine, autonomic and
behavioral functions. The purpose of this dissertation research project
was to investigate the role of neurons in, and afferents to, the PVN in
mediating stress-induced neuroendocrine responses, specifically increased
PRA, plasma renin concentration (PRC), and corticosterone levels. Male
Sprague-Dawley rats were used for these experiments. The stressor
consisted of a conditioned emotional (fear) response paradigm, which has
been shown to cause increases in renin and corticosterone secretion. In
this paradigm, each rat receives a footshock 10 minutes following
placement in a chamber, and then was returned to his home cage. This
procedure is repeated for three consecutive days. On the fourth day,
instead of receiving shock 10 minutes after being placed in the test
chamber, the rat is removed and sacrificed by decapitation. Trunk blood
was collected for hormone assays. The experimental control rats were
treated identically to rats in the treatment group with the exception that
shock was not administered at any time.
1

2
1. Effect of Electrolytic PVN Lesions on Stress-Induced Renin and
Corticosterone Secretion. Bilateral electrolytic lesions in the PVN
prevented the stress-induced increase in PRA, PRC and corticosterone
levels. In contrast, electrolytic lesions in the thalamic nucleus
reuniens, dorsal and caudal to the PVN, did not prevent the stress-induced
increase in either PRA, PRC or corticosterone levels. Since the majority
of the diencephalic corticotropin releasing factor (CRF) neurons are
located in the parvocellular subnuclei of the PVN, it is not surprising
that electrolytic PVN lesions prevented the stress-induced increase in
corticosterone secretion. However, the finding that the stress-induced
increases in PRA and PRC were also prevented was unexpected.
2. Effect of Ibotenic Acid Lesions of Cells in the PVN on Stress
Induced Renin and Corticosterone Secretion. Ibotenic acid is a selective
neurotoxin that destroys cell bodies while leaving fibers of passage
intact. To determine whether cell bodies in the PVN or fibers of passage
mediate stress-induced increases in PRA, PRC and corticosterone levels,
the cells in the PVN were selectively destroyed by injection of ibotenic
acid. The corticosterone, PRA and PRC responses to stress were blocked
by ibotenic acid injection. These observations suggest that neuronal cell
bodies in the PVN mediate the stress-induced increase in PRA, PRC and
corticosterone.
3. Effect of 6-Hydroxydopamine-Induced Lesions in the PVN on
Stress-Induced Renin and Corticosterone Secretion. Since the data
indicated that cell bodies in the PVN mediate stress-induced renin and

3
corticosterone secretion, and the PVN is known to have an extensive
catecholaminergic innervation, the next approach was to determine if
catecholaminergic innervation of PVN neurons plays an important role in
the neuroendocrine response to stress. This was accomplished by injection
of the neurotoxin 6-hydroxydopamine (6-0HDA) into the PVN. 6-0HDA is
taken up by the catecholaminergic nerve endings resulting in degeneration
of these nerve terminals. Lesion placement and damage to noradrenergic
nerve terminals was verified by immunocytochemical methods, using an
antibody against dopamine E-hydroxylase. The 6-0HDA injections prevented
the stress-induced increase in PRA, PRC and corticosterone levels,
suggesting that intact catecholaminergic afferents to the PVN are critical
for the stress-induced increase in renin and corticosterone secretion.
4. Effect of Microinj ection of the E-Adrenoceptor Antagonist
Sotalol into the PVN on Stress-Induced Renin and Corticosterone Secretion.
The results from the experiment with 6-0HDA suggest that catecholaminergic
nerve terminals in the PVN are involved in mediating the effect of stress
on renin and corticosterone secretion. In previous studies, the H.-
adrenoceptor antagonist propranolol attenuated the stress- induced increase
in PRA. Since propranolol is known to cross the blood-brain barrier, it
is possible that &-receptors, located in the PVN, mediate the effect of
stress on renin and corticosterone secretion. To test this hypothesis,
the H.-adrenoceptor antagonist sotalol was microinj ected into the PVN.
Sotalol prevented the stress-induced increase in corticosterone levels,
but did not attenuate the stress-induced increase in PRA or PRC. The
results suggest that a catecholaminergic input to cells in the PVN mediate

4
the effect of stress on renin and corticosterone secretion. Furthermore,
stress-induced increases in corticosterone levels are mediated via &
receptors, whereas stress-induced increases in PRA and PRC are mediated
by different receptors.

CHAPTER II
LITERATURE REVIEW
A. RENIN
1. Introduction
The renin-angiotensin system plays a role in the maintenance of
sodium balance (Parfrey et al., 1981) and the regulation of blood pressure
and volume (Fujii and Vatner, 1985). Renin is the enzyme responsible for
the conversion of angiotensinogen to angiotensin I (AI), a decapeptide.
AI is further converted to the octapeptide angiotensin II (All) by a
peptidyl dipeptidase, converting enzyme. This reaction is slow in vitro,
but rapid in vivo due to the activity of tissue-bound converting enzyme
present on the luminal aspect of vascular endothelial cells throughout the
body (Caldwell et al., 1976). Renin is the rate-limiting enzyme in the
synthesis of All. All is the physiologically active component of the
system and a potent pressor agent. All causes vasoconstriction by a
direct action on vascular smooth muscle (Fujii and Vatner, 1985) and
indirectly via the sympathetic nervous system (Peach et al., 1971).
2. Description of Renin
Renin is a glycoprotein with a molecular weight of about 40,000
(Galen et al., 1979). Specifically, it is an aspartyl proteinase (Dzau
and Pratt, 1986) that cleaves a leucine-leucine peptide bond between
5

6
residues 10 and 11 of angiotensinogen. Renin has a circulation half
life of 4-15 minutes (Michelakis and Mizukoshia, 1971; Gordon and Sull
ivan, 1969; Gutman et al., 1973; Oates et al., 1974; Oparil et al., 1970;
De Vito et al., 1977) and is metabolized by the liver and kidneys.
Inactive forms of renin (prorenin and activation intermediates) have been
found in the plasma of humans (Leckie, 1981), dogs (Wilczynski and Osmond,
1983) and rats (Osmond et al., 1982). Prorenin is a circulating, high
molecular weight form of renin with no intrinsic enzymatic activity
(Sealey et al., 1979; lnagami and Murakami, 1980). Prorenin release
following bilateral nephrectomy was reported to be under F..-receptor
regulation (Wilczynski and Osmond, 1986). Nielsen and Poulsen (1988)
reported that the kidney is the main source of inactive renin.
3. Physiological Aspects of the Angiotensins
a. Angiotensinogen (renin substrate)
The glycoprotein angiotensinogen (approximate molecular weight
60,000) is the substrate for renin. The main source of angiotensinogen
is the liver. lmmunocytochemical studies (Richoux et al., 1983) and
Northern Blot analysis with complementary mRNA sequences for angioten
sinogen (lngelfinger et al., 1986) have shown that the liver contains and
synthesizes angiotensinogen. Angiotensinogen is widely distributed in the
blood and other extracellular fluids (Reid and Ramsay, 1975; Horky et al.,
1971; Keeton and Campbell, 1980).
Plasma levels of angiotensinogen can be decreased by adrenalectomy
(Carretero and Gross, 1967), hypophysectomy (Goodwin et al., 1970) and

7
diseases that impair liver function (Ayers, 1967). The administration of
glucocorticoids, mineralocorticoids and sodium replacement can prevent the
decrease in angiotensinogen levels produced by adrenalectomy (Nasjletti
and Masson, 1969). Nephrectomy increases plasma levels of angiotensinogen
(Hiwada et al., 1976; Carretero and Gross, 1967; Bing and Poulsen, 1969).
This increase is thought to be mediated by adrenocortical steroids.
Eggena and Barrett (1988) demonstrated an additive effect of nephrectomy
and dexamethasone on the stimulation of renin substrate secretion.
Freeman and Rostorfer (1972) observed that adrenalectomy caused a 70%
reduction of the increase in plasma angiotensinogen caused by nephrectomy.
Surgical stress, which stimulates the release of adrenocortical steroids,
also increases plasma levels of angiotensinogen for several days in
rabbits (Campbell et al., 1973; Romero et al., 1970; Morris et al., 1977).
These studies suggest that an intact pituitary-adrenal axis is necessary
for adequate substrate production.
b. Angiotensin I (AI)
The amino terminal decapeptide AI is cleaved from angiotensinogen
by renin in the blood. Angiotensin I has been reported to act in a
similar manner to All with respect to the adrenal medulla (stimulating
the release of catecholamines) and the central nervous system (inducing
thirst), but with lower potency (Fitzsimons, 1971; Peach, 1974).
c. Angiotensin II (AII)
All has many physiological effects, all of which are involved in
the maintenance of blood pressure and plasma volume. All is a potent

8
pressor agent, acting directly on the vascular smooth muscle to cause
vasoconstriction (Skeggs et al., 1956) and indirectly, via the sympathetic
nervous system (Peach et al., 1971; Scroop et al., 1971). Similarly, All
increases cardiac contractility in vitro by a direct action on the
myocardium and indirectly via norepinephrine release from the cardioac
celerator nerves (Blumberg et al., 1975). All acts on the peripheral
arterioles to maintain arterial pressure in low cardiac output states
(Davis and Freeman, 1976).
All stimulates the secretion of aldosterone by a direct action on
the zona glomerulosa of the adrenal cortex (Bartter et al., 1961; Biron
et al., 1961). Blockade of All receptors decreases aldosterone production
to nondetectable levels (Davis and Freeman, 1976). All also stimulates
vasopressin secretion when administered intraventricularly in rats and
dogs (Share, 1979; Keil et al., 1975) or intravenously in conscious dogs
(Reid et al., 1982; Bonjour and Malvin, 1970), conscious rats (Knepel and
Meyer, 1980) and humans (Uhlich et al., 1975; Padfield and Morton, 1977).
Aldosterone and vasopressin act at the distal tubules of the kidney to
increase sodium and water reabsorption, respectively. Together, they act
to restore blood pressure and volume in response to the physiological
stress of hypotension, hemorrhage and/or hypovolemia.
All inhibits renin release when it binds to the juxtaglomerular
(JG) cells (Dzau and Pratt, 1986), and increases the concentration of
angiotensinogen in the plasma of dogs (Blair-West et al., 1974; Beaty et
al., 1976) and rats (Nasjletti and Masson, 1973; Khayyall et al., 1973).
All also stimulates secretion of angiotensinogen in vitro from rat
hepatocytes (Klett and Hackenthal, 1987). It is likely that the

9
stimulation of angiotensinogen by All occurs via enhanced glucocorticoid
secretion. All stimulates glucocorticoid secretion by a direct action on
the adrenal cortex (Kaplan, 1965) and indirectly by potentiating the
effect of CRF on ACTH secretion (Keller-Wood et al., 1986; Maran and
Yates, 1977; Spinedi and Negro-Vilar, 1983). Reid et al. (1978) suggested
that All stimulates angiotensinogen production to provide a positive
feedback mechanism to prevent lack of substrate supply during periods of
elevated renin secretion.
4. Regulation of Renin Secretion
a. Introduction
Renin secretion from the kidney is regulated by a number of
neural, chemical and mechanical factors. In general, factors that cause
a decrease in either renal perfusion pressure, blood volume or plasma
sodium concentration will stimulate renin secretion.
Renin is secreted by the granular juxtaglomerular cells located in
the afferent arterioles of the kidney (Taugner et al., 1979; Taugner et
al., 1984). The juxtaglomerular cells are in close proximity to the
macula densa, a specialized portion of the distal tubule that is sensitive
to changes in sodium reabsorption across the tubular epithelium. The
macula densa monitors the ionic environment in the tubular fluid and
relays the message to either increase or decrease renin secretion.
Studies by ltoh et al. (1985) suggest that adenosine, via the activation
of A1 receptors, may be a signal from the macula densa to the juxta
glomerular cells to inhibit renin release. It is reported that a decrease
in the amount of sodium (Churchill et al., 1978), potassium (Shade et al.,

10
1972) or chloride (Rostand et al., 1985) that crosses the macula densa
cells will stimulate renin secretion.
Renin secretion is also stimulated by a decrease in renal arterial
pressure. The idea that a "pressor substance" (later to be known as
renin) was secreted in response to a decrease in renal blood flow was
first postulated by Goldblatt et al. (1934). This idea was further
developed by Tobian et al. (1959) who suggested the existence of a renal
baroreceptor that was responsible for the inverse relationship between
blood pressure and renin secretion.
Studies by Fahri et al. (1982) have shown that stimulation of renin
secretion following a decrease in renal arterial pressure occurs via an
intrarenal baroreceptor interacting with a renal B-receptor. Their
studies have demonstrated that elevating plasma epinephrine concentration
without changing blood pressure does not increase plasma renin activity.
However, if epinephrine secretion accompanies a stimulus (e.g., hemor
rhage) which also lowers blood pressure, then renin secretion will be
increased. This system allows for plasma renin activity to be either
stimulated or unaffected by high physiological levels of circulating
epinephrine, depending on whether or not a hypotensive threat is involved.
b. Role of the Sympathetic Nervous System in Renin Secretion
The sympathetic innervation of the rat kidney consists of
sympathetic cell bodies in paravertebral (thoracic T6 through lumbar L4)
and prevertebral (renal, greater splanchnic, and celiac) ganglia
(Sripairoj thikoon and Wyss, 1987). Stimulation of the renal nerves

increases renin secretion (Di Bona, 1985; Blair et al., 1985).
11
This
stimulatory effect is mediated, at least in part, via norepinephrine
released from the endings of the postganglionic sympathetic neurons that
innervate the kidneys (Johnson et al., 1971; Taber et al., 1976).
Circulating epinephrine can also stimulate renin secretion. The effect
of stimulation of the renal nerves can be enhanced by activating beta (,8)
receptors (Vander, 1965). Catecholamines are believed to stimulate renin
secretion via a ,B-adrenergic mechanism since /3-antagonists have been shown
to inhibit their effect both in vivo (Loeffler et al., 1972; Pettinger and
Keeton, 1975) and in vitro (Weinberger et al., 1975; Vandongen et al.,
1973). The renin stimulating potency profile is isoproterenol >
epinephrine > norepinephrine according to studies in conscious dogs by
Johnson et al. (1979). Studies by Healy et al. (1985) and Osborn et al.
(1981) suggest that the sympathetic nervous system stimulates renin
release by activating B1 receptors on the juxtaglomerular cells.
c. Role of Extra-Renal Beta Receptors in Renin Secretion
Although there is evidence supporting intrarenal /3 receptor
regulation of renin secretion, this does not exclude a role of extrarenal
/3-adrenergic receptors that provide a powerful stimulatory effect on renin
secretion. For example, Reid et al. (1972) observed increases in plasma
renin activity and renin secretion following isoproterenol infusion into
the femoral artery of dogs. Renal perfusion pressure was kept constant
with an aortic clamp. However, infusion of the same doses of isoproter
enol into the renal artery had no effect on plasma renin activity or renin
secretion, suggesting that an extrarenal ,B receptor mechanism is involved

12
in regulating renin secretion. Johnson et al. (1979) obtained similar
results with infusion of epinephrine and norepinephrine into the inferior
vena cava (IVG) and renal artery. Epinephrine infusion into the IVG
resulted in a 3. 5-fold increase in plasma renin activity while IVG
infusion of norepinephrine resulted in a 1.5-fold increase in plasma renin
activity. Infusion of epinephrine or norepinephrine directly into the
renal artery to achieve similar arterial concentrations resulted in no
increases in plasma renin activity. Fahri et al. (1982) suggested that
the effect of epinephrine in the intact conscious dog can be explained as
the combination of an extrarenal and intrarenal action. The extrarenal
event is a generalized vasodilation that results in a drop in blood
pressure which is sensed by the renal baroreceptor. The intrarenal action
is mediated through intrarenal /3 receptors which affect the stimulus-
response curve of the renal baroreceptor. At the present time the
location of the extrarenal /3 receptor has not been established.
d. Role of Brain Serotonin (5-HT) Neurons in Renin Secretion
In 1980, Zimmermann and Ganong demonstrated that injection of the
5-HT precursors 5-hydroxytryptophan (5-HTP) or tryptophan to anesthetized
dogs increased plasma renin activity. In humans, the 5-HT precursor L
tryptophan was reported to increase renin secretion (Modlinger et al.,
1979), and the 5-HT antagonist cyproheptadine inhibited the secretion of
renin induced by furosemide (Epstein and Hamilton, 1977). In rats, the
5-HT releasers p-chloroamphetamine (PGA) and fenfluramine dose-dependently
increased plasma renin activity via release of 5-HT from serotonergic
nerve terminals (Van de Kar et al., 1981; 1985). The 5-HT2 antagonist

13
LY53857 dose-dependently inhibited the increase in plasma renin activity
and plasma renin concentration following administration of fenfluramine
and MK-212, a 5-HT agonist, suggesting that stimulation of 5-HT2 receptors
enhances renin release (Lorens and Van de Kar, 1987).
The effect of PCA on plasma renin activity was prevented by either
pretreatment with the 5-HT synthesis inhibitor p-chlorophenylalanine
(PCPA) or by a chemical lesion of the dorsal raphe nucleus with the 5-HT
neurotoxin, 5, 7-dihydroxytryptamine (Van de Kar et al., 1982). A
mechanical lesion in the mediobasal hypothalamus, as well as postero
lateral deafferentation of the hypothalamus (separating the hypothalamus
from the midbrain) also blocked the effect of PCA on plasma renin activity
(Karteszi et al., 1982). Anterolateral section through the retrochias
matic area was ineffective in blocking the effect of PCA on plasma renin
activity (Karteszi et al., 1982). These results suggest that a serotoner
gic pathway, with cells in the dorsal raphe nucleus in the midbrain and
terminals in the mediobasal hypothalamus, stimulates renin secretion.
1. Role of Beta Receptors in the Serotonergic
Stimulation of Renin Secretion
There is evidence for f3 receptor involvement in mediating the
serotonergic stimulation of renin secretion. Van de Kar and Richardson
Morton (1986) demonstrated that the B1 and B2 antagonist sotalol and the
B1 antagonist atenolol prevented the increase in plasma renin activity
after the injection of PCA and fenfluramine. These findings supported
those of Alper and Ganong (1984) who demonstrated that pretreatment with
the sotalol and propranolol prevented the increase in plasma renin

14
activity produced by PCA. These studies suggested a possible role for
the sympathetic nervous system in mediating the increase in renin
secretion following the administration of 5-HT releasers.
However, in rats which were adrenal medullectomized and sympathec
tomized by chronic injections of 6-0HDA, there was no inhibition of the
PCA- and fenfluramine-induced increase in plasma renin activity. Renal
norepinephrine content was reduced to undetectable levels, suggesting that
it is not likely that the serotonergic stimulation of renin secretion is
mediated via the sympathetic nervous system or adrenal catecholamines (Van
de Kar and Richardson Morton, 1986). These results seem to be in
contradiction with the fact that the ~-adrenergic receptor antagonists
sotalol and atenolol prevented the PCA- or fenfluramine-induced increase
in plasma renin activity. However studies by Johnson (1984) and Johnson
et al. (1979), as described earlier, provide evidence that~ receptors
that mediate the increase in plasma renin activity are located outside the
kidney, suggesting an entirely different mechanism of stimulating renin
secretion. In addition, many beta receptor antagonists are known to bind
to 5-HT receptors (Middlemiss et al., 1977) and it is possible that
sotalol and atenolol are present in sufficient concentrations in the brain
to block the 5-HT receptors and inhibit the serotonin-mediated increase
in plasma renin activity (Lemmer et al., 1985; Garvey and Ram, 1975). To
summarize, the data suggest that brain 5-HT neurons regulate renin
secretion via a mechanism that is not related to the autonomic sympathetic
nervous system or adrenal catecholamines.

e. Role of the Hypothalamic Paraventricular Nucleus
in Renin Secretion
15
The PVN plays a role in neurogenic and genetic models of hyperten
sion (Ciriello et al., 1984; Zhang and Ciriello, 1985). The PVN is a
logical central site to influence renin secretion as it receives afferent
information from the baroreceptor region as well as having efferent
projections to the sympathetic areas of the spinal cord (Sawchenko and
Swanson, 1982).
Electrolytic lesions in the PVN have been shown to prevent the
increase in plasma renin activity produced by the 5-HT releasing drug PCA
as well as preventing the increase in plasma renin activity that follows
immobilization stress (Gotoh et al., 1987). Previous studies (Gotoh et
al., 1985) demonstrated that bilateral knife cuts behind the hypothalamic
paraventricular nucleus (PVN) reduced the renin response to immobilization
and the gravitational stress of head-up tilting, suggesting a role for the
PVN in both stress-induced and 5-HT- stimulated renin secretion.
Electrical stimulation of the PVN in conscious rats increased
plasma renin activity in a frequency-related manner. There were no
significant changes reported in heart rate, mean arterial pressure or
renal vascular resistance at any of the frequencies that produced an
increase in plasma renin activity (Porter, 1986). These results suggest
that neurons in the PVN stimulate renin secretion.
Additional studies by Porter (1988) demonstrated the relationship
between renal perfusion pressure and plasma renin activity during
continuous low-level stimulation of the PVN. He found that for any given

16
decrease in renal perfusion pressure, the plasma renin activity increase
was greater during the ongoing PVN stimulation. The effect of stimula-
tion of the PVN was prevented by both renal denervation and pretreatment
with propranolol. These studies suggest that stimulation of the PVN
elicits increased responsiveness of the kidneys to decreases in renal
perfusion pressure, and that this effect is mediated via the sympathetic
nervous system.
f. Other Hypothalamic Areas Involved in Renin Secretion
The lateral and posterior hypothalamic areas have long been known
as sites where pressor responses can be elicited with electrical
stimulation (Brody et al., 1986; Ciriello and Calaresu, 1977). Therefore
it is not surprising that increases in plasma renin activity are also
observed following electrical stimulation of the lateral (Zanchetti and
Stella, 1975) and posterior (Natcheff et al., 1977) hypothalamus. Unlike
the studies by Porter (1986) involving the PVN, these increases in plasma
renin activity were accompanied by increases in blood pressure and
required intact renal nerves. It is likely that the lateral and posterior
hypothalamus exert their cardiovascular effects through long descending
neuronal projections to the NTS (Kuypers et al., 1973) or ventrolateral
medulla (Brody et al., 1986).
Gotoh et al. (1987) reported that bilateral electrolytic lesions
of the ventromedial nucleus prevented the increase in plasma renin
activity produced by each of the following stimuli: the 5-HT-releaser PCA,
immobilization, head-up tilt under inactin anesthesia, and a low-sodium
diet. The authors hypothesize that the stimuli that increase plasma renin

17
activity via the ventromedial nucleus act by increasing sympathetic
activity since the P-adrenergic receptor antagonist propranolol blocks the
renin response to PCA, immobilization and head-up tilt. The PCA response
is also abolished by another P-adrenergic receptor antagonist, sotalol,
and by ganglionic blockade (Alper and Ganong, 1984). In addition,
stimulation of the ventromedial nucleus increases blood pressure (Bunag
and Inoue, 1985), while bilateral lesions of this nucleus prevent the
development of experimental hypertension (Brody and Johnson, 1980).
g. Extrahypothalamic Pathways Involved in Renin Secretion
Other central nervous system sites play a role in renin release.
Electrical stimulation of the ventrolateral medulla (Richardson et al.,
1974), dorsal medulla (Passo et al., 1971) and mesencephalic dorsal
periaqueductal grey (Ueda et al., 1967) produced increases in plasma renin
activity that were mediated via the renal nerves.
h. Renin and Angiotensin in the Brain
Administration of renin or All into the cerebral ventricles
stimulates vasopressin secretion (Keil et al., 1975), increases drinking
behavior and increases blood pressure (Reid and Ramsay, 1975). The
anterior forebrain and nuclei of the anteroventral third cerebral
ventricle (AV3V) play a role in the All-induced stimulation of drinking
behavior, vasopressin release and increased arterial pressure. These
brain areas are richly innervated by catecholaminergic fibers arising from
cell bodies in the A2 area of the nucleus tractus solitarius, and other
brainstem regions involved with central cardiovascular regulation (Saper

18
et al., 1983; Palkovits et al., 1974; Meldrum et al., 1984). Bellin et
al. (1987) have shown that rats with 6-0HDA injections into the lateral
cerebral ventricle had significantly attenuated drinking and blood
pressure responses following both central and systemic All administration.
Rats pretreated with desmethylimipramine (DMI) prior to injection of 6-
0HDA (to protect noradrenergic terminals) had no thirst or pressor
response deficits following All administration. These results suggest a
role for norepinephrine in mediating All-induced drinking and blood
pressure responses.
The distributions of angiotensin I and II, angiotensinogen,
angiotensin-converting enzyme (ACE) and renin in the brain have been
studied by various biochemical and irnrnunocytochemical techniques
(Brownfield et al., 1982; Changaris et al., 1977; Fuxe et al., 1976; Fuxe
et al., 1980; Basso et al., 1982; Fischer-Ferraro et al., 1971), although
the physiological role for these substances is not clear. Ganong et al.
(1979) suggested that All-like irnrnunoreactive material in the brain may
be involved in modulating the release and action of brain catecholamines.
The presence of a brain renin-angiotensin system is highly
debatable. Fuxe et al. (1980) demonstrated renin-like irnrnunoreactivity
in the paraventricular, periventricular and supraoptic nuclei in the rat
and mouse. Dabsys et al. (1988) demonstrated an increase in angiotensin
converting enzyme activity levels in the amygdaloid complex following
bilateral destruction of the ventral noradrenergic pathway with 6-0HDA.
There was a concomitant increase in arterial blood pressure that
significantly correlated with the ACE activity levels in the amygdala.
Studies in nephrectomized animals revealed All-like irnrnunoreactivity in

19
brain tissue (Phillips and Stenstrom, 1983) and significant elevations in
angiotensinogen content in the hypothalamus and midbrain (Gregory et al.,
1982). However, the All-like material from rat brain has been shown to
be different from authentic All by both gel filtration (Meyer et al.,
1982) and anion exchange HPLC techniques (Pohl et al., 1988). These
studies indicated that the All-like material has a molecular weight of
5000 7000 (compared to 1046 for All) and is more polar but less
positively charged than All.
Although indirect studies suggest the presence of each of the
components of the renin-angiotensin system in the brain (Printz et al.,
1982) there is no evidence to support a functional brain renin-angiotensin
system. Brownfield et al. (1982) have demonstrated that although
angiotensin and converting enzyme immunoreactivities are present in the
brain, they are not co-distributed, suggesting that there is no brain
pathway for the formation of angiotensin.
i. Role of Brain Catecholamines in Renin Secretion
Norepinephrine in the brain was proposed to inhibit secretion of
renin from the kidney through changes in sympathetic activity and
secretion of vasopressin (Blair et al. , 1977; Reid et al. , 1978). The
antihypertensive drug clonidine is an alpha (a) 2-agonist with a biphasic
effect on blood pressure: a transient increase followed by a prolonged
decrease. lntracerebroventricular injection of clonidine lowers plasma
renin activity at a dose which is ineffective when given intravenously
(Reid et al., 1975). Blair et al. (1977) have demonstrated that
administration of L-DOPA to dogs pretreated with the peripheral decar-

20
boxylase inhibitor carbidopa resulted in a drop in blood pressure and an
inhibition of renin secretion. Since this treatment also increases brain
catecholamine content, this study suggests that catecholamines formed from
L-DOPA can act within the central nervous system to cause a decrease in
renin secretion. In this study the decrease in plasma renin activity was
dependent upon the presence of the renal nerves.
The 5-HT releaser fenfluramine has a biphasic effect on plasma
renin activity: an initial increase followed by a sustained decrease.
Studies by Van de Kar et al. (1985) suggested that the initial increase
in plasma renin activity following fenfluramine administration is mediated
via release of serotonin, while the delayed, long-lasting suppression of
plasma renin activity is mediated via a catecholaminergic mechanism.
However, the long-term suppression of plasma renin activity following
fenfluramine administration is not dependent on the peripheral sympathetic
nervous system, as it was not prevented by adrenal enucleation combined
with 6-0HDA-induced sympathectomy (Van de Kar et al., 1988). To
determine whether a 2 -adrenoceptors mediate the suppressive effects of
fenfluramine, the a 2-antagonists yohimbine and rauwolscine were injected
prior to fenfluramine administration. Neither a 2-antagonist prevented the
long-term suppressive effect of fenfluramine on plasma renin activity or
plasma renin concentration. The data suggest that fenfluramine does not
inhibit renin secretion by activating a 2-adrenoceptors in the CNS. This
is in contrast to clonidine and a-methyl DOPA, which inhibit sympathetic
outflow to the kidneys through activation of central a 2-adrenoceptors.
Privitera et al. (1979) have shown that low doses of intracister
nally-injected propranolol produced dose-dependent decreases in plasma

21
renin activity and mean arterial pressure (MAP), whereas identical doses
given intravenously had no significant effect on plasma renin activity or
MAP. A concomitant reduction in circulating norepinephrine levels was
also seen. This study suggests that propranolol could be acting at a
central ~-adrenergic site to suppress renin release.
It is clear that central catecholamines play a role in mediating
the suppression of plasma renin activity, but the site(s) of action and
type of receptors involved have yet to be identified.
B. CORTICOSTERONE
1. Introduction
In 1856 Brown-Sequard discovered that the adrenal glands are essential
to life. By the 1930's it became clear that the adrenal cortex is the
source of two vital classes of hormones: one, the glucocorticoids,
regulate carbohydrate and protein metabolism, and the ability to tolerate
prolonged stress (Selye, 1946; Selye, 1943; Cori and Cori, 1927; Britton
and Silvette, 1931; Long et al., 1940). The second class is the
mineralocorticoids, which control water and electolyte metabolism and the
ability of the kidney to reabsorb sodium (Loeb et al., 1933; Harrop et
al., 1933). Aldosterone is the primary salt-retaining hormone of the
adrenal cortex and is synthesized in the glomerulosa layer of cortical
cells. Cortisol (in humans) and corticosterone (in rats) are the main
glucocorticoids secreted by the adrenal gland and are synthesized in the
fasciculata layer. The fasiculata layer of cells in the adrenal cortex
is stimulated by adrenocorticotropin hormone (ACTH) from the anterior lobe
of the pituitary gland (Li et al., 1943; Sayers et al., 1943). Ingle et

22
al., (1938) found that ACTH secretion from the anterior pituitary is
inhibited by secretions of the adrenal cortex. The role of the hypothal
amus in integrating stimulatory and inhibitory inputs to the adenohypo
physis was studied by Harris (1948) and Rasmussen (1938). Characteriza
tion of the hypothalamic factor that stimulates corticotropin secretion
(CRF) was completed by Vale et al. (1981).
2. Corticotropin-Releasing Factor (CRF)
CRF from ovine hypothalamic tissue is a 41-amino acid peptide that
releases ACTH and ~-endorphin from anterior pituitary cells (Vale et al.,
1981; Plotsky, 1985; Rivier et al., 1983; Rivier and Vale, 1983). This
peptide is 5-10 times more potent in releasing ACTH and ~-endorphin than
vasopressin, which was originally thought to act as a corticotropin
releasing factor (Saffran and Schally, 1977). Swanson et al. (1983)
described the localization of CRF immunoreactive cells and fibers in three
distinct systems in the rat brain. The first group is in the hypothalamic
paraventricular nucleus (PVN). Approximately 2,000 CRF-stained cells are
found in each of the eight subdivisions of the PVN, although most (80%)
of the cells are concentrated in the parvocellular division (especially
the periventricular and medial parts), and about 15% are found in the
magnocellular division where oxytocinergic cells predominate. These CRF
containing neurons are the only ones that demonstrated increased staining
intensity following adrenalectomy. The second group of CRF-containing
neurons are involved in mediating autonomic reponses. They include the
central nucleus of the amygdala, bed nucleus of the stria terminalis,
substantia innominata, lateral hypothalamic area, medial and lateral

23
preoptic areas, central periaqueductal grey, laterodorsal tegmental
nucleus, locus coeruleus, dorsal vagal complex, parabrachial nucleus and
regions containing the Al and AS catecholamine cell groups. The third
group of CRF- immunoreactive cells are found in the cerebral cortex,
especially layers II and III of the neocortex, and limbic regions such as
the cingulate gyrus, prefrontal areas and areas bordering the rhinal
fissure (Swanson et al., 1983). CRF-like immunoreactive neurons were also
observed in the cerebellum and spinal cord (Olschowka et al., 1982).
Outside the central nervous system, CRF-like immunoreactivity has been
found in endocrine cells of the pancreas and gastrointestinal system, also
in the liver, pituitary, adrenal, lung and placenta (Petrusz et al.,
1985).
The parvocellular neurons of the PVN contain CRF (Merchenthaler et
al., 1982; Liposits et al., 1983), which is transported along axons to the
median eminence where it is released into the hypophyseal portal vessels.
Makara et al. (1981) have shown that the CRF fibers in the stalk-median
eminence region either originate from or run through the PVN. In the
pituitary gland, CRF is then bound to specific membrane receptors of ACTH
containing cells (Wynn et al., 1984; Leroux and Pelletier, 1984). ACTH
release is stimulated which, in turn, increases steroid hormone production
in the adrenal cortex. The glucocorticoids then exert negative feedback
effects on both the hypothalamus (Yates and Urquhart, 1962) and anterior
pituitary (Russell et al., 1969; Mulder and Smelik, 1977) via glucocor
ticoid receptors (Gustafsson et al., 1983). ACTH immunoreactive fibers
are found in most parts of the parvocellular division of the PVN and in
the oxytocinergic cells of the magnocellular division (Sawchenko et al.,

24
1982; Swanson and Sawchenko, 1983).
Afferent connections to CRF neurons include presynaptic boutons
containing immunoreactive ACTH 1-39 (Liposits et al., 1985) and CRF
(Liposits et al., 1985), suggesting an autoregulatory process in the
control of CRF release. Parvocellular CRF neurons also receive input from
magnocellular neurons in the same nucleus (Leranth et al., 1983) and send
processes to these magnocellular elements (Liposits et al., 1985). CRF
neurons have been shown to have angiotensin II reactivity in adrenalec
tomized, colchicine-treated rats (Lind et al., 1985).
Rivier and Vale (1983) suggested that vasopressin may act
synergisticallywith CRF under certain physiological conditions to promote
ACTH secretion. In addition, in vitro studies have shown that vasopressin
acts directly on the anterior pituitary corticotropes to stimulate ACTH
secretion (Vale et al., 1983). Together, these results suggest that
vasopressin may act synergistically with CRF under certain physiological
conditions to promote ACTH secretion.
Sawchenko (1987) demonstrated that adrenalectomy (ADX) enhanced
both CRF and vasopressin immunoreactivity suggesting that they may be
colocalized in parvocellular neurosecretory neurons of the PVN. Other
studies support this observation (Tramu et al., 1983; Kiss et al., 1984;
Sawchenko et al., 1984; Piekut and Joseph, 1985). Dual immunocytochemical
studies have shown the co-localization of CRF and vasopressin in the
parvocellular neuronal perikarya in the PVN. In general, CRF immunos
tained cells are concentrated in the medial parvocellular part of the PVN,
while the vasopressin neurons are predominantly in the magnocellular part
of the PVN. However, in adrenalectomized, colchicine-treated rats, a

25
dense accumulation of vasopressin-imrnunoreactive cells were seen in the
medial parvocellular part of the PVN, in a similar distribution to that
seen for CRF- containing cell bodies (Piekut and Joseph, 1986). Extra
hypothalamic sites of ADX-enhanced CRF imrnunoreactivity included the
cerebral cortex, arnygdala and the bed nucleus of the stria terminalis.
These extra-hypothalamic sites contained no vasopressin imrnunoreactivity
colocalized with the CRF (Sawchenko, 1987). Hypophysectomy as well as ADX
has been shown to enhance CRF imrnunostaining in PVN cell bodies (Bugnon
et al., 1983; Merchenthaler et al., 1983; Paull and Gibbs, 1983; Swanson
et al., 1983). Sawchenko (1987) reported that hypophysectomy produced
results comparable to ADX-enhanced CRF and vasopressin staining in the
PVN. Low doses of dexamethasone attenuated and high doses prevented the
ADX-induced enhancement of CRF and vasopressin imrnunoreactivity, with the
adrenal steroid potency profile as dexamethasone > corticosterone >
deoxycorticosterone > aldosterone. These results suggest that adrenal
steroids, particularly glucocorticoids, play a primary role as regulators
of peptide expression.
3. Other Factors Affecting ACTH Release
Other neuropeptides that stimulate the release of ACTH include
angiotensin II (All) (Ramsay et al., 1978; Steele et al., 1981) and
oxytocin (Beny and Baertschi, 1980; Pearlmutter et al., 1974). In vitro
dose-response studies by Spinedi and Negro-Vilar (1983) ranked the
corticotropin-releasing activity as follows: CRF > vasopressin > oxytocin
> AII. The stimulatory effects of AII may involve the stimulation of AII
receptors in the anterior pituitary (Hauger et al., 1982; Mukherjee et

26
al., 1982) or the hypothalamic PVN (Saavedra et al., 1986). The density
of the All binding sites in the PVN have been shown to increase following
repeated immobilization stress (Castren and Saavedra, 1988). It is
possible that All receptors in the PVN play a role in modulating stress
induced corticosterone secretion.
4. Role of Brain Catecholamines in Corticosterone Secretion
a. Introduction
In general, the ascending noradrenergic pathways appear to relay
afferent visceral information to the forebrain (Cunningham and Sawchenko,
1988). There has been considerable controversy as to whether catechol
amines play an excitatory or inhibitory role in corticosterone secretion.
Norepinephrine and/or epinephrine have been implicated as stimulators of
the secretion of virtually every anterior pituitary hormone (Sawchenko and
Swanson, 1982). With respect to ACTH secretion, brain catecholamines were
originally postulated to have an inhibitory role (van Loon et al., 1971;
Weiner and Ganong, 1978; Cuello et al., 1973; Scapagnini et al., 1970).
However, in recent years more accurate and reliable anatomical and
pharmacological studies suggest a stimulatory role for catecholamines in
corticosterone secretion (Szafarczyk et al., 1985; Feldman et al., 1988;
Alonso et al., 1986; Guillaume et al., 1987).
b. Anatomical Evidence
The predominantly noradrenergic (Sawchenko and Swanson, 1981)
nucleus of the solitary tract (NTS) sends fiber projections to the
parvocellular PVN, substantia innominata, central nucleus of the amygdala

27
and bed nucleus of the stria terminalis (Ricardo and Koh, 1978), all of
which contain CRF-stained cells (Swanson et al., 1983). Swanson et al.
(1983) suggested that the nucleus of the solitary tract, which receives
direct vagal and glossopharyngeal afferents (Beckstead and Norgren, 1979)
could serve as a major relay point of visceral sensory information to CRF
cell bodies in the basal forebrain. Likewise the parabrachial nucleus,
which receives a large input from the NTS (Norgren, 1978), and the locus
coeruleus also project to many of the sites that contain CRF-immunoreac
tive cell bodies, including the parvocellular PVN, the bed nucleus of the
stria terminalis and the central nucleus of the amygdala. These nuclei
may also be involved in the relay of visceral sensory information to CRF
cell bodies (Swanson et al., 1983).
Another fiber projection is described from the predominantly
noradrenergic cells in the Al region of the ventrolateral medulla (which
receives an input from the NTS) to the magnocellular (vasopressinergic)
and parvocellular PVN. Reciprocal connections have also been described
from the parvocellular PVN to the dorsal vagal complex and spinal cord
(Hosoya and Matsushita, 1979; Swanson and Kuypers, 1980). Electrophysio
logical studies by Kannan and Yamashita (1985) have also demonstrated
reciprocal connections between neurons in the NTS region and the PVN.
Studies by Skofitsch et al. (1985) have demonstrated binding sites for CRF
in sensory areas of the rat hindbrain and spinal cord, specifically in the
posterior part of the NTS, the substantia gelatinosa nervi trigemini and
laminae I and II of the spinal cord. Since these areas are rich in nerve
endings of primary sensory neurons, this observation suggests a possible
role for CRF in mediating peripheral sensory processes.

28
Cunningham and Sawchenko (1988) used an immunofluorescence double
labeling procedure to determine which anterogradely labeled fibers and
terminals in the PVN also displayed dopamine P-hydroxylase immunoreac
tivity, indicating the presence of catecholaminergic neurons. Specifical
ly, projections from the Al (caudal ventrolateral medulla) region were
found to synapse on magnocellular vasopressinergic neurons in the PVN,
while projections from the A2 (medial part of the NTS) region were located
primarily throughout the parvicellular division of the PVN. The
projections were the most dense in the dorsal medial parvicellular region,
which is known to contain a large population of CRF- immunoreactive
neurons. A less-dense projection was found in the magnocellular division
of the PVN. The A6 (locus coeruleus) projections were found almost
exclusively in the parvicellular division of the PVN, specifically in the
periventricular zone, which contains dopamine, somatostatin and thyrotrop-
in-releasing hormone-containing neurons. These findings provide
anatomical evidence that the noradrenergic cells in the medial NTS
innervate CRF-immunoreactive cells in the PVN.
Liposits et al. (1986) showed that PNMT-immunoreactive axon
terminals have synaptic connections with dendrites, somata and spinous
structures of CRF-immunoreactive neurons. The parvocellular subnuclei
demonstrated a more intense adrenergic innervation than the magnocellular
subnuclei. The mammalian anterior pituitary does not receive catechol
aminergic innervation, unlike the neural and intermediate lobes which
receive noradrenergic and dopaminergic innervation (Bjorklund et al.,
1967; Saavedra et al., 1975).

c. Role of the Ventral Noradrenergic Bundle in
Corticosterone Secretion
29
The ventral noradrenergic bundle (VNAB) originates from the
medullary Al and A2 cell groups, and the pontine locus coeruleus. It
extensively innervates the hypothalamus, conveying most of the catechol
aminergic innervation to the PVN (Moore and Bloom, 1979; Palkovits, 1981;
Swanson and Sawchenko, 1980). The VNAB is thought to be the source for
the noradrenergic modulation of ACTH secretion (Szafarczyk et al., 1985;
Feldman et al., 1988). Also, afferents from the epinephrine-containing
cell groups (Cl-C3) reach the hypothalamus via the VNAB (Ungerstedt,
1971).
Injections of 6-0HDA into the VNAB significantly reduced CRF levels
in hypophysial portal blood (HPB) as well as producing significant
depletion of hypothalamic norepinephrine and epinephrine (Guillaume et
al., 1987). Similar results were obtained by Eckland et al. (1988), who
found that 6-0HDA injections into the VNAB or the lateral ventricles
caused a reduction of HPB CRF concentration. However, HPB norepinephrine
was reduced only with the i.c.v. 6-0HDA injections and was unchanged in
the VNAB-lesioned group.
Further support for a stimulatory role of norepinephrine in CRF
secretion is provided by Plotsky (1987). The increase in hypophysial
portal immunoreactive CRF (irCRF) following electrical stimulation of the
VNAB was prevented by i.c.v. pretreatment with the a 1-adrenoceptor
antagonist corynanthine, but not by the ~-adrenergic antagonist propran-
olol. Corynanthine also blocked the dose-dependent increases in irCRF

30
following icv administration of norepinephrine (0.1-5.0 nmol). In
contrast, 5 nmol or greater doses of norepinephrine resulted in a dose
dependent inhibition of irCRF release that could be blocked by propran
olol, but was unaffected by corynanthine. This suggests that low doses
of norepinephrine stimulate CRF release via an a-receptor mechanism,
whereas higher doses of norepinephrine inhibit CRF release via a &
receptor.
d. Stimulation of CRF/Corticosterone Secretion by Epinephrine
and Norepinephrine
Microinjections of norepinephrine (40 nmoles) into the PVN
significantly increased serum corticosterone levels. Epinephrine (40
nrnoles) microinjections into the PVN were even more efficacious than
norepinephrine in increasing serum corticosterone levels, whereas dopamine
(40 nrnoles) had no effect (Leibowitz et al., 1986).
Intravenous infusion of either !-epinephrine or the P-adrenoceptor
agonist 1-isoproterenol into pentobarbital anesthetized rats resulted in
a dose-dependent increase in plasma corticosterone levels. The effect of
epinephrine and isoproterenol on plasma corticosterone levels was
associated with a parallel and dose-related increase in plasma ACTH
(Berkenbosch et al., 1983).
Tilders et al. (1985) found that intravenous infusion of epineph
rine increased plasma ACTH and corticosterone to levels that are
comparable to those induced by stress. The stimulation of ACTH secretion
by epinephrine appears to be mediated via CRF and P-adrenergic receptors
since it was prevented by i.p. injections of propranolol or administration

31
of CRF antiserum.
In vitro studies with incubated rat hypothalami demonstrated that
norepinephrine produced a dose-dependent stimulation of CRF release which
was prevented by the ~-adrenoceptor antagonists propranolol and timolol.
The norepinephrine-induced CRF release was not affected by the a1
-
adrenoceptor antagonists thymoxamine, prazosin or corynanthine or the a2-
adrenoceptor antagonist idazoxan (Tsagarakis et al., 1988). This study
suggests a stimulatory role for norepinephrine on CRF release that is
mediated by~ receptors.
e. Role of Dopamine Neurons in Corticosterone Secretion
The PVN is uniformly innervated with dopaminergic neurons. The
dopaminergic innervation of the PVN originates from cells of the arcuate
nucleus (Al2), zona incerta (A13) and periventricular nucleus (Al4)
(Hokfelt et al., 1984). Immunocytochemical studies indicated that both
dopamine and norepinephrine terminals synapatically contact magnocellular
neurons on their dendrites or cell bodies (Decavel et al., 1987). In
addition, tyrosine hydroxylase immunoreactivity has been reported in both
fibers and neurons of the PVN (Swanson et al., 1981; Liposits et al.,
1986).
Dopamine has been shown to stimulate corticosterone secretion.
Injection (i.p.) of the dopamine agonist pergolide caused a dose-dependent
increase in plasma corticosterone levels, which was blocked by pretreament
with the dopamine antagonists spiperone and haloperidol. Spiperone also
partially inhibited the increase in plasma corticosterone levels following
administration of quipazine, a serotonin agonist, although much higher

32
doses were required than for pergolide. In addition, the peripheral
dopamine antagonist domperidone did not prevent the increase in plasma
corticosterone levels by pergolide (Fuller and Snoddy, 1981). These
results support a role for central dopamine receptors in corticosterone
secretion. Similar results were obtained by Jezova et al (1985), who
found that subcutaneous injection of the dopamine agonist apomorphine
increased plasma ACTH and corticosterone levels. Haloperidol pretreatment
completely inhibited the apomorphine-induced stimulation in plasma ACTH.
5. Role of 5-HT Neurons in Corticosterone Secretion
Many studies support the hypothesis that stimulation of serotonergic
receptors leads to increased corticosterone secretion in unstressed
animals (Buckingham and Hodges, 1979; Fuller and Snoddy, 1980; Fuller,
1981; Jones et al., 1976; Van de Kar et al., 1982). Gibbs and Vale (1983)
reported that administration of the 5-HT uptake inhibitor fluoxetine to
anesthetized rats caused increased release of CRF-41 into hypophyseal
portal blood. Lorens and Van de Kar (1987) demonstrated that plasma
corticosterone levels were increased by the 5-HT agonist MK-212, the 5-
HT1A agonists ipsapirone and 8-hydroxy-2-(di-N-propylamino)tetralin (8-0H-
DPAT) , and the serotonin releaser fenfluramine. The 5-HT2 antagonist
LY53857 did not prevent the effect of MK-212 and fenfluramine on
corticosterone secretion. These data suggest a possible role for 5-HT1A
receptors in the stimulation of corticosterone secretion. This conclusion
was confirmed by Koenig et al. (1987) and Gilbert et al. (1988).
The dorsal and median raphe nuclei in the midbrain are known to be
the major sources of serotonergic pathways to the hypothalamus (Azmitia

33
and Segal, 1978; Parent et al., 1981; Van de Kar et al., 1980). In
particular, the median raphe nuclei are the primary source of 5-HT fibers
to the suprachiasmatic nucleus, anterior hypothalamic area and medial
preoptic area, while the anterolateral hypothalamic area and arcuate
nucleus receive 5-HT inputs from both the dorsal and median raphe nuclei
(Van de Kar and Lorens, 1979).
With respect to the PVN, a relatively light serotonergic innerva
tion has been described (Aghajanian et al., 1973; Steinbusch, 1981).
Immunocytochemical studies by Sawchenko et al. (1983) found that the major
source of ascending serotonergic projections to the PVN were from the
dorsal and median raphe nuclei and the medial lemniscus. A significant
decrease in PVN serotonin was reported following surgical destruction of
the dorsal raphe nucleus (Palkovits et al., 1977).
Immunocytochemical studies by Liposits (1987) demonstrated a
moderate serotonergic innervation to the PVN, with a prominent distribu
tion to the parvocellular subnuclei. Serotonergic axons were found to
overlap with CRF-immunoreactive axons. Further ultrastructural examina
tion revealed that serotonin-containing terminals formed axo-dendritic and
axo-somatic synapses with CRF-immunoreactive neurons. This study suggests
that serotonin can influence CRF-containing neurons via synaptic
transmission.
Midline injections of 5,7-dihydroxytryptamine (5,7-DHT) into the
raphe nuclei produced significant depletions of 5-HT in the hypothalamus,
and prevented the increase in corticosterone following stimulation of
cortical and limbic areas (dorsal hippocampus, basolateral amygdala,
mesencephalic reticular formation and medial septal nucleus). The ether-

34
induced increase in corticosterone levels was not affected by this lesion.
Injection of 5, 7-DHT into the PVN also prevented the adrenocortical
response to dorsal hippocampal and photic stimulation. These results
suggest that 5-HT in the PVN mediates the stimulation of corticosterone
secretion following either peripheral or central stimuli (Feldman et al.,
1987).
In contrast, Van de Kar et al. (1982) demonstrated that the PCA
induced stimulation of corticosterone secretion was not mediated by either
the dorsal or median raphe nucleus. In addition, posterolateral cuts
which interrupted caudal inputs to the hypothalamus did not block the
serotonergic stimulation of corticosterone secretion by the serotonin
releasing drug PCA (Van de Kar et al., 1985).
Serotonin has been shown to play a role in mediating the diurnal
variation in basal plasma corticosterone in rats. Plasma corticosterone
levels peak at the onset of the dark cycle in the rat (Krieger and Hauser,
1978; Wilkinson et al., 1979). Depletion of brain serotonin with p-
chlorophenylalanine (PCPA) raised the morning low and prevented the
evening increase in plasma corticosterone (Scapagnini et al., 1971;
Vernikos-Danellis et al., 1973). Similar results were observed with rats
fed a tryptophan-free diet for 21 days (Vernikos-Danellis et al., 1977).
In humans, plasma cortisol and ACTH levels also follow a diurnal rhythm,
except that the highest levels are found in the morning (0600h), with the
lowest values at night (2200h) (Watabe et al., 1987). This study
demonstrated a similar diurnal pattern for plasma CRF, with peak values
at 0600h and significantly lower levels at 1800h and 2200h. These results
suggest that the diurnal rhythm in CRF secretion plays a role in

35
regulating the diurnal rhythm in cortisol and ACTH secretion.
In conclusion, the studies presented in this section demonstrate the
presence of CRF cell bodies in the PVN, and a role for brain catechol
amines in the stimulation of CRF secretion.
C. STRESS
1. Introduction
a. Definition
Stress has been defined by Hans Selye (1973) as the "non-specific
response of the organism to any demand made upon it" (Selye, 1973). By
this definition, the word "stress" describes a response, but it is often
used interchangeably with the stimulus or "stressor" that elicits the
response (Mills, 1985). The stress response can occur following a wide
variety of stimuli. Stressors can be classified as physiological,
environmental or psychological. Physiological stressors include surgery,
injection of foreign substances such as proteins, diseases, anesthesia,
loss of blood, exercise and trauma. Environmental stressors include
prolonged exposure to cold or heat, noise, radiation, pollutants and
chemicals. Psychological stressors include a threatening predator,
intense competition among members of the same species, prolonged conflict,
learning how to avoid a painful stimulus (such as electric shock),
conditions characterized by novelty, anticipation, fear, unpredictability
and change (Kidman, 1984; Gray, 1971; Mills, 1985).
b. Cannon's Alarm Reaction
Cannon (1914) described the role of the sympathetic nervous system

36
in the "fight or flight" emergency or "alarm reaction". This reaction
permits the body to perform more strenuous muscle activity than normally
possible. Cannon noted that when animals underwent stressful situations
there was a marked increase in the activity of sympathetic nerves. This
stimulation of the autonomic nervous system mobilizes the body's resources
to deal with the stressful situation. The alarm reaction is thought to
be transmitted from the hypothalamus through the reticular formation to
the spinal cord to evoke massive catecholamine release from the sym
pathetic nerve endings (Guyton, 1981).
c. Selye's General Adaptation Syndrome
Selye (1936) was the first to describe a biological stress syndrome
produced by diverse noxious agents. His description subsequently became
known as the "general adaptation syndrome", a three-stage process that
describes the long-term adjustments elicited by prolonged stress. The
first stage is the "alarm reaction", when the body is exposed to diverse
stimuli to which it has not adapted. Following the initial alarm
reaction, a "stage of resistance" occurs in which resistance to the
original stressor (which continues to exist) is increased, but resistance
to other stressors is decreased. If the stressor continues for a long
enough time, the stage of resistance develops into a final "stage of
exhaustion", with a catastrophic decline in resistance to all other types
of stressors. If the stress reaction continues undiminished, death ensues
(Selye, 1976).
In cases of prolonged stress, Selye (1946) described a major role
for glucocorticoids during the alarm reaction. Glucocorticoids increase

37
sugar deposition in the liver and facilitate the conversion of proteins
and fats into sugars. They enhance the responsiveness of the blood
vessels to epinephrine and norepinephrine. This will provide the body
with energy sources to be rapidly utilized in the event of sudden
strenuous activity. Glucocorticoids also lower the body's resistance to
infection and have anti-inflammatory properties, including decreased
numbers of lymphocytes and eosinophils, decreased thymus weight and
inhibition of antibody formation. Stress appears to play a major role as
a suppressor of immune function.
Other physiological changes induced by prolonged stress include a
depression of thyroid activity, inhibition of body growth, and suppression
of sexual and reproductive behavior (Gray, 1971). In simple terms, the
body copes with prolonged stress by closing down any process that detracts
from immediate energy mobilization.
d. Sympathetic and Endocrine Stress Responses
The biological stress response is believed to be mediated by the
stimulation of two systems (Kidman, 1984). One, the pituitary-adrenal
axis, is activated by sensory nerves which relay information regarding
external stressors to the hypothalamus. Hypothalamic neurons contain CRF
and release it into the portal circulation, stimulating the anterior
pituitary gland to produce and release ACTH. In response, the adrenal
cortex produces corticosteroids. The glucocorticoids liberated into the
bloodstream inhibit the synthesis and/or release of ACTH from the
pituitary by a negative feedback mechanism.
The second system is the sympathetic adrenal medullary axis, which

38
plays an important role in the alarm reaction described by Cannon. In
this system, once the sensory nerves relay the information regarding the
external stressors to the hypothalamus, it relays a message through the
reticular formation to the spinal cord to evoke massive catecholamine
release from the sympathetic nerve endings and adrenal medulla.
e. Physiological Changes in Response to Stress
Selye (1950) emphasized the fact that the physiological changes
(primarily in the adrenal cortex) in response to stressful stimuli were
the same irrespective of the stimulus used. This suggests that whether
the stressor is physiological, environmental or psychological, the
biological stress response would always be the same. This general
hypothesis allows for comparison of physiological responses following a
variety of different stressors.
Since stress is abstractly defined, it is helpful to have
quantifiable parameters that are known to change when animals or humans
are exposed to a stressor. In the experiments presented in this
dissertation, the neuroendocrine responses to stress were measured with
respect to plasma renin activity and plasma corticosterone levels. The
stressor was a behavioral paradigm known as conditioned fear, or
conditioned emotional response (CER), designed to produce psychological
stress.
B. The Conditioned Fear Paradigm
a. Description
Each rat was transported from his home cage to the stress chamber.

39
Ten minutes following placement in the chamber, the experimental
(stressed) rat received an inescapable foot shock and was returned to his
home cage. This procedure was repeated for three consecutive days. In
previous experiments it was apparent that the stressed rats had learned
that their placement in the chamber will be followed by a foot shock. By
the third day, in contrast to the control animals, the stressed rats
defecated, urinated and demonstrated freezing (no movement) behavior. On
the fourth day, after 10 minutes in the test chamber, instead of receiving
shock the rat was removed, transported to a third area outside the stress
room and immediately sacrificed by decapitation. The control rats were
treated identically except that shock was not administered at any time.
A more detailed description of this procedure can be found in the Methods
section.
b. Pavlov's Dog: Classical Conditioning
Ivan Pavlov (1927) presented studies on classical conditioning in
which a dog was repeatedly exposed to a situation where an initially
neutral stimulus (e.g., a tone or light flash) was followed directly by
a stimulus of biological significance to the animal (food). The latter
"unconditioned stimulus" (UCS) elicited a range of "unconditioned
responses" (UCR) . As the pairings of stimuli were continued, the
originally neutral stimulus became a "conditioned stimulus" (CS), capable
of eliciting some part of the responses formerly elicited exclusively by
the UCS. This "conditioned response" (CR) in Pavlov's dogs was saliva
tion.

40
c. Definition of Fear
Fear can be defined as an emotional response to an aversive
stimulus. In this respect, it is classified as an emotion. The common
element that emotions share is that they represent a reaction to a
"reinforcing event" or signals of reinforcing events. Reinforcing events
include both rewards and punishments. An example of a punishment would
be the removal of a reward, or the failure of an expected reward to occur.
The emotional responses will depend on the type of reinforcing event and
the animal's knowledge of these events (Gray, 1971).
d. Measures of Fear
Hall (1941) established the link between the study of fearfulness
and the use of defecation as a measure of fear. He developed a standard
test (the Open Field) for the measurement of emotional defecation. Parker
(1939) demonstrated that defecation occured in response to a variety of
situations which one would expect to elicit fear. He recorded the amount
of defecation elicited by six different situations: the Open Field test,
a buzzer, sudden dropping, a tilting box that caused the rat to slide down
an inclined plane, immobilization and forced swimming. High positive
correlations were obtained between the scores on every test. Further
factor analysis indicated a general factor (presumably fear) which all the
tests were measuring. These results support the statement by Selye (1950)
that physiological changes in response to stressful stimuli are the same
irrespective of the alarming stimulus used.

41
e. Emotional Factors
There are physiological events that occur within an animal to
influence its behavior in a given environment. In 1890, William James
and Carl Lange independently expressed a theory for the interrelationship
between physiological events and emotional responses. The James-Lange
theory, as it is known, defines emotions as the perception of the
physiological changes that occurred in response to a stimulus. For
example, the feeling of being frightened would consist solely of feeling
your heart pound, your breathing become faster, and so on. Unfortunately,
it is not possible to isolate specific physiological functions and
correlate them exclusively with a particular emotional state. However,
the role of emotional factors should not be overlooked. Studies by Mason
et al. (1976) recognize the sensitivity of the pituitary-adrenal cortical
system to psychological influences. Departing from Selye's non-specific
physiological response hypothesis (1950), Mason et al. (1976) proposed
that there are similar psychological reactions to various external
stimuli. Factors such as heat, cold, fasting or exercise do not produce
an adrenal cortical response if emotional arousal is carefully avoided
(Mason et al., 1976). To illustrate, on the first day of starvation,
food-deprived monkeys that could watch other monkeys being fed in nearby
cages had an elevation of blood corticosteroids that occurred too early
to be attributed to a physiological response due to a lack of food.
However when the monkeys were carefully adapted to their surroundings,
kept in a soundproof room, and maintained on regular "feeding" (non
caloric, fruit-flavored pellets) schedules, there were none of the usual

42
changes in glucocorticosteroids associated with fasting periods. Mason
concluded that there is a considerable degree of emotional disturbance,
due to discomfort, inherent in the laboratory setting. Kidman (1984)
expanded this idea to a "cognitive theory of stress" which says that, in
spite of our best efforts to control hunger, temperature and other
environmental pressures, we still become frustrated over perceived threats
to our welfare and security. Thus, the stress response consists of a
variety of physiological and psychological factors. These factors are
most likely integrated through the central nervous system, in particular
the hypothalamus, as it coordinates the interaction between the autonomic
nervous system, the endocrine system and many forms of emotional behavior.
3. Cardiovascular Changes in Response to Classical Conditioning
Stressful conditions have been implicated in the etiology of
cardiovascular disease and hypertension (Hollenberg et al., 1981; Manuck
et al., 1981; Schulte et al., 1984). Changes in heart rate and arterial
blood pressure can be classically conditioned (Dykman and Gantt, 1956).
A neutral stimulus that elicits no cardiovascular response (conditioned
stimulus) is paired with another stimulus (unconditioned stimulus) that
evokes a vigorous cardiovascular response (unconditioned response). After
several pairings of the conditioned and unconditioned stimulus, the
previously neutral conditioned stimulus will eventually cause cardiovas
cular changes in anticipation of the unconditioned stimulus (Cohen and
Randall, 1984).
Smith et al. (1984) have demonstrated that the perifornical area of
the hypothalamus (referred to as HAGER, the Hypothalamic Area Controlling

43
Emotional Responses) is responsible for the cardiovascular responses that
accompany emotional behavior in monkeys. Their operational definition of
emotion included cardiovascular responses (elevated arterial blood
pressure) and a behavioral response (suppression of lever pressing). 'When
the hypothalamic area (HAGER) was electrically stimulated, it produced the
same cardiovascular pattern as that associated with the emotional
situation. The cardiovascular but not the behavioral portion of the
emotional response was lost following ablation of the HAGER. Afferents
to the HAGER include the amygdala and septal area as well as brain stem
autonomic areas such as the central grey, nucleus tractus solitarius, and
dorsal motor nucleus of the vagus. Efferents include a direct projection
to the intermediolateral cell column cells of the thoracic cord, an
important site with respect to sympathetic control of cardiovascular
function. Other efferent projection sites include the raphe nuclei, the
parabrachial complex, the hypothalamic PVN and the periaqueductal grey.
Cardiovascular changes have been observed with the Sidman avoidance
behavioral testing (Galosy et al., 1982). The Sidman avoidance paradigm
(Sidman, 1953) is used frequently to evaluate cardiovascular changes
associated with the postponement of electric shock contingent upon an
appropriate reponse. The paradigm involves delivery of electric shock at
regular intervals unless a response (such as lever pressing) occurs to
delay the shock. If the animal presses the lever at least once during the
interval, it can avoid all shocks (Turkkan et al., 1982).
Anderson and Brady (1971, 1972) observed decreased heart rate and
increased pulse and arterial blood pressure during the anticipatory pre
avoidance period. Once the avoidance test was underway, the heart rate

44
rapidly increased above pre-avoidance levels while pulse and arterial
blood pressure remained elevated. Further pharmacological studies
indicated that the ~-adrenoceptor blocker propranolol partially attenuated
the heart rate elevations during avoidance, but did not affect the
increases in blood pressure in either avoidance or pre-avoidance periods
(Anderson and Brady, 1976). In similar studies, the a-antagonist
phenoxybenzamine blocked the elevation in blood pressure during the
avoidance period but did not affect pre-avoidance changes in heart rate
and arterial blood pressure (Anderson et al., 1976). From these and other
studies, Galosy et al. (1982) suggested that reduced heart rate during the
pre-avoidance period is dominated by parasympathetic mechanisms, whereas
the arterial blood pressure levels may be mediated by peripheral vascular
mechanisms which do not involve a receptors, perhaps cholinergically-
mediated shifts in blood flow (Anderson et al., 1976). During the
avoidance period, a combination of increased sympathetic and decreased
parasympathetic activity leads to increased heart rates, while elevated
arterial pressure is due to continued sympathetic peripheral vasoconstric
tion (Galosy et al., 1982).
In humans, the stress response, as elicited by a demanding
arithmetic test (Brod et al., 1959), consisted of the following:
vasodilatation in skeletal muscle, coronary vessels and brain, while most
other resistance vessels and veins underwent a neurogenic vasoconstric
tion. Increased sympathetic and decreased parasympathetic influences on
the heart resulted in tachycardia, increased cardiac ouput and slightly
increased mean arterial pressure. Neuroendocrine responses included
increased epinephrine, ACTH, corticosteroids, vasopressin and renin-

45
angiotensin-aldosterone by means of sympathetic nerve stimulation of the
kidney (Brod, 1970). Brod et al. (1962) demonstrated that this cir
culatory pattern exists in hypertension, especially the early stages.
Eliasson (1984) observed that the limbic-hypothalamic pattern of stress
responses is nearly identical in animals and man. He proposed that modern
humans develop hypertension because they are socially deprived of the
acting-out behavior which is a large part of the stress or defense
reaction. However, the autonomic and hormonal changes associated with
the stress reaction are not suppressible, therefore hypertension results.
4. Neuroendocrine Changes in Response to Stress
a. Stress-Induced Renin Secretion
The physiological response to stress involves the release of
catecholamines from the adrenal medulla and sympathetic nerve endings.
Stimulation of the renal nerves increases renin secretion (Di Bona, 1985;
Blair et al., 1985), and this stimulatory effect is mediated, at least in
part, via norepinephrine released at the endings of the postganglionic
sympathetic neurons that innervate the kidneys (Davis and Freeman, 1976;
Reid et al, 1978; Ganong and Barbieri, 1982). Circulating epinephrine and
norepinephrine can also stimulate renin secretion.
Many investigators have reported increases in plasma renin activity
in response to a variety of stressors. Clamage et al. (1976) reported
increases in plasma renin activity in rats following 30 minutes exposure
to either novel "open field" environment or the presence of a hungry cat.
Jindra et al. (1980) observed increases in plasma renin activity after
acute and repeated immobilization stress. Paris et al. (1987) reported

46
increased plasma renin activity by the following stressors: 12 minute
conditioned fear paradigm; 20 minute immobilization; 20 minute forced
swimming in cold water; 2, 12 and 22 minutes of intermittent footshock;
and a 3 minute conditioned fear paradigm with 0, 10, 30 and 60 minute
recovery. Blair et al. (1976) reported an elevation of plasma renin
activity in baboons during avoidance performance. Gotoh et al. (1985)
demonstrated that bilateral knife cuts behind the hypothalamic paraven
tricular nucleus (PVN) reduced the renin response to immobilization and
the gravitational stress of head-up tilting. Plasma renin activity
clearly can be increased by many different types of stressors.
Sigg et al. (1978) found that restraint causes an increase in plasma
renin activity which was not blocked by pretreatment with the &-adrenergic
antagonists dl-propranolol or sotalol. However, propranolol pretreatment
prevented the increase in plasma renin activity following swim stress and
ether stress (Pettinger et al., 1972) and intermittent footshock (Leenen
and Shapiro, 1974). Also, Van de Kar et al. (1985) have shown that
propranolol significantly inhibits the increase in plasma renin activity
following a three-minute conditioned fear paradigm. Evidence for a role
of central catecholamines in stress-induced renin secretion was provided
by Lightman et al. (1984) who demonstrated that injections of the
catecholamine neurotoxin 6-0HDA into the dorsal noradrenergic bundle
markedly attenuated the increase in plasma renin activity in response to
hemorrhage. Kobayashi et al. (1975) have reported that afferent fibers
from the locus coeruleus to the PVN travel in the dorsal noradrenergic
bundle. These studies suggest that central catecholaminergic projections
to the PVN may be involved in the renin response to stress.

b. Stress-Induced Corticosterone Secretion
1. Introduction
47
Corticosterone levels have been shown to increase in response to
several stressors. For example, Armario et al. (1986) demonstrated that
the corticosterone response to stress is directly related to the intensity
of the stress stimulus. The stressors used in their study were graded
levels of low intensity stressors (either transfer to a novel cage or
room, or a loud noise). These results are in agreement with findings by
Kant et al. (1983), who demonstrated that increases in plasma cor
ticosterone levels were related to the intensity of footshock. They found
that corticosterone secretion was very sensitive not only to an obvious
stressor (footshock) but also to any change in the environment accompany
ing the procedure. Simply placing the rat in the shock box without
delivery of any current elicited a strong corticosterone response. These
studies suggest that the increase in corticosterone levels in response to
stress can represent a sensitive index of the intensity of emotional
arousal experienced by the animals.
Dohanics et al. (1986) found that the adrenocortical response to
ether stress occurs independently of neurohypophyseal function, and that
the full corticosterone and ACTH response to ether or ether plus
laparotomy stress requires not only an intact PVN but also an intact
neurointermediate lobe.
2. Stress-Induced Secretion of CRF
CRF has been shown to mediate stress-induced ACTH release in rats

48
(Rivier and Vale, 1983). CRF also has been shown to initiate other
biological actions in response to stress. For example, injections of CRF
into the cerebral ventricles produced significant increases in plasma
concentrations of epinephrine, norepinephrine, glucagon and glucose,
elevated mean arterial pressure and heart rate, and inhibited gastric acid
secretion. Pretreatment with the ganglionic blocking drug chlorisondamine
completely abolished the increases in plasma catecholamines and glucose,
heart rate and blood pressure (Lenz et al., 1987). CRF has been shown to
activate brain catecholamine systems. Studies by Dunn and Berridge (1987)
demonstrated that injections of CRF into the cerebral ventricles of mice
resulted in increased dihydroxyphenylacetic acid (DOPAC):dopamine ratios
in prefrontal cortex, septum, hypothalamus and brain stem compared to
saline controls. The ratio of 3-methoxy-4-hydroxyphenylethyleneglycol
(MHPG):norepinephrine was increased in the prefrontal cortex and
hypothalamus. DOPAC and MHPG are the major catabolites of dopamine and
norepinephrine, respectively. CRF release has been measured in conscious,
unrestrained rats using a push-pull cannula system implanted in the median
eminence (Ixart et al., 1987). Basal CRF release occurs in a pulsatile
pattern with peaks about every 45 minutes (peak mean 9.0 ± 0.7 pg vs.
trough 4.1 ± 0.3 pg). Ether elicited an increase in CRF which lasted
about 45 minutes with peak values of 54.3 ± 3.2 pg.
3. Stress-Induced Secretion of ACTH
Gibbs (1984) found that ACTH secretion increased in response to
restraint, cold or ether stress. However, Armario et al. (1985) have
found dissociations between ACTH and corticosterone responses: the ACTH

49
response to restraint stress, unlike the corticosterone response, was the
same in control and in chronically stressed rats. This suggests that the
increased corticosterone levels from restraint stress are not necessarily
due only to increased pituitary ACTH release.
The increase in ACTH following hemorrhage was prevented by PVN
lesions, while the responses of renin, epinephrine, norepinephrine, mean
arterial blood pressure and heart rate to hemorrhage were not affected
(Darlington et al., 1988). However, these lesions consisted of creating
an island of the PVN and dorsal hypothalamus by lowering a triangular
(Halasz-type) knife into the third ventricle to the level of the PVN and
then rotating. This may have disrupted fiber pathways to and from the
PVN without actual damage to PVN cell bodies.
Makara et al. (1986) proposed that ACTH hypersecretion following
adrenalectomy is driven primarily by CRF- and/or vasopressin-producing
neurons in and around the PVN, while either (a) other sources of CRF-41,
(b) increased pituitary sensitivity, or (c) other hypothalamic factors
may restore the stress-induced ACTH release in the absence of the PVN.
The effect of bilateral lesions of the PVN on circadian rhythm and
ether stress responsiveness of plasma ACTH and corticosterone were studied
in chronically cannulated female Sprague-Dawley rats. Three weeks after
PVN lesions, ACTH values had dropped by at least half, and the diurnal
maximum and amplitude for corticosterone were diminished by one third.
The peaks for both hormones following ether stress were only half as high
as the control peaks (Ixart et al., 1982). These results support the role
of the PVN as a mediator of stress- induced ACTH and corticosterone
secretion.

50
4. Role of Vasopressin in Stress-Induced Corticosterone Secretion
Vasopressin promotes the retention of water by the kidney, to
maintain plasma volume. There are two major vasopressin systems in the
hypothalamus. One system has cell bodies in the PVN and supraoptic
nuclei, projecting through the internal zone of the median eminence to
the pars neuralis of the pituitary gland. The other system consists of
cell bodies in the PVN with nerve terminals in the median eminence
(Silverman and Zimmerman, 1983). One difference between these systems is
that adrenalectomy resulted in increased staining intensity of vasopressin
immunoreactive nerve fibers in the external zone of the median eminence,
whereas the fibers of the neural lobe or intenal zone of the median
eminence were not affected (Zimmerman et al., 1977).
Vasopressin has been shown by Tilders et al. (1985) to have an
important role in the stress-induced activation of the pituitary-adrenal
system, possibly by potentiating the effects of CRF. Pretreatment of rats
with an antiserum to vasopressin inhibited the ACTH response to both
restraint and formalin stress. Vasopressin has also been shown to
potentiate the activity of CRF (Gillies et al., 1982). Vasopressin
secretion is stimulated by hemorrhage (Robertson, 1977), hypoglycemia
(Bayliss and Robertson, 1980), electric shock (Husain et al., 1979) and
ether (Gibbs, 1984).
Further evidence for the involvement of vasopressin in the ACTH
response to stress is that the vasopressin antagonist dpTyr(Me)arginine
vasopressin decreases ether stress-induced ACTH release, and administra
tion of both the vasopressin antagonist and CRF antiserum completely

51
prevented stress-induced ACTH secretion (Rivier and Vale, 1983). However,
some studies reported that dpTyr(Me)arginine vasopressin does not block
either the intrinsic ACTH-releasing activity or its synergistic effects
with CRF (Baertschi et al., 1984; Knepel et al., 1984) and, therefore,
should be avoided in studies involving ACTH release (Tilders et al.,
1985).
5. Role of Catecholamines in Stress-Induced Corticosterone
Secretion
Controversy still exists as to whether catecholamines play an
excitatory or inhibitory role in stress-induced corticosterone secretion.
i. Evidence for an Inhibitory Role of Catecholamines
Brain norepinephrine has been postulated to inhibit ACTH secretion
(van Loon et al., 1971; Weiner and Ganong, 1978). Eisenberg (1975)
administered reserpine (systemically) or phenoxybenzamine (centrally) and
found an augmentation of the plasma corticosterone response to ether.
This suggests that norepinephrine may function in the central nervous
system to inhibit the stress response. However, reserpine is not a
selective noradrenergic depletor as it will also deplete serotonin stores
(Bugnon et al., 1983).
Suemaru et al. (1985) observed significant increases in serum ACTH
and corticosteorne levels 15 minutes after the onset of either ether
laparotomy or cold-restraint stress. The hypothalamic CRF concentration
showed a rapid increase 2.5 minutes following ether-laparotomy stress,
followed by a significant decrease after 15 minutes, and a subsequent

52
increase. Norepinephrine concentration in the hypothalamus was reduced,
whereas dopamine concentration was increased. Epinephrine concentrations
did not show a significant change throughout the stress procedure. From
these observations they concluded that norepinephrine in the hypothalamus
may not be involved in stimulating hypothalamic CRF secretion in the early
phase of acute stress.
Glavin et al. (1985) depleted brain norepinephrine in rats by
injecting the dopamine-E.-hydroxylase inhibitor FI.A-63 (to prevent the
conversion of dopamine to norepinephrine) followed by R04-1284, a
reserpine-like vesicular depletor of norepinephrine, dopamine and
serotonin. This treatment produced significant depletion of norepineph-
rine in all brain regions examined. Norepinephrine depletion did not
prevent the increase in plasma corticosterone levels following cold-
restraint stress. However, in the non- stressed rats, there was no
significant difference in plasma corticosterone levels between the
norepinephrine-depleted and saline-treated rats. The authors suggest that
norepinephrine does not mediate cold restraint stress-induced increases
in plasma corticosterone levels, nor does it play an inhibitory role with
respect to corticosterone secretion. However, this study combines two
non-related stimuli (cold vs. immobilization) as stressors without
addressing how this could affect hormonal responses.
Reinstein et al. (1985) have demonstrated that acute, uncontroll
able stress (restraint and intermittent footshock for one hour) depleted
norepinephrine in the rat brain (increased norepinephrine turnover).
Pretreatment with supplemental tyrosine (to prevent norepinephrine
depletion) prevented hypothalamic norepinephrine depletion and suppressed

53
the increase in plasma corticosterone, suggesting an inhibitory role for
norepinephrine. However, these corticosterone data were reported as
percent of non-stressed control values, which were quite high (37 .4
ug/dl). In conclusion, there is weak evidence in these studies to support
an inhibitory role for norepinephrine in corticosterone secretion.
ii. Evidence for a Stimulatory Role of Catecholamines
in Stress-Induced Gorticosterone Secretion
Gibson et al. (1986) found that bilateral injections of 6-0HDA into
the PVN prevented the restraint stress-induced increase in plasma
corticosterone levels. Pretreatment with the a 1-adrenoceptor antagonist
prazosin, at a dose which did not influence resting hormone levels,
inhibited the increase in plasma corticosterone following restraint
stress. The authors suggest an excitatory role for norepinephrine via a
receptors in CRF release during acute stress. Similarly, Feldman et al.
(1986) injected 6-0HDA into the PVN and found that PVN norepinephrine
depletion did not affect basal levels or ether stress-induced increases
in corticosterone levels, but it did significantly inhibit the adrenal
response to photic and acoustic stimuli and sciatic nerve stimulation,
suggesting that noradrenergic terminals in the PVN play a role in the
activation of adrenocortical responses following afferent neural stimuli.
Smythe et al. (1983) support the hypothesis that stress-induced ACTH
release from the pituitary is stimulated by hypothalamic noradrenergic
neuronal pathways. They measured norepinephrine activity in the
mediobasal hypothalamus by assessing the ratio of 3,4-dihydroxyphenyl
ethyleneglycol (DHPG) /norepinephrine. Hypothalamic norepinephrine

54
activity was rapidly increased by cold swim stress in the rat, accompanied
by increased ACTH/corticosterone release into the blood. Norepinephrine
activity and ACTH release were also stimulated in rats following ether
stress. The significant correlation between serum corticosterone and the
ratio of hypothalamic DHPG/norepinephrine (shown by linear regression
analysis of the combined data for control and stressed rats in their
studies) is not evidence, but suggests that hypothalamic norepinephrine
neuronal activity may play an important stimulatory role in stress-induced
ACTH release.
Studies by Johnston et al. (1985) support the data presented by
Smythe et al. (1983). Norepinephrine, dopamine and 5-HT concentrations
as well as the concentrations of their respective metabolites MHPG, DOPAC
and 5-hydroxyindole-3-acetic acid (5-HIAA) were measured by HPLC. The
metabolism of norepinephrine, dopamine and 5-HT in different rat brain
nuclei was estimated by the ratio of the metabolite to its respective
amine. Ether stress produced large increases in plasma ACTH, accompanied
by significant increases in norepinephrine metabolism in the PVN, arcuate
and dorsomedial nuclei. Serotonin metabolism was decreased in the PVN,
supraoptic nucleus and caudal division of the arcuate nucleus, while
significant increases in 5-HT metabolism were detected in the suprachias-
matic nucleus and rostral division of the arcuate nucleus. Dopamine
metabolism was selectively increased in the rostral division of the
arcuate nucleus. The authors interpret the data as suggesting a
stimulatory role for norepinephrine, possibly in the PVN with respect to
ACTH secretion.
Studies by Mermet and Gonon (1988) used differential normal-pulse

55
voltammetry combined with carbon fiber electrodes to measure in vivo
extracellular catechols released by noradrenergic nerve terminals in the
PVN. In rats pretreated with the monoamine oxidase inhibitor pargyline,
ether stress produced an increase in the catechol signal corresponding to
extracellular norepinephrine. Electrical stimulation of the ventral
noradrenergic bundle also caused an immediate increase in the signal.
The increase in norepinephrine release following ether stress was
immediate and lasted as long as the stimulus duration. No dopamine
release was recorded from either the striatum or the Al3 dopaminergic cell
body group caudal to the PVN. These data suggest a facilitatory action
of norepinephrine on PVN cell bodies which are involved in the hormonal
responses to stress.
Immobilization stress (varying from 15 to 180 minutes) significant
ly increased MHPG-S04 levels and decreased norepinephrine levels in the
following rat brain regions: hypothalamus, amygdala, hippocampus,
thalamus, pons, medulla and cerebral cortex, with the most rapid and
profound increase in MHPG-S04 found in the hypothalamus. However, the
basal ganglia demonstrated the reverse process- increases in norepineph-
rine levels with transient decreases in MHPG-S04 levels. There was a
concomitant increase in plasma corticosterone levels following immobiliza
tion stress (Tanaka et al., 1982). This study suggests that norepineph
rine released in the hypothalamus during immobilization stress may play
a role in the stress-induced increase in corticosterone levels.
Beaulieu et al. (1987) demonstrated that bilateral lesions of the
central nucleus of the amygdala significantly inhibited the stress-induced
ACTH secretion following immobilization. These lesions also reduced the

56
stress-induced noradrenergic activity in the anterior and lateral
hypothalamic areas, the PVN and arcuate nucleus, and the bed nucleus of
the stria terminalis. These results suggest that norepinephrine plays a
stimulatory role in ACTH secretion and is somehow regulated by neurons in
the central nucleus of the amygdala.
As stated earlier, the ventral noradrenergic bundle (VNAB) is
thought to be the source for the noradrenergic modulation of ACTH
secretion (see CORTICOSTERONE: Role of the VNAB). Szafarczyk et al.
(1985) have shown that injection of 6-0HDA into the VNAB obliterates the
circadian patterns for ACTH and corticosterone secretion, and also caused
an 80% inhibition of the ACTH response to stress.
Injections of 6-0HDA into the VNAB also prevented the increase in
corticosterone secretion following exposure to photic and acoustic stimuli
and sciatic nerve stimulation (Feldman et al., 1988). However, basal
levels of corticosterone and ether-induced increases in corticosterone
were not affected. The 6-0HDA-injected rats had average norepinephrine
depletions of 72% in the mediobasal hypothalamus and 78% in the paraven-
tricular nucleus when compared to vehicle-injected rats. These data
support previous studies (Feldman et al., 1986) in which 6-0HDA injections
into the PVN did not affect basal levels or ether-induced increases in
corticosterone levels, but did significantly inhibit the adrenal response
to photic and acoustic stimulation and sciatic nerve stimulation. These
studies suggest that norepinephrine plays a role in the activation of
adrenocortical responses following afferent neural stimuli.
In contrast, Velley et al. (1988) have shown that 6-0HDA lesions
in the locus coeruleus did not prevent the effect of novel environment

57
stress on corticosterone, although there was a significant (53%) depletion
of hypothalamic norepinephrine. However, this can be explained by the fact
that about 90% of the norepinephrine supply to the PVN originates from the
lower brainstem Al and A2 cell groups, while the locus coeruleus
projection to the PVN is only about 8% (Sawchenko and Swanson, 1982).
iii. Role of Epinephrine in Stress-Induced
Corticosterone Secretion
Szafarczyk et al. (1987) demonstrated a stimulatory role for
epinephrine (and norepinephrine) on corticosterone release when
intracerebroventricular (icv) injections of epinephrine or norepinephrine
induced ACTH release comparable to that occurring under conditions of
ether stress. Epinephrine was more efficacious than norepinephrine on an
equimolar basis. The stimulating effect of norepinephrine was reversed
by icv pretreatment with prazosin, an a 1- adrenoceptor antagonist, whereas
the effect of epinephrine was inhibited by either prazosin or the .&
blocker propranolol. These results suggest that norepinephrine triggers
ACTH release via a1-adrenoceptors, whereas epinephrine acts via both a 1-
and B-adrenoceptors. It should be noted that they also administered a
combined pretreatment of prazosin and propranolol, and this combined
treatment was no more effective than either antagonist alone. They also
demonstrated that a unilateral juxta-PVN injection of either epinephrine
or norepinephrine produced an increase in plasma ACTH concentration with
a time course similar to that for an icv injection, and pretreatment with
anti-rCRF-41 serum caused a 50-60% reduction in the ACTH response to
either catecholamine. Since the majority of diencephalic CRF neurons are

58
located in the parvocellular subnuclei of the PVN (Swanson et al., 1983;
Liposits et al., 1985), these data suggest a role for a1
and B receptors
in the PVN in the regulation of corticosterone secretion.
Additional evidence was obtained by Spinedi et al. (1988) who
demonstrated that central inhibition of the enzyme that converts
norepinephrine to epinephrine, phenylethanolamine-N-methyltransferase
(PNMT), produced a selective decrease in hypothalamic epinephrine levels,
significantly decreased the plasma ACTH response to ether stress, and at
later times also caused a selective decrease in CRF content. In addition,
the altered ACTH response to ether stress was not due to changes in
arginine vasopressin levels or changes in pituitary responses to CRF.
These results support a stimulatory role of central epinephrine neurons
on CRF release.
6. Role of Serotonin (5-HT) in Stress-Induced
Corticosterone Secretion
The role of serotonin as a mediator of stress-induced corticosterone
secretion is not clear. Electrolytic lesions in the dorsal raphe nucleus
prevented the stress-induced increase in corticosterone levels as well as
increases in plasma renin activity and plasma renin concentration
following a ten-minute conditioned fear paradigm (Richardson Morton et
al., 1986). Other studies from our laboratory have shown that injection
of the neurotoxin 5,7-dihydroxytryptamine into the dorsal and median raphe
nucleus did not prevent the stress-induced increase in renin and
corticosterone secretion. Furthermore, i. p. injection of the 5-HT2
antagonist LY53857 did not prevent the stress-induced increase in renin

59
and corticosterone secretion (Richardson Morton et al., 1986), whereas low
doses of the 5-HT1 agonist buspirone and ipsapirone inhibited the stress
induced increase in corticosterone and renin secretion (Van de Kar et al.,
1985; Urban et al., 1986; Lorens et al., 1989). These studies suggest
that a non-serotonergic (or possibly 5-HT1) mechanism is involved in
stress-induced renin and corticosterone secretion.
Feldman et al. (1987) reported that injection of the serotonergic
neurotoxin 5, 7-dihydroxytryptamine (5, 7-DHT) into the PVN blocked the
adrenocortical responses to cortical and extra-hypothalamic limbic
stimulation, but had no effect on the adrenocortical response to ether
stress. Immobilization and cold-exposure stress resulted in four-fold
increases in ACTH-like imrnunoreactivity in plasma. This increase was
potentiated by i.p. injections of the 5-HT reuptake inhibitor fluoxetine
and significantly reduced by the 5-HT antagonists metergoline and
cinanserin (Bruni et al., 1982). These results suggest that 5-HT may play
a role in stress-induced ACTH release. In contrast, Fuller and Snoddy
(1977) found that pretreatment with fluoxetine did not significantly alter
the increase in plamsa corticosterone levels following swim stress or
insulin-induced hypoglycemia. However, fluoxetine did enhance the
corticos terone response produced by an inj ec ti on of L- 5-hydroxytryptophan.
These results suggest that serotonergic neural pathways are involved in
mediating the adrenocortical response to some stressful stimuli (i
mmobilization and cold exposure) but not others (swim stress and insulin
induced hypoglycemia).
Beaulieu et al. (1986) support the studies by Bruni et al. (1982).
They found that bilateral lesions of the central nucleus of the amygdala

60
resulted in significantly decreased immobilization stress-induced ACTH
secretion with a concomitant increase in serotonergic activity in the PVN
and arnygdaloid nuclei. Additional studies by Beaulieu et al. (1987)
suggest a serotonergic involvement in immobilization stress. In these
studies, bilateral lesions of the central nucleus of the amygdala in
unstressed rats increased the serotonergic activity in the PVN, dor
somedial nucleus, anterior hypothalamic area (AHA), cortical and medial
amygdaloid nuclei, and the lateral part of the basal amygdaloid nucleus.
Immobilization stress applied to lesion rats resulted in a decrease in
serotonergic activity to control levels in the PVN, AHA and the dor
somedial nucleus, and below control levels in the ventromedial nucleus.
The serotonergic activity remained elevated in each of the amygdaloid
areas measured, which are rich in glucocorticoid receptors (Beaulieu et
al., 1986). Glucocorticoids have been reported to down-regulate their own
receptors in a site-specific manner in the amygdala, septum and hippocarn
pus, but not in the hypothalamus or pituitary (Sapolsky et al., 1984).
Also noradrenergic cell bodies (Al-A7), serotonergic cell bodies (Bl-B9),
and PNMT-immunoreactive nerve cells of the adrenergic cell groups (Cl-C3)
have been shown to have strong glucocorticoid receptor immunoreactivity
(Harfstrand et al., 1986). De Kloet et al. (1986) have shown that
corticosterone selectively controls the 5-HT1 receptor density in the
dorsal raphe and dorsal hippocampus. These studies suggest that serotonin
may modulate the negative feedback of glucocorticoids on stress-induced
ACTH secretion. A moderate to strong nuclear glucocorticoid receptor
immunoreactivity has been demonstrated in the majority of CRF-immunoreac
tive neurons of the parvocellular PVN, septohypothalamic thalamus, bed

61
nucleus of the stria terminalis, and the medial and central amygdaloid
nuclei (Cintra et al., 1987).
C. Effect of Peripheral Sympathectomy on Stress-Induced Renin and
Corticosterone Secretion
The physiological response to stress involves the release of
catecholamines from the adrenal medulla and sympathetic nerve endings.
These catecholamines stimulate the renal nerves to increase renin output.
It is generally accepted that the sympathetic nervous system mediates the
stress response from the brain to the kidney (Golin et al., 1988; Reid et
al., 1978). Studies by Van de Kar et al. (1985) have shown that the &
adrenoceptor antagonist propranolol significantly inhibits but does not
completely block the stress-induced increase in plasma renin activity.
It should be noted, however, that this effect could be mediated by either
central or peripheral &-receptors since propranolol is known to cross the
blood brain barrier (Giarcovich et al., 1984; Lemmer et al., 1985). To
determine whether stress-induced renin secretion is mediated by the
peripheral sympathetic nervous system, rats were adrenal enucleated and
given weekly injections of 6-0HDA (i.p.). The adrenal medulla was removed
since it does not effectively take up 6-0HDA. The effectiveness of the
sympathectomy/adrenal enucleation was verified by determination of plasma
epinephrine (less than 1% of control) and norepinephrine (less than 10%
of control). The stressor induced a significant increase in plasma renin
activity and concentration. Sympathectomized-adrenal enucleated rats
showed no inhibition of the stress - induced increase in plasma renin
activity/concentration or plasma corticosterone levels (Richardson Morton

62
et al., 1988). The data suggest that the sympathetic nervous system and
adrenal catecholamines are not the only mediators of the stress-induced
increase in renin secretion.
5. Central Nervous System Pathways Involved in the
Stress Response
a. Sensory Afferents
With respect to the role of the central nervous system in the
conditioned fear response, it is important to consider how the sensory
stimulus (pain) is transmitted to the cerebral cortex for analysis.
Specialized nerve fibers found throughout the skin serve as end organs
for pain. These fibers enter the spinal cord and ascend through the
lateral spinothalamic tract to the posteroventral nucleus of the thalamus.
From here the fibers travel to the appropriate loci in the cerebral
cortex, depending on the somatic origin of the pain message.
The ascending reticular activating system (ARAS) is a network of
interconnective fibers that also travel from the top of the spinal cord
to the thalamus, and eventually to the cortex. This tract receives major
sensory inputs, and conveys information regarding alertness and excite
ment.
There appears to be a close connection between pain sensation and
the ARAS. The midbrain and brainstem reticular formations receive many
pain afferents from the spinothalamic tract. Studies in animals (Olds
and Olds, 1965) and humans (Gray, 1971) have suggested that stimulation
through electrodes in the periventricular midbrain reticular system
generates pain sensation. Other pain reception fibers have been found in

63
the thalamic portion of the ARAS. Lesions in the thalamic intralaminar
nuclei and the thalamic nucleus centrum medianum have been shown to
alleviate intractable pain in humans (Gray, 1971).
b. The Limbic System
1. Septo-Hippocampal System
The limbic system, in particular the septo-hippocampal system,
appears to play a major role in pain reception. The hippocampus emits a
theta wave (4-9 cps rhythm) as a response to a painful stimulus in the
rat, and in the emotional states of disappointment and frustration in
humans (Gray, 1971). This theta wave can be abolished by lesions in the
medial nuclei of the septum, and the thalamic nucleus centrum medianum,
suggesting that the pain input route most likely travels from the midbrain
reticular formation, through the thalamus, to reach the septum and
hippocampus (Gray, 1971).
2. Amygdala
The amygdala is a limbic structure with many interconnections to the
hippocampus (Krettek and Price, 1978). The amygdaloid complex has been
shown to be involved in the regulation of fear-motivated behavior
(Blanchard and Blanchard, 1972; Spevack et al., 1975). In addition, the
amygdala appears to play a role in the stress response via the pituitary-
adrenal cortical axis. The central nucleus of the amygdala, in par-
ticular, contains large numbers of CRF-containing cell bodies and
terminals (Swanson et al., 1983; Moga and Gray, 1985). Injection of CRF
into the lateral ventricles of rats or dogs elicits autonomic and

64
behavioral changes that are characteristic of the stress or defense
reaction (Brown et al., 1982; Lenz et al., 1987; Britton
et al., 1982). Electrical stimulation of the amygdaloid complexes results
in an increase in adrenal glucocorticoid secretion in rats (Redgate and
Fahringer, 1973), cats (Redgate, 1970) and monkeys (Mason, 1959). The
amygdaloid complex is an essential part of the ACTH controlling mechanism
in rats (Knigge, 1961; Allen and Allen, 1974). Specifically it has been
shown to be necessary for the hypersecretion of ACTH following adrenalec
tomy (Allen and Allen, 1975). In these studies, lesions which destroyed
the direct medial-projecting amgdalo-hypothalamic fibers blocked the
hypersecretion of ACTH following adrenalectomy, while lesions of the
septal region, stria terminalis and fornices did not.
Lesions in the amygdaloid central nucleus impaired one-way active
avoidance learning (Werka et al., 1978), produced marked deficits in
passive avoidance performance (Mcintyre and Stein, 1973; Grossman et al.,
1975), and significantly decreased ACTH secretion in response to
immobilization stress (Beaulieu et al., 1987). Axons from cells in the
olfactory bulb terminate in the amygdala (Powell et al., 1965) and the
medial hypothalamus receives axons from cells in the amygdala (Cowan et
al., 1965). Bilateral amygdalectomy significantly reduced the adrenocor
tical responses to sciatic nerve and olfactory stimulation, without
affecting the corticosterone response to ether or to photic and acoustic
stimulation (Feldman and Conforti, 1981). This amygdalo-hypothalamic
pathway is believed to play a role in the adrenocortical response to a
number of somatosensory stimuli (Feldman et al., 1975). The amygdala has
also been shown to play an important role in regulating the cardiovascular

65
system during Sidman avoidance stress testing (Galosy et al., 1982).
3. Hypothalamic Ventromedial Nucleus
With respect to the central neural circuits involved in the
fight/flight behavior, de Molina and Hunsperger (1962) described a pathway
in cats that descends from the amygdala along the stria terminalis to the
ventromedial hypothalamus (VMN) with connections to the midbrain central
grey. Stimulation of sites within the hypothalamus, amygdala and midbrain
central grey could produce defensive attack while other loci immediately
adjacent to those stimulated would produce flight. When the VMN was
removed, stimulation of the amygdala no longer produced defensive attack,
and removal of the central grey prevented the flight behavior following
hypothalamic stimulation.
In addition, Kling et al. (1958) reported that cats made docile by
lesions in the amygdala could be made ferocious by subsequent lesions in
the VMN. The converse did not apply - cats first made ferocious by
lesions in the hypothalamus were not affected by a subsequent amygdalec
tomy.
Based on these and other observations, Gray (1971) suggested that
the midbrain central grey is the ultimate controlling area for either
flight or defensive behavior. In addition, there is a tonic inhibitory
input to the central grey from the VMN. Further modulation of this
inhibitory input would come from the amygdala and the septo-hippocampal
system. Input from the amygdala would prevent the inhibition, resulting
in fight/flight behavior, and the septo-hippocampal input would intensify
passive avoidance behavior. The resulting behavior (either passive

66
avoidance or fight/flight) would depend on which of the inputs to the VMN
is stronger under a particular set of environmental conditions. This
scheme would provide a basis for the observation that, in times of danger,
both freezing and fight/flight patterns may be stimulated simultaneously
and alternate rapidly with each other.
Neurons in the VMN have been shown to play a role in stress-induced
renin secretion. Gotoh et al. (1988) found that bilateral electrolytic
lesions in the VMN prevented the increase in plasma renin activity
following immobilization stress. Lesions in the VMN led to excessive
levels of food intake and adiposity (Hetherington and Ranson, 1942), and
elicited a variety of metabolic and behavioral disturbances termed the VMN
syndrome, which has become a model for the study of obesity and motivated
behavior (Weingarten et al., 1985; Stellar, 1954).
c. Role of Brain Catecholamines in Stress
A variety of stressors including foot shock (Bliss et al., 1968;
Thierry et al., 1968, Iimori et al., 1981), immobilization (Corrodi et
al., 1968; Tanaka et al., 1981) and ether stress (Vellucci, 1977) have
been shown to decrease norepinephrine concentration and increase
norepinephrine turnover in the rat brain.
The metabolism of norepinephrine in the rat brain results in the
production of the sulfate conjugates MHPG-S04 , the major metabolite, and
DHPG-S04 (Schanberg et al., 1968; Sugden and Eccleston 1971). The
functional activity of central noradrenergic neurons has been shown to be
directly related to the production of MHPG-S04 (Stone, 1975). Levels of
MHPG-S04 in rat brain represent a useful index for norepinephrine turnover

67
in discrete brain regions (Kohno, 1981) as well as in the whole brain
(Korf et al., 1973; Stone, 1976).
Rats subjected to a mild psychological stressor (exposure to
responses of shocked rats) had decreased norepinephrine and increased
MHPG-S04 concentrations preferentially in the hypothalamus and amygdala.
Rats subjected to footshock stress had decreased norepinephrine and
increased MHPG-S04 not only in the hypothalamus and amygdala but also the
pons, medulla oblongata and cerebral cortex (Iimori et al., 1981). This
study suggests that the noradrenergic response to stress varies with the
nature or intensity of the stressor.
Brain noradrenergic systems, in particular the locus coeruleus
noradrenergic system, have been shown to play a role in nociception,
anxiety and fear (Charney and Redmond, 1983). The locus coeruleus
receives innervation from sensory (pain) pathways in the spinal cord, and
relays information to the cerebral cortex and limbic structures (hypothal
amus, hippocampus, amygdala) (Kuhar, 1986).
Rasmussen and Jacobs (1986) have recorded the electrical activity
of noradrenergic neurons in the locus coeruleus of freely moving cats.
During GER training, locus coeruleus units showed a large increase in
activity in response to a stimulus paired with a noxious air puff, whereas
no increase in unit activity is seen in response to a stimulus not paired
with the air puff. Also, electrical stimulation of the locus coeruleus
in monkeys elicited behavioral responses identical to those seen in
experimentally-induced or naturally-occurring fear states (Bertrand,
1969).
In humans, drugs that increase noradrenergic function have been

68
shown to produce anxiety. The a 2-adrenoceptor antagonists yohimbine and
piperoxane induced anxiety states in normal subjects (Soffer, 1954;
Holmberg and Gershon, 1961). These studies support the role of brain
norepinephrine in mediating the response to anxiety and fear.
The Kety hypothesis (Kety, 1970) proposed that forebrain nor
epinephrine enhances cell firing in neurons that receive afferent
environmental information to "affectively important events". Further,
norepinephrine would produce a "persistent facilitation" of these inputs
which would reinforce the memory process for these events. The extensive
innervation of the limbic system (especially the hippocampus) by the locus
coeruleus-norepinephrine system suggests that noradrenergic neurons may
play a role in the memory or cognitive process of stress.
D. HYPOTHALAMIC PARAVENTRICULAR NUCLEUS (PVN)
A. Introduction
The PVN is a bilateral, triangular-shaped nucleus at the dorsal
end of the third ventricle in the hypothalamus. It is a morphologically
complex structure, with eight clearly distinguishable subdivisions, which
contain as many as thirty different putative neurotransmitters. Three
major efferent projection sites, arising from separate neuron groups,
include (1) the median eminence, (2) the posterior pituitary and (3) the
autonomic centers in the brain stem and spinal cord (Swanson and
Sawchenko, 1983; Saper et al., 1976). This suggests that the PVN serves
as an integrator of endocrine, autonomic and behavioral functions in a
variety of physiological systems.

69
B. Morphology
The PVN is composed of distinct parvocellular and magnocellular
divisions (Krieg, 1932). Swanson and Kuypers (1980) have further
characterized the nucleus into eight distinct subdivisions, three
magnocellular and five parvocellular. The anterior magnocellular part of
the PVN is ventromedial to the descending column of the fornix as it
enters the hypothalamus from the septum. The medial magnocellular
division is just caudal to the anterior magnocellular level. The largest
magnocellular division is the posterior magnocellular part. It first
appears in the ventromedial PVN, just caudal to the medial magnocellular
division. After dorsolateral expansion it forms a compact cell mass at
the lateral margin of the nucleus. Hatton et al., (1976) further
subdivide this group by defining two distinct populations of large cells
in the medial and lateral halves of the posterior magnocellular region.
The lateral group contains cells with multiple nucleoli that are larger
and more uniform in orientation, when compared to the medial group. The
posterior magnocellular cells number from 1300-2000 on either side of the
brain (Olivecrona, 1957; Bodian and Maren, 1951; Swanson and Sawchenko,
1983).
There are five distinct parts of the parvocellular division. The
periventricular part, medial to the rest of the PVN, contains small,
vertically-oriented fusiform cells. The anterior parvocellular group is
not well-defined but consists of small- to medium-sized cells at the level
of the anterior and medial magnocellular parts. Extending through the
caudal half of the nucleus is the medial parvocellular part, which

70
consists of densely packed cells. The dorsal parvocellular part is also
in the caudal half of the nucleus and consists of horizontally-oriented,
medium-sized cells in an elliptical group. The lateral parvocellular part
of the PVN is in the caudal third of the nucleus and is the lateral
expansion of the parvocellular part (the wings). These cells are
horizontally-oriented, medium-sized and fusiform shaped. At its maximum
extent the lateral parvocellular PVN extends laterally over the descending
column of the fornix, about 750-1000 um from the midline (Swanson and
Sawchenko, 1983).
Cells in the parvocellular division project primarily to the
external lamina of the median eminence (Wiegand and Price, 1980; Swanson
et al., 1980) and to the brainstem (dorsal vagal complex) and spinal cord
(Hosoya and Matsushita, 1979; Swanson and Kuypers, 1980). The PVN is also
known to project to those segments of the thoracic cord thought to contain
sympathetic preganglionic neurons which innervate the kidney (Swanson and
Mc Kellar, 1979). Cells in the magnocellular division project primarily
to the posterior level of the pituitary gland (Sherlock et al., 1975).
The parvocellular division of the PVN contains about 7000 neurons
in the following distribution: anterior 1650; dorsal 600; lateral 1300;
medial 2175; periventricular 1275 (Sawchenko and Swanson, 1981).
C. Role of the PVN in Behavior
The PVN plays a major role in the control of feeding. Bilateral
damage to the PVN results in increased levels of food intake and excesive
weight gain (Leibowitz et al., 1981). This eating behavior can be
elicited by hypothalamic injections of norepinephrine and other a-

71
noradrenergic agonists (Leibowitz, 1975). In particular, injection of
the a2-agonist clonidine into the PVN initiates a feeding response in
satiated rats (Marino et al., 1983). Goldman et al. (1985) observed a
potent feeding response in satiated animals following injection of
norepinephrine or clonidine into the PVN. This effect was blocked by the
a-antagonist phentolamine as well as the a2-antagonists rauwolscine and
yohimbine. These results suggest that a2 receptors in the PVN play a role
in mediating feeding behavior. Leibowitz et al. (1984) have demonstrated
that these a2 noradrenergic receptors fluctuate in density with respect to
the circadian cycle, exhibiting a unimodal peak at the onset of the dark
cycle, when feeding normally occurs. This shift in a 2 receptor density may
be correlated with plasma corticosterone levels, which also peak at the
onset of the dark cycle (Krieger and Hauser, 1978; Wilkinson et al.,
1979). Norepinephrine-stimulated feeding behavior was prevented by
adrenalectomy and restored by administration of corticosterone (Leibowitz
et al., 1984).
D. Afferent Connections to the PVN
Descending afferent connections to the PVN include a large input
from the limbic system structures, especially the lateral septa! nucleus
and the ventral portion of the subicular cortex, as well as input from
the amygdala, in particular, the medial nucleus (Silverman et al., 1981;
Tribollet and Dreifuss, 1981). The PVN receives afferent sensory
information from the telencephalic limbic system. The bed nucleus of the
stria terminalis receives a large input from the amygdala (Krettek and
Price, 1978) and smaller projection from the ventral part of the subiculum

72
(Swanson and Cowan, 1977). The bed nucleus has been shown to project to
the PVN by autoradiographic studies (Swanson and Cowan, 1979). This
projection innervates all parts of the parvocellular division and the
oxytocinergic parts of the magnocellular division (Sawchenko and Swanson,
1981). Electrophysiological studies have shown that stimulation of the
amygdala or septum can have inhibitory (and sometimes excitatory) effects
on the PVN magnocellular neurons (Renaud and Arnauld, 1979; Koizumi and
Yamashita, 1972; Pittman et al., 1981).
Anterograde autoradiographic techniques were used to describe brain
stem afferents to various PVN regions (McKellar and Loewy, 1981). The
parabrachial nuclei and locus coeruleus project to the posterior,
periventricular, parvocellular and dorsal divisions. The ventral medulla
projects to the lateral, medial, posterior, dorsal and parvocellular
divisions. The Al catecholamine cell group projects to the posterior,
lateral, medial and dorsal divisions, while the NTS projects to the
parvocellular and dorsal divisions. The projections from the Al and A2
(NTS) catecholaminergic cell groups to the PVN have been further
characterized by an immunofluorescence double-labeling procedure to
determine which anterogradely labeled fibers and terminals in the PVN also
displayed immunoreactive dopamine ~-hydroxylase, indicating the presence
of catecholaminergic neurons (Cunningham and Sawchenko, 1988). (See Role
of Brain Catecholamines in Corticosterone Secretion)
Studies using retrogradely-transported horseradish peroxidase
identified cell bodies of neurons that project to the PVN. Labelled cell
bodies were found in the mediobasal hypothalamus, the limbic system
(lateral septum, posterornedial arnygdala and ventral subiculum) and the

73
brain stem, including the dorsal raphe nucleus, locus coeruleus,
parabrachial nuclei, NTS and lateral reticular nucleus (Tribollet and
Dreifuss, 1981).
E. Efferent Projections of the PVN
Descending autonomic projections of the PVN were investigated using
the anterograde transport technique of Phaseolus vulgaris leuco-agglutinin
(Luiten et al., 1985). Two major fiber bundles were described. One
(fiber bundle 1) leaves the PVN dorsally and runs caudally in a dorsal
periventricular position to reach the midbrain periaqueductal grey, where
a number of terminal boutons are observed. This bundle then courses
laterally to join the lateral lemniscus. Some projections innervate the
dorsal raphe nucleus, the parabrachial nucleus, locus coeruleus and
pontine raphe. The second group of fibers (fiber bundle 2) runs medially
and caudally from the PVN and travels in a position dorsal to the
transition of the internal capsule and medial forebrain bundle. In the
midbrain, fiber bundle 2 moves laterally to cover the dorso-lateral pole
of the substantia nigra, and has some terminations in the ventral aspects
of the reticular formation. At the level of the pons, fiber bundle 2
moves ventrally to join the lateral lemniscus (and bundle 1) to reach the
floor of the brainstem. Posterior to the pons, all descending PVN
efferents are now in a ventral position adjacent to the pyramidal tracts.
While in the medulla, some fibers branch off to terminate in the reticular
and medullary raphe nuclei. Nearly all of the bundle fibers traverse the
lateral reticular nucleus, nucleus ambiguus, parvocellular reticular
nucleus and area postrema, with an especially dense amount of varicosities

74
and terminal boutons in the NTS and dorsal motor nucleus of the vagus.
F. Catecholaminergic Innervation of the Hypothalamic PVN
There is substantive evidence supporting extensive catecholaminergic
innervation of the PVN (Carlsson et al., 1962; Lindvall and Bjorklund,
1974; Hokfelt et al., 1974). Most of the adrenergic and noradrenergic
innervation to the PVN is provided by (1) the Al and Cl cell groups of the
ventral medulla, (2) the A2 and C2 cell groups in the medial NTS and (3)
the A6 cell group, the locus coeruleus of the pontine central gray. The
locus coeruleus receives major fiber projections from the Al region, and
projects to the most medial parts of the parvocellular PVN (Swanson et
al., 1981).
G. Neuroactive Substances in the PVN
The colocalization of CRF and vasopressin in the parvocellular PVN
has already been presented (see CORTICOSTERONE: CRF). These are only two
of many neuroactive substances found in the PVN by immunohistochemical
studies. Swanson and Sawchenko (1983) have compiled a thorough listing:
Cell Bodies include oxytocin and vasopressin in the magnocellular PVN
(Swaab et al., 1975; Vandesande and Dierickx, 1975); somatostatin in the
periventricular part of the parvocellular PVN (Dierickx and Vandesande,
1979); dopamine in the periventricular and adjoining parvocellular PVN
(Hokfelt et al., 1973); met- and leu-enkephalin in the magnocellular and
pavocellular PVN (Rossier et al., 1979; Finley et al., 1981); neurotensin
in the medial and periventricular parts of the parvocellular PVN (Kahn et
al., 1980); dynorphin coexists with vasopressin in the magnocellular PVN
(Watson et al., 1982); substance Pin the parvocellular PVN (Ljungdahl et

75
al., 1978); glucagon in the magnocellular PVN (Tager et al., 1980); renin
in the oxytocinergic parts of the magnocellular PVN (Fuxe et al., 1980);
and ACTH and i-endorphin in the magnocellular PVN (Joseph and Sternberger,
1979; 'Watkins, 1980). The PVN is also the major source of atrial
natriuretic peptide (ANF)-containing cells that project to the median
eminence (Palkovits et al., 1987). Double-labelling studies with antisera
to GAD and CRF in the PVN have indicated that a sub-population of CRF
containing neurons contain the inhibitory neurotransmitter GABA (Meister
et al., 1987).
Nerve terminals and fibers in addition to norepinephrine,
epinephrine and serotonin (previously described) include "cholinoceptive"
acetylcholine neurons (Kimura et al., 1981); ACTH, i-endorphin and a-MSH
in the parvocellular and oxytocinergic divisions of the magnocellular PVN,
all of which originate in the arcuate nucleus (Joseph, 1980; Bloom et al.,
1978; Jacobowitz and O'Donohue, 1978); VIP in the periventricular part of
the parvocellular PVN (Sims et al., 1980); GABA (Perez de la Mora et al.,
1981); TRH in primarily parvocellular PVN (Hokfelt et al., 1975); and
sparse fibers of LHRH (Jennes and Stumpf, 1980); bradykinin (Correa et
al., 1979); prolactin (Fuxe et al., 1977) and angiotensin II (Fuxe et al.,
1976; Changaris et al., 1978).
a. Neuropeptide Y
The distribution of neuropeptide Y (NPY) immunoreactivity in the
PVN as well as its coexpression in noradrenergic and adrenergic brain stem
groups has been determined with the retrograde transport-double immunohis-
tochemical techniques (Sawchenko et al., 1985). Antisera to phenyl-

76
ethanolamine-N-methyltransferase (PNMT) was used as a marker for
adrenergic neurons, and dopamine-&-hydroxylase (DBH) antisera was used as
a marker for adrenergic and noradrenergic neurons. The results indicated
a dense NPY input to most parts of the parvocellular PVN in a similar, yet
more extensive distribution than that stained by DBH antisera. In the
magnocellular division, NPY-stained fibers were evenly distributed
throughout areas containing oxytocinergic and vasopressinergic fibers.
Catecholaminergic fibers were preferentially associated with vasopres
sinergic fibers. Combined retrograde transport-double irnrnunohistochemical
labeling studies demonstrated that NPY irnrnunoreactivity is extensively
contained within the adrenergic Cl, C2 and C3 cell groups, with somewhat
less irnrnunoreactivity in the noradrenergic cell groups, mostly limited to
neurons in the Al cell group.
Leibowitz et al. (1986) found that NPY injected directly into the
PVN increased plasma corticosterone levels within 15 minutes of infusion.
The NPY response was similar to that for epinephrine and norepinephrine
(See CORTICOSTERONE: Stimulation by Norepinephrine and Epinephrine).
Also, NPY injected into the third ventricle results in robust feeding
behavior in satiated rats (Allen et al., 1975) in a similar manner to
adrenergic neurotransmitters (See PVN: Role in Behavior). Studies by Sahu
et al. (1988) suggest that NPY neurons in the brainstem project to the PVN
and mediate the increased sensitivity of the feeding response elicited by
NPY.

CHAPTER III
METHODS
A. Animals
Male Sprague-Dawley rats (200-250 g) were purchased from Sasco-
King Animal Laboratories (Oregon, WI). The rats were housed, two per
cage, in a temperature (20-22°C), humidity (50-55%) and illumination (12:12
hour light/dark cycle; lights on at 0700 h) controlled room. Water and
food (Wayne Lab Blox, Lab Mills Inc. , Chicago, IL) were available ad
libitum.
B. Surgery
All surgery was performed under pentobarbital anesthesia (50
mg/kg, i.p.). The rats received atropine sulfate (0.4 mg/kg, i.m.) to
reduce respiratory tract secretion and ampicillin ( 50 mg/kg, i. m.) to
prevent infection. Chloramphenicol (1% ophthalmic ointment) was topically
applied around the wound margin. The rats were euthanized by decapitation
with a guillotine in an area outside the rat room. Trunk blood was
collected into centrifuge tubes containing 0.5 ml 0.3 M ethylenediamine
tetra acetic acid (EDTA; pH 7.4).
77

78
c. Electrolytic Destruction of the PVN
Bilateral electrolytic lesions in the PVN were produced by passing
1. 0 mA (D. C.) current for 10 seconds through an intracranial cathode. The
anode was clipped to the wound margin. The cathode was a stainless steel
pin (0.25 mm diameter) insulated with Epoxylite except for 0.5 mm of its
tip. A Kopf stereotaxic apparatus and a Grass DC constant current lesion
maker were used. The coordinates for the lesion site were 6.0 mm rostral
to lambda, 0.5 mm lateral to the midsagittal suture, and 8.3 mm ventral
to the skull surface, with the incisor bar set at -4. 0 mm. Control
animals were treated identically except that the electrode was not lowered
into the skull. The rats were allowed to recuperate from surgery for 14
days, after which time they were subjected to a conditioned emotional
(fear) response paradigm (GER). The CER procedure is described in detail
below. The rats were sacrificed by decapitation immediately following the
last CER procedure. Trunk blood was collected and assayed as described
below. The brains were removed and saved for histological verification
of the lesion placement. Eight rats were included in the sham/control as
well as the sham/stress group, while 12 rats per group were used in both
the lesion/control and lesion/stress group.
D. Electrolytic Destruction of the Nucleus Reuniens
Bilateral electrolytic lesions of the nucleus reuniens were
produced by passing 1.0 mA current for 5 seconds through an intracranial
cathode while the anode was clipped to the wound margin. The coordinates
for the lesion site were 5.35 mm rostral to lambda, 0.4 mm lateral to the
midsagittal suture and 8. 0 mm ventral to the skull surface, with the

79
incisor bar set at -4.0 mm. Eight rats were included in the sham/control
as well as the sham/stress group, 10 rats were used in the lesion/control
and 12 rats were used in the lesion/stress group. The rest of the
protocol for this experiment was identical to that described in the
previous section (Electrolytic Destruction of the PVN).
E. Microiniections of Ibotenic Acid into the PVN
Bilateral stereotaxic microinjections of ibotenic acid (Sigma,
St. Louis, MO) were made using a Kopf stereotaxic apparatus with a 5 µl
Hamilton syringe and a Trent Wells hydraulic microdrive, to allow for a
slow delivery rate. Ibotenic acid (10 µg/µl) was dissolved in 0.01 M
phosphosaline buffer (PBS, pH 7.4), and injected in a volume of 0.3 µl
per side over five minutes (Fitzsimons and Ciriello, 1986). The injection
coordinates (incisor bar at -4.0 mm) were 5.7 mm anterior to lambda, 0.5
mm lateral to the midsagittal suture and 8. 5 mm ventral to the skull
surface. Control animals received an injection of the vehicle. After
completion of the injection (five minutes), the needle was left in situ
for an additional five minutes to prevent dorsal diffusion of the
neurotoxin along the needle track. The rats were allowed to recuperate
from surgery for 14 days, after which time they were subjected to the CER
procedure. The rats were sacrificed by decapitation immediately following
the CER procedure, and trunk blood was collected and later assayed. Nine
rats were included in the vehicle/control and eight rats in the vehicle/
stress group, while 12 rats per group were used in both the ibotenic
acid/control and ibotenic acid/stress group.

80
F. Microinjections of 6-Hydroxydopamine into the PVN
Bilateral stereotaxic microinjections of 6-0HDA-HBr (Regis, Morton
Grove, IL) were made using a Kopf stereotaxic apparatus with a bilateral
cannula constructed of two 26 gauge stainless steel tubes (injectors) with
a 1. 0 mm distance between centers. The cannula was connected with
flexible tubing to two 25 ul Hamilton syringes and a Sage Instruments
syringe pump, model 355. 6-0HDA was dissolved in 0.1% ascorbic acid in
physiological saline, to prevent autooxidation. In addition, the vials
of drug solution were kept on ice and covered with aluminum foil to
prevent light exposure. 6-0HDA was injected bilaterally over a five
minute period, in a concentration of 8.0 µg free base in 1.5 µl saline
containing 0.1% ascorbic acid. Control animals received an injection of
the vehicle (0.1% ascorbic acid in physiological saline). After completion
of the injection, the needle was left in situ for an additional five
minutes to inhibit dorsal diffusion of the neurotoxin along the needle
track. The rats were injected with the monoamine oxidase inhibitor
pargyline ( 30 mg/kg, i. p. ; Sigma, St. Louis, MO) 30 minutes prior to
surgery, to augment the neurotoxic effect of 6-0HDA. The coordinates for
the injection site were 5.7 mm rostral to lambda, 0.5 mm lateral to the
midsagittal suture and 8. 7 mm ventral to the skull surface, with the
incisor bar set at -4.0 mm. The rats were allowed to recuperate from
surgery for 14 days, after which time they were subjected to the CER
paradigm, described below. The rats were sacrificed by decapitation
immediately following the last CER procedure. Trunk blood was collected
and assayed as described below. The extent of damage to noradrenergic
nerve terminals was determined by immunocytochemical techniques (see

81
below). Eight rats were included in the vehicle/control as well as the
vehicle/stress group, while 12 rats per group were used in both the 6-
0HDA/control and 6-0HDA/stress group.
G. Microinjections of Sotalol into the PVN
Rats were placed in a Kopf stereotaxic apparatus and implanted with
chronic bilateral guide cannulae (Plastic Products, Roanoke, VA). The
guide cannulae were constructed of two 26 gauge stainless steel tubes with
a 1.0 mm distance between centers. The inner stylets were constructed of
33 gauge stainless steel wire. The rats were allowed to recuperate from
surgery for 14 days, and were handled each day following the surgery to
condition them for handling during the injection. At the end of the 14
day period, the rats were subjected to the CER procedure, described below.
In this particular study, the rats were transported in their home cage to
the CER room, wrapped in a small towel, and held while the dust cap and
stylets were removed and replaced with a 33 gauge bilateral injector. The
rats received an injection of either vehicle or sotalol, or no injection
at all. All injections were performed approximately three minutes before
placement in the stress chamber, and 13 minutes before sacrifice. The
volume (0.5 µl per side) was injected over a 60 second interval. The
injector remained in situ for an additional 60 seconds to help prevent
dorsal diffusion of the drug. Sotalol was injected bilaterally (0.5 µl
per side), using two 25 ul Hamilton syringes and a CMA microinjection
pump. The concentration of sotalol was 2.0 µg in 0.5 µl of physiological
saline. Vehicle rats received an injection of saline. The non-injected
rats had their dust caps and stylets removed and injectors placed, but no
injection occured. The rats were sacrificed by decapitation immediately

82
following the CER procedure. Trunk blood was collected and assayed as
described below. Fourteen rats were included in the vehicle/control and
vehicle/stress groups. The non-injected/control and non-injected/stress
groups had six rats each. Sixteen rats were included in the sotalol/-
control and eighteen rats were in the sotalol/stress group.
H. Histology
Lesion placement and drug injection sites were verified histologi
cally. The brains were preserved in buffered formalin for 48 hours. The
brains were then infiltrated with sucrose by successive four hour
intervals in 5% and 10% sucrose solutions, respectively. The brains were
stored in a 20% sucrose solution until they were cut on a cryostat (40
micron sections) and stained by the cresyl violet method and inspected by
a microscope to verify the position of the tip of the needle or extent of
lesion damage.
I. Immunocytochemistry
The brains from the 6-0HDA-treated rats were fixed for five hours
in a solution containing 4% paraformaldehyde-0.05% glutaraldehyde in 0.10
M phosphate buffer, pH 7.5 at 4°C. They were then transferred to a 15%
sucrose PBS solution where they were stored overnight at 4°C (Furness et
al., 1978). The brains were then mounted in a cryostat, frozen and cut
into 40 micron sections. These sections were floated in 0.05 M PBS and
mounted on slides and allowed to air dry. The slides were then placed in
air-tight slide boxes and kept at 0°C until processed (approximately 48
hrs). Sections were then washed and incubated in immunocytochemistry

83
(ICC) buffer consisting of phosphosaline buffer (PBS), 1.0% normal goat
serum (from second antibody host species), 0.1% gelatin, 0.01% thimerosal,
O - 0.002% neomycin, and 0 - 0.2% triton X-100. Sections were incubated
consecutively in the primary antibody, rabbit anti-bovine adrenal
medullary dopamine ~-hydroxylase (DBH, 1:1000 dilution) for 12 hours in
humidity-controlled chambers at 4°C. The DBH antibody was obtained from
two sources: (1) Eugene-Tech, Allendale, NJ and (2) Dr. Eric Schweitzer,
Madison, WI. The two DBH antibodies gave nearly identical results when
tested on control rats. This incubation was followed by addition of a
1:50 dilution of goat anti-rabbit IgG followed by an incubation with 1:200
peroxidase-antiperoxidase (rabbit PAP) solution for 30 minutes at room
temperature. Sections were washed between each immune incubation with
0.02 M PBS. Bound peroxidase was visualized by incubating sections in 25
mg/100 ml diaminobenzidine tetrahydrochloride (DAB-4HC1) and 0.01%
hydrogen peroxide for 5-7 minutes. To increase sensitivity this solution
also included 2.5% nickel ammonium sulfate. Sections were washed, mounted
with cover slips, and viewed on a Leitz Ortholux photomicroscope.
J. Stress (GER) Procedure
In each of the experiments, the rats had at least a two-week post
operative recovery period before being subjected to the conditioned
emotional response (GER) paradigm. The GER was performed in a ten-inch
square, clear Plexiglass chamber with a grid floor composed of stainless
steel rods (0.3 inches in diameter) spaced 0.5 inches apart. The chamber
was illuminated by a 7.5 W incandescent lamp. The chamber was located
inside a larger, rectangular sound-attenuating box equipped with a two-

84
way mirror (13" x 9") so that the rat's behavior could be observed. The
sound-proof box also had a small fan in the back wall for ventilation.
Scrambled current (0.8 mA D.C.) shock for 10 seconds was delivered through
the grid floor by a Grayson-Stadler shock generator.
The rats were assigned at random before testing to a particular
treatment group (control or stress) such that cage mates were members of
the same group. Cage mates were tested sequentially and at the same time
each day (between 10:00 and 15:00 hours) on four consecutive days.
On each day, the rats were transported ten feet from their living
quarters to the stress chamber. Ten minutes following placement in the
chamber, the experimental animals received an inescapable foot shock, then
were returned to their home cage. This procedure was repeated for three
consecutive days. The behavior of the rats was observed on days 3 and 4
of the CER paradigm. It was apparent from these observations that the
stressed rats had learned that their placement in the chamber would be
followed by a foot shock. By the third day, in contrast to the control
animals, the stressed rats defecated, urinated and demonstrated freezing
(no movement) behavior. On the fourth day, after 10 minutes in the test
chamber, instead of receiving shock the rats were removed, transported to
a third area outside the stress room and immediately sacrificed by
decapitation. The control rats were treated identically except that shock
was not administered at any time.
The stress chambers were cleaned carefully after each rat had been
tested. Fecal boli and urine were removed, as well as washing the grid
floor and sides of the plexiglass chamber. This was to prevent the
analgesic effect described by Fanselow (1985) that can occur when an

85
unstressed rat is placed in a chamber containing the odor of a rat that
has received an electric shock. Rats have been shown to release an
innately recognized odor in response to many types of aversive events
(Brown, 1979). In particular, aversive electric shock elicits an odor
that conspecifics can discriminate from the odors of unstressed rats
(Valenta and Rigby, 1968). These odors have been shown to interfere with
the acquisition of one-way shock avoidance (Dua and Dobson, 1974).
K. Biochemical Determinations
1. Plasma Renin Activity (PRA)
PRA was measured by radioimmunoassay for generated angiotensin I
(Haber et al., 1969; Stockigt et al., 1971). The pH of the samples was
reduced to pH 6.5 by addition of 0.5 ml of 0.5 M phosphate buffer pH 6.0.
The conversion of angiotensin I to angiotensin II was prevented by
addition of 0.025 ml of 8-hydroxyquinoline (8-HQ; 10% suspension in 0.3
M EDTA; 3.4 mM final concentration; Mallinckrodt, St. Louis, MO) and 0.020
ml of phenylmethylsulfonyl fluoride (PMSF; 5 g in 150 ml ethanol; 2.5 mM
final concentration; Sigma, St. Louis, MO). Plasma samples were incubated
for three hours at 37°C to generate angiotensin I (AI). The incubation was
stopped by immersion of the samples in boiling water for three minutes.
The radioimmunoassay of AI was performed with an antiserum against AI at
a dilution of 1:30,000, and a total binding of 35%. The antibody was
added to the samples in a volume of 0.3 ml (AI antiserum in a 0.1 M Tris
buffer, pH 8.0, containing 0.1% gelatin) and incubated overnight at 4°c.
The tracer ( 125I labelled AI) was added to the samples the following
morning, in a volume of 0.4 ml per tube, approximately 10,000 counts per

86
minute per tube. On the next day, the antibody-bound (iodinated AI)
tracer was separated from the free tracer by addition of O. 5 ml of a
suspension containing activated charcoal and dextran T500 (Pharmacia,
Piscataway, NJ) and centrifugation at 2000 x g for 15 minutes. The pellet
containing the unbound tracer was counted on a micromedic 4/200 plus
automatic gamma counter which was connected via a buffer (Quadram
microfazer) to an IBM PC-AT computer. The data were analyzed by a program
(RIA-AID) that was purchased from Robert Maciel Associates (Arlington,
MA). The sensitivity limit of the RIA is 10 pg AI per tube and the intra
assay variability was 4.0%. The inter-assay variability was 7.4% (Van de
Kar et al., 1984). The tracer (AI; Beckman, Arlington Heights, IL) was
iodinated via the chloramine T (Sigma, St. Louis, MO) method (Greenwood
et al., 1963). Phosphate buffer (0.05 M; 0.02 ml), 12 µl AI (0.5 µg/µl)
and 20 µl chloramine T (3.5 mg/ml phosphate buffer) were added to the vial
containing the 125I and mixed for three seconds. This was immediately
followed by addition of 25 µl of sodium metabisulfite (4. 5 mg/ml phosphate
buffer; J. T. Baker Chemical Co. , Phillipsburg, NJ) to terminate the
reaction. Chromatography of the iodinated AI was performed first by ion
exchange (Biorad AG-1X4, 200-400 mesh) chromatography. The column
consisted of a siliconized Pasteur pipette (5 3/4"; Scientific Products)
with a small amount of glass wool placed in the tip of the pipette to
support the anion exchange res in. Fractions (12 drops per tube) were
collected from the column and the radioactivity in each tube was measured.
The radioactive peak fractions eluted off this column were then placed on
a Sephadex G-15 column for gel filtration and eluted with 0.05 N acetic
acid containing 0.1% bovine serum albumin (Sigma, St. Louis, MO). Two ml

87
fractions were collected off the Sephadex column and measured for
radioactivity. The tubes containing the radioactive AI were pooled and
stored at -10°c.
2. Plasma Renin Concentration (PRC)
Renin is the enzyme responsible for the conversion of the
substrate angiotensinogen, produced by the liver, to
Angiotensin I. The measurement of plasma renin activity reflects the
concentration of renin as well as renin substrate (angiotensinogen) in
plasma. The normal angiotensinogen concentration is only about or less
than Km (Reid et al., 1978) and thus much less than the concentration
required to generate AI at maximum velocity (Vmax). Studies by Van de
Kar et al. (1981) have demonstrated that 5-HT dependent changes in plasma
renin activity are not accompanied by changes in plasma angiotensinogen
levels. However, since electrolytic lesions in the PVN could cause a
reduction in corticosterone levels, this could lead to decreased secretion
of angiotensinogen from the liver. This would cause plasma renin activity
values to be low even when renin levels were unaffected. The assay for
plasma renin concentration involves the addition of a saturating
concentration of exogenous renin substrate to the samples such that renin
is working at Vmax. The measurement of plasma renin concentration
eliminates the possibility that decreased substrate levels would be
misinterpreted as decreased levels of renin.
The renin substrate was plasma from nephrectomized rats which
received dexamethasone (0.2 mg/rat) 24 hours before sacrifice. The plasma
samples (0.2 ml) received 0.1 ml of nephrectomized plasma, 0.1 ml of 0.5

88
M phosphate buffer pH 6.0, and 5 µl each of PMSF and 8-hydroxyquinoline
(to a final concentration of 2.5 mM and 3.4 mM, respectively). The
mixture was incubated at 37°C for one hour. The reaction was stopped by
immersion of the tubes in a boiling water bath for three minutes. The
radioimmunoassay of AI is as described for PRA.
3. Plasma Angiotensinogen
This radioimmunoassay is similar to that for PRC except that a
saturating amount of the enzyme renin (from kidney homogenates) is added
to the plasma samples before incubation to allow conversion of endogenous
renin substrate to AI (Menard and Catt, 1972). Therefore any differences
in the production of AI will be attributed to differences in substrate,
not enzyme levels.
The plasma samples (0.1 ml) received 0.1 ml of 0.5 M phosphate
buffer pH 6.0, 0.1 ml renin (from rat kidneys homogenized in cold water
1 g/ml at a 1:1,000 dilution), and 5 µl each of PMSF and 8-HQ. The
mixture was incubated at 37°C for one hour. The reaction was stopped by
immersion of the tubes in a boiling water bath for three minutes. The
remainder of the radioimmunoassay for AI is as described for PRA.
4. Plasma Corticosterone
Corticosterone radioimmunoassays on unextracted plasma (5 and 10
µl) were performed using procedures and antiserum from Radioassay Systems
Laboratories (Carson, CA). The binding proteins were denatured by
incubation of the sample tubes and assay buffer in a hot water (80°C) bath
for twenty minutes. The standards were made of serial dilutions of

89
corticosterone (Steraloids, Wilton, NH) in assay buffer. The antiserum
(catalog # 1472, lot # 3R3-38) was used at a final dilution of 1:10,500
and total binding of 40%. The incubation of the samples (or standard)
with the antiserum and tracer (corticosterone l,2,6,7- 3H(N)-; NET-399,
New England Nuclear) occurred overnight at 4°C. The antibody-bound tracer
was separated from the unbound tracer by addition of a charcoal-Dextran
T70 suspension, centrifugation, and decanting of the supernatant into
scintillation vials containing 5 ml of a scintillation cocktail (4 g of
Omnifluor in 1 liter of toluene). The samples containing the supernatant
and scintillation cocktail were counted in a Packard liquid scintillation
counter. The data were stored on a diskette by a Diskstore (United
Technologies, Packard) and were analyzed using the RIA-AID software
(Robert Maciel Associates, Arlington, MA). The intra- and inter-assay
variability were 4.5% and 11.9% respectively (Van de Kar et al., 1985).
L. Statistics
The data are reported as mean ± S. E .M. (standard error of the
mean). The sample mean was the average of the number of animals in each
group (n). The standard error of the mean was calculated as described
below. Statistical analyses of the data was performed by a two-way
analysis of variance (ANOVA). Individual group means were compared by
Student Newman-Keul's multiple range test (Steel and Torrie, 1960).
The most common measurement of central tendency for given samples
of a population is the arithmetic average or mean. In addition to the
measure of central tendency, it is important to be aware of how the
samples differ in variability. This measure is the variance, or its

90
square root, the standard deviation (S.D.). This variance will provide
information as to whether an observation is an ordinary or an unusual
value in a specified population, giving a small value when the observa
tions cluster closely about a central value, and a larger value when they
are spaced widely. The sum of squares is the sum of the squared
deviations from the arithmetic mean. The quantity (n-1) is known as
degrees of freedom. Sum of squares is divided by degrees of freedom to
give unbiased estimates of the population variances. Since sample means
are also subject to variation, the standard error of the mean is also
reported. It is calculated by dividing the S.D. by the square root of the
number of observations in the mean.
1. Analysis of Variance (ANOVA)
Analysis of variance is defined by Steel and Torrie (1960) as
" ... an arithmetic process for partitioning a total sum of squares into
components associated with recognized sources of variation." Using this
type of test, the data are classified according to their treatment group.
With respect to the present studies, a two-way analysis of variance was
the appropriate test, since the animals were subjected to two separate
treatments: (1) surgical (or drug) and (2) behavioral (stress).
The error mean square is an average of the components contributed
by the treatments. It is an estimate of a common variation among
observations of the same treatment group. Treatment mean square is an
independent estimate of the variance when the null hypothessis is true.
F is defined as the ratio of two independent estimates of the same
variance, thus, the F value is obtained by dividing the treatment mean

91
square by the error mean square. Once F has been calculated, it is
compared with the tabular F at the appropriate degrees of freedom at the
.05 (5%) probability level. If the calculated F is greater than the
tabulated F, then the null hypothesis (no difference between population
means) can be rejected and it can be concluded that there are real
differences among treatment means. With respect to the two-way ANOVAS of
the present studies, three F values were obtained: Fa for the surgical or
drug treatment, Fb for the stress treatment, and Fa x b for the interaction
between the drug and stress treatments. The F value reported was the one
that accounts for interaction between both the stress and surgical/drug
treatment groups.
2. Student-Newman-Keuls' Test
Multiple comparison tests can be used to compare each treatment
mean with every other treatment mean. Student-Newman-Keuls' test is a
multiple range test " ... for judging the significance of a set of
differences; it permits decisions as to which differences are significant
and which are not ... Error rate is seen to apply neither on an experiment
wise nor a per-comparison basis since a particular range test may be made
for one or more comparisons in a particular experiment" (Steel and Torrie,
1960). This test also takes into account the number of treatments in the
experiment. To perform the Newman-Keuls' test, the mean square error from
the ANOVA is divided by the harmonic mean (average number of n per group).
The square root of this number is Sx, the standard error of the mean. Next
a tabular value, q, is selected. This value is based on a .05 level of
probability, and the appropriate p (number of treatment means) and degrees

92
of freedom. Q is multiplied by Sx to give a range of significance. If the
distance between two means is greater than the range of significance, then
the difference between the two means is significant at the 0.05 probabil
ity level.

CHAPTER IV
RESULTS
A. Electrolytic Lesions in the PVN
Figure 1 is a coronal section of an intact PVN, stained with cresyl
violet. Figure 2 shows an electrolytic lesion in the PVN. These lesions
did not extend caudally beyond the PVN, and the dorsomedial nucleus was
intact. There was some damage to the caudal aspect of the anterior
hypothalamic area, as in Figure 2. This was not evident in all cases.
The lesions did not extend laterally to the fornix. Only lesions which
destroyed 90% or more of the PVN and did not extend dorsally into the
thalamus were considered in the statistical evaluation of hormonal data.
Following histological verification, the lesion control/group consisted
of five rats, while the lesion/stress group consisted of seven rats with
accurate and complete PVN lesions. One rat did not survive the anesthesia
in the sham/control group, making the total for this group seven rats.
Electrolytic lesions in the PVN prevented the stress-induced
increases in plasma renin activity (PRA) (Figure 3), plasma renin
concentration (PRC) (Figure 4), and in plasma corticosterone levels
(Figure 5). By comparison, rats with lesions which missed the PVN (n=S),
but destroyed areas rostral to the PVN, showed no inhibition of the effect
of stress on PRA (24.0 ± 3.2), PRC (33.7 ± 6.2) and corticosterone (29.3
± 6.1) levels.
93

FIGURE 1
CORONAL SECTION OF RAT BRAIN WITH AN INTACT PARAVENTRICULAR NUCLEUS
FIGURE 2
CORONAL SECTION OF RAT BRAIN WITH AN ELECTROLYTIC LESION IN THE PARAVENTRICULAR NUCLEUS
94

~ > 20 ~ /"\
L
0 £ <( I")
z """15 z E w """ 0::: <( 10 <( 01 2 c (/) '-.../
<( 5 _J o_
SHAM LESION
FIGURE 3
* D CONTROL ~ STRESS
ELECTROLYTIC PVN LESION
95
The effect of stress on plasma renin activity in rats with electrolytic PVN lesions. The data represent mean± S.E.M., n=7 for the sham/control group, n=6 for the sham/stress group, n=5 for the lesion/control group, and n=7 for the lesion/stress group.
* Significant difference from the corresponding control group, p < 0.05. A minimal difference of 5.7 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of the lesion F(l, 21) -10.789; stress F(l,21) = 14.617; and lesion*stress interaction F(l,21) -9. 031.

96
FIGURE 4
z 30 0 D CONTROL f-
25 * <( ~STRESS (l'.'. f-z "' w L 20 u _c
z "" -0 ~ 15 u
z <(
z ()> w c 10 (l'.'. \.._,./
<( 2 5 (/)
:5 0... 0
SHAM ELECTROLYTIC LESION PVN LESION
The effect of stress on plasma renin concentration in rats with electrolytic PVN lesions. The data represent mean± S.E.M., n=7 for the sham/control group, n=6 for the sham/stress group, n=5 for the lesion/control group, and n=7 for the lesion/stress group.
* Significant difference from the corresponding control group, p < 0.05. A minimal difference of 4.5 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a highly significant effect of the lesion F(l,21) = 16.713; stress F(l,21) = 26.858; and lesion*stress interaction F(l,21) - 17.588.
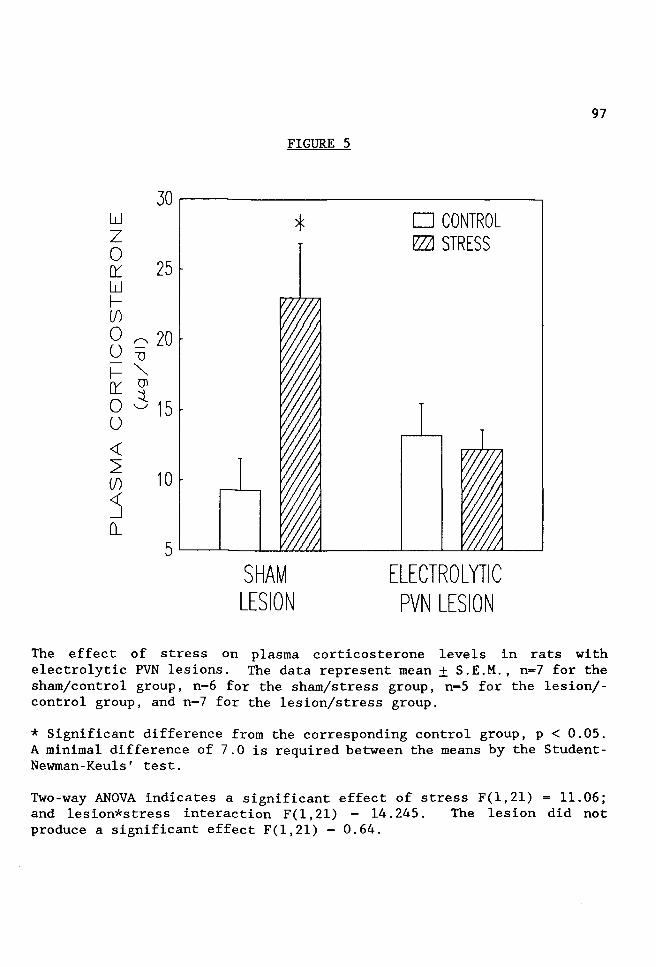
w z 0 ()'.'. 25 w f---(j)
0 """' 20 u 1J
- "' f--- Ol ()'.'. ::t 0 'J 15 u <(
~ 10
s o_
FIGURE 5
SHAM LESION
* D CONTROL ~ STRESS
ELECTROLYTIC PVN LESION
97
The effect of stress on plasma corticosterone levels in rats with electrolytic PVN lesions. The data represent mean± S.E.M., n=7 for the sham/control group, n=6 for the sham/stress group, n=5 for the lesion/control group, and n=7 for the lesion/stress group.
* Significant difference from the corresponding control group, p < 0.05. A minimal difference of 7.0 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of stress F(l,21) = 11.06; and lesion*stress interaction F(l, 21) = 14. 245. The lesion did not produce a significant effect F(l,21) = 0.64.

98
B. Electrolyic Lesions in the Nucleus Reuniens
In a control experiment, lesions in the thalamic nucleus reuniens
were made, dorsal and caudal to the PVN. These lesions were small,
discrete and did not extend to the region of the third ventricle.and the
PVN (Figure 6). Following histological verification of placement in the
nucleus reuniens, five rats were included in the lesion/control and six
rats included in the lesion/stress group. There were six rats in the
sham/stress group, and eight rats in the sham/control group.
Electrolytic lesions in the nucleus reuniens did not prevent the
stress-induced increase in plasma renin activity (Figure 7) or plasma
renin concentration (Figure 8). The stress-induced increase in plasma
corticosterone levels was attenuated but not completely blocked by nucleus
reuniens lesions (Figure 9). It is possible that this lesion interrupted
afferent fibers to the PVN that are involved in stimulating CRF neurons
in the PVN. Stressed rats in which the lesions missed both the nucleus
reuniens and the PVN (n=S), had hormonal values similar to the sham/stress
group and the group in which the nucleus reuniens was destroyed (PRA 14.3
± 3.6; PRC 13.5 ± 1.8; and corticosterone 18.2 ± 1.7).

FIGURE 6
CORONAL SECTION OF RAT BRAIN WITH AN ELECTROLYTIC LESION IN THE THALAMIC NUCLEUS REUNIENS
99

>f-> 20 f- ~
L u _c
<( n z ""' 15 z E w ""' 0::: <( 10 <( O'> 2 c ~ ~ 5 _J
Q_
FIGURE 7
D CONTROL
~ STRESS
SHAM LESION
*
ELECTROLYTIC LESION NUCLEUS
REUNIENS
100
The effect of stress on plasma renin activity in rats with electrolytic lesions in the nucleus reuniens. The data represent mean± S.E.M., n-8 for the sham/control group, n=6 for the sham/stress group, n=5 for the lesion/control group and n=6 for the lesion/stress group.
* Significant difference from the corresponding control group, p < 0.05. A minimal difference of 6.4 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of stress F(l,21) = 11.58; but not of the lesion F(l,21) = 0.761 or lesion*stress interaction F(l,21) - 0.92.

z 0 I-<{ ~ 1-z~ W L
20
~ ~ 15 o E u """ z <{
z w 0:::
<{ 2 (/)
:5 CL
(J>
c 10
5
SHAM LESION
FIGURE 8
* D CONTROL
~STRESS
ELECTROLYTIC LESION NUCLEUS
REUNIENS
101
The effect of stress on plasma renin concentration in rats with electrolytic lesions in the nucleus reuniens. The data represent mean± S.E.M., n=8 for the sham/control group, n=6 for the sham/stress group, n=S for the lesion/control group and n=6 for the lesion/stress group.
* Significant difference from the corresponding control group, p < 0.05. A minimal difference of 7.4 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of stress F(l,21) = 10.291; but not of the lesion F(l,21) - 0.042 or lesion*stress interaction F(l,21) = 1.109.

w z 0 0::: 25 w 1-(f)
0 ~ 20 u ""O
I- "" 0::: (Jl
0 3 15 u <t
~ 10
s Q_
SHAM LESION
FIGURE 9
* D CONTROL r'Z2l STRESS
ELECTROLYTIC LESION NUCLEUS
REUNIENS
102
The effect of stress on plasma corticosterone levels in rats with electrolytic lesions in the nucleus reuniens. The data represent mean± S.E.M., n=7 for the sham/control group, n=6 for the sham/stress group, n=5 for the lesion/control group and n=6 for the lesion/stress group.
* Significant difference from the corresponding control group, p < 0.05. A minimal difference of 6.5 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of stress F(l,20) - 10.29; but not of the lesion F(l,20) = 0.601 or lesion*stress interaction F(l,20) = 1. 83.

103
c. Injection of Ibotenic Acid into the PVN
Figure 10 is a section of an intact PVN stained with cresyl violet.
Figure 11 shows the needle tracks and destruction of neurons in the PVN
following an injection of ibotenic acid. Acceptable lesions were
restricted to the PVN and, compared with control sections, showed greater
than 50% cell loss. Following histological verification of the injection
site and evaluation of the hormonal data, there were seven rats included
in both the vehicle/control and vehicle/stress group. Four rats in the
ibotenic acid/control and six rats in the ibotenic acid/stress group were
considered accurate PVN lesions.
Ibotenic acid-induced lesions in the PVN prevented the stress
induced increase in plasma renin activity (Figure 12) and plasma renin
concentration (Figure 13). Stress-induced increases in corticosterone
levels also were prevented by ibotenic acid lesions of neurons in the PVN
(Figure 14). By comparison, stressed rats with ibotenic acid injections
which missed or caused unnoticeable damage to neurons in the PVN (n=6) had
PRA (28.9 ± 3.8), PRC (35.5 ± 4.5) and corticosterone levels (16.9 ± 1.9)
which were indistinguishable from the vehicle/stress group. Thus far, the
data suggested that cell bodies in the PVN are involved in stress-induced
renin and corticosterone secretion.
104
FIGURE 10
INTACT PARAVENTRICULAR NUCLEUS

PARAVENTRICULAR NUCLEUS FOLLOWING BILATERAL INJECTION OF IBOTENIC ACID
105

~ 35 > - /'"""'\
0 ~ 30 <{ 6 z ..c 25 - t')
Q"' o::: E 20
<{ "' 2 <{ 15 (f) 0\ <{ c _J '-"' Q_ 10
*
VEHICLE
FIGURE 12
D CONTROL fZZJ STRESS
IBOTENIC ACID
106
The effect of stress on plasma renin activity in rats with ibotenic acid lesions in the PVN. The data represent mean ± S. E .M., n=7 for both vehicle groups, n=4 for the ibotenic acid/control group, and n=6 for the ibotenic acid/stress group.
* Significant difference from the corresponding control group, p < 0.05. A minimal difference of 10.5 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of the drug F(l,20) = 4.959; stress F(l,20) = 9.488; and drug*stress interaction F(l,20) = 6.063.
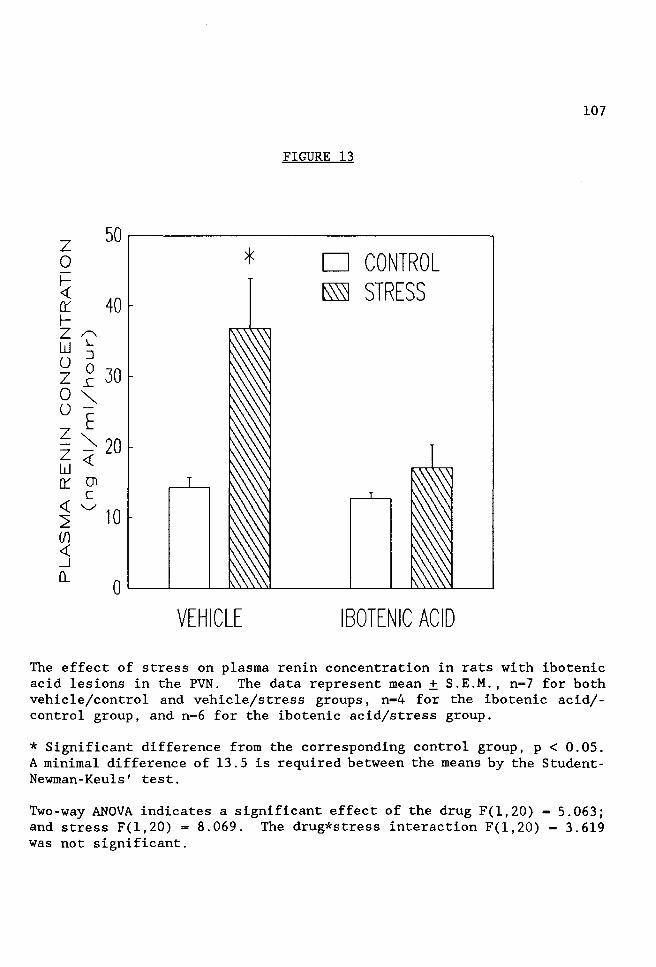
z 0
~ er 1-z /"-w L
40
u :J
z ~ 30
0 "" u -E
~ "" 20 w <( er (JI
c ~ '-/ 10 (/) <( _J
o_
*
VEHICLE
FIGURE 13
D CONTROL ~ STRESS
IBOTENIC ACID
107
The effect of stress on plasma renin concentration in rats with ibotenic acid lesions in the PVN. The data represent mean± S.E.M., n=7 for both vehicle/control and vehicle/stress groups, n=4 for the ibotenic acid/control group, and n=6 for the ibotenic acid/stress group.
* Significant difference from the corresponding control group, p < 0.05. A minimal difference of 13.5 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of the drug F(l,20) = 5.063; and stress F(l,20) = 8.069. The drug*stress interaction F(l,20) - 3.619 was not significant.

w z 0 tr 15 w 1-(/) o; u " I- Ol 10 i:r ::i. 0 ....._.,, u <(
2 (/) <( _J
()_
5
*
VEHICLE
FIGURE 14
D CONTROL f7ZJ STRESS
IBOTENIC ACID
108
The effect of stress on plasma corticosterone levels in rats with ibotenic acid lesions in the PVN. The data represent mean± S.E.M., n=7 for both vehicle/control and vehicle/stress, n=4 for the ibotenic acid/control group, and n=6 for the ibotenic acid/stress group.
* Significant difference from the corresponding control group, p < 0.05. A minimal difference of 4.3 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of the drug F(l,21) = 12.273; and stress F(l, 21) 19. 645. The drug*stress interaction was not significant F(l,21) = 3.198.

109
o. Injection of 6-0HDA into the PVN
Figure 15A shows a coronal section of the PVN following vehicle
injection and staining with dopamine ~-hydroxylase (DBH) antiserum.
Figure 15B shows a coronal section of the PVN following 6-0HDA injection
and staining with DBH antiserum, showing loss of DBH immunoreactivity.
In general, there was a 60-80% reduction in DBH staining (as compared to
vehicle rats) in a 500-1000 micron area that encompassed the PVN and
anterior hypothalamus, but did not spread as far as the mediobasal
hypothalamus. Following immunocytochemical and histological verification,
the vehicle/control and 6-0HDA/control groups consisted of seven rats
each, the vehicle/stress group consisted of six rats, and the 6-0H
DA/stress group had five rats.
The stress (GER) procedure produced a significant increase in plasma
renin acivity and concentration as well as increasing plasma corticos
terone levels in rats which received an injection of vehicle or non
injected animals. Injection of 6-0HDA into the PVN prevented the effect
of stress on plasma renin activity (Figure 16) and plasma renin concentra-
ti on (Figure 17) . In addition the effect of stress on plasma cor-
ticosterone levels was also blocked (Figure 18). Since 6-0HDA injections
into the PVN prevented the stress - induced increase in corticosterone
levels, it is possible that this caused decreased secretion of the renin
substrate (angiotensinogen) from the liver. This would cause PRA values
to be low even though renin levels were unaffected. To test this, in

110
addition to measurements of PRC, plasma angiotensinogen levels were
determined (Figure 19). There were no differences observed between the
groups. This suggested that neither stress nor 6-0HDA injections affected
plasma levels of renin substrate. Thus far the data suggest that
catecholaminergic neurons or nerve terminals in the PVN play a role in the
stress-induced increase in renin and corticosterone secretion.

FIGURE lSA
CORONAL SECTION FROM RAT INJECTED WITH VEHICLE AND STAINED WITH DBH ANTISERUM
CORONAL SECTION FROM RAT INJECTED WITH 6-0HDA AND STAINED WITH DBH ANTISERUM,
SHOWING LOSS OF DBH IMMUNOREACTIVITY
111

>I-> - /'""'\ I- Cf)
25
0 ~ 20 <( 0 z _c
z I") 15 w"" o: E <( "" 10 2 <t (f) Q)
<( c 5 _J '-.,,/
Q_
*
VEHICLE
FIGURE 16
D CONTROL ~ STRESS
6-0HDA
112
The effect of stress on plasma renin activity in rats with 6-0HDA lesions. The data represent mean ± S.E.M.; n=7 for vehicle/control, n=6 for vehicle/stress, n=7 for 6-0HDA/control and n=5 for 6-0HDA/stress.
*Significant difference from the corresponding control group, p < 0.05. A minimal difference of 5.48 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of the drug F(l,21) = 13.576; stress F(l,21) = 35.595; and drug*stress interaction F(l,21) = 12.427.
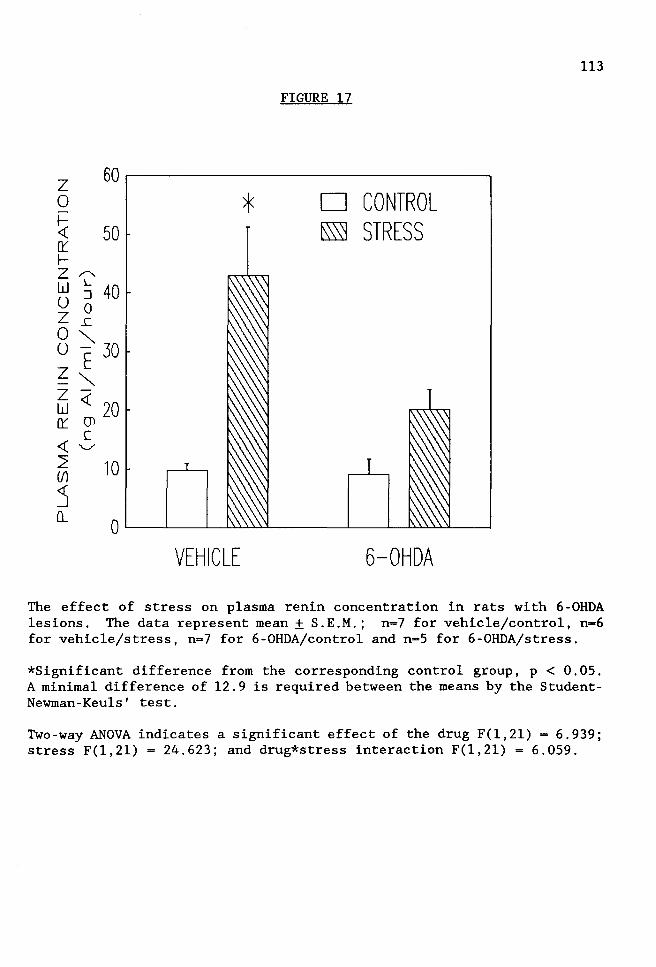
z 0
~ O'.'. f-
50
z /"w L
0 g 40 z .r::
8 ""' 30 z E - ""' z -w <{ 20 O'.'. (Jl
<{ ~ 2 (/)
s 0...
10
*
VEHICLE
FIGURE 17
D CONTROL ~ STRESS
6-0HDA
113
The effect of stress on plasma renin concentration in rats with 6-0HDA lesions. The data represent mean± S.E.M.; n=7 for vehicle/control, n-6 for vehicle/stress, n=7 for 6-0HDA/control and n-5 for 6-0HDA/stress.
*Significant difference from the corresponding control group, p < 0.05. A minimal difference of 12.9 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of the drug F(l,21) = 6.939; stress F(l,21) = 24.623; and drug*stress interaction F(l,21) = 6.059.

~ 30 0 O'.'. w 25 I-(/)
0 -; 20 u "" I- Ol
~ 3 15 u
~ 10 (/) <t: _J 5 D...
*
VEHICLE
FIGURE 18
D CONTROL f7ZJ STRESS
6-0HDA
114
The effect of stress on plasma corticosterone levels in rats with 6-0HDA lesions. The data represent mean± S.E.M.; n-7 for vehicle/control, n-6 for vehicle/stress, n=7 for 6-0HDA/control and n-5 for 6-0HDA/stress.
*Significant difference from the corresponding control group, p < 0.05. A minimal difference of 10.0 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of stress F(l,21) = 9.422. There were no significant effects of either the drug F(l,21) = 0.194 or drug*stress interaction F(l,21) = 3.371.

FIGURE 19
120 ~----------~
z w (.'.)
0 z,,.......
100
(/) ~ 80 z"' w-1- E 0 .-- 60 (.'.) . zo <{ ci; <{ 5 40 2 (/) s 20 Q_
VEHICLE
D CONTROL f7ZJ STRESS
6-0HDA
115
The effect of stress on plasma angiotensinogen levels in rats with 6-0HDA lesions. The data represent mean± S.E.M.; n-7 for vehicle/control, n=6 for vehicle/stress, n=7 for 6-0HDA/control and n=5 for 6-0HDA/stress.
No significant differences found among the groups, p < 0.05. A minimal difference of 33.l would be required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates no significant effects of the drug F(l, 21) -1. 395; stress F(l, 21) = 0. 072; and drug*stress interaction F(l, 21) -0. 721.

116
E. Injection of Sotalol into the PVN
Figure 20 shows a coronal section of the PVN stained with cresyl
violet following a bilateral injection of sotalol into the PVN. Only rats
in which the tip of the cannula was immediately dorsal to the border of
the PVN, or inside the dorsal PVN, were acceptable. In several rats, the
cannulae ended inside the PVN, producing large mechanical lesions. These
rats were excluded from the statistical analysis of the hormonal data.
Following histological verification of the injection site, eight rats were
used in the vehicle/control, five rats in vehicle/stress and non
injected/stress, and four rats in the non-injected/control group. There
were six rats used in the sotalol/control group and eight rats in the
sotalol/stress group.
Figures 21 and 22 show the effect of stress on plasma renin activity
and plasma renin concentration, respectively, in rats injected with either
vehicle or sotalol, or in non-injected rats. Stress-induced increases in
PRA and PRC were not prevented by injection of sotalol. However, the
stress-induced increase in corticosterone levels was prevented by
injections of sotalol into the PVN (Figure 23).

FIGURE 20
CORONAL SECTION OF THE PARAVENTRICULAR NUCLEUS FOLLOWING BILATERAL INJECTION OF SOTALOL
117

>~
> ~,....._ OL <( .c
z~ ZE w"" (t <(
JO
25
20
15 <( Ol
25 10 (/) <( _J 5 ()_
*
VEHICLE
FIGURE 21
D CONTROL ~STRESS
*
SOTALOL
118
*
UNINJECTED
The effect of stress on plasma renin activity in rats injected with either vehicle or sotalol, or in non-injected rats. The data represent mean± S.E.M.; n=8 for vehicle/control, n=4 for vehicle/stress, n=6 for sotalol/control, n=7 for sotalol/stress, n=4 for non-injected/control and n=5 for non-injected/stress.
*Significant difference from the corresponding control group, p < 0.05. A minimal difference of 8.42 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of stress F(l,28) - 32.526; but not of the drug F(2,28) = 0.871 or drug*stress interaction F(2,28) -0.528.
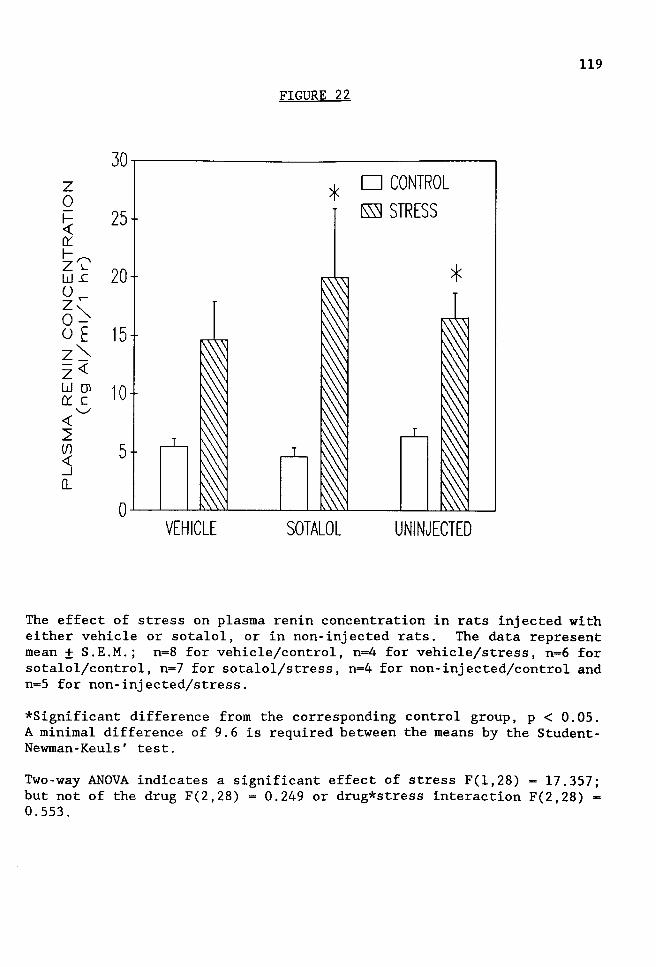
z 0 ~ 25 O'.: f-,........
~ ~ 20 0 z r-0~ o E 15 z"'--<( z WOl 10 O'.: c <( 2
...._,,,,
(f) 5 ~ o_
FIGURE 22
*
VEHICLE SOTALOL
D CONTROL ~ STRESS
*
UNINJECTED
119
The effect of stress on plasma renin concentration in rats injected with either vehicle or sotalol, or in non-injected rats. The data represent mean± S.E.M.; n=8 for vehicle/control, n=4 for vehicle/stress, n=6 for sotalol/control, n=7 for sotalol/stress, n=4 for non-injected/control and n=5 for non-injected/stress.
*Significant difference from the corresponding control group, p < 0.05. A minimal difference of 9.6 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of stress F(l,28) - 17.357; but not of the drug F(2,28) = 0.249 or drug*stress interaction F(2,28) -0.553.
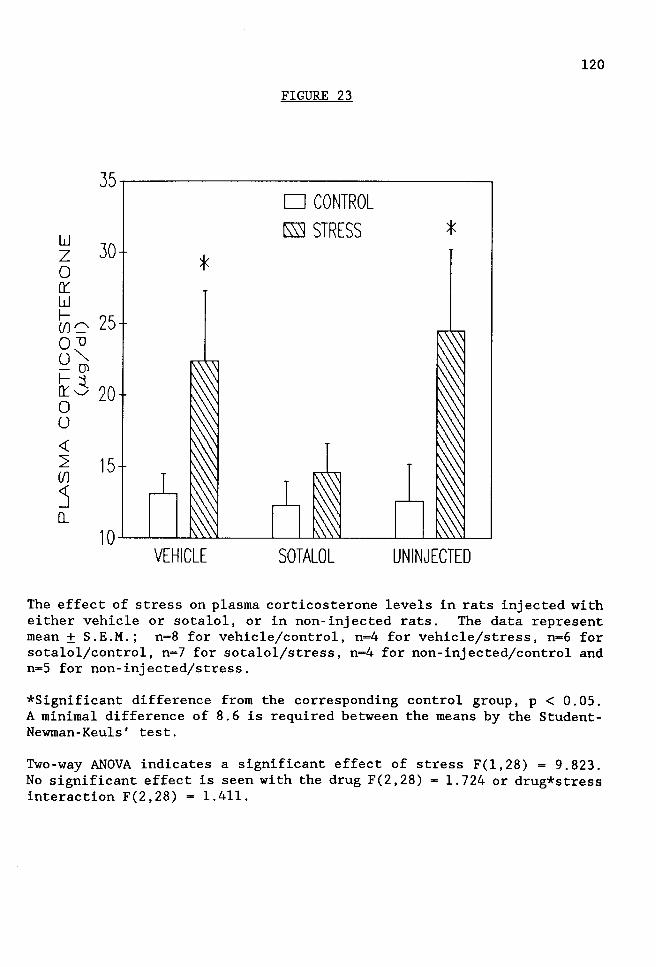
FIGURE 23
35....---------------,
~ 30 0 O'.'. w I-(/) 0 25 Q""CJ u"'-- (])
~3 20 0 u <{
2 15 (/)
s 0...
*
VEHICLE
D CONTROL
~STRESS
SOTALOL
*
UN INJECTED
120
The effect of stress on plasma corticosterone levels in rats injected with either vehicle or sotalol, or in non-injected rats. The data represent mean± S.E.M.; n=8 for vehicle/control, n=4 for vehicle/stress, n-6 for sotalol/control, n=7 for sotalol/stress, n=4 for non-injected/control and n=S for non-injected/stress.
*Significant difference from the corresponding control group, p < 0.05. A minimal difference of 8.6 is required between the means by the StudentNewman-Keuls' test.
Two-way ANOVA indicates a significant effect of stress F(l,28) = 9.823. No significant effect is seen with the drug F(2,28) - 1.724 or drug*stress interaction F(2,28) = 1.411.

121
F. Comparison of Normal Plasma Pools to Experimental Controls
Pooled plasma samples for "normal" and "high" PRA or "normal" and
"high" corticosterone levels are run routinely with each respective assay.
The pooled plasma samples are compiled from rat plasma that has been
previously assayed and classified within the range of "normal" or "high"
based on its corticosterone or PRA value. A random sample of these
values, taken from assays run between 1986 and 1989, gave the following
values (mean± standard error of the mean):
"normal" PRA 7.6 ± 0.5 (n 23) "high" PRA 23.6 ± 2.8 (n 23)
"normal" corticosterone 8.9 ± 1.1 (n 13) "high" corticosterone 20.9 ± 2.0 (n 10)
The pooled values for "normal" PRA and corticosterone are very
similar to previously published values for PRA and corticosterone in
untested control animals, shown below.
PRA Corticosterone
Untested Control Rats
7.1 ± 1.2 8.9 ± 2.1
(Van de Kar et al., 1985) (Van de Kar et al., 1985)
The normal plasma pool values were compared to the means of the
sham- or vehicle-treated groups in each of the experiments of this
dissertation. ANOVA and Newman-Keuls' Multiple Range Test were run to
determine if significant differences between the groups exist. With
respect to plasma renin activity, the data are shown on the following
page:

Group Number
4
1
5
2
6
3
Table 1.
Comparison of Mean Values: Normal PRA Pool versus Experimental
Sham- or Vehicle-Treated Groups
Description Mean + Std.Error
6-0HDA Expt. Vehicle/Control 3.6 ± 0.8
Electrolytic PVN Sham/Control 4.5 ± 0.6
Sotalol Expt. Vehicle/Control 6.3 ± 0.5 + Non-injected/Control
Electrolyt. N. Reun. Sham/Cont. 6.4 ± 0.4
Normal PRA Pool 7.6 ± 0.5
Ibotenic Expt. Vehicle/Control 9.2 ± 0.9
Standard Error of Treatment Means - 0.74
122
n
7
7
12
8
23
7

123
Table 2
Newman-Keuls' Multiple Range Test for PRA
Treatment vs. Treatment Difference Sig .OS
Group 4 Group 1 0.94
Group 4 Group 5 2. 71 * Group 4 Group 2 2.82 * Group 4 Group 6 4.06 * Group 4 Group 3 5.62 * Group 1 Group 5 1. 77
Group 1 Group 2 1. 88
Group 1 Group 6 3.11 * Group 1 Group 3 4.68 * Group 5 Group 2 0.11
Group 5 Group 6 1. 34
Group 5 Group 3 2.91 * Group 2 Group 6 1. 23
Group 2 Group 3 2.80 * Group 6 Group 3 1.56

124
The means of the experimental vehicle- or sham-treated goups were
not significantly elevated above the normal PRA plasma pool. However,
two experimental groups had PRA values lower than the normal PRA pool
levels.
With respect to corticosterone, the data are as follows:
Group Number
3
4
6
1
2
5
Table 3
Comparison of Mean Values: Normal Corticosterone Pool versus Experimental
Sham- or Vehicle-Treated Groups
Description Mean + Std.Error
Ibotenic Expt. Vehicle /Control 6.5 ± 0.5
6-0HDA Expt. Vehicle/Control 7.1 ± 0.5
Normal Corts Pool 8.9 ± 1.1
Electrolyt. PVN Sham/Control 9.3 ± 2.3
Electrolyt. N. Reun. Sham/Cont. 12.8 ± 0.9
Sotalol Expt. Vehicle/Control 12.9 ± 1.2 + Non-injected Control
Standard Error of Treatment Means= 0.74
!!
7
6
13
7
7
12

125
Table 4
Newrnan-Keuls' Multiple Range Test for Corticosterone
Treatment vs. Treatment Difference Sig .05
Group 3 Group 4 0.49
Group 3 Group 6 2.36
Group 3 Group 1 2.75
Group 3 Group 2 6.31 * Group 3 Group 5 6.34 * Group 4 Group 6 1. 86
Group 4 Group 1 2.26
Group 4 Group 2 5.81 * Group 4 Group 5 5.85 * Group 6 Group 1 0.39
Group 6 Group 2 3.95
Group 6 Group 5 3.98
Group 1 Group 2 3.55
Group 1 Group 5 3.58
Group 2 Group 5 0.03
None of the vehicle- or sham-treated experimental groups were
significantly different from the normal corticosterone plasma pool.

CHAPTER V
DISCUSSION
A. Overview
The following conclusions can be drawn from the results presented:
(1) Neurons in the PVN mediate the stress-induced increase in renin and
corticosterone secretion. (2) Catecholaminergic nerve terminals in the
PVN mediate the effect of stress on renin and corticosterone secretion.
(3) Stress-induced corticosterone secretion appears to be mediated by ~
receptors in the PVN, while (4) stress-induced renin secretion is mediated
by a receptor that is not ~-adrenergic. Stress-induced renin secretion
may be mediated by a different catecholamine receptor, a neuropeptide or
other substance(s) colocalized in catecholaminergic nerve terminals.
B. Justification of the Stress Paradigm
Abundant evidence was presented in the literature review to
demonstrate that renin and corticosterone secretion can be elevated by
many different stressors. The stress paradigm used in these experiments
was a conditioned fear paradigm that involved exposing the rats to an
initially neutral stimulus (the stress chamber) which was followed after
ten minutes by an aversive stimulus (footshock). The latter unconditioned
stimulus elicited a range of unconditioned responses, including neuroen
docrine (increased renin and corticosterone levels) and behavioral
(defecation, urination, freezing) responses. As the pairings of stimuli
126

127
were continued, the originally neutral stimulus became a conditioned
stimulus, capable of eliciting the same responses as the unconditioned
stimulus.
1. Classical Conditioning of Fear Stimuli
Classical conditioning can account for the wide range of stimuli
that can produce fear in animals and humans (Watson, 1924). An experiment
by Hunt and Otis (1953) demonstrated that one symptom of fear, defecation,
can be classically conditioned. They presented a rat with a flashing
light for three minutes, at the end of which time the animal received an
electric foot shock. Eight conditioning trials were run. At first,
defecation occurred primarily when the shock was delivered, but it
increasingly began to occur during the presentation of the flashing light.
A similar phenomenon was observed with the conditioned fear
paradigm used in the experiments of this dissertation. Behavioral
observations were made throughout the testing procedure, and recorded on
the third and fourth (final) days. Rats were monitored for defecation
and urination, as well as exploratory, freezing or grooming behaviors.
On the first day, the rats were placed in the stress chamber for a ten
minute interval, at the end of which time they received a mild foot shock.
The rats were tested in this manner for four consecutive days, the only
exception being that on the final day, no foot shock was given. On the
first day in the stress chamber the rats did not defecate until the time
of the foot shock. However by the third day (and in some cases the
second), as soon as the rats were placed in the stress chamber they began

128
to defecate. The defecation suggested that the rats had paired the
placement in the chamber with the impending foot shock, supporting the
hypothesis that classical conditioning is an integral part of the ability
of previously neutral stimuli to elicit fear. Lorens et al. (1989)
reported the number of fecal boli as a defecation score, a quantitative
measurement of a behavioral response to conditioned fear.
In the experiments of this dissertation, the control animals
explored the test chamber and groomed themselves. The stressed animals,
after the first or second day of conditioning, would sometimes explore,
and sometimes groom, but for the most part would defecate, urinate and
exhibit freezing behavior. Freezing behavior consists of tense and silent
immobility, and is seen in many species in times of danger. The
observation of freezing behavior in response to the CS represents another
similarity between my experiments and those presented by Hunt and Otis
(1953). Normally a rat responds to an electric foot shock with a great
increase in activity - running around frantically, jumping, and trying to
escape the shock. However, in my experiments and the experiments of Hunt
and Otis (1953), the stimulus followed by shock caused a radically
different behavior - immobility. This is quite an exception to the usual
pattern of the CS (in my case, the stress chamber) eliciting a similar
(though not exact) reaction to that of the UCS (the footshock). Gray
(1971) suggested the possibility that freezing and more active forms of
escape and avoidance behavior are controlled by separate brain systems,
and represent a combination of learning and innate behavior.
The process of producing electrolytic lesions or injecting
neurotoxins is an invasive procedure that could, theoretically, interrupt

129
fiber pathways that are involved in the learning/conditioning process.
Because this surgical intervention occurs two weeks prior to the
behavioral conditioning paradigm, it is important to look for potential
differences in the behavior of sham or vehicle-injected rats compared with
rats with electrolytic or chemical lesions in the PVN. Behavioral
observations during the conditioning process indicated that the lesions
did not prevent defecation, urination and freezing in the stressed rats.
This suggests that the lesions did not interfere with the conditioning
process.
C. Electrolytic and Ibotenic Acid Lesions in the PVN
The data suggest that neurons in the PVN mediate the effect of
stress on renin secretion and confirm results by others (Rivier and Vale,
1983; Dohanics et al., 1986) that neurons in the PVN mediate the effect
of stress on corticosterone secretion. This conclusion is based on the
finding that both complete electrolytic lesions in the PVN and selective
destruction of cells in the PVN prevented the stress-induced elevation in
plasma renin activity, plasma renin concentration and plasma cort
icosterone levels, whereas electrolytic lesions in the thalamic nucleus
reuniens were ineffective. Ibotenic acid was selected for its ability to
selectively destroy cell bodies while leaving fibers of passage intact
(Markowska et al., 1985; Winn et al., 1984). Magnocellular neurons in the
PVN have been reported to be resistant to the neurotoxic effects of kainic
acid (Zhang and Ciriello, 1985). Herman and Wiegand (1986) have reported
that ibotenic acid selectively destroyed parvocellular neurons in a dose
dependent manner while sparing magnocellular neurons. They reported that

130
loss of parvocellular neurons occurred at 0.5, 0.75, 1.0 and 2.0 µg doses
of ibotenic acid. Loss of magnocellular neurons (mostly vasopressinergic)
was observed only after the 2 .0 µg dose. Their protocol involved
unilateral injections (compared to our protocol of bilateral injections)
directed towards the central portion of the PVN at the level of the
posterior magnocellular division. In our study, the drug volume was
higher (0.3 µl compared to 0.1 µl) and the injection site had to be at
least 0.5 mm lateral to the midsagittal suture. Injections of ibotenic
acid closer to the midline led to diffusion of the drug into the third
ventricle, resulting in death of the rats. We did not observe selective
sparing of the magnocellular neurons after injections of ibotenic acid.
The effect of stress on corticosterone secretion is most likely
propagated by stimulation of CRF neurons in the PVN. The majority of the
diencephalic CRF-neurons are located in the parvocellular neurons in the
dorsal and medial subnuclei of the PVN (Liposits et al., 1987; Liposits
and Paull, 1985). A small number of CRF-neurons also are present in the
magnocellular divisions of the nucleus (Swanson and Sawchenko, 1983;
Piekut and Joseph, 1986). Dohanics et al. (1986) reported that the full
corticosterone and ACTH response to ether or ether plus laparotomy stress
requires not only an intact PVN, but also an intact neurointermediate
lobe. Therefore, our data are consistent with the hypothesis that CRF
neurons in the parvocellular PVN are activated during the CER procedure
to stimulate ACTH and thus corticosterone secretion. Furthermore, the
data with corticosterone support the histological verification of the
accuracy of the lesions in the PVN. The finding that basal corticosterone
concentration was not altered by the PVN lesions is in agreement with

131
previous studies by Meyerhoff et al. (1987) and Makara et al. (1986), who
reasoned that in the absence of CRF input, the cells in the pituitary can
maintain a basal level of ACTH but cannot respond to stimuli which would
increase ACTH secretion. Similar findings were also reported using CRF
antisera (Nakani et al., 1985; Ono et al., 1985; Rivier et al., 1982).
Nucleus reuniens lesions inhibited the effect of stress on
corticosterone but not renin secretion. This suggests that fibers which
course through the nucleus reuniens and innervate the CRF neurons in the
PVN may have been disconnected by this lesion. Since no inhibition of
stress-induced renin secretion occurred in these rats, the data suggest
that the fibers, and possibly cells, which mediate the effect of stress
on renin secretion are different from the fibers and cells which mediate
stress-induced corticosterone secretion.
The nature of the PVN neurons which mediate the effect of stress
on renin secretion is not known. It is clear, both from the present
results and from studies by Porter (1988), that cells in the PVN play an
important role in the regulation of renin secretion. Electrolytic lesions
in the PVN prevent the increase in plasma renin activity after injection
of the serotonin releasing drug p-chloroamphetamine (Gotoh et al., 1987).
Furthermore, these lesions also prevent the increase in plasma renin
activity that follows immobilization stress, although a reduction in
plasma renin substrate (angiotensinogen) levels could account for this
effect (Gotoh et al., 1987). It is important to note that, in the present
study, a saturating concentration of exogenous renin substrate was added
in the plasma renin concentration assay to control for possible variation
of renin substrate in the plasma samples from the experimental rats. In

132
contrast, the plasma renin activity assay was a measure of the production
of angiotensin I from endogenous renin, combined with endogenous renin
substrate. Since plasma renin activity and concentration were equally
affected by the lesions, the changes were clearly due to changes in renin,
not renin substrate concentration in plasma.
D. Injection of 6-0HDA into the PVN
The main conclusion of the 6-0HDA experiment is that catecholaminer
gic nerve terminals in the PVN mediate the effect of stress on both renin
and corticosterone secretion. This conclusion is based on the observation
that injection of 6-0HDA into the PVN inhibited the stress-induced
increase in renin and corticosterone secretion. 6-0HDA is a neurotoxin
that is taken up by the catecholaminergic nerve endings and produces
degeneration of the nerve terminals (Breese and Traylor, 1970; Uretsky and
Iversen, 1970). 6-0HDA is considered to be selective for catecholaminer
gic neurons. Uretsky and Iversen (1970) reported that doses of 6-0HDA
that decreased brain norepinephrine and dopamine levels did not affect
brain serotonin or GABA. Shor-Posner et al. (1986) have reported that
injection of 6-0HDA into the PVN, in the same dose as in my experiment,
produced a 75% reduction in norepinephrine, 56% reduction of epinephrine
and 47% reduction in dopamine concentration in the PVN. The catechol
amines were determined by HPLC and electrochemical detection. Histochemi
cal analysis also revealed a significant decrease in catecholamine
fluorescence in the PVN. This study suggests that 6-0HDA can cause a
degeneration of noradrenergic, adrenergic and dopaminergic elements in the
PVN. Further support was provided by Fety et al. (1984), who injected 6-
0HDA into the fourth ventricle of rats to study the biochemical reponses

133
of the adrenergic (Cl, C2) cell groups versus the noradrenergic (Al, A2
and A6) cell groups. Their results indicated that the adrenergic cell
groups are sensitive to the neurotoxic effects of 6-0HDA since they
exhibited changes in tyrosine hydroxylase and dopamine-&-hydroxylase in
a similar manner to the noradrenergic cell groups.
In the 6-0HDA experiment of this dissertation, norepinephrine
depletion was verified immunocytochemically by using an antibody for
dopamine-&-hydroxylase. No attempt was made to differentiate between loss
of norepinephrine-, dopamine- or epinephrine-containing nerve terminals.
Consequently, these results cannot indicate which catecholamine mediates
the effects of stress on renin or corticosterone secretion.
The suggestion that catecholaminergic nerve terminals in the PVN
mediate stress-induced corticosterone secretion is supported by studies
involving the ventral noradrenergic bundle (VNAB), which conveys most of
the noradrenergic (Moore and Bloom, 1979; Palkovits, 1981; Swanson and
Sawchenko, 1980) and adrenergic (Ungerstedt, 1971) afferents to the PVN.
Injections of 6-0HDA into the VNAB inhibited the ACTH stress response
(Szafarczyk et al. , 1985), prevented the increase in corticosterone
secretion following photic, acoustic and sciatic nerve stimulation
(Feldman et al., 1988), and significantly reduced CRF levels in hypophys
ial portal blood (Guillaume et al., 1987). In addition, Feldman et al.
(1986) demonstrated that 6-0HDA injections into the PVN significantly
inhibit the adrenal response to photic, acoustic and sciatic nerve
stimulation. Together, these results suggest a role for catecholaminergic
afferents to the PVN in the activation of adrenocortical responses to
stress.

134
The dorsal noradrenergic bundle (DNAB) may play a role as a
mediator of stress-induced renin secretion. Some afferent fibers from
the locus coeruleus travel to the PVN in the DNAB (Kobayashi et al., 1975;
Loizou, 1969). Lightman et al. (1984) demonstrated that injections of 6-
0HDA into the DNAB markedly attenuate the increase in plasma renin
activity in response to hemorrhage, while injections of 6-0HDA into the
VNAB had no effect. This study, in combination with my results, suggests
that catecholaminergic afferents to the PVN mediate the renin response to
stress.
E. Injection of Sotalol into the PVN
Since the data suggest that catecholaminergic nerve terminals in
the PVN mediate the stress-induced increase in renin and corticosterone
secretion, the next question to address was whether or not E-adrenergic
receptors mediate this effect. Previous studies demonstrated that the &
adrenoceptor antagonist propranolol (1 mg/kg, i.p.) significantly
attenuated the stress-induced increase in plasma renin activity (Van de
Kar et al., 1985). Since propranolol is known to cross the blood-brain
barrier (Garvey and Ram, 1975; Lemmer et al., 1985), and the PVN contains
a high concentration of~ receptors (Wanaka et al., 1989), it is possible
that a E-receptor, located in the PVN, could be involved in mediating the
effect of stress on renin and corticosterone secretion. In the present
experiment, sotalol was selected instead of propranolol because it is
devoid of local anesthetic activity and is both a &1 and E2 antagonist
(Fitzgerald, 1984). Injection of sotalol into the PVN prevented the
stress-induced increase in corticosterone secretion but did not prevent

135
the stress-induced increase in renin secretion. This suggests that a &
adrenergic receptor in the PVN plays a role in the corticosterone response
to stress. However, many B-adrenoceptor antagonists also bind to
serotonin receptors in the brain (Conway et al., 1978). Since only one
dose of sotalol was administered, it is possible that this dose (15 nmol)
could affect serotonergic as well as B-adrenergic receptors. Other
investigators have injected much higher doses of sotalol and propranolol
in the brain. Injections of sotalol (400 nmol) or propranolol (120 nmol)
into the perif ornical region of the anterior hypothalamus did not prevent
the feeding response induced by either epinephrine (20 nmol) or norepi
nephrine (20 nmol). This feeding response was prevented by injections of
the a-adrenoceptor antagonists phentolamine (30 or 60 nmol), tolazoline
(100 nmol) or phenoxybenzamine (15 nmol) in the same brain region
(Leibowitz, 1975). Injection of propranolol (116 nmol) or timolol (95
nmol) into the cerebral ventricles prevented the effect of air stress on
renal sympathetic nerve activity in spontaneously hypertensive rats. In
addition, icv injection of propranolol (4 or 25 nmol) produced an
inhibition of epinephrine-stimulated ACTH release (Szafarczyk et al.,
1987). In my study, the fact that sotalol inhibited the stress-induced
increase in corticosterone secretion but not the stress-induced increase
in renin secretion argues for a relative selectivity in receptor binding.
Other investigators have provided evidence for the role of &-
receptors in the regulation of corticosterone secretion. Szafarczyk et
al. (1987) demonstrated a stimulatory role for epinephrine and norepineph-
rine on corticosterone release. Injection (icv) of epinephrine or
norepinehrine induced ACTH release comparable to that occuring under

136
(ether) stress conditions. Epinephrine was more efficacious than
norepinephrine on an equimolar basis. The stimulating effect of
norepinephrine was reversed by icv pretreatment with prazosin, an a 1
antagonist, whereas the effect of epinephrine was inhibited by either
prazosin or propranolol. These results suggest that norepinephrine
stimulates ACTH release via a 1 receptors, whereas epinephrine acts via both
a 1- and &-adrenoceptors. In vitro studies support the role of &-adrenergic
receptors in CRF release. Norepinephrine stimulated the release of
immunoreactive CRF from cultured neonatal rat hypothalamic cells. This
effect was blocked by propranolol but not by prazosin. In addition, the
E.-adrenoceptor agonist isoproterenol significantly increased CRF while the
a-adrenoceptor agonist phenylephrine was ineffective, except at high
concentrations (Widmaier et al., 1989). Together, these data strongly
suggest a stimulatory role for &-receptors in the regulation of cor
ticosterone secretion.
To speculate as to which neurotransmitter could be involved in
mediating the effect of stress on corticosterone secretion, several
studies should be considered. Leibowitz et al. (1986) demonstrated that
norepinephrine injections into the PVN dose-dependently increased serum
corticosterone levels, while dopamine had no effect. Epinephrine in the
PVN was even more efficacious than norepinephrine in increasing serum
corticosterone levels. Further, a study by Spinedi et al. (1988)
demonstrated that central inhibition of phenylethanolamine-N-methyltrans
ferase (PNMT), producing a selective decrease in hypothalamic epinephrine
levels, significantly decreased the plasma ACTH response to ether stress,
and at later times also caused a selective decrease in CRF content. These

137
results support a stimulatory role of central epinephrine-containing
neurons in CRF release. Sawchenko et al. (1987) estimated that at least
40% of the retrogradely- labeled catecholaminergic cell groups that project
to the PVN could be adrenergic (PNMT-immunoreactive). These results
suggest that stress-induced increases in corticosterone levels could be
mediated via adrenergic nerve terminals activating B-receptors in the PVN.
Stress-induced increases in plasma renin activity and concentration
are mediated by a different receptor. It is possible that an a-adrenergic
receptor could mediate the stress-induced increase in renin secretion.
However, pharmacological studies suggest that activation of brain a 2-
adrenoceptors lowers renin secretion by inhibiting sympathetic outflow to
the kidneys. Injection of clonidine (icv) prevented the air jet stress
induced increase in renal sympathetic nerve activity in spontaneously
hypertensive rats. This effect was reversed by icv injection of the a 2-
adrenergic antagonists yohimbine and rauwolscine (Koepke and DiBona,
1986). !CV injection of clonidine lowered plasma renin activity at a dose
which is ineffective when given intravenously (Reid et al., 1975). Blair
et al. (1977) demonstrated that administration of L-DOPA to dogs
pretreated with the peripheral decarboxylase inhibitor carbidopa results
in a drop in blood pressure and an inhibition of renin secretion. Since
this treatment increases brain catecholamine content, the results suggest
that catecholamines formed from L-DOPA can act within the central nervous
system to cause a decrease in renin secretion. In this study the decrease
in plasma renin activity was dependent upon the presence of the renal
nerves. These studies suggest that central a 2-adrenoceptors are involved
in decreasing renin secretion. However, it is possible that central a1
-

138
adrenoceptors could mediate the stress-induced increase in renin
secretion. Ganong et al. (1982) suggested that the activation of central
a 1 receptors increases renin secretion in anesthetized dogs. However,
these data are not very convincing since they only reported renin values
following icv injection of the a 1-agonists methoxamine and phenylephrine,
without the appropriate vehicle controls.
The PVN has been shown to project to those segments of the thoracic
cord containing sympathetic preganglioninc neurons innervating the kidney
(Swanson and Mc Kellar, 1979). Other studies have suggested that the
sympathetic nervous system mediates the stress response from the brain to
the kidney (Golin et al., 1988). However, in our laboratory, (see STRESS:
Effect of Peripheral Sympathectomy) sympathectomized-adrenal enucleated
rats showed no inhibition of the stress-induced increase in plasma renin
activity/concentration or plasma corticosterone levels (Richardson Morton
et al., 1988). This suggests that the sympathetic nervous system and
adrenal catecholamines are not the only mediators of the stress-induced
increase in renin secretion. An alternate theory is that cell bodies in
the PVN are stimulated to secrete a renin-releasing factor to the
bloodstream to promote renin release from the kidneys (Van de Kar et al.,
1987).
If a and~ receptors in the PVN do not mediate stress-induced renin
secretion, another possibility is that a neuropeptide, coreleased at the
catecholaminergic nerve terminals in the PVN, mediates stress-induced
renin secretion. A likely candidate is neuropeptide Y, which is
coexpressed in noradrenergic and adrenergic brain stem cell groups and is
present in noradrenergic and adrenergic nerve terminals in the PVN

139
(Sawchenko et al., 1985). Intraventricular injection of NPY has been
shown to increase plasma ACTH and corticosterone secretion (Harfstrand et
al., 1987) and plasma angiotensin II levels (Harfstrand et al., 1986).
However, Fuxe et al. (1983) reported that intracisternal injection of NPY
causes hypotension, which could also increase renin secretion by
activation of the intra-renal stretch receptor (Fahri et al., 1982).
Yet another neuropeptide of interest is galanin, due to its
preferential coexistance with noradrenergic cell groups that project to
the PVN. Combined immunohistochemical-retrograde transport studies have
demonstrated galanin-immunoreactivity coexisting with dopamine-&
hydroxylase immunoreactivity in the Al and A6 cell groups, with no
evidence for galanin coexistence in adrenergic (PNMT-immunoreactive)
neurons that project to the PVN. Galanin inputs were confined to the
anterior, periventricular, dorsal and ventral subdivisions of the
parvocellular PVN, with only a sparse projection to the oxytocinergic
neurons of the magnocellular division of the PVN (Levin et al, 1987).
F. Hypothetical Organization of Brain Pathways Mediating
Stress-Induced Renin and Corticosterone Secretion
The neural pathways mediating stress-induced renin and cort
icosterone secretion are likely to be complex and multi-synaptic, due to
the many inputs which must be integrated. First, there is a sensory
component that arises during behavioral conditioning. When the rat
receives the foot shock as part of the conditioned fear paradigm, the
sensory message is transmitted from the nerve fibers in the skin to the
spinal cord, where it travels in the lateral spinothalamic tract to the

140
posteroventral nuclei of the thalamus. The spinothalamic fibers are
accompanied by fibers that terminate in the reticular formation (spino
reticular) and the periaqueductal gray and tectum (spinotectal). The
periaqueductal grey has been shown to play a role in flight or defensive
behavior (de Molina and Hunsperger, 1962).
The pain input route most likely continues from the midbrain
reticular formation, through the thalamus, to reach the septum and
hippocampus. The septo-hippocampal system plays a major role in pain
reception. The hippocampus emits a theta wave (4-9 cps rhythm) as a
response to a painful stimulus in the rat, and in the emotional states of
disappointment and frustration in humans. Lesions in the medial nuclei
of the septum and the thalamic nucleus centrum medianum abolish the theta
wave emitted by the hippocampus in response to a painful stimulus (Gray,
1971).
Damage to the hippocampus has been shown to facilitate acquisition
of shuttle box avoidance (Green et al., 1967; Van Hoesen et al., 1972)
possibly due to a deficit in a behavioral inhibitory system (Douglas,
1967; Gray, 1982). These studies, and others (Squire and Zola-Morgan,
1983), suggest a critical role of the hippocampal formation in learning
and memory. The hippocampal formation and septa! region share extensive
bidirectional connections. The lateral, medial and posterior divisions
of the septa! regions are associated with the hippocampal formation while
the ventral division (bed nuclei) is primarily related to the amygdala
(Swanson et al., 1987). The subicular complex and the entorhinal area of
the hippocampus receive an extensive sensory input from the cortex,
including olfactory, visual, auditory and somatosensory. Additional

141
sensory information is relayed through two routes, one via the thalamus,
extrahippocampal cortex and amygdala and the other via ascending pathways
from the brainstem and basal forebrain. The hippocampus also receives
considerable serotonergic input from the raphe nuclei (Azmitia, 1978;
Lorens and Guldberg, 1974; Steinbusch, 1981). Immunoreactive-CRF neurons
have been demonstrated in the hippocampus (Swanson, 1983). However, the
axonal projections of these cells are relatively short, suggesting that
they might function as interneurons (Kohler, 1986).
The hippocampus has many interconnections to another limbic
structure, the amygdala (Krettek and Price, 1978). The amygdaloid complex
has been shown to be involved in the regulation of fear-motivated behavior
(Blanchard and Blanchard, 1972; Spevack et al., 1975). The amygdala
appears to play a role in the stress response via the pituitary-adrenal
cortical axis . The central nucleus of the amygdala, in particular,
contains large numbers of CRF-containing cell bodies and terminals
(Swanson et al., 1983; Moga and Gray, 1985). Both the central and medial
amygdaloid nuclei project to the thalamic nucleus reuniens (Aggleton and
Mishkin, 1984). This helps to explain why lesions in the nucleus reuniens
prevented the effect of stress on corticosterone secretion.
Most amygdaloid nuclei project to the bed nucleus of the stria
terminalis (BNST) and the hypothalamus. Since the hypothalamus also
receives a substantial projection from the BNST, there exists a di
synaptic route for the amygdala to modulate hypothalamic function (Krettek
and Price, 1978). The BNST also receives fibers from the dorsal and
median raphe nucleus (Azmitia and Segal, 1978), as well as a massive
projection from the hippocampus (Swanson and Cowan, 1977). To summarize,

142
the incoming sensory information and "learning"/associative processes that
accompany the conditioned fear paradigm are likely conveyed to the PVN via
limbic regions of the telencephalon since direct neocortical projections
to the hypothalamus cannot be demonstrated (Swanson et al., 1987). The
hippocampus, amygdala and ventral septal region project heavily to the
BNST, which in turn projects to the CRF regions of the PVN. The question
that remains is: how does this input to the PVN become integrated with the
direct nordrenergic (A2) and adrenergic (Cl-C3) input via the VNAB (brain
stem to the CRF neurons in the PVN)?
Swanson et al. (1983) suggested that the nucleus of the solitary
tract, which receives direct vagal and glossopharyngeal afferents
(Beckstead and Norgren, 1979) could serve as a major relay point of
visceral sensory information to CRF cell bodies in the basal forebrain.
Likewise the parabrachial nucleus, which receives a large input from the
NTS (Norgren, 1978), and the locus coeruleus also project to many of the
sites that contain CRF-immunoreactive cell bodies, including the
parvocellular PVN, the bed nucleus of the stria terminalis and the central
nucleus of the amygdala. As mentioned above, these nuclei may also be
involved in the relay of visceral sensory information to CRF cell bodies
(Swanson et al., 1983). This hypothetical pathway could provide a direct
stimulus for corticosterone secretion.
Another possibility is that the noradrenergic input to the PVN is
serving as a neuromodulator to "set a bias" to amplify or buffer incoming
stimuli to the cells. Even though norepinephrine might have little effect
on the resting excitability of the CRF neurons, it could have profound

143
effects on the amount of CRF released following an action potential
(Nicoll, 1982). For example, pretreatment with norepinephrine (5 µ.M)
before iontophoretically-applied glutamate (50 nA) in a pyramidal
(hippocampal) cell has been shown to evoke a higher frequency and longer
duration train of action potentials than following glutamate alone
(Madison and Nicoll, 1986).
Wallace et al. (1989) demonstrated that the amygdala innervates the
locus coeruleus (A6) and NTS (A2) noradrenergic cells, and the C2
adrenergic cells. Most of the rostral A6 and A2 neurons had amygdaloid
terminal contacts. This pathway could be involved in integrating the
sensory/associative processes with the stimulatory catecholaminergic
afferents to the PVN.
The possibility exists that serotonergic neurons play a role in
transmitting the stress stimulus. The dorsal raphe nucleus (B7) is one
of the nine 5-HT cell groups originally described by Dahlstrom and Fuxe
(1965), which innervates the forebrain. Studies from our laboratory have
shown that electrolytic lesions in the dorsal raphe nucleus prevent the
stress-induced increase in plasma renin activity using a three-minute CER
(Van de Kar et al., 1984). This experiment was repeated, extending the
CER to ten minutes so that corticosterone levels could be evaluated.
Confirming the previous results, we found that electrolytic lesions in the
dorsal raphe nucleus prevented the stress - induced increase in plasma renin
activity and concentration and corticosterone levels (Richardson Morton
et al., 1986). There could be a connection between the fact that both
dorsal raphe lesions and PVN lesions prevent the stress-induced increase
in plasma renin activity, plasma renin concentration and corticosterone

levels.
144
Sawchenko et al. (1983) have demonstrated that serotonergic
neurons in the midbrain (especially the dorsal raphe) innervate the PVN.
Although the serotonergic fiber density in the PVN is low compared with
immediately surrounding areas, there is a distinct projection to the
medial parts of the parvocellular division. Liposits et al. (1987) have
demonstrated by electron microscopic methods that serotonergic nerve
terminals innervate CRF-immunoreactive parvocellular cells in the PVN.
Injections (i.p.) of the 5-HT2 antagonist, LY53857, did not prevent
the stress-induced increase in renin or corticosterone secretion
(Richardson Morton et al., 1986). However, injection of the 5-HT1A
agonists buspirone and ipsapirone significantly attenuated the stress
induced increase in plasma renin activity, plasma renin concentration and
corticosterone levels (Urban et al., 1986; Van de Kar et al., 1985; Lorens
et al. , 1989). Both buspirone and ipsapirone inhibit the firing of
serotonergic neurons in the dorsal raphe (Basse-Tomusk, 1986; VanderMaelen
et al., 1986). Thus, serotonergic neurons in the dorsal raphe nucleus
could be involved in stress-induced renin and corticosterone secretion.
There are fiber projections from the locus coeruleus to the dorsal raphe
nucleus (Cowan and Park, 1986), with direct noradrenergic nerve terminal
innervation of serotonin nerve cells in the dorsal raphe (Baraban and
Aghajanian, 1981), and projections from the dorsal raphe to the locus
coeruleus (Conrad et al., 1974). These interconnections could provide a
relay for the noradrenergic bundles to influence hypothalamic neuroen
docrine responses.
Another possibility is that the dorsal raphe serotonergic neurons,
which also project to the hippocampus (Swanson et al., 1987), provide a

145
tonic inhibition of the "behavioral inhibition" system of the hippocampus.
In this case, administration of 5-HT1A anxiolytic agents or electrolytic
lesions in the dorsal raphe nucleus would decrease the input to the septo
hippocampal system, thus reducing the activity of the septo-hippocampal
behavioral system, ultimately decreasing the behavioral and possibly
neuroendocrine stress response (Kuhar, 1986).
Clearly, the neural pathways mediating stress-induced renin and
corticosterone secretion are complex and multi-synaptic. From this
dissertation, it can be concluded that stress-induced corticosterone
secretion appears to be mediated by Preceptors in the PVN, while stress
induced renin secretion is mediated by a receptor that is not P-adrener
gic. Stress- induced renin secretion may be mediated by a different
catecholamine receptor, a neuropeptide or other substance(s) colocalized
in catecholaminergic nerve terminals.
The following figure is a schematic diagram of the hypothetical
brain pathways mediating stress-induced renin and corticosterone
secretion.
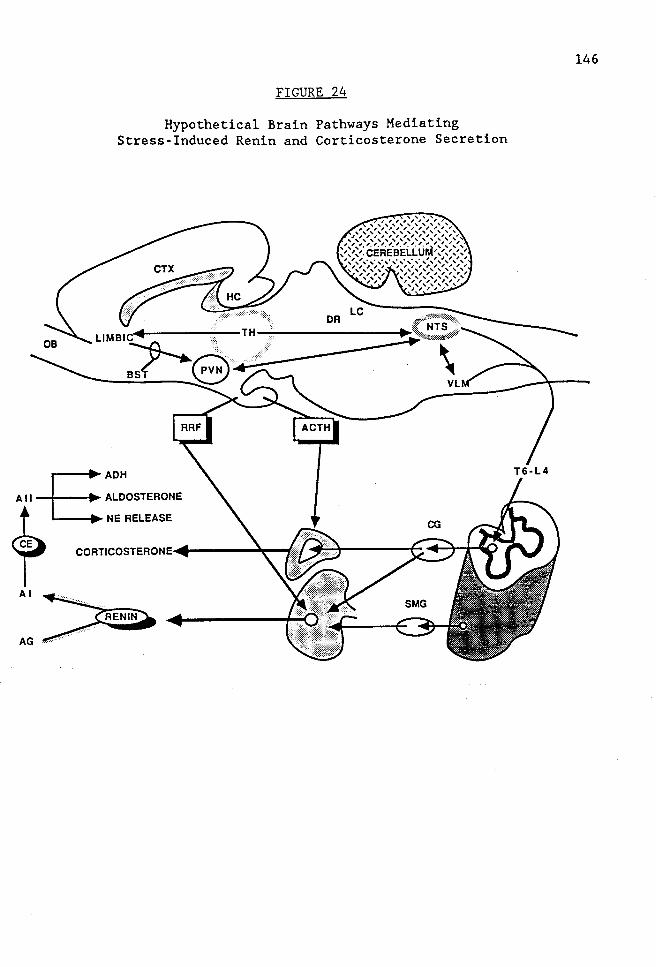
FIGURE 24
Hypothetical Brain Pathways Mediating Stress-Induced Renin and Corticosterone Secretion
~ AG~
146

LITERATURE CITED
Aggleton, J. P. and Mishkin, M.: Projections of the amygdala to the thalamus in the cynomolgus monkey. J. Comp. Neurol. 222: 56-68, 1984.
Aghajanian, G. K., Kuhar, M. J., and Roth, R.H.: Serotonin-containing neuronal perikarya and terminals: Differential effects of p-chlorophenylalanine. Brain Research 54: 85-101, 1973.
Allen, J. P. and Allen, C. F.: Role of the amygdaloid complexes in the stress-induced release of ACTH in the rat. Neuroendocrinology 15: 220-230, 1974.
Allen, J. P. and Allen, C. F.: Amygdalar participation in tonic ACTH secretion in the rat. Neuroendocrinology 19: 115-125, 1975.
Allen, L. G., Kalra, P. S., Crowley, W. R., and Kalra, S. P.: Comparison of the effects of neuropeptide Y and adrenergic transmitters on LH release and food intake in male rats. Life Sciences 37: 617-623, 1985.
Alonso, G., Szafarczyk, A., Balmefrezol, M., and Assenmacher, I.: Immunocytochemical evidence for stimulatory control by the ventral noradrenergic bundle of parvocellular neurons of the paraventricular nucleus secreting corticotropin releasing hormone and vasopressin in rats. Brain Research 397: 297-307, 1986.
Alper, R. H. and Ganong, W. F.: Pharmacological evidence that the sympathetic nervous system mediates the increase in secretion of renin produced by p-chloroamphetamine. Neuropharmacology 11: 1237-1240, 1984.
Anderson, D. E. and Brady, J. V.: Preavoidance blood pressure elevations accompanied by heart rate decreases in the dog. Science 172: 595-597, 1971.
Anderson, D. E. and Brady, J. V.: Differential preparatory cardiovascular responses to amine and operative behavioral conditioning. Condit. Refl. 7: 82-96, 1972.
Anderson, D. E. and Brady, J. V.: Cardiovascular responses to avoidance conditioning in the dog: effects of beta adrenergic blockade. Psychosom. Med. 38: 181-189, 1976.
Anderson, D. E., Yingling, J. E., and Brady, J. V.: Cardiovascular responses to avoidance conditioning in the dog: effect of alpha adrenergic blockade. Pav. J. Biol. Sci. 11: 150-161, 1976.
Armario, A., Montero, J. L., and Balasch, J.: Sensitivity of corticosterone and some metabolic variables to graded levels of low intensity stresses in adult male rats. Physiology and Behavior 37: 559-561, 1986.
147

148
Armario, A., Restrepo, C., Castellanos, J. M., and Balasch, J.: Dissociation between Adrenocorticotropin and Corticosterone Responses to Restraint after previous chronic exposure to stress. Life Sciences 36: 2085-2092, 1985.
Ayers, C. R.: Plasma renin activity and renin-substrate concentration in patients with liver disease. Gire. Res. 20: 594-598, 1967.
Azmitia, E. C. The serotonin-producing neurons in the midbrain median and dorsal raphe nuclei. In L. L. Iversen, S. D. Iversen and S. H. Snyder (Eds.), Handbook of Psychopharmacology: Chemical Pathways in the Brain. New York: Plenum Press, 1978.
Azmitia, E. C. and Segal, M.: An autoradiographic analysis of differential ascending projections of the dorsal and median raphe nuclei in the rat. J. Comp. Neurol. 179: 641-668, 1978.
Baertschi, A. J., Gahwiler, B., Antoni, F., Holmes, M. C., and Makara, G. B.: No role of vasopressin in stress-induced ACTH secretion? Nature 308: 85-86, 1984.
Baraban, J. M. and Aghajanian, G. K.: Noradrenergic innervation of serotonergic neurons in the dorsal raphe: Demonstration by electron microscopic autoradiography. Brain Research 204: 1-11, 1981.
Bartter, F. C., Casper, A. C., Delea, C. S., and Slater, J. D. H.: On the role of the kidney in control of adrenal steroid products. Metabolism 10: 1006-1020, 1961.
Basse-Tomusk, A. and Rebec, G. V.: Ipsapirone depresses neuronal activity in the dorsal raphe nucleus and the hippocampal formation. Europ. J. Pharmacol. 130: 141-143, 1986.
Basso, N., Gripson, D., Ruiz, P., and Taquini, A. C.: Regional distributions of angiotensin in the central nervous system of the rat: effect of DOCA-salt treatment. Clin. Sci. 63: 1495-1525, 1982.
Bayliss, P. H. and Robertson, G. L.: Rat vasopressin response to insulin-induced hypoglycemia. Endocrinology 107: 1975-1979, 1980.
Beaty, 0., Sloop, C. H., Schmid, H. E., and Buckalew, V. M.: Renin response and angiotensinogen control during graded hemorrhage and shock in the dog. Am. J. Physiol. 231: 1300-1307, 1976.
Beaulieu, S., Di Paolo, T., and Barden, N.: Control of ACTH secretion by the central nucleus of the amygdala: implication of the serotoninergic system and its relevance to the glucocorticoid delayed negative feedback mechanism. Neuroendocrinology 44: 247-254, 1986.

149
Beaulieu, S., Di Paolo, T., Cote, J., and Barden, N.: Participation of the central amygdaloid nucleus in the response of adrenocorticotropin secretion to immobilization stress: Opposing roles of the noradrenergic and dopaminergic systems. Neuroendocrinology 45: 37-46, 1987.
Beckstead, R. M. and Norgren, R.: An autoradiographic examination of the central distribution of the trigeminal, facial, glossopharyngeal, and vagus nerve in the monkey. J. Comp. Neurol. 184: 455-472, 1979.
Bellin, S. I., Bhatnagar, R. K., and Johnson, A. K.: Periventricular noradrenergic systems are critical for angiotensin-induced drinking and blood pressure responses. Brain Research 403: 105-112, 1987.
Benetos, A., Gavras, I., and Gavras, H.: Norepinephrine applied in the paraventricular hypothalamic nucleus stimulates vasopressin release. Brain Res. 381: 322-326, 1986.
Beny, J. L. and Baertschi, A. J.: Oxytocin: major tropin-releasing factor secreted from diabetes insipidus rat pituitary in vitro. Neuroendocrinology 31: 261-264, 1980.
corticoposterior
Berkenbosch, F., Vermes, I., Buijs, R. M., and Tilders, F. J. H.: Vasopressin is not involved in the catecholamine-induced release of ACTH, alpha-MSH and beta-endorphin from the rat pituitary gland. Neuroendocrinology 37: 117-121, 1983.
Bertand, M. The Behavioral Repertoire of the Stump Tail Macque. New York: Karger, 1969.
Bing, J. and Poulsen, K.: Experimentally-induced changes in plasma angiotensinogen and plasma renin. Acta Pathol. Microbiol. Scand. 77: 389-398, 1969.
Biron, P., Koiw, E., Nowaczynski, W., Broulet, J., and Genest, J.: The effects of intravenous infusion of valine-5-angiotensin II and other pressor agents on urinary electrolytes and corticosteroids including aldosterone. J. Clin. Invest. 40: 338-347, 1961.
Bjorklund, A., Falck, B., and Rosengren, E.: Monoarnines in the pituitary gland of the pig. Life Sciences 6: 2103-2110, 1967.
Blair, M. L. , Chen, Y. H. , and Izzo, J. L. : Influence of renal perfusion pressure on alpha- and beta-adrenergic stimulation of renin release. Arn. J. Physiol. 248: E317-E326, 1985.
Blair, M. L., Feigl, E. 0., and Smith, 0. A.: Elevation of plasma renin activity during avoidance performance in baboons. Arn. J. Physiol. 231: 772-776, 1976.
Blair, M. L., Reid, I. A., and Ganong, W. F.: Effect of L-DOPA on plasma renin activity with and without inhibition of extracerebral DOPA decarboxylase in dogs. J. Pharmacol. Exp. Ther. 202: 209-215, 1977.

150
Blair-West, J. R., Reid, I. A., and Ganong, W. F.: Stimulation of angiotensinogen release by raised blood angiotensin concentration in the dog. Clin. Sci. Mol. Med. 46: 665-669, 1974.
Blanchard, D. C. and Blanchard, R. J.: Innate and conditioned reactions to threat in rats with amygdaloid lesions. J. Comp. Physiol. Psychol. 81: 281-290, 1972.
Bliss, E. L., Ailion, J., and Zwanziger, J.: Metabolism of norepinephrine, serotonin and dopamine in rat brain with stress. J. Pharmacol. Exp. Ther. 168: 258-263, 1968.
Bloom, F. E., Battenberg, E., Rossier, J., Ling, N., andGuillemin, R.: Neurons containing beta-endorphin exist separately from those containing enkephalin: Immunocytochemical studies. Proc. Natl. Acad. Sci. (USA) 75: 1591-1595, 1978.
Blumberg, A. L., Ackerly, J. A., and Peach, M. J.: Differentiation of neurogenic and myocardial angiotensin II receptors in isolated rabbit atria. Circ. Res. 36: 719-726, 1975.
Bodian, D. and Maren, T. H.: Effect of neuro- and adenohypophysectomy on retrograde degeneration in the hypothalamic nuclei of the rat. J. Comp. Neurol. 94: 485-511, 1951.
Bonjour, J. P. and Malvin, R. L.: Stimulation of ADH release by the renin-angiotensin system. Am.J. Physiol. 218: 1555-1559, 1970.
Breese, G. R. and Traylor, T. D.: Effect of 6-hydroxydopamine on brain norepinephrine and dopamine: Evidence for selective degeneration of catecholamine neurons. Journal of Pharmacology and Experimental Therapeutics 174(3): 413-420, 1970.
Britton, D. R., Koob, G. F., Rivier, J., and Vale, W.: Intraventricular corticotropin-releasing factor enhances behavioral effects of novelty. Life Sciences 31: 363, 1982.
Britton, S. W. and Silvette, H.: Some effects of cortico-adrenal extract and other substances on adrenalectomized animals. Am. J. Physiol. 99: 15-32' 1931.
Brod, J. Haemodynamics and emotional stress. In M. Koster,H. Musaph and P. Visser (Eds.), Psychosomatics in Essential Hypertension. Bihl. Psychiat., 1970.
Brod, J., Fencl, V., Hejl, Z., and Jirka, J.: Circulatory changes underlying blood pressure elevation during acute emotional stress (mental arithmetic) in normotensive and hypertensive subjects. Clin. Sci. 18: 269-279, 1959.

151
Brod, J., Fencl, V., Hejl, Z., Jirka, J., and Ulrych, M.: General and regional haemodynamic patterns underlying essential hypertension. Clin. Sci. 23: 339-349, 1962.
Brody, M. J., Alper, R.H., O'Neill, T. P., and Porter, J.P. 1. Central neural control of the cardiovascular system. In A. Zanchetti and R. C. Tarazi (Eds.), Handbook of hypertension: Pathophysiology of hypertension- regulatory mechanisms. : Elsevier Science Publishers, 1986.
Brody, M. J. and Johnson, A. K. Role of the anteroventral third ventricle region in fluid and electrolyte balance, arterial pressure regulation, and hypertension. In L. Martini and W. F. Ganong (Eds.), Frontiers in Neuroendocrinology. New York: Raven Press, 1980.
Brown, M. R., Fischer, L.A., Spiess, J., Rivier, C., Rivier, J., and Vale, W.: Corticotropin-releasing factor: actions on sympathetic nervous system and metabolism. Endocrinology 111: 928, 1982.
Brown, R. J. Mammalian social odors: A critical review. In J. S. Rosenblatt,R. A. Hinde, L. Beer and M. Busnel (Eds.), Advances in the study of behavior. New York: Academic Press, 1979.
Brown-Sequard, C. E.: Recherches experimentales sur la physiologie et la pathologie des capsules surrenales. C.R. Acad. Sci. (Paris) 43: 422-425, 1856.
Brownfield, M. S., Reid, I. A., Ganten, D., and Ganong, W. F.: Differential Distribution of Immunoreactive Angiotensin and Angiotensin-Converting Enzyme in Rat Brain. Neuroscience 7: 1759-1769, 1982.
Bruni, J. F., Hawkins, R. L., and Yen, S. S. C.: Serotonergic mechanism in the control of beta-endorphin and ACTH release in male rats. Life Sciences 30: 1247-1254, 1982.
Buckingham, J.C. and Hodges, J. R.: Hypothalamic receptors influencing the secretion of corticotropin-releasing hormone in the rat. J. Physiol. (London) 290: 421-431, 1979.
Bugnon, C., Hadji, Y., Iassemis, M., Fellman, D., and Cardot, J.: Reserpine-induced depletion of corticoliberin (CRF)-like immunoreactivity in the zona externa of the rat median eminence. Brain Research 275: 198-201, 1983.
Bunag, R. D. and Inoue, A. : Differentiated pressor and sympathetic response to dual brain stimulation: Ventromedial hypothalamus versus locus coeruleus. Proc. Soc. Exp. Biol. Med. 178: 90-100, 1985.
Caldwell, P. R. B., Seegal, B. C., and Hsu, K. C.: Angiotensin-Converting Enzyme: Vascular Endothelial Localization. Science 191: 1050-1051 .• 1976.

152
Campbell, D. J., Skinner, S. L., and Day, A. J.: Cellophane perinephritis hypertension and its reversal in rabbits. Circ. Res. 33: 105-112, 1973.
Cannon, W. B.: The emergency function of the adrenal medulla in pain and the major emotions. Am. J. Physiol. 33: 356-372, 1914.
Carlsson, A., Falck, B., and Hillarp, N.: Cellular localization of brain monoamines. Acta Physiol. Scand. 56: 1-27, 1962.
Carretero, 0. and Gross, F.: Renin substrate in plasma under various experimental conditions in the rat. Am J Physiol 213(3): 695-700, 1967.
Castren, E. and Saavedra, J.M.: Repeated stress increases the density of angiotensin II binding sites in rat paraventricular nucleus and subfornical organ. Endocrinology 122(1): 370-372, 1988.
Changaris, D., Demers, L., Keil, L., and Severs, W. Immunopharmacology of angiotensin I in brain. In J. Buckley and C. Ferrario (Eds.), Central Actions of Angiotensin and Related Hormones. New York: Pergamon Press, 1977.
Changaris, D. G., Keil, L., and Severs, W. B.: Angiotensin II immunohistochemistry of the rat brain. Neuroendocrinology 25: 257-274, 1978.
Charney, D. S. and Redmond, D. E.: Neurobiological mechanisms in human anxiety--Evidence supporting central noradrenergic hyperactivity. Neuropharmacology 22: 1531-1536, 1983.
Churchill, P. C., Churchill, M. C., and McDonald, F. D.: Renin secretion and distal tubule Na+ in rats. Am. J. Physiol. 235(6): F611-F616, 1978.
Cintra, A., Fuxe, K., Harfstrand, A., Agnati, L. F., Wikstrom, A. C., Okret, S., Vale, W., and Gustafsson, J. A.: Presence of glucocorticoid receptor immunoreactivity in corticotropin releasing factor and in growth hormone releasing factor immunoreactive neurons of the rat di- and telencephalon. Neuroscience Letters 77: 25-30, 1987.
Ciriello, J. and Calaresu, F.: Lateral reticular nucleus: a site of somatic and cardiovascular integration in the cat. Am. J. Physiol. 233: RlOO, 1977.
Ciriello, J., Kline, R. L., Zhang, T. X., and Caverson, M. M.: Lesions of the paraventricular nucleus alter the development of spontaneous hypertension in the rat. Brain Research 310: 355-359, 1984.
Clamage, D. M., Sanford, C. S., Vander, A. J., and Mouw, D.R.: Effect of psychosocial stimuli on plasma renin activity in rats. Am J Physiol 231: 1290-1294, 1976.
Cohen, D. H. and Randall, D. C.: Classical conditioning of cardiovascular responses. Ann. Rev. Physiol. 46: 187-197, 1984.

153
Conrad, L. C. A., Leonard, C. M., and Pfaff, D. W.: Connections of the median and dorsal raphe nuclei in the rat: An autoradiographic and degeneration study. J. Comp. Neurol. 156: 179-206, 1974.
Conway, J., Greenwood, D. T., and Middlemiss, D. N.: Central nervous actions of beta-adrenoceptor antagonists. Clin. Sci. Mol. Med. 54: 119-124, 1978.
Cori, C. F. and Cori, G. T.: The fate of sugar in the animal body. VII. The carbohydrate metabolism of adrenalectomized rats and mice. J. Biol. Chem. 74: 473-494, 1927.
Correa, F. M., Innis, R. B., Uhl, G. R., and Snyder, S. H.: Bradykinin-like immunoreactive neuronal systems localized histochemically in rat brain. Proc. Natl. Acad. Sci. (USA) 76: 1489-1493, 1979.
Corrodi, H., Fuxe, K., and Hokfelt, T.: The effect of immobilization stress on the activity of central monoamine neurons. Life Sciences 7: 107-112, 1968.
Cowan, R. L. and Park, M. L.: PHAL Mapping of the Locus Coeruleus to Dorsal Raphe Nucleus Projection in the Rat. Neuroscience Abstracts 12: 139' 1986.
Cowan, W. M., Raisman, G., and Powell, R. P. S.: The connections of the amygdala. J. Neurol. Neurosurg. Psychiat. 28: 137-151, 1965.
Cuello, A. C., Scapagnini, U., Licko, V., Preziosi, P., and Ganong, W. F. : Effect of dihydroxyphenylserine on the increase in plasma corticosterone in rats treated with alpha-methyl-para-tyrosine. Neuroendocrinology 13: 115-122, 1973.
Cunningham, E. T. and Sawchenko, P. E.: Anatomical specificity of noradrenergic inputs to the paraventricular and supraoptic nuclei of the rat hypothalamus. The Journal of Comparative Neurology 274: 60-76, 1988.
Dabsys, S. M., Balda, M. S., Pirola, C. J., Finkielman, S., and Nahmod, V. E.: Angiotensin converting enzyme activity in the amygdaloid complex in a neurogenic hypertensive model. Clin. and Exper. Hyper. Al0(4): 605-615, 1988.
Dahlstrom, A. and Fuxe, K.: Evidence for the existence of monoamine-containing neurons in the central nervous system IV. Acta Physiol. Scand. 64(247): 1-36, 1965.
Darlington, D. N., Shinsako, J., and Dallman, M. F.: Paraventricular lesions: hormonal and cardiovascular responses to hemorrhage. Brain Research 439: 289-301, 1988.
Davis, J. O. and Freeman, R.H.: The use of angiotensin II blockade to study adrenal steroid secretion. Fed. Proc. 35: 2508-2511, 1976.

154
De Kloet, E. R., Sybesma, H., and Reul, M. H. M.: Selective control by corticosterone of serotonin-1 receptor capacity in raphe-hippocampal system. Neuroendocrinology 42: 513-521, 1986.
de Molina, A. F. and Hunsperger, R. W.: Organization of the subcortical system governing defence and flight reactions in the cat. J. Physiol. 160: 200-213, 1962.
De Vito, E., Koninckx, E., Cabrerea, R., and Nolly, H.: Half-life of circulating renin under different experimental conditions. Mayo Clin. Proc. 52: 424-426, 1977.
Decavel, C., Geffard, M., and Calas, A.: Comparative study of dopamineand noradrenaline-immunoreactive terminals in the paraventricular and supraoptic nuclei of the rat. Neuroscience Letters 77: 149-154, 1987.
DiBona, G. F.: Neural regulation of renal tubular sodium reabsorption and renin secretion. Fed. Proc. 44: 2816-2822, 1985.
Dierickx, K. and somatostatin neurons 349-359, 1979.
Vandesande, F.: Immunochemical in the rat hypothalamus. Cell
localization of Tiss. Res. 201:
Dohanics, J., Kapocs, G., Janaky, T., Kiss, J. Z., Rappay, G., Laszlo, F. A., and Makara, G. B.: Mechanism of restoration of ACTH release in rats with long-term lesions of the paraventricular nuclei. J. Endocr. 111: 75-82, 1986.
Douglas, R. J.: The hippocampus and behavior. Psychological Bulletin 67: 416-442, 1967.
Dua, J. K. and Dobson, M. J.: Role of olfactory cues in acquisition and extinction of avoidance. Journal of Experimental Psychology 103: 461-465, 1974.
Dunn, A. J. and Berridge, C. W.: Corticotropin-releasing factor administration elicits a stress-like activation of cerebral catecholaminergic systems. Pharmacol. Biochem. Behav. 27(4): 685-691, 1987.
Dykman, R. A. and Gantt, W. H.: Relation of experimental tachycardia to amplitude of motor activity and intensity of the motivating stimulus. Am. J. Physiol. 185: 495-498, 1956.
Dzau, V. J. and Pratt, R. R. Renin-Angiotensin System: Biology, Physiology and Pharmacology. In H. A. Fozzard (Ed.), The Heart and Cardiovascular System. New York: Raven Press, 1986.
Eckland, D. J. A., Todd, K., Jessop, D. S., Biswas, S., and Lightman, S. L.: Differential effects of hypothalamic catecholamine depletion on the release of arginine vasopressin and CRF-41 into hypothalamohypophyseal portal blood. Neuroscience Letters 90: 292-296, 1988.

155
Eggena, P. and Barrett, J. D.: Renin substrate release in response to perturbations of renin-angiotensin system. Am. J. Physiol. 254: E389-E393, 1988.
Eisenberg, R. M.: Further evidence of a central alpha-adrenergic inhibitory influence on the hypothalamo-pituitary-adrenal axis in the rat. Neuroendocrinology 17: 154-166, 1975.
Eliasson, K.: Stress and Catecholamines. Acta Med. Scand. 215: 197-204, 1984.
Epstein, S. and Hamilton, S.: Cyproheptadine inhibition of stimulated plasma renin activity. J. Clin. Endocrinol. Metab. 45: 1235-1237, 1977.
Fahri, E. R., Cant, J. R., and Barger, A. C.: Interactions between intrarenal epinephrine receptors and the renal baroreceptor in the control of PRA in conscious dogs. Circ. Res. 50: 477-485, 1982.
Fanselow, M. S.: Odors released by stressed rats produce opioid analgesia in unstressed rats. Behavioral Neuroscience 99(3): 589-592, 1985.
Feldman, S. and Conforti, N.: Amygdalectomy inhibits adrenocortical responses to somatosensory and olfactory stimulation. Neuroendocrinology 32: 330-334, 1981.
Feldman, S., Conforti, N., and Chowers, I.: Adrenocortical responses following sciatic nerve stimulation in rats with partial hypothalamic deafferentations. Acta endocr., Copenh. 80: 625-629, 1975.
Feldman, S., Conforti, N., and Melamed, E.: Norepinephrine depletion in the paraventricular nucleus inhibits the adrenocortical responses to neural stimuli. Neuroscience Letters 64: 191-195, 1986.
Feldman, S., Conforti, N., and Melamed, E.: Paraventricular nucleus serotonin mediates neurally stimulated adrenocortical secretion. Brain Res. Bull. 18: 165-168, 1987.
Feldman, S., Conforti, N., and Melamed, E.: Involvement of ventral noradrenergic bundle in corticosterone secretion following neural stimuli. Neuropharmacology 27(2): 129-133, 1988.
Fety, R., Lambas-Senas, L., Chamba, G., and Renaud, B.: Changes in tyrosine hydroxylase and dopamine-beta-hydroxylase activities but not in phenylethanolamine-N-methyltransferase activity within central adrenaline neurons after 6-hydroxydopamine administration. Biochemical Pharmacology 33(12): 1887-1891, 1984.
Finley, J. C. W., Maderdrut, J. L., and Petrusz, P.: The immunocytochemical localization of enkephalin in the central nervous system of the rat. J. Comp. Neurol. 198: 541-565, 1981.

156
Fischer-Ferraro, C., Nahmod, V. E., Goldstein, D. J., and Finkielman, S.: Angiotensin and renin in rat and dog brain. J. Exp. Med. 133: 353-361, 1971.
Fitzgerald, J. D. Beta-adrenoceptor antagonists. In P. A. Van Zwieten (Ed.), Handbook of Hypertension. Pharmacology of antihypertensive drugs. Amsterdam: Elsevier Science Publishers, 1984.
Fitzsimons, C. L. and Ciriello, J.: Selective Destruction of Parvocellular Neurons in the Paraventricular Nucleus Decreases Norepinephrine Turnover Rates in Kidney of Spontaneously Hypertensive Rats. Fed. Proc. 45: 875, 1986.
Fitzsimons, J. T.: The effect on drinking of peptide precursors and of shorter chain peptide fragments of angiotensin II injected into the rat's diencephalon. J. Physiol. 214: 295-303, 1971.
Freeman, R.H. and Rostorfer, H. H.: Hepatic changes in renin substrate biosynthesis and alkaline phosphatase activity in the rat. Am. J. Physiol. 223: 364-370, 1972.
Fujii, A. M. and Vatner, S. F.: Direct versus indirect pressor and vasoconstrictor actions of angiotensin in conscious dogs. Hypertension 7: 253-261, 1985.
Fuller, R. W.: Serotonergic stimulation of pituitary-adrenocortical function in rats. Neuroendocrinology 32: 118-127, 1981.
Fuller, R. W. and Snoddy, H. D.: Elevation of plasma corticosterone by swim stress and insulin-induced hypoglycemia in control and fluoxetine-pretreated rats. Endocrine Research Communications 4(1): 11-23, 1977.
Fuller, R. W. and Snoddy, H. D.: Effect of serotonin-releasing drugs on serum corticosterone concentration in rats. Neuroendocrinology 31: 96-100, 1980.
Fuller, R. W. and Snoddy, H. D.: Elevation of serum corticosterone concentrations in rats by pergolide and other dopamine agonists. Endocrinology 109: 1026-1032, 1981.
Furness, J. B., Heath, J. W., and Costa, M.: Aqueous aldehyde (Faglu) methods for the fluorescence histochemical localization of catecholamines and for ultrastructural studies of central nervous tissue. Histochemistry 57: 285-295, 1978.
Fuxe, K. , Agnati, L. MerloPich, E., Hokfelt, 189-192, 1983.
F.' T.'
Harfstrand, A., Zini, I., Tatemoto, and Mutt, V.: Acta Physiol. Scand.
K.' 118:

157
Fuxe, K., Ganten, D., Hokfelt, T., and Bohne, P.: Immunohistochemical evidence for the existence of angiotensin II containing nerve terminals in the brain and spinal cord of the rat. Neuroscience Letters 2: 229-234, 1976.
Fuxe, K., Ganten, D., Hokfelt, T., Locatelli, V., Poulsen, K., Stock, G., Rix, E., and Taugner, R.: Renin-like immunocytochemical activity in the rat and mouse brain. Neuroscience Letters 18: 245-250, 1980.
Fuxe, K., Hokfelt, T., Eneroth, P., Gustafsson, J. A., and Skett, P.: Prolactin: Localization in nerve terminals of the rat hypothalamus. Science 196: 899-900, 1977.
Galen, F. X., Devaux, C., Guyenne, T., Menard, J., and Corvol, P.: Multiple Forms of Human Renin- Purification and Characterization. The Journal of Biological Chemistry 254(11): 4848-4855, 1979.
Galosy, R. A., Crawford, I. L., and Thompson, M. E. Behavioral stress and cardiovascular regulation: Neural mechanisms. In Smith, Galosy and Weiss (Eds.), Circulation, Neurobiology, and Behavior. : Elsevier Science Publishing Co., 1982.
Ganong, W. F.: Neurotransmitters and pituitary function: regulation of ACTH secretion. Fed. Proc. 39: 2923-2930, 1980.
Ganong, W. F. and Barbieri, C. Neruoendocrine components in the regulation of renin secretion. In: Frontiers Neuroendocrinology, 1982.
Ganong, W. F., Chalett, J., Jones, H., Kaplan, S. L., Karteszi, M., Stith, R. D., and Van de Kar, L. D.: Further characterization of putative alpha-adrenergic receptors in brain that affect blood pressure and the secretion of ACTH, GH and renin in dogs. Endocrinologia Experimentalis 16: 191-205, 1982.
Ganong, W. F. , Rudo 1 ph, C. conponents in the regulation Hypertension 1: 207-218, 1979.
D., and Zimmermann, H.: Neuroendocrine of blood pressure and renin secretion.
Garvey, H. L. and Ram, N.: Comparative antihypertensive effects and tissue distribution of beta adrenergic blocking drugs. J. Pharmacol. Exp. Ther. 194: 220-233, 1975.
Giarcovich, S. and Enero, M.A.: Decreased brain serotonergic activity after acute propranolol. Eur. J. Pharmacol. 100: 123-125, 1984.
Gibbs, D. M.: Dissociation of oxytocin, vasopressin and corticotropin secretion during different types of stress. Life Sciences 35: 487-491, 1984.

158
Gibbs, D. M. and Vale, W.: Effect of the serotonin reuptake inhibitor fluoxetine on corticotropin-releasing factor and vasopressin secretion into hypophysial portal blood. Brain Research 280: 176-179, 1983.
Gibson, A., Hart, S. L., and Patel, S.: Effects of 6-hydroxydopamine-induced lesions of the paraventricular nucleus, and of prazosin, on the corticosterone response to restraint in rats. Neuropharmacology 25(3): 257-260, 1986.
Gilbert, F., Brazell, C., Tricklebank, M. D., and Stahl, S. M.: Activation of the 5-HTlA receptor subtype increases rat plasma ACTH concentration. European Journal of Pharmacology 147: 431-439, 1988.
Gillies, G.E., Linton, E.A., Lowry, P.J.: Corticotropin releasing activity of the new CRF is potentiated several times by vasopressin. Nature 299: 355-357, 1982.
Glavin, G. B.: Selective noradrenaline depletion markedly alters stress responses in rats. Life Sciences 37: 461-465, 1985.
Goldblatt, H., Lynch, J., Hanzal, R. F., and Summerville, W.W.: Studies on experimental hypertension. I. The production of persistent elevation of systolic blood pressure by means of renal ischemia. J. Exp. Med. 59: 347-379, 1934.
Goldman, C. K., Marino, L., and Leibowitz, S. F.: Postsynaptic alpha-2 noradrenergic receptors mediate feeding induced by paraventricular nucleus injection of norepinephrine and clonidine. European Journal of Pharmacology 115: 11-19, 1985.
Golin, R. M. A., Gotoh, E., Said, S. I., and Ganong, W. F.: Pharmacological evidence that the sympathetic nervous system mediates the increase in renin secretion produced by immobilization and head-up tilt in rats. Neuropharmacology 27(12): 1209-1213, 1988.
Goodwin, F. J. , Kirshman, J. D. , Sealey, J. E. , and Laragh, J. H. : Influence of the pituitary gland on sodium conservation, plasma renin and renin substrate concentrations in the rat. Endocrinology 86: 824-834, 1970.
Gordon, D. B. and Sullivan, P. H.: Acute effect of over transfusion on plasma renin activity in the rat. Endocrinology 85: 341-343, 1969.
Gotoh, E., Bahnson, T. D., Alper, R. H., and Ganong, W. F.: Role of the hypothalamus in the regulation of renin secretion in rats. The Physiologist 28(4): 41.14, 1985.
Gotoh, E., Golin, R. M. A., and Ganong, W. F.: Relation of the ventromedial nuclei of the hypothalamus to the regulation of renin secretion. Neuroendocrinology 47: 518-522, 1988.

159
Gotoh, E., Murakami, K., Bahnson, T. D., and Ganong, W. F.: Role of brain serotonergic pathways and hypothalamus in regulation of renin secretion. Am. J. Physiol. 253: Rl79-Rl85, 1987.
Gray, J. A. The Psychology of Fear and Stress. London: Weidenfeld and Nicolson, 1971.
Gray, J. A. The Neuropsychology of Anxiety. Oxford: Clarendon Press, 1982.
Green, R. H., Beatty, W. S., and Schwartzbaum, J. S.: Comparative effects of septo-hippocampal and caudate lesions on avoidance behavior in rats. Journal of Comparative and Physiological Psychology 64: 444-452, 1967.
Greenwood, F. C., Hunter, W. M., and Glover, J. S.: The preparation of 131I-labelled human growth hormone of high specific radioactivity. Biochem. J. 89: 114-123, 1963.
Gregory, T. J., Wallis, C. J., and Printz, M. P.: Regional changes in rat brain angiotensin following bilateral nephrectomy. Hypertension 4: 827-838, 1982.
Grossman, S. P., Grossman, L., and Walsh, L.: Functional organization of the rat amygdala with respect to avoidance behavior. J. Comp. Physiol. Psychol. 88: 829-850, 1975.
Guillaume, V., Conte-Devolx, B., bute, N. , Grino, M. , Alonso, tropin-releasing factor release mediated by brain catecholamines. 143-146, 1987.
Szafarczyk, A., Malaval, F., Pares-HerG., and Oliver, C.: The corticoin rat hypophysial portal blood is Neuroendocrinology 46:
Gustafsson, J. A., Okret, S., Wikstrom, A. C., Anderson, B., Radojcic, M., Wrange, 0., Sachs, W., and Doupe, A. J. On the use of poly- and monoclonal antibodies in studies on the structure and function of the glucocorticoid receptor. In H. Eriksson and J. A. Gustafsson (Eds.), Steroid hormone receptors: structure and function. Amsterdam: Elsevier, 1983.
Gutman, F. D. , Tagawa, H. , Haber, E. , and Barger, A. C. : Renal arterial pressure, renin secretion and blood pressure control in trained dogs. Amer. J. Physiol. 224: 66-72, 1973.
Guyton, A. C. The autonomic nervous system: the adrenal medulla. In (Ed.), Medical Physiology. Philadelphia: W.B. Saunders Co., 1981.
Haber, C., Loerner, D., Page, L. B., Kliman, B., and Purnode, A.: Application of a radioimmunoassay for angiotensin I to the physiologic measurements of plasma renin activity in normal human subjects. Journal of Clinical Endocrinology 29: 1329-1355, 1969.

160
Hall, C. S.: Temperament: a survey of animal studies. Psycho!. Bull. 38: 909-943, 1941.
Harfstrand, A., Eneroth, P., Agnati, L., and Fuxe, K.: Further studies on the effects of central administration of neuropeptide Y on neuroendocrine function in the male rat: relationship to hypothalamic catecholamines. Regulatory Peptides 17: 167-179, 1987.
Harfstrand, A., Fuxe, K., Agnati, L. F., Eneroth, P., Zini, I., Zoli, M., Andersson, K., and von Euler, G.: Studies on neuropeptide Y-catecholamine interactions in the hypothalamus and in the forebrain of the male rat. Relationship to neuroendocrine function. Neurochem. Int. 8(3): 355-376, 1986.
Harfstrand, A., Fuxe, K., Cintra, A., Agnati, L. F., Zini, I., 'Wikstrom, A. C., Okret, S., and Yu, Z. Y.: Glucocorticoid receptor immunoreactivity in monoaminergic neurons of rat brain. Proc. Natl. Acad. Sci. (USA) 83: 9779-9783, 1986.
Harris, G. 'W.: Neural Control of the Pituitary Gland. Physiol. Rev. 28: 139, 1948.
Harrop, G. A., Soffer, L. J., Ellsworth, R., and Trescher, J. H.: Studies on the suprarenal cortex. III. Plasma electrolytes and electrolyte excretion during suprarenal insufficiency in the dog. J. Exp. Med. 58: 17-38, 1933.
Hatton, G. I., Hutton, U. E., Hoblitzell, E. R., and Armstrong, 'W. E.: Morphological evidence for two populations of magnocellular elements in the rat paraventricular nucleus. Brain Research 108: 187-193, 1976.
Hauger, R. L., Aguilera, G., Baukal, A., and Catt, K. J.: Characterization of angiotensin II receptors in the anterior pituitary gland. Mol. Cell. Endocrinol. 25: 203-212, 1982.
Healy, D. P., Munzel, P. A., and Insel, P. A.: Localization of the beta-1 and beta-2-adrenergic receptors in rat kidney by autoradiography. Gire. Res. 57: 278-284, 1985.
Herman, J. P. and 'Wiegand, S. J.: Ibotenate-induced cell death in the hypothalamic paraventricular nucleus: differential susceptibility of magnocellular and parvicellular neurons. Brain Res. 383: 367-372, 1986.
Hetherington, A. 'W. and Ranson, S. 'W.: The spontaneous activity and food intake of rats with hypothalamic lesions. Am. J. Physiol. 136: 609-617, 1942.
Hiwada, K., Tanaka, H., and Kokubu, T.: The influence of nephrectomy, uretal ligation, and of estradiol on plasma renin substrate in unilaterally nephrectomized rats. Pflugers Arch. 365: 177-182, 1976.

161
Hokfelt, T., Fuxe, K., and Goldstein, M.: Immunohistochemical studies on monoamine-containing cell systems. Brain Research 62: 461-469, 1973.
Hokfelt, T., Fuxe, K., Goldstein, M., and Johansson, O.: Immunohistochemical evidence for the existence of adrenaline neurons in the rat brain. Brain Research 66: 239-251, 1974.
Hokfelt, T., Fuxe, K., Johansson, 0., Jeffcoate, S., Distribution of thyrotropin-releasing hormone (TRH) in the system as revealed with immunohistochemistry. Eur. J. 389-392, 1975.
and White, N.: central nervous Pharmacol. 34:
Hokfelt, T., Johansson, 0., and Goldstein, M.: Chemical anatomy of the brain. Science 225: 1326-1334, 1984.
Hollenberg, N. K., Williams, G. H., and Adams, D. F.: Essential hypertension: Abnormal renal vascular and endocrine responses to a mild psychologic stimulus. Hypertension 3: 11-17, 1981.
Holmberg, G. and Gershon, S. : Autonomic and psychiatric effects of yohimbine hydrochloride. Psychopharmacologia 2: 93-106, 1961.
Horky, K., Rojo-Ortega, J. M., Rodriguez, J., Boucher, R., and Genest, J.: Renin, renin substrate and angiotenin I - converting enzyme in the lymph of rats. Am. J. Physiol. 220(2): 307-311, 1971.
Hosoya, Y. and Matsushita, M.: Identification and distribution of the spinal and hypophysial projection neurons in the paraventricular nucleus of the rat. A light and electron microscopic study with the horseradish peroxidase method. Exp. Brain Res. 35: 315-331, 1979.
Hunt, H. F. and Otis, L. S.: Conditioned and unconditioned emotional defecation in the rat. J. Comp. Physiol. Psychol. 46: 378-382, 1953.
Husain, M. K., Manger, W. M., Rock, T. W., Weiss, R. J., and Frantz, A. G.: Vasopressin release due to manual restraint in the rat: Role of body compression and comparison with other stressful stimuli. Endocrinology 104: 641-644, 1979.
Iimori, K., Tanaka, M., Kohno, Y., Ida, Y., Nakagawa, R., Hoaki, Y., Tsuda, A., and Nagasaki, N.: Psychological stress enhances noradrenaline turnover in specific brain regions in rats. Pharmacol. Biochem. Behav. 16(4): 637-640, 1982.
Inagami, T. and Murakami, K.: Prorenin. Biomed. Res. 1: 456-475, 1980. Ingelfinger, J. R., Pratt, R. E., Ellison, K., and Dzau, V. J.: Sodium regulation of angiotensinogen mRNA expression in rat kidney cortex and medulla. J. Clin. Invest. 78: 1311-1315, 1986.

162
Ingle, D. J., Higgins, G. M., and Kendall, E. C.: Atrophy of the adrenal cortex in the rat produced by administration of large amounts of cortin. Anat. Rec. 71: 363-372, 1938.
Itoh, S., Carretero, A., and Murray, R. D.: Possible role of adenosine in the macula densa mechanism of renin release in rabbits. J. Clin. Invest. 76: 1412-1417, 1985.
Ixart, G. , Alonso, G. , Szafarczyk, A. , Malaval, F. , Nouguier- Soule, J. , and Assenmacher,!.: Adrenocorticotropic Regulations after Bilateral Lesions of the Paraventricular or Supraoptic Nuclei and in Brattleboro Rats. Neuroendocrinology 35: 270-276, 1982.
Ixart, G., Barbanel, G., Conte-Devoix, B., Grino, M., Oliver, C., and Assenmacher, I.: Evidence for basal and stress-induced release of corticotropin releasing factor in the push-pull cannulated median eminence of conscious free-moving rats. Neuroscience Letters 74: 85-89, 1987.
Jacobowitz, D. M. and O'Donohue, T.: Alpha-melanocyte stimulating hormone: Immunohistochemical identification and mapping in neurons of rat brain. Proc. Natl. Acad. Sci. (USA) 75: 6300-6304, 1978.
James, W. The Principles of Psychology. New York: Henry Holt, 1890.
Jennes, L. and Stumpf, W. E.: LHRH-systems in the brain of the golden hamster. Cell Tiss. Res. 209: 239-256, 1980.
Jezova, D., Jurcovicova, J., Vigas, M., Murgas, K., and Labrie, F.: Increase in plasma ACTH after dopaminergic stimulation in rats. Psychopharmacology 85: 201-203, 1985.
Jindra, A., Kvetnansky, R., Belova, T. I., andSudakov, K. V. Effect of acute and repeated immobilization stress on plasma renin activity, catecholamines and corticosteroids in Wistar and August Rats. In E. Usdin and I. Kopin (Eds.), Catecholamines and Stress. North Holland: Elsevier, 1980.
Johnson, J. A., Davis, J. 0., and Witty, R. T.: Effects of catecholamines and renal nerve stimulation on renin release in the nonfiltering kidney. Circ. Res. 29: 646-653, 1971.
Johnson, M. D.: Circulating epinephrine stimulates renin secretion in anesthetized dogs by activation of extrarenal adrenoceptors. Am. J. Physiol. 246: F676-F684, 1984.
Johnson, M. D., Shier, D. N., and Barger, A. C.: Circulating catecholamines and control of plasma renin activity in conscious dogs. Am. J. Physiol. 236(3): H463-H470, 1979.

163
Johnston, C. A., Spinedi, E. J., and Negro-Vilar, A.: Effect of acute ether stress on monoamine metabolism in median eminence and discrete hypothalamic nuclei of the rat brain and on anterior pituitary hormone secretion. Neuroendocrinology 41: 83-88, 1985.
Jones, M. T., Millhouse, E. W., and Burden, J.: Effects of various putative neurotransmitters on the secretion of corticotropin-releasing hormone from the rat hypothalamus in vitro -a model of the neurotransmitters involved. J. Endocr. 69: 1-10, 1976.
Joseph, S. A.: Immunoreactive adrenocorticotropin in rat brain: A neuroanatomical study using antiserum generated against synthetic ACTH. Am. J. Anat. 158: 533-548, 1980.
Joseph, S. A. and Sternberger, L. A.: The unlabeled antibody method. Contrasting color staining of beta-lipotropin and ACTH-associated hypothalamic peptides without antibody removal. J. Histochem. Cytochem. 27: 1430-1437, 1979.
Kahn, D., Abrams, G. M., Zimmerman, E. A., Carraway, R., and Leeman, S. E.: Neurotensin neurons in the rat hypothalamus: An immunohistochemical study. Endocrinology 107: 47- 54, 1980.
Kannan, H. and Yamashita, H.: Connections of neurons in the region of the nucleus tractus solitarius with the hypothalamic paraventricular nucleus: their possible involvement in neural control of the cardiovascular system in rats. Brain Research 329: 205-212, 1985.
Kant, G. J., Mougey, E. H., Pennington, L. L., and Meyerhoff, J. L.: Graded Footshock Stress Elevates Pituitary Cyclic AMP and Plasma Bendorphin, B-LPH, Corticosterone and Prolactin. Life Sciences 33: 2657-2663, 1983.
Kaplan, N. M.: The biosynthesis angiotensin II, adrenocorticotropin, 2029-2039, 1965.
of adrenal steroids: effects of and potassium. J. Clin. Invest. 44:
Karteszi, M., Van de Kar, L. D., Makara, G., Stark, E., and Ganong, W.F.: Evidence that the mediobasal hypothalamus is involved in serotonergic stimulation of renin secretion. Neuroendocrinology 34: 323-326, 1982.
Keeton, T. K. and Campbell, W. B.: The pharmacologic alteration of renin release. Pharmacological Reviews 32(2): 81-227, 1980.
Keil, L. C., Summy-Long, J., and Severs, W. B.: Release of vasopressin by angiotensin II. Endocrinology 96: 1063-1065, 1975.
Keller-Wood, M., Kimura, B., Shinsako, J., and Phillips, M. I.: Interaction between CRF and angiotensin II in control of ACTH and adrenal steroids. Am. J. Physiol. 250: R396- R402, 1986.

164
Kety, S. S. The biogenic amines in the central nervous system: Their possible roles in arousal, emotion and learning. In F. 0. Schmitt (Ed.), The Neurosciences Second Study Program. New York: Rockefeller University Press, 1970.
Khayyall, M., MacGregor, J., Brown, J. J., Lever, A. F., and Robertson, J.I.S.: Increase of plasma renin-substrate concentration after infusion of angiotensin in the rat. Clin. Sci. 44: 87-90, 1973.
Kidman, A.: Stress, cognition and the nervous system. Neurochem. Int. 6(6): 715-720, 1984.
Kimura, H., McGeer, P. L., and McGeer, E. G.: The central cholinergic system studied by choline acetyltransferase immunohistochemistry in the cat. J. Comp. Neurol. 200: 151-201, 1981.
Kiss, J. Z., Mezey, E., and Skirboll, L.: Corticotropin-releasing factor -immunoreactive neurons of the paraventricular nucleus become vasopressin positive after adrenalectomy. Proc. Natl. Acad. Sci. (USA) 81: 1854-1858, 1984.
Klett, C. and Heckenthal, E.: and secretion by angiotensin 2027-2047, 1987.
Induction of angiotensinogen synthesis II. Clin. and Exper. Hyper. A9(12):
Kling, A. and Hutt, P. J.: Effect of hypothalamic lesions on the amygdala syndrome in the cat. A.M.A. Arch. Neurol. Psychiat. 79: 511-517, 1958.
Knepel, W., Homolka, L., Vlaskovska, M., and Nutto, D.: Stimulation of adrenocorticotropinjbeta-endorphin release by synthetic ovine corticotropin-releasing factor in vitro. Neuroendocrinology 38: 344-350, 1984.
Knepel, W. and Meyer, D. K.: Role of the renin-angiotensin system in isoprenaline-induced vasopressin release. J. Cardiovasc. Pharmacol. 2: 815-824, 1980.
Knigge, K. M.: Adrenocortical response to stress in rats with lesions in hippocampus and amygdala. Proc. Soc. Exp. Biol. Med. 108: 18-21, 1961.
Kobayashi, R. M., Palkovits, M., Jacobowitz, D. M., and Kopin, I. J.: Biochemical Mapping of the Noradrenergic Projection from the Locus Coeruleus. Neurology 25: 223-233, 1975.
Koenig, J. I. , Gudelsky, G. A. , and Meltzer, H. Y. : Stimulation of corticosterone and beta-endorphin secretion in the rat by selective 5-HT receptor subtype activation. Europ. J. Pharmacol. 137: 1-8, 1987.
Koepke, J. P. and DiBona, G. F.: Central adrenergic receptor control of renal function in conscious hypertensive rats. Hypertension 8: 133-141, 1986.

16S
Kohno, Y., Tanaka, M., Nakagawa, R., Toshima, N., and Nagasaki, N.: Regional distribution and production rate of 3-methoxy-4- hydroxyphenylethyleneglycol sulphate (MHPG-S04) in rat brain. J. Neurochem. 36: 286-289, 1981.
Koizumi, K. and Yamashita, H.: Studies of antidromically identified neurosecretory cells of the hypothalamus by intracellular and extra-cellular recordings. J. Physiol. (London) 221: 683-70S, 1972.
Korf, J., Aghajanian, G. K., and Roth, R.H.: Stimulation and destruction of the locus coeruleus: Opposite effects on 3-methoxy-4-hydroxyphenylglycol sulfate levels in the rat cerebral cortex. Eur. J. Pharmacol. 21: 30S-310, 1973.
Krettek, J. E. and Price, J. L.: Amygdaloid projections to subcortical structure within the basal forebrain and the brainstem in the rat and the cat. J. Comp. Neurol. 178: 2SS-280, 1978.
Krieg, W. J. S.: The hypothalamus of the albino rat. J. Comp. Neurol. SS: 19-89, 1932.
Krieger, D. T. and Hauser, H.: Comparison of synchronization of circadian corticosteroid rhythms by photoperiod and food. Proc. Natl. Acad. Sci. (USA) 1975: 1577-1581, 1978.
Kuhar, M. J.: Neuroanatomical substrates of anxiety: a brief survey. T.I.N.S. July: 307-311, 1986.
Kuypers, H. G. J. M. and Maisky, V. A.: Retrograde axonal transport of horseradish peroxidase from spinal cord to brainstem cell groups in the cat. Neuroscience Letters 1: 9, 1973.
Kvetnansky, R., Kopin, I. J., and Saavedra, J.M.: Changes in epinephrine in individual hypothalamic nuclei after immobilization stress. Brain Research lSS: 387-390, 1978.
Leckie, B. J.: Inactive renin: an attempt at a perspective. Clinical Science 60: 119-130, 1981.
Leenin, F. H. and Shapiro, A. P.: Effects of intermittent electric shock on plasma renin activity in rats. Proc. Soc. Exp. Biol. Med. 146: 534-S38, 1974.
Leibowitz, S. F.: Pattern of drinking and feeding produced by hypothalamic norepinephrine injection in the satiated rat. Physiol. Behav. 14: 731-742, 1975.
Leibowitz, S. F., Diaz, S., and Spencer, L.: Adrenergic and neuropeptide Y systems in the hypothalamic paraventricular nucleus affect circulating levels of corticosterone and glucose. Neuroscience Abstracts 12(2): 782, 1986.

166
Leibowitz, S. F., Hammer, N. J., and Chang, K.: Hypothalamic paraventricular nucleus lesions produce overeating and obesity in the rat. Physiol. Behav. 27: 1031-1040, 1981.
Leibowitz, S. F., Roland, C. R., Hor, L., and Squillari, V.: Noradrenergic feeding elicited via the paraventricular nucleus is dependent upon circulating corticosterone. Physiol. Behav. 32: 8S7-864, 1984.
Lemmer, B., Winkler, H., Ohm, T., and Fink, M.: Chronopharmacokinetics of beta-receptor blocking drugs of different lipophilicity (propranolol, metoprolol, sotalol, atenolol) in plasma and tissues after single and multiple dosing in the rat. Naunyn-Schmiedeberg's Arch Pharmacol 330: 42-49, 198S.
Lenz, H. J., Raedler, A., Greten, H., and Brown, M. R.: CRF initiates biological actions within the brain that are observed in response to stress. Am. J. Physiol. 2S2: R34-R39, 1987.
Leranth, C., Antoni, F. A., and Palkovits, M.: Ultrastructural demonstration of ovine CRF-like immuno-reactivity (oCRF-LI) in the rat hypothalamus: processes of magnocellular neurons establish membrane specializations with parvocellular neurons containing oCRF-LI. Regul. Peptides 6: 179-188, 1983.
Leroux, P. and Pelletier, G.: Radioautographic study of binding and internalization of corticotropin-releasing factor by rat anterior pituitary corticotrophs. Endocrinology 114: 14-21, 1984.
Levin, M. C., Sawchenko, P. E., Howe, P. R. C., Bloom, S. R., and Polak, J.M.: Organization of galanin-immunoreactive inputs to the paraventricular nucleus with special reference to their relationship to catecholaminergic afferents. The Journal of Comparative Neurology 261: 562-S82, 1987.
Li, C. H. , Evans, H. M. , and Simpson, M. E. : Adrenocorticotropin hormone. J. Biol. Chem. 149: 413-424, 1943.
Lightman, S. L., Todd, K., and Everitt, B. J.: Ascending noradrenergic projections from the brainstem: Evidence for a major role in the regulation of blood pressure and vasopressin secretion. Experimental Brain Research SS: 14S-151, 1984.
Lind, R. W., Swanson, L. W., Bruhn, T. 0., and Ganten, D.: The distribution of angiotensin II- immunoreactive cells and fibers in the paraventriculo-hypophysial system of the rat. Brain Research 338: 81-89, 198S.
Lindvall, O. and Bjorklund, A.: The organization of ascending catecholamine neuron systems in the rat brain. Acta Physiol. Scand. 412: 1-48, 1974.

167
Liposits, Z., Gores, T., Setalo, G., Lengvari, I., Flerko, B., Vigh, S., and Schally, A.V.: Ultrastructural characteristics of immunolabelled, corticotropin releasing factor (CRF)-synthesizing neurons in the rat brain. Cell Tissue Res. 229: 191-196, 1983.
Lipos its, Z. , Lengvari, I. , Vigh, S. , Gores, T. , Setalo, G. , and Flerko, B. The paraventriculo- infundibular corticotropin releasing factor (CRF)-Immunoreactive System of the Rat- I. Morphological Aspects. In K. W. McKerns and V. Pantie (Eds.), Neuroendocrine Correlates of Stress. New York: Plenum Press, 1985.
Liposits, Z. and Paull, W. K.: Ultrastructural alterations of the paraventriculo-infundibular corticotropin releasing factor (CRF)-immunoreactive neuronal system in long term adrenalectomized rats. Peptides 6: 1021-1036, 1985.
Liposits, Z., Phelix, C., and Paull, W. K.: Electron microscopic analysis of tyrosine hydroxylase, dopamine-beta-hydroxylase and phenylethanolamine-N-methyl transferase immunoreactive innervation of the hypothalamic paraventricular nucleus in the rat. Histochemistry 84: 105-120, 1986.
Liposits, Z., Phelix, C., and Paull, W. K.: Adrenergic innervation of corticotropin releasing factor (CRF)- synthesizing neurons in the hypothalamic paraventricular nucleus of the rat. Histochemistry 84: 201-205, 1986.
Liposits, Z., Phelix, C., and Paull, W. K.: Synaptic interaction of serotonergic axons and corticotropin releasing factor (CRF) synthesizing neurons in the hypothalamic paraventricular nucleus of the rat. A light and electron microscopic immunocytochemical study. Histochemistry 86: 541-549, 1987.
Liposits, Z., Uht, R. M., Harrison, R. W., Gibbs, F. P., Paull, W. K., and Bohn, M. C.: Ultrastructural localization of glucocorticoid receptor (GR) in hypothalamic paraventricular neurons synthesizing corticotropin releasing factor (CRF). Histochemistry 87: 407-412, 1987.
Ljungdahl, A., Hokfelt, T., and Nilsson, G.: Distribution of substance P-like immunoreactivity in the central nervous system of the rat. I. Cell bodies and nerve terminals. Neuroscience 3: 861-944, 1978.
Loeb, R. F., Atchley, D. W., Benedict, E. M., and Leland, J.: Electrolyte balance studies in adrenalectomized dogs with particular reference to the excretion of sodium. J. Exp. Med. 57: 775-792, 1933.
Loeffler, J. R., Stockigt, J. R., and Ganong, W. F.: Effect of alphaand beta-adrenergic blocking agents on the increase in renin secretion produced by stimulation of the renal nerves. Neuroendocrinology 10: 129-138, 1972.

168
Loizou, L. A.: Projections of the nucleus locus coeruleus in the albino rat. Brain Research 15: 563-566, 1969.
Long, C. N. H., Katzin, B., and Fry, E. G.: Adrenal cortex and carbohydrate metabolism. Endocrinology 26: 309-344, 1940.
Lorens, S. A. and Guldberg, H. C.: Regional 5-hydroxytryptamine following selective midbrain raphe lesions. Brain Research 78: 45-56, 1974.
Lorens, S. A., Mitsushio, H., and Van de Kar, L. D. Effects of the 5-HTlA agonist ipsapirone on the behavioral, endocrine and neurochemical responses to conditioned fear. In T. Archer,P. Bevan,C. Broekkamp and A. Cools (Eds.), Behavioral Pharmacology of 5-HT. : In Press, 1989.
Lorens, S. A. and Van de Kar, L. D.: Differential effects of serotonin (5-HTlA and 5-HT2) agonists and antagonistson renin and corticosterone secretion. Neuroendocrinology 45: 305-310, 1987.
Luiten, P. G. M., ter Horst, G. J., Karst, H., and Steffens, A. B.: The course of paraventricular efferents to autonomic structures in medulla and spinal cord. Brain Research 329: 374-378, 1985.
Madison, D. V. and Nicoll, R. A.: Actions of noradrenaline recorded intracellularly in rat hippocampal CAl pyramidal neurons, in vitro. J. Physiol. 372: 221-244, 1986.
Makara, G. B., Stark, E., Kapocs, G., and Antoni, F. A.: Long-term effects of hypothalamic paraventricular lesion on CRF content and stimulated ACTH secretion. Am. J. Physiol. 250: E319-E324, 1986.
Makara, G. B., Stark, E., Karteszi, M., Palkovits, M., and Rappay, G.: Effects of paraventricular lesions on stimulated ACTH release and CRF in stalk-median eminence of the rat. Am. J. Physiol. 240: E441-E446, 1981.
Manuck, S. B., Giordani, B., McQuaid, K. J., and Garritz, S. J.: Behaviorally-induced cardiovascular reactivity among sons of reported hypertensive normotensive patients. J. Psychosom. Res. 25: 261, 1981.
Maran, J. W. and Yates, F. E.: Cortisol secretion during intrapituitary infusion of angiotensin II in conscious dogs. Am. J. Physiol. 233: E273-E285, 1977.
Marino, L.A., DeBellis, M. D., and Leibowitz, S. F.: Alpha-2 adrenergic receptors in the paraventricular nucleus mediate feeding induced by norepinephrine and clonidine. Soc. Neurosci. Abstr. 9: 467, 1983.
Markowska, A., Bakke, H.K., Walther, B., and Ursin, H.: Comparison of electrolytic and ibotenic acid lesions in the lateral hypothalamus. Brain Research 328: 313-323, 1985.

Mason, J. W.: Plasma 17-corticosteroid stimulation of the amygdaloid complex in Physiol. 196: 44-48, 1959.
169
levels during electrical conscious monkeys. Am. J.
Mason, J. W., Maher, J. T., Hartley, L. H., Mougey, E. H., Perlow, M. J., and Jones, L.G. Selectivity of corticosteroid and catecholamine response to various natural stimuli. In G. Serban (Ed.), Psychopathology of Human Adaptation. New York: Plenum Press, 1976.
Mcintyre, M. and Stein, D. G.: Differential effects of one- vs. two-stage amygdaloid lesions on activity, exploratory, and avoidance behavior in the albino rat. Behav. Biol. 9: 451-465, 1973.
McKellar, S. and Loewy, A. D.: Organization of some brain stem afferents to the paraventricular nucleus of the hypothalamus in the rat. Brain Res 217: 351-357, 1981.
Meister, B., Hokfelt, T., and Staines, W.: Presence of glutamic acid decarboxylase in some corticotropin-releasing factor- (CRF-) containing neurons of the paraventricular nucleus. Acta Physiol. Scand. 131: 471-473, 1987.
Meldrum, M. J., Xue, C. S., Badino, L., and Westfall, T. C.: Angiotensin facilitation of noradrenergic neurotransmission in central tissues of the rat: effects of sodium restriction. Journal of Cardiovascular Pharmacology 6: 989-995, 1984.
Menard, J. and Catt, K. J.: Measurement of renin activity, concentration and substrate in rat plasma by radioimmunoassay of angiotensin I. Endocrinology 90: 422-430, 1972.
Merchenthaler, I., Vigh, S., Petrusz, P., and Schally, A. V.: Immunocytochemical localization of corticotropin releasing factor (CRF) in the rat brain. Am. J. Anat. 165: 385, 1982.
Merchenthaler, I., Vigh, S., Petrusz, P., and Schally, A. V.: The paraventriculo-infundibular corticotropin releasing factor (CRF) pathway as revealed by immunocytochemistry in long-term hypophysectomized or adrenalectomized rats. Regulatory Peptides 5: 295-305, 1983.
Mermet, C. C. and Gonon, F. G.: Ether stress stimulates noradrenaline release in the hypothalamic paraventricular nucleus. Neuroendocrinology 47: 75-82, 1988.
Meyer, D. K., Phillips, M. I., and Eiden, L.: Studies on the presence of angiotensin II in rat brain. J. Neurochem. 38: 816-820, 1982.
Meyerhoff, J. L., Mougey, E. H., and Kant, G. J.: Paraventricular lesions abolish the stress-induced rise in pituitary cyclic adenosine monophosphate and attenuate the increase in plasma levels of proopiomelanocortin-derived peptides and prolactin. Neuroendocrinology 46: 222-230, 1987.

170
Michelakis, A. M. andMizukoshi, H.: Distribution and disappearance rate of renin in man and dog. J. Clin. Endocrinol. Metab. 33: 27-34, 1971.
Middlemiss, D. N., Blakeborough, L., and Leather, S. R.: Direct evidence for an interaction of beta-adrenergic blockers with the serotonin receptor. Nature 267: 289-290, 1977.
Mills, F. J.: The endocrinology of stress. Aviat. Space Environ. Med. 56: 642-650, 1985.
Modlinger, R. S., Schonmuller, J.M., and Arora, S. P.: Stimuation of aldosterone, renin and cortisol by tryptophan. J. Clin. Endocrinol. Metab. 48: 599-603, 1979.
Moga, M. M. and Gray, T. S.: Evidence for corticotropin-releasing factor, neurotensin and somatostatin from the central nucleus of the amygdala to the parabrachial nucleus in the rat. J. Comp. Neurol. 241: 275, 1985.
Moore, R. Y. and Bloom, F. E.: Central catecholamine neuron systems: anatomy and physiology of the norepinephrine and epinephrine systems. Ann. Rev. Neurosci. 2: 113-168, 1979.
Morris, B. J., Davis, J. 0., Zatzman, M. L., and Williams, G. M.: The renin-angiotensin-aldosterone system in rabbits with congestive heart failure produced by aortic constriction. Gire. Res. 40: 275-282, 1977.
Mukherjee, A., Kulkarni, P., Mccann, S. M., and Negro-Vilar, A.: Evidence for the presence and characterization of angiotensin II receptors in rat anterior pituitary membranes. Endocrinology 110: 665-667, 1982.
Mulder, G. H. and Smelik, P. G.: A perfusion technique for the study of sites of action of glucocorticoids in rat hypothalamus-pituitary-adrenal system in vitro. Endocrinology 100: 1143-1152, 1977.
Nakani, T., Audhya, T., Kani, N., and Hollander, C. S.: Evidence for a role of endogenous corticotropin-releasing factor in cold, ether, immobilization and traumatic stress. Proc. Natl. Acad. Sci. (USA) 82: 1247-1251, 1985.
Nasjletti, A. and Masson, G. M. C.: Effects of corticosteroids on plasma angiotensinogen and renin activity. Am. J. Physiol. 217(5): 1396-1400, 1969.
Nasjletti, A. and Masson, G. M. C.: Stimulation of angiotensinogen formation by renin and angiotensin. Proc. Soc. Exp. Biol. Med. 142: 307-310, 1973.
Natcheff, N., Logofeton, A., and Tzaneva, N.: Hypothalamic control of plasma renin activity. Pflugers Archiv. 371: 279-285, 1977.

171
Nicoll, R. A.: Neurotransmitters can say more than just "yes" or "no". Trends in NeuroSciences 5(11): 269-374, 1982.
Nielsen, A. H. and Poulsen, K.: Evidence for a predominant renal secretion and clearance of inactive plasma renin, studied by in vivo inhibition of protein synthesis. Journal of Hypertension 6: 129-137, 1988.
Norgren, R.: Projections from the nucleus of the solitary tract in the rat. Neuroscience 3: 207-218, 1978.
Oates, H. F., Fretten, J. A., and Stokes, G. S.: Disappearance rate of circulating renin after bilateral nephrectomy in the rat. Clin. Exp. Pharmacol. Physiol. 1: 547-549, 1974.
Olds, J. and Olds, M. In F. Barron {Ed.), New Directions in Psychology. New York: Holt, Rinehart and Winston, 1965.
Olivecrona, H.: Paraventricular nucleus and pituitary gland. Acta Physiol. Scand. 40(136): 1-178, 1957.
Olschowka, J. A., O'Donohue, T. L., Mueller, G. P., and Jacobowitz, D. M.: Hypothalamic and extrahypothalamic distribution of CRF-like immunoreactive neurons in the rat brain. Neuroendocrinology 35: 305-308, 1982.
Ono, N., Samson, W. K., Mc Donald, J. K., Lumpkin, M. D., Bedran de Castro, I. C., and Mc Cann, S. M.: Effect of intravenous and intraventricular injection of antisera directed against corticotropin-releasing factor in the secretion of anterior pituitary hormones. Proc. Natl. Acad. Sci. (USA) 88: 7787-7790, 1985.
Oparil, S., Vassaux, C., Sander, C. A., and Haber, E.: Role of renin in acute postual homeostasis. Circulation 41: 89-95, 1970.
Osborn, J. L., DiBona, G. F., and Thames, M. D.: Beta-1 receptor mediation of renin secretion elicited by low-frequency renal nerve stimulation. J. Pharmacol. Exp. Ther. 216: 265-269, 1981.
Osmond, D. H., Wilczynski, E. A., and Loh, A. Y.: Peculiarities of plasma prorenin measurements in man, dog, and rat, and their theoretical implications. Clin. Exp. Hypertension 4: 2213-2230, 1982.
Padfield, P. L. and Morton, J. J.: Effects of angiotensin II on arginine-vasopressin in physiological and pathological situations in man. J. Endocr. 74: 251-259, 1977.
Palkovi ts, M. : Catecholamines in the hypothalamus: An anatomical review. Neuroendocrinology 33: 123-128, 1981.
Palkovits, M., Brownstein, M., Saavedra, J. M., and Axelrod, J.: Norepinephrine and dopamine content of hypothalamic nuclei of the rat. Brain Research 77: 137-149, 1974.

172
Palkovits, M., Eskay, R. L., and Antoni, F. A.: Atrial natriuretic peptide in the median eminence is of paraventricular nucleus origin. Neuroendocrinology 46: 542-544, 1987.
Palkovits, M., Saavedra, J. M., Jacobowitz, D. M., Kizer, J. S., Zaborszky, I., and Brownstein, M. J.: Serotonergic innervation of the forebrain: effect of lesions on serotonin and tryptophan hydroxylase levels. Brain Research 130: 121-134, 1977.
Parent, A., Descarries, L., and Beaudet, A.: Organization of ascending serotonin systems in the adult rat brain. A radioautographic study after intraventricular administration of 3H 5-hydroxytryptamine. Neuroscience 6: 45-138, 1981.
Parfrey, P. S., Markandu, N. D., Roulston, J. E., Jones, B. E., Jones, J. C., and MacGregor, G. A.: Relation between arterial pressure, dietary sodium intake, and renin system in essential hypertension. Br. Med. J. 283: 94-97, 1981.
Paris, J. M., Lorens, S. A., Van de Kar, L. D., Urban, J. H., Richardson-Morton, K. D., and Bethea, C. L.: A comparison of acute stress paradigms: Hormonal responses and hypothalamic serotonin. Physiol Behav 39(1): 33-43, 1987.
Passo, S. S., Assaykeen, T. A., Goldfien, A., and Ganong, W. F.: Effect of alpha- and beta-adrenergic blocking agents on the increase in renin secretion produced by stimulation of the medulla oblongata in dogs. Neuroendocrinology 7: 97-104, 1971.
Paull, W. K. and Gibbs, F. P.: The corticotropin releasing factor (CRF) neurosecretory system in intact, adrenalectomized, and adrenalectomized-dexamethasone treated rats. Histochemistry 78: 303-316, 1983.
Pavlov, I. P. Conditioned Reflexes, translated and edited by G.V. Anrep. Oxford: Oxford University Press, 1927.
Peach, M. J. Adrenal Medulla. In I. M. Page and F. M. Bumpus (Eds.), Angiotensin. New York: Springer-Verlag, 1974.
Peach, M. J., Bumpus, F. M., and Khairallah, P.A.: Release of adrenal catecholamines by angiotensin I. J. Pharmacol. Exp. Ther. 176(2): 366-376, 1971.
Pearlmutter, A. F., Rapino, E., and Saffran, M.: A semiautomated in vitro assay for CRF: activities of peptides related to oxytocin and vasopressin. Neuroendocrinology 15: 106-119, 1974.
Perez de la Mora, M., Possani, L. D., Tapia, R., Teran, L., Palacios, R., Fuxe, K., Hokfelt, T., and Ljungdahl, A.: Demonstration of central gamma-aminobutyrate-containing nerve terminals by means of antibodies against glutamate decarboxylase. Neuroscience 6: 875-895, 1981.

173
Petrusz, P., Merchenthaler, I., Maderdrut, J. L., and Heitz, P. U.: Central and peripheral distribution of corticotropin-releasing factor. Fed. Proc. 44: 229-23S, 198S.
Pettinger, W. A., Augusto, L., and Leon, A. S. Alteration of renin release by stress and adrenergic receptor and related drugs in unanesthetized rats. In C. M. Bloor (Ed.), Comparative Pathophysiology of Circulatory Disturbances. New York: Plenum Press, 1972.
Pettinger, W. A. and Keeton, T. K.: Altered renin release and propranolol potentiation of vasodilatory drug hypotension. J. Clin. Invest. SS: 236-243, 197S.
Pettinger, W. A., Keeton, T. K., Campbell, W. B., and Harper, D. C.: Evidence for a renal alpha-adrenergic receptor inhibiting renin release. Circ. Res. 38: 338-346, 1976.
Phillips, M. I. and Stenstrom, B.: An analysis of angiotensin II and III in the brain with high pressure liquid chromatography. Physiologist 26: A47, 1983.
Piekut, D. T. and Joseph, S. A.: Relationship of CRF-immunostained cells and magnocellular neurons in the paraventricular nucleus of rat hypothalamus. Peptides 6: 873-882, 198S.
Piekut, D. T. and Joseph, S. A.: Co-existance of CRF and vasopressin immunoreactivity in parvocellular paraventricular neurons of rat hypothalamus. Peptides 7: 891-898, 1986.
Pittman, Q. J., Blume, H. W., and Renaud, L. P.: Connections of the hypothalamic paraventricular nucleus with the neurohypophysis, median eminence, amygdala, lateral septum and midbrain periaqueductal gray: An electrophysiological study in the rat. Brain Research 21S: lS-28, 1981.
Plotsky, P. M.: Hypophyseotropic regulation of adenohypophyseal adrenocorticotropin secretion. Fed.Proc. 44: 207-213, 198S.
Plotsky, P. M.: Facilitation of immunoreactive corticotropin-releasing factor secretion into the hypophysial-portal circulation after activation of catecholaminergic pathways or central norepinephrine injection. Endocrinology 121: 924-930, 1987.
Pohl, M., Carayon, A., Cesselin, F., and Hamon, M.: Angiotensin II-like material extracted from the rat brain is distinct from authentic angiotensin II. J. Neurochem. Sl: 1407-1413, 1988.
Porter, J. P.: Low-Level Electrical Stimulation of the Paraventricular Nucleus Increases Plasma Renin Activity in Conscious Rats. Neuroscience Abst. 12: S39, 1986.

174
Porter, J. P.: Electrical stimulation of the paraventricular nucleus increases plasma renin activity. Am. J. Physiol. 254: R325-R330, 1988.
Porter, J. P.: The renin response to aortic occlusion is enhanced by stimulation of the hypothalamus. Hypertension 12: 52-58, 1988.
Powell, T. P. S., Cowan, W. M., and Raisman, G.: The central olfactory connections. J. Anat. 99: 791-813, 1965.
Printz, M. P., Ganten, D., Unger, T., and Phillips, M. I.: Minireview: The brain renin-angiotensin system 1982. Exp. Brain. Res. 4(supp): 4-52, 1982.
Privitera, P. J. , Webb, J. G. , and Walle, T. : Effects of centrally administered propranolol on plasma renin activity, plasma norepinephrine and arterial pressure. Eur J Pharmacol 54: 51-60, 1979.
Ramsay, D. J., Keil, L. C., Sharpe, M. C., and Shinsako, J.: Angiotensin II infusion increases vasopressin, ACTH and 11-hydroxy-corticosteroid secretion. Am. J. Physiol. 234: R66-R71, 1978.
Rasmussen, A. T.: Innervation of the hypophysis. Endocrinology 23: 263-278, 1938.
Rasmussen, K. and Jacobs, B. L.: Single unit activity of locus coeruleus neurons in the freely moving cat. II. Conditioning and pharmacologic studies. Brain Research 371: 335-344, 1986.
Redgate, E. S.: ACTH release evoked by electrical stimulation of brain stem and limbic system sites in the cat: the absence of ACTH release upon infundibular area stimulation. Endocrinology 4: 806-823, 1970.
Redgate, E. S. and Fahringer, E. E.: A comparison of the pituitary adrenal activity elicited by electrical stimulation of preoptic, amygdaloid and hypothalamic sites in the rat brain. Neuroendocrinology 12: 334-343, 1973.
Reid, I. A., Brooks, V. L., Rudolph, C. D., and Keil, L. C.: Analysis of the actions of angiotensin on the central nervous system of conscious dogs. Am. J. Physiol. 243: R82-R91, 1982.
Reid, I. A., MacDonald, D. M., Pachnis, B., and Ganong, W. F.: Studies concerning the mechanism of suppression of renin secretion by clonidine. J. Pharmacol. Exp. Ther. 192: 713-721, 1975.
Reid, I. A., Morris, B. J., and Ganong, W. F.: The renin-angiotensin system. Ann. Rev. Physiol. 40: 377-410, 1978.
Reid, I. A. and Ramsay, D. J.: The effect of intracerebroventricular administration of renin on drinking and blood pressure. Endocrinology 97: 536-542, 1975.

175
Reid, I. A., Schrier, R. W'., and Earley, L. E.: An effect of extrarenal beta adrenergic stimulation on the release of renin. J. Clin. Invest. 51: 1861-1869, 1972.
Reinstein, D. K., Lehnert, H., and W'urtman, R. J.: Dietary tyrosine suppresses the rise in plasma corticosterone following acute stress in rats. Life Sciences 37: 2157-2163, 1985.
Renaud, L. P. and Arnauld, E.: Supraoptic phasic neurosecretory neurons: response to stimulation of amygdala and dorsal hippocampus and sensitivity to microiontophoresis of amino acids and noradrenaline. Soc. Neurosci. Abstr. 5: 233, 1979.
Ricardo, J. A. and Koh, E.T.: Anatomical evidence of direct projections from the nucleus of the solitary tract to the hypothalamus, amygdala and other forebrain structures in the rat. Brain Research 153: 1-26, 1978.
Richardson, D., Stella, A., Keonetti, G., Barotorelli, A., and Zanchetti, A.: Mechanisms of renal release of renin by electrical stimulation of the brain stem of the cat. Circ. Res. 34: 425-434, 1974.
Richardson Morton, K. D., Johnson, M. D., and Van de Kar, L. D.: Effect of stress on renin secretion is not blocked by adrenal enucleation combined with chemical sympathectomy. Fed. Proc. 47: 3877, 1988.
Richardson Morton, K. D. , Van de Kar, L. D. , and Lorens , S . A. : Stress-induced renin and corticosterone release: effect of dorsal raphe lesions and 5-HT2 antagonist. Fed. Proc. 45: 3019, 1986.
Richardson Morton, K. D., Van de Kar, L. D., Lorens, S. A., Paris, J. M., Urban, J. H., and Kunimoto, K.: The effect of stress on renin and corticosterone secretion is blocked by electrolytic lesions in the mesencephalic dorsal raphe and hypothalamic paraventricular nuclei. Neurosci. Abst. 12: 290.15, 1986.
Richoux, J. P., Cordonnier, J. L., Bouhnik, J., Clauser, E., Corvol, P., Menard, J., and Grignon, G.: Imrnunocytochemical localization of angiotensinogen in rat liver and kidney. Cell Tissue Res. 233: 439-451, 1983.
Rivier, C., Rivier, J., and Vale, W.: Inhibition of adrenocorticotropic hormone secretion in the rat by imrnunoneutralization of corticotropin-releasing factor. Science 218: 377-378, 1982.
Rivier, C. and Vale, W'.: Interaction (CRF) and arginine vasopressin (AVP) docrinology 113: 939-942, 1983.
of corticotropin-releasing factor on ACTH secretion in vivo. En-
Rivier, C. and Vale, W.: Modulation of stress-induced ACTH release by corticotropin-releasing factor, catecholamines and vasopressin. Nature 305: 325-327, 1983.

176
Rivier, J., Speiss, J., and Vale, W.: Characterization of rat hypothalainic corticotropin-releasing factor. Proc. Natl. Acad. Sci. 80: 4851-4855, 1983.
Robertson, G. L.: The regulation of vasopressin function in health and disease. Rec. Prog. Horm. Res. 33: 333-374, 1977.
Romero, J.C., Lazar, J. D., and Hoobler, S. W.: Effects of renal artery constriction and subsequent contralateral nephrectomy on the blood pressure, plasma renin activity, and plasma renin substrate concentration in rabbits. Lab. Invest. 22: 581-587, 1970.
Rossier, J., Battenberg, E., Pittman, Q., Bayon, A., Koda, L., Miller, R., Guilleman, R., and Bloom, F.: Hypothalainic enkephalin neurons may regulate the neurohypophysis. Nature 277: 653-655, 1979.
Rostand, S. G., Work, J., and Luke, R. G.: Effect of reduced chloride reabsorption on renin release in the isolated rat kidney. Pflugers Arch. 405: 46-51, 1985.
Russell, D. M., Dhariwal, A. P. S., Mc Cann, S. M., and Yates, F. E.: Inhibition by dexamethasone of the in vivo pituitary response to corticotropin releasing factor (CRF). Endocrinology 85: 512-521, 1969.
Saavedra, J. M., Israel, A., Plunkett, L. M., Kurihara, M., Shigematsu, K., and Correa, F.M.A.: Quantitative distribution of angiotensin II binding sites in rat brain by autoradiography. Peptides 7: 679, 1986.
Saavedra, J. M., Palkovits, M., Kizer, J. S., Brownstein, M., and Zivin, J. A.: Distribution of biogenic amines and related enzymes in the rat pituitary gland. Journal of Neurochemistry 25: 257-260, 1975.
Saffran, M. and Schally, A. V.: The status of the corticotropin releasing factor (CRF). Neuroendocrinology 24: 359-375, 1977.
Sahu, A., Kalra, S. P., Crowley, W. R., and Kalra, P. S.: Evidence that NPY-containing neurons in the brainstem project inot selected hypothalainic nuclei: implication in feeding behavior. Brain Research 457: 376-378, 1988.
Saper, C. B., Loewy, A. D., Swanson, L. W., and Cowan, W. M.: Direct hypothalamo-autonomic connections. Brain Research 117: 305-312, 1976.
Saper, C. B., Reis, D. J., and Joh, T.: Medullary catecholamine inputs to the anteroventral third ventricular cardiovascular regulatory region in the rat. Neuroscience Letters 42: 285-291, 1983.
Sapolsky, R. M., Krey, L. C., and Mc Ewen, B. S.: Stress down-regulates corticosterone receptors in a site-specific manner in the brain. Endocrinology 114: 287-292, 1984.

177
Sawchenko, P. E.: Adrenalectomy-induced enhancement of CRF and vasopressin immunoreactivity in parvocellular neurosecretory neurons: anatomic, peptide and steroid specificity. The Journal of Neuroscience 7(4): 1093-1106, 1987.
Sawchenko P.E.: Evidence for differential regulation of corticotropinreleasing factor and vasopressin immunoreactivities in parvocellular neurosecretory and autonomic-related projections of the paraventricular nucleus. Brain Research 437: 253-263, 1987.
Sawchenko, P. E. and Swanson, L. W.: A method for tracing biochemically defined pathways in the central nervous system using combined fluorescence retrograde transport and immunohistochemical techniques. Brain Research 210: 31-51, 1981.
Sawchenko, P. E. and Swanson, L. W.: Central noradrenergic pathways for the integration of hypothalamic neuroendocrine and autonomic responses. Science 214(6): 685-687, 1981.
Sawchenko, P. E. and Swanson, L. W.: The distribution and cells of or1g1n of some afferent projections to the paraventricular and supraoptic nuclei in the rat. Soc Neurosci 7: 325, 1981.
Sawchenko, P. E. and Swanson, L. W.: The organization of noradrenergic pathways from the brainstem to the paraventricular and supraoptic nuclei in the rat. Brain Research Reviews 4: 275-325, 1982.
Sawchenko, P. E. and Swanson, L. W.: Immunohistochemical identification of neurons in the paraventricular nucleus of the hypothalamus that project to the medulla or to the spinal cord in the rat. J. Comp. Neurol. 205: 260-272, 1982.
Sawchenko, P. E. and Swanson, L. W.: Localization, colocalization and plasticity of corticotropin-releasing factor immunoreactivity in rat brain. Fed Proc 44: 221-227, 1985.
Sawchenko, P. E., Swanson, L. W., Grzanna, R., Howe, P. R. C., Bloom, S. R., and Polak, J.M.: Colocalization of neuropeptide Y immunoreactivity in brainstem catecholaminergic neurons that project to the paraventricular nucleus of the hypothalamus. The Journal of Comparative Neurology 241: 138-153, 1985.
Sawchenko, P. E., Swanson, L. W., and Joseph, S. A.: The distribution and cells of origin of ACTH (1-39)-stained varicosities in the paraventricular and supraoptic nuclei. Brain Res. 232: 265-274, 1982.
Sawchenko, P. E., Swanson, L. W., Steinbusch, H.W.M., and Verhofstad, A.A.J.: The distribution and cells of origin of serotonergic inputs to the paraventricular and supraoptic nuclei of the rat. Brain Res. 277: 355-360, 1983.

178
Sawchenko, P. E., Swanson, L. W., and Vale, W. W.: Co-expression of corticotropin-releasing factor and vasopressin immunoreactivity in parvocellular neurosecretory neurons of the adrenalectomized rat. Proc. Natl. Acad. Sci. (USA) 81: 1883-1887, 1984.
Sayers, G., White, A., and Long, C. N. H.: Preparation and properties of pituitary adrenotropic hormone. J. Biol. Chem. 149: 425-436, 1943.
Scapagnini, U., Moberg, G. P., Van Loon, G. R., De Groot, J., and Ganong, W. F.: Relation of brain 5-HT content to the diurnal variation in plasma corticosterone in the rat. Neuroendocrinology 7: 90-96, 1971.
Scapagnini, U., Van Loon, G. R., Moberg, G. P., and Ganong, W. F.: Effect of alpha-methyl-para-tyrosine on the circadian variation of plasma corticosterone in rats. European J. Pharmacol. 11: 266-268, 1970.
Schanberg, S. M., Schildkraut, J. J., Breese, G. R., and Kopin, I. J.: Metabolism of norepinephrine-H3 in rat brain- -Identification of conjugated 3-methoxy-4-hydroxyphenylglycol as the major metabolite. Biochem. Pharmac. 17: 247-254, 1968.
Schulte, W., Neus, H., Thones, M., and von Eiff, A. W.: Basal blood pressure variability and reactivity of blood pressure to emotional stress in essential hypertension. Basic Res. Cardiol. 79: 9, 1984. Scroop, G. C., Katie, F., Joy, M. D., and Lowe, R. D.: Importance of central vasomotor effects in angiotensin-induced hypertension. Brit. Med. J. 1: 324-326, 1971.
Sealey, J. E., Atlas, S. A., Laragh, J. H., Silverberg, M., and Kaplan, A. P.: Initiation of plasma prorenin activation by Hageman factordependent conversion of plasma prekallikrein to kallikrein. Proc. Natl. Acad. Sci. 76: 5914-5918, 1979.
Selye, H.: A syndrome produced by diverse nocuous agents. Nature 138: 32, 1936.
Selye, H.: An attempt at a natural classification of the steroids. Nature 151: ]662-663, 1943.
Selye, H.: The general adaptation syndrome and the diseases of adaptation. Journal of Clinical Endocrinology 6: 117-231, 1946.
Selye, H. The Physiology and Pathology of Exposure to Stress. Montreal: Medical Publishers, Inc., 1950.
Selye, H.: Stress and aerospace medicine. Aerospace Med. 44(2): 190-193, 1973.
Selye, H. Stress in Health and Disease. London: Butterworths, 1976.

179
Shade, R. E., Davis, J. 0., Johnson, J. A., and Witty, R. T.: Effects of arterial infusion of sodium and potassium on renin secretion in the dog. Circ. Res. 31: 719-727, 1972.
Share, L.: Interrelations between vasopressin and the renin-angiotensin system. Fed. Proc. 38: 2267-2271, 1979.
Sherlock, D. A., Field, P. M., and Raisman, G.: Retrograde transport of horseradish peroxidase in the magnocellular neurosecretory system of the rat. Brain Research 88: 403-414, 1975.
Shor-Posner, G., Azar, A. P., Jhanwar-Uniyal, M., Filart, R., and Leibowitz, S. F.: Destruction of noradrenergic innervation to the paraventricular nucleus: Deficits in food intake, macronutrient selection, and compensatory eating after food deprivation. Pharmacology Biochemistry and Behavior 25: 381-392, 1986.
Sidman, M.: Avoidance conditioning with brief shock and no extroceptive warning signal. Science 118: 157-158, 1953.
Sigg, E. B., Keim, K. L., and Sigg, T. D.: On the Mechanism of Renin Release by restraint stress in rats. Pharmac. Biochem. Behav. 8(1): 47-50, 1978.
Silverman, A. J., Hoffman, D. L., and Zimmerman, E. A.: The descending afferent connections of the paraventricular nucleus of the hypothalamus. Brain Res. Bull. 6: 47-61, 1981.
Silverman, A. J. and Zimmerman, E. A.: Magnocellular neurosecretory system. Ann. Rev. Neurosci. 6: 357-380, 1983.
Sims, K. B., Hoffman, D. L., Said, S. I., and Zimmerman, E. A.: Vasoactive intestinal polypeptide (VIP) in mouse and rat brain: An immunocytochemical study. Brain Research 186: 165-183, 1980.
Skeggs, L. T., Kahn, J. R., and Shumway, N. P.: The preparation and function of the hypertensin-converting enzyme. J. Exp. Med. 103: 295-299, 1956.
Skofitsch, G., Insel, T. R., and Jacobowitz, D. M.: Binding sites for corticotropin releasing factor in sensory areas of the rat hindbrain and spinal cord. Brain Research Bulletin 15: 519-522, 1985.
Smith, 0. A., DeVito, J. L., and Astley, C. A.: Organization of central nervous system pathways influencing blood pressure responses during emotional behavior. Clin. and Exper. Hyper. A6(1&2): 185-204, 1984.
Smythe, G. A. , Bradshaw, J. E. , and Vining, R. F. : Hypothalamic Monoamine Control of Stress-Induced Adrenocorticotropin Release in the Rat. Endocrinology 113(3): 1062-1071, 1983.

180
Soffer, A.: Regatine and benadine in the diagnosis of pheochromocytoma. Med. Clin. North Am. 38: 375-384, 1954.
Spevack, A. A., Campbell, C. T., and Drake, L.: Effect of amygdalectomy on habituation and CER in rats. Physiol. Behav. 15: 199-207, 1975.
Spinedi, E., Johnston, C. A., Chisari, A., and Negro-Vilar, A.: Role of central epinephrine on the regulation of corticotropin-releasing factor and adrenocorticotropin secretion. Endocrinology 122(5): 1977-1983, 1988.
Spinedi, E. and Negro-Vilar, A.: Angiotensin II and ACTH release: Site of action and potency relative to corticotropin releasing factor and vasopressin. Neuroendocrinology 37: 446-453, 1983.
Squire, L. R. and Zola-Morgan, S. The neurology of memory: the case for correspondence between the findings for man and non-human primate. In J. A. Deutsch (Ed.), The Physiological Basis of Memory. New York: Academic Press, 1983.
Sripairojthikoon, W. and Wyss, J.M.: Cells of origin of the sympathetic renal innervation in rat. Am. J. Physiol. 252: F957-F963, 1987.
Steel, R. G. D. and Torrie, J. H. Principles and Procedures of Statistics with Special Reference to the Biological Sciences. New York: Mc Graw Hill, 1960.
Steele, M. K., Negro-Vilar, A., and Mccann, S. M.: Effect of angiotensin II on in vivo and in vitro release of anterior pituitary hormones in the female rat. Endocrinology 109: 893-899, 1981.
Steinbusch, H. W. M.: Distribution of serotonin-immunoreactivity in the central nervous system of the rat--cell bodies and terminals. Neuroscience 6: 557-618, 1981.
Stellar, E.: The psychology of motivation. Psychol. Rev. 61: 5-22, 1954.
Stockigt, J. R., Collins, R. D., and Biglieri, E.G.: Determination of Plasma renin concentration by angiotensin I radioimmunoassay. Circulation Research 28/29: 175-180, 1971.
Stone, E. A.: Effect of stress on sulfated glycol metabolites of brain norepinephrine. Life Sciences 16: 1725-1730, 1975.
Stone, E. A.: Central noradrenergic activity and the formation of glycol sulfate metabolites of norepinephrine. Life Sciences 19: 1491-1498, 1976.
Suemaru, S., Hashimoto, K., and Ota, Z.: Brain corticotropin-releasing factor (CRF) and catecholamine responses in acutely stressed rats. Endocrinol. Japon. 32(5): 709-718, 1985.

181
Sugden, R. F. and Eccleston, D.: Glycol sulphate ester formation from [14C]noradrenaline in brain and the influence of a COMT inhibitor. J. Neurochem. 18: 2461-2468, 1971.
Swaab, D. F., Nijveldt, F., and Pool, C. W.: Distribution of oxytocin and vasopressin cells in the rat supraoptic and paraventricular nucleus. J. Endocrinol. 67: 461-462, 1975.
Swanson, L. W. The hippocampus and the concept of the limbic system. In W. Siefert (Ed.), Neurobiology of the hippocampus. New York: Academic Press, 1983.
Swanson, L. W. The hypothalamus. In A. Bjorklund,T. Hokfelt and L. W. Swanson (Eds.), Handbook of Chemical Neuroanatomy: Integrated systems of the CNS. Amsterdam: Elsevier, 1987.
Swanson, L. W. and Cowan, W. M.: An autoradiographic study of the organization of the efferent connections of the hippocampal formation in the rat. J. Comp. Neural. 172:49-84, 1977.
Swanson, L. W. and Cowan, W. M.: The connections of the septal region in the rat. J. Comp. Neural. 186: 621-656, 1979.
Swanson, L. W., Kohler, C., and Bjorklund, A. The limbic region. I: The septohippocampal system. In A. Bjorklund,T. Hokfelt and L. W. Swanson (Eds.), Handbook of Chemical Neuroanatomy. Integrated Systems of the CNS. Amsterdam: Elsevier Science Publishers B.V., 1987.
Swanson, L. W. and Kuypers, H. G. J.M.: The paraventricular nucleus of the hypothalamus: Cytoarchitectonic subdivisions and the organization of projections to the pituitary, dorsal vagal complex and spinal cord as demonstrated by retrograde fluorescence double-labeling methods. J. Comp. Neural. 194: 555-570, 1980.
Swanson, L. W. and McKellar, S.: The distribution of oxytocin- and neurophysin-stained fibers in the spinal cord of the rat and monkey. J. Comp. Neurol. 188: 87-106, 1979.
Swanson, L. W. and Sawchenko, P. E.: Paraventricular nucleus: a site for the integration of neuroendocrine and autonomic mechanisms. Neuroendocrinology 31: 410-417, 1980.
Swanson, L. W. and Sawchenko, P. E.: Hypothalamic Integration: Organization of the paraventricular and supraoptic nuclei. Ann. Rev. Neurosci. 6: 269-324, 1983.
Swanson, L. W., Sawchenko, P. E., Berod, A., Hartman, B. K., Helle, K. B., and Vanorden, D. E.: An immunohistochemical study of the organization of catecholaminergic cells and terminal fields in the paraventricular and supraoptic nuclei of the hypothalamus. Journal of Comparative Neurology 196: 271-285, 1981.

182
Swanson, L. W., Sawchenko, P. E., Rivier, J., and Vale, W. W.: Organization of ovine corticotropin-releasing factor immunoreactive cells and fibers in the rat brain: An immunohistochemical study. Neuroendocrinology 36: 165-186, 1983.
Swanson, L. W., Sawchenko, P. E., Wiegand, S. J., and Price, J. L.: Separate neurons in the paraventricular nucleus project to the median eminence and to the medulla or spinal cord. Brain Research 197: 207-212, 1980.
Szafarczyk, A., Alonso, G., Ixart, G., Malaval, F., and Assenmacher, I.: Diurnal-stimulated and stress-induced ACTH release in rats is mediated by ventral noradrenergic bundle. Am. J. Physiol. 249: E219-E226, 1985.
Szafarczyk, A., Malaval, F., Laurent, A., Gibaud, R., and Assenmacher, I.: Further evidence for a central stimulatory action of catecholamines on adrenocorticotropin release in the rat. Endocrinology 121(3): 883-892, 1987.
Tager, H., Hohenboken, M., Markese, J., and Dinerstein, R. J.: Identification and localization of glucagon-related peptides in rat brain. Proc. Natl. Acad. Sci. (USA) 77: 6229-6233, 1980.
Taher, M. S., McLain, L. G., McDonald, K. M., and Schrier, R. W.: Effects of beta-adrenergic blockade on renin response to renal nerve stimulation. J. Clin. Invest. 57:459-465, 1976.
Tanaka, M., Kohno, Y., Nakagawa, R., Ida, Y., Takeda, S., and Nagasaki, N.: Time-related differences in noradrenaline turnover in rat brain regions by stress. Pharmacol. Biochem. Behav. 16(2): 315-319, 1982.
Taugner, C., Poulsen, K., Hackenthal, E., and Taugner, R.: Immunocytochemical localization of renin in mouse kidney. Histochemistry 62: 19-27, 1979.
Taugner, R., Buhrle, C. P., and Nobiling, R.: Ultrastructural changes associated with renin secretion from the juxtaglomerular apparatus of mice. Cell Tiss. Res. 237: 459-472, 1984.
Thierry, A. M., Jovey, F., Glowinski, J., and Kety, S. S.: Effects of stress on the metabolism of norepinephrine, dopamine and serotonin in the central nervous system in the rat. I. Modifications of norepinephrine turnover. J. Pharmacol. Exp. Ther. 163: 163-171, 1968.
Tilde rs, F. J. H. , Berkenbosch, F. , Vermes, I. , Linton, E. A. , and Smelik, P. G.: Role of epinephrine and vasopresson in the control of the pituitary-adrenal response to stress. Fed. Proc. 44(1): 155-160, 1985.
Tobian, L., Tomboulian, A., and Janacek, J.: The effect of high perfusion pressures on the granulation of juxtaglomerular cells in an isolated kidney. J. Clin. Invest. 38: 605-610, 1959.

183
Tramu, G., Croix, C., and Pillez, A.: Ability of the CRF immunoreactive neurons of the paraventricular nucleus to produce a vasopressin-like material. Neuroendocrinology 37: 467-469, 1983.
Tribollet, E. and Dreifuss, J. J.: Localization of neurones projecting to the hypothalamic paraventricular nucleus area of the rat: A horseradish peroxidase study. Neuroscience 6: 1315-1328, 1981.
Tsagarakis, S., Holly, J. M. P., Rees, L. H., Besser, G. M., and Grossman, A.: Acetylcholine and norepinephrine stimulate the release of corticotropin-releasing factor-41 from the rat hypothalamus in vitro. Endocrinology 123: 1962-1969, 1988.
Turkkan, J. S., Brady, J. V., and Harris, A. H. Animal studies of stressful interactions: A behavioral-physiological overview. In L. Goldberger and S. Bregnitz (Eds.), Handbook of Stress-Theoretical and Clinical Aspects. New York: Macmillan Publishing Co., 1982.
Ueda, H., Yasuda, H., Takabatake, Y., Tizuka, M., Tizuka, T., Ihori, M., Yammoto, M., and Sakamoto, Y.: Increased renin release evoked by mesencephalic stimulation in the dog. Jpn. Heart J. 8: 498-506, 1967.
Uhlich, E., Weber, P., Eigler, J., and Groschel-Stewart, U.: Angiotensin-stimulated AVP release in humans. Klin. Wschr. 53: 177-180, 1975.
Ungerstedt, U.: Stereotaxic mapping of the monoamine pathways in the rat brain. Acta Physiol. Scand. 82(367): 1, 1971.
Urban, J. H., Van de Kar, L. D., Lorens, S. A., and Bethea, C. L.: Effect of the anxiolytic drug buspirone on prolactin and corticosterone secretion in stressed and unstressed rats. Pharmacology Biochemistry & Behavior 25(2): 457-462, 1986.
Uretsky, N. J. and Iversen, L. L.: Effects of 6-Hydroxydopamine on Catecholamine containing neurones in the rat brain. Journal of Neurochemistry 17: 269-278, 1970.
Vale, W., Spiess, J., Rivier, C., and Rivier, J.: Characterization of a 41-residue ovine hypothalamic peptide that stimulates secretion of corticotropin and B-endorphin. Science 213: 1394, 1981.
Vale, W., Vaughan, J., Smith, M., Yamamoto, G., Rivier, J., and Rivier, C.: Effects of synthetic ovine corticotropin-releasing factor, glucocorticoids, catecholamines, neurohypophysial peptides, and other substances on cultured corticotropic cells. Endocrinology 113: 1121-1131, 1983.
Valenta, J. G. and Rigby, M. K.: Discrimination of the odor of stressed rats. Science 161: 599-601, 1968.

184
Van de Kar, L. D., Karteszi, M., Bethea, C. L., and Ganong, W. F.: Serotonergic stimulation of prolactin and corticosterone secretion is mediated by different pathways from the mediobasal hypothalamus. Neuroendocrinology 41: 380-384, 1985.
Van de Kar, L. D. and Lorens, S. A.: Differential serotonergic innervation of individual hypothalamic nuclei and other forebrain regions by the dorsal and median raphe nuclei. Brain Res 162: 45-54, 1979.
Van de Kar, L. D., Lorens, S. A., McWilliams, C., Kunimoto, K., Urban, J. H., and Bethea, C. L.: Role of midbrain raphe in stress-induced renin and prolactin secretion. Brain Res. 311: 333-341, 1984.
Van de Kar, L. D., Lorens, S. A., Urban, J. H., Richardson, K. D., Paris, J., and Bethea, C. L.: Pharmacological studies on stress-induced renin and pro lac tin secretion: effects of benzodiazepines, naloxone, propranolol and diisopropylfluorophosphate (DFP). Brain Res. 345: 257-263, 1985.
Van de Kar, L. D., Lorens, S. A., Vodraska, A., Allers, G., Green, M., Van Orden, D. E., and Van Orden, L. S.: Effect of selective midbrain and diencephalic 5, 7-dihydroxytryptamine lesions on serotonin content in individual preoptico-hypothalamic nuclei and on serum luteinizing hormone level. Neuroendocrinology 31: 309-315, 1980.
Van de Kar, L. D., Morton, K. D. R., and Gulati, A.: Fenfluramine-induced suppression of renin secretion is not mediated by peripheral catecholamines or by alpha-2 receptors. Fed. Proc. 47: 3876, 1988.
Van de Kar, L. D., Richardson, K. D., and Urban, J. H.: Serotonin and norepinephrine-dependent effects of fenfluramine on plasma renin activity in conscious rats. Neuropharmacol. 24: 487-494, 1985.
Van de Kar, L. D. and Richardson-Morton, K. D.: Serotonergic regulation of the release of renin is not mediated by the autonomic nervous system but involves beta adrenoceptors. Neuropharmacology 25(5): 487-492, 1986.
Van de Kar, L. D., Urban, J. H., Brownfield, M. S., and Simmons, W. H.: Partial characterization of a renin-releasing factor from plasma and hypothalamus. Hypertension 9(6): 598-606, 1987.
Van de Kar, L. D., Urban, J. H., Lorens, S. A., and Richardson, K. D.: The non-benzodiazepine anxiolytic buspirone inhibits stress-induced renin secretion and lowers heart rate. Life Sci. 36: 1149-1155, 1985.
Van de Kar, L. D., Wilkinson, C. W., and Ganong, W. F.: Pharmacological evidence for a role of brain serotonin in the maintenance of plasma renin activity in unanesthetized rats. J. Pharmacol. Exp. Ther. 219: 85-94, 1981.

185
Van de Kar, L. D., Wilkinson, C. W., Skrobik, Y., Brownfield, M. S., and Ganong, W. F.: Evidence that serotonergic neurons in the dorsal raphe nucleus exert a stimulatory effect on the secretion of renin but not of corticosterone. Brain Research 235: 233-243, 1982.
Van Hoesen, G. W., Wilson, L. M., Mac Dougall, J.M., and Mitchell, J. C.: Selective hippocampal complex deafferentation and de-efferentation and avoidance behavior in rats. Physiology and Behavior 8: 873-879, 1972.
Van Loon, G. R., Scapagnini, U., Moberg, G. P., and Ganong, W. F.: Evidence for central adrenergic neural inhibition of ACTH secretion in the rat. Endocrinology 89: 1464-1469, 1971.
Vander, A. J.: Effect of catecholamines and the renal nerves on renin secretion in anesthetized dogs. Am. J. Physiol. 209(3): 659-662, 1965.
Vander, A. J. and Geelhoed, G. W.: Inhibition of renin secretion by angiotensin II. Proc. Soc. Exp. Biol. Med. 120: 339-403, 1965.
VanderMaelen, C. P., Matheson, G. K., Wilderman, R. C., and Patterson, L. A.: Inhibition of serotonergic dorsal raphe neurons by systemic and iontophoretic administration of buspirone, a non-benzodiazepine anxiolytic drug. Eur. J. Pharmacol. 129: 123-130, 1986.
Vandesande, F. and Dierickx, K.: Identification of the vasopressin producing and of the oxytocin producing neurons in the hypothalamic neurosecretory system of the rat. Cell. Tiss. Res. 164: 153-162, 1975.
Vandongen, R., Peart, W. S., and Boyd, G. W.: Adrenergic stimulation of renin secretion in the isolated perfused kidney. Circulation Research 32: 290-296, 1973.
Velley, L., Mormede, P., and Kempf, E.: Neurochemical lesion of the nucleus locus coeruleus increases neophobia in a specific exploration task but does not modify endocrine response to moderate stress. Pharmacol. Biochem. Behav. 29: 1-7, 1988.
Vellucci, S. V.: The effects of ether stress of betamethasone treatment on the concentrations of noradrenaline and dopamine in various regions of the rat brain. Br. J. Pharmacol. 60: 601-605, 1977.
Vernikos-Danellis, J., Berger, P.A., and Barchas, J. D.: Brain serotonin and pituitary-adrenal function. Prog. Brain Res. 39: 301-309, 1973.
Vernikos-Danellis, J., Kellar, K. J., Kent, D., Gonzales, C., Berger, P. A., and Barchas, J. D.: Serotonin involvement in pituitary-adrenal function. Annals of the New York Academy of Sciences 297: 518-526, 1977.
Wallace, D. M., Magnuson, D. J., and Gray, T. S.: The amygdalo-brainstem pathway: selective innervation of dopaminergic, noradrenergic and adrenergic cells in the rat. Neuroscience Letters 97: 252-258, 1989.

186
Wanaka, A., Kiyama, H., Murakami, T., Matsumoto, M., Kamada, T., Malbon, C. C., and Tohyama, M.: Immunocytochemical localization of beta-adrenergic receptors in the rat brain. Brain Research 485: 125-140, 1989.
Watabe, T., Tanaka, K., Kumagae, M., Itoh, S., Hasegawa, M., Horiuchi, T., Miyabe, S., and Ohno, H.: Diurnal rhythm of plasma immunoreactive corticotropin-releasing factor in normal subjects. Life Sciences 40: 1651-1655, 1987.
Watkins, W. B. : Presence of adrenocorticotropin and beta-endorphin immunoreactivities in the magnocellular neurosecretory system of the rat hypothalamus. Cell. Tiss. Res. 207: 65-80, 1980.
Watson, J. B. Behaviorism. New York: Norton, 1924.
Watson, S. J., Akil, H., Fischli, W., Goldstein, A., Zimmerman, E., Nilaver, G., and van Wiersma Greidanus, T.: Dynorphin and vasopressin: Common localization in magnocellular neurons. Science 216: 85-87, 1982.
Weinberger, M. H., Aoi, W., and Henry, D. P.: Direct effects of beta-adrenergic stimulation on renin release by the rat kidney slice in vitro. Circ Res 37: 318-324, 1975.
Weiner, R. I. and Ganong, W. F.: Role of brain monoamines and histamine in regulation of anterior pituitary secretion. Physiol. Rev. 58: 905-975, 1978.
Weingarten, H. P., Chang, P., and McDonald, T. J.: Comparison of the metabolic and behavioral disturbances following paraventricular- and ventromedial-hypothalamic lesions. Brain Research Bulletin 14: 551-559, 1985.
Werka, T., Skar, J., and Ursin, H.: Exploration and avoidance in rats with lesions in amygdala and piriform cortex. J. Comp. Physiol. Psycho!. 92: 672-681, 1978.
Widmaier, E. P., Lim, A. T., and Vale, W.: Secretion of corticotropin-releasing factor from cultured rat hypothalamic cells: Effects of catecholamines. Endocrinology 124: 583-590, 1989.
Wiegand, S. J. and Price, J. L.: The cells of origin of the afferent fibers to the median eminence in the rat. J. Comp. Neurol. 192: 1-19, 1980.
Wilczynski, E. A. and Osmond, D. H.: Plasma prorenin in humans and dogs species differences and further evidence of a systemic activation cascade. Hypertension 5: 277-285, 1983.
Wilczynski, E. A. and Osmond, D. H.: Extrarenal prorenin in plasma requires an activator of renal origin. Hypertension 8: Suppl II,85-88, 1986.

187
Wilczynski, E. A. and Osmond, D. H.: Evidence for beta-adrenergic regulation of renal and extrarenal plasma prorenin and renin in dogs. Proc. Soc. Exp. Bio.and Med. 182: 208-214, 1986.
Wilkinson, C. W., Shinsako, J., and Dallman, M. F.: Daily rhythms in adrenal responsiveness to adrenocorticotropin are determined primarily by the time of feeding in the rat. Endocrinology 104: 350-359, 1979.
Winn, P., Tarbuck, A., and Dunnett, S. B.: Ibotenic acid lesions of the lateral hypothalamus: comparison with the electrolytic lesion syndrome. Neuroscience 12: 225-240, 1984.
Wynn, P. C., Hauger, R. L., Holmes, M. C., Millan, M. A., Catt, K. J., and Aguilera, G.: Brain and pituitary receptors for corticotropin releasing factor: localization and differential regulation after adrenalectomy. Peptides 5: 1077-1084, 1984.
Yates, F. E. and Urkuhart, J. : Control of plasma concentration of adrenocortical hormones. Physiol. Rev. 42: 359-443, 1962.
Zanchetti, A. and Stella, A.: Neural control of renin release. Clin. Sci. Mol. Med. 48: 215-223, 1975.
Zhang, T. X. and Ciriello, J.: Effect of paraventricular nucleus lesions on arterial pressure and heart rate after aortic baroreceptor denervation in the rat. Brain Research 341: 101-109, 1985.
Zhang, T. X. and Ciriello, J.: Kainic acid lesions of paraventricular nucleus neurons reverse the elevated arterial pressure after aortic baroreceptor denervation in the rat. Brain Research 358: 334-338, 1985.
Zimmerman, E. A., Stillman, M. A., Recht, L. D., Antunes, J. L., and Carmel, P. W.: Vasopressin and corticotropin releasing factor: an axonal pathway to portal capillaries in the zona externa of the median eminence containing vasopressin and its interaction with adrenal corticoids. Ann. N.Y. Acad. Sci. 297: 405-417, 1977.
Zimmerman, H. and Ganong, W. F.: Pharmacological evidence that stimulation of central serotonergic pathways increases renin secretion. Neuroendocrinology 30: 101-107, 1980.

APPROVAL SHEET
The dissertation submitted by Kathy Richardson Morton has been read and approved by the following committee:
Louis D. Van de Kar, Ph.D. Associate Professor Department of Pharmacology Loyola University Stritch School of Medicine
Mark S. Brownfield, Ph.D. Associate Professor Department of Comparative Biosciences University of Wisconsin School of Veterinary Medicine
Stephen B. Jones, Ph.D. Associate Professor Department of Physiology Loyola University Stritch School of Medicine
Alexander G. Karczmar, M.D., Ph.D. Professor Emeritus Department of Pharmacology Loyola University Stritch School of Medicine
Stanley A. Lorens, Ph.D. Professor Department of Pharmacology Loyola University Stritch School of Medicine
T. Celeste Napier, Ph.D. Assistant Professor Department of Pharmacology Loyola University Stritch School of Medicine
The final copies have been examined by the director of the dissertation and the signature which appears below verifies the fact that any necessary changes have been incorporated and that the dissertation is now given final approval by the Committee with reference to content and form.
The dissertation is therefore accepted requirements for the degree of Doctor of
in partial fulfillment Philosophy.
,/ h~ {,
Director's Signature
of the